Embed Size (px)
Citation preview

Oncogenic herpesvirus hijacks components of the DNArepair machinery to promote its own replication
Xiaofan Li, Ph.D.Bhaduri-McIntosh laboratory

The DNA Damage Response cascade and ATM
Yosef Shiloh et al., Nature Reviews, 2013
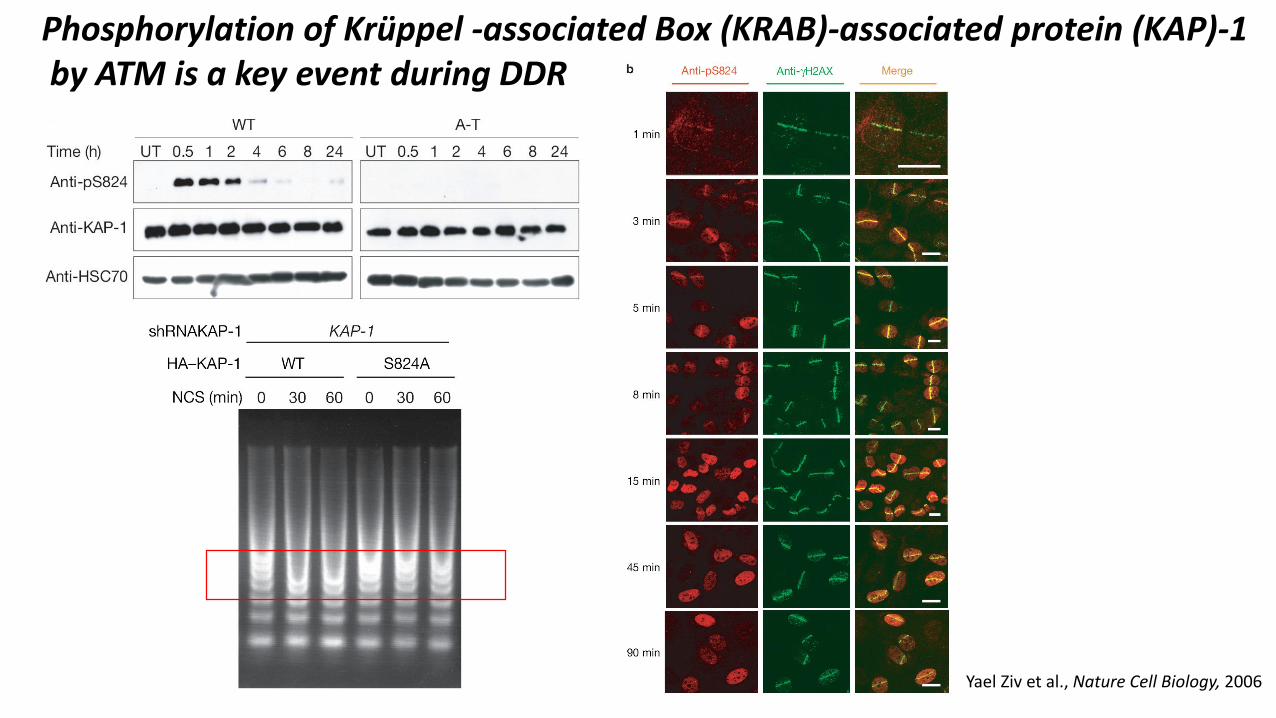
Phosphorylation of Krüppel -associated Box (KRAB)-associated protein (KAP)-1by ATM is a key event during DDR
Yael Ziv et al., Nature Cell Biology, 2006
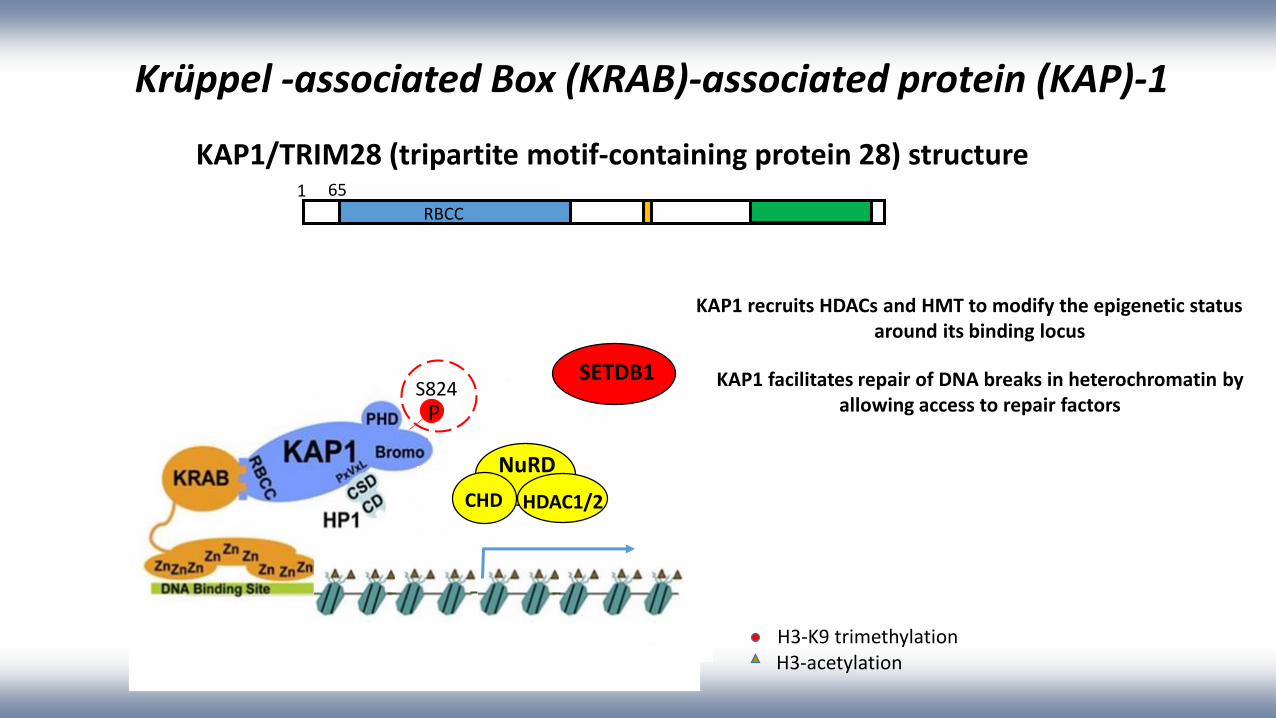
Krüppel -associated Box (KRAB)-associated protein (KAP)-1
376 493 628 801 835
651RBCC
BromoHP1 box
KAP1/TRIM28 (tripartite motif-containing protein 28) structure
KAP1 recruits HDACs and HMT to modify the epigenetic status around its binding locus
KAP1
DAPI
PS824
NuRD
CHD HDAC1/2
SETDB1
PS824
H3-K9 trimethylation H3-acetylation
KAP1 facilitates repair of DNA breaks in heterochromatin by allowing access to repair factors
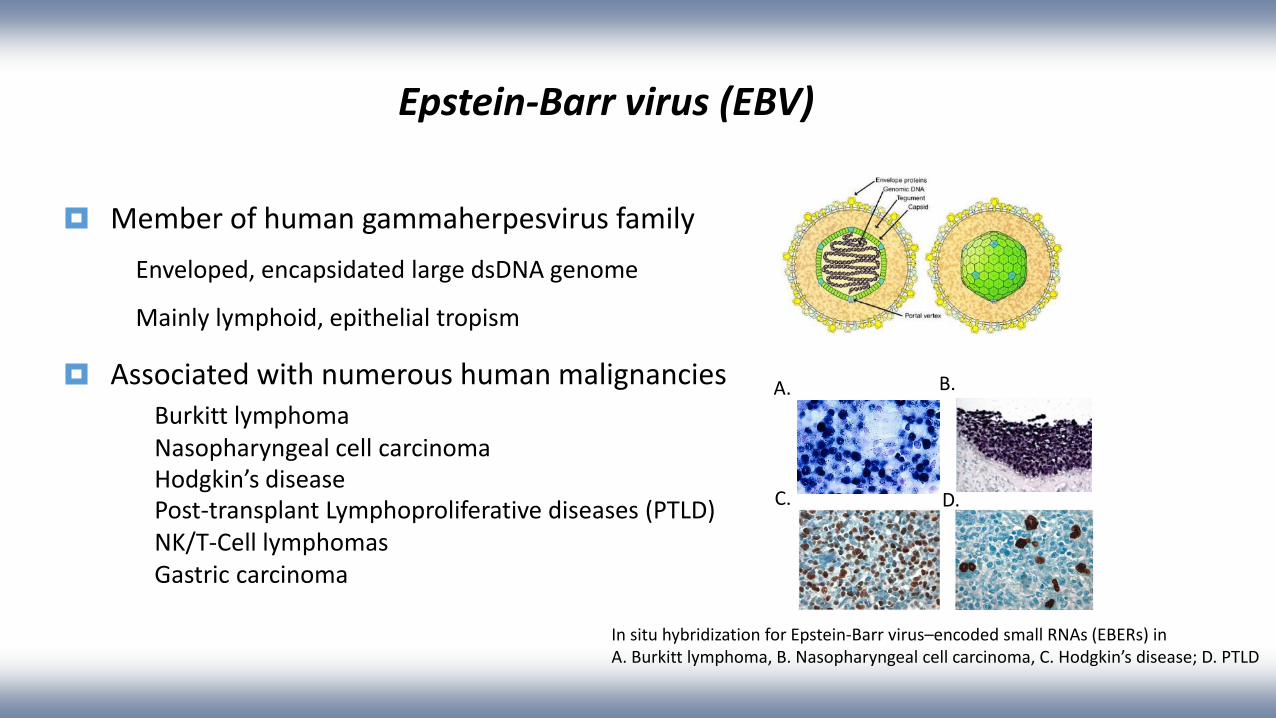
Epstein-Barr virus (EBV)
Member of human gammaherpesvirus family
Enveloped, encapsidated large dsDNA genome
Mainly lymphoid, epithelial tropism
Associated with numerous human malignanciesBurkitt lymphoma Nasopharyngeal cell carcinoma Hodgkin’s diseasePost-transplant Lymphoproliferative diseases (PTLD)NK/T-Cell lymphomasGastric carcinoma
A. B.
C. D.
In situ hybridization for Epstein-Barr virus–encoded small RNAs (EBERs) inA. Burkitt lymphoma, B. Nasopharyngeal cell carcinoma, C. Hodgkin’s disease; D. PTLD

Lytic (re)activationLatent infection
Latency to lytic switch of EBV
Limited viral genes expressed
Viral genome duplicates in S-phase
No progeny production
HDACi, TGF-β, hypoxia, etc
ImmediateEarly
Early Late

Cellular components that regulate latency-to-lytic switch
-STAT3
-PCBP2
-KRAB-ZFPs: large family of transcriptional repressors
thought to function via the KAP1 corepressor
Derek Daigle, et al., JVI, 2010; Erik Hill, et al., JVI, 2013;
Siva Koganti, et al., JVI, 2015;
Mechanisms underlying EBV latency to lytic switch
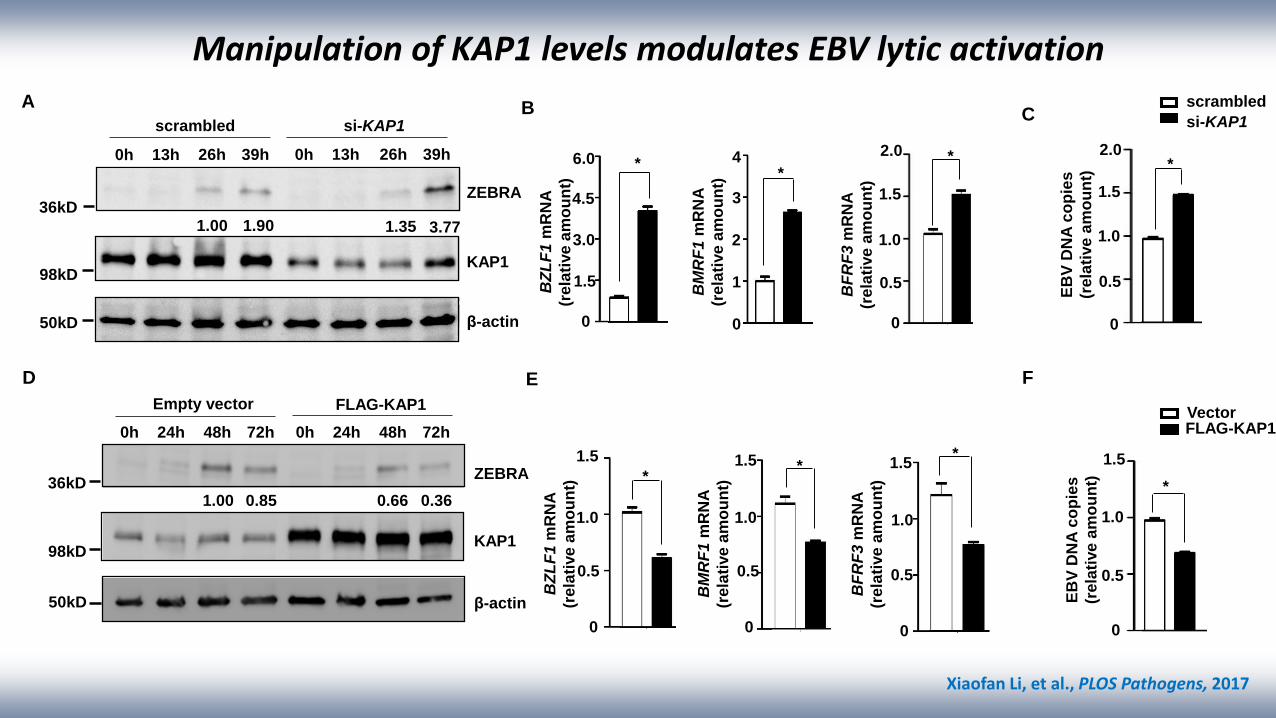
Manipulation of KAP1 levels modulates EBV lytic activation
CA
E
0.5
1.0
1.5
0
0.5
1.0
1.5
0
0.5
1.0
1.5
0
BZ
LF
1m
RN
A
(rela
tive
am
ou
nt)
BM
RF
1m
RN
A
(rela
tive
am
ou
nt)
BF
RF
3m
RN
A
(rela
tive
am
ou
nt)
ZEBRA
KAP1
β-actin
0h 24h 48h 72h
Empty vector FLAG-KAP1
0h 24h 48h 72h
50kD
98kD
36kD1.00 0.85 0.66 0.36
D F
0h 13h 26h 39h
scrambled
ZEBRA
KAP1
β-actin
0h 13h 26h 39h
si-KAP1
50kD
98kD
36kD1.00 3.771.90 1.35
0.5
1.0
1.5
2.0
0
BZ
LF
1m
RN
A
(rela
tive
am
ou
nt)
BM
RF
1m
RN
A
(rela
tive
am
ou
nt)
BF
RF
3m
RN
A
(rela
tive
am
ou
nt)
0
1.5
*
3.0
4.5
6.0
0
1
2
3
4
0.5
1.0
1.5
0
EB
V D
NA
co
pie
s
(rela
tive
am
ou
nt)
0.5
1.0
1.5
2.0
0
EB
V D
NA
co
pie
s
(rela
tive
am
ou
nt)
B
**
*
**
*
*
scrambled
si-KAP1
VectorFLAG-KAP1
Xiaofan Li, et al., PLOS Pathogens, 2017
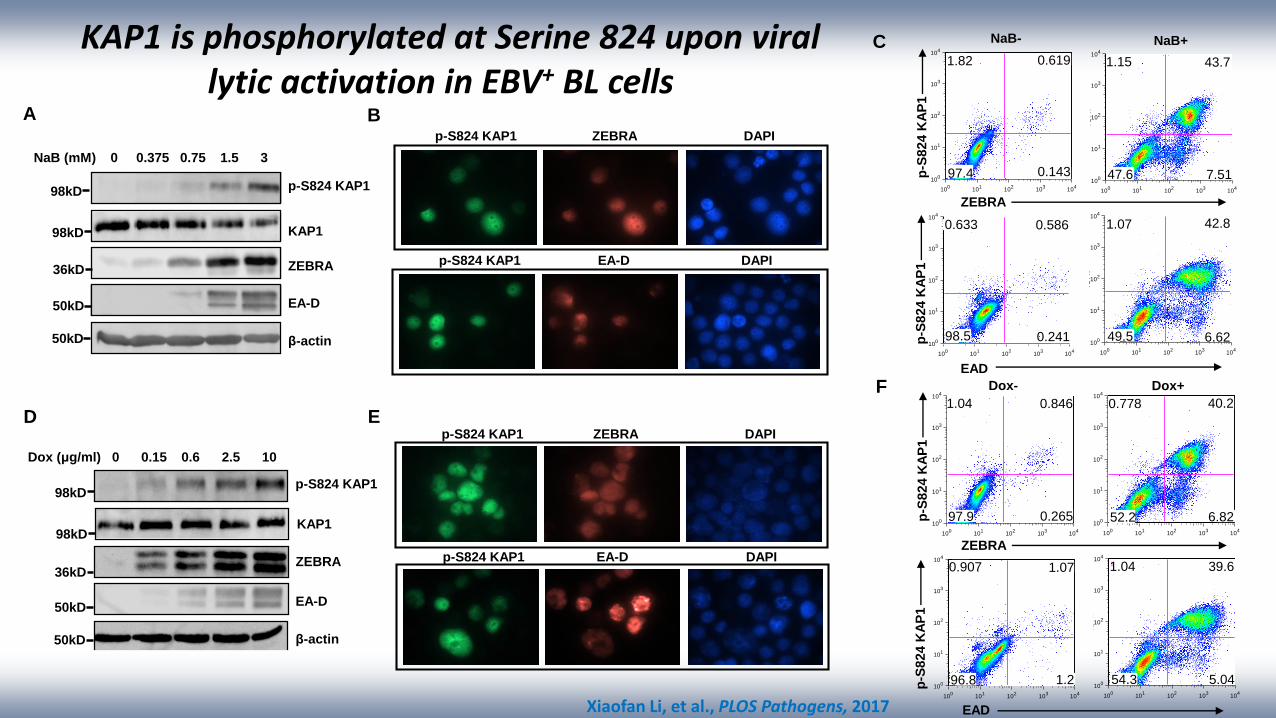
A
p-S824 KAP1
KAP1
β-actin
ZEBRA
EA-D
98kD
98kD
36kD
50kD
50kD
NaB (mM) 0 0.375 0.75 1.5 3
p-S824 KAP1
KAP1
β-actin
ZEBRA
EA-D
98kD
98kD
36kD
50kD
50kD
Dox (μg/ml) 0 0.15 0.6 2.5 10
p-S824 KAP1 ZEBRA DAPI
p-S824 KAP1 EA-D DAPI
B
D E
F
100
101
102
103
104
100
101
102
103
104
006
FL1-H
FL
4-H
0.778 40.2
6.8252.2
100
101
102
103
104
100
101
102
103
104
005
FL1-H
FL
4-H
1.04 0.846
0.26597.9
100
101
102
103
104
100
101
102
103
104
010
FL1-H
FL
4-H
1.04 39.6
5.0454.3
100
101
102
103
104
100
101
102
103
104
009
FL1-H
FL
4-H
0.907 1.07
1.296.8
1.04 0.846
97.9 0.265
0.778 40.2
52.2 6.82
0.907 1.07
96.8 1.2
1.04 39.6
54.3 5.04
ZEBRA
p-S
824 K
AP
1
EAD
p-S
824 K
AP
1
Dox- Dox+
EAD
100
101
102
103
104
100
101
102
103
104
012
FL1-H
FL
4-H
1.07 42.8
6.6249.5
100
101
102
103
104
100
101
102
103
104
007
FL1-H
FL
4-H
1.82 0.619
0.14397.4
100
101
102
103
104
100
101
102
103
104
008
FL1-H
FL
4-H
1.15 43.7
7.5147.6
100
101
102
103
104
100
101
102
103
104
011
FL1-H
FL
4-H
0.633 0.586
0.24198.5
1.82 0.619
97.4 0.143
1.15 43.7
47.6 7.51
0.633 0.586
98.5 0.241
1.07 42.8
49.5 6.62
ZEBRA
p-S
824 K
AP
1
NaB- NaB+
p-S
824 K
AP
1
KAP1 is phosphorylated at Serine 824 upon virallytic activation in EBV+ BL cells
C
p-S824 KAP1 ZEBRA DAPI
p-S824 KAP1 EA-D DAPI
Xiaofan Li, et al., PLOS Pathogens, 2017
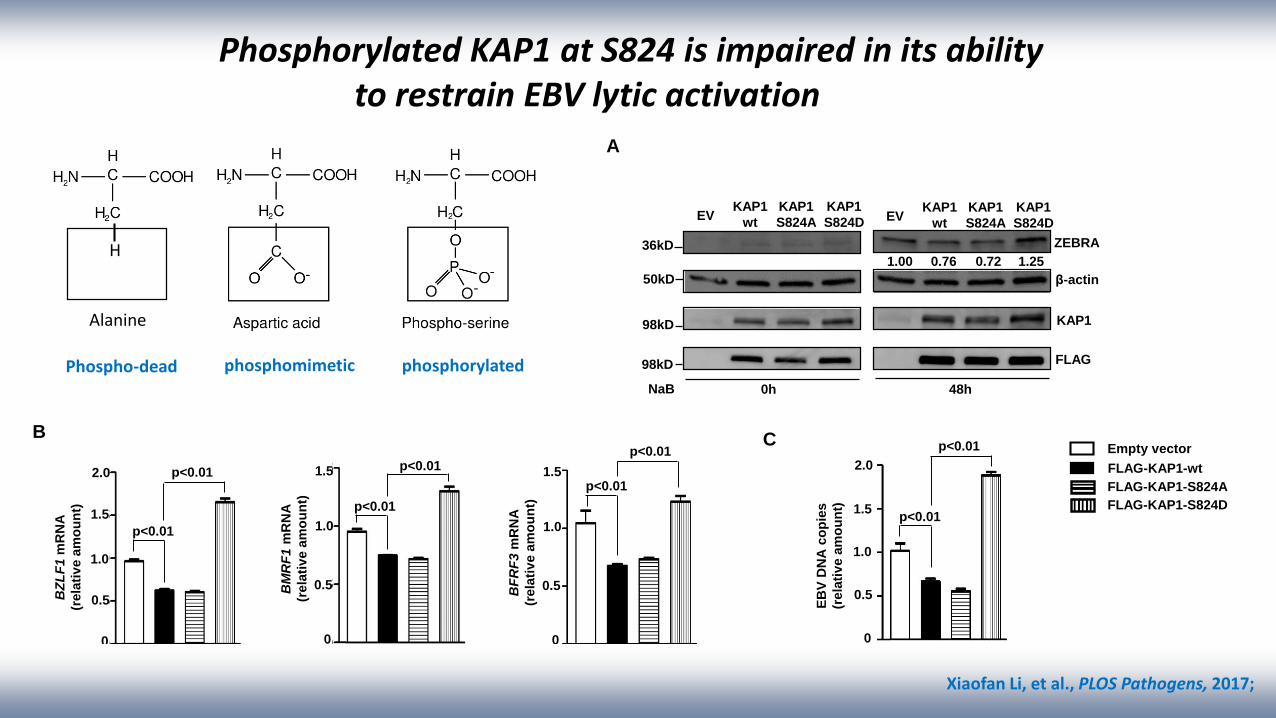
36kD
50kD
98kD
98kD
ZEBRA
β-actin
KAP1
FLAG
0h 48h
1.00 0.76 0.72 1.25
NaB
EVKAP1
wt
KAP1
S824A
KAP1
S824D
A
0.5
1.0
1.5
0
BM
RF
1m
RN
A
(rela
tiv
e a
mo
un
t)
p<0.01
p<0.01
BF
RF
3m
RN
A
(rela
tiv
e a
mo
un
t)
0.5
1.0
1.5
0
p<0.01
p<0.01
0.5
1.0
1.5
2.0
0
BZ
LF
1m
RN
A
(rela
tiv
e a
mo
un
t)
p<0.01
p<0.01
B
0.5
1.0
1.5
2.0
0
EB
V D
NA
co
pie
s
(rela
tiv
e a
mo
un
t)
p<0.01
p<0.01
C
Figure 4
EVKAP1
wt
KAP1
S824A
KAP1
S824D
Phosphorylated KAP1 at S824 is impaired in its ability to restrain EBV lytic activation
C
H
Alanine
Phospho-dead phosphomimetic phosphorylated
Xiaofan Li, et al., PLOS Pathogens, 2017;
FLAG-KAP1-wt
Empty vector
FLAG-KAP1-S824A
FLAG-KAP1-S824D
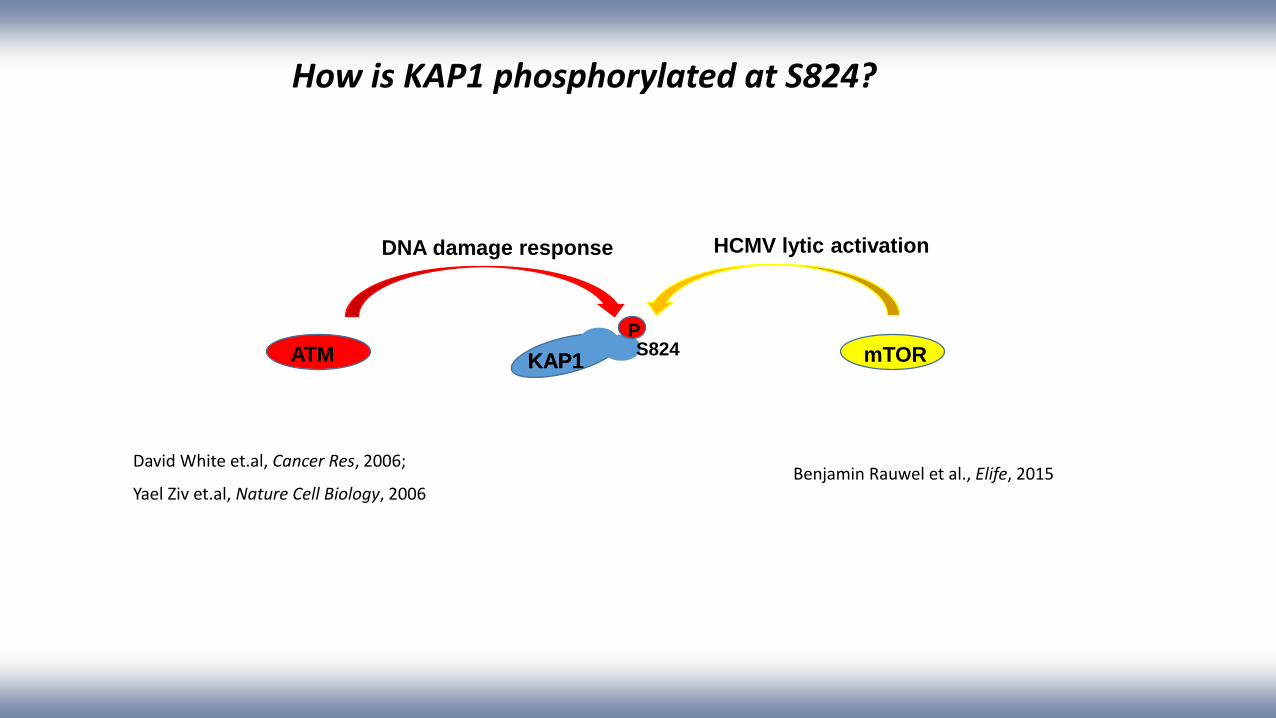
How is KAP1 phosphorylated at S824?
PS824
KAP1KAP1ATM mTOR
HCMV lytic activationDNA damage response
David White et.al, Cancer Res, 2006;
Yael Ziv et.al, Nature Cell Biology, 2006Benjamin Rauwel et al., Elife, 2015
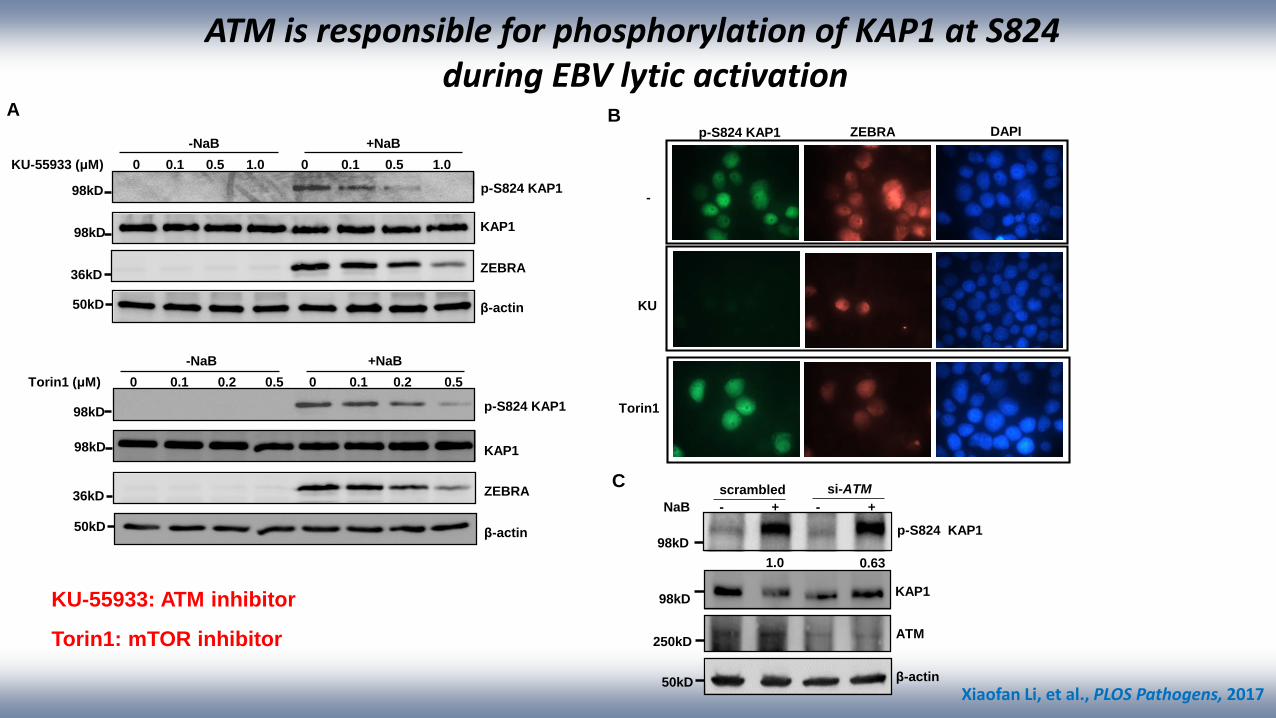
ATM is responsible for phosphorylation of KAP1 at S824 during EBV lytic activation
A
C
p-S824 KAP1
KAP1
β-actin
ATM
98kD
NaB - + - +
scrambled si-ATM
1.0 0.63
98kD
250kD
50kD
p-S824 KAP1
KAP1
β-actin
ZEBRA
KU-55933 (μM) 0 0.1 0.5 1.0 0 0.1 0.5 1.0
-NaB +NaB
98kD
98kD
36kD
50kD
p-S824 KAP1
KAP1
β-actin
ZEBRA
Torin1 (μM) 0 0.1 0.2 0.5 0 0.1 0.2 0.5
-NaB +NaB
98kD
98kD
36kD
50kD
B
Torin1
KU
p-S824 KAP1 ZEBRA DAPI
-
Xiaofan Li, et al., PLOS Pathogens, 2017
KU-55933: ATM inhibitor
Torin1: mTOR inhibitor
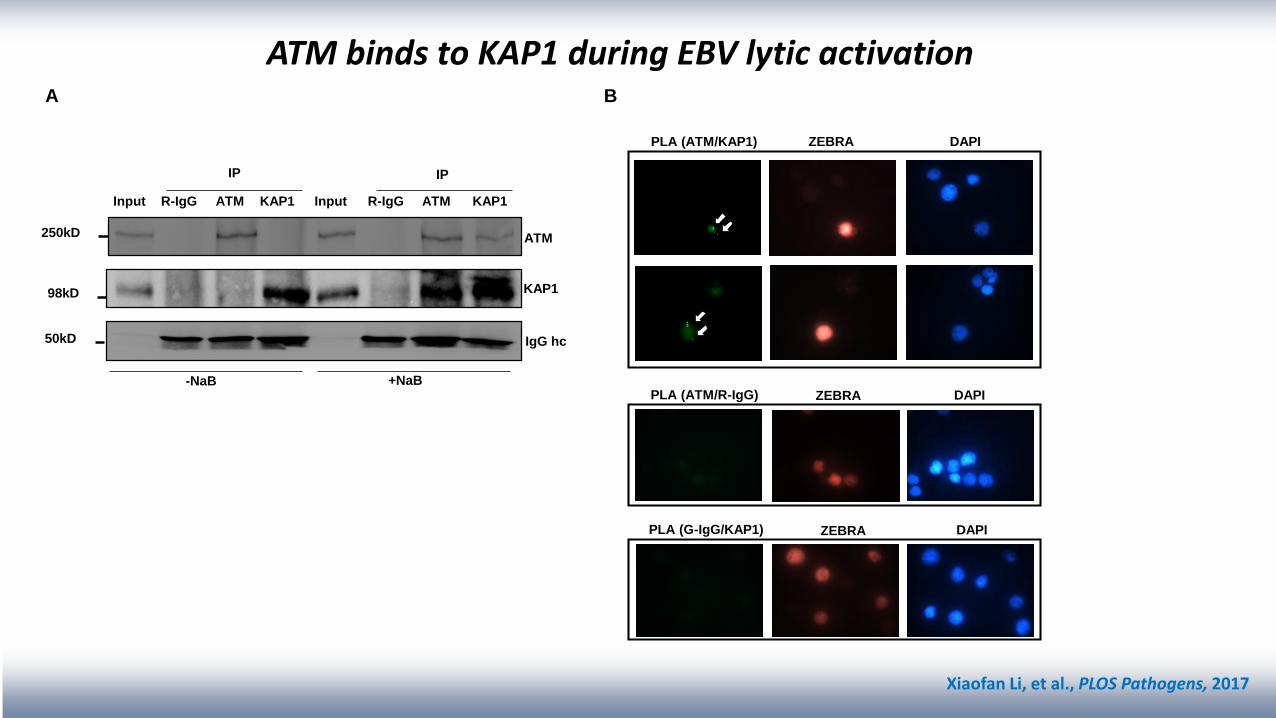
Input R-IgG ATM KAP1 Input R-IgG ATM KAP1
IP IP
ATM
KAP1
IgG hc
250kD
98kD
50kD
-NaB +NaB
DAPI
BA
PLA (ATM/KAP1) ZEBRA DAPI
DAPIPLA (ATM/R-IgG) ZEBRA
PLA (G-IgG/KAP1) ZEBRA
ATM binds to KAP1 during EBV lytic activation
Xiaofan Li, et al., PLOS Pathogens, 2017

KAP1KAP1 S824
p
ZEBRA
ATMViral lytic product
(IE, E, L)
EBV genome EBV genome
Viral lytic product
(IE, E, L)
i. ii.
Working model
Chloroquine
F
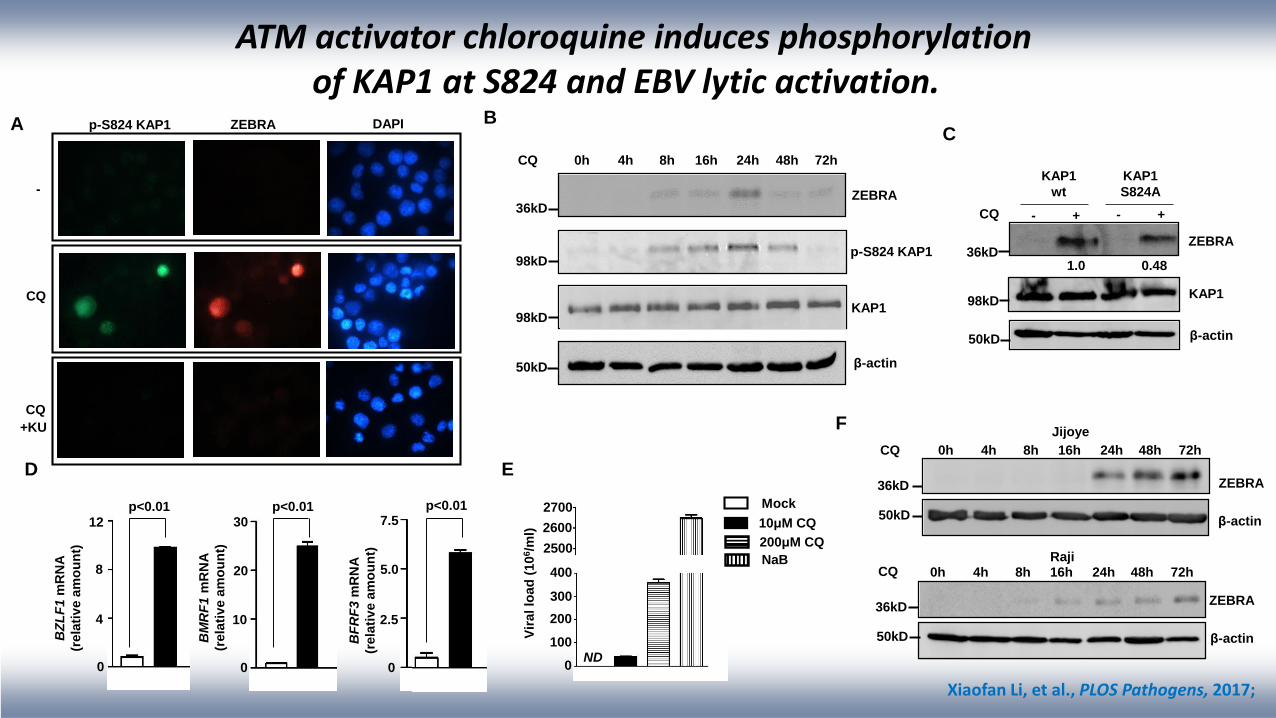
ATM activator chloroquine induces phosphorylation of KAP1 at S824 and EBV lytic activation.
C
CQ 0h 4h 8h 16h 24h 48h 72h
ZEBRA
β-actin
p-S824 KAP1
KAP1
36kD
98kD
98kD
50kD
B
ZEBRA
β-actin
KAP1
CQ - + - +
KAP1
wt
KAP1
S824A
1.0 0.48
98kD
50kD
36kD
A
-
CQ
ZEBRA DAPIp-S824 KAP1
CQ
+KU
BZ
LF
1m
RN
A
(rela
tiv
e a
mo
un
t)
0
4
8
12
0
10
20
30
BM
RF
1m
RN
A
(rela
tiv
e a
mo
un
t)
0
2.5
5.0
7.5
BF
RF
3m
RN
A
(rela
tiv
e a
mo
un
t)
p<0.01 p<0.01 p<0.01
D E
2700
2600
2500
400
300
100
0
200
Vir
al lo
ad
(10
6/m
l)
NDG
ZEBRA
β-actin
CQ 0h 8h 24h 72h4h 16h 48h
36kD
50kD
0h 8h 24h 72h4h 16h 48h
Jijoye
Raji
ZEBRA
β-actin
CQ
36kD
50kD
Xiaofan Li, et al., PLOS Pathogens, 2017;
10μM CQ
Mock
NaB
200μM CQ
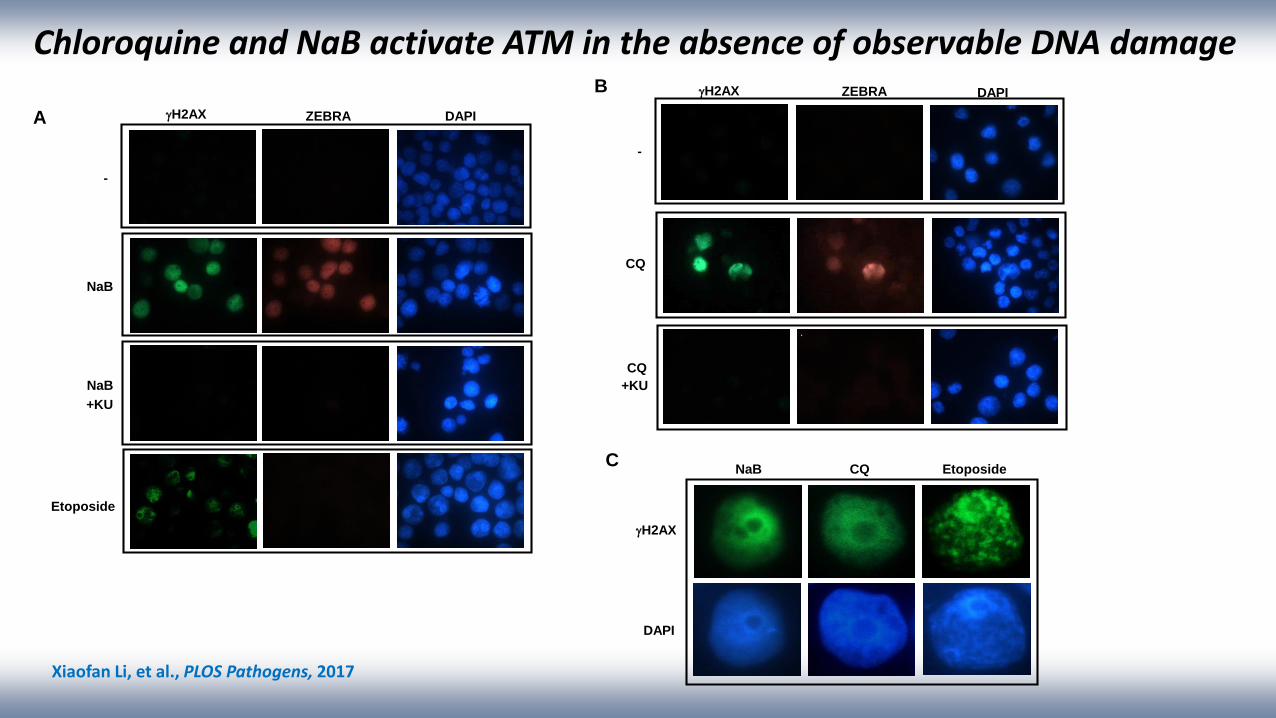
ZEBRA DAPIgH2AX
-
NaB
NaB
+KU
Etoposide
A
B
NaB EtoposideCQ
gH2AX
DAPI
C
-
gH2AX ZEBRA DAPI
CQ
+KU
CQ
Chloroquine and NaB activate ATM in the absence of observable DNA damage
Xiaofan Li, et al., PLOS Pathogens, 2017

Conclusion
The cancer-causing virus EBV exploits a cellular mechanism (involving ATM and KAP1)
that repairs breaks in heterochromatin to promote its own replication

Acknowledgements
Collaborators:
Jizu Zhi, Ph.D.FDA, USA
Funding:
NIAID, NIHStony Brook Research Foundation
Sumita Bhaduri-McIntosh, M.D., Ph.D.
Siva Koganti, Ph.D.
Eric Burton
Lab members:
Ramon Perez
Sandeepta Burgula, Ph.D
Salvatore Spadaro






![REVIEW Open Access The modulation of apoptosis by oncogenic … · 2017. 8. 25. · transmissible oncogenic pathogen [4], and in 1932, Shope and Hurst demonstrated the oncogenic activity](https://img.dokumen.tips/doc/110x75/60a5adee03abc344316eb0df/review-open-access-the-modulation-of-apoptosis-by-oncogenic-2017-8-25-transmissible.jpg)






















