Embed Size (px)
Citation preview

Ostracod palaeoecology and environmental change in the Laptev andKara seas (Siberian Arctic) during the last 18 000 years
ANNA STEPANOVA, EKATERINA TALDENKOVA AND HENNING A. BAUCH
Stepanova, A., Taldenkova, E. & Bauch, H. A. 2012: Ostracod palaeoecology and environmental change in theLaptev and Kara seas (Siberian Arctic) during the last 18 000 years. Boreas, 10.1111/j.1502-3885.2012.00254.x.ISSN 0300-9483.
Fossil ostracod assemblages were investigated in five AMS 14C-dated cores from various water depths of theLaptev and Kara seas ranging from the upper continental slope (270 m) to the present-day shelf depth (40 m). Sixfossil assemblages were distinguished. These represent the varying environmental conditions at the North Siberiancontinental margin since about 18 ka. In the cores from the shelf the ostracod assemblages reflect the gradualtransition from an estuarine brackish-water environment to modern marine conditions since 12.3 ka, as inducedby the regional early Holocene transgression. The core from the upper continental slope dates back to c. 17.6 kaand contains assemblages that are absent in the shelf cores. The assemblage older than 10 ka stands out as aspecific community dominated by relatively deep-water Arctic and North Atlantic species that also containseuryhaline species. Such an assemblage provides evidence for past inflows of Atlantic-derived waters from as earlyas c. 17.2 ka, probably facilitated by upwelling in coastal polynyas, and a considerable riverine freshwaterinfluence with enhanced surface water stratification owing to the proximity of the palaeocoastline until earlyHolocene times. In all studied cores, relative increases in euryhaline species dominant in the inner-shelf regions arerecorded in the mid–late Holocene sediments (<7 ka), which otherwise already contain modern-like ostracodassemblages with relatively deep-water species. This observation suggests euryhaline species to be largely sea-ice-and/or iceberg-rafted and therefore may provide evidence for a climate cooling trend.
Anna Stepanova (e-mail: [email protected]), Borisyak Paleontological Institute RAS, Profsoyuznaya,123, Moscow, 117997, Russia; Ekaterina Taldenkova (e-mail: [email protected]), Geographical Faculty,Lomonosov Moscow State University, Leninskie Gory, 1, Moscow, 119991, Russia; Henning A. Bauch (e-mail:[email protected]), Mainz Academy for Science, Humanities and Literature, c/o GEOMAR Helmholtz Centre forOcean Research, Wischhofstrasse 1-3, 24148 Kiel, Germany; received 7th February 2011, accepted 27th January2012.
The late Quaternary palaeoceanographic record of theArctic is characterized by marked shifts in climate andsea level (Bauch et al. 1999, 2001; Spielhagen et al.2004). These changes are strongest at the interfacebetween marine and terrestrial systems, owing to thehigh instability of this environment related to the inputof terrigenous material, high organic productivity, pro-nounced seasonality and large changes in salinity (Steinet al. 2001, 2004).
Ostracods represent an important group of benthicorganisms inhabiting a wide range of aquatic environ-ments, from freshwater pools to marine deep-seabasins. Ostracods rapidly invade newly created habitatsand are able to track their preferred environmental andclimatic optima (Griffiths 2001). The taxonomic com-position of ostracod assemblages is strongly dependenton environmental conditions. Certain index taxa arerestricted to a specific range of environmental param-eters and can reflect changes of water-mass propertieseven on decadal time scales, making ostracod assem-blage analysis a very precise instrument (AlvarezZarikian et al. 2000; Cronin et al. 2002; Cronin &Vann 2003). For this reason they are frequentlyused as palaeo-environmental indicators of bottom-water salinity and temperature (Cronin 1977, 1981;McDougall et al. 1986; Stepanova 2006).
Previous studies of recent ostracods from the Laptevand Kara seas have covered environments ranging fromfreshwater lagoons and brackish-water estuaries to rela-tively deep-water parts of the upper continental slope(Stepanova et al. 2003, 2004, 2007). Distribution pat-terns of ostracods in these river-influenced marginal seasshow a clear relationship with salinity and water depth.
Using these data on recent Arctic ostracod assem-blages, we carried out a detailed study of fossil ostra-cods from five AMS 14C-dated sediment cores from theLaptev and Kara seas (Bauch et al. 2001; Simstich et al.2004; Taldenkova et al. 2005, 2008, 2010) in order toanalyse the transition from Lateglacial to modern envi-ronments in the High Arctic continental marginal area.These regions underwent a transformation from terres-trial to marine environments under postglacial sea-levelrise and faced climate-induced changes in water-masscirculation, freshwater influence and extent of sea icecover (Bauch et al. 1999, 2001; Polyakova & Stein2004; Stein et al. 2004; Polyakova et al. 2005;Taldenkova et al. 2005, 2008, 2009, 2010; Darby et al.2006). Our inferences about past environmentalchanges are based on the analysis of ostracod abun-dance and taxonomy, the ecology of index and domi-nant species, and the range of individual speciestolerance to changing salinity and water depth.
bs_bs_banner
DOI 10.1111/j.1502-3885.2012.00254.x © 2012 The AuthorsBoreas © 2012 The Boreas Collegium

Materials and methods
Materials and sampling
Five core sections were analysed, four from differentparts of the Laptev Sea and one from the Kara Sea(Fig. 1; Table S1 in Supporting Information). Coresfrom the Laptev Sea were obtained in 1998 duringthe TRANSDRIFT V expedition aboard the R/V
‘Polarstern’. Three cores from the Laptev Sea shelf arelocated in the palaeovalleys of the biggest rivers of theregion. The eastern cores include core PS51/138-12from the Lena palaeovalley (water depth 45 m) andcore PS51/135-4 from the Yana palaeovalley (waterdepth 51 m). Core PS51/159-10 was retrieved fromthe Khatanga palaeovalley (water depth 60 m) on thewestern outer shelf, whereas core PS51/154-11 origi-nates from the western continental slope (water depth
aimyr PeninsulaK o t e l n y y
Lena Delta
Tiksi
Anabar
Olenek
Yana 71
10
10 10
10
10
10
10
10
10
10
10
10
10
10
10
10
10
10
10
1010
10
1010 10
10
10
20
20
20
20
20
20 2
0
20
20
20
20
20
20
20
20
20
20
2020
20
20
20
30
30
30
30 30
30
30 30
30
30
30
30
40
40
40
40
40
40
40
40
40
40
40
50
50
50
50
50
50
50
50
Taimyr PeninsulaK o t e l n y i
Tiksi
Khata
nga
Anabar
Olenek
Yana
105 110 1 120 125 130 135 140 145
105 110 1 120 125 140 145
72
73
74
75
76
77
78
72
73
74
75
76
77
10
10 10
10
10
10
10
10
10
10
10
10
10
10
10
10
10
10
10
1010
10
1010 10
10
10
20
20
20
20
20
20 2
0
20
20
20
20
20
20
20
20
20
20
2020
20
20
20
30
30
30
30 30
30
30 30
30
30
30
30
40
40
40
40
40
40
40
40
40
40
40
50
50
50
50
50
50
50
50
71
T
10
135
100
130
100
LenaDelta
PS51/138-12
PS51/135-4PS51/159-10
PS51/154-11
154
Barents-Kara Ice Sheet
Laptev Sea
Fram Strait
Arctic Ocean
Kara Sea
07
159
138
135
A
Laptev Sea
1 5
C
RussiaB
C5
80 82 84
69
70
71
72
73
74
75
76
77
-10
-10
-10
-10
-10
-25-25
-25
-25
-25
-50
-50
-50
-50
-50
-50
-100
2
787674 E
N
Ob’
Yenisei
BP00/07-5
Kara SeaB
1 5
Severnaya Zemlya
100 km
100 km100 km
Fig. 1. A. Distribution of Atlantic-derived waters and ice-sheet boundaries during the Last Glacial Maximum (Svendsen et al. 2004). Numbersin the figure correspond to the studied core sites: 07=BP00/07-5; 154=PS51/154-11; 159=PS51/159-10; 135=PS51/135-4; 138=PS51/138-12. B.Site locations in the Kara Sea. C. Site locations in the Laptev Sea. This figure is available in colour at http://www.boreas.dk.
2 Anna Stepanova et al. BOREAS

270 m). Core BP00/07-5 was obtained from the easternKara Sea shelf (water depth 43 m) during expeditionSIRRO-2000 aboard the R/V ‘Akademik BorisPetrov’. It originates from a topographic depression, aformer Yenisei valley now filled with sediments (Sim-stich et al. 2004). Hereafter the cores will be referred toas PS138, PS135, PS159, PS154 and BP07.
Cores were sampled in 2–3 cm thick slices at differentintervals: cores PS138 and BP07 were sampled at2–10 cm step intervals, whereas cores PS159 and PS154were sampled continuously. Samples from these coreswere freeze-dried and weighed. Core PS135 was origi-nally sampled onboard at 30-cm intervals. These large-size samples (approximately 400 mL) were washedonboard without weighing. Later, the core wasre-sampled at 10-cm intervals (~100 mL). These lattersamples were freeze-dried, weighed and washed over a63-mm mesh-size sieve. Ostracods were picked fromsample residues, identified and counted. All ostracodvalves are stored at the Borisyak Paleontological Insti-tute of the Russian Academy of Sciences (Moscow);counts and distribution data are available at:doi:10.1594/PANGAEA.735539.
Core chronologies
The chronology of all cores is based on radiocarbondates of marine biogenic carbonate of bivalves, ostra-cods and foraminifers (Table S1) (Bauch et al. 2001;Simstich et al. 2004; Taldenkova et al. 2008, 2009,2010). Dates were obtained by accelerator mass spec-trometry (AMS) at the Leibniz Laboratory in Kiel.Reservoir age corrections were applied as follows:-370 years for the Laptev Sea (Bauch et al. 2001) and440 years for the Kara Sea (Simstich et al. 2004). Forconverting the AMS 14C dates to calendar years BPwe used the Fairbanks 0107 program (Fairbanks et al.2005). Application of the Fairbanks 0107 program forthe dates from the Kara Sea showed insignificant agedifferences from the originally obtained age estima-tions of Simstich et al. (2004) based on the calibrev.4.3 program (Stuiver & Reimer 1993) (Table S1).All ages noted in the text are calculated calendarages.
Ages between dated samples were estimated by linearinterpolation and assuming a modern age for the sedi-ment surface (Fig. 2). When calculating the age modelfor core PS154 we excluded the two reversed dates from31 and 39 cm depth obtained on bivalves and gastro-pods (Table S1), as these are probably the result of theburrowing of molluscs into the sediment accumulatedunder low sedimentation rates. The sediment sectionbetween 51 and 115 cm is strongly bioturbated andshows almost the same age (Table S1). Therefore, onlythe youngest and the oldest dates were taken for agemodel calculations, and the date obtained on foramini-fers from 73 cm was assigned to 51 cm core depth.
Micropalaeontological analysis
For ostracod assemblage characterization we comparethe number of species per sample, the total abundanceof specimens, and the abundance of different ecologicalgroups and individual taxa (Figs 3–7). For core PS135(Fig. 3) we produced species percentages instead of thespecies abundances because of the two sets of samplesused (see ‘Materials and sampling’): the large-sizesamples lack dry bulk weight information, while thesmall-size samples contain few ostracod valves, pre-sumably because of a certain degree of dissolution insediment during storage. For the relative percentage,samples with fewer than 10 valves were excluded fromthe calculation. Since the number of specimens in mostof the small-size samples was less than 5 valves persample, when compared with the original large-sizesamples that average 50–70, we produced species per-centage plots only from the latter set of samples.
In general, ostracod occurrence in marine sedimentsis rare compared with other fossil groups. Ostracodsgrow discontinuously, and their juvenile stages are pre-served. It has been shown, for instance, that the numberof juvenile valves (population-age structure) is ameasure characterizing the energy type of an environ-ment together with the preservation of an ostracodassemblage (Whatley 1983) (Figs 3–7). Whatley (1983)distinguished three types of assemblages, interpretationof which can vary: (i) with up to 80% of juveniles ofdifferent instars (life assemblage in a low-energy envi-ronment); (ii) with at least 25% of juveniles of olderinstars (either a life assemblage in a high-energy envi-ronment when smaller juveniles and dead valves arewashed out or a thanatocoenosis with current velocitiessufficient to carry larger allochtonous valves); and (iii)100% juvenile percentage (mass burial indicating acatastrophic event or low-energy environment wherethe current velocity is low and smaller allochtonousvalves could be introduced). Although the percentageof juveniles in our samples is quite variable, indicatinga rather high-energy environment, ostracod valve pres-ervation was generally good, without traces of corro-sion. The exception is in core PS154, where theextremely poorly resolved interval corresponding to9–7 ka contained only few ostracods, and those were ofrelatively poor preservation.
The Shannon–Weaver diversity index (Shannon &Weaver 1963) was calculated for all cores (Fig. 8). It is
defined as H S p pi ii
S
( )=− ( )=∑ ln
1
, where pi is the pro-
portion of the ith species and S is the total number ofspecies. The Shannon-Weaver diversity index is rela-tively insensitive to sample size and low total abun-dance (Alvarez Zarikian et al. 2009). Low index values(typically around 1.5) indicate low diversity andhigh dominance in association with stressful condi-tions, whereas high values (around 3.5) correspond to
Ostracod palaeoecology and environmental change in the Laptev and Kara seas, last 18 000 years 3BOREAS

5908(CB)
1061(CT)
- ,Core Base inter/extrapolated age
- ,Core Top extrapolated age
- Core Catcher, extrapolated age
0
100
200
300
400
500
0 2 4 6 8 10 12
11 335(CB)
11 426(CC)
11 225
10 358
9537
9015
7589
69775252
184
371.5
De
pth
(cm
)
0 2 4 6 8
648
2778
3810
5598
6925
7978
80
(CB)
0
100
200
300
400
500
0 2 4 6 8 10 12
1205(CT)
2682
81938040
10 226
10 521
11 274
17
117
11305(CB)
17 582(CC)
0
100
200
300
400
500
600
0 2 4 6 8 10 12 14 16 18
700
32795398
10 208
10 44211 16511 187
11 250
12 175
13 661
13 941
14 856
15 33815 372
5
120
Age (cal. ka)
17 208(CB)
17 582(CC)
0
100
200
300
400
500
600
BP00-07/5
PS51/154-11
0
100
200
300
400
500
0
2 4 6 8 10 12 14
12 167
10 475
10 216
9541
6750
5337514
12 230(CB)
12 631(CC)
115
13
PS51/159-10
PS51/135-4PS51/138-12
De
pth
(cm
)
De
pth
( cm
)D
ep
th( c
m)
De
pth
(cm
)
Age (cal. ka)
Age (cal. ka)
Age (cal. ka)
Age (cal. ka)
Se
dim
en
tatio
n ra
te (c
m k
a )-1
Se
dim
en
tatio
n ra
te (c
m k
a )-1
Se
dim
en
tatio
n ra
te (c
m k
a )-1
Se
dim
en
tatio
n ra
te (c
m k
a )-1
Se
dim
en
tatio
n ra
te (c
m k
a )-1
8018
Fig. 2. Age–depth relationships in the studied cores. This figure is available in colour at http://www.boreas.dk.
4 Anna Stepanova et al. BOREAS
0
20
00
40
00
60
00
80
00
10
00
0
12
00
0
Paracyprideis pseudopunctillata
Rabilimis septentrionalis
Heterocyprideis sorbyana
Cytheromorpha macchesneyi
Cytheropteron sulense
Palmenella limicola
Acanthocythereis dunelmensis
Cytheropteron elaeni
Cluthia cluthaeJonesia sp.1
Cytheropteron suzdalskyi
Semicytherura complanata
Sarsicytheridea bradii
Cytheropteron sp.
Roundstonia globulifera
Sclerochilus sp.
PS
51
/13
5-4
, e
aste
rn L
ap
tev S
ea
sh
elf,
Ya
na
pa
lae
ova
lley (
76
.2°
N,
13
3.2
° E
, w
ate
r d
ep
th 5
1 m
)Age (cal. a)
Total abundance, valves per 100g *
Number of species *
Brackishwater, %
Euryhaline, %
Shallow-water marine,%
Juvenile valves, %
02
04
06
08
02
04
06
02
04
02
04
02
02
04
0
Cytheropteron montrosiense
20
40
60
20
20
20
20
40
20
40
20
40
10
20
55
50
100
51
05
0
50
100
50
100
50
100
Ostr
aco
d s
pe
cie
s (
%)
620
0
10 3
00
11 2
00
11 4
00
Fig
.3.
Ost
raco
dsp
ecie
sco
mpo
siti
onin
core
PS5
1/13
5-4
(Yan
apa
laeo
valle
y):t
otal
abun
danc
epe
r10
0g
dry
bulk
wei
ght,
dive
rsit
y,pe
rcen
tage
ofsp
ecie
s,ju
veni
leva
lves
,and
ecol
ogic
algr
oups
.*=
Onl
ysa
mpl
esw
ith
dry
bulk
wei
ghtd
ata
wer
eus
edfo
rca
lcul
atio
nof
the
tota
labu
ndan
cean
dnu
mbe
rof
spec
ies,
thes
ein
clud
ein
terv
als
wit
hve
rylo
wab
unda
nce.
Sam
ples
wit
hlo
wab
unda
nce
wer
eno
tus
edfo
rca
lcul
atin
gth
esp
ecie
spe
rcen
tage
(see
‘Mat
eria
lsan
dsa
mpl
ing’
).T
his
figur
eis
avai
labl
ein
colo
urat
http
://w
ww
.bor
eas.
dk.
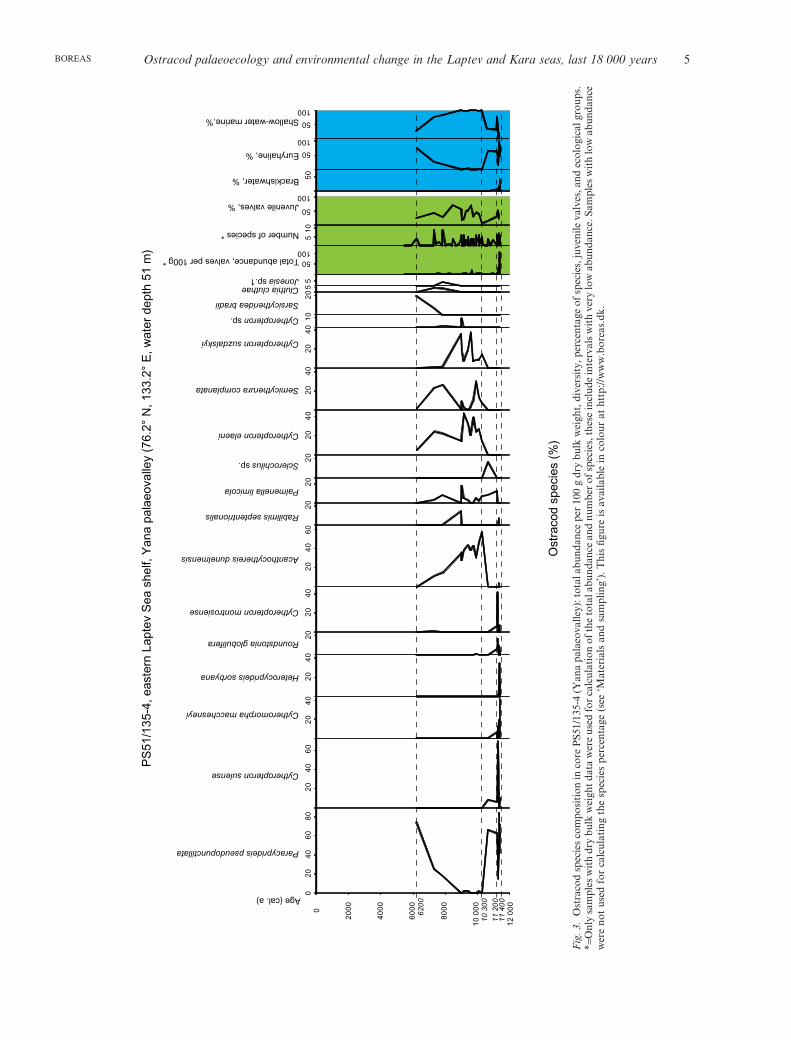
Ostracod palaeoecology and environmental change in the Laptev and Kara seas, last 18 000 years 5BOREAS

01
02
03
04
05
06
0
0
20
00
40
00
60
00
80
00
10
00
0
12
00
0
Paracyprideis pseudopunctillata
10
20
30
40
50
60
70
Rabilimis septentrionalis
10
20
Loxoconcha venepidermoidea
10
20
Heterocyprideis sorbyana
10
20
Cytheromorpha macchesneyi
10
20
Cytheropteron sulense
5Pteroloxa cumuloidea
5
Palmenella limicola
10
20
30
40
Acanthocythereis dunelmensis
5
Eucythere sp.
10
20
Cytheropteron elaeni
10Cluthia cluthae
5Jonesia sp.1
10
20
Cytheropteron suzdalskyi
10
20
30
Semicytherura complanata
10
20
30
40
Sarsicytheridea bradii
5Cytheropteron nodosoalatum
5Cytheropteron champlainum
5
Roundstonia globulifera
5Sclerochilus sp.
5Candona sp.
5Undetermined
PS
51
/13
8-1
2,
ea
ste
rn L
ap
tev S
ea
sh
elf,
Le
na
pa
lae
ova
lley (
75
.1°
N,
13
0.9
° E
, w
ate
r d
ep
th 4
5 m
)
1650
8200
10 2
00
11 3
00
50
150Total abundance, valves per 100g
10Number of species
20Freshwater and brackishwater/100g
50
Euryhaline/100g
50
150
Shallow-water marine/100g
2Deep-water marine/100g
50
Juvenile valves, %
Age (cal. a)
Ostr
aco
ds (
va
lve
s 1
00
g
)-1
Fig
.4.
Ost
raco
dsp
ecie
sco
mpo
siti
onin
core
PS5
1/13
8-12
(Len
apa
laeo
valle
y):t
otal
abun
danc
e,ab
unda
nce
ofec
olog
ical
grou
psan
dab
unda
nce
ofsp
ecie
spe
r10
0g
dry
bulk
wei
ght,
dive
rsit
y,pe
rcen
tage
ofju
veni
leva
lves
.Thi
sfig
ure
isav
aila
ble
inco
lour
atht
tp://
ww
w.b
orea
s.dk
.
6 Anna Stepanova et al. BOREAS

51
0Sarsicytheridea bradii
0
20
00
40
00
60
00
80
00
10
00
0
12
00
0
Rabilimis mirabilis
5
Cytheropteron laptevensis
5
Bythocythere spp.
5
Cytheropteron spp.
51
0Sarsicytheridea macrolaminata
5
Acanthocythereis dunelmensis
Pteroloxa sp.
5
Cytheretta teshekpukensis
5Semicytherura concentrica
10
20
Cytheropteron montrosiense
Krithe glacialis
Palmenella limicola
5Argilloecia sp.
51
0
Cytheropteron biconvexa
51
0Roundstonia globulifera
Cytheropteron paralatissimum
5
Elofsonella concinna
Heterocyprideis sorbyana
5Cytheromorpha macchesneyi
51
0Cytheropteron nodosum
51
0
Cytheropteron suzdalskyi
51
01
5
Cluthia cluthae
51
0
Rabilimis spp.
10
10
20
Paracyprideis pseudopunctillata
5
Cytheropteron tumefactum
Cytheropteron sulense
51
0
Undetermined
51
0
Cytheropteron porterae
Cytheropteron nodosoalatum
5
Sarsicytheridea sp.
5
Cytheropteron arcuatum
10
20
30
10
20
30
Cytheropteron elaeni *
10
20
12
30
0
11
200
620
0Age (cal. a)
50
100
150
Semicytherura complanata *
0
50
100
55
Total abundance, valves per 100 g
10Number of species
100200300
10
30
20
100
300
Brackishwater/100g
Euryhaline/100g
Deep-water/100g
Shallow-water/100g
55
5
PS
51/1
59-1
0, w
este
rn L
apte
v S
ea s
helf, K
hata
nga p
ala
eovalle
y (7
6.8
° N
, 116.0
° E
, w
ate
r depth
60 m
)
50
100Juvenile valves, %
500
10
10
10
0
Ostr
aco
ds (
va
lve
s 1
00
g )
-1
Fig
.5.
Ost
raco
dsp
ecie
sco
mpo
siti
onin
core
PS5
1/15
9-10
(Kha
tang
apa
laeo
valle
y):t
otal
abun
danc
e,ab
unda
nce
ofec
olog
ical
grou
psan
dsp
ecie
spe
r10
0g
dry
bulk
wei
ght,
dive
rsit
y,pe
rcen
tage
ofju
veni
leva
lves
.*N
ote
that
the
abun
danc
esof
spec
ies
Sem
icyt
heru
raco
mpl
anat
aan
dC
ythe
ropt
eron
elae
niar
esh
own
ona
smal
ler
scal
e.T
his
figur
eis
avai
labl
ein
colo
urat
http
://w
ww
.bor
eas.
dk.
Ostracod palaeoecology and environmental change in the Laptev and Kara seas, last 18 000 years 7BOREAS

01
00
20
00
40
00
60
00
80
00
10
00
0
12
00
0
14
00
0
16
00
0
18
00
0
Cytheropteron laptevensis
10
Cytheropteron paralatissimum
10Cluthia cluthae
10
20
30
Krithe glacialis
10
30
50
70
Cytheropteron pseudomontrosiense*1
03
05
0
Paracyprideis pseudopunctillata*
10Cytheropteron tumefactum
10Cytheromorpha spp.
10
30
50
Acanthocythereis dunelmensis*
10
20
Heterocyprideis sorbyana
10Bythocythere sp. 1
10Cytheropteron porterae
5Cytheropteron dimlingtonensis
5Sclerochilus sp. 1
10Cytheropteron biconvexa
10Roundstonia globulifera
5Cytheropteron nodosoalatum
5Cytheropteron occultum
5Cytheropteron inflatum
10Bythocythere constricta
10
20
30
40
Rabilimis mirabilis
10
20
Sarsicytheridea punctillata
10Pteroloxa sp.
5Elofsonella concinna
5Cytheropteron elaeni
10Cytheropteron arcuatum
10
20
Semicytherura complanata
5Cytheropteron montrosiense
10Cytheropteron discoveria
5Rabilimis septentrionalis
5Palmenella limicola
10Cytheropteron sulense
5Pseudocythere caudata
5Cytheropteron suzdalskyi
5Cytheropteron nodosum
5Cytheropteron perlaria
5Cytheropteron champlainum
5Eucythere spp.
5Loxoconcha spp.
5Sarsicytheridea spp.
5Semicytherura spp.
5Argilloecia spp.
5Cytheropteron spp.
10Polycope spp.
5Freshwater species
5Undetermined
1160
5400
17 2
00
100
15
10Brackishwater/100 g
50Euryhaline/100 g
50Shallow-water marine/100 g
50
100
Deep-water marine/100 g
5Freshwater /100 g
50100
Total abundance, valves per 100g
Number of species
Juvenile valves, %
PS
51
/15
4-1
1,
we
ste
rn L
ap
tev S
ea
co
ntin
en
tal slo
pe
(7
7.3
° N
, 1
20
.6°
E,
wa
ter
de
pth
27
0 m
)
Age (cal. a)
Ostr
aco
ds (
va
lve
s 1
00
g
)-1
Fig
.6.
Ost
raco
dsp
ecie
sco
mpo
siti
onin
core
PS5
1/15
4-11
(wes
tern
uppe
rco
ntin
enta
lslo
peof
the
Lap
tev
Sea)
:tot
alab
unda
nce,
abun
danc
eof
ecol
ogic
algr
oups
and
spec
ies
per
100
gdr
ybu
lkw
eigh
t,di
vers
ity,
perc
enta
geof
juve
nile
valv
es.H
ighl
ight
edis
the
inte
rval
ofa
sand
slid
eev
enta
tabo
ut15
ka.*
Not
eth
atth
eab
unda
nces
ofsp
ecie
sP
arac
ypri
deis
pseu
dopu
ncti
llata
,Cyt
hero
pter
onps
eudo
mon
tros
iens
ean
dA
cant
hocy
ther
eis
dune
lmen
sis
are
show
non
asm
alle
rsc
ale.
Thi
sfig
ure
isav
aila
ble
inco
lour
atht
tp://
ww
w.b
orea
s.dk
.
8 Anna Stepanova et al. BOREAS
10
20
30
40
50
Semicytherura complanata
01
02
03
04
05
06
07
00
10
00
20
00
30
00
40
00
50
00
60
00
70
00
80
00
Paracyprideis pseudopunctillata
10
20
30
Cytheropteron elaeni1
02
0
Cytheropteron suzdalskyi
10Roundstonia globulifera
10
20
Cytheromorpha macchesneyi
5Bythocythere spp.
5Semicytherura sp.
10Cytheropteron arcuatum
10
Palmenella limicola
01
0
Heterocyprideis sorbyana
5Cytheropteron dimlingtonensis
5
Sarsicytheridea punctillata
5Sarsicytheridea bradii
5Krithe glacialis
5Elofsonella concinna
5
Cytheropteron tumefactum
5
Cytheropteron biconvexa
10
20
30
Cluthia cluthae
10
Acanthocythereis dunelmensis
10
Rabilimis mirabilis
5Cytheropteron montrosiense
5Sclerochilus sp.
5Argilloecia spp.
5Cytheropteron porterae
5Cytheropteron champlainum
5Cytheropteron nodosoalatum
5Jonesia acuminata
Age (cal. a)B
P00/0
7-5
, easte
rn K
ara
Sea s
helf (
74.7
° N
, 81.1
° E
, w
ate
r depth
43 m
)
330
0
730
0
Total abundance, valves per 100 g
Number of species
Brackishwater/100 g
Euryhaline/100 g
Shallow-water marine/100 g
Deep-water marine/100 g
Juveniles, %
50
150
10
50
50
100
20
20
50
100
100
Ostr
acods (
valv
es 1
00 g
)
-1
Fig
.7.
Ost
raco
dsp
ecie
sco
mpo
siti
onin
core
BP
00/0
7-5
(eas
tern
Kar
aSe
ash
elf)
:tot
alab
unda
nce,
abun
danc
eof
ecol
ogic
algr
oups
and
spec
ies
per
100
gdr
ybu
lkw
eigh
t,di
vers
ity,
perc
enta
geof
juve
nile
valv
es.T
his
figur
eis
avai
labl
ein
colo
urat
http
://w
ww
.bor
eas.
dk.
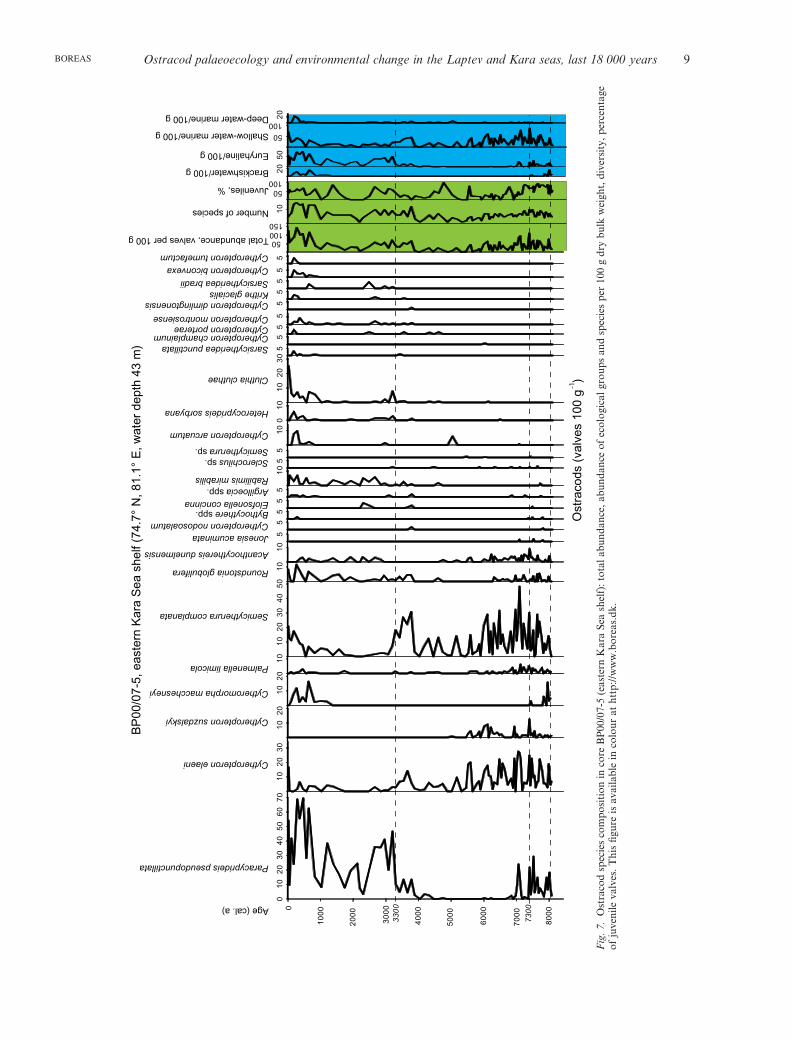
Ostracod palaeoecology and environmental change in the Laptev and Kara seas, last 18 000 years 9BOREAS

low-dominance, high-diversity assemblages and morestable conditions (Shannon & Weaver 1963).
According to the distribution of modern ostracods indifferent environments and their ecological preferences,primarily salinity and water depth, we divided ostracodspecies into six groups: freshwater, brackish water,euryhaline, marine shallow-water, marine relativelydeep water, and a specific group of marine relativelydeep-water North Atlantic species restricted toAtlantic-derived waters (Table S2; for details see alsoStepanova et al. 2007). Generally, these ecologicalgroups correspond to the following salinity ranges:marine relatively deep-water taxa are found in watersof >30 psu; marine shallow-water taxa in waters of>18 psu; euryhaline species tolerate a wide salinityrange between 5 and 35 psu; brackish-water speciescorrespond to the salinity range 5–26 psu; and, finally,freshwater species are found in waters with salinitylower than 7 psu (Stepanova et al. 2007).
Regional setting
Modern environments of the Laptev and Kara seas
The Kara and Laptev seas are marginal Arctic seaslargely represented by vast shallow shelves gentlysloping northwards down to water depths of 100 m. Aremarkable feature of the shelf topography in both seasis the presence of several palaeo-river valleys formedduring the Pleistocene sea-level lowstand and stret-ching towards the shelf break. These palaeovalleyswere the major areas of sediment accumulation duringthe Holocene transgression and contain evidence forthe transformation of the shelf from a terrestrial to amarine environment (Bauch et al. 1999, 2001; Dittmerset al. 2003; Stein et al. 2004).
Bottom-water temperature in the seas is below 0°Cnearly all year round. Only in the outer shelf troughsand at the continental slope does the temperature riseto 1.0–1.5°C owing to the inflow of warmer Atlantic-derived waters (Dobrovol’skii & Zalogin 1982; Pavlovet al. 1996). Atlantic surface waters enter the ArcticOcean north of Spitsbergen (Fram Strait BranchWater, FSBW) and through the Barents Sea troughs(Barents Sea Branch Water, BSBW), submerge beneathcolder polar waters and flow eastwards along the Eura-sian continental slope (Rudels et al. 2004; Fig. 1A). Inthe Laptev and Kara seas these waters remain at theslopes, although periodically reversed bottom currentsmay bring them onto the shelf along the river palaeo-valleys (Dmitrenko et al. 2001).
The seas are ice-covered for 9–10 months of the year,with a continuous fast ice cover bounding the shallowinner-shelf zone approximately along the 20–25 misobaths. Further to the north, in the middle shelf area,the fast ice edge is often separated from the drift iceby wind-induced open-water areas termed polynyas
(Pavlov & Pfirman 1995; Dmitrenko et al. 2000). Insummer, the seas are largely ice-free with the drift icemargin located far northwards, although in some yearsthe remnants of fast ice and pack ice can persist duringthe summer in the western Laptev Sea close to theTaimyr Peninsula (Reimnitz et al. 1995). The importantmechanism of sedimentation in the near-shore zone isthe resuspension of sediments by autumn storms andreversed currents, entrainment into the newly formed icein the freshened coastal waters, and ice-rafting to theseasonal drift ice margin in distant offshore regions(Eicken et al. 1997; Wegner et al. 2005).
The eastern parts of the Kara and Laptev seas aresubject to strong runoff influence by the Yenisei, Ob’and Lena rivers. As a result of the considerable riverrunoff in summer and ice formation in winter, salinityis the most variable feature affecting the spatial distri-bution of ostracods. Surface water salinities rangefrom less than 5 psu in the estuaries down to 28–30 psuin the river-distal areas. Because of the prevention ofvertical mixing by the low-density freshwater outflow,bottom-water salinities are less variable. They rangefrom 10–15 psu in the outer river estuaries in the KaraSea up to 34–35 psu on the outer shelf and upper con-tinental slope (Polyak et al. 2002; Stepanova et al.2003, 2007).
Recent ostracod assemblages from surface sediments
Modern ostracod assemblages from the Laptev andKara seas show large changes in composition withincreasing distance from the coast and decreasingriver influence as a result of changes in water depth,food supply and water mass properties (Table S2;Stepanova et al. 2003, 2007).
River estuaries of the Ob’ and Yenisei rivers areinhabited by freshwater species in their inner parts,where summer bottom water salinities are less than2 psu, and by a brackish water assemblage in theirouter parts, where salinity increases up to 26 psu.
The mixed euryhaline–marine assemblage is domi-nated by the euryhaline species Paracyprideis pseudop-unctillata, and Heterocyprideis sorbyana occurs in theinner-shelf zone of the Kara and Laptev seas down to20–30 m water depths, where bottom-water salinitiesrange between 26 and 32 psu.
A taxonomically diverse marine assemblage com-prises shallow-water marine taxa. These characterizenorthern parts of the Kara and Laptev shelves andupper continental slope with stable bottom environ-ments and a salinity higher than 32 psu.
In the middle-shelf zone, shallow-water marinespecies could reach up to 100%, while the relative abun-dance of euryhaline species and the total abundancedecrease.
Taxonomic diversity and percentages of relativelydeep-water species including the North Atlantic ones
10 Anna Stepanova et al. BOREAS

increase on the outer-shelf and upper continentalslope. Atlantic-derived waters influence the taxonomiccomposition of modern assemblages in the easternKara Sea and western Laptev Sea, but they do notreach the eastern Laptev Sea. We found North Atlan-tic species only at sites from the Kara Sea and fromthe Khatanga valley of the Laptev Sea (Stepanovaet al. 2003, 2007).
Abundant euryhaline species that dominate theinner-shelf assemblage are also found at greater waterdepths, where they are identified as part of an ice-raftedassemblage (Cronin et al. 1994; Jones et al. 1998, 1999;Stepanova et al. 2003, 2007). They are possiblyentrained into the newly formed ice during autumnstorms and the freeze-up period and transported to thedistal open-sea areas (Nürnberg et al. 1994; Eickenet al. 1997).
Fossil ostracod assemblages from marinesediment cores: abundance, taxonomyand age ranges
Six fossil assemblages have been distinguished in thestudied cores: Estuarine brackish water (1), Inner-shelfeuryhaline–marine (2), Middle-shelf shallow-watermarine (3), Outer-shelf marine (4), Pre-Holocene uppercontinental slope marine (5), and Holocene upper con-tinental slope marine (6a, b). A sequence of assem-blages from estuarine to modern normal marine ones(assemblages 1–4 in Figs 3–5, 7–9) is observed in allstudied sediment cores from palaeoriver valleys on themiddle-outer shelf, namely PS138, PS135, PS159 andBP07. Relatively deep-water ostracod assemblages 5and 6 from the deepest core PS154 on the western uppercontinental slope of the Laptev Sea (Figs 6, 8, 9) reflecta transition from a glacial-like near shore environmentto a modern offshore relatively deep-water marine one.In Table S3 we list the most abundant and ecologicallycharacteristic taxa for all six assemblages and give their(palaeo)ecological interpretation together with taxo-nomic notes based on our own data and publishedevidence.
Some minor disadvantages of studying ostracods inshelf–upper slope environments arise from the fact thatthey usually occur in lower abundance compared with,for instance, benthic foraminifers, and that in theseenvironments they may represent mixed faunas withallochthonous elements. Marginal marine environ-ments are known for a number of post-mortem tapho-nomic processes (Boomer & Eisenhauer 2002) and theycommonly contain allochthonous elements either fromland or from open-marine habitats. It is important todistinguish between these elements in the assemblage(Whatley 1983; Boomer & Eisenhauer 2002; Frenzelet al. 2010). For the Kara and Laptev seas, staining ofsurface samples with Rose Bengal showed that most
samples contain 90–100% of live organisms, and at thesame time they have a juvenile portion of about25–50%, which implies washout of valves of dead andsmaller juvenile valves (Whatley 1983; Stepanova et al.2007). In the studied core samples the preservation ofvalves is generally good, and most samples hold a juve-nile percentage of 25–50%; this indicates either a high-energy environment from which smaller juvenile oradult dead valves were washed out or a thanatocoenosiswith current velocities sufficient to carry larger valves(Whatley 1983). A direct comparison with modern shelfassemblages with a high ratio of live organisms andsimilar juvenile percentage (25–50%) convinces us of theautochthoneity of fossil assemblages and high-energyenvironments. Another pattern is periodically observedin assemblage 5 on the upper continental slope core.High fluctuations of juvenile ratio of up to 100% corre-spond to periodic sudden events, such as slides, whenostracods were overwhelmed and entombed in thesediment (Whatley 1983).
Shelf cores
Lithologically, all shelf cores obtained from palaeorivervalleys (see ‘Modern environments of the Laptev andKara seas’) represent relatively uniform sedimentsequences composed largely of bioturbated marine siltyclay to clayey silt, with the weight percentage of thecoarse fraction (>63 mm) not exceeding 4–5%, exceptfor the middle part of core PS138 where it rises to10–20% (Stein et al. 2004; Taldenkova et al. 2005,2008).
Although the general trend in ostracod assemblageevolution from estuarine to modern normal marine(assemblages 1–4) is the same for all studied shelf cores(Table S3; Figs 8, 9), there are certain differencesbetween similar assemblages from different locations.Therefore, our attention was also drawn to the corre-lation between variations in the taxonomic composi-tion of assemblages and geographical locations of thecores.
At the end of the Late Pleistocene and in the earlyHolocene, as a result of the fast sea-level rise and pro-ximity of the coastline, accumulation rates in the riverpalaeovalleys on the outer shelf were high (Bauch et al.1999, 2001; Taldenkova et al. 2005, 2009), and environ-mental changes were fast and abrupt, as indicated bythe succession of several fossil ostracod assemblages inthe studied shelf cores. None of these cores reveals atransition from terrestrial to marine deposits. Thelower age of the oldest estuarine brackish-water assem-blage (1), which is present in all cores from the LaptevSea, ranges in accordance with the water depth of thesites, from 12.3 ka in core PS159 (60 m), to 11.4 ka incore PS135 (51 m) and to 11.3 ka in core PS138 (45 m).The estimated duration of the estuarine phase is uncer-tain and ranges from 200 to 1000 years (Fig. 8). In the
Ostracod palaeoecology and environmental change in the Laptev and Kara seas, last 18 000 years 11BOREAS

Kara Sea core this assemblage is missing, as the age ofthe oldest studied sediments is 8 ka.
Assemblage (1) is characterized by a low taxonomicdiversity (Fig. 8). It is represented mainly by euryha-
line and brackish water species, as well as by someshallow-water marine ones (Table S3; Figs 3–5, 8, 9).Total abundance is relatively low, on average 10–40specimens per 100 g of sediment (Figs 3–5). The
Fossil assemblages
inner-shelf, marineeuryhaline-2
estuarine, brackish water1
middle-shelf, marineshallow-water3 Holocene upper continental slope, marine6
outer shelf, marine
Pre-Holocene upper continental slope, marine
4
5
Laptev SeaKara Sea
outer-shelfPS159 (60 m)
Western
upper continentalslope PS154
(270 m)
outer-shelfPS135 (51 m)
middle-shelfPS138 (45 m)
Eastern
middle-shelfBP07 (43 m)
11 200
10 300
1
2
3
11 400
6200
6
12 000
11 2002
4
3
12 3001
6200
Age (
cal. a
)
4
3
5
Eastern
8
0
0
0
1
0
0
0
0
3300
0
2000
4000
6000
8000
12 000
14 000
16 000
10 000
2
10 200
1
2
3
11 300
8200
0
2000
4000
6000
8000
12 000
14 000
16 000
10 000
8000
7300
0.5
1.5
2.5
Shannon-Weaver diversity
10 000
1160
18 000
17 200
1650
18 000
0.5
1.5
2.5
1.5 2.50.5
0.5
1.5
2.5
0.5
1.5
2.5
500
Fig. 8. Comparison of fossil ostracod assemblage evolution in different parts of the Laptev and Kara seas based on species diversity definedby the Shannon–Weaver index.
12 Anna Stepanova et al. BOREAS

Fig. 9. Comparison of the changes in the relative representation of different ecological groups during the Postglacial sea-level rise in differentparts of the Laptev Sea. North Atlantic species belong to the group of deep-water species, but their percentage is also shown separately in orderto highlight the periods of enhanced influence of Atlantic-derived waters at the continental slope. LG=Lateglacial; B-A=Bølling-Allerød;YD=Younger Dryas; EHol=early Holocene; MHol=middle Holocene; LHol=late Holocene. This figure is available in colour athttp://www.boreas.dk.
Ostracod palaeoecology and environmental change in the Laptev and Kara seas, last 18 000 years 13BOREAS

Shannon–Weaver diversity index is generally slightlylower for sediments older than 10 ka, averaging 0.5–1in all cores from the Laptev Sea shelf (Fig. 8). Thepercentage of juvenile valves in lower parts of the coresaverages 25–50%. According to Whatley (1983), thismay imply a high-energy environment. Altogether,this indicates a harsh and unstable environment of theestuarine zone with very strong freshwater influence.
Assemblage (1) is replaced upcore by assemblage (2),which is practically devoid of any brackish water taxa.This transition occurred at around 12 ka in core PS159,at 11.2 ka in core PS135, and at 10.2 ka in core PS138(Fig. 8).
The inner-shelf, euryhaline–marine assemblage (2) istaxonomically more diverse than assemblage (1),reflecting a gradual transition to normal marine condi-tions (Table S3; Figs 3–5, 7–9). It is dominated byshallow-water marine species and the euryhalinespecies P. pseudopunctillata. In cores PS159 and BP07from the west some relatively deep-water species arealso present (Figs 5, 7). They reach up to 30–50% incore PS159 (Fig. 9) and represent Arctic (Krithe glacia-lis, Argilloecia spp., Rabilimis mirabilis) as well asNorth Atlantic (Cytheropteron biconvexa, C. porterae,Bythocythere spp.) taxa (Table S3). The high percent-age of relatively deep-water Arctic and North Atlanticspecies in the early Holocene inner-shelf assemblagecould be explained by the proximity to the shelf breakand the occasional penetration of Atlantic-derivedwaters from the west into the shelf palaeovalleys withwind-induced reversed currents (Dmitrenko et al.2001). The percentage of juvenile valves in the easternpart of the Laptev Sea is very low, less than 25%, whilefarther west and in the Kara Sea it is 50–80%. Lowjuvenile ratios in the east reflect low total abundanceand indicate a high-energy environment. In the westernLaptev Sea, the higher ratios evidence a lower energyenvironment, while in the Kara Sea, we probablyobserve a life assemblage with up to 80% represented byjuveniles of different instar stages (Whatley 1983).
The average total abundance of ostracods is low,fewer than 50 specimens per 100 g of sediment(Figs 3–5, 7, 9). The diversity index is highly variableand ranges between 0 and 1.5 (Fig. 8). Similar to itsmodern analogue from surface sediments (Stepanovaet al. 2003, 2007), the inner-shelf assemblage character-izes river-affected coastal regions down to depths of20–30 m, with considerable seasonal changes includingfast ice cover in winter and the formation of a thicksurface-freshened water layer in summer owing to riverrunoff and ice melting.
The time duration of the existence of assemblage (2)ranges from 800 to 900 years in cores PS159 and PS135,to 2000 years in core PS138 (Fig. 8). This regional dif-ference could be attributable to the stronger fresheninginfluence of the Lena River discharge compared withthe Khatanga and Yana rivers. In core BP07, the age of
the initiation of this assemblage is unknown, but theduration of its existence exceeds 700 years.
A relatively poor middle-shelf shallow-water marineassemblage (3) occurs in all four studied cores from theLaptev and Kara shelves. It is dominated by variousshallow-water marine species, while euryhaline andbrackish-water species are rare or absent (Table S3). Inthe eastern Laptev Sea, the transition to assemblage 3 ismarked by the increasing proportion of shallow-watermarine species and indicates the establishment ofmodern-like conditions (Figs 3, 4). Taxonomic differ-ences within assemblage (3) from the studied cores aredefined largely by the distribution of Atlantic-derivedwaters in the study area. Unlike the case for the easternLaptev Sea, relatively deep-water species includingNorth Atlantic species occur in the western Laptev andeastern Kara sea shelf sediments and include some ofthe taxa found in the underlying sediments (assemblage2) together with some other Arctic (Sarsicytherideapunctillata) and North Atlantic (Cytheropteron arcua-tum) forms. These species may indicate an influence ofAtlantic-derived waters, probably brought via reversedbottom currents to these two sites during the Holocene.
The total abundance is in general low, fewer than 50specimens per 100 g of sediment. However, we observethe evident increase in ostracod abundance during thetransition to middle-shelf conditions, which is mostpronounced in the continuously studied core PS159,where the peak total abundance of 250 specimens per100 g of sediment is recorded at 10.2 ka (Figs 7, 9). Thispeak abundance suggests a high seasonal productivityand probably a short time period when the core sitewas located at the fast sea-ice margin. Juvenile ratiosrise in the east, compared with the underlying layer,to 25–50%, and fluctuate a lot in the west, rangingbetween 20 and 100%. Such a distribution of juvenileratios reflects the stabilization of environments in theeast and fluctuating parameters in the west, such asAtlantic water inflow and food supply. The diversityindex for assemblages 2–3 is less than 1.5. In general,the tendency in all cores is towards an increase ofdiversity upcore, which indicates a stabilization of theenvironments.
In the western cores PS159 and BP07, assemblage (3)is replaced upcore by the outer-shelf marine assemblage(4), at 7.2 ka in the Khatanga palaeovalley and at3.3 ka in the eastern Kara Sea shelf (Figs 5, 7–9). Theassemblages from the two cores are almost identical,although the core from the Kara Sea actually comesfrom a shallower environment than PS159. The taxo-nomic composition of assemblage (4) is somewhat con-tradictory. On one hand we observe an increase oftaxonomic diversity owing to the introduction of rela-tively deep-water species, including North Atlanticforms (Table S3; Figs 5, 7). On the other hand, wenotice a significant increase in euryhaline species,primarily P. pseudopunctillata (Table S3). In fact,
14 Anna Stepanova et al. BOREAS

this represents the second, recurring introduction ofthis species, as it almost disappeared in the previousassemblage (3).
The total abundance in core PS159 is fewer than 50specimens per 100 g of sediment, but abruptly risesto 250 specimens in the uppermost layers, possiblybecause of the better preservation of valves (Figs 5, 9).In the Kara Sea, this assemblage is characterized by anincreased total abundance of up to 80 specimens per100 g of sediment (Fig. 7). The juvenile ratio in bothcores averages 25–50% and indicates a rather high-energy, modern-like environment (Figs 5, 7). TheShannon–Weaver diversity index for assemblage 4 isrelatively high (1.5–2), reflecting water-depth increaseand the increasing influence of Atlantic-derived waters.
Western Laptev Sea upper continental slope
Owing to its deeper location on the western upper con-tinental slope of the Laptev Sea, core PS154 (270 m) isdistinguished from the other studied shelf cores. Corelithology together with the occurrence of ice/iceberg-rafted debris (IRD) and authigenic concretions are dis-cussed in Taldenkova et al. (2008, 2010). The coresequence younger than 15 ka consists largely of biotur-bated clayey silt. Sandy lenses occur in the older partof the core, with a prominent sandy layer (up to 70 wt%of >63-mm fraction) at about 15 ka. IRD peaks arerecorded in the oldest (>c. 16.2 ka) and youngest(<7 ka) parts of the core. Prior to shelf flooding, thissite was located relatively close to the coast at a dis-tance of about 50 km (Taldenkova et al. 2008). Water-mass distribution and topography of the sea floor weredifferent from those on the shelf, and thus the fossilassemblages (5) and (6) recognized here (Figs 6, 9) havedifferent components compared with the shelf cores.
The oldest ostracod-bearing sample at 673 cm coredepth dates back to c. 17.2 ka and contains only fourvalves belonging to Cluthia cluthae, Cytheropteronlaptevensis and C. paralatissimum. The overlying nearly1-m-thick sediment sequence is barren of ostracods.Ostracods are constantly present since c. 15.6 ka.
The taxonomic composition of the oldest pre-Holocene upper continental slope assemblage (5)(Table S3; Figs 6, 8, 9), dating back to 15.6–10 ka, issimilar to that of assemblage (4), but has many distin-guishing features. It includes two deep-water Arcticspecies (Krithe glacialis, Rabilimis mirabilis), which areespecially abundant around 15.6–15 ka, when fewNorth Atlantic species are present. Various NorthAtlantic species are constantly present, and becomequite abundant after c. 15 ka, evidencing the continu-ous presence of Atlantic-derived waters. We presumethat the bottom-water temperature did not change sig-nificantly, as even at present Atlantic-derived watersarrive in the Laptev Sea considerably chilled, down to1°C northwards of New Siberian Islands (Dmitrenko
et al. 2010). The highest relative abundances of NorthAtlantic species, implying the strongest inflows ofchilled Atlantic-derived waters, are observed at 15–14.5and 11.5–10 ka (Fig. 9). The second and third mostabundant ecological groups of ostracods in assemblage(5) are represented by two euryhaline species andshallow-water middle-shelf species (Figs 6, 9). At about15 ka, an evident slide event is suggested by a sedimentunit with a 70% sand fraction (Taldenkova et al. 2008,2010). This sediment layer contains valves of brackishwater and even freshwater ostracod species (Table S3;Fig. 6). Periodic smaller peaks in the occurrence ofeuryhaline and brackish-water species might be indi-cative of smaller, unidentified slide events and/orenhanced freshwater influence.
The total abundance within this assemblage is dis-tributed very irregularly, but is typically fewer than 50specimens, with several peaks of up to 120 specimensper 100 g of sediment at about 14.5, 13 and 12 ka(Figs 6, 9). Juvenile ratios exhibit high-amplitude fluc-tuations of 0–100%, giving additional evidence for ahigh-energy environment and high mortality ratesowing to catastrophic slide events (Fig. 6). TheShannon–Weaver diversity index is higher (1–2) in sedi-ments older than 10 ka (Fig. 8), reflecting a highly vari-able total abundance and taxonomic diversity causedby an active hydrodynamic regime, the proximity of thecoast, and temporal catastrophic events such as slides.The exception is the oldest time interval of this section(15–15.6 ka), when the diversity index is below 1, andthe ostracod association is composed almost entirely ofK. glacialis.
The transition to the Holocene upper continentalslope marine assemblage (6) occurred at about 10 ka.The overall composition of this assemblage is less taxo-nomically diverse than that of assemblage (5) owing tothe absence of many shallow-water taxa (Table S3;Fig. 6). It was subdivided into two intervals: 10–5.4 ka(sub-assemblage 6a) and 5.4–2 ka (sub-assemblage 6b).The first sub-assemblage is dominated by the relativelydeep-water species R. mirabilis and the North Atlanticspecies Cytheropteron tumefactum, and some shallow-water marine taxa are also present (Cytheropteronsuzdalskyi and C. montrosiense). The second sub-assemblage is characterized by a combination of thedeep-water marine species R. mirabilis, K. glacialis andSarsicytheridea punctillata, the euryhaline species Het-erocyprideis sorbyana and the North Atlantic taxonC. tumefactum. Other species are present in minor quan-tities. Sub-assemblage 6b is very similar to the modernostracod assemblage of this area (Stepanova et al. 2003).The total abundance of ostracods here is slightly higherthan in the underlying assemblage and averages 30–60specimens per 100 g of sediment (Fig. 8). Juvenile ratiosare very low in sub-assemblage (6a), reflecting theoverall low total abundance. They rise to 10–50% insub-assemblage (6b) and indicate the establishment of a
Ostracod palaeoecology and environmental change in the Laptev and Kara seas, last 18 000 years 15BOREAS

modern-like environment. The Shannon–Weaver diver-sity index varies between 1.5 and 2 for assemblage (6),indicating stabilization of the environment.
Besides the possibility of ice-rafting from the innershelf areas (see ‘Recent ostracod assemblages fromsurface sediments’), the abundance of euryhalinespecies in assemblage (6) can be explained by thehigh tolerance of these species to environmentalchanges (Figs 6, 9). The range of their salinity/depth/temperature/nutrition requirements is very wide whencompared with purely marine species. That is why wefind euryhaline species abundant both in shallowinner-shelf environments and on the outer shelf /uppercontinental slope.
Reconstruction of past environmental changesbased on ostracod assemblage analysis
Whereas there are several investigations of Quaternarydeep-sea ostracod assemblages from the Central ArcticOcean (Cronin et al. 1994, 1995, 2010), records fromArctic shelves are rare. So far, the most detailed data onfossil ostracods from Arctic shelf sediments come fromborehole sections in northern Alaska (McDougall et al.1986). However, that investigation lacks sufficient agecontrol, as only three radiocarbon dates were availablefor these up to 8-m-long borehole sediment sequences.In this respect, our data on the timing and character ofostracod assemblage variations in the Laptev and Karaseas represent the first record that allows both the trendand the rate of environmental changes in these mar-ginal Arctic seas to be addressed under the last postgla-cial sea-level rise.
Lateglacial and deglaciation (c. 17.6–11 ka)
Siberian shelves to the east of Taimyr Peninsula aswell as the southeastern Kara Sea were sub aeriallyexposed during the Last Glacial Maximum (LGM). Atthis time, an ice sheet partly covered the Kara Seashelf and the vast territory farther west, affecting theinfluence of North Atlantic water on this region(Svendsen et al. 2004; Fig. 1A). Sea level during theLGM was about 120 m lower than modern valuesprior to the shelf flooding (Fairbanks 1989), and theLaptev Sea upper continental slope was located veryclose to the palaeocoastline, giving rise to conditionscharacterized by a cold-water marine environmentsubject to meltwater and fluvial influence (Bauch et al.2001; Spielhagen et al. 2005; Taldenkova et al. 2008,2010). Enhanced meltwater inputs resulted in strongwater stratification, as evidenced by abundant authi-genic concretions of vivianite and rhodochrositeformed under anaerobic conditions and the generalabsence of benthic remains during the time periodbetween 17.6 and 15.4 ka (Taldenkova et al. 2010).
At the same time, ostracod species with North Atlan-tic affinities were identified here as early as c. 17.2 ka,giving clear evidence for the presence of Atlantic-derived waters in the Arctic during lateglacial times. Itfurther implies that ostracods were able to settle andsurvive in the harsh environments of the early deglacialperiod with considerable meltwater input and strongwater stratification. Scarce findings of ostracods arerecorded simultaneously with extremely high abun-dances of small-sized subpolar planktic foraminifersand the occurrence of some benthic foraminiferalspecies, including the species Cassidulina neoteretisindicative of Atlantic-derived waters (Taldenkova et al.2010). Rare subpolar planktic foraminifers were iden-tified in the sediment sequence interval of c. 17.6–15.6 ka, giving evidence for subsurface inflows ofAtlantic-derived waters to the site, but the event at c.17.2 ka probably corresponds to enhanced upwelling ofAtlantic-derived waters in a coastal polynya main-tained in winter by offshore winds from the Siberianmainland (Taldenkova et al. 2008, 2010).
The continuous presence of North Atlantic species inthe record since c. 15.6 ka suggests that the westernLaptev Sea slope was constantly under the influence ofthe subsurface inflow of Atlantic-derived waters(Figs 6, 9). The most abundant occurrence of thesespecies corresponds to the transition from the Lategla-cial to the Bølling–Allerød period (Fig. 9). It furtherindicates a strong penetration of subsurface inflow ofAtlanic-derived water along the Eurasian continentalslope, as is also supported by the occurrence of thehighest relative abundances of C. neoteretis in corePS154 (Taldenkova et al. 2008, 2010); the latter arecomparable to the reported values of this speciesaround Svalbard (Slubowska et al. 2005; Slubowska-Woldengen et al. 2007, 2008).
The total abundance of ostracods within assemblage(5) is distributed very irregularly, with several peaksfound at about 14.7–14.5, 13 and 12 ka (Figs 6, 9).These peaks may correspond to enhanced surface-water productivity owing to climate amelioration and astronger fluvial influence possibly resulting in surface-water warming. Interestingly, the maximum totalabundance of ostracods as well as benthic foraminifersin core PS154 centred at 13 ka corresponds to the localfreshwater event indicated by the low d18O spike inplanktic foraminifers identified from the eastern LaptevSea continental slope by Spielhagen et al. (2005). Incore PS154, the abovementioned abundance peaks arelargely represented by shallow-water marine andeuryhaline/brackish-water taxa. This confirms the pos-sibility of enhanced freshwater influence at the expenseof the open-sea water masses, including Atlantic-derived waters (Figs 6, 9). Unlike the 15-ka slide event(Fig. 6), when the valves of freshwater, brackish waterand euryhaline ostracods were probably displaceddownslope from a nearby coast, the younger deglacial
16 Anna Stepanova et al. BOREAS

peaks of brackish water and euryhaline species at 13and 12 ka after the onset of shelf flooding are a result ofice-rafting processes. Valves of these species can beentrained into the newly formed ice in the freshenednear shore zone during autumn storms and then trans-ported to the seasonal drift-ice limit (cf. Eicken et al.1997). The fact that these valves were abundant in corePS154 during deglacial times may imply that thesummer drift-ice margin was located close to the site.Because the latter was still not far from the formercoastline prior to extensive shelf flooding in the earlyHolocene, it would mean that the overall climate con-ditions remained relatively cool at this time.
On the outer shelf, the transition into the earlyHolocene (12.3–11 ka) was marked by rapid inunda-tion and the occurrence of brackish-water ostracodassemblages typical of unstable estuarine and inner-shelf environments (Stepanova 2006). The studiedsites were located in river estuaries at water depths ofabout 5–10 m and with a bottom-water salinity ofaround 18–20 psu (Taldenkova et al. 2005). In thewestern Laptev Sea, deep-water ostracod species andspecies with North Atlantic affinities were alsopresent, where they accounted for up to 50% ofassemblage (2) in core PS159 (Fig. 9). We supposethat the deeper water species were transported to shal-lower sites with wind-induced reversed bottom cur-rents (Dmitrenko et al. 2001; Wegner et al. 2005).None of these ostracod taxa were found on theeastern Laptev Sea shelf (Figs 3, 4, 9), probably owingto the dominance of fresh water discharged from theLena River. It is noteworthy that at about 11.3 kaseveral planktic foraminiferal species were found incore PS135 (Taldenkova et al. 2005), which probablymeans that the Yana River runoff was less strongthan that of the Lena around this time.
Early–middle Holocene (11–6 ka)
On the western continental slope, the major changefrom a fluvial-affected to modern marine environmentoccurred at around 10 ka. This change was accompa-nied by a sharp reduction in sedimentation rates fromabout 120 to 5 cm ka-1 owing to the onset of extensiveshelf flooding (Fig. 2; Bauch et al. 2001), and resulted ina less good preservation of ostracod valves, of whichonly a few are found between 9 and 7 ka (Figs 6, 9).Because the number of shallow-water and euryhalineostracods is clearly lower, their absence on the conti-nental slope may indicate climate warming with reducedsea-ice cover in summer, and a northward displacementof the seasonal drift-ice margin. Because the sea-icemarginal zone is known to be an area of high seasonalproductivity (Polyak et al. 2002), a spatial change couldhave contributed to the drastically diminished sedimen-tation rates. The offshore shift of the sea-ice marginalzone is supported by the almost complete absence of
ice-rafted debris in the sediments of core PS154 duringthe early Holocene (Taldenkova et al. 2010).
As is evidenced by the high percentage of NorthAtlantic species, a continuous influence of Atlanticwater is observed on the western Laptev Sea slopeduring the early Holocene (Fig. 9). These data gener-ally agree with other interpretations indicating anenhanced influence of modified Atlantic watersbetween 11 and 9 ka (e.g. Stein et al. 2001, 2004). Awarmer climate at this time has also been reconstructedby various authors using proxy records for terrestrial(MacDonald et al. 2000; Naidina & Bauch 2011),coastal marine (Duplessy et al. 2001; Lubinski et al.2001; Ivanova et al. 2002) and deep Arctic Ocean(Cronin et al. 2010) environments.
However, the taxonomic diversity of Atlantic water-indicative ostracod species was higher during deglacialtimes than in the early Holocene. Certain species of thisgroup (Pseudocythere caudata, Cytheropteron dimling-tonensis, C. nodosoalatum, C. paralatissimum, C. bicon-vexa) disappeared from the record around 10–11 ka,whereas other species (C. tumefactum, C. occultum,C. pseudomontrosiense, C. arcuatum) were continu-ously present also in the Holocene record, and the onlynewly invading North Atlantic species was C. perlaria(Fig. 6). Slightly earlier, at around 12 ka, the Atlanticwater-indicative benthic foraminifer C. neoteretisalso disappeared from the record of core PS154, onlyto re-appear in the mid-Holocene at about 5 ka(Taldenkova et al. 2008, 2010). The observed changesin the taxonomic composition of microfossils in theearly Holocene might possibly be related to the vari-ability in the modified Atlantic water masses enteringthe Laptev Sea (Fig. 1A); that is, to a possibly strongerinfluence of the BSBW after the flooding of the for-merly glaciated over-deepened Barents Sea shelf(Lubinski et al. 2001) at the expense of the FSBW.
On the shelf, changes in ostracod assemblage com-position, which are similar in all studied cores fromboth the Laptev and the Kara seas, reflect rapid trans-gressive inundation of the region and the transitionfrom estuarine–brackish-water to normal marine con-ditions (Figs 3–5, 7–9; Bauch et al. 2001; Taldenkovaet al. 2005, 2008, 2009). Age–depth relationship plotsfor the studied cores (Fig. 2) show that there is anevident drop in sedimentation rates owing to the south-ward migration of coastline and major depocentres(Bauch et al. 2001) that occurred, depending on the sitelocation and water depth, between 10.2 and 8.2 ka(Figs 1, 2). In core BP07 from the Kara Sea, sedimen-tation remained steady and relatively high during thelast 8 ka, because the core spans a shorter time intervalthan the other shelf cores, and also the site is locatedin a sea-floor depression that was gradually filledwith sediments during the Holocene. Along with thedrop in sedimentation rates in the western Laptev Seacore PS159, relatively deep-water species that were
Ostracod palaeoecology and environmental change in the Laptev and Kara seas, last 18 000 years 17BOREAS

abundant during early stages of inundation becamealmost absent later (Figs 5, 9). Between 10.5 and 6 kathey gave way to shallow-water marine species notwith-standing the water depth increase (Fig. 9). This couldbe due to the relatively weak estuarine-like water circu-lation on the shelf in the early Holocene when thestrength of reversed bottom currents was probablyreduced. An increase in the percentage of the euryha-line species P. pseudopunctillata between 9 and 6 ka inthe eastern Laptev Sea (Figs 3, 4, 9) might be explainedby the enhanced influence of river runoff from the Lenaand Yana initiated by climate warming.
Late Holocene (6 ka–recent)
Because global and regional sea level stabilized ataround 5–6 ka (Fairbanks 1989; Bauch et al. 1999,2001), the ostracod assemblages at all studied sitesacquired their modern-like composition at this time(Figs 3–7, 9; Stepanova et al. 2003, 2007; Stepanova2006).
On the western upper continental slope and the outershelf of the Laptev Sea, the youngest ostracod assem-blage is characterized by a significant amount ofshallow-water and euryhaline species co-occurring withdeep-water species, including the North Atlantic ones(Table S3; Figs 6, 9). Given the greatest distance fromthe coast and water depths since the beginning of therecord at about 17.6 ka, the re-introduction of ostracodspecies characteristic of inner-shelf environments to theouter shelf and upper continental slope must be linkedprimarily to rafting by sea ice and probably also byicebergs, which implies climate cooling resulting inthe onshore shift of the seasonal drift-ice limit andthe growth of local ice caps (e.g. Severnaya Zemlya).The latter assumption is supported by a considerableincrease in IRD in cores PS154 and PS159 since 7.2 ka,pointing to this local source of iceberg-rafted material(Taldenkova et al. 2010). Interestingly, the high-resolution record of IRD in core PS159 shows a sub-millennial periodicity with the peaks centred at about7.2, 6.4, 5.4, 3 and 2 ka (Taldenkova et al. 2010), whichis roughly similar to the timing of the peaks in euryha-line ostracod species abundance (Fig. 9).
Not only in the cores from the western Laptev Seaouter shelf and slope (influenced by iceberg-rafting fromSevernaya Zemlya) does the percentage of euryhalinespecies increase, it is also true for the Kara Sea mid-shelfcore BP07 and cores from the eastern Laptev Sea outerand mid-shelf. We assume that this points to a south-ward shift of the summer drift-ice limit owing to climatecooling. As a result, more ostracod valves from thenear-shore regions that had been entrained in the newlyformed ice during autumn storms and then transportedto the drift-ice margin (cf. Eicken et al. 1997) later foundthemselves released into the sediments of the outershelf and upper continental slope rather than being
transported farther offshore to the Arctic Ocean andNorth Atlantic. The increase in the reintroduced near-shore species is especially pronounced in core BP07(Fig. 7). Here, the share of euryhaline P. pseudopunctil-lata in the upper part of the core (assemblage 4) is evenhigher than in the lowermost assemblage (2), whichcorresponds to an otherwise more fluvially affected envi-ronment, as is also clearly indicated by the lower valuesof the d18O composition of calcareous microfossils fromthe same core (Simstich et al. 2004, 2005).
In general, the tendency in all studied cores from theshelf is towards the increase of diversity upcore, whichindicates stabilization of environments, depth increaseand the establishment of diverse and stable ecolo-gical niches during the course of the Holocene. TheShannon–Weaver diversity index remains relativelylow for all identified assemblages. For late Holoceneostracod assemblages it ranges between 1.5 and 2 forassemblages (3)–(4) (outer shelf) and further decreasesfor the upper continental slope assemblage (6) (from1.5–2 down to 0) (Fig. 8). Such a difference may beexplained by variable nutrient supply. The latter, prob-ably, decreased on the continental slope owing to thecombination of the considerable distance from thecoastline and the southward migration of the seasonaldrift-ice margin induced by cooling.
Conclusions
The late Pleistocene–Holocene shelf history of theLaptev and eastern Kara seas is reconstructed based onostracod assemblages from four sediment cores recov-ered from mid outer shelves and a core from thewestern Laptev Sea upper continental slope. The fossilrecords show large faunal changes associated withrapid inundation of the shelves and coastal retreatduring postglacial sea-level rise, together with variabil-ity in freshwater runoff. In sediments younger than7 ka we observe climatic-driven rather than sea-level-driven water-mass changes, as reflected in the increasedrepresentation of ice-rafted euryhaline taxa in other-wise modern-like assemblages, along with an increasingIRD content.
Ostracod assemblage analysis indicates three majorperiods of environmental change characterized by sixecologically different assemblages: estuarine brackish-water assemblage (1), inner-shelf euryhaline–marineassemblage (2), middle-shelf shallow-water marineassemblage (3), outer-shelf marine assemblage (4), pre-Holocene upper continental slope marine assemblage(5), and Holocene upper continental slope marineassemblage (6a, b):
• The Lateglacial and deglaciation (c. 17.6–11 ka) arecharacterized by assemblage (5) on the upper con-tinental slope of the Laptev Sea, indicative of a
18 Anna Stepanova et al. BOREAS

cold-water marine environment strongly influencedby both a subsurface inflow of Atlantic-derivedwater and riverine freshwater input. Ostracods withNorth Atlantic affinities occur as early as c. 17.2 kaand are continuously present after c. 15.6 ka, indi-cating that Atlantic-derived water upwelled to thesite facilitated by coastal polynyas and offshorewinds. After about 14.7 ka, the increased totalabundance of ostracods with peaks at 14.7–14.5, 13and 12 ka probably reflects enhanced surface-waterproductivity as a result of climate amelioration, agrowing fluvial influence and offshore shifts in theposition of the seasonal sea-ice marginal zone. Onthe outer shelf, this period (starting from 12.3 ka) ischaracterized by the transition from estuarinebrackish water (with water depths of about 5–10 mand bottom-water salinities of 18–20 psu) to inner-shelf environments.
• The early–middle Holocene (11–6 ka) is dominatedon the western upper continental slope of theLaptev Sea by sub-assemblage (6a), introduced ataround 10 ka and indicative of climate warming,reduced sea-ice cover and the continuous presenceof Atlantic-derived waters. On the shelves, assem-blages (2) and (3) indicate a rapid inunda-tion process and the transition to normal marineconditions.
• The late Holocene (6 ka–recent) corresponds tosub-assemblage (6b) on the western upper continen-tal slope of the Laptev Sea, and to assemblages (3)and (4) on the shelves. They reflect the stabilizationof depositional environments and the establishmentof modern-like conditions. The simultaneousincrease in the percentage of shallow-water marineand euryhaline species recorded in all cores suggeststhat these species are mostly sea-ice- and/or iceberg-rafted. This observation probably provides evi-dence for a cooling trend that was accompanied bythe southward migration of the seasonal sea-icemargin and the regional re-growth of ice caps.
Acknowledgements. – This research was supported by the BMBF andRussian Ministry for Education and Science (OSL fellowshipprojects) and the Russian Foundation for Basic Research (projects08-05-00849 and 11-05-01091). It is a contribution to the APEX(Artic Paleoclimate and its Extremes) program. We are grateful to theIntegrated Ocean Drilling Program at Texas A&M University forhosting the first author and providing office support and analyticalinstrumentation during the completion of this paper. The paper ben-efited greatly from comments made by T. M. Cronin (USGS) on anearlier version. The helpful remarks and suggestions of E. M. Brou-wers (USGS) and the two anonymous reviewers are very muchappreciated.
References
Alvarez Zarikian, C. A., Blackwelder, P. L., Hood, T., Nelsen, T. A.& Featherstone, C. 2000: Ostracods as indicators of naturaland anthropogenically-induced changes in coastal marine
environments. Coasts at the Millenium, Proceedings of the 17thInternational Conference of the Coastal Society, Portland, USA,July 9–12, 2000, 896–905.
Alvarez Zarikian, C. A., Stepanova, A. Y. & Grützner, J. 2009:Glacial–interglacial variability in deep sea ostracod assemblagecomposition at IODP Site U1314 in the subpolar North Atlantic.Marine Geology 258, 69–87.
Bauch, H. A., Kassens, H., Erlenkeuser, H., Grootes, P. M. &Thiede, J. 1999: Depositional environment of the Laptev Sea(Arctic Siberia) during the Holocene. Boreas 28, 201–204.
Bauch, H. A., Mueller-Lupp, T., Taldenkova, E., Spielhagen, R. F.,Kassens, H., Grootes, P. M., Thiede, J., Heinemeier, J. &Petryashov, V. 2001: Chronology of the Holocene Transgression atthe North Siberian margin. Global and Planetary Change 31, 125–139.
Benson, R. H., DelGrosso, R. M. & Steinbeck, P. L. 1983: Ostracodedistribution and biofacies, Newfoundland continental slope andrise. Micropaleontology 29, 430–453.
Boomer, I. & Eisenhauer, G. 2002: Ostracod faunas as palaeoenvi-ronmental indicators in marginal marine environments. InHolmes, A. & Chivas, A. R. (eds.): The Ostracoda. Applications inQuaternary research, 135–151. Geophysical Monograph 131.
Brouwers, E. M. 1994: Systematic paleontology of Quaternary Ostra-code assemblages from the Gulf of Alaska, Part 3: Family Cytheru-ridae. US Geological Survey, Professional Paper 1544, 43 pp.
Brouwers, E. M., Cronin, T. M., Horne, D. J. & Lord, A. R. 2000:Recent shallow marine ostracods from high latitudes: implicationsfor late Pliocene and Quaternary palaeoclimatology. Boreas 29,127–142.
Cronin, T. M. 1977: Champlanian Sea foraminifera and ostracoda: asystematic and paleontological synthesis. Géographie Physique etQuaternaire 31, 107–122.
Cronin, T. M. 1981: Paleoclimatic implications of late Pleistocenemarine ostracods from the St. Lawrence Lowlands. Micropaleon-tology 27, 384–418.
Cronin, T. M. & Vann, C. D. 2003: The sedimentary record ofclimatic and anthropogenic influence on the Patuxent Estuary andChesapeake Bay ecosystems. Estuaries 26, 196–209.
Cronin, T. M., Briggs, W. M. Jr, Brouwers, E. M., Whatley, R. C.,Wood, A. & Cotton, M. 1991: Modern Arctic Podocopid data-base. US Geological Survey, Open-File Report 91-385, 1–51.
Cronin, T. M., Dwyer, G. S., Schwede, S. B., Vann, C. D. & Dowsett,H. 2002: Climate variability from the Florida Bay sedimentaryrecord: possible teleconnections to ENSO, PNA and CNP. ClimateResearch 19, 233–245.
Cronin, T. M., Gemery, L., Briggs, W. M. Jr, Jakobsson, M., Polyak,L. & Brouwers, E. M. 2010: Quaternary sea ice history in the ArcticOcean based on a new Ostracode sea-ice proxy. Quaternary ScienceReviews 29, 3415–3429.
Cronin, T. M., Holtz, T. R., Stein, R., Spielhagen, R., Fütterer, D. &Wollenburg, J. 1995: Late Quaternary paleoceanography of theEurasian Basin, Arctic Ocean. Paleoceanography 10, 259–281.
Cronin, T. M., Holtz, T. R. & Whatley, R. C. 1994: Quaternarypaleoceanography of the deep Arctic Ocean based on quantitativeanalysis of Ostracoda. Marine Geology 119, 305–332.
Darby, D., Polyak, L. & Bauch, H. A. 2006: Past glacial and inter-glacial conditions in the Arctic Ocean and marginal seas – a review.Progress in Oceanography 71, 129–144.
Dittmers, K., Niessen, F. & Stein, R. 2003: Holocene sedimentbudget and sedimentary history for the Ob and Yenisei estuaries.In Stein, R., Fahl, K., Fütterer, D. K., Galimov, E. M. & Stepan-ets, O. V. (eds.): Siberian River Run-Off in the Kara Sea: Charac-terisation, Quantification, Variability and EnvironmentalSignificance, 457–470. Proceedings in Marine Sciences, Elsevier,Amsterdam.
Dmitrenko, I. A., Gribanov, V. A., Volkov, D. L., Berezovskaya, S.L. & Kassens, H. 2000: The role of riverine runoff in the interan-nual variability of the fast ice distribution in the Russian Arctic.Meteorologiya i Gidrologiya 3, 85–94 (in Russian).
Dmitrenko, I. A., Hoelemann, J., Kirillov, S. A., Berezovskaya, S. L.& Kassens, H. 2001: Role of barotropic sealevel changes in currentformation on the eastern shelf of the Laptev Sea. Doklady EarthSciences 377, 243–249.
Ostracod palaeoecology and environmental change in the Laptev and Kara seas, last 18 000 years 19BOREAS

Dmitrenko, I. A., Kirillov, S. A., Tremblay, L. B., Bauch, D.,Hölemann, J. A., Krumpen, T., Kassens, H., Wegner, C., Hein-emann, G. & Schröder, D. 2010: Impact of the Arctic Ocean Atlan-tic water layer on Siberian shelf hydrography. Journal ofGeophysical Research 115, C08010. doi: 10.1029/2009JC006020.
Dobrovol’skii, A. D. & Zalogin, B. S. 1982: Morya SSSR (Seas of theUSSR). 192 pp. Moscow University Press, Moscow (in Russian).
Duplessy, J. C., Ivanova, E. V., Murdmaa, I. O., Paterne, M. &Labeyrie, L. 2001: Holocene palaeoceanography of the NorthernBarents Sea and variations of the northward heat transport by theAtlantic Ocean. Boreas 30, 2–16.
Eicken, H., Reimnitz, E., Alexandrov, V., Martin, T., Kassens, H. &Viehoff, T. 1997: Sea-ice processes in the Laptev Sea and theirimportance for sediment export. Continental Shelf Research 17,205–233.
Fairbanks, R. G. 1989: A 17,000-year glacio-eustatic sea level record:influence of glacial melting rates on the Yonger Dryas event anddeep-ocean circulation. Nature 342, 637–642.
Fairbanks, R. G., Mortlock, R. A., Chiu, T., Cao, L., Kaplan, A.,Guilderson, T. P., Fairbanks, T. W., Bloom, A. L., Grootes, P. M.& Nadeau, M. J. 2005: Radiocarbon calibration curve spanning 0to 50,000 years BP based on paired 230Th/ 234U/ 238U and 14C dateson pristine corals. Quaternary Science Reviews 24, 1781–1796.
Frenzel, P., Keyser, D. & Viehberg, F. A. 2010: An illustrated keyand (paleo)ecological primer for Postglacial to Recent Ostracoda(Crustacea) of the Baltic Sea. Boreas 39, 567–575.
Griffiths, H. I. 2001: Ostracod evolution and extinction – its bios-tratigraphic value in the European Quaternary. QuaternaryScience Reviews 20, 1743–1751.
Ivanova, E. V., Murdmaa, I. O., Duplessy, J.-C. & Paterne, M. 2002:Late Weichselian to Holocene paleoenvironments of the BarentsSea. Global and Planetary Change 34, 69–78.
Jones, R., Whatley, R. C. & Cronin, T. 1998: The zoogeographicaldistribution of deep water Ostracoda in the Arctic Ocean. InCrasquin-Soleau, S., Braccini, E. & Lethiers, F. (eds.): What AboutOstracoda!, 83–90. Third European Ostracodologists Meeting,Paris, Bierville, 8–12 July 1996. Elf Exploration, Production Edi-tions, Pau.
Jones, R. L., Whatley, R. C., Cronin, T. M. & Dowsett, H. J. 1999:Reconstructing late Quaternary deep-water masses in the easternArctic Ocean using benthonic Ostracoda. Marine Micropaleonto-logy 37, 251–272.
Lev, O. M. 1983: Neogene-Quaternary ostracod assemblages. InGramberg, I. S. & Kulakov, Y. N. (eds.): The Main Problems of theLate Cenozoic Paleogeography of the Arctic, 104–142. Nedra, Len-ingrad (in Russian).
Lord, A., Robinson, J. E. & Moutzourides, S. G. 1993: Ostracodafrom Holstenian deposits in the Hamburg Area. Geology JournalA138, 127–145.
Lubinski, D. J., Polyak, L. & Forman, S. L. 2001: Freshwater andAtlantic water inflows to the deep northern Barents and Kara seassince ca 13 14C ka: foraminifera and stable isotope. QuaternaryScience Reviews 20, 1851–1879.
MacDonald, G. M., Velichko, A. A., Kremenetski, C. V., Borisova,O. K., Goleva, A. A., Andreev, A. A., Cwynar, L. C., Riding, R.T., Forman, S. L., Edwards, T. W. D., Aravena, R., Hammarlund,D., Szeicz, J. M. & Gattaulin, V. N. 2000: Holocene treeline historyand climate change across Northern Eurasia. Quaternary Research53, 302–311.
McDougall, K., Brouwers, E. M. & Smith, P. 1986: Micropaleonto-logy and sedimentology of the PB borehole series, Prudhoe Bay,Alaska. US Geological Survey Bulletin 1598, 1–62.
Naidina, O. D. & Bauch, H. A. 2011: Early to Middle Holocenepollen record from the Laptev Sea (Arctic Siberia). QuaternaryInternational 229, 84–88.
Nürnberg, D., Wollenburg, I., Dethleff, D., Eicken, H., Kassens, H.,Letzig, T., Reimnitz, E. & Thiede, J. 1994: Sediments in Arctic seaice: implications for entrainment, transport and release. MarineGeology 119, 185–214.
Pavlov, V. K. & Pfirman, S. L. 1995: Hydrographic structure andvariability of the Kara Sea: Implications for pollutant distribution.Deep-Sea Research II 42, 1369–1390.
Pavlov, V. K., Timokhov, L. A., Baskakov, G. A., Kulakov, M. Y.,Kurashov, V. K., Pavlov, P. V., Pivovarov, S. V. & Stanovoy, V. V.1996: Hydrometeorological regime of the Kara, Laptev and East-Siberian Seas. Applied Physics Laboratory, University of Washing-ton, Technical Memorandum 1-96, 1–179.
Polyak, L., Korsun, S., Febo, L., Stanovoy, V., Khusid, T., Hald, M.,Paulsen, B. E. & Lubinski, D. A. 2002: Benthic foraminiferalassemblages from the southern Kara Sea, a river-influenced arcticmarine environment. Journal of Foraminiferal Research 32, 252–273.
Polyakova, Y. I. & Stein, R. 2004: Holocene paleoenvironmentalimplications of diatom and organic carbon records from the south-eastern Kara Sea (Siberian Margin). Quaternary Research 62, 256–266.
Polyakova, Y. I., Bauch, H. A. & Novichkova, T. S. 2005: Pastchanges in Laptev Sea water masses deduced from diatom andaquatic palynomorph assemblages. Global and Planetary Change48, 208–222.
Reimnitz, E., Eicken, H. & Martin, T. 1995: Multiyear fast ice alongthe Taymyr Peninsula, Siberia. Arctic 48, 359–367.
Rudels, B., Jones, E. P., Schauer, U. & Eriksson, P. 2004: Atlanticsources of the Arctic Ocean surface and halocline waters. PolarResearch 23, 181–208.
Schornikov, E. I. & Zenina, M. A. 2006: Benthic ostracod fauna ofthe Kara, Laptev and East-Siberian seas (data from the expeditionscarried out by the Pacific Oceanography Institute of the EasternBranch of the Russian Academy of Sciences). In Sergienko, V.I.,Semiletov, I.P. (eds.): Marine investigations of DVO RAS in theArctic. Proceedings of the Arctic Regional Center IV, 156–211.Dal’nauka, Vladivostok (in Russian).
Shannon, C. E. & Weaver, W. 1963: The Mathematical Theory ofCommunication. 144 pp. University of Illinois Press, Champaign.
Simstich, J., Erlenkeuser, H., Harms, I., Spielhagen, R. & Stanovoy,V. 2005: Modern and Holocene hydrographic characteristics of theshallow Kara Sea shelf (Siberia) as reflected by stable isotopes ofbivalves and benthic foraminifera. Boreas 34, 252–263.
Simstich, J., Stanovoy, V., Bauch, D., Erlenkeuser, H. & Spielhagen,R. F. 2004: Holocene variability of bottom water hydrographyon the Kara Sea shelf (Siberia) depicted in multiple single-valveanalyses of stable isotopes in ostracods. Marine Geology 206, 147–164.
Slubowska, M. A., Koç, N., Rasmussen, T. L. & Klitgaard-Kristensen, D. 2005: Changes in the flow of Atlantic water into theArctic Ocean since the last deglaciation: Evidence from the north-ern Svalbard continental margin, 80°N. Paleoceanography 20,PA4014, doi:10.1029/2005PA001141.
Slubowska-Woldengen, M., Koç, N., Rasmussen, T. L., Klitgaard-Kristensen, D., Hald, M. & Jennings, A. E. 2008: Time-slice recon-structions of ocean circulation changes at the continental marginsof the Nordic and Barents Seas during the last 16,000 cal yr B.P.Quaternary Science Reviews 27, 1476–1492.
Slubowska-Woldengen, M., Rasmussen, T. L., Koç, N., Klitgaard-Kristensen, D., Nilsen, F. & Solheim, A. 2007: Advection of Atlan-tic Water to the western and northern Svalbard shelf since 17,500cal yr BP. Quaternary Science Reviews 26, 463–478.
Spielhagen, R. F., Baumann, K.-H., Erlenkeuser, H., Nowaczyk, N.R., Nørgaard-Pedersen, N., Vogt, C. & Weiel, D. 2004: ArcticOcean deep-sea record of northern Eurasian ice sheet history.Quaternary Science Reviews 23, 1455–1483.
Spielhagen, R. F., Erlenkeuser, H. & Siegert, C. 2005: History offreshwater runoff across the Laptev Sea (Arctic) during the Lastdeglaciation. Global and Planetary Change 48, 187–207.
Stein, R., Boucsein, B., Fahl, K., Garcia De Oteyza, T., Knies, J. &Niessen, F. 2001: Accumulation of particulate organic carbon atthe Eurasian continental margin during late Quaternary times:controlling mechanisms and paleoenvironmental significance.Global and Planetary Change 31, 87–104.
Stein, R., Dittmers, K., Fahl, K., Kraus, M., Matthiessen, J.,Niessen, F., Pirrung, M., Polyakova, Y., Schoster, F., Steinke, T.& Fütterer, D. 2004: Arctic (palaeo) river discharge and environ-mental change: evidence from the Holocene Kara Sea sedimentaryrecord. Quaternary Science Reviews 23, 1485–1511.
20 Anna Stepanova et al. BOREAS

Stepanova, A., Taldenkova, E. & Bauch, H. A. 2003: Recent Ostra-coda of the Laptev Sea (Arctic Siberia): taxonomic compositionand some environmental implications. Marine Micropaleontology48, 23–48.
Stepanova, A., Taldenkova, E. & Bauch, H. A. 2004: Ostracodspecies of the genus Cytheropteron from Late Pleistocene,Holocene and Recent sediments of the Laptev Sea (Arctic Siberia).Revista Española de Micropaleontologia 36, 83–108.
Stepanova, A., Taldenkova, E., Simstich, J. & Bauch, H. A. 2007:Comparison study of the modern ostracod associations in the Karaand Laptev seas: Ecological aspects. Marine Micropaleontology 63,111–142.
Stepanova, A. Y. 2006: Late Pleistocene-Holocene and Recent Ostra-coda of the Laptev Sea and their importance for paleoenvironmen-tal reconstructions. Paleontological Journal 40, Supplement 2,91–204.
Stuiver, M. & Reimer, P. J. 1993: Extended 14C data base and revisedCALIB 3.0 14C ages calibration program. Radiocarbon 35, 215–230.
Svendsen, J. I., Alexanderson, H., Astakhov, V. I. et al. 2004: LateQuaternary ice sheet history of eastern Eurasia. QuaternaryScience Reviews 23, 1229–1271.
Taldenkova, E., Bauch, H. A., Gottschalk, J., Nikolaev, S., Rostovt-seva, Y., Pogodina, I., Ovsepyan, Y. & Kandiano, E. 2010: Historyof ice-rafting and water mass evolution at the northern Siberiancontinental margin (Laptev Sea) during Late Glacial and Holocenetimes. Quaternary Science Reviews 29, 3919–3935.
Taldenkova, E., Bauch, H. A., Stepanova, A., Dem’yankov, S. &Ovsepyan, A. 2005: Last postglacial environmental evolution ofthe Laptev Sea shelf as reflected in molluscan, ostracodal, andforaminiferal faunas. Global and Planetary Change 48, 223–251.
Taldenkova, E., Bauch, H. A., Stepanova, A., Strezh, A.,Dem’yankov, S. & Ovsepyan, Y. 2008: Postglacial to Holocenehistory of the Laptev Sea continental margin: paleoenvironmentalimplications of benthic assemblages. Quaternary International 183,40–60.
Taldenkova, E. E., Bauch, H. A., Stepanova, A. Y., Pogodina, I. A.,Ovsepyan, Y. S. & Simstich, J. 2009: Paleoenvironmental changesof the Laptev and Kara sea shelves during the Postglacial trans-gression (inferred from fossil ostracods and foraminifers). InKassens, H., Lisitzin, A. P., Thiede, J., Polyakova, Y. I.,Timokhov, L. A. & Frolov, I. E. (eds.): System of the Laptev Seaand the Adjacent Arctic Seas: Modern and Past Environments, 384–409. Moscow University Press, Moscow (in Russian with Englishabstract).
Wegner, C., Hölemann, J. A., Dmitrenko, I., Kirillov, S. & Kassens,H. 2005: Seasonal variations in Arctic sediment dynamics – evi-dence from 1-year records in the Laptev Sea (Siberian Arctic).Global and Planetary Change 48, 126–140.
Whatley, R. C. 1983: Some simple procedures for enhancing the useof Ostracoda in paleoenvironmental analysis. Norwegian Petro-leum Directorate, Bulletin 2, 129–146.
Whatley, R. C., Eynon, M. & Moguilevsky, A. 1996: Recent Ostra-coda of the Scoresby Sund fjord system, East Greenland. RevistaEspanola de Micropaleontologia 28, 5–23.
Whatley, R. C., Eynon, M. & Moguilevsky, A. 1998: The depthdistribution of Ostracoda from the Greenland Sea. Journal ofMicropalaeontology 17, 15–32.
Supporting information
Additional Supporting Information may be found inthe online version of this article:
Table S1. Radiocarbon (AMS) and calendar ages ofthe studied cores.
Table S2. Some characteristic ostracod species fromdifferent environments in the Laptev and Kara seas(Stepanova et al. 2003, 2004, 2007; Stepanova 2006).I=index species; d=dominant species; r=rare species
Table S3. Description of ostracod assemblages and(paleo)ecological references for ostracod taxa based onour own data and published evidence. f=freshwater;br=brackish water; eury=euryhaline; sm=shallow-watermarine; dm=deep-water marine; Atl.=deep-watermarine with North Atlantic affinities.
Please note: Wiley-Blackwell are not responsible for thecontent or functionality of any supporting materialssupplied by the authors. Any queries (other thanmissing material) should be directed to the correspond-ing author for the article.
Ostracod palaeoecology and environmental change in the Laptev and Kara seas, last 18 000 years 21BOREAS





























