Embed Size (px)
Citation preview

Q 1986 by The Amerrcan Society of Biological Chemists, Inc. THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 261, No. 2. Issue of January 15,gp: 631-637 1986
rlnted tn LkS.A.
The Purified Q Subunits of Go and Gi from Bovine Brain Require pr for Association with Phospholipid Vesicles”
(Received for publication, May 14, 1985)
Paul C. Sternweis$ From the Department of Pharmacology, University of Texas Health Science Center at Dallas, Dallas, Texas 75235
The purified G-proteins from bovine brain were ex- amined for potential solubility in the absence of deter- gent. The isolated a, and ai subunits migrated through sucrose with rates consistent with the existence of monomeric species either in the presence or the absence of cholate. The fly subunits or holo-G-proteins aggre- gated extensively if cholate was absent. AI3+, Mg2+, and F- prevented the aggregation of a. and ai caused by the addition of fly and could also prevent the aggre- gation of as when G. was examined at higher tempera- ture.
The association of subunits with phospholipid vesi- cles was examined. Whereas By associated totally with phospholipid vesicles, purified a. showed little inter- action. a. did bind to vesicles containing By (& vesicles) in a saturable fashion that indicated a stoichiometric association between the subunits. Treatment with guanosine 6‘-(3-O-thio)triphosphate could partially dissociate a, that was bound to fly vesicles.
These data suggest that B y may be an anchor for association of Q subunits with membranes and that regulation by these proteins may not be limited to the plasma membrane. This possibility and its implications are discussed. The reversible association of a,, to vesicles may provide a very sensitive system for the study of the interactions between these subunits.
G,,‘, Gi, Go, and Gt comprise a family of homologous pro- teins (G-proteins) that are associated with membranes and are involved in signal transduction (for reviews, see Ref. 1- 5). Three of these proteins are known to regulate enzymes that appear to reside in the same membranes as the respective G-proteins. Thus, G , and Gi mediate stimulation or inhibition of adenylate cyclase by hormones, a process thought to occur in the plasma membrane. G, stimulates cGMP-dependent
* This investigation was supported by National Institutes of Health Grant GM31954 and March of Dimes Foundation Grant 5-435. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Recipient of Searle Scholars Award. The abbreviations used are: G, and Gi, regulatory proteins that
mediate stimulation and inhibition of adenylate cyclase, respectively; Go, a similar GTP-binding protein of unknown function; G, (also called transducin), the GTP-binding protein from rod outer segments; G-proteins (sometimes referred to as N-proteins), any GTP-binding protein that resembles a family of homologous proteins composed of G., G , Go, and G,; cy, 6, and y, the subunits of the G-proteins in decreasing size; Goa or a,, example of nomenclature to designate a specific subunit of the indicated G-protein; GTPyS, guanosine 5’-(3- 0thio)triphosphate; Hepes, 4-(2-hydroxyethyl)-l-piperazineethane- sulfonic acid; SDS, sodium dodecyl sulfate; AMF indicates the pres- ence of 50 gM AlC13,lO mM MgSO,, and 10 mM NaF.
phosphodiesterase in response to activation of rhodopsin by light; all three of the proteins involved in this process are associated with disc membranes in rod outer segments of vertebrates. Go is a fourth G-protein (6) which has been shown to interact with muscarinic receptors (7). The function of Go remains to be determined.
The G-proteins show remarkable similarities in both struc- ture and mechanisms of action (2-7). Each of the G-proteins contains three subunits (a, @, and y), binds guanine nucleo- tides with high affinity, and possesses a GTPase activity that is implicated in function. The binding site for guanine nu- cleotide is contained in the a subunit (6, 8-11). Experiments with purified subunits have suggested that the a subunits are the active species in regulation (10-12). Activation of the purified proteins coincides with dissociation of the subunits @,lo, 11,13,14). The fly subunits have been suggested to act as inhibitors of activation of the N subunits in hormone- regulated systems (3, 12, 15, 16) or as stimulators in the photoreceptor system (17). The /3y complex enhances the interaction of a, with muscarinic receptors (7) and seems to be essential for binding and activation of at by rhodopsin (17).
While the a subunits vary in size and appear to differ functionally, as, at, and ai have been shown to have similar amino acid compositions and patterns of proteolytic cleavage (18). The proteolytic profiles of the P-polypeptides were in- distinguishable (18). More recent evidence with antibodies has confirmed both similarities and differences in structure. Thus, antisera raised against G, recognized the P but not the y subunits of Gi and G, (19). Roof and colleagues (20) have also observed these results, but demonstrated that only the larger f l subunit, fl36, of the two found in G,, Gi, and Go was cross-reactive with antibodies for pt. They also demonstrated that a,,, but not ai or as, was recognized strongly by an antiserum raised to G, if the subunits were examined by blotting in the absence of SDS.
The similarity in the structural properties and mechanistic behavior of the G-proteins is marked by another major difference between G, and the other G-proteins. Whereas G, can be dissociated from membranes by the addi- tion of guanine nucleotides (Zl), the other G-proteins appear more firmly associated with membranes and require detergent for extraction (1, 3-6). G,, Go, and Gi have, therefore, always been purified in detergent. This study examines in detail the behavior of purified Gi and Go from bovine brain in the absence of detergent. While purified Go was reported to ag- gregate in the absence of detergent (61, the isolated ar subunits of this protein and Gi but not the fly subunits remain mon- odisperse under these conditions. This potential solubility of a,, has been suggested by others (22). These data and initial experiments that examine the association of these subunits
63 I

632 Solubility of a but Not Py Subunits of Go and Gi
with phospholipid vesicles are presented and the implications discussed.
EXPERIMENTAL PROCEDURES
Preparation of Membranes and G-proteins-Membranes from bo- vine brain (6), bovine rod outer segments (23), and the G.-deficient (cyc-) mutant of the S49 lymphoma cell (24) were prepared as described. The two major GTP-binding proteins from bovine brain (Gi and Go) were purified by the method of Sternweis and Robishaw (6). G. was also purified from bovine brain using the same procedures as those described for Gi and Go, but with a final step of purification on hydroxylapatite (20). G, was prepared from bovine rod outer segments (25). The isolated a and By subunits of Gi and Go and the By subunits of Gt were prepared by chromatography of the purified proteins through heptylamine-Sepharose in the presence of Ala+, M$+, and F- (AMF) as reported (16) with modifications (20). The a subunits of Go and Gi were concentrated by pressure filtration through an Amicon PM-30 membrane and stored either on ice or frozen at -80 "C. The AMF that remained in the samples helped to stabilize the proteins during storage and was rendered ineffective by dilution during experimentation. The By subunits were filtered free of AMF by passage through (2-50 Sephadex (Pharmacia) into 20 mM Na- Hepes, pH 8.0, 1 mM EDTA, 1 mM dithiothreitol, and 25 mM sodium cholate prior to storage on ice or frozen at -80 "C.
Sucrose Density Gradients-The different G-proteins and their subunits were sedimented through gradients of sucrose (520%) in 200 mM NaHepes, pH 8.0, 2 mM EDTA, and 1 mM dithiothreitol. Sodium cholate (25 mM), NaCl (100 mM), NaF (10 mM), MgSO, (IO mM), and AICls (50 pM) were included in specific experiments as indicated. Gradients were subjected to centrifugation in an SW60 rotor (Beckman) for 15 h at 50,000 rpm and 1 "C (except Fig. 5). Gradients were separated into approximately 20 fractions and ana- lyzed. Values of ~ 2 0 , ~ for the G-proteins were determined directly from the migration of calibrating proteins, which included horse heart lactic dehydrogenase ( S ~ O , ~ , 7.321, pig heart malic dehydrogenase ( ~ 2 0 , ~ .
4.3), horseradish peroxidase 3.47), and horse heart myoglobin ( S Z ~ , ~ , 2.0). Standard proteins were not included in experiments where fractions were analyzed for subunits of G-proteins by SDS-polyacryl- amide gel electrophoresis; the migration distances of G-proteins, as determined by assays of activity, were the same as when calibration proteins were included.
Incorporation of By into Phospholipid Vesicles-Egg phosphatidyl- choline (10 mg/ml) and sodium cholate (5 mg/ml) were sonicated to clarity in Solution A 200 mM NaHepes, pH 8.0, 1 mM EDTA, and 1 mM dithiothreitol. An aliquot (200 pl) of this mixture was combined with 50 pl of purified By (1 mg/ml) a t 0 'C. Every 10 min, this mixture was diluted with 200 pl of Solution A until a total volume of 1.25 ml was obtained. The diluted sample was applied to a column (7 ml) of Ultrogel AcA-34 (LKB) and eluted with Solution A; fractions of 0.5 ml were collected. Both the phospholipid and protein were obtained in fractions corresponding to the excluded volume of the column; these fractions were pooled for use, and the preparation referred to as 07 vesicles. Phospholipid vesicles without By were made by the same procedure.
Intermtion of a Subunits with By Vesicles-A column (7 ml) of Ultrogel AcA-34 was utilized to separate phospholipid vesicles from free a subunits. Thus, the vesicles were essentially excluded from the gel matrix, while the a subunits were included. Samples of phospho- lipid vesicles (either with or without By) were mixed with a subunits in a total volume of 0.5 ml of Solution A. The samples were incubated successively for 15 min at 20 'C and at least 30 min at 0 "C, applied to AcA-34, and eluted with Solution A. Fractions of 0.5 ml were collected and analyzed for GTPyS-binding activity. Binding sites (a subunit) that eluted at the excluded volume of the column along with phospholipid were considered associated with the vesicles.
Assays-The a subunits of Gi and Go were measured by their ability to bind GTPrS (6, 26). The By subunits were assayed by their capability to increase the rate of deactivation of G. (E), and G, was measured by reconstitution of adenylate cyclase activity in cyc- membranes (13). One unit of G. activity is defined as the amount of G. that will reconstitute the production of 1 nmol of cyclic AMP/ min. The presence of subunits of G-proteins was also assessed by electrophoresis of samples through SDS-polyacrylamide gels using the discontinuous system described by Laemmli (27) and staining with silver (28). Samples for electrophoresis were alkylated with N- ethylmaleimide by published procedures (6) . Proteins were deter-
mined by staining with amido black (29), with bovine serum albumin as the standard. Phospholipid was measured as phosphate after hydrolysis with perchloric acid (30).
RESULTS
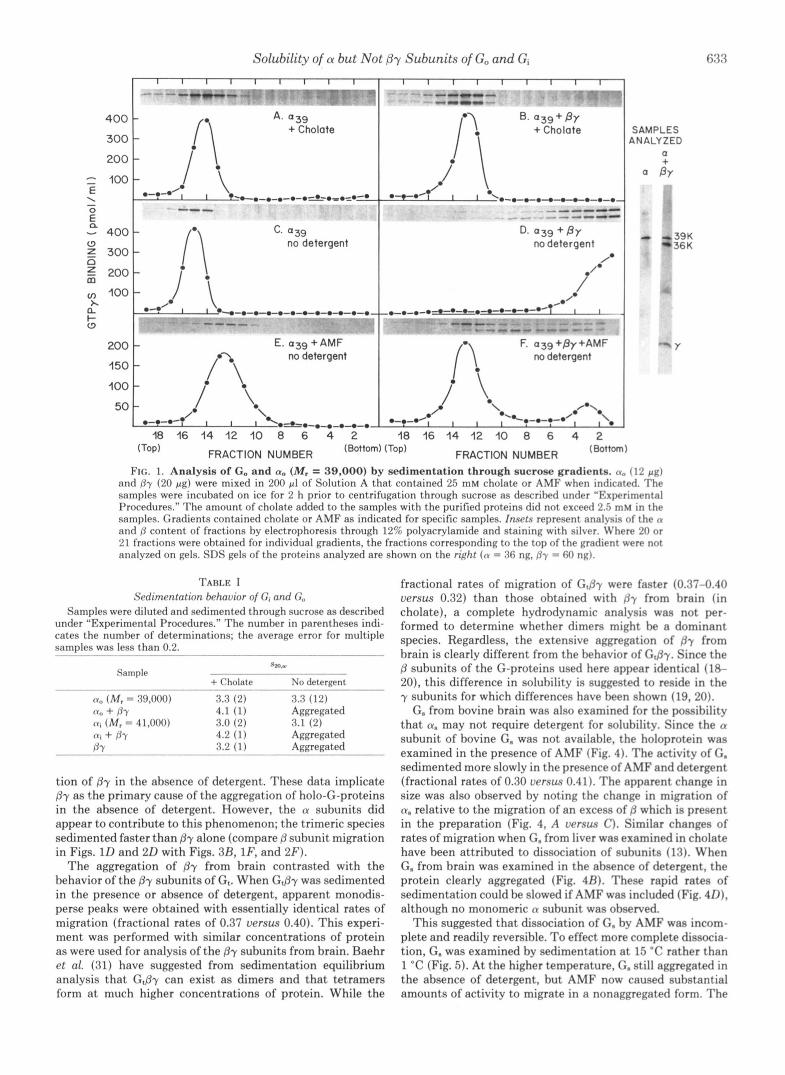
Three of the G-proteins are purified in detergents after extraction from membranes. The exception to this is G,, which can be dissociated from membranes of rod outer segments with light and GTP. Attempts to extract the G-proteins from membranes of bovine brain in the absence of detergent have failed. Therefore, it was somewhat unexpected but of great interest to find that the purified a subunits of Go and Gi from brain remain soluble in the absence of any detergent. This behavior is demonstrated for a, in Fig. 1 where potential aggregation of the subunit was examined by sedimentation through sucrose. In either the presence or absence of cholate, the migration rate of a,, was identical. This migration distance is consistent with a, existing as a monomeric species in either condition. Table I shows sedimentation values that were determined when standards were included in other experi- ments.
The behavior of a, contrasts with the behavior of holo-Go. When By was added to a,, the trimeric holoprotein was formed as evinced by a small increase in the rate of sedimentation of a, (GTPyS-binding sites) through sucrose in the presence of cholate (Fig. 1B and Table I). If the detergent was not included, the Go formed aggregates with very large sedimen- tation rates (Fig. 1D). This aggregation could be reversed by inclusion of A13+, M$+, and F- (AMF) in the gradient, a condition believed to cause dissociation of G-proteins into a and Py subunits (11, 13, 16). Thus, in AMF (Fig. lF), a large portion of the a, subunit migrated slowly with a sedimentation rate consistent with that of a monomeric species; however, the bulk of the /3 subunits still aggregated (Fig. lF, gel).
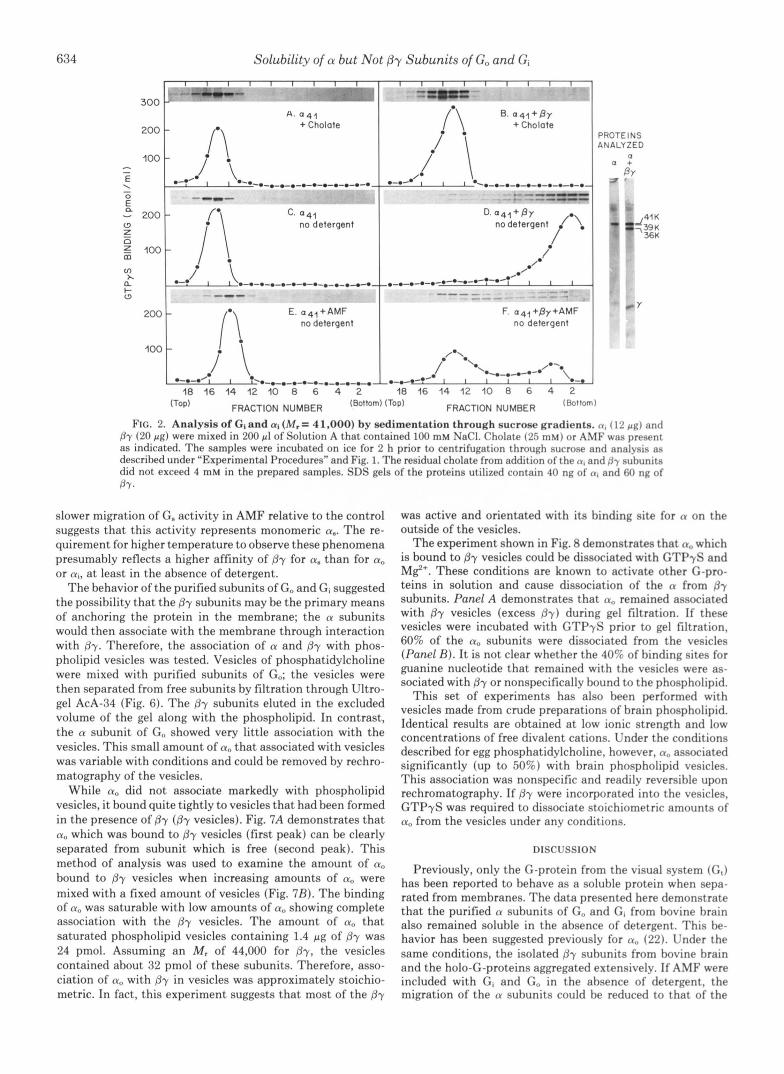
The same set of experiments was performed with purified ai from brain (Fig. 2). Purified ai also remained monomeric in the absence of detergent, but aggregated extensively when the /3y subunits were included. In the presence of cholate, the apparent trimeric species (Gi) remained soluble, but sedi- mented faster than (Y, alone (Table I). The presence of AMF was again effective in reversing the aggregation of ai in the presence of By, but in the absence of detergent (Fig. 2F). The apparent dissociation of ai and Py was incomplete and prob- ably reflects an equilibrium of association among the compo- nents. This result was obtained with gradients run in the presence of 100 mM NaCl, a condition thought to stabilize the dissociated form of G-proteins (11). In the absence of NaC1, ai showed poor reversal if examined as in Fig. 2F; this con- trasts with the good reversal observed with a. in the absence of NaCl (Fig. 1F). The difference in behavior of a, and ai may reflect a higher affinity of /3y for (yi than a,,. This is consistent with the isolation of a, but not ai during purification of G- proteins from bovine brain in the absence of activators (6).
The solubility of the Py subunits from brain was also examined (Fig. 3). In cholate, Py migrated as an apparent single species with a sedimentation value of 3.2 (Table I). This is similar to values obtained for from rabbit liver (11) and suggests that the primary species may be a dimer of P and y. If detergent is removed from the gradient, the 07 subunits sedimented at higher rates. The gel strips show only the B subunits in Fig. 3, B and C, as the y subunit stained poorly and was not convincingly observed after apparent aggregation and further dilution through the gradient. There- fore, while y aggregated (based on its disappearance), it was not clear that it distributed evenly with the P subunit during aggregation. As expected, AMF had no effect on the aggrega-
Solubility of CY but Not 0-y Subunits of Go and Gi 633
400
300
200
= 100 E \
0 -
- 400 Z 300 n
5 0
2 200 100
h a i- (3
ANALYZED SAMPLES
a + " BY
18 16 14 12 10 8 6 4 2 18 16 14 12 10 8 6 4 2 (Top) FRACTION NUMBER FRACTION NUMBER
FIG. 1. Analysis of Go and a, (MI = 39,000) by sedimentation through sucrose gradients. (Y, (12 pg) and Py (20 pg) were mixed in 200 pl of Solution A that contained 25 mM cholate or AMF when indicated. The samples were incubated on ice for 2 h prior to centrifugation through sucrose as descrihed under "Experimental Procedures." The amount of cholate added to the samples with the purified proteins did not exceed 2.5 mM in the samples. Gradients contained cholate or AMF as indicated for specific samples. Insets represent analysis of the n and /j cont.ent of fractions by electrophoresis through 12% polyacrylamide and staining with silver. IVhere 21) or 21 fract.ions were ohtained for individual gradients, the fractions corresponding to the top of the gradient were not analyzed on gels. SDS gels of the proteins analyzed are shown on the rcght (CY = 36 ng. /jy = 61) ng).
36 K 39K
TABLE I Sedimentation behavior of C; and G"
Samples were diluted and sedimented through sucrose as described under "Experimental Procedures." The numher in parentheses indi- cates the numher of determinations; the average error for multiple samples was less than 0.2. ~
Sample Sm.u
+ Cholate No detergent "
(Y" (M, = 39,000) 3.3 (2) 3.3 (12) n" + /j-Y 4.1 (1) Aggregated
1I; + /fy 4.2 (1) Aggregated (I, (M. = 41,000) 3.0 (2) 3.1 (2)
BY 3.2 - (1) Aggregated ~~
tion of Py in the absence of detergent. These data implicate By as the primary cause of the aggregation of holo-G-proteins in the absence of detergent. However, the n subunits did appear to contribute to this phenomenon; the trimeric species sedimented faster than By alone (compare p subunit migration in Figs. ID and 2 0 with Figs. 3R, lF, and 2 F ) .
The aggregation of By from brain contrasted with the behavior of the By subunits of G,. When G,@y was sedimented in the presence or absence of detergent, apparent monodis- perse peaks were obtained with essentially identical rates of migration (fractional rates of 0.37 uersu.9 0.40). This experi- ment was performed with similar concentrations of protein as were used for analysis of the (37 subunits from brain. Baehr et al. (31) have suggested from sedimentation equilibrium analysis that G,@y can exist as dimers and that tetramers form a t much higher concentrations of protein. While the
fractional rates of migration of G,py were faster (0.37-0.40 uersw 0.32) than those obtained with dy from brain (in cholate), a complete hydrodynamic analysis was not per- formed to determine whether dimers might be a dominant species. Regardless, the extensive aggregation of fly from brain is clearly different from the behavior of G,py. Since the B subunits of the G-proteins used here appear identical (18- 20), this difference in solubility is suggested to reside in the y subunits for which differences have been shown (19, 20).
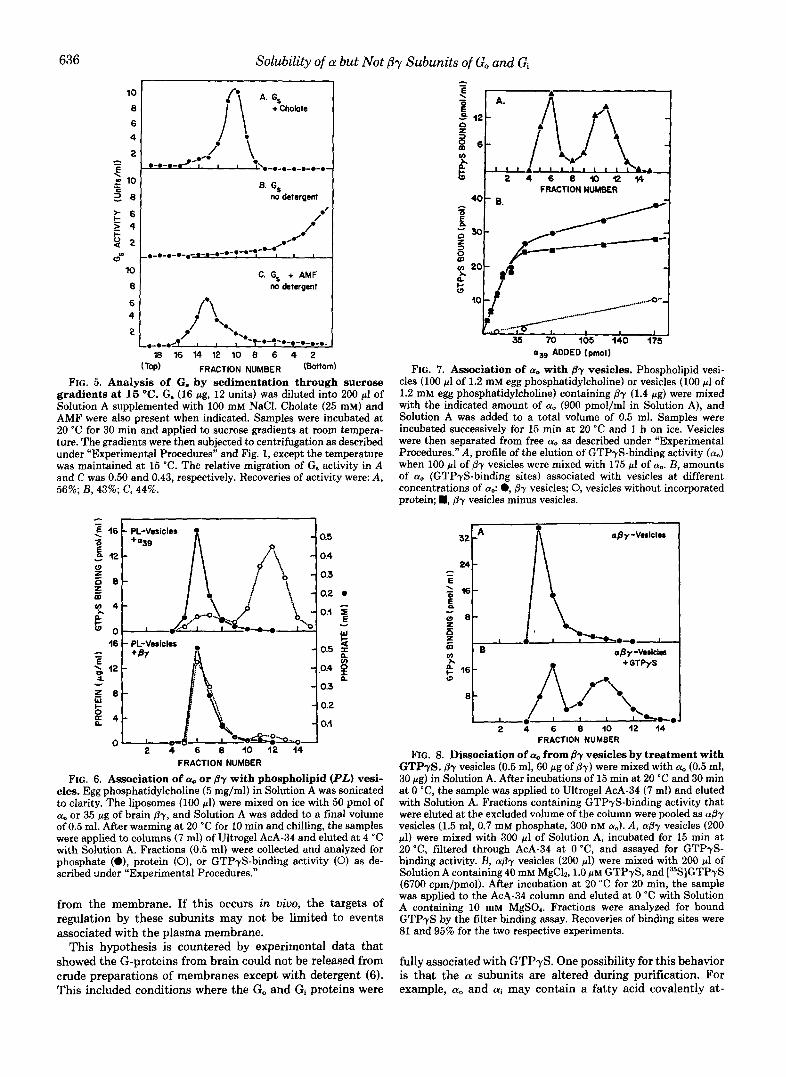
G. from bovine brain was also examined for the possibility that n. may not require detergent for solubility. Since the rn subunit of bovine G. was not available, the holoprotein was examined in the presence of AMF (Fig. 4). The activity of G. sedimented more slowly in the presence of AMF and detergent (fractional rates of 0.30 uersus 0.41). The apparent change in size was also observed by noting the change in migration of n. relative to the migration of an excess of /j which is present in the preparation (Fig. 4, A uersu.9 C ) . Similar changes of rates of migration when G. from liver was examined in cholate have been attributed to dissociation of subunits (13). When G, from brain was examined in the absence of detergent, the protein clearly aggregated (Fig. 4R). These rapid rates of sedimentation could be slowed if AMF was included (Fig. 4 0 ) , although no monomeric n subunit was observed.
This suggested that dissociation of G. by AMF was incom- plete and readily reversible. To effect more complete diasocia- tion, G. was examined by sedimentation a t 15 "C rather than 1 "C (Fig. 5). At the higher temperature, G. still aggregated in the absence of detergent, hut AMF now caused substantial amounts of activity to migrate in a nonaggregated form. The
634 Solubility of CY but Not P-y Subunits of Go and Gi
I 0 4 1
0
'41 no detergent
m v, a t- W
.-.-.-.-.-.-.-.-.-.-.~-.- "- 200 E. o41+AMF
no detergent
100
i ' \ 'ROTEINS 4NALYZED
0 a +
S ' BY
41 K 3 9 K 36K
(TOP) FRACTION NUMBER (Bottom) 'Top) FRACTION NUMBER (80ttom)
FIG. 2. Analysis of GI and a1 (M,= 41.000) by sedimentation through sucrose gradients. (1, (12 p g ) and B-y (20 p g ) were mixed in 200 pl of Solution A that contained 100 mM NaCI. Cholate (25 mM) or AMF was present as indicated. The samples were incubated on ice for 2 h prior to centrifugation through sucrose and analysis as described under "Experimental Procedures" and Fig. 1. The residual cholate from addition of the n, and f ly nuhunits did not exceed 4 mM in the prepared samples. SDS gels of the proteins utilized contain 40 ng of (I, and 60 ng of BY.
slower migration of G. activity in AMF relative to the control suggests that this activity represents monomeric a". The re- quirement for higher temperature to observe these phenomena presumably reflects a higher affinity of By for a. than for a, or ai, a t least in the absence of detergent.
The behavior of the purified subunits of G, and Gi suggested the possibility that the by subunits may be the primary means of anchoring the protein in the membrane; the a subunits would then associate with the membrane through interaction with By. Therefore, the association of a and by with phos- pholipid vesicles was tested. Vesicles of phosphatidylcholine were mixed with purified subunits of Go; the vesicles were then separated from free subunits by filtration through Ultro- gel AcA-34 (Fig. 6). The By subunits eluted in the excluded volume of the gel along with the phospholipid. In contrast, the a subunit of G,, showed very little association with the vesicles. This small amount of a, that associated with vesicles was variable with conditions and could be removed by rechro- matography of the vesicles.
While a, did not associate markedly with phospholipid vesicles, it bound quite tightly to vesicles that had been formed in the presence of By (By vesicles). Fig. 7A demonstrates that a, which was bound to by vesicles (first peak) can be clearly separated from subunit which is free (second peak). This method of analysis was used to examine the amount of CY,,
bound to By vesicles when increasing amounts of CY, were mixed with a fixed amount of vesicles (Fig. 7R). The binding of a, was saturable with low amounts of a, showing complete association with the by vesicles. The amount of a. that saturated phospholipid vesicles containing 1.4 pg of By was 24 pmol. Assuming an M, of 44,000 for By, the vesicles contained about 32 pmol of these subunits. Therefore, asso- ciation of a, with By in vesicles was approximately stoichio- metric. In fact, this experiment suggests that most of the By
was active and orientated with its binding site for a on the outside of the vesicles.
The experiment shown in Fig. 8 demonstrates that a, which is bound to By vesicles could be dissociated with GTPyS and M$'. These conditions are known to activate other G-pro- teins in solution and cause dissociation of the a from By subunits. Panel A demonstrates that a, remained associated with By vesicles (excess B y ) during gel filtration. If these vesicles were incubated with GTPyS prior to gel filtration, 60% of the a, subunits were dissociated from the vesicles (Panel B ) . It is not clear whether the 40% of binding sites for guanine nucleotide that remained with the vesicles were as- sociated with By or nonspecifically bound to the phospholipid.
This set of experiments has also been performed with vesicles made from crude preparations of brain phospholipid. Identical results are obtained a t low ionic strength and low concentrations of free divalent cations. Under the conditions described for egg phosphatidylcholine, however, a, associated significantly (up to 50%) with brain phospholipid vesicles. This association was nonspecific and readily reversible upon rechromatography. If By were incorporated into the vesicles, GTPyS was required to dissociate stoichiometric amounts of a, from the vesicles under any conditions.
DISCUSSION
Previously, only the G-protein from the visual system (G,) has been reported to behave as a soluble protein when sepa- rated from membranes. The data presented here demonstrate that the purified a subunits of G, and G, from bovine brain also remained soluble in the absence of detergent. This be- havior has been suggested previously for a,, (22). Under the same conditions, the isolated By subunits from bovine brain and the holo-G-proteins aggregated extensively. If AMF were included with Gi and G, in the absence of detergent, the migration of the a subunits could be reduced to that of the

Solubility of CY but Not P-y Subunits of Go and Gi
- -~ """"" -.
B. P y no detergent
I I I I I I I I I - "" - "- . C. P y + AMF
no detergent
I I I I I I I I I 18 16 14 12 10 8 6 4 2
(TOP) FRACTION NUMBER
-B
-Y
-B
-P
FIG. 3. Analysis of f3-y from bovine brain by uedimentation through sucrose gradients. Brain fly (30 p g ) was mixed with 200 p1 of Solution A that was supplemented with 25 mM cholate or AMF as indicated. After incubation for 2 h a t 0 "C, the samples were applied to gradients of sucrose, sedimented, and analyzed as described under "Experimental Procedures" and Fig. 1. Residual cholate in prepared samples was about 3 mM. The fly subunits were assayed hy their ahility to rapidly deactivate G. that had heen stimulated with AMF. Thus, the presence of ($7 is represented by a decrease in the reconstitution of adenylate cyclase activity by G.. The presence of (f and y in each fraction was also examined by analysis on SDS gels of 1.5% polyacrylamide. Only the region is shown for R and C, as the y suhunit was too dilute to distinguish convincingly under these conditions.
monomeric species. AMF has been shown to activate G- proteins, and this activation is thought to be accompanied by dissociation of the a from Py subunits. This dissociation can be easily observed in Figs. 1F and 2F as the P-y subunits remained aggregated and migrated a t rates faster than those of the (Y suhunits.
Studies were also carried out with G. from bovine brain. If AMF were utilized to activate G. prior to and during sedimen- tation in the absence of detergent, the aggregation of G. activity could be prevented. This suggests that a. alone may also be soluble and that this property may be common to all members of the G-protein family. The fact that higher tem- perature was required to prevent aggregation of G. and that salt was required a t low temperature to prevent aggregation of Gi predicts that the affinity of (Y suhunits for brain By in the absence of detergent follows the order a. > ai > a,.
The apparent hydrophilicity of purified (Y" was also shown by its lack of association with phospholipid vesicles. This again contrasted with the behavior of By which associated totally with these vesicles. If By were initially incorporated
I- Q 0
s
60
4 0 .
D. Gs +AMF no detergent
(Top) FRACTION NUMBER (Bottom)
FIG. 4. Analysis of G. by uedimentation through sucrose gradients at 1 O C . C . (22 pg. 37 units) was diluted into 200 pl of Solution A supplemented with 100 mM NaCl. Cholate (25 mM) and AMF were also present when indicated. After incuhation on ice for 3 h, samples were applied to sucrose gradients. subjected to centrifuga- tion, and analyzed as described under "Experimental Procedures" and Fig. 1. Residual cholate in the samples was 4 mM. The preparation of G. utilized is shown to the right (silver stnin of 70 ng). 'The gel for I ) was lost during processing. The relative migration of C. artivitv in A and C was 0.41 and 0.30, respectively. Recoveries of activity were: A , 107%; R, 12276, c. 1206; I ) , m n .
into the phospholipid vesicles, a, then associated with the vesicles in a saturable fashion that suggested a stoichiometric interaction between a, and Sy. The n,, that hound to (fy vesicles could be partially dissociated hy activation of the protein with GTPyS, a condition helieved to cause separation of the subunits. The reason for the lack of complete dissocia- tion of the a, is unclear; the suhunit remaining with the vesicles does have bound GTPyS. I t is possible that conditions of activation have partially denatured the a, and induced some hydrophobic behavior in the suhunit. Alternatively, the activated a, may interact significantly with /3y under the conditions utilized, and the partial result is an expression of the equilibrium that was achieved. So far, activated a, does not appear to interact with phospholipid vesicles or fly vesi- cles. Further experimentation will he required to determine the answer.
The data from analysis on sucrose gradients and association with phospholipid vesicles suggest that Py may be the anchor by which a, and ai associate with membranes. Activation of G, and Gi may then lead to release of the activated n suhunits
636 Solubility of a but Not ,8y Subunits of Go and Gi
B. G, no detergent I
C. Gs + AMF no detergent
(Top) FRACTION NUMBER (Bottom)
FIG. 5. Analysis of G. by sedimentation through sucrose gradients at 15 "C. G. (16 pg, 12 units) was diluted into 200 pl of Solution A supplemented with 100 mM NaCI. Cholate (25 mM) and AMF were also present when indicated. Samples were incubated at 20 "C for 30 min and applied to sucrose gradients a t room tempera- ture. The gradients were then subjected to centrifugation as described under "Experimental Procedures" and Fig. 1, except the temperature was maintained at 15 "C. The relative migration of G. activity in A and C was 0.50 and 0.43, respectively. Recoveries of activity were: A, 56%; B, 43%; C, 44%.
I FRACTION NUMBER I
as9 ADDED (pmol)
FIG. 7. Association of a. with fly vesicles. Phospholipid vesi- cles (100 p1 of 1.2 mM egg phosphatidylcholine) or vesicles (100 p1 of 1.2 mM egg phosphatidylcholine) containing By (1.4 pg) were mixed with the indicated amount of a. (900 pmol/ml in Solution A), and Solution A was added to a total volume of 0.5 ml. Samples were incubated successively for 15 min at 20 "C and 1 h on ice. Vesicles were then separated from free a. as described under "Experimental Procedures." A, profile of the elution of GTPyS-binding activity (a,,) when 100 p l of By vesicles were mixed with 175 p1 of ao. B, amounts of a,, (GTPyS-binding sites) associated with vesicles at different concentrations of a,: 0, By vesicles; 0, vesicles without incorporated protein; H, p-y vesicles minus vesicles.
c , I
24
FRACTION NUMBER
FIG. 6. Association of a,, or By with phospholipid (PL) vesi- cles. Egg phosphatidylcholine (5 mg/ml) in Solution A was sonicated to clarity. The liposomes (100 pl) were mixed on ice with 50 pmol of a, or 35 pg of brain (37, and Solution A was added to a final volume of 0.5 ml. After warming at 20 "C for 10 min and chilling, the samples were applied to columns (7 ml) of Ultrogel AcA-34 and eluted at 4 'C with Solution A. Fractions (0.5 ml) were collected and analyzed for phosphate (e), protein (01, or GTPyS-binding activity (0) as de- scribed nnder "Experimental Procedures."
from the membrane. If this occurs in uiuo, the targets of regulation by these subunits may not be limited to events associated with the plasma membrane.
This hypothesis is countered by experimental data that showed the G-proteins from brain could not be released from crude preparations of membranes except with detergent ( 6 ) . This included conditions where the Go and Gi proteins were
"
FRACTION NUMBER
FIG. 8. Dissociation of a, from fly vesicles by treatment with GTPyS. By vesicles (0.5 ml, 60 pg of B-y) were mixed with a. (0.5 ml, 30 pg) in Solution A. After incubations of 15 min at 20 "C and 30 min at 0 "C, the sample was applied to Ultrogel AcA-34 (7 ml) and eluted with Solution A. Fractions containing GTPyS-binding activity that were eluted at the excluded volume of the column were pooled as a0-y vesicles (1.5 ml, 0.7 mM phosphate, 300 nM ao). A, aBy vesicles (200 pl) were mixed with 300 pl of Solution A, incubated for 15 min at 20 "C, filtered through AcA-34 at 0 "C, and assayed for GTPyS- binding activity. B, a& vesicles (200 pl) were mixed with 200 ~1 of Solution A containing 40 mM MgC12,l.O p~ GTPyS, and [36S]GTPyS (6700 cpm/pmol). After incubation at 20 "C for 20 min, the sample was applied to the AcA-34 column and eluted at 0 "C with Solution A containing 10 mM MgSO,. Fractions were analyzed for bound GTPyS by the filter binding assay. Recoveries of binding sites were 81 and 95% for the two respective experiments.
fully associated with GTP+. One possibility for this behavior is that the a subunits are altered during purification. For example, cu, and ai may contain a fatty acid covalently at-

Solubility of a but Not P-y Subunits of Go and Gi 637
tached near their N termini as described for several proteins including p60"" and HaSV p21 (32). The attachment of fatty acid to a specific N-terminal region may direct localization of proteins to plasma membranes (33). Thus, a covalently bound fatty acid could help keep the subunits associated with the membrane; deacylation of the fatty acid during extraction and purification could then lead to the observed behavior of the purified subunits. Alternatively, it is possible that the acti- vated a: subunits dissociate from &, but bind to target pro- teins in the membrane fraction; these targets need not be limited to plasma membranes. Finally, the association of activated a: with the high concentrations of By in these membranes may be strong enough to prevent observation of large amounts of free a subunit. Thus, the dissociation of only small amounts ( 3 4 % ) of GTPyS-bindings sites from mem- branes (6) may actually reflect an equilibrium of dissociated a subunit. If this is the case, and only small amounts of activated a dissociate from the membrane and play a role in regulation, the event will be hard to observe in vivo.
Further experimentation with whole cells and their derived membranes is clearly required to verify that dissociation of a subunits from plasma membranes may be involved in regu- lation. However, the experimental usefulness of the soluble a subunits is immediately clear. The reversible binding of a to By vesicles should prove to be a very sensitive means to explore dissociation of a and j3y subunits under a variety of conditions. Transitory events involving dissociation will also be accessible to study by examining subunit exchange of radiolabeled a: subunits. The inclusion of receptors in this system will allow further exploration of the mechanism by which these proteins activate the G-proteins.
Acknowledgments-I am grateful for the superb technical assist- ance provided by Barbara Creighton and Hilda Holman. I also thank Drs. Vincent Florio, Iok-Hou Pang, and Elliott Ross for critically reading the manuscript and Nancy Bryant for skillful assistance in its preparation.
REFERENCES 1. Ross, E. M., and Gilman, A. G. (1980) Annu. Reu. Biochem. 49,
2. Stryer, L., Hurley, J. B., and Fung, B. K. K. (1981) Curr. Top.
3. Gilman, A. G. (1984) Cell 36,577-579 4. Gilman, A. G. (1984) J. Clin. Invest. 73, 1-4 5. Spiegel, A. M., Gierschik, P., Levine, M. A., and Downs, R. W.
533-564
Membr. Tramp. 15,93-108
(1985) N . Engl. J. Med. 312,26-33 6. Sternweis, P. C., and Robishaw, J. D. (1984) J. Bwl. Chem. 259,
7. Florio, V. A., and Sternweis, P. C. (1985) J. Bwl. Chem. 260,
8. Bokoch, G. M., Katada, T., Northup, J. K., Hewlett, E. L., and
9. Pfeuffer, T. (1977) J. Bioi. Chem. 252,7224-7234
13806-13813
3477-3483
Gilman, A. G. (1983) J. Bbl . Chem. 258,2072-2075
10. Fung, B. K. K., Hurley, J. B., and Stryer, L. (1981) Proc. Natl.
11. Northup, J. K., Smigel, M. D., Sternweis, P. C., and Gilman, A.
12. Katada, T., Bokoch, G. M., Smigel, M. D., Ui, M., and Gilman,
13. Sternweis, P. C., Northup, J. K., Smigel, M. D., and Gilman, A.
14. Hanski, E., Sternweis, P. C., Northup, J. K., Dromerick, A. W.,
15. Northup, J. K., Sternweis, P. C., and Gilman, A. G. (1983) J.
16. Katada, T., Bokoch, G. M., Northup, J. K., Ui, M., and Gilman,
17. Fung, B. K.-K. (1983) J. Biol. Chem. 258, 10495-10502 18. Manning, D. R., and Gilman, A. G. (1983) J. Biol. Chem. 258,
19. Gierschik, P., Godina, J., Simons, C., Birnbaumer, L., and Spie- gel, A. (1985) Proc. Natl. Acad. Sci. U. s. A. 82,727-731
20. Roof, D., Applebury, M., and Sternweis, P. C. (1985) J. Biol. Chem. 260, 16242-16249
21. Godchaux, W., 111, and Zimmerman, W. F. (1979) J. Bwl. Chem.
22. Neer. E. J.. Lok. J. M.. and Wolf. L. G. (1984) J. Bwl. Chem.
Acad. Sci. U. S. A. 78, 152-156
G. (1983) J . Bid. Chem. 258,11369-11376
A. G. (1984) J. Bwl. Chem. 259, 3586-3595
G. (1981) J. Biol. Chem. 256, 256,11517-11526
and Gilman, A. G. (1981) J. Bwl. Chem. 256, 12911-12919
Bwl. Chem. 258,11361-11368
A. G. (1984) J. B i d Chem. 259,3568-3577
7059-7063
254,7874-7884
269,14222-14229 '
. .
23. Papermaster, D. S. and Drever. W. J. (1974) Biochemistry 13. 2438-2444.
Gilman, A. G. (1977) J. Biol. Chem. 252,5761-5775
Chem. 254, 11669-11677
Chem. 267,11416-11423
- , . . - I
24. Ross, E. M., Maguire, M. E., Sturgill, J. W., Biltonen, R. L., and
25. Baehr, W., Devlin, M. J., and Applebury, M. L. (1979) J. Biol.
26. Northup, J. K., Smigel, M. D., and Gilman, A. G. (1982) J. Biol.
27. Laemmli, U. K. (1970) Nature 227,680-685 28. Wray, W., Boulikas, T., Wray, V. P., and Hancock, R. (1981)
29. Schaffner, W. and Weissman, C. (1973) Anal. Biochem. 56,502-
30. Ames, B. N. (1966) Methods Enzymol. 8, 115-118 31. Baehr, W., Morita, E. A., Swanson, R. J., and Applebury, M. L.
32. Sefton, B. M., Trowbridge, I. S., Cooper, J. A., and Skolnick, E.
33. Pellman, D., Garber, E. A., Cross, F. R., and Hanafusa, H. (1985)
Anal. Biochem. 118,197-203
514
(1982) J. Biol. Chem. 257,6452-6460
M. (1982) Cell 31,465-474
Nature 314,374-377





























