Embed Size (px)
Citation preview

1521-009X/44/6/871–876$25.00 http://dx.doi.org/10.1124/dmd.116.070235DRUG METABOLISM AND DISPOSITION Drug Metab Dispos 44:871–876, June 2016U.S. Government work not protected by U.S. copyright
p38 MAP Kinase Links CAR Activation and Inactivation in theNucleus via Phosphorylation at Threonine 38 s
Takeshi Hori, Rick Moore, and Masahiko Negishi
Pharmacogenetics Section, Reproductive and Developmental Biology Laboratory, National Institute of Environmental HealthSciences, National Institutes of Health, Research Triangle Park, North Carolina
Received February 25, 2016; accepted April 1, 2016
ABSTRACT
Nuclear receptor constitutive androstane receptor (CAR, NR1I3),which regulates hepatic drug and energy metabolisms as well as cellgrowth and death, is sequestered in the cytoplasmas its inactive formphosphorylated at threonine 38. CAR activators elicit dephosphory-lation, and nonphosphorylated CAR translocates into the nucleus toactivate its target genes. CAR was previously found to require p38mitogen-activated protein kinase (MAPK) to transactivate the cyto-chromeP450 2B (CYP2B) genes. Herewe havedemonstrated that p38
MAPK forms a complex with CAR, enables it to bind to the responsesequence, phenobarbital-responsive enhancer module (PBREM),within the CYP2B promoter, and thus recruits RNA polymerase II toactivate transcription. Subsequently, p38 MAPK elicited rephosphor-ylation of threonine 38 to inactivate CAR and exclude it from thenucleus. Thus, nuclear p38 MAPK exerted dual regulation by sequen-tially activating and inactivating CAR-mediated transcription throughphosphorylation of threonine 38.
Introduction
The nuclear receptor constitutive androstane receptor (CAR, NR1I3),a member of the thyroid and steroid hormone receptor superfamily,activates genes encoding for enzymes and transporters that metabolizeand excrete therapeutic drugs and is activated by these drugs andxenobiotics (Kobayashi et al., 2015). With these functions, CARcritically regulates hepatic capability for drug disposition. CAR gainsits drug responsiveness by suppressing its high constitutive activityby phosphorylation at threonine 38 within the DNA-binding domain(DBD) (Mutoh et al., 2009). Phosphorylation of CAR abolishes itsDNA binding ability and is sequestered in the cytoplasm. Phenobarbital,the classic drug that indirectly activates CAR, elicits a cell signal thatstimulates protein phosphatase 2A (PP2A) to dephosphorylate threonine38 for activation (Mutoh et al., 2013). Thus, the cell signal–mediatedprocess of CAR activation as it occurs in the cytoplasm is now welldocumented. Here we have investigated p38 mitogen-activated proteinkinase (MAPK) that regulates CAR in the nucleus.Phosphorylation has long been investigated in both ligand-dependent
and -independent regulation of nuclear receptors. It is involved indegradation, cofactor recruitment, and dimerization (Shao and Lazar,1999; Tremblay et al., 1999; Hong et al., 2003; Picard et al., 2008). Theamino acid residues targeted for studies reside within the activationfunction 1 (AF-1) or ligand-binding domain (LBD) region and have
been investigated in a given but not in any other nuclear receptors.Compared with these regions, much less emphasis has been placed onphosphorylation within the DBD. One phosphorylation site resideswithin a protein kinase C (PKC) motif in the DBD and is conserved in41 out of 48 human nuclear receptors. Phosphorylation of this conservedsite is the most well characterized within threonine 38 of CAR (Mutohet al., 2009; Mutoh et al., 2013). CAR is, in fact, phosphorylated atthreonine 38 in mouse hepatocytes to regulate its drug activation.Recently, estrogen receptor alpha (ERa) phosphorylated at the corre-sponding serine 216 was also found in mouse neutrophils and appears toregulate their infiltration into the uterus (Shindo et al., 2013). Althoughtheir phosphorylation in tissues in vivo have not yet been confirmed,studies with phosphomimetic mutants suggest that these residues inhepatocyte nuclear factor 4 alpha (HNF-4a), vitamin D receptor (VDR),peroxisome proliferator-activated receptor alpha, retinoid X receptoralpha (RXRa), and farnesoid X receptor may regulate various function-alities of these nuclear receptors, such as cytoplasmic retention, degrada-tion, and transactivation (Hsieh et al., 1993; Sun et al., 2007; Gineste et al.,2008). Therefore, conserved phosphorylation has provided us with theopportunity to uniformly investigate nuclear receptors but has only beenstudied as the target of PKC or dephosphorylation by PP2A.p38 MAPK is activated in response to various extracellular stimuli,
such as growth factors, UV radiation, inflammatory cytokines, oxidativestress, and hyperosmosis (Freshney et al., 1994; Han et al., 1994; Rouseet al., 1994; Huot et al., 1997; Zhang and Jope, 1999). In liver cells,phosphorylated p38 MAPK is accumulated in the nucleus to activatevarious transcription factors and protein kinases. We have recentlydemonstrated that p38 MAPK is essential for CAR to activate thecytochrome P450 2B6 (CYP2B6) gene in human hepatoma HepG2-derived cells (Saito et al., 2013b). In the present study, we revealed that
This work was supported by the Intramural Research Program of the NationalInstitutes of Health and National Institute of Environmental Health Sciences [GrantZ01ES71005-01].
dx.doi.org/10.1124/dmd.116.070235.s This article has supplemental material available at dmd.aspetjournals.org.
ABBREVIATIONS: CAR, constitutive androstane receptor; ChIP, chromatin immunoprecipitation; DBD, DNA-binding domain; DMSO, dimethylsulfoxide; ER, estrogen receptor; FBS, fetal bovine serum; GST, glutathione S-transferase; HNF, hepatocyte nuclear factor; HRP, horseradishperoxidase; LBD, ligand-binding domain; MAPK, mitogen-activated protein kinase; PBREM, phenobarbital-responsive enhancer module; PCR,polymerase chain reaction; PGC, peroxisome proliferator-activated receptor -g coactivator; PP2A, protein phosphatase 2A; RXR, retinoid Xreceptor; SRC, steroid receptor coactivator; TBS, Tris-buffered saline; TCPOBOP, 1,4-bis[2-(3,5-dichloropyridyloxy)]benzene; VDR, vitamin Dreceptor.
871
http://dmd.aspetjournals.org/content/suppl/2016/04/12/dmd.116.070235.DC1Supplemental material to this article can be found at:
at ASPE
T Journals on June 18, 2018
dmd.aspetjournals.org
Dow
nloaded from
p38 MAPK forms a complex with CAR to promote the CAR-mediatedtranscription in the nucleus. Moreover, p38 MAPK linked CARtransactivation and inactivation by stimulating phosphorylation ofthreonine 38.
Materials and Methods
Materials. Phenobarbital sodium salt, 1,4-bis[2-(3,5-dichloropyridyloxy)]benzene (TCPOBOP), anisomycin, SB 239063, anti-FLAG M2 affinity gel,anti-FLAG M2-horseradish peroxidase (HRP) antibodies, phosphatase inhibitorcocktails 2 and 3, and L-glutathione reduced were purchased from Sigma-Aldrich(St. Louis, MO); anti-p38a (ab7952) and anti-phospho-RNA polymerase II CTD(ab5131) from Abcam (Cambridge, MA); anti-phospho-p38 MAPK (Thr180/Tyr182; #4511) and anti-p38 MAPK from Cell Signaling Technology (Danvers,MA); anti-RXRa (sc-553 X) and HRP-conjugated anti-mouse or rabbit IgGantibodies from Santa Cruz Biotechnology (Dallas, TX); and anti-V5 fromInvitrogen/ThermoFisher Scientific (Carlsbad, CA). Anti-phospho-Thr38 peptideantibodies for CAR were produced in our previous work (Mutoh et al., 2009).Anti-phospho-Ser51 peptide antibodies (GFFRR-pS-MKRKALFTC) for VDRwere produced by AnaSpec Inc. (San Jose, CA). A Lipofectamine 2000 reagentand Dynabeads Protein G were obtained from Life Technologies/ThermoFisherScientific (Grand Island, NY); TaqMan Gene Expression Assays (probe andprimer sets) for CYP2B6 (AssayID: Hs00167937_m1) (FAM), Cyp2b10(AssayID: Mm00456591_m1) (FAM), Cyp2c55 (AssayID: Mm00472168_m1)(FAM), and human and mouse GAPDH (FAM) from Applied Biosystems(Foster, CA); cOmplete mini protease inhibitor cocktail tablets from RocheDiagnostics Corp. (Indianapolis, IN); recombinant active p38 MAPK (#14-587)from Millipore UK Limited (Dundee, UK).
Animals. Both Car+/+and Car2/2 mice in C3H/HeNCrlBR background wereproduced in house (Yamamoto et al., 2004). Phenobarbital (100 mg/kg body
weight) in phosphate-buffered saline (PBS), TCPOBOP (3mg/kg bodyweight) indimethyl sulfoxide (DMSO) in corn oil, or a control solution, was intraperito-neally injected into 7- to 8-week-old male mice for a treatment of 6 hours, fromwhich liver RNAs and nuclei were prepared for real-time polymerase chainreaction (PCR) and chromatin immunoprecipitation (ChIP) assays, respectively.Mice were maintained under the standard condition at the National Instituteof Environmental Health Sciences, and animal experiments were conductedaccording to protocols approved by the animal ethics committee at NIEHS/National Institutes of Health.
Plasmid Construction. Mouse CAR cDNA (GenBank accession no. NM009803.5) was previously cloned into pGEX-4T-3 vector (GE Healthcare,Piscataway, NJ) for glutathione S-transferase (GST)-CAR fusion proteinsand pCR3 vector (Invitrogen). CAR Thr48Ala (T48A) and CAR Thr48Asp(T48D) mutants were generated by a site-directed mutagenesis method with thefollowing primers: 59-GGCTTCTTCAGACGAgCAGTCAGCAAAACCATT-39and 59 -AATGGTTTTGCTGACTGcTCGTCTGAAGAAGCC-3 9 forCAR T48A; 59-GGCTTCTTCAGACGAgatGTCAGCAAAACCATT-39 and59-AATGGTTTTGCTGACatcTCGTCTGAAGAAGCC-39 for CAR T48D.Mouse p38a cDNA (NM 011951.3) was cloned into pcDNA3.1 vector(Invitrogen). The FLAG tag was inserted into the 59-flanking region of CAR inpCR3 or p38a in pcDNA3.1.
Cell Cultures. Primary hepatocytes were isolated using a collagenase two-stepperfusion method as described previously (Honkakoski et al., 1996). Hepatocytes(6� 105 cells/ml per well) were seeded on collagen-coated wells and cultured inWilliams’ medium E containing 10% fetal bovine serum (FBS) and penicillin/streptomycin. After 3 hours of seeding, cells were treatedwith 2mMphenobarbitalor 250 nM TCPOBOP for 12 hours in the presence or absence of 20 mMSB 239063, and total RNA was extracted for real-time PCR analysis. Huh-7 cellsor HepG2-derived Ym17 cells were cultured in minimum essential medium(Invitrogen) or Dulbecco’s modified Eagle’s medium (Invitrogen), respectively,
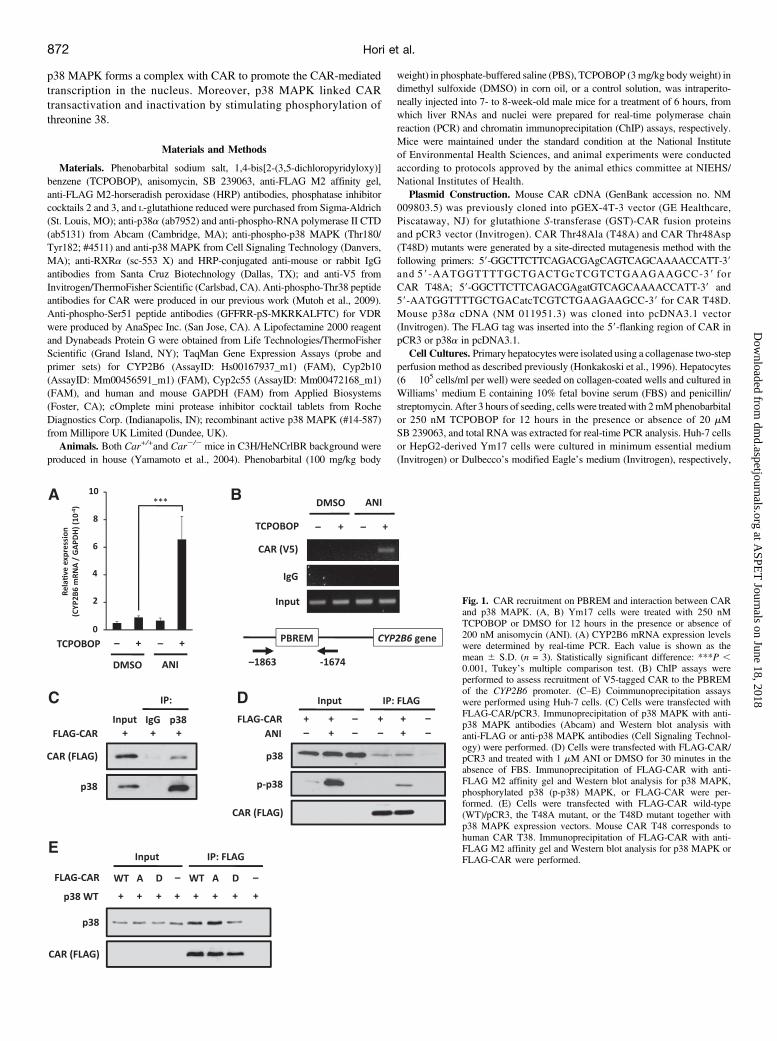
Fig. 1. CAR recruitment on PBREM and interaction between CARand p38 MAPK. (A, B) Ym17 cells were treated with 250 nMTCPOBOP or DMSO for 12 hours in the presence or absence of200 nM anisomycin (ANI). (A) CYP2B6 mRNA expression levelswere determined by real-time PCR. Each value is shown as themean 6 S.D. (n = 3). Statistically significant difference: ***P ,0.001, Tukey’s multiple comparison test. (B) ChIP assays wereperformed to assess recruitment of V5-tagged CAR to the PBREMof the CYP2B6 promoter. (C–E) Coimmunoprecipitation assayswere performed using Huh-7 cells. (C) Cells were transfected withFLAG-CAR/pCR3. Immunoprecipitation of p38 MAPK with anti-p38 MAPK antibodies (Abcam) and Western blot analysis withanti-FLAG or anti-p38 MAPK antibodies (Cell Signaling Technol-ogy) were performed. (D) Cells were transfected with FLAG-CAR/pCR3 and treated with 1 mM ANI or DMSO for 30 minutes in theabsence of FBS. Immunoprecipitation of FLAG-CAR with anti-FLAG M2 affinity gel and Western blot analysis for p38 MAPK,phosphorylated p38 (p-p38) MAPK, or FLAG-CAR were per-formed. (E) Cells were transfected with FLAG-CAR wild-type(WT)/pCR3, the T48A mutant, or the T48D mutant together withp38 MAPK expression vectors. Mouse CAR T48 corresponds tohuman CAR T38. Immunoprecipitation of FLAG-CAR with anti-FLAG M2 affinity gel and Western blot analysis for p38 MAPK orFLAG-CAR were performed.
872 Hori et al.
at ASPE
T Journals on June 18, 2018
dmd.aspetjournals.org
Dow
nloaded from

supplemented with 10% FBS and penicillin/streptomycin at 37�C with 5% CO2.Plasmids were transfected into Huh-7 or Ym17 cells with a Lipofectamine 2000reagent according to the manufacturer’s instructions.
Real-Time PCR. Total RNAwas extracted frommouse livers, hepatocytes, orYm17 cell using TRIzol reagent (Life Technologies), with which cDNAs weresynthesized using a High Capacity cDNA Archive kit (Life Technologies). Real-time PCR was conducted using a TaqMan Universal PCR Master mix andTaqMan probes and primers with a 7900HT Fast Real-Time PCR System(Applied Biosystems).
Immunoprecipitation. Huh-7 cells were lysed in a lysis buffer [20 mM Tris-HCl (pH 7.5), 0.5 mM EDTA, 100 mM NaCl, 1% Triton X-100, 10% glycerol,and protease inhibitor cocktails 2 and 3]. After sonication and centrifugation, theresulting supernatant was incubated with Dynabeads with a given antibody or ananti-FLAG M2 affinity gel (FLAG gel). Dynabeads or FLAG gel were washedthree to four times in a lysis buffer or Tris-buffered saline [TBS; 25 mMTris-HCl(pH 7.4), 140 mMNaCl, and 2.7 mMKCl], respectively. Washed FLAG gel washeat-treated in 2� SDS sample buffer [157 mMTris-HCl (pH 6.8), 4% SDS, 25%glycerol, and 0.01% bromophenol blue]. Washed Dynabeads were incubatedand centrifuged in 0.1 M glycine buffer (pH 2.0) to elute proteins that weresubsequently heat-treated in the above-mentioned 2� SDS sample buffer.
ChIP Assays. ChIP assay was performed using ChIP-IT Express kit (ActiveMotif, Carlsbad, CA) as described previously (Saito et al., 2013a; Gotoh andNegishi, 2015) with some modifications. Sheared chromatin (10 mg) obtainedfrom mouse livers or Ym17 cells was incubated with the indicated antibodies(0.3–1 mg; anti-phospho RNA polymerase II, anti-RXRa, anti-phospho-p38,anti-V5, or normal IgG) and magnetic beads. After washing beads, DNA waseluted and purified. DNA fragments were amplified by PCR with the followingspecific primers: 59-TTACTGTGTGTAAAGCACTTC-39 and 59-GACAAA-CAGTCCTATTTGTAAG-39 [for the phenobarbital-responsive enhancermodule (PBREM) of the CYP2B6 gene; PBREM/CYP2B6] (the amplicon is
from –1863 to –1674; –1863/–1674); 59-GCTAATGCCTGTCTGGAT-CAGGA-39 and 59-GGAATACTGACCCAAGTTCAGTG-39 (PBREM/Cyp2b10) (–2434/–2232); 59-AAGGGAATGAGGAGTGAGC-39 and 59-CAA-GAAGCCCACAAGGAGAG-39 (TATA box/Cyp2b10) (–149/+75);59-GCTTCTCTTTGCCCTCGATA-39 and 59-ACCCAAGTCCCCTGTACCT-TAC-39 (direct repeat 4 (DR4)/Cyp2c55) (–1860/–1623); 59-GGCCAGAGTC-CATTCAGAAG-39 and 59-GAGCTTCCCTCTCCCAGAGT-39 (TATA box/Cyp2c55) (–115/+114).
In Vitro Kinase Assay. Huh-7 cells, which expressed ectopic FLAG-p38MAPK, were treated with 1 mM anisomycin for 10 minutes. These cells werelysed in the above-mentioned lysis buffer containing 2.5 mMNa4P2O7 and 1 mMNa3VO4. Anti-FLAG M2 affinity gels, which were pretreated with dimethylpimelimidate (1 mg/ml) and triethanolamine (100 mM), were incubated withlysates for 3 hours at 4�C and were washed four to five times with TBS. Resultinggels were used as an enzyme source for active p38 MAPK. For substrates, GST-CAR wild-type and the T48A mutant were expressed and purified as previouslyreported (Mutoh et al., 2009). In kinase reaction, GST-CAR (1 mg) and FLAG-p38 MAPK-bound gel were mixed in a kinase buffer consisting of 25 mM Tris-HCl (pH 7.5), 2.5 mM Na4P2O7, 1 mM Na3VO4, 10 mM MgCl2, and 1 mMdithiothreitol. Reaction was started by adding 200 mMATP, incubated for 1 hourat 37�C, and stopped by adding 1� SDS sample buffer. Phosphorylated CARwasdetected by Western blot analysis using anti-pThr38 CAR antibodies and anti-rabbit IgG (light chain–specific)–HRP antibodies (Jackson ImmunoResearchLaboratories, West Grove, PA).
Bacterially expressed active p38 MAPK (0.36 mg) was incubated with GST-CAR (0.5 mg) and ATP (250 mM) in the above-mentioned kinase buffer for6 hours at 37�C, and then Western blot analysis was performed to detectphosphorylated CAR.
Western Blot. Proteins were separated in a 10% SDS-polyacrylamide gel andwere transferred onto a polyvinylidene difluoride membrane. Membranes were
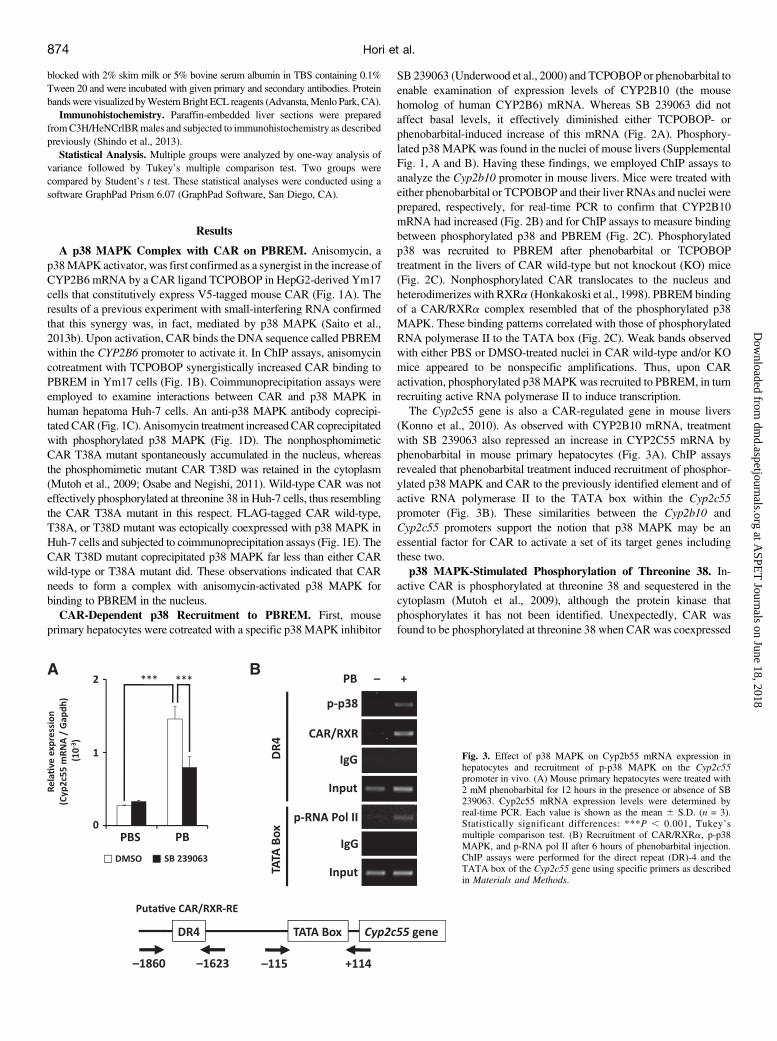
Fig. 2. Effect of p38 MAPK on Cyp2b10 mRNA expression inhepatocytes and recruitment of p-p38 MAPK on the Cyp2b10promoter in vivo. (A) Mouse primary hepatocytes were treated with2 mM phenobarbital or 250 nM TCPOBOP for 12 hours in thepresence or absence of SB 239063. Cyp2b10 mRNA expressionlevels were determined by real-time PCR. Each value is shown asthe mean 6 S.D. (n = 3). Statistically significant differences: **P,0.01 and ***P , 0.001, Tukey’s multiple comparison test. (B)Effect of treatment with phenobarbital (100 mg/kg) or TCPOBOP(3 mg/kg) for 6 hours on Cyp2b10 mRNA expression in livers.Each value is shown as the mean 6 S.D. (n = 3). Statisticallysignificant difference: ***P , 0.001, Student’s t test. (C)Recruitment of CAR/RXRa, p-p38 MAPK, and phosphorylatedRNA polymerase II (p-RNA pol II) after 6 hours of phenobarbital orTCPOBOP injection. ChIP assays were performed for the PBREMand the TATA box of the Cyp2b10 gene using specific primersas described in Materials and Methods. PB or TC indicatesphenobarbital or TCPOBOP, respectively.
p38 MAPK Links CAR Activation and Inactivation 873
at ASPE
T Journals on June 18, 2018
dmd.aspetjournals.org
Dow
nloaded from
blocked with 2% skim milk or 5% bovine serum albumin in TBS containing 0.1%Tween 20 and were incubated with given primary and secondary antibodies. Proteinbandswere visualized byWesternBright ECL reagents (Advansta,Menlo Park, CA).
Immunohistochemistry. Paraffin-embedded liver sections were preparedfromC3H/HeNCrlBRmales and subjected to immunohistochemistry as describedpreviously (Shindo et al., 2013).
Statistical Analysis. Multiple groups were analyzed by one-way analysis ofvariance followed by Tukey’s multiple comparison test. Two groups werecompared by Student’s t test. These statistical analyses were conducted using asoftware GraphPad Prism 6.07 (GraphPad Software, San Diego, CA).
Results
A p38 MAPK Complex with CAR on PBREM. Anisomycin, ap38MAPK activator, was first confirmed as a synergist in the increase ofCYP2B6 mRNA by a CAR ligand TCPOBOP in HepG2-derived Ym17cells that constitutively express V5-tagged mouse CAR (Fig. 1A). Theresults of a previous experiment with small-interfering RNA confirmedthat this synergy was, in fact, mediated by p38 MAPK (Saito et al.,2013b). Upon activation, CAR binds the DNA sequence called PBREMwithin the CYP2B6 promoter to activate it. In ChIP assays, anisomycincotreatment with TCPOBOP synergistically increased CAR binding toPBREM in Ym17 cells (Fig. 1B). Coimmunoprecipitation assays wereemployed to examine interactions between CAR and p38 MAPK inhuman hepatoma Huh-7 cells. An anti-p38 MAPK antibody coprecipi-tated CAR (Fig. 1C). Anisomycin treatment increasedCAR coprecipitatedwith phosphorylated p38 MAPK (Fig. 1D). The nonphosphomimeticCAR T38A mutant spontaneously accumulated in the nucleus, whereasthe phosphomimetic mutant CAR T38D was retained in the cytoplasm(Mutoh et al., 2009; Osabe and Negishi, 2011). Wild-type CAR was noteffectively phosphorylated at threonine 38 in Huh-7 cells, thus resemblingthe CAR T38A mutant in this respect. FLAG-tagged CAR wild-type,T38A, or T38D mutant was ectopically coexpressed with p38 MAPK inHuh-7 cells and subjected to coimmunoprecipitation assays (Fig. 1E). TheCAR T38D mutant coprecipitated p38 MAPK far less than either CARwild-type or T38A mutant did. These observations indicated that CARneeds to form a complex with anisomycin-activated p38 MAPK forbinding to PBREM in the nucleus.CAR-Dependent p38 Recruitment to PBREM. First, mouse
primary hepatocytes were cotreated with a specific p38 MAPK inhibitor
SB 239063 (Underwood et al., 2000) and TCPOBOP or phenobarbital toenable examination of expression levels of CYP2B10 (the mousehomolog of human CYP2B6) mRNA. Whereas SB 239063 did notaffect basal levels, it effectively diminished either TCPOBOP- orphenobarbital-induced increase of this mRNA (Fig. 2A). Phosphory-lated p38 MAPK was found in the nuclei of mouse livers (SupplementalFig. 1, A and B). Having these findings, we employed ChIP assays toanalyze the Cyp2b10 promoter in mouse livers. Mice were treated witheither phenobarbital or TCPOBOP and their liver RNAs and nuclei wereprepared, respectively, for real-time PCR to confirm that CYP2B10mRNA had increased (Fig. 2B) and for ChIP assays to measure bindingbetween phosphorylated p38 and PBREM (Fig. 2C). Phosphorylatedp38 was recruited to PBREM after phenobarbital or TCPOBOPtreatment in the livers of CAR wild-type but not knockout (KO) mice(Fig. 2C). Nonphosphorylated CAR translocates to the nucleus andheterodimerizes with RXRa (Honkakoski et al., 1998). PBREMbindingof a CAR/RXRa complex resembled that of the phosphorylated p38MAPK. These binding patterns correlated with those of phosphorylatedRNA polymerase II to the TATA box (Fig. 2C). Weak bands observedwith either PBS or DMSO-treated nuclei in CAR wild-type and/or KOmice appeared to be nonspecific amplifications. Thus, upon CARactivation, phosphorylated p38MAPKwas recruited to PBREM, in turnrecruiting active RNA polymerase II to induce transcription.The Cyp2c55 gene is also a CAR-regulated gene in mouse livers
(Konno et al., 2010). As observed with CYP2B10 mRNA, treatmentwith SB 239063 also repressed an increase in CYP2C55 mRNA byphenobarbital in mouse primary hepatocytes (Fig. 3A). ChIP assaysrevealed that phenobarbital treatment induced recruitment of phosphor-ylated p38 MAPK and CAR to the previously identified element and ofactive RNA polymerase II to the TATA box within the Cyp2c55promoter (Fig. 3B). These similarities between the Cyp2b10 andCyp2c55 promoters support the notion that p38 MAPK may be anessential factor for CAR to activate a set of its target genes includingthese two.p38 MAPK-Stimulated Phosphorylation of Threonine 38. In-
active CAR is phosphorylated at threonine 38 and sequestered in thecytoplasm (Mutoh et al., 2009), although the protein kinase thatphosphorylates it has not been identified. Unexpectedly, CAR wasfound to be phosphorylated at threonine 38 when CAR was coexpressed
Fig. 3. Effect of p38 MAPK on Cyp2b55 mRNA expression inhepatocytes and recruitment of p-p38 MAPK on the Cyp2c55promoter in vivo. (A) Mouse primary hepatocytes were treated with2 mM phenobarbital for 12 hours in the presence or absence of SB239063. Cyp2c55 mRNA expression levels were determined byreal-time PCR. Each value is shown as the mean 6 S.D. (n = 3).Statistically significant differences: ***P , 0.001, Tukey’smultiple comparison test. (B) Recruitment of CAR/RXRa, p-p38MAPK, and p-RNA pol II after 6 hours of phenobarbital injection.ChIP assays were performed for the direct repeat (DR)-4 and theTATA box of the Cyp2c55 gene using specific primers as describedin Materials and Methods.
874 Hori et al.
at ASPE
T Journals on June 18, 2018
dmd.aspetjournals.org
Dow
nloaded from
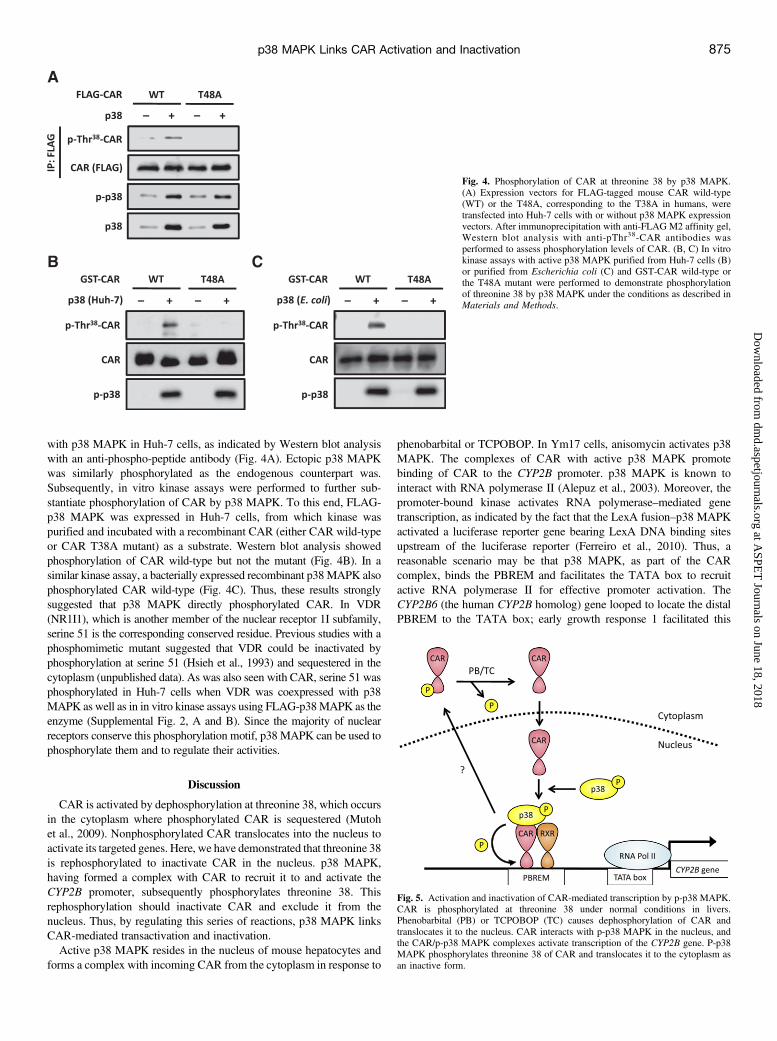
with p38 MAPK in Huh-7 cells, as indicated by Western blot analysiswith an anti-phospho-peptide antibody (Fig. 4A). Ectopic p38 MAPKwas similarly phosphorylated as the endogenous counterpart was.Subsequently, in vitro kinase assays were performed to further sub-stantiate phosphorylation of CAR by p38 MAPK. To this end, FLAG-p38 MAPK was expressed in Huh-7 cells, from which kinase waspurified and incubated with a recombinant CAR (either CAR wild-typeor CAR T38A mutant) as a substrate. Western blot analysis showedphosphorylation of CAR wild-type but not the mutant (Fig. 4B). In asimilar kinase assay, a bacterially expressed recombinant p38MAPK alsophosphorylated CAR wild-type (Fig. 4C). Thus, these results stronglysuggested that p38 MAPK directly phosphorylated CAR. In VDR(NR1I1), which is another member of the nuclear receptor 1I subfamily,serine 51 is the corresponding conserved residue. Previous studies with aphosphomimetic mutant suggested that VDR could be inactivated byphosphorylation at serine 51 (Hsieh et al., 1993) and sequestered in thecytoplasm (unpublished data). As was also seen with CAR, serine 51 wasphosphorylated in Huh-7 cells when VDR was coexpressed with p38MAPK as well as in in vitro kinase assays using FLAG-p38MAPK as theenzyme (Supplemental Fig. 2, A and B). Since the majority of nuclearreceptors conserve this phosphorylation motif, p38 MAPK can be used tophosphorylate them and to regulate their activities.
Discussion
CAR is activated by dephosphorylation at threonine 38, which occursin the cytoplasm where phosphorylated CAR is sequestered (Mutohet al., 2009). Nonphosphorylated CAR translocates into the nucleus toactivate its targeted genes. Here, we have demonstrated that threonine 38is rephosphorylated to inactivate CAR in the nucleus. p38 MAPK,having formed a complex with CAR to recruit it to and activate theCYP2B promoter, subsequently phosphorylates threonine 38. Thisrephosphorylation should inactivate CAR and exclude it from thenucleus. Thus, by regulating this series of reactions, p38 MAPK linksCAR-mediated transactivation and inactivation.Active p38 MAPK resides in the nucleus of mouse hepatocytes and
forms a complex with incoming CAR from the cytoplasm in response to
phenobarbital or TCPOBOP. In Ym17 cells, anisomycin activates p38MAPK. The complexes of CAR with active p38 MAPK promotebinding of CAR to the CYP2B promoter. p38 MAPK is known tointeract with RNA polymerase II (Alepuz et al., 2003). Moreover, thepromoter-bound kinase activates RNA polymerase–mediated genetranscription, as indicated by the fact that the LexA fusion–p38 MAPKactivated a luciferase reporter gene bearing LexA DNA binding sitesupstream of the luciferase reporter (Ferreiro et al., 2010). Thus, areasonable scenario may be that p38 MAPK, as part of the CARcomplex, binds the PBREM and facilitates the TATA box to recruitactive RNA polymerase II for effective promoter activation. TheCYP2B6 (the human CYP2B homolog) gene looped to locate the distalPBREM to the TATA box; early growth response 1 facilitated this
Fig. 5. Activation and inactivation of CAR-mediated transcription by p-p38 MAPK.CAR is phosphorylated at threonine 38 under normal conditions in livers.Phenobarbital (PB) or TCPOBOP (TC) causes dephosphorylation of CAR andtranslocates it to the nucleus. CAR interacts with p-p38 MAPK in the nucleus, andthe CAR/p-p38 MAPK complexes activate transcription of the CYP2B gene. P-p38MAPK phosphorylates threonine 38 of CAR and translocates it to the cytoplasm asan inactive form.
Fig. 4. Phosphorylation of CAR at threonine 38 by p38 MAPK.(A) Expression vectors for FLAG-tagged mouse CAR wild-type(WT) or the T48A, corresponding to the T38A in humans, weretransfected into Huh-7 cells with or without p38 MAPK expressionvectors. After immunoprecipitation with anti-FLAG M2 affinity gel,Western blot analysis with anti-pThr38-CAR antibodies wasperformed to assess phosphorylation levels of CAR. (B, C) In vitrokinase assays with active p38 MAPK purified from Huh-7 cells (B)or purified from Escherichia coli (C) and GST-CAR wild-type orthe T48A mutant were performed to demonstrate phosphorylationof threonine 38 by p38 MAPK under the conditions as described inMaterials and Methods.
p38 MAPK Links CAR Activation and Inactivation 875
at ASPE
T Journals on June 18, 2018
dmd.aspetjournals.org
Dow
nloaded from

looping, possibly enabling the promoter to recruit RNA polymerase IIfor effective promoter activation (Inoue and Negishi, 2008, 2009). Inaddition to early growth response 1, various other transcription factorswere shown to interact with and activate the CYP2B promoters togetherwith CAR, such as peroxisome proliferator-activated receptor -gcoactivator-1 alpha (PGC-1a), steroid receptor coactivator-1 (SRC-1),HNF-4a, and CCAAT/enhancer-binding protein alpha (Shiraki et al.,2003; Li et al., 2008; Benet et al., 2010). Since p38 MAPK can activatethese factors (Lim et al., 1998; Knutti et al., 2001; Guo et al., 2007; Qiaoet al., 2006; Fernandez-Marcos and Auwerx, 2011; Antoon et al., 2012),it may enable CAR to bind to PBREM and to loop the promoter byphosphorylating one or more of them. In addition to the CYP2B genes,p38 MAPK similarly regulated the Cyp2c55 gene in mouse primaryhepatocytes. This indicates that the regulation observedwith p38MAPKmay be a general mechanism by which CAR activates hepatic genes.We now know that CAR can be phosphorylated in the nucleus.
Phosphorylated CAR moves back to the cytoplasm, possibly beingretained for reactivation by dephosphorylation. Thus, it is possible thatCAR can be recycled through phosphorylation, dephosphorylation, andrephosphorylation of threonine 38 during drug inductions. Figure 5schematizes the hypothesis that p38 MAPK plays dual functions andlinks the CAR-mediated activation and inactivation of the CYP2B genesthrough phosphorylation/dephosphorylation of threonine 38. The phos-phorylation of ERa is also known to link its activation and inactivation(Zhou and Slingerland, 2014). However, unlike CAR, which can bereused, phosphorylated ERa is degraded through ubiquitination and,therefore, cannot be recycled.In conclusion, our present study shed light on phosphorylation within
the DBD that has long been ignored. Our present findings with CAR andp38 MAPK can be used to understand biologic roles of this phosphor-ylation among members of the nuclear receptor superfamily.
Acknowledgments
The authors thank the NIEHS DNA sequencing and histology cores.
Authorship ContributionsParticipated in research design: Hori, Negishi.Conducted experiments: Hori, Moore.Performed data analysis: Hori.Wrote or contributed to the writing of the manuscript: Hori, Negishi.
References
Alepuz PM, de Nadal E, Zapater M, Ammerer G, and Posas F (2003) Osmostress-induced tran-scription by Hot1 depends on a Hog1-mediated recruitment of the RNA Pol II. EMBO J 22:2433–2442.
Antoon JW, Bratton MR, Guillot LM, Wadsworth S, Salvo VA, and Burow ME (2012) Inhibitionof p38-MAPK alters SRC coactivation and estrogen receptor phosphorylation. Cancer Biol Ther13:1026–1033.
Benet M, Lahoz A, Guzmán C, Castell JV, and Jover R (2010) CCAAT/enhancer-binding proteinalpha (C/EBPalpha) and hepatocyte nuclear factor 4alpha (HNF4alpha) synergistically cooperatewith constitutive androstane receptor to transactivate the human cytochrome P450 2B6(CYP2B6) gene: application to the development of a metabolically competent human hepatic cellmodel. J Biol Chem 285:28457–28471.
Fernandez-Marcos PJ and Auwerx J (2011) Regulation of PGC-1a, a nodal regulator of mito-chondrial biogenesis. Am J Clin Nutr 93:884S–890.
Ferreiro I, Barragan M, Gubern A, Ballestar E, Joaquin M, and Posas F (2010) The p38 SAPK isrecruited to chromatin via its interaction with transcription factors. J Biol Chem 285:31819–31828.
Freshney NW, Rawlinson L, Guesdon F, Jones E, Cowley S, Hsuan J, and Saklatvala J (1994)Interleukin-1 activates a novel protein kinase cascade that results in the phosphorylation ofHsp27. Cell 78:1039–1049.
Gineste R, Sirvent A, Paumelle R, Helleboid S, Aquilina A, Darteil R, Hum DW, Fruchart JC,and Staels B (2008) Phosphorylation of farnesoid X receptor by protein kinase C promotes itstranscriptional activity. Mol Endocrinol 22:2433–2447.
Gotoh S and Negishi M (2015) Statin-activated nuclear receptor PXR promotes SGK2 de-phosphorylation by scaffolding PP2C to induce hepatic gluconeogenesis. Sci Rep 5:14076.
Guo H, Gao C, Mi Z, Zhang J, and Kuo PC (2007) Characterization of the PC4 binding domain andits interactions with HNF4alpha. J Biochem 141:635–640.
Han J, Lee JD, Bibbs L, and Ulevitch RJ (1994) A MAP kinase targeted by endotoxin andhyperosmolarity in mammalian cells. Science 265:808–811.
Hong YH, Varanasi US, Yang W, and Leff T (2003) AMP-activated protein kinase regulatesHNF4alpha transcriptional activity by inhibiting dimer formation and decreasing protein sta-bility. J Biol Chem 278:27495–27501.
Honkakoski P, Moore R, Gynther J, and Negishi M (1996) Characterization of phenobarbital-induciblemouse Cyp2b10 gene transcription in primary hepatocytes. J Biol Chem 271:9746–9753.
Honkakoski P, Zelko I, Sueyoshi T, and Negishi M (1998) The nuclear orphan receptor CAR-retinoid X receptor heterodimer activates the phenobarbital-responsive enhancer module of theCYP2B gene. Mol Cell Biol 18:5652–5658.
Hsieh JC, Jurutka PW, Nakajima S, Galligan MA, Haussler CA, Shimizu Y, Shimizu N, WhitfieldGK, and Haussler MR (1993) Phosphorylation of the human vitamin D receptor by proteinkinase C. Biochemical and functional evaluation of the serine 51 recognition site. J Biol Chem268:15118–15126.
Huot J, Houle F, Marceau F, and Landry J (1997) Oxidative stress-induced actin reorganizationmediated by the p38 mitogen-activated protein kinase/heat shock protein 27 pathway in vascularendothelial cells. Circ Res 80:383–392.
Inoue K and Negishi M (2008) Nuclear receptor CAR requires early growth response 1 to activatethe human cytochrome P450 2B6 gene. J Biol Chem 283:10425–10432.
Inoue K and Negishi M (2009) Early growth response 1 loops the CYP2B6 promoter for syner-gistic activation by the distal and proximal nuclear receptors CAR and HNF4alpha. FEBS Lett583:2126–2130.
Knutti D, Kressler D, and Kralli A (2001) Regulation of the transcriptional coactivator PGC-1 viaMAPK-sensitive interaction with a repressor. Proc Natl Acad Sci USA 98:9713–9718.
Kobayashi K, Hashimoto M, Honkakoski P, and Negishi M (2015) Regulation of gene expressionby CAR: an update. Arch Toxicol 89:1045–1055.
Konno Y, Kamino H, Moore R, Lih F, Tomer KB, Zeldin DC, Goldstein JA, and Negishi M (2010)The nuclear receptors constitutive active/androstane receptor and pregnane x receptor activate theCyp2c55 gene in mouse liver. Drug Metab Dispos 38:1177–1182.
Li L, Chen T, Stanton JD, Sueyoshi T, Negishi M, and Wang H (2008) The peripheral benzodi-azepine receptor ligand 1-(2-chlorophenyl-methylpropyl)-3-isoquinoline-carboxamide is a novelantagonist of human constitutive androstane receptor. Mol Pharmacol 74:443–453.
Lim CP, Jain N, and Cao X (1998) Stress-induced immediate-early gene, egr-1, involves activationof p38/JNK1. Oncogene 16:2915–2926.
Mutoh S, Osabe M, Inoue K, Moore R, Pedersen L, Perera L, Rebolloso Y, Sueyoshi T,and Negishi M (2009) Dephosphorylation of threonine 38 is required for nuclear translocationand activation of human xenobiotic receptor CAR (NR1I3). J Biol Chem 284:34785–34792.
Mutoh S, Sobhany M, Moore R, Perera L, Pedersen L, Sueyoshi T, and Negishi M (2013)Phenobarbital indirectly activates the constitutive active androstane receptor (CAR) by inhibitionof epidermal growth factor receptor signaling. Sci Signal 6:ra31.
Osabe M and Negishi M (2011) Active ERK1/2 protein interacts with the phosphorylated nuclearconstitutive active/androstane receptor (CAR; NR1I3), repressing dephosphorylation and se-questering CAR in the cytoplasm. J Biol Chem 286:35763–35769.
Picard N, Charbonneau C, Sanchez M, Licznar A, Busson M, Lazennec G, and Tremblay A (2008)Phosphorylation of activation function-1 regulates proteasome-dependent nuclear mobility andE6-associated protein ubiquitin ligase recruitment to the estrogen receptor beta. Mol Endocrinol22:317–330.
Qiao L, MacDougald OA, and Shao J (2006) CCAAT/enhancer-binding protein alpha mediatesinduction of hepatic phosphoenolpyruvate carboxykinase by p38 mitogen-activated protein ki-nase. J Biol Chem 281:24390–24397.
Rouse J, Cohen P, Trigon S, Morange M, Alonso-Llamazares A, Zamanillo D, Hunt T,and Nebreda AR (1994) A novel kinase cascade triggered by stress and heat shock that stim-ulates MAPKAP kinase-2 and phosphorylation of the small heat shock proteins. Cell 78:1027–1037.
Saito K, Moore R, and Negishi M (2013a) Nuclear receptor CAR specifically activates the two-pore K+ channel Kcnk1 gene in male mouse livers, which attenuates phenobarbital-inducedhepatic hyperplasia. Toxicol Sci 132:151–161.
Saito K, Moore R, and Negishi M (2013b) p38 Mitogen-activated protein kinase regulates nuclearreceptor CAR that activates the CYP2B6 gene. Drug Metab Dispos 41:1170–1173.
Shao D and Lazar MA (1999) Modulating nuclear receptor function: may the phos be with you. JClin Invest 103:1617–1618.
Shindo S, Moore R, Flake G, and Negishi M (2013) Serine 216 phosphorylation of estrogenreceptor a in neutrophils: migration and infiltration into the mouse uterus. PLoS One 8:e84462.
Shiraki T, Sakai N, Kanaya E, and Jingami H (2003) Activation of orphan nuclear constitutiveandrostane receptor requires subnuclear targeting by peroxisome proliferator-activated receptorgamma coactivator-1 alpha. A possible link between xenobiotic response and nutritional state. JBiol Chem 278:11344–11350.
Sun K, Montana V, Chellappa K, Brelivet Y, Moras D, Maeda Y, Parpura V, Paschal BM,and Sladek FM (2007) Phosphorylation of a conserved serine in the deoxyribonucleic acidbinding domain of nuclear receptors alters intracellular localization. Mol Endocrinol 21:1297–1311.
Tremblay A, Tremblay GB, Labrie F, and Giguère V (1999) Ligand-independent recruitment ofSRC-1 to estrogen receptor beta through phosphorylation of activation function AF-1. Mol Cell3:513–519.
Underwood DC, Osborn RR, Kotzer CJ, Adams JL, Lee JC, Webb EF, Carpenter DC, BochnowiczS, Thomas HC, and Hay DW, et al. (2000) SB 239063, a potent p38 MAP kinase inhibitor,reduces inflammatory cytokine production, airways eosinophil infiltration, and persistence. JPharmacol Exp Ther 293:281–288.
Yamamoto Y, Moore R, Goldsworthy TL, Negishi M, and Maronpot RR (2004) The orphannuclear receptor constitutive active/androstane receptor is essential for liver tumor promotion byphenobarbital in mice. Cancer Res 64:7197–7200.
Zhang L and Jope RS (1999) Oxidative stress differentially modulates phosphorylation of ERK,p38 and CREB induced by NGF or EGF in PC12 cells. Neurobiol Aging 20:271–278.
Zhou W and Slingerland JM (2014) Links between oestrogen receptor activation and proteolysis:relevance to hormone-regulated cancer therapy. Nat Rev Cancer 14:26–38.
Address correspondence to: Dr. Masahiko Negishi, National Institute of Environ-mental Health Sciences, National Institutes of Health, Research Triangle Park, NorthCarolina, 27709. E-mail: [email protected]
876 Hori et al.
at ASPE
T Journals on June 18, 2018
dmd.aspetjournals.org
Dow
nloaded from





























