Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, Nov. 2011, p. 6244–6256 Vol. 193, No. 220021-9193/11/$12.00 doi:10.1128/JB.00175-11Copyright © 2011, American Society for Microbiology. All Rights Reserved.
Localization Pattern of Conjugation Machinery in aGram-Positive Bacterium�†
Theresa Bauer,1 Thomas Rosch,1,2 Mitsuhiro Itaya,3 and Peter L. Graumann1,2*Mikrobiologie, Fachbereich fur Biologie, Universitat Freiburg, Schanzle Strasse 1, 79104 Freiburg, Germany1; Spemann Graduate School of
Biology and Medicine, Albert Ludwigs University Freiburg, Freiburg, Germany2; and Laboratory of Genome Designing Biology,Institute for Advanced Biosciences, Keio University, 403-1 Nipponkoku, Daihoji, Tsuruoka, Yamagata 997-0017, Japan3
Received 8 February 2011/Accepted 6 September 2011
Conjugation is an efficient way for transfer of genetic information between bacteria, even between highlydiverged species, and a major cause for the spreading of resistance genes. We have investigated the subcellularlocalization of several conserved conjugation proteins carried on plasmid pLS20 found in Bacillus subtilis. Weshow that VirB1, VirB4, VirB11, VirD2, and VirD4 homologs assemble at a single cell pole, but also at othersites along the cell membrane, in cells during the lag phase of growth. Bimolecular fluorescence complemen-tation analyses showed that VirB4 and VirD4 interact at the cell pole and, less frequently, at other sites alongthe membrane. VirB1 and VirB11 also colocalized at the cell pole. Total internal reflection fluorescencemicroscopy showed that pLS20 is largely membrane associated and is frequently found at the cell pole,indicating that transfer takes place at the pole, which is a preferred site for the assembly of the activeconjugation apparatus, but not the sole site. VirD2, VirB4, and VirD4 started to localize to the pole or themembrane in stationary-phase cells, and VirB1 and VirB11 were observed as foci in cells resuspended in freshmedium but no longer in cells that had entered exponential growth, although at least VirB4 was still expressed.These data reveal an unusual assembly/disassembly timing for the pLS20 conjugation machinery and suggestthat specific localization of conjugation proteins in lag-phase cells and delocalization during growth are thereasons why pLS20 conjugation occurs only during early exponential phase.
Conjugation was one of the first tools to be used to revealthe nature of genetic material, and later it was used to gaininsight into molecular genetics. Although conjugation has beenstudied for several decades, many key questions about themechanism of DNA transfer between cells remain to be solved.Moreover, much less is known about the conjugation machin-ery and mode of conjugation in Gram-positive bacteria (12, 21,27), a large bacterial phylum containing important humanpathogens. The aim of this study was to shed light on themolecular mechanism of conjugation in the Gram-positivemodel bacterium Bacillus subtilis.
Several proteins have been shown to be essential or impor-tant for conjugation in Gram-negative systems, which togetherare thought to set up a type IV secretion system that cantranslocate proteins as well as DNA (7). Some of the charac-terized systems are the Escherichia coli F plasmid (IncF) Trasystem, E. coli RP4 (IncP) Trb system, E. coli R388 (IncW)Trw system, Shigella Collb-P9 (IncI) Tra system, Agrobacteriumtumefaciens VirB system, and Legionella pneumophila Dot/Icmsystem (8, 11). Conjugation systems are also found on integra-tive conjugative elements (ICEBs1 from Bacillus subtilis). Inthis work, we have used the Vir nomenclature, because it iswidely used and refers to the best-studied system in A. tume-faciens, which can transfer a copy of DNA from the transfer
region of the tumor-inducing (Ti) plasmid as single-strandedDNA (ssDNA) into plant cells; these then become trans-formed. There are several conserved (and thus centrally im-portant) proteins: VirD2 is a so-called relaxase. It introduces anick in the T-DNA border sequence and remains covalentlybound at the 5� end of the DNA transfer intermediate (15, 17,18). After guiding the ssDNA to the coupling protein VirD4,VirD2 is cotransferred into the recipient cell (10). In all knowncases (except for systems in Streptomyces), an ssDNA/proteincomplex is translocated into the other cell. There are threecomponents containing ATPase motifs, VirB11, VirB4, andVirD4. VirB11 and VirD4 contain Walker A and B motifs(ATP binding site) and belong to the superfamily of P-loopATPases (11). VirB11 in A. tumefaciens is an ATPase thatforms homomultimers and possibly provides energy for DNAtransfer and pilus biogenesis. Dihybrid screens have showninteractions with VirB4 and with VirB9 (8, 38). For Helicobac-ter pylori HP0525, a homolog of VirB11, the crystal structurewas solved and it showed a hexameric assembly of the struc-ture. The pore, which is formed by the multimeric protein inthe ADP-bound form, has an external diameter of 100 Å andan internal diameter of 50 Å. It has been proposed that nu-cleotide binding and hydrolysis lead to conformational changesto facilitate substrate export (33).
VirD4 from A. tumefaciens is called a coupling protein (CP).The proposed functions are the recruitment of the ssDNA andprotein substrate to the conjugation machinery and their trans-location. In dihybrid and biochemical assays, a close contactwith VirE2 (single-strand binding protein [SSB]) was detected(5). CPs of Gram-negative bacteria are known to have 2 amino-terminal transmembrane helices, a small periplasmic domain,
* Corresponding author. Mailing address: Mikrobiologie, Fachbereichfur Biologie, Universitat Freiburg, Schanzle Strasse 1, 79104 Freigburg,Germany. Phone: 49 (0) 7612032630. Fax: 49 (0) 7612032773. E-mail:[email protected].
† Supplemental material for this article may be found at http://jb.asm.org/.
� Published ahead of print on 23 September 2011.
6244
on August 25, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

and a large carboxy-terminal region in the cytoplasm. TheX-ray crystal structure of the soluble C-terminal part of the E.coli VirD4 homolog TrwB from plasmid R388 shows a ring-likestructure, similar to F1 ATPase, with a channel diameter of 20Å (20). Purified VirD4 was detected in the soluble as well as inthe membrane fractions, while exclusively protein from thesoluble fraction showed ATPase activity. It was proposed thatVirD4 has a translocase function, which is supported by thefact that it bears sequence homologies to DNA translocases,like SpoIIIE and FtsK. The mechanism of this process is un-known, although there are hints that interactions occur withparts of the mating pair formation (Mpf) complex (19, 30), andso it has been suggested that VirD4 recruits the transfer sub-strate and delivers the DNA/protein complex to the conjuga-tion apparatus (4, 32, 34, 35).
VirB4 has homologies to the P-loop ATPase HerA. TheVirB4 protein is postulated to energize the substrate export byATP-driven conformational changes. It is essential for DNAexport and seems to interact with the second Mpf-ATPase,VirB11 (4, 38). In A. tumefaciens, VirB11 interacted withVirB4, VirB8, and VirB10 and with itself (4, 11, 38). LikeVirB11, VirB4 is postulated to provide energy for pilus bio-genesis.
VirB1 belongs to the periplasmic factors (11) and contains alytic transglycosylase domain (LT domain; belonging to thelysozyme superfamily). LTs catalyze the cleavage of the beta-1,4-glycosidic bond between N-acetylmuramic acid (MurNAc)and N-acetyl-D-glucosamine (GlcNAc). The proposed func-tions are peptidoglycan hydrolase activity and pilus/channelassembly. Dihybrid screens showed interactions with VirB1,VirB4, VirB8, VirB9, VirB10, and VirB11 (38).
We have chosen to study pLS20 as a model for conjugativeplasmids in Gram-positive bacteria. pLS20 is a plasmid discov-ered in a natural Bacillus subtilis isolate (natto) (36) and ispartitioned through a ParM-like active mechanism to daughtercells during each cell cycle, but it can also propagate via con-jugation. Conjugation was shown to be independent of RecAprotein and was most efficient during the early phase of loga-rithmic growth (22), in contrast to the acquisition of DNAthrough natural competence in B. subtilis, which is restricted tothe entry into stationary-phase growth (9). pLS20 is 65 kb insize and can also mobilize nonconjugative plasmids, and it hasbeen studied because it can conjugate into other Bacillusstrains and can potentially be used for the transfer of largeDNA fragments. Replication of pLS20 occurs via a novelmechanism: the replication region shows no similarity withother known plasmid replicons (31) and therefore has beensuggested to belong to a new class of theta replicons, estab-lishing an average of 1 to 3 copies per cell (26, 29). Its segre-gation employs actin-like protein Alp7a, which appears to pushplasmids toward opposite cell poles via the formation of highlydynamic filaments (16). Although a miniversion of pLS20 hasbeen used to visualize the segregation pattern, nothing isknown about the localization of the full-length pLS20 plasmidor the localization of parts of its conjugation machinery. Weshow that full-length pLS20 behaves differently from the mini-plasmid, and its localization pattern appears to be a mixture ofthat of bipolarly positioned low-copy-number plasmids and ofan additional extremely polarly located plasmid copy (or cop-ies). We provide evidence that the conjugation machinery as-
sembles at a single cell pole or at a defined site along thelateral cell membrane. Most interestingly, we found that theconjugation machinery assembles in cells during extended sta-tionary phase and during lag phase but disassembles as cellscommence exponential growth, in correlation with the transferactivity of the plasmid.
MATERIALS AND METHODS
Bacterial strains and media. Bacillus strains (see Table S2 in the supplementalmaterial) were grown in LB medium at 37°C for conjugation assays and at 30°Cfor microscopy. Selection pressure for the inserted fusions was always maintainedwith appropriate antibiotics. Because of the high stability of pLS20 (22), antibi-otic was never added for maintenance of the plasmid. The fusion of VirD4 andcyan fluorescent protein (CFP), expressed from the chromosome (strains TCR3and TB15), was induced with 0.01 mM isopropyl-�-D-thiogalactopyranoside(IPTG), and the inducible fusion of VirB4 and yellow fluorescent protein (YFP;TCR04) was grown in LB medium supplemented with 0.5% xylose. For micros-copy, cells were grown until stationary phase for 10 h and were resuspended intofresh LB medium (time point, 0 h).
Conjugation assays. Mating experiments were performed as described before(22). To compare the number of trancipients in a single growth experiment, thenumber of trancipients at each time point was calculated. These numbers oftrancipients were normalized to equal cell numbers at each time point of theexperiment. Given that numbers of trancipients are relative numbers, they can beused to compare different growth stages of one strain. To determine the exacttransfer rate, a dilution series (10�5, 10�6, and 10�7) of samples, taken formating experiments at different growth stages, was plated on LB agar to deter-mine the definite number of cells during a conjugation experiment. Differentamounts of mating mixtures were spread on plates supplemented with antibioticsthat selected for the according trancipient.
Constructions of plasmids and strains. To create C-terminal green fluorescentprotein (GFP), YFP, CFP, mCherry or TAP Tag fusions of VirB11 (ORF 13),VirD4 (ORF 24), VirB4 (ORF 28), VirB1 (ORF 30), and VirD2 (ORF 34) forsingle-crossover integration into the plasmid, the 3� regions of the genes wereamplified by PCR using primers 1 to 10 (see Table S1 in the supplementalmaterial). PCR products of the C-terminal regions of virB11, virD4, virB4, andvirD2 were cloned with the indicated enzymes into the multiple cloning site (mcs)of modified pSG1164y (25), generating pTB01, pTB02 and pTB03, pTB05, TB13,and TB14 (see Table S2 in the supplemental material). The plasmid expressingan mCherry C-terminal fusion to virB1 was constructed by exchanging dnaX withvirB1 in pCT2 (14), using EcoRI and XhoI restriction sites and generating pTB04(virB1-mCherry).
pLS20neo was a kind gift from and constructed by Shinya Kaneko (TokyoInstitute of Technology, Yokohama-shi Kanagawa, Japan) by replacing the catgene of pLS20cat (22) with a neo gene by using in vitro gene assembly technology(unpublished results). To introduce the lacO repeat into pLS20neo, a 1-kb regionof pLS20neo (bp 61101 to 62100) was amplified by PCR, using primers 11 and 12(see Table S1 in the supplemental material), and cloned into the HindIII re-striction site of pAT12 (39), resulting in pTB06.
To generate the IPTG-inducible thrC::Physpank::virD4-cerulean cfp constructpTCR3, virD4 was first subcloned in pFD2, encoding the fluorescent proteincerulean-CFP. virD4 was amplified by PCR using primer pair 14/17 andpLS20neo as template DNA, digested with KpnI and XhoI, and cloned intopFD2. In a second PCR, virD4 was amplified together with GFP, using pTCR2as template DNA and primer pair 18/19, digested with NheI and SphI and clonedinto NheI and SphI sites of pDP150, creating plasmid pTCR3. To generate thexylose-inducible amyE::Pxyl::virB4-yfp construct pTCR4, virB4 was amplified byPCR using primer pair 20/21 and pLS20neo as template DNA, digested withKpnI and XhoI, and cloned into KpnI- and XhoI-digested pSG1193.
To generate PY79 containing pLS20 (AKR04), PY79 was transformed withpLS20neo and transformants were selected with kanamycin. AKR04 was trans-formed with the plasmids pTB01, pTB02 pTB03, pTB04, pTB05, pTB06, pTB10,and pTB14, and transformants were selected with the appropriate antibiotics togenerate strains TB01, TB02, TB03, TB04, TB05, TB06, TB10, and TB14. Tocreate a strain expressing a LacI-CFP fusion (AKR05), PY79 was transformedwith chromosomal DNA of PG26. Transformants were selected with lincomycinand erythromycin.
pLS20neo with the inserted lacO repeat from strain TB06 was conjugated asdescribed previously (22) into strain AKR05 (lacI-cfp), as recipient, resulting instrain TB07. Strains TB08 and TB09 were created by conjugation with strainCDS4 (dnaX-mCherry) (14). Trancipients of TB07 were selected with chloram-
VOL. 193, 2011 LOCALIZATION OF CONJUGATION MACHINERY IN B. SUBTILIS 6245
on August 25, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

phenicol, lincomycin, and erythromycin, and trancipients of TB08 and TB09were selected by spectinomycin and chloramphenicol. Strain TB12 (pLS-cfpdnaX-mCherry) was generated by transformation of chromosomal DNA fromstrain CDS4 (dnaX-mCherry) in TB07.
For the bimolecular fluorescence complementation (BiFC) experiment, gfpfrom construct 1164 virB4gfp was replaced with the C-terminal part of YFP (Yc)by using primer pair 15/16 and PstI and SpeI restriction sites, resulting in pTB10.In pSW02 (dnaX-yn; kind gift from S. Welling), dnaX was exchanged with virD4by using the EcoRI and XhoI restriction sites (primers 13 and 14), resulting inpTB11. PY79/pLS20neo was transformed with pTB10, and transformants ofTB10 were selected by chloramphenicol resistance. TB10 was transformed withpTB11, and transformants were selected with spectinomycin and chloramphen-icol, resulting in TB11. TCR1 and TCR2 were created by conjugation of TB10with SW02 and JS96 (13), and trancipients were selected with spectinomycin andchloramphenicol. TB15 was created by conjugation of TCR03 and TB14, andtrancipients were selected by chloramphenicol, lincomycin, and erythromycin.For the simultaneous visualization of VirB1 and VirB11 in the same cell, TB01(virB11-yfp) was transformed with pTB04 and transformants were selected withspectinomycin and chloramphenicol, resulting in strain TB16.
To generate a C-terminally truncated fusion of VirD2 and GFP, 500 bp lying120 bp upstream of the virD2 stop codon was PCR amplified with primer pair24/25 and pLS20neo as template, digested with ApaI and ClaI, and cloned intoApaI- and ClaI-digested pSG1164.
FISH. To localize double-stranded pLS20 and ssDNA intermediates producedduring the process of conjugation in situ, a denaturing and nondenaturing genefluorescence in situ hybridization (FISH) assay was performed as described inreference 3, using the 5� region of the virD2 gene as a probe. Details andvariations are provided in the supplemental material.
Image acquisition. Fluorescence microscopy was performed with an OlympusAX70 microscope and an AXIO Observer A1 (Zeiss) or an AXIO Imager(Zeiss) with a Cool Snap ES2 camera (Photometrics), using total internal reflec-tion fluorescence (TIRF) objectives with a numerical aperture of 1.45. Cells weregrown in LB medium and mounted on thin agarose pads (1% prepared in S750
minimal medium supplemented with 0.1% Casamino Acids) on object slides.Membranes were stained with FM4-64 (2.5 �g/ml). Images were processed byusing Metamorph 6.3 and ImageJ 1.43 software (Image Processing and Analysis).FRAP experiments were performed on a Zeiss Axio Observer Z with a CascadeII 518 camera (Photometrics) and a 405-nm laser beam.
Western blot analysis. For Western blot analysis of TAP-tagged VirB4, cul-tures were grown in liquid media with the appropriate antibiotics. Equal numbersof cells were harvested at each time point during exponential growth. Blottingwith specific antisera was performed using a semidry procedure. Antibodiesagainst the CBP-epitope of the TAP tag were used to detect the fusion protein,and anti-MreB antiserum was used as a control.
RESULTS
Genes present on pLS20. pLS20 is a 65-kb plasmid that wasisolated by Tanaka et al. (36) from B. subtilis (natto) IFO3335and was characterized as a natural fertility plasmid. Growthstage-dependent kinetics of conjugational transfer frequenciesin liquid compared to solid media were determined. pLS20transfers poorly on solid media, or in minimal media, butshows maximal transfer kinetics of (3.3 � 0.78) � 10�3 tran-cipients/donor/15 min during the transition from fresh inocu-lation of the culture to exponential growth. We have system-atically analyzed all open reading frames (ORFs) present onthe plasmid, with special interest for potential conjugationproteins. A cartoon of pLS20 is shown in Fig. S1 of the sup-plemental material. We found that pLS20 carries genes for allconserved ATPase motif proteins: orf13, virB11; orf24, virD4;orf28, virB4. A BLAST search showed strongest homology ofVirD4 to the TraG/TraD system from Clostridium perfringens.VirD4 is predicted to have 2 N-terminal transmembrane do-mains (based on SMART and EMBL databases) or even 4transmembrane domains (HMMTOP database). For VirB4,BLAST searches showed highest homologies to a predictedTraC-like protein from Enterococcus faecalis T1 and a VirB4
component from Lactobacillus casei. orf30 encodes a VirB1ortholog with highest homologies to putative lipoproteins fromListeria spp. or Bacillus thuringiensis, as well as significant ho-mologies to the secreted cell wall DL-endopeptidase Yddh/CwIT from B. subtilis. VirB1 contains one predicted transmem-brane helix at the N terminus (HMMTOP, residues 24 to 48;SMART, 25 to 47). Catalytic residues of the N-acetyl-D-glu-cosamine binding site of LTs are conserved. The C-terminalregion of the protein contains conserved domains with homol-ogies to cell wall-associated hydrolyases and invasion-associ-ated secreted endopeptidases and bacteriophage-like ami-dases. VirB1 thus appears to be able to confer glycosidase aswell as peptidase activity. orf34 has similarity to relaxases andis thus likely to encode a VirD2 ortholog. It contains no ap-parent transmembrane domains and is likely a soluble protein.More proteins found on pLS20 are described in the supple-mental material.
Localization pattern of pLS20 in live bacterial cells. Wewanted to visualize a conjugative plasmid from a Gram-posi-tive organism and proteins involved in conjugation to gainfurther insights into this process. The group of J. Pogliano hascharacterized an actin-like partitioning system on pLS20 andhas used a minimal variant of the plasmid to visualize itslocalization (16). We wanted to visualize the full-length plas-mid, which may behave differently from its minimal version,and to colocalize it with the DNA replication machinery todetermine its place of replication. To do this, we used the lacOarray (integrated close to the replication region in pLS20)/LacI-CFP system, which yielded fully conjugation-proficientpLS20. Figure 1A shows that exponentially growing cells con-tain 3 to 4 copies (up to 5) of pLS20, which agrees well with themeasured copy number of 3 (26). In exponential growth phase,all cells contained several pLS20 foci. Of these, 30% of thecells showed a focus at one pole and 3% at both poles (Table1). These data suggest that pLS20 plasmids do not undergocohesion but are rapidly distributed within the cell. In station-ary-phase cells, only one or two copies are detectable (Fig. 1C),showing that the copy number is growth phase-dependent anddrops as cells exit exponential growth. Of stationary-phasecells, 42% showed a focus at one pole, 46% showed foci at bothpoles, and 12% contained one or two foci at nonpolar positionsat the membrane; only 27% of all cells contained additionalfoci between the poles. Thus, pLS20 is predominantly found atthe cell pole, but also at additional membrane-proximal sitesaway from a pole, in stationary-phase cells and in cells duringlag phase. These data suggest that the cell pole plays a domi-nant role for pLS20 during early growth or growth arrest.Additionally, most pLS20 copies appear to localize at the cellmembrane. To verify this, we used TIRF microscopy, whichvisualizes fluorescence that is very close to the cell surface.Figure 1E shows that a large number of plasmids were stillseen in the TIRF mode, in which some signals were lost thatwere not close to the membrane plane imaged.
We also followed the localization of pLS20 in sporulatingcells. B. subtilis spores all contained several pLS20 copies (Fig.1D), showing that pLS20 is inherited through sporulation withhigh efficiency.
To obtain information on the mode of replication of theconjugative plasmid, we colocalized pLS20 and DnaX, a cen-tral component of the replication machinery. We found over-
6246 BAUER ET AL. J. BACTERIOL.
on August 25, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
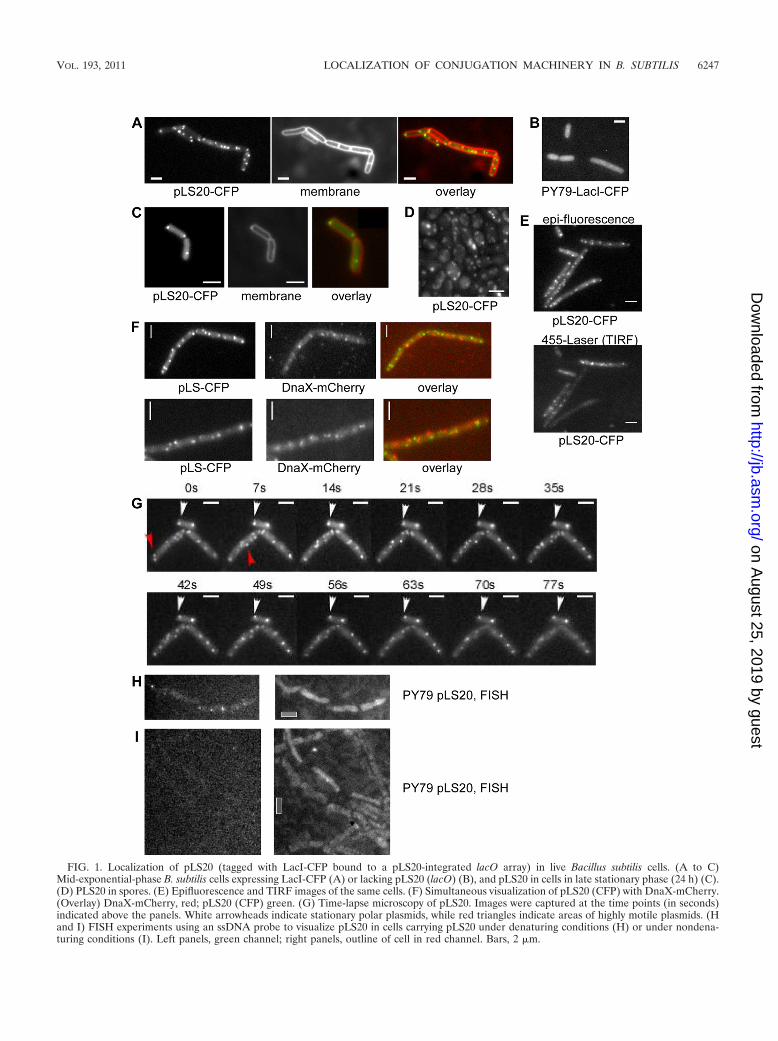
FIG. 1. Localization of pLS20 (tagged with LacI-CFP bound to a pLS20-integrated lacO array) in live Bacillus subtilis cells. (A to C)Mid-exponential-phase B. subtilis cells expressing LacI-CFP (A) or lacking pLS20 (lacO) (B), and pLS20 in cells in late stationary phase (24 h) (C).(D) PLS20 in spores. (E) Epifluorescence and TIRF images of the same cells. (F) Simultaneous visualization of pLS20 (CFP) with DnaX-mCherry.(Overlay) DnaX-mCherry, red; pLS20 (CFP) green. (G) Time-lapse microscopy of pLS20. Images were captured at the time points (in seconds)indicated above the panels. White arrowheads indicate stationary polar plasmids, while red triangles indicate areas of highly motile plasmids. (Hand I) FISH experiments using an ssDNA probe to visualize pLS20 in cells carrying pLS20 under denaturing conditions (H) or under nondena-turing conditions (I). Left panels, green channel; right panels, outline of cell in red channel. Bars, 2 �m.
VOL. 193, 2011 LOCALIZATION OF CONJUGATION MACHINERY IN B. SUBTILIS 6247
on August 25, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
lapping pLS20-CFP and DnaX-mCherry signals in only 2% ofthe cells, while 98% of the signals were nonoverlapping (Fig.1F). Note that most experiments were performed with rich(LB) medium at 30°C (because cells carrying pLS20 do notgrow and conjugate well in minimal medium [data not shown]),where cells have overlapping rounds of chromosomal replica-tion and therefore multiple replication machineries. Thesedata indicate that pLS20 may not employ the chromosomalreplication machinery for its own propagation, in agreementwith its unique replication mechanism.
Time-lapse microscopy revealed two different populations ofpLS20. Polar pLS20-CFP foci were generally static and did notmove during the entire experiment, while nonpolar plasmidswere generally mobile and moved between the 7-s time inter-vals (Fig. 1G; also see Movie S1 in the supplemental material).These data suggest that polar pLS20 is stably anchored to themembrane, while internal plasmids are free to diffuse within acertain area of the cell.
Nondenaturing FISH has revealed which of the two DNAstrands is translocated by the A. tumefaciens conjugation ma-chinery (3). We also performed FISH experiments on pLS20,which verified that under denaturing conditions, the plasmid isfound at the cell poles and at the lateral membrane (Fig. 1H).However, we were not able to detect a specific signal undernondenaturing conditions (Fig. 1I), showing that ssDNA frompLS20 transferred during conjugation is less accessible forFISH than in the Gram-negative system.
VirB4 and VirD4 localize as discrete foci at the cell mem-brane and to the cell pole during late stationary phase andduring lag phase. We generated fluorescent protein fusions toseveral conserved conjugation proteins encoded on pLS20. Fu-sions were integrated via single-crossover recombination intopLS20. To test if the fusions were functional, transformantswere used as donors to transfer pLS20 into a new recipient.Because only a single copy was transferred, the resultantconjugants contained solely recombinant pLS20 copies. Con-jugation efficiencies of resultant strains were very similar towild-type levels (Table 2), showing that all protein fusionsgenerated are fully functional, and they were expressed as solesources of the protein within cells.
For VirB4, we found homogeneous fluorescence in �95% ofall cells during mid-exponential growth (Fig. 2A), showing thatthe protein is expressed. This was verified by Western blotanalysis (Fig. 3D). Note that during exponential growth, B.subtilis PY79 cells form almost exclusively chains (Fig. 2A, graytriangle), and few single cells (swimmers, 5 to 10% of all cells[white triangles]). During mid-exponential growth, few singlecells (less than 1% of all cells) were observed that containedsingle VirB4-GFP foci, and these were generally at the cell
membrane (Fig. 2B; note that in this image small cells arehighly overrepresented). During late exponential phase, nomore cells containing VirB4-GFP foci were detectable, al-though single cells were still present (data not shown) and,likewise, only homogeneous fluorescence was observed in cellsduring early stationary phase (10 h after inoculation in freshmedium) (Fig. 2C). To address the question, whether singlecells containing VirB4-GFP foci during mid-log-phase growthmay not yet have commenced an active cell cycle, we moni-tored the localization of VirB4-GFP in cells expressing DnaX-mCherry, a central component of the replication machinery.Small cells that displayed VirB4-GFP very rarely containedDnaX-mCherry foci (however, due to a reason unclear to us,many [about 60%] of the cells had high mCherry backgroundfluorescence), while all cell chains contained DnaX-mCherryfoci but no VirB4-GFP foci (Fig. 2D). Colocalization of dis-crete DnaX-mCherry foci and VirB4-GFP was only observedin less than 1% of all cells analyzed (Fig. 2D). Thus, cells thatare actively replicating their chromosomes and are thereforecommencing exponential growth do not contain VirB4-GFPassemblies, while cells that probably have not yet initiatedreplication frequently do contain assemblies. These findingsraised the possibility that B. subtilis cells assemble VirB4 mol-ecules at the cell membrane as they transition from the sta-tionary phase to growth (lag phase) and disassemble them asthey induce an active cell cycle. We therefore monitoredVirB4-GFP fluorescence in cells freshly resuspended in richmedium (note that in minimal medium, we observed extremelypoor conjugation activity and few cells containing VirB4-YFPfoci). Strikingly, 20 to 30% of the cells contained a single focusor, rarely, up to 2 foci. In 75% of these cases, a focus waslocated at a cell pole, while in 25% a focus was positioned at arandom position, but in 90% of all cases, the focus was clearlylocated at the membrane (Fig. 2E). Therefore, VirB4 assem-bles at the membrane, and frequently at the pole, during lagphase. Interestingly, this is the time when conjugation activityis highest, and in fact, the number of cells showing VirB4-GFPfoci correlated with the conjugation activity of the culture (Fig.3A and B). We wished to find out if VirB4 assembles at somepoint during stationary phase, or if foci are freshly induced asthe cell obtains new medium. Intriguingly, 24 h after inocula-
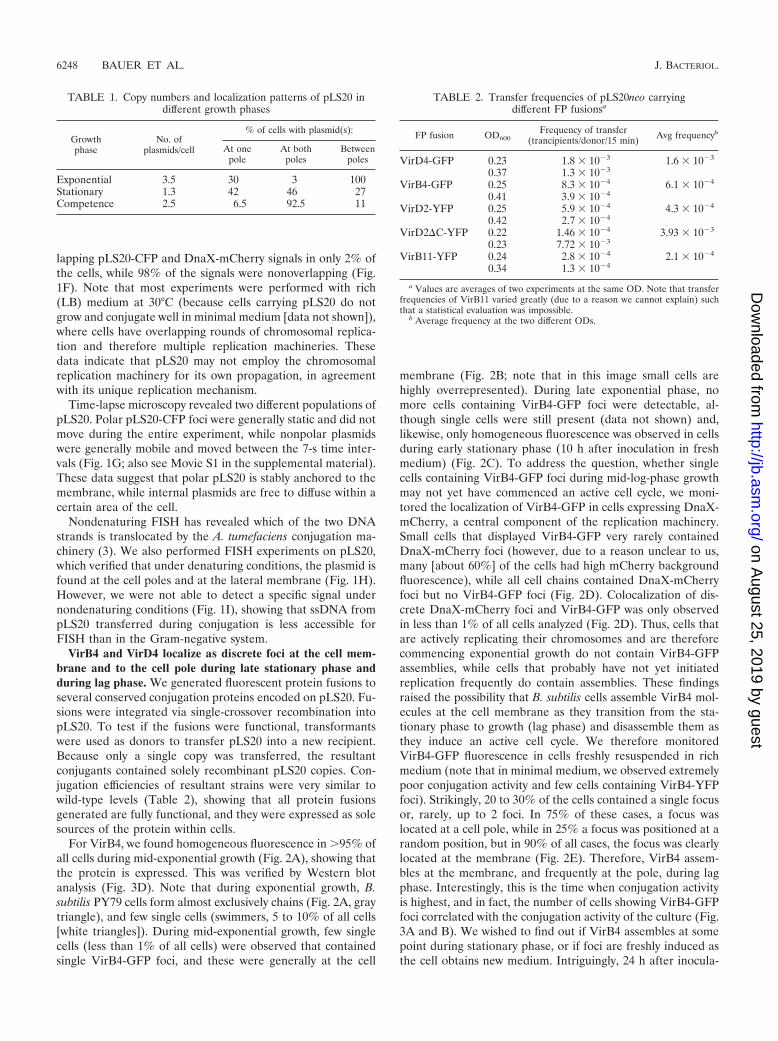
TABLE 1. Copy numbers and localization patterns of pLS20 indifferent growth phases
Growthphase
No. ofplasmids/cell
% of cells with plasmid(s):
At onepole
At bothpoles
Betweenpoles
Exponential 3.5 30 3 100Stationary 1.3 42 46 27Competence 2.5 6.5 92.5 11
TABLE 2. Transfer frequencies of pLS20neo carryingdifferent FP fusionsa
FP fusion OD600Frequency of transfer
(trancipients/donor/15 min) Avg frequencyb
VirD4-GFP 0.23 1.8 � 10�3 1.6 � 10�3
0.37 1.3 � 10�3
VirB4-GFP 0.25 8.3 � 10�4 6.1 � 10�4
0.41 3.9 � 10�4
VirD2-YFP 0.25 5.9 � 10�4 4.3 � 10�4
0.42 2.7 � 10�4
VirD2�C-YFP 0.22 1.46 � 10�4 3.93 � 10�3
0.23 7.72 � 10�3
VirB11-YFP 0.24 2.8 � 10�4 2.1 � 10�4
0.34 1.3 � 10�4
a Values are averages of two experiments at the same OD. Note that transferfrequencies of VirB11 varied greatly (due to a reason we cannot explain) suchthat a statistical evaluation was impossible.
b Average frequency at the two different ODs.
6248 BAUER ET AL. J. BACTERIOL.
on August 25, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

tion of the culture (i.e., about 14 h after reaching stationaryphase), 10% of the cells contained a single polar VirB4-GFPfocus and 3% had a focus at the lateral membrane; 1% con-tained two polar foci (Fig. 2F). Thus, VirB4 also assemblesduring stationary phase in a subset of the cells, where cells are
not active for conjugation (Fig. 3A), and in additional cellswhen fresh medium is supplied, where conjugation is induced.
We wanted to investigate if the localization of VirB4 wasassociated with swimmers (motile single cells) or chain form-ers. Because during exponential phase most PY79 cells are
FIG. 2. (A to D) Localization of VirB4 and VirD4. Images show VirB4-GFP in the mid-exponential growth phase (A, B, and D), earlystationary phase (10 h) (C), or with simultaneous visualization of VirB4-GFP and DnaX-mCherry during mid-exponential growth (D). Graytriangles indicate cell chains expressing DnaX-mCherry, and white triangles indicate small cells with VirB4-GFP foci. (E and F) VirB4-GFP at lagphase (1 to 2 h after inoculation) (E) or at mid-stationary phase (24 h) (F). Polar VirB4-GFP assemblies are indicated by white triangles. (G toJ) VirD4-GFP during lag phase (G), early stationary phase (10 h) (H), or mid-stationary phase (24 h) (I), and simultaneous visualization ofVirD4-GFP and DnaX-mCherry during mid-exponential growth (J). Gray triangles indicate cell chains expressing DnaX-mCherry (faint foci), andwhite triangles indicate small cells with VirD4-GFP foci. Bars, 2 �m.
VOL. 193, 2011 LOCALIZATION OF CONJUGATION MACHINERY IN B. SUBTILIS 6249
on August 25, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

chain formers, we overproduced SwrA from the amylase locus,which results in the formation of a majority of swimmers(�90%) (24). However, even under these conditions, mostcells during lag phase showed VirB4-GFP foci, and few cellscontained foci during exponential phase (data not shown).Because the pattern of localization was not considerably dif-ferent between chain formers and swimmers, it is clearly asso-ciated with the growth phase and not with motility.
To test if several proteins from the presumed conjugationmachinery follow a similar localization pattern, we monitoredthe subcellular localization of VirD4. Indeed, VirD4-GFP wasvisible as a membrane-proximal focus in cells during lag phaseand again frequently at a single cell pole (Fig. 2G). Few cellscontaining foci were seen during mid-exponential growth (Fig.2J), and no foci were observed during early stationary phase(Fig. 2H). However, discrete foci were easily detectable after24 h (mid-stationary phase) (Fig. 2I). As for VirB4, DnaX-mCherry and VirD4-GFP showed exclusive localization andwere present in the same cell only in extremely few cases (Fig.
2J), revealing that VirD4 assembles at the membrane duringlate stationary phase and even more so during lag phase (whenearly stationary phase cells are resuspended in fresh medium,VirD4-GFP also assembles within 60 to 120 min, like VirB4-GFP), but it disassembles as cells enter exponential growth.These data suggest that the entire conjugation machinery or atleast the central part of it may assemble and disassemble at adefined subcellular site within stationary/lag-phase cells.
To test for the specificity of localization, we expressedVirB4-YFP or VirD4-CFP from ectopic loci on the chromo-some in B. subtilis cells lacking pLS20. In both cases, homoge-neous cytosolic fluorescence was observed throughout allgrowth phases (Fig. 4A and B, respectively), and no mem-brane-attached foci were detectable even in lag-phase cells(under low or high inducer concentrations). As a control, bothectopically induced fusions were induced in cells carryingpLS20. In both cases, conjugation proteins localized in lag-phase cells and no longer in exponentially growing cells (seeFig. S4 in the supplemental material). Thus, VirB4 and VirD4
FIG. 3. Transfer kinetics of pLS20 and localization relative to growth phase. (A) Conjugation frequency of pLS20 at different time points duringgrowth. OD600, optical density at 600 nm. (B) Number of cells showing VirB4-GFP foci at different time points during growth (localizationfrequency). (C) Number of cells showing VirB11-YFP foci at different time points during growth (localization frequency). (D) Western blot atdifferent time points during growth (hours after inoculation are indicated) of VirB4-TAP by using TAP tag-specific antibodies (or against MreB,loading control).
6250 BAUER ET AL. J. BACTERIOL.
on August 25, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

FIG. 4. Colocalization of VirB4 and VirD4 and their interaction. (A) VirB4-YFP expressed from the amylase locus during lag phase in theabsence of pLS20. (B) VirD4-CFP expressed from the thr locus during lag phase in the absence of pLS20. (C) Cells expressing VirB4-mCherryfrom the original gene locus on pLS20 and VirD4-CFP from the thr locus during lag phase. White arrowheads indicate colocalization of VirD4and VirB4. (D to H) BiFC experiments, with VirD4-Yn expressed during lag phase (D), VirB4-Yc and VirD4-Yn expressed during lag phase (whitelines indicate septa between cells) (E), VirB4-Yc and VirD4-Yn expressed during mid-exponential phase (F), VirB4-Yc and MreC-Yn expressedduring lag phase (G), and VirB4-Yc and DnaX-Yn expressed during lag phase (H). White arrowheads indicate small cells with a BiFC focus.(I) VirD2-CFP expressed during lag phase. (J and K) VirD2-CFP during early stationary phase (10 h after inoculation) (J) or VirD2-CFP duringearly exponential phase (K). White arrowheads indicate single cells containing a VirD2-CFP focus; gray arrowheads indicate large cells lackingVirD2-CFP signals. (L) Cells expressing truncated VirD2-GFP. Bars, 2 �m.
VOL. 193, 2011 LOCALIZATION OF CONJUGATION MACHINERY IN B. SUBTILIS 6251
on August 25, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

require at least a part of the pLS20 conjugation machinery toobtain their specific localization in lag-phase cells. These datareinforce the idea that two central components of the conju-gation machinery specifically assemble at the membrane tomediate conjugation during the early growth phase.
VirB4 and VirD4 colocalize and interact at defined sites atthe membrane. In order to investigate if VirB4 and VirD4localize to the same site(s), we expressed an mCherry fusion ofVirB4 from its original locus on pLS20 and VirD4-CFP froman ectopic locus on the chromosome. VirD4-CFP was ex-pressed at a low inducer concentration. With full induction,polar foci were not detectable, due to high background fluo-rescence. During lag phase and until early exponential phase (3h after inoculation), VirB4-mCherry and VirD4-GFP foci wereobserved, 80% of which colocalized at a single cell pole or atother sites along the cell membrane or within the cell (Fig. 4C),indicating that VirB4 and VirD4 assemble at the same site(s)within cells during lag phase. To test if both proteins form afunctional complex, we employed BiFC. The N-terminal partof YFP was fused to VirD4, and the C-terminal part was fusedto VirB4, both expressed from their original locus on pLS20.Each construct introduced into pLS20 by itself did not showany fluorescence (Fig. 4D and data not shown), but whenexpressed together, clear foci were detectable, mostly close tothe cell membrane and at a cell pole, in lag-phase cells (Fig.4E). Mid-exponential-phase growing cells did not show anyfoci; only a few small cells showed a fluorescent focus (Fig. 4F).During early stationary phase, no foci were detectable (datanot shown). As controls, we performed BiFC with VirB4-Ycand DnaX-Yn, but no foci were found even during lag phase(Fig. 4H). We also used the membrane protein MreC as acontrol, because MreC localizes at many positions along themembrane and also to the cell poles (13). However, no BiFCfoci were detected (Fig. 4G), showing that VirB4 and VirD4specifically interact at single defined sites at the cell membranein lag-phase cells, suggesting that they functionally interact inthe conjugation apparatus.
VirD2, VirB1, and VirB11 also localize to the membrane ina growth phase-dependent manner, but with different pat-terns. The finding that most lag-phase cells contain a polarlylocalized or membrane-localized plasmid copy suggested thatthe entire conjugation machinery may be present at these sites,such that ssDNA can be directly threaded into the exportchannel. If this were true, we would expect the VirD2 relaxaseprotein to also be present at this site, although it was possiblethat the two theoretical proteins required might not be re-solved against background fluorescence. However, we clearlyobserved VirD2-CFP foci at a single cell pole or at otherpositions in early-lag-phase cells (Fig. 4I). During mid-expo-nential phase, foci were only observed in the few remainingsingle cells, while in cell chains, only faint diffuse fluorescencewas detectable (Fig. 4K). Thus, like VirB4 and VirD4, therelaxase is present at the presumed sites of transfer at themembrane only during commencement of growth. In contrastto VirB4 and VirD4, VirD2-CFP foci were again detectable inearly-stationary-phase cells (10 h) (Fig. 4J), indicating thatearly steps in conjugation may already commence at this timepoint, while the entire machinery assembles at a later timepoint (24 h). The C-terminal region of A. tumefaciens VirD2has been shown to be important for efficient DNA transfer and
for interaction with the type IV secretion system (37). Wetested whether this also holds true for B. subtilis by integratinga truncated GFP fusion into the original virD2 locus. Interest-ingly, truncated VirD2-GFP still localized to the pole or to thelateral membrane (Fig. 4L), indistinguishably from wild-typeVirD2, and the conjugation frequency of pLS20 carrying trun-cated VirD2 was not considerably affected (Table 2), showingthat the mode of action of pLS20 VirD2 differs markedly fromthat of the Gram-negative system.
The third essential ATPase motif protein, VirB11, showed adifferent localization pattern. The fusion protein started toaccumulate as discrete foci at the cell pole and at the lateralmembrane in lag-phase cells, 1 h after inoculation (Fig. 5A).These foci became brighter, and 2 h after inoculation, VirB11-YFP formed additional irregular foci along the entire cellmembrane (Fig. 5B). As cells entered exponential growth (3 hafter inoculation), VirB11-YFP foci localized in a punctatehelical pattern throughout the membrane and also at the newlyformed septum (Fig. 5C) (showing that cells had commencedand completed a cell cycle), where other Vir proteins are rarelyobserved (Fig. 2 and 4). During this early exponential phase,most cells showed a focus or a gradient accumulation ofVirB11-YFP at a single cell pole (Fig. 5C). In many cells, ahelical pattern of VirB11-YFP foci was apparent. In mid-ex-ponential growth, 20% of all cells exclusively showed the he-lical pattern of localization (but no more polar foci), whichdisappeared during late exponential phase (Fig. 3C). Thus,VirB11 also accumulates at the presumed conjugation siteduring lag phase and also throughout the membrane duringexponential growth, when the above-mentioned conjugationproteins have already disassembled. To find out if VirB11diffuses through the cell membrane or is somewhat anchored atits specific sites, we performed FRAP (fluorescence recoveryafter photobleaching) analysis. In contrast to the membraneproteins ATPase and succinate dehydrogenase from B. subtilis,which show half-time recoveries of 3 to 4 min (23), VirB11-YFP fluorescence did not markedly recover over a 35-minperiod (Fig. 5D). These data show that VirB11 is somewhatstatically positioned within the membrane and does not showconsiderable diffusion rates.
VirB1 is a protein that is assumed to be associated with thecell wall. It has a predicted transmembrane helix at its Nterminus, so the protein including its C terminus are suppos-edly outside the cell. Due to this fact, we fused VirB1 tomCherry, which in contrast to GFP variants can be transportedacross the membrane. Strikingly, VirB1 accumulated at the cellpoles and at septa but also showed faint and homogeneouslocalization along the lateral sides of the cells during lag phase(Fig. 5F). However, accumulation at the poles was only ob-served for a subset of the cells (about 10%), indicating thatconjugation employing VirB1 occurs only in a minority of thecells. Different from most other investigated conjugation pro-teins, and similar to VirB11, VirB1-mCherry still localized incells during mid-exponential phase (Fig. 5G).
We also succeeded in generating a strain expressing VirB11-YFP and VirB1-mCherry, which is functional (several otherdual labels were not functional and/or were mislocalized). Fig-ure 5H shows that both signals frequently colocalized (in about75% of the cells having either signal), and in the remainingcells, either one of the constructs was visible at a position
6252 BAUER ET AL. J. BACTERIOL.
on August 25, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

where the other was not present. Therefore, VirB11 and VirB1are present largely at the same position, like VirB4 and VirD4,suggesting that the conjugation machinery coassembles at thesame subcellular sites.
Synthesis of VirB4 is temporally regulated. As mentionedabove, VirB4 and VirD4 coassembled at the membrane only incells resuming growth, but not during exponential growth orearly stationary phase. This was also true for VirB1 and VirB11
FIG. 5. Localization of VirB11 and VirB1. (A) VirB11-YFP 1 h after inoculation. (B) VirB11-YFP 2 h after inoculation. (C) VirB11-YFP 3 hafter inoculation. (D) FRAP analysis of VirB11-YFP during early exponential phase. The dashed circle marks the area of bleaching, and numbersin top right corners of panels indicate minutes after bleaching. (E) Simultaneous visualization of VirB11-YFP and DnaX-mCherry in cells 4 h afterinoculation (mid-log phase). White triangles indicate cells containing DnaX-mCherry foci and VirB11-YFP foci. (F) VirB1-mCherry 2 h afterinoculation. (G) VirB1-mCherry 4 h after inoculation (mid-exponential phase). White lines indicate septa between cells. (H) Colocalization ofVirB11-YFP and VirB1-mCherry (several sites are indicated by white triangles). Please note that there was no bleeding of YFP into the mCherrychannel, and vice versa (data not shown). Bars, 2 �m.
VOL. 193, 2011 LOCALIZATION OF CONJUGATION MACHINERY IN B. SUBTILIS 6253
on August 25, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

(Fig. 5). VirD2 accumulates in early stationary phase and dur-ing lag phase, but not during exponential growth. The assemblyof the entire machinery completely coincides with conjugationactivity (Fig. 3A to C), suggesting that conjugation is driventhrough assembly of the machinery during transition from non-growth to growth. In this respect, it is interesting to note thatPY79 cells carrying pLS20 have a reproducibly longer lagphase than cells lacking the plasmid (see Fig. S3 in the sup-plemental material), which was most noticeable for cells grow-ing at 30°C. These data suggest that pLS20 alters the physiol-ogy of the host cells during the transition from the stationaryphase to growth, extending the period in which it can be trans-ferred into recipient cells.
We wanted to analyze if VirB4 assembles and disassemblesdue to induction and arrest of protein synthesis, and so weperformed Western blot analysis with a VirB4-TAP tag fusion(GFP antiserum resulted in high background in cells carryingpLS20, precluding analysis of all generated FP fusions) thatwas expressed from the original virB4 locus. Figure 3D showsthat VirB4-TAP was undetectable in stationary-phase cells butwas rapidly induced upon resuspension in fresh medium.VirB4 disappeared during mid-exponential phase. Thus, VirB4is synthesized at the time of assembly but disappears later thanthe time of disassembly of VirB4-YFP foci. VirD4-GFP wasdetectable throughout the cells during mid-exponential growthbased on homogeneous cytosolic staining, and VirB11-YFPstill localized to the membrane in a punctate helical patternduring mid-exponential phase, suggesting that also in thesecases, disassembly of the conjugation machinery is not drivenby the degradation of these components. It will be interestingto identify the factor(s) driving the unusual behavior of pLS20.
DISCUSSION
Our work provides evidence that the conjugation machineryin the Gram-positive model organism B. subtilis assembles at asingle site at the membrane, frequently at the cell pole. Wefound that the pLS20 plasmid as well as VirD2, VirB4, VirD4,and VirB1 localize at the pole, in cells during extended sta-tionary phase, and in an even more pronounced fashion in cellsentering growth. VirB11 protein is present at many sites alongthe cell membrane and is also frequently concentrated at asingle cell pole. Using BiFC, we showed that VirD4 and VirB4functionally interact, suggesting that the type IV secretion sys-tem-like apparatus operates from a single defined subcellularsite. However, VirD2, VirB4, and VirD4 no longer localized incells that had commenced exponential growth, although theyappeared to be continued to be synthesized. The assembly anddisassembly of the machinery occur in parallel with the induc-tion and decline of conjugation activity of pLS20 (22). Whencells from early stationary phase, in which all monitored con-jugation proteins are delocalized except for VirD2, are resus-pended into fresh medium, the conjugation machinery assem-bles within as little as 60 min and is functional, revealing rapidassembly kinetics. Thus, our work provides striking evidencefor a machinery that commences to assemble in non-exponen-tially growing cells, to become active during the transition togrowth, and is then rendered inactive as an active cell cyclebecomes operative.
In agreement with these findings, we found that pLS20 is
mostly membrane associated and frequently found at the cellpole. Cells contain on average 3 to 4 plasmids during exponen-tial growth, but only 1 to 2 during stationary phase. It is likelythat one plasmid is present at the conjugation machinery, suchthat conjugation can occur very rapidly. Indeed, the 65-kbplasmid can conjugate in as little as 2 min (see Fig. S2 in thesupplemental material). Corroborating the identification of anentirely novel replication mechanism, we found that pLS20rarely colocalized with the chromosomal replication machin-ery. PLS20 codes for several replication proteins (a primase, aligase, and an SSB paralog) and may use only a few compo-nents of the host replication setup. Interestingly, we found thatpolar pLS20 was generally statically located at its positionwhile nonpolar plasmids were mobile, suggesting that the con-jugation machinery stably anchors the plasmid, even in cellsthat are not actively conjugating (i.e., in the absence of recip-ient cells).
Our work also provides evidence for different patterns oflocalization of Vir proteins, as summarized in Table 3. VirB11,the third ATPase motif protein besides VirB4 and VirD4,accumulated at the cell pole soon after resuspension of sta-tionary-phase cells into fresh medium and later during expo-nential growth localized to many discrete foci along the entirecell membrane, although a higher concentration was seen to-ward a single cell pole. The putative cell wall hydrolase VirB1was present at the entire surface of the cell pole, while VirB4,VirD2, and VirD4 formed discrete foci. Our findings mayreconcile data from A. tumefaciens and other bacteria in whichit has also been reported that some proteins localized all alongthe lateral cell membrane (2) while others were only present atthe cell pole (3, 28). It is interesting that the VirB4 componentof a conjugative transposon from B. subtilis also localizes to thecell poles (6), indicating that transposon conjugation may alsooccur at the cell pole. Our data suggest that the pLS20 conju-gation machinery is composed of a core machinery and differ-ent modules that are also present at other places in the cell,where they are nonactive. Core proteins VirB1, VirB4, andVirD4 have also been identified on another conjugative plas-mid in a Gram-positive bacterium, and they also interact witheach other (1), suggesting that many components of and con-nections within the conjugation machineries are conserved.PLS20 VirB11 protein did not diffuse through the cell mem-brane but showed a rather static arrangement. These datasuggest that at least VirB11 is anchored by an unknown factorand/or is attached to the cell membrane.
We asked ourselves why a conjugative plasmid may regulate
TABLE 3. Patterns of localization of Vir proteins: localization atthe pole and membrane as foci
Protein
Localizationa during growth phase
Lag Mid-exponential
Earlystationary
Latestationary
VirB4 � � � �VirD4 � � � �VirB11 � (�) (helices) � �VirD2 � � � VirB1b � � � �
a �, present; �, absent.b Did not form discrete foci.
6254 BAUER ET AL. J. BACTERIOL.
on August 25, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

its transfer to another cell at a time when there are fewersiblings around (dilution into fresh medium) and not when celldensity is high and many recipient cells are available (i.e., thetime point when competence is induced). We speculate that alarge plasmid that requires at least 2 min for transfer (see Fig.S2 in the supplemental material) and has an active partitioningsystem may distribute to recipients early during growth, whennutrients are readily available. During growth, it is efficientlypropagated through its active mitotic-like segregation machin-ery and has no need for conjugation. It is also possible that thecell wall becomes amenable for the assembly of the complexwhen it commences to remodel and extend, while stationary-phase cells are known for their increased robustness andchanges in peptidoglycan architecture, which may hinder con-jugation. Also, B. subtilis cells grow as predominantly nonmo-tile chains of cells, facilitating cell/cell contact, but becomehighly motile when they enter stationary phase.
Intriguingly, we found that cells carrying pLS20 have anextended lag phase. PLS20-carrying cells commenced expo-nential growth considerably later than plasmid-free cells, whilethe doubling times of both strains were indistinguishable.These findings indicate that the presence of the plasmid mayalter the response of cells to the addition of fresh medium. Thisway, the plasmid may increase the time to find a suitablerecipient cell. It will be interesting to determine if the plasmidinterferes with the physiology of the host cell simply by beinga metabolic burden or by changing the activity of host proteinsor the transcription of host genes.
It will be also illuminating to identify the trigger that inducesthe assembly of the conjugation machinery and the signal(s)that leads to disassembly. Interestingly, we found that VirD2accumulates at the cell pole in early-stationary-phase cells,while VirD4 and VirB4 assemble about 12 h later, duringextended stationary phase. These findings suggest that earlysteps in conjugation are induced as cells exit growth and thatthe core conjugation machinery is set up during continuedstationary phase. The entire machinery then appears to be setup as cells gain a new nutritional supply and the incentive tocommence growth. Thus, the Gram-positive conjugation ma-chinery appears to be built in a somewhat hierarchical order. Itwill also be important to identify the factors that induce mem-brane targeting of VirD4 and VirB4. On their own, they local-ize throughout the cytosol and, therefore, are apparentlysoluble proteins, although VirD4 contains 2 predicted trans-membrane helices at its N terminus. Most of the proteins wehave analyzed in this study appear to be synthesized from earlygrowth into stationary phase, so there appears to be an assem-bly factor for the machinery that needs to be identified.
ACKNOWLEDGMENTS
We thank Shinya Kaneko, Tokyo Institute of Technology, Nagat-suta, Yokohama-shi Kanagawa, Japan, for providing pLS20neo beforepublication and Stefanie Welling for providing the DnaX-Yc construct.
This work was supported by the Deutsche Forschungsgemeinschaft(SFB 746) and the SGBM Freiburg.
REFERENCES
1. Abajy, M. Y., et al. 2007. A type IV secretion-like system is required forconjugative DNA transport of broad-host-range plasmid pIP501 in Gram-positive bacteria. J. Bacteriol. 189:2487–2496.
2. Aguilar, J., J. Zupan, T. A. Cameron, and P. C. Zambryski. 2010. Agrobac-terium type IV secretion system and its substrates form helical arrays around
the circumference of virulence-induced cells. Proc. Natl. Acad. Sci. U. S. A.107:3758–3763.
3. Atmakuri, K., E. Cascales, O. T. Burton, L. M. Banta, and P. J. Christie.2007. Agrobacterium ParA/MinD-like VirC1 spatially coordinates early con-jugative DNA transfer reactions. EMBO J. 26:2540–2551.
4. Atmakuri, K., E. Cascales, and P. J. Christie. 2004. Energetic componentsVirD4, VirB11 and VirB4 mediate early DNA transfer reactions required forbacterial type IV secretion. Mol. Microbiol. 54:1199–1211.
5. Atmakuri, K., Z. Ding, and P. J. Christie. 2003. VirE2, a type IV secretionsubstrate, interacts with the VirD4 transfer protein at cell poles of Agrobac-terium tumefaciens. Mol. Microbiol. 49:1699–1713.
6. Berkmen, M. B., C. A. Lee, E. K. Loveday, and A. D. Grossman. 2010. Polarpositioning of a conjugation protein from the integrative and conjugativeelement ICEBs1 of Bacillus subtilis. J. Bacteriol. 192:38–45.
7. Cascales, E., and P. J. Christie. 2004. Definition of a bacterial type IVsecretion pathway for a DNA substrate. Science 304:1170–1173.
8. Cascales, E., and P. J. Christie. 2003. The versatile bacterial type IV secre-tion systems. Nat. Rev. Microbiol. 1:137–149.
9. Chen, I., and D. Dubnau. 2004. DNA uptake during bacterial transforma-tion. Nat. Rev. Microbiol. 2:241–249.
10. Christie, P. J. 1997. Agrobacterium tumefaciens T-complex transport appa-ratus: a paradigm for a new family of multifunctional transporters in eubac-teria. J. Bacteriol. 179:3085–3094.
11. Christie, P. J., K. Atmakuri, V. Krishnamoorthy, S. Jakubowski, and E.Cascales. 2005. Biogenesis, architecture, and function of bacterial type IVsecretion systems. Annu. Rev. Microbiol. 59:451–485.
12. de Antonio, C., M. E. Farias, M. G. de Lacoba, and M. Espinosa. 2004.Features of the plasmid pMV158-encoded MobM, a protein involved in itsmobilization. J. Mol. Biol. 335:733–743.
13. Defeu Soufo, H. J., and P. L. Graumann. 2006. Dynamic localization andinteraction with other Bacillus subtilis actin-like proteins are important forthe function of MreB. Mol. Microbiol. 62:1340–1356.
14. Defeu Soufo, C., et al. 2008. Cell-cycle-dependent spatial sequestration of theDnaA replication initiator protein in Bacillus subtilis. Dev. Cell 15:935–941.
15. de la Cruz, F., L. S. Frost, R. J. Meyer, and E. L. Zechner. 2010.Conjugative DNA metabolism in Gram-negative bacteria. FEMS Micro-biol. Rev. 34:18–40.
16. Derman, A. I., et al. 2009. Phylogenetic analysis identifies many uncharac-terized actin-like proteins (Alps) in bacteria: regulated polymerization, dy-namic instability and treadmilling in Alp7A. Mol. Microbiol. 73:534–552.
17. Francia, M. V., et al. 2004. A classification scheme for mobilization regionsof bacterial plasmids. FEMS Microbiol. Rev. 28:79–100.
18. Garcillan-Barcia, M. P., M. V. Francia, and F. de la Cruz. 2009. The diver-sity of conjugative relaxases and its application in plasmid classification.FEMS Microbiol. Rev. 33:657–687.
19. Gilmour, M. W., J. E. Gunton, T. D. Lawley, and D. E. Taylor. 2003.Interaction between the IncHI1 plasmid R27 coupling protein and type IVsecretion system: TraG associates with the coiled-coil mating pair formationprotein TrhB. Mol. Microbiol. 49:105–116.
20. Gomis-Ruth, F. X., et al. 2001. The bacterial conjugation protein TrwBresembles ring helicases and F1-ATPase. Nature 409:637–641.
21. Grohmann, E., G. Muth, and M. Espinosa. 2003. Conjugative plasmid trans-fer in gram-positive bacteria. Microbiol. Mol. Biol. Rev. 67:277–301.
22. Itaya, M., N. Sakaya, S. Matsunaga, K. Fujita, and S. Kaneko. 2006. Con-jugational transfer kinetics of pLS20 between Bacillus subtilis in liquid me-dium. Biosci. Biotechnol. Biochem. 70:740–742.
23. Johnson, A. S., S. van Horck, and P. J. Lewis. 2004. Dynamic localization ofmembrane proteins in Bacillus subtilis. Microbiology 150:2815–2824.
24. Kearns, D. B., and R. Losick. 2005. Cell population heterogeneity duringgrowth of Bacillus subtilis. Genes Dev. 19:3083–3094.
25. Kidane, D., H. Sanchez, J. C. Alonso, and P. L. Graumann. 2004. Visual-ization of DNA double-strand break repair in live bacteria reveals dynamicrecruitment of Bacillus subtilis RecF, RecO and RecN proteins to distinctsites on the nucleoids. Mol. Microbiol. 52:1627–1639.
26. Koehler, T. M., and C. B. Thorne. 1987. Bacillus subtilis (natto) plasmidpLS20 mediates interspecies plasmid transfer. J. Bacteriol. 169:5271–5278.
27. Kopec, J., A. Bergmann, G. Fritz, E. Grohmann, and W. Keller. 2005. TraAand its N-terminal relaxase domain of the Gram-positive plasmid pIP501show specific oriT binding and behave as dimers in solution. Biochem. J.387:401–409.
28. Kumar, R. B., and A. Das. 2002. Polar location and functional domains of theAgrobacterium tumefaciens DNA transfer protein VirD4. Mol. Microbiol.43:1523–1532.
29. Kuroki, A., N. Ohtani, K. Tsuge, M. Tomita, and M. Itaya. 2007. Conjuga-tional transfer system to shuttle giant DNA cloned by Bacillus subtilis ge-nome (BGM) vector. Gene 399:72–80.
30. Llosa, M., S. Zunzunegui, and F. de la Cruz. 2003. Conjugative couplingproteins interact with cognate and heterologous VirB10-like proteins whileexhibiting specificity for cognate relaxosomes. Proc. Natl. Acad. Sci. U. S. A.100:10465–10470.
31. Meijer, W. J., A. J. de Boer, S. van Tongeren, G. Venema, and S. Bron. 1995.
VOL. 193, 2011 LOCALIZATION OF CONJUGATION MACHINERY IN B. SUBTILIS 6255
on August 25, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

Characterization of the replication region of the Bacillus subtilis plasmidpLS20: a novel type of replicon. Nucleic Acids Res. 23:3214–3223.
32. Pansegrau, W., and E. Lanka. 1996. Enzymology of DNA transfer by con-jugative mechanisms. Prog. Nucleic Acid Res. Mol. Biol. 54:197–251.
33. Savvides, S. N., et al. 2003. VirB11 ATPases are dynamic hexameric assem-blies: new insights into bacterial type IV secretion. EMBO J. 22:1969–1980.
34. Schroder, G., et al. 2002. TraG-like proteins of DNA transfer systems and ofthe Helicobacter pylori type IV secretion system: inner membrane gate forexported substrates? J. Bacteriol. 184:2767–2779.
35. Szpirer, C. Y., M. Faelen, and M. Couturier. 2000. Interaction between theRP4 coupling protein TraG and the pBHR1 mobilization protein Mob. Mol.Microbiol. 37:1283–1292.
36. Tanaka, T., and T. Koshikawa. 1977. Isolation and characterization of
four types of plasmids from Bacillus subtilis (natto). J. Bacteriol. 131:699–701.
37. van Kregten, M., B. I. Lindhout, P. J. Hooykaas, and B. J. van der Zaal.2009. Agrobacterium-mediated T-DNA transfer and integration by minimalVirD2 consisting of the relaxase domain and a type IV secretion system.translocation signal. Mol. Plant Microbe Interact. 22:1356–1365.
38. Ward, D. V., O. Draper, J. R. Zupan, and P. C. Zambryski. 2002. Peptidelinkage mapping of the Agrobacterium tumefaciens vir-encoded type IV se-cretion system reveals protein subassemblies. Proc. Natl. Acad. Sci. U. S. A.99:11493–11500.
39. Webb, C. D., et al. 1998. Use of time-lapse microscopy to visualize rapidmovement of the replication origin region of the chromosome during the cellcycle in Bacillus subtilis. Mol. Microbiol. 28:883–892.
6256 BAUER ET AL. J. BACTERIOL.
on August 25, 2019 by guest
http://jb.asm.org/
Dow
nloaded from





























