Embed Size (px)
Citation preview

Oroxylin A, a classical natural product, shows
a novel inhibitory effect on angiogenesis induced
by lipopolysaccharide
Xiuming Song1, Yan Chen1, Yajing Sun1, Biqi Lin2, Yansu Qin1, Hui Hui1,
Zhiyu Li1, Qidong You1, Na Lu1, Qinglong Guo1
1State Key Laboratory of Natural Medicines, Jiangsu Key Laboratory of Carcinogenesis and Intervention,
China Pharmaceutical University, 24 Tongjiaxiang, Nanjing 210009, P.R. China
2School of Pharmacy, China Pharmaceutical University, Nanjing 210009, P.R. China
Correspondence: Na Lu, e-mail: [email protected]; Qinglong Guo, e-mail: [email protected]
Abstract:
Background: There is an obvious relationship among angiogenesis and inflammation. From previous study, we learn that oroxylin
A possesses anti-angiogenic activity in vitro and in ovo. It also has an inhibitory effect on inflammation. But whether oroxylin A sup-
presses the inflammation-induced angiogenesis is still unknown. Our present study focuses on the role of oroxylin A in targeting
LPS-induced angiogenesis, inflammatory and related pathways.
Methods: The effects of oroxylin A on angiogenesis were investigated by transwell assay, tube formation assay, rat aortic ring assay
and chorioallantoic membrane (CAM) model. Western blotting analysis was used to detect the expression of certain proteins.
Results: We found that oroxylin A inhibited LPS-induced migration and tube formation of human umbilical vein endothelial cells
(HUVECs), as well as microvessel sprouting from rat aotric ring in vitro and the angiogenesis of chicken chorioallantoic membrane
(CAM) model in ovo. The results also indicated that oroxylin A could inhibit the expression of LPS acceptor toll-like receptor 4
(TLR4) and the activities of its downstream mitogen-activated protein kinases (MAPKs), including reducing expressions of the
phosphorylation of JNK, p38, and ERK. Moreover, oroxylin A prevented NF-kB dimers from translocating to the nucleus.
Conclusions: Taken together, oroxylin A can suppress the angiogenesis induced by LPS and it may affect the LPS/TLR4 signaling
pathway.
Key words:
Scutellaria baicalensis Georgi (Lamiaceae), oroxylin A, lipopolysaccharide, angiogenesis, toll like receptor 4 (TLR4)
Abbreviations: CAM – chorioallantoic membrane, DMSO –
dimethyl sulfoxide, ERK – extracellular signal-regulated ki-
nases, FBS – fetal bovine serum, HUVECs – human umbilical
vein endothelial cells, IKK – IkB kinase, JNKs – c-Jun N-
terminal kinases, LPS – lipopolysaccharide, MAPKs – mito-
gen-activated protein kinases, p38 – p38 mitogen-activated
protein kinases, TLRs – toll-like receptors
Introduction
Angiogenesis occurs in many physiological and
pathological processes such as reproductive functions
in adults, wound healing, tumor development, some
Pharmacological Reports, 2012, 64, 1189�1199 1189
Pharmacological Reports2012, 64, 1189�1199ISSN 1734-1140
Copyright © 2012by Institute of PharmacologyPolish Academy of Sciences

chronic inflammatory diseases and so on [23]. A link
between angiogenesis and inflammation has been
strongly supported. It seems that most inflammations,
especially chronic inflammations, are linked to angio-
genesis [2, 10]. In inflammations, vascular remodel-
ing occurs and contributes to inflammatory diseases,
proceeding to other diseases for the next step. Chronic
inflammation also promotes angiogenesis by certain
inflammatory factors. Angiogenesis, a significant risk
factor for the development of inflammations, plays an
important role in inflammatory diseases. Thus, target-
ing angiogenesis may be effective in the development
of some diseases caused by inflammation.
Lipopolysaccharide (LPS), an important structural
component of the outer membrane of Gram-negative
bacteria, acts as a mediator in inflammatory responses
and it also induces angiogenesis [19] as an exogenous
factor in several chronic inflammatory diseases. LPS
promotes angiogenesis in many stages in which the
toll-like receptors (TLRs) play a central role. TLRs
are receptors involved in the immune response and
can recognize LPS as their ligand [16]. LPS/TLR in-
tracellular signaling pathway depends on the activity
of tumor necrosis factor (TNF) receptor associated
factor 6 (TRAF6) [19]. Different TLRs use different
combinations of adaptor proteins to determine down-
stream signaling and TLR4 is critical for signal trans-
duction utilizing all the adaptor proteins in the path-
way [14]. In the process of angiogenesis stimulated
by LPS, the activation of TLR4 can induce the up-
regulation of chemokines and adhesion molecules in
MAPKs pathway and stimulate NF-kB leading to the
synthesis and secretion of various cytokines, metabo-
lites, and enzymes that in turn mediate their different
biological effects [22].
The Chinese herbs, Huang-Chi, Scutellaria bai-
calensis Georgi, has been widely used to treat several
diseases such as inflammation, diarrhea, and pyro-
genic infections and so on. Oroxylin A is one of the
major flavonoids which are the most common active
ingredients in the root of Scutellaria baicalensis
Georgi [11]. It has been widely studied and proved to
possess a wide spectrum of pharmacological proper-
ties, such as pro-apoptotic [12] and anti-invasive [15,
25] activities. Several previous reports suggested that
oroxylin A was a potential antiinflammatory agent [11].
But whether oroxylin A can inhibit LPS-mediated in-
duction of angiogenesis in some chronic inflamma-
tory diseases is still unknown. What’s more, the mo-
lecular mechanisms remain poorly understood and
warrant further investigation.
In the present study, we isolated oroxylin A from Scutel-
laria baicalensis Georgi and identified its structure. The ef-
fect of oroxylin A on LPS-induced angiogenesis in vitro
and in ovo was also examined, which depended on TLR4,
phosphorylation of MAPKs and activation of NF-kB.
Materials and Methods
Cell lines, chemicals and biochemicals
Human umbilical vein endothelial cells (HUVECs)
were isolated from human umbilical cord veins by
collagenase treatment as described previously [9].
The harvested cells were grown in medium 199
(Gibco, Grand Island, NY) containing endothelial cell
growth supplement (ECGS, 30 µg/ml; Sigma, St.
Louis, MO), epidermal growth factor (EGF, 10 ng/ml;
Sigma, St. Louis, MO), 20% fetal bovine serum (FBS,
Gibco, Grand Island, NY), 100 U/ml penicillin, and
100 µg/ml streptomycin, pH 7.4. The cells were incu-
bated in a humidified atmosphere of 95% air + 5%
CO2 at 37°C. After 3–5 passages, HUVECs were col-
lected for use in all experiments.
The drug was dissolved in dimethyl sulfoxide
(DMSO) as a stock solution, stored at –20°C, and di-
luted with medium before experiments. The final
DMSO concentration did not exceed 0.1% throughout
the study. The control groups were treated with 0.1%
DMSO in the corresponding experiments. Escheri-
chia coli (055:B5) LPS was supplied by Sigma
Chemical Co. (St. Louis, MO). Primary antibodies for
TLR4 (H-80), p38, p-p38, p-ERK1/2, JNK, p-JNK,
NF-kB, IkB, lamin A and b-actin were obtained from
Santa Cruz Biotechnology (Santa Cruz, CA). Primary
antibody for ERK1/2 and p-IKK were from Bioworld
(St. Louis Park, MN). Primary antibody for IKK and
p-IkB were from Cellsignaling (Danvers, MA).
IRDyeTM800 conjugated secondary antibodies were
obtained from Rockland Inc. (Philadelphia, PA).
Plant material
The roots of Scutellaria baicalensis Georgi (Radix
Scutellariae) were purchased from Bozhou, Anhui,
China (latitude: 33° 52’ 38’’ N, longitude: 115° 46’
13’’ E). A voucher specimen was deposited at Depart-
ment of Natural Medicinal Chemistry and identified
by Dr. F. Feng, China Pharmaceutical University.
1190 Pharmacological Reports, 2012, 64, 1189�1199

Extraction and isolation
Oroxylin A was isolated and purified from Radix Scu-
tellariae according to the established methods [24] with
slight modifications. In brief, dried Scutellaria bai-
calensis Georgi roots were chopped into small pieces,
immersed, and extracted with 10-fold v/w acetone
twice at room temperature for 2 weeks. After filtration,
the residues were reflux-extracted with 4-fold v/w of
50% aqueous ethanol twice for 6 h. The acetone ex-
tracts were subjected to column chromatography on
silica gel eluted with CHCl3 and CHCl3-MeOH (9:1,
v/v), and rechromatographed on silica gel eluted with
hexane-acetone (1:1, v/v) to yield oroxylin A.
Identification and content determination
The structure of oroxylin A was identified using IR, UV,
TOF/MS, and 1H-NMR. The data were consistent with
that had been reported before [7]. The purity level of
oroxylin A exceeded 99% as determined by HPLC with
Evaporative Light Scattering Detector (HPLC ELSD).
Endothelial cell migration assay
Chemotactic motility of HUVECs was assayed using
a Transwell chamber (Millicell, Billerico, MA) as de-
scribed previously [18]. Briefly, cells were incubated
in M199 containing 1% FBS in the absence or pres-
ence of oroxylin A (1, 10 and 100 µM) for 60 min.
Then, cells were trypsinized and suspended at a final
concentration of 5 × 105 cells/ml in M199 containing
1% FBS. The fresh M199 medium containing 1%
FBS and 1 µg/ml LPS was placed in the lower wells.
Cell suspension was loaded into each of the upper
wells. After 4 h, cells were fixed with methanol, stain-
ing with crystal violet, releasing the bound dye with
33% glacial acetic acid, and measuring the optical
density (OD) of the solution at 570 nm by using an
enzyme immunosorbent assay reader.
Tube formation assay
An in vitro capillary tube formation assay was per-
formed as described earlier [18]. Briefly, a 96-well
plate was coated with 80 µl matrigel (Becton Dickin-
son, Bedford, MA), which was allowed to solidify
and polymerize at 37°C for 30 min. HUVECs were
incubated in medium containing 1% FBS in the ab-
sence or presence of oroxylin A (1, 10 and 100 µM)
for 60 min. These cells were harvested after trypsin
treatment and suspended in medium containing 1%
FBS before seeding and planting onto matrigel and
then LPS (1 µg/ml) was added. Following 8 h of incu-
bation, the plate was examined for capillary tube for-
mation under an inverted microscope and photo-
graphed and tubular structures were quantified by
manual counting the tube numbers, and five randomly
chosen fields were analyzed for each well.
Rat aortic ring assay
Rat aortic ring assay was performed as described pre-
viously [1]. The thoracic aorta was dissected from
male Sprague-Dawley rats (6 weeks old) and cut into
1-mm long rings and set in 24-well plates. The disks
were treated as described previously [1]. Clotting me-
dia contained M199+ (M199 with 100 U/ml penicillin
and 100 µg/ml streptomycin) plus 0.3% fibrinogen
and 0.5% e-amino-n-caproic acid (ACA; Sigma, St.
Louis, MO). Growth media consisted of M199+ with
20% FBS and 0.5% ACA. Then, the growth media
were added to the wells with or without 1 µg/ml LPS
and various concentrations of oroxylin A (8, 40 and
200 µM). Plates were then stored in incubator at 37°C
and 5% CO2. After 7 days, the sprouting microvessels
in five randomly chosen fields were counted and pho-
tographed under a microscope for each group.
Chicken chorioallantoic membrane (CAM) assay
Anti-angiogenic effect of oroxylin A on CAM was as-
sayed according to the method described with modifi-
cation [13]. Briefly, fertilized chicken eggs were incu-
bated at 37°C for 9 days. After this incubation, a small
hole was punched on the broad side of the egg, and
a window was carefully created through the egg shell.
Sterilized filter paper disks (5 × 5 mm) saturated with
or without LPS (1 µg/ml) and oroxylin A (1.5, 15 and
150 ng/egg) were placed on the CAMs. They were
then incubated at 37°C for another 2 days. After 48 h
of incubation, shells of fertilized chicken eggs were
opened. Then, an appropriate volume of 10% fat
emulsion (Intralipose, 10%) was injected into the em-
bryo chorioallantois for observing the density and
length of vessels toward the CAM face. Neovascular
zones under the filter paper disks were observed and
photographed with a digital camera at 5× magnifica-
tion. The number of newly growth vessels were
counted on digitalized pictures.
Pharmacological Reports, 2012, 64, 1189�1199 1191
Oroxylin A inhibits LPS induced angiogenesisXiuming Song et al.

Western blot analysis
Human umbilical vein endothelial cells were pre-
treated for 2 h with various concentrations of oroxylin
A (1, 10 and 100 µM) and stimulated with LPS
(1 µg/ml) for 60 min. After stimulation, cells were
collected and lysed in lysis buffer [50 mM Tris-Cl, pH
7.6, 150 mM NaCl, 1 mM EDTA, 1% (m/v) NP-40,
0.2 mM PMSF, 0.1 mM NaF and 1 mM DTT], and
lysates were clarified by centrifugation at 4°C for
15 min at 13,000 × g. The concentration of protein in
the supernatants was detected using bicinchoninic
acid (BCA) assay with a Varioskan multimode micro-
plate spectrophotometer (Thermo, Marietta, OH).
Then, equal amount of total proteins was separated by
SDS-PAGE and transferred onto the PVDF mem-
branes (Millipore, Billerica, MA). The blots were in-
cubated with specific antibodies against indicated pri-
mary antibodies overnight at 4°C followed by
IRDyeTM800 conjugated secondary antibody for 1 h at
37°C. Detection was performed by the Odyssey Infra-
red Imaging System (LI-COR Inc., Lincoln, NE).
Statistical analysis
All data in different experimental groups were expressed
as the mean ± SEM. These data shown in the study were
obtained in at least three independent experiments. Sta-
tistical analyses were performed using an unpaired,
two-tailed Student’s t-test. All comparisons were made
relative to LPS-treated groups; p value less than 0.05
was considered as statistically significant.
Results
Oroxylin A reduces LPS-induced migration
of HUVECs
Transwell assay was used to evaluate the effect of orox-
ylin A on the migration of HUVECs. As shown in
Figure 1, LPS stimulated HUVECs migration compared
to the control. When treated with 1, 10 and 100 µM
oroxylin A, fewer cells migrated to the lower side of the
filter in the Transwell Chamber. Oroxylin A signifi-
cantly inhibited the migration of HUVECs in a con-
centration-dependent manner. The inhibition of migra-
tion is reduced by 11%, 44% and 59% at 1, 10 and 100
µM, respectively, compared with the LPS group.
Oroxylin A suppresses LPS-stimulated tube
formation of HUVECs
Tube formation assay was performed to evaluate the ef-
fects of oroxylin A on LPS-induced tube formation of
HUVECs on a matrigel substratum. As shown in Figure
2, LPS promoted formation of capillary-like tubes, and
the number of tubes was much larger than control group.
Tube formation of cells treated with oroxylin A before
seeding and plating onto matrigel was inhibited in
a concentration-dependent manner. Quantitative analysis
revealed that treatment with oroxylin A resulted in 27%,
65% and 80% inhibition of LPS-induced tube formation
at 1, 10, and 100 µM, respectively.
Oroxylin A inhibits LPS-stimulated
angiogenesis in vitro
Several stages in angiogenesis, including endothelial
cell proliferation, migration, and tube formation, can
be mimicked in the rat aortic ring model. We utilized
the model to evaluate the effect of oroxylin A on
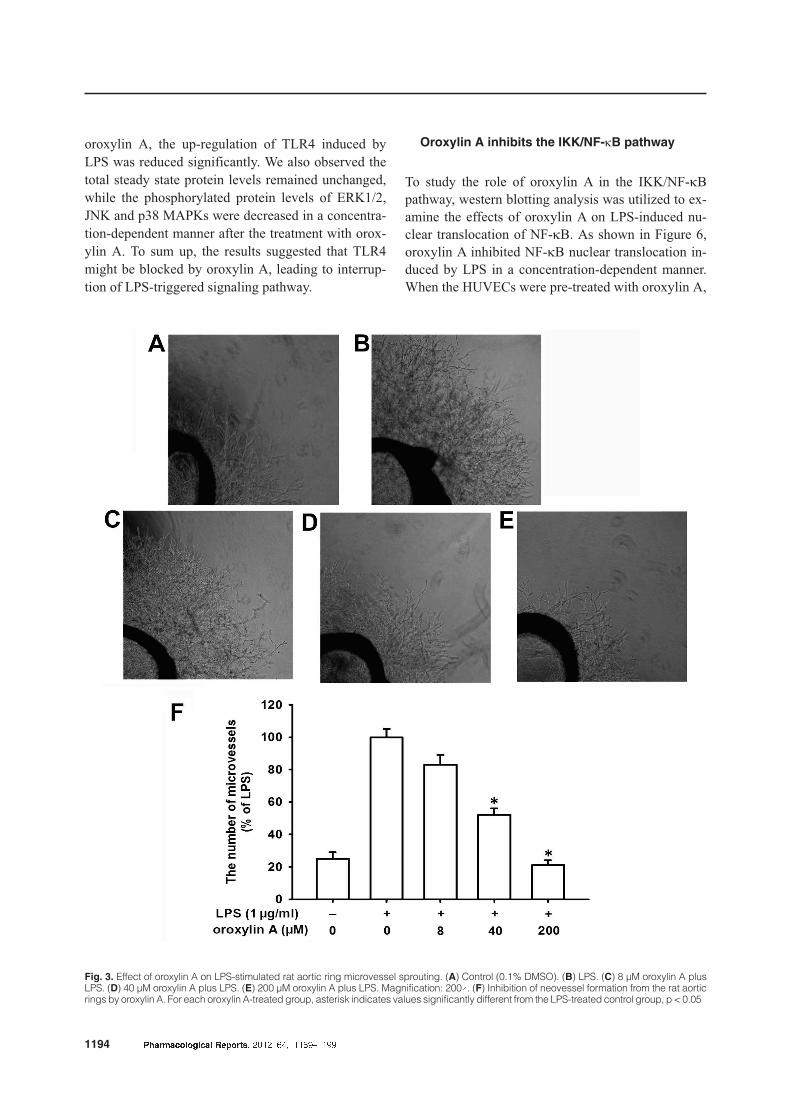
LPS-induced angiogenesis. As shown in Figure 3,
oroxylin A at 8, 40 and 200 µM resulted in the inhibi-
tion of microvessel growth by 17%, 48% and 79%, re-
spectively. At the tested concentrations, oroxylin A
suppressed the formation of microvessel outgrowth
from explants of rat aorta in a concentration-depend-
ent manner.
1192 Pharmacological Reports, 2012, 64, 1189�1199
Fig. 1. Effect of oroxylin A on LPS-stimulated migration of HUVECs.Inhibition of LPS-stimulated migration of HUVECs by oroxylin A.Optical density (OD) of the solution was measured at 570 nm. Foreach oroxylin A-treated group, asterisk indicates values significantlydifferent from the LPS-treated control group, * p < 0.05, ** p < 0.01

Oroxylin A inhibits LPS-stimulated angiogenesis
in ovo
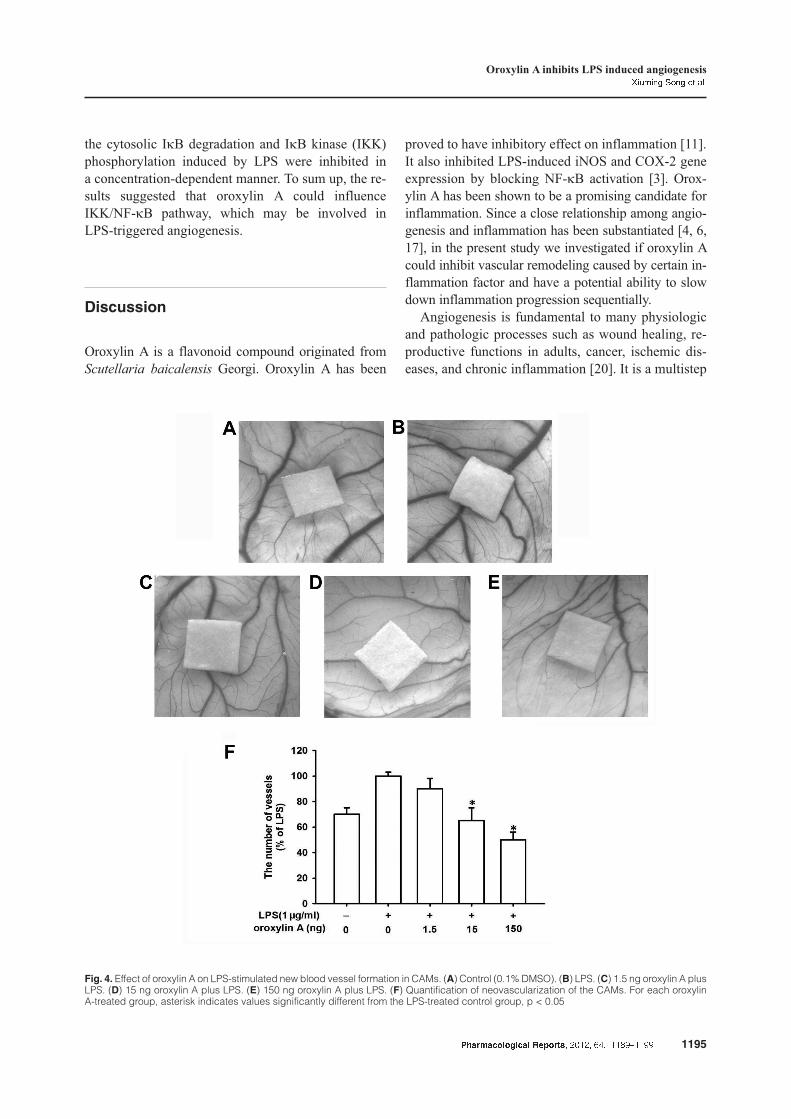
To study the anti-angiogenesis capacity of oroxylin A
in ovo, CAM model was applied. From this model, we
investigated the process of new blood vessel forma-
tion and vessel responses to anti-angiogenic agents.
As shown in Figure 4, more and thicker new blood
vessels were formed in the LPS group compared with
the control group. When oroxylin A was added, the
quantity of vessels was reduced by 10%, 35% and
50% at 1, 10, and 100 µM, respectively. It displayed
that oroxylin A inhibited the in ovo angiogenesis in
a concentration-dependent manner.
Influence of oroxylin A on TLR4 and its signaling
pathways
In this part, western blotting analysis was utilized to
evaluate the molecular mechanism of oroxylin A an-
tagonizing LPS-induced angiogenesis in HUVECs.
LPS promotes angiogenesis through activation of its
cell surface receptor TLR4. So we investigated the ef-
fects of oroxylin A on the interaction between LPS
and TLR4 as well as the activation of its downstream
signaling molecules including ERK1/2, JNK and p38
MAPKs. As shown in Figure 5, level of TLR4 was
apparently induced by addition of exogenous LPS to
HUVECs. When the HUVECs were pre-treated with
Pharmacological Reports, 2012, 64, 1189�1199 1193
Oroxylin A inhibits LPS induced angiogenesisXiuming Song et al.
Fig. 2. Effect of oroxylin A on LPS-stimulated tube formation of HUVECs. (A) Control (0.1% DMSO). (B) LPS. (C) 1 µM oroxylin A plus LPS.(D) 10 µM oroxylin A plus LPS. (E) 100 µM oroxylin A plus LPS. Magnification: 100´. (F) Inhibition of LPS-stimulated tube formation of HUVECsby oroxylin A. For each oroxylin A-treated group, asterisk indicates values significantly different from the LPS-treated control group, p < 0.05
oroxylin A, the up-regulation of TLR4 induced by
LPS was reduced significantly. We also observed the
total steady state protein levels remained unchanged,
while the phosphorylated protein levels of ERK1/2,
JNK and p38 MAPKs were decreased in a concentra-
tion-dependent manner after the treatment with orox-
ylin A. To sum up, the results suggested that TLR4
might be blocked by oroxylin A, leading to interrup-
tion of LPS-triggered signaling pathway.
Oroxylin A inhibits the IKK/NF-kB pathway
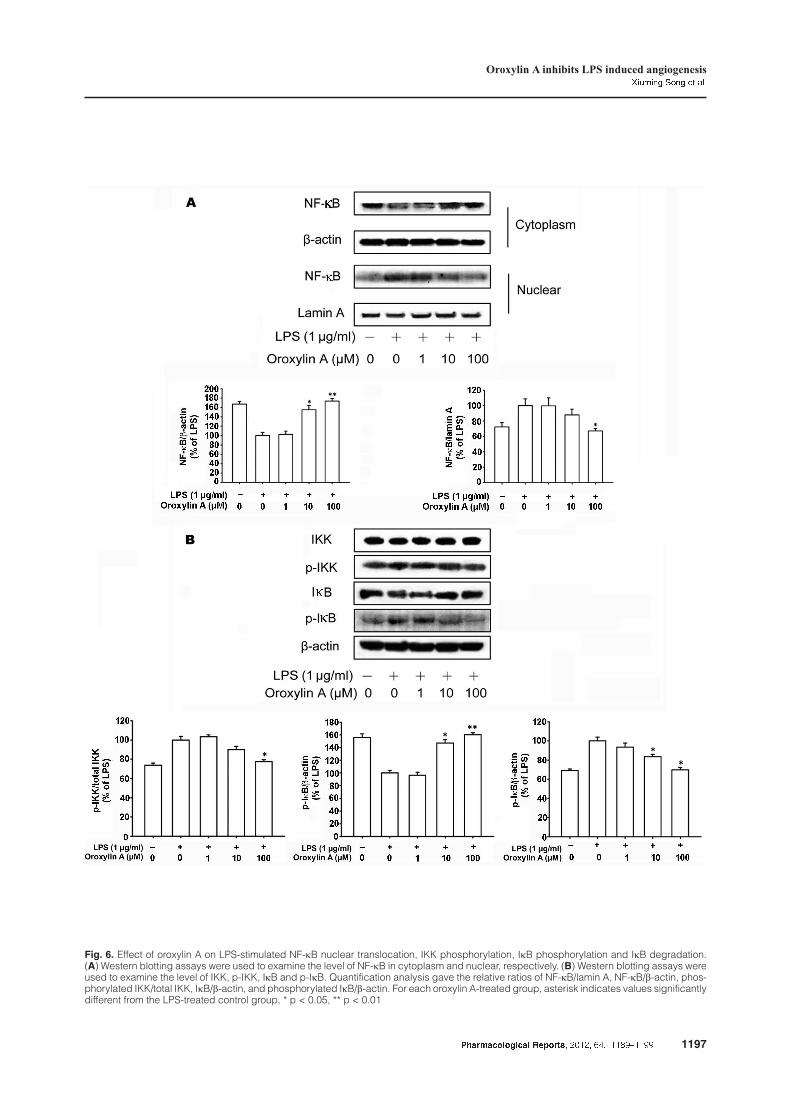
To study the role of oroxylin A in the IKK/NF-kB
pathway, western blotting analysis was utilized to ex-
amine the effects of oroxylin A on LPS-induced nu-
clear translocation of NF-kB. As shown in Figure 6,
oroxylin A inhibited NF-kB nuclear translocation in-
duced by LPS in a concentration-dependent manner.
When the HUVECs were pre-treated with oroxylin A,
1194 Pharmacological Reports, 2012, 64, 1189�1199
Fig. 3. Effect of oroxylin A on LPS-stimulated rat aortic ring microvessel sprouting. (A) Control (0.1% DMSO). (B) LPS. (C) 8 µM oroxylin A plusLPS. (D) 40 µM oroxylin A plus LPS. (E) 200 µM oroxylin A plus LPS. Magnification: 200´. (F) Inhibition of neovessel formation from the rat aorticrings by oroxylin A. For each oroxylin A-treated group, asterisk indicates values significantly different from the LPS-treated control group, p < 0.05
the cytosolic IkB degradation and IkB kinase (IKK)
phosphorylation induced by LPS were inhibited in
a concentration-dependent manner. To sum up, the re-
sults suggested that oroxylin A could influence
IKK/NF-kB pathway, which may be involved in
LPS-triggered angiogenesis.
Discussion
Oroxylin A is a flavonoid compound originated from
Scutellaria baicalensis Georgi. Oroxylin A has been
proved to have inhibitory effect on inflammation [11].
It also inhibited LPS-induced iNOS and COX-2 gene
expression by blocking NF-kB activation [3]. Orox-
ylin A has been shown to be a promising candidate for
inflammation. Since a close relationship among angio-
genesis and inflammation has been substantiated [4, 6,
17], in the present study we investigated if oroxylin A
could inhibit vascular remodeling caused by certain in-
flammation factor and have a potential ability to slow
down inflammation progression sequentially.
Angiogenesis is fundamental to many physiologic
and pathologic processes such as wound healing, re-
productive functions in adults, cancer, ischemic dis-
eases, and chronic inflammation [20]. It is a multistep
Pharmacological Reports, 2012, 64, 1189�1199 1195
Oroxylin A inhibits LPS induced angiogenesisXiuming Song et al.
Fig. 4. Effect of oroxylin A on LPS-stimulated new blood vessel formation in CAMs. (A) Control (0.1% DMSO). (B) LPS. (C) 1.5 ng oroxylin A plusLPS. (D) 15 ng oroxylin A plus LPS. (E) 150 ng oroxylin A plus LPS. (F) Quantification of neovascularization of the CAMs. For each oroxylinA-treated group, asterisk indicates values significantly different from the LPS-treated control group, p < 0.05

1196 Pharmacological Reports, 2012, 64, 1189�1199
Fig. 5. Effect of oroxylin A on LPS-stimulated activation of TLR4 and its signaling pathway. (A) Western blotting assays were used to examinethe expression of TLR4 and phosphorylation of ERK1/2, JNK and p38 MAPKs. (B) Quantification analysis gave the relative ratios of TLR4/b-actin, phosphorylated JNK/total JNK, phosphorylated ERK1/2/total ERK1/2, and phosphorylated p38 MAPK/total p38 MAPK. For each oroxylinA-treated group, asterisk indicates values significantly different from the LPS-treated control group, p < 0.05
Pharmacological Reports, 2012, 64, 1189�1199 1197
Oroxylin A inhibits LPS induced angiogenesisXiuming Song et al.
Fig. 6. Effect of oroxylin A on LPS-stimulated NF-kB nuclear translocation, IKK phosphorylation, IkB phosphorylation and IkB degradation.(A) Western blotting assays were used to examine the level of NF-kB in cytoplasm and nuclear, respectively. (B) Western blotting assays wereused to examine the level of IKK, p-IKK, IkB and p-IkB. Quantification analysis gave the relative ratios of NF-kB/lamin A, NF-kB/b-actin, phos-phorylated IKK/total IKK, IkB/b-actin, and phosphorylated IkB/b-actin. For each oroxylin A-treated group, asterisk indicates values significantlydifferent from the LPS-treated control group, * p < 0.05, ** p < 0.01

process of sprouting of new capillaries from preexist-
ing blood vessels involving the proliferation and mi-
gration of endothelial cells, remodeling of the extra-
cellular matrix, maturation of the newly blood vessels
and all kinds of cytokine production [8]. Angiogene-
sis and inflammation complement each other. Devel-
opment of angiogenesis is a multistep process which
can be activated by inflammatory agents. Inflamma-
tion promotes angiogenesis by many ways and it fur-
ther potentiates endothelial proliferation and angio-
genesis, while angiogenesis also contributes to the pa-
thology of inflammations. Vascular remodeling occurs
in many chronic inflammatory disorders, such as in-
flammatory bowel disease [21].
In our study, transwell assay and tube formation as-
say were employed to demonstrate the effects of orox-
ylin A on LPS-induced migration and tube formation
of HUVECs in vitro. The results showed that oroxylin
A remarkably suppressed the LPS-stimulated migra-
tion and tube formation of HUVECs. Rat aortic ring
assay was used to investigate if oroxylin A could in-
hibit the microvessels sprouting induced by LPS from
rat aorta. Then, we used CAM model to illustrate that
oroxylin A inhibited new vessel formation and vascu-
lar remodeling in ovo. Our results demonstrated that
oroxylin A could inhibit the microvessel outgrowth
from rat aorta in vitro and it also had anti-angiogenic
activity in ovo. These results implied that oroxylin
A could act as a probable anti-angiogenic compound
in inflammation.
LPS, a potent inflammatory mediator, plays an im-
portant role in inflammation and indeed has angio-
genic potential [5]. Its activities are transmitted
through TLRs. In fact, LPS directly causes endothe-
lial sprouting and angiogenesis utilizing the stimula-
tion of TLRs and TRAF6 [19]. Involvement of vari-
ous intracellular signaling pathways, such as MAPKs,
have been reported downstream TLR4. Activation of
MAPKs induces certain effects in LPS-induced
angiogenesis. Therefore, further experiments were
performed to determine whether oroxylin A could
affect activation of MAPKs. When HUVECs were
pretreated with oroxylin A, the upregulated expres-
sions of TLR4 and phosphorylated JNK, ERK and
p38 MAPKs induced by LPS were blocked, without
affecting their total proteins. This indicated that the
inhibitive effects of oroxylin A on the responsibility
of HUVECs to LPS might derive partly from inhibit-
ing TLR4 signal transduction. As one of the most im-
portant transcription factors, NF-kB plays a pivotal
role in LPS-induced angiogenesis due to its ability to
induce transcription of pro-inflammatory genes [19].
In our study, we found oroxylin A could prevent
NF-kB dimers from translocating to the nucleus due
to the inhibition of IKK phosphorylation and the sub-
sequent IkB degradation. This may prevent certain
target genes downstream NF-kB from being activated
in angiogenesis. Our findings suggested that inhibi-
tory effect of oroxylin A on LPS-induced angiogene-
sis in inflammation was also due to inhibition of
NF-kB pathway. Taken the results together, we con-
jecture that oroxylin A can influence LPS-induced
angiogenesis by inhibiting the expression of TLR4,
phosphorylation of MAPKs and activation of NF-kB.
In summary, we demonstrated that oroxylin A in-
hibited angiogenesis induced by LPS in vitro and in
ovo through down-regulating the activation of
LPS/TLR4 signaling pathway. Oroxylin A, as a novel
angiogenesis inhibitor, probably is beneficial in the
treatment of inflammatory disorders. Our findings
provide new insights into the molecular mechanisms
of oroxylin A-induced suppression of angiogenesis
and suggest possible applications for oroxylin A in in-
flammation diseases.
Conflict of interest:
None declared.
Acknowledgments:
This work was supported by the National Natural Science
Foundation of China (No. 30973556 and 81001452), Program
for Changjiang Scholars and Innovative Research Team in
University (IRT1193), the Project Program of State Key Laboratory
of Natural Medicines, China Pharmaceutical University
(No. JKGZ201101), the National Science & Technology Major
Project (No. 2012ZX09304-001) and Natural Science Foundation
of Jiangsu province (No. BK2009297 and No. BK2010432).
References:
1. Berger AC, Wang XQ, Zalatoris A, Cenna J, Watson JC:
A murine model of ex vivo angiogenesis using aortic
disks grown in fibrin clot. Microvasc Res, 2004, 68,
179–187.
2. Bonnet CS, Walsh DA: Osteoarthritis, angiogenesis and
inflammation. Rheumatology (Oxford), 2005, 44, 7–16.
3. Chen Y, Yang L, Lee TJ: Oroxylin A inhibition of
lipopolysaccharide-induced iNOS and COX-2 gene ex-
pression via suppression of nuclear factor-kappaB activa-
tion. Biochem Pharmacol, 2000, 59, 1445–1457.
4. Clavel G, Valvason C, Yamaoka K, Lemeiter D, Laroche
L, Boissier MC, Bessis N: Relationship between angio-
1198 Pharmacological Reports, 2012, 64, 1189�1199

genesis and inflammation in experimental arthritis. Eur
Cytokine Netw, 2006, 17, 202–210.
5. Harmey JH, Bucana CD, Lu W, Byrne AM, McDonnell
S, Lynch C, Bouchier-Hayes D et al.: Lipopoly-
saccharide-induced metastatic growth is associated with
increased angiogenesis, vascular permeability and tumor
cell invasion. Int J Cancer, 2002, 101, 415–422.
6. Haywood L, McWilliams DF, Pearson CI, Gill SE,
Ganesan A, Wilson D, Walsh DA: Inflammation and
angiogenesis in osteoarthritis. Arthritis Rheum, 2003, 48,
2173–2177.
7. Huang WH, Chien PY, Yang CH, Lee AR: Novel synthe-
sis of flavonoids of Scutellaria baicalensis Georgi. Chem
Pharm Bull (Tokyo), 2003, 51, 339–340.
8. Ichihara E, Kiura K, Tanimoto M: Targeting angiogene-
sis in cancer therapy. Acta Med Okayama, 2011, 65,
353–362.
9. Jaffe EA, Nachman RL, Becker CG, Minick CR: Culture
of human endothelial cells derived from umbilical veins.
Identification by morphologic and immunologic criteria.
J Clin Invest, 1973, 52, 2745–2756.
10. Kobayashi H, Lin PC: Angiogenesis links chronic in-
flammation with cancer. Methods Mol Biol, 2009, 511,
185–191.
11. Li C, Lin G, Zuo Z: Pharmacological effects and phar-
macokinetics properties of Radix Scutellariae and its bio-
active flavones. Biopharm Drug Dispos, 2011, 32,
427–445.
12. Li HN, Nie FF, Liu W, Dai QS, Lu N, Qi Q, Li ZY et al.:
Apoptosis induction of oroxylin A in human cervical
cancer HeLa cell line in vitro and in vivo. Toxicology,
2009, 257, 80–85.
13. Lu N, Yang Y, You QD, Ling Y, Gao Y, Gu HY, Zhao L
et al.: Gambogic acid inhibits angiogenesis through sup-
pressing vascular endothelial growth factor-induced tyro-
sine phosphorylation of KDR/Flk-1. Cancer Lett, 2007,
258, 80–89.
14. Lu YC, Yeh WC, Ohashi PS: LPS/TLR4 signal transduc-
tion pathway. Cytokine, 2008, 42, 145–151.
15. Lu ZJ, Lu N, Li CL, Li F, Zhao K, Lin B, Guo QL:
Oroxylin A inhibits matrix metalloproteinase-2/9 expres-
sion and activation by up-regulating tissue inhibitor of
metalloproteinase-2 and suppressing the ERK1/2 signal-
ing pathway. Toxicol Lett, 2012, 209, 211–220.
16. Makela SM, Osterlund P, Julkunen: TLR ligands induce
synergistic interferon-beta and interferon-lambda1 gene
expression in human monocyte-derived dendritic cells.
Mol Immunol, 2011, 48, 505–515.
17. Mazibrada J, Ritta M, Mondini M, De Andrea M,
Azzimonti B, Borgogna C, Ciotti M et al.: Interaction
between inflammation and angiogenesis during different
stages of cervical carcinogenesis. Gynecol Oncol, 2008,
108, 112–120.
18. Min JK, Han KY, Kim EC, Kim YM, Lee SW, Kim OH,
Kim KW et al.: Capsaicin inhibits in vitro and in vivo
angiogenesis. Cancer Res, 2004, 64, 644–651.
19. Pollet I, Opina CJ, Zimmerman C, Leong KG, Wong F,
Karsan A: Bacterial lipopolysaccharide directly induces
angiogenesis through TRAF6-mediated activation of
NF-kappaB and c-Jun N-terminal kinase. Blood, 2003,
102, 1740–1742.
20. Risau W: Mechanisms of angiogenesis. Nature, 1997,
386, 671–674.
21. Scaldaferri F, Vetrano S, Sans M, Arena V, Straface G,
Stigliano E, Repici A et al.: VEGF-A links angiogenesis
and inflammation in inflammatory bowel disease patho-
genesis. Gastroenterology, 2009, 136, 585–595.
22. Schumann RR, Pfeil D, Lamping N, Kirschning C,
Scherzinger G, Schlag P, Karawajew L et al.: Lipopoly-
saccharide induces the rapid tyrosine phosphorylation of
the mitogen-activated protein kinases erk-1 and p38 in
cultured human vascular endothelial cells requiring the
presence of soluble CD14. Blood, 1996, 87, 2805–2814.
23. Sherer DM, Abulafia O: Angiogenesis during implanta-
tion, and placental and early embryonic development.
Placenta, 2001, 22, 1–13.
24. Shih HC, Hsu CS, Yang LL: In vitro study of the toco-
lytic effect of oroxylin A from Scutellaria baicalensis
root. J Biomed Sci, 2009, 16, 27.
25. Sun Y, Lu N, Ling Y, Gao Y, Chen Y, Wang L, Hu R
et al.: Oroxylin A suppresses invasion through down-
regulating the expression of matrix metalloproteinase-2/9
in MDA-MB-435 human breast cancer cells. Eur J Phar-
macol, 2009, 603, 22–28.
Received: September 29, 2011; in the revised form: April 16, 2012;
accepted: May 25, 2012.
Pharmacological Reports, 2012, 64, 1189�1199 1199
Oroxylin A inhibits LPS induced angiogenesisXiuming Song et al.
























![POLSKIEJ AKADEMII NAUK - min-pan.krakow.pl · TABELA 1.2. Klasyfikacja rosyjskich wêgli kamiennych (GOST 25543-88) Typ Nazwa typu Vdaf [%] Gruboœæ warstwy plastycznej Y [mm] D](https://img.dokumen.tips/doc/110x75/5f9dda6452a8a7421359560a/polskiej-akademii-nauk-min-pan-tabela-12-klasyfikacja-rosyjskich-wgli-kamiennych.jpg)




