Embed Size (px)
Citation preview

E X P E R I M E N T A L C E L L R E S E A R C H 3 1 4 ( 2 0 0 8 ) 2 7 7 4 – 2 7 8 7
ava i l ab l e a t www.sc i enced i rec t . com
www.e l sev i e r. com/ loca te /yexc r
Research Article
O-GlcNAc-glycosylation of β-catenin regulates its nuclearlocalization and transcriptional activity
Ria Sayata, Brian Lebera, Vanja Grubaca, Lesley Wiltshirea, Sujata Persada,b,⁎aDepartment of Biochemistry and Biomedical Sciences, McMaster University, Hamilton, Ontario, Canada L8N 3Z5bDepartment of Pediatrics, University of Alberta, Edmonton, Alberta, Canada T6G 2J3
A R T I C L E I N F O R M A T I O N
⁎ Corresponding author. 1-130A Dentistry PhaFax: +1 780 407 3350.
E-mail address: [email protected]
0014-4827/$ – see front matter © 2008 Elsevidoi:10.1016/j.yexcr.2008.05.017
A B S T R A C T
Article Chronology:Received 6 February 2008Revised version received30 April 2008Accepted 21 May 2008Available online 6 June 2008
β-catenin plays a role in intracellular adhesion and regulating gene expression. The latter roleis associated with its oncogenic properties. Phosphorylation of β-catenin controls itsintracellular expression but mechanism/s that regulates the nuclear localization of β-cateninis unknown. We demonstrate that O-GlcNAc glycosylation (O-GlcNAcylation) of β-cateninnegatively regulates its levels in the nucleus.We show that normal prostate cells (PNT1A) havesignificantly higher amounts of O-GlcNAcylated β-catenin compared to prostate cancer (CaP)cells. The total nuclear levels of β-catenin are higher in the CaP cells than PNT1A but only aminimal fraction of the nuclear β-catenin in the CaP cells are O-GlcNAcylated. Increasing thelevels of O-GlcNAcylated β-catenin in the CaP cells with PUGNAc (O- (2-acetamido-2-deoxy-D-gluco-pyranosylidene) amino-N-phenylcarbamate) treatment is associated with a progressivedecrease in the levels of β-catenin in the nucleus. TOPFlash reporter assay and mRNAexpressions of β-catenin's target genes indicate that O-GlcNAcylation of β-catenin results in adecrease in its transcriptional activity. We define a novel modification of β-catenin thatregulates its nuclear localization and transcriptional function.
© 2008 Elsevier Inc. All rights reserved.
Keywords:β-cateninO-GlcNAcylationNuclear localizationTranscriptional activityOncogenic properties
Introduction
β-catenin is a multifunctional protein that serves a structuralrole at the adherens junctions [1] and a regulatory function asa transcriptional co-activator mediating wnt/wingless canoni-cal pathway signal transduction. Activation of this pathwayresults in stabilization and accumulation of β-catenin in thecytosol, and subsequent localization of the protein into thenucleus. In the nucleus, it interacts with the T-cell Factor(TCF)/lymphoid enhancer factor (LEF) family of transcriptionfactors and activates transcription of Wnt target genes, someof which are involved in cell proliferation, invasion andmetastasis [2–5]. Deregulated Wnt signalling results in,increased levels of β-catenin in the nucleus and constitutive
rmacy Centre, University
(S. Persad).
er Inc. All rights reserved
transcription of Wnt target genes. Increased transcriptionalactivity of β-catenin has been associated with the develop-ment and progression of many cancers.
Under normal conditions, cytosolic levels of β-catenin aretightly regulated by the ubiquitin–proteasome system, whichrequires the targeted phosphorylation of highly conservedserine and threonine residues at the NH2-terminal region ofthe protein, by glycogen synthase kinase-3 beta (GSK-3β) [6].This phosphorylation of β-catenin requires association withaxin and the product of the adenomatous polyposis coli (APC)tumor suppressor gene [5].
The phosphorylation of β-catenin is key in the regulation ofits intracellular levels and hence its transcriptional activity.However, in order for β-catenin to fulfill its transcriptional role
of Alberta, 1134-89 Avenue, Edmonton, Alberta, Canada T6G 2N8.
.

2775E X P E R I M E N T A L C E L L R E S E A R C H 3 1 4 ( 2 0 0 8 ) 2 7 7 4 – 2 7 8 7
it has to localize into the nucleus. It is still unclear as to themechanisms that regulate the nuclear localization of β-catenin, as the protein does not use the conventional importinnuclear transport system nor does it contain a nuclearlocalization sequence (NLS) [7]. Further, no mandatory carrierproteins have been identified for the transport of β-cateninfrom the cytoplasm into the nucleus. In this study, we querywhether other post-translational mechanisms may regulatethe nuclear trafficking of β-catenin.
β-catenin was found to be O-GlcNAcylated when breastcancer (MCF-7) cells were exposed to thapsigargin (TG), a drugthat induces cytoplasmic O-GlcNAcylation by activating a path-way localized at the endoplasmic reticulum [8]. While thefunctional significanceof thismodificationofβ-catenin remainsunknown, it was suggested that O-GlcNAcylation regulates itstrafficking to the cell surface of epithelial cells [8]. O-GlcNAcyla-tion is a reversible regulatory post-translational modificationfound on serine and threonine residues of nuclear andcytoplasmic proteins. It involves a single O-linked attachmentof anN-acetylglucosamine to serine/threonine residueswithoutany further modification [9,10]. The enzymes O-GlcNAc-trans-ferase (OGT) and beta-N-acetylglucosaminidase are responsiblefor adding and removing the O-GlcNAc group from proteins,respectively [11,12].O-GlcNAcmodificationsaremoreanalogousto phosphorylation than to other types of glycosylations [13,14].However, the relationship between serine/threonine O-GlcNA-cylation andphosphorylation appears to be complex and little isknown about its regulation and significance.
In this study we investigated the involvement of O-GlcNAcylation of β-catenin in regulating its subcellularlocalization and transcriptional activity. Our results indicatethat O-GlcNAcylation of β-catenin is inversely related to itsnuclear localization and transcriptional function. This novelfinding highlights O-GlcNAcylation as a new level of regula-tion of β-catenin transcriptional function.
Experimental procedures
Cell culture and PUGNAc treatment
Human CaP cells DU-145 and LNCaP (ATCC) were grown inDulbecco's Modified Eagle's medium (DMEM) supplementedwith 10% FBS. Human immortalized prostate cell line PNT1A(European Cell Culture Association) was grown in RPMI-1640media supplemented with 10% FBS. MCF-7 breast cancer cells(ATCC) were grown in DMEM supplemented with 10% FBS. Allcells were maintained in 5% CO2 at 37 °C.
PUGNAc (Toronto Research Chemicals) was dissolved insterile Milli-Q water to a final concentration of 20 mM, anddiluted to a final concentration of 100 μM in DMEM. Prior toPUGNAc treatment, DU-145 cells were incubated for 18 h withmedia containing DMEM/1% FBS. Thereafter, cells weretreated with PUGNAc (100 μM) in the absence of any serumfor different time intervals.
Whole cell lysis and nuclear extracts
Whole cell lysates from cells were obtained by incubatingwith100 μl of lysis buffer [10mMTris–HCl pH 7.5, 1mM EDTA, 1mM
EGTA, 1% Triton X-100, 2 mM phenylmethylsulfonyl fluoride(PMSF), 80 ng/ml aprotinin, 40 ng/ml chymostatin, 40 ng/mlantipain, 40 ng/ml leupeptin, 40 ng/ml pepstatin] per 106 cellson ice for 30 min. Cellular debris and nuclei were removed bycentrifugation. Nuclear and cytosolic extracts were separatedusing the NE-PER kit as described by the manufacturers(Pierce). Protein concentrations were determined using theBradford assay (Bio-Rad).
Isolation and determination of O-GlcNAcylated proteins
O-GlcNAcylated proteins were isolated by precipitation withWheat germ agglutinin (WGA)-agarose beads (Sigma) wherebyWGA-agarose beadswere incubatedwith 100 μg of cell lysate in400–500 μL of lysis buffer andmixed by rotation. Thereafter, thebeads were pelleted then washed with lysis buffer. Complexesisolated with WGA-agarose were separated on 8% SDS-PAGEgels and the gels were then used for immunoblotting withvarious antibodies. Alternatively, we also determined levels ofO-GlcNAcylated proteins by immunoprecipitation of the pro-teinswith their respective antibodies (anti-β-catenin; anti-Sp1;anti-TCF-4) and identification of the O-GlcNAc-modificationwith WGA. Briefly, 100 μg of cell lysate was immunoprecipi-tated with 4 μl of antibody (anti-β-catenin; anti-Sp1; anti-TCF-4) and the immunocomplexes separated by protein A/G plusagarose (Santa Cruz Biotech). Isolated complexes were sepa-rated on tricine polyacrylamide gels and analyzed by Westernblot for O-GlcNAc with horseradish peroxidase labeled WGA(WGA-HRP) (Vector Labs) at a dilution of 1/10,000.
Western blot
Equivalent protein amounts were resolved by SDS-PAGEelectrophoresis and cell equivalents were resolved on tricinepolyacrylamide gels. Proteins were transferred onto polyviny-lidene difluoride (PVDF) membrane, the PVDF blocked in 5%BSA and incubated with the relevant antibody. Western blotanalysis was done using the enhanced chemiluminescencedetection system (Perkin-Elmer) and developed on film. Anti-bodies to the following proteins were used for this study: β-Catenin (BD Transduction Laboratories), β-Tubulin (Santa CruzBiotechnology), Cyclin D1 (Santa Cruz Biotechnology), VEGF A(Santa Cruz), Lamin B (Calbiochem), Sp1 (Santa Cruz Biotech-nology), TCF4 (Upstate Biotechnology), Actin (ICNBiomedicals),and α,β Tubulin (Cell Signaling), APC (Cell Signaling).
Pulse-chase analysis
DU-145 cells were cultured to 70–80% confluence. Cells weretreated with or without PUGNAc (100 μM) for 14 h. Pulse-chaseanalysis of β-catenin was done as reported previously [15].Briefly, cells were pulsed for 1 h at 37°C using [35S] Promix(Amersham Pharmacia Biotech), 100 μCi/ml. Cells were thenchased with DMEM/10% FBS for the indicated time points.Immunocomplexes were isolated with anti-β-catenin anti-body complexed to protein G Sepharose (Santa Cruz Biotech)and separated on 7.5% SDS-PAGE gels, stainedwith Coomassieblue, destained, and incubated with Amplify (AmershamPharmacia Biotech) fluorographic reagent and exposed toBioflex Econofilm (Clonex).

2776 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 4 ( 2 0 0 8 ) 2 7 7 4 – 2 7 8 7
Luciferase assay for β-catenin/TCF promoter activities
β-catenin/TCF-induced transcriptional activity was deter-mined by using a β-catenin/TCF promoter-luciferase reporter
Fig. 1 – Nuclear levels and transcriptional function of β-catenin ihigh levels ofO-GlcNAcylatedβ-catenin in normal prostate cells (Pcancer (CaP) cell lines (DU-145 and LNCaP) are present in the O-Glcblot analysis (WB) of whole cell lysates with anti-β-catenin antibmethods: 1. precipitation of O-GlcNAcylated proteins with WGAantibody and 2. immunoprecipitation of β-catenin with anti-β-cspecifically withWGA-HRP. Figure is a representative experimenO-GlcNAcylated and totalβ-catenin in the nuclear and cytosolic frPNT1A. O-GlcNAcylated-β-catenin was determined by two methseparated from cell equivalents of 106 cells using the NE-PER kithigher level of β-catenin in the nucleus of the CaP cell lines comcytosolic levels of β-catenin. On the other hand the level of the O-and nuclear fractions of the PNT1A cells compared tomuch lowerDU-145 and LNCaP cells. Lamin B is used as a marker for nucleacytosolicmarker that indicates the level of cytosolic contaminatioof three separate experiments. (C) Comparison of the relativeβ-caCaP cell lines (DU-145 and LNCaP) was done by TOPFLASH-lucifeconstruct (pFOPFlash) served as a negative control for TOPFlashnormalized to pRenilla luciferase activity. β-Catenin/TCF activityPNT1A. The reporter assay shows a pattern that is parallel to thethe pattern of O-GlcNAcylated β-catenin in the cells. *p<0.05 signthe average of three separate experiments done in triplicate.
construct, pTOPFlash, by techniques reported previously [16]and according to themanufacturer's instructions (Stratagene).Briefly, a dual luciferase reporter assay was performedwhereby cells (DU-145, LNCap and PNT1A) were transfected
s inversely related to its O-GlcNAcylation. (A) While there areNT1A), very little of the total cellularβ-catenin in the prostateNAcylated form. Totalβ-cateninwas determined byWesternody. O-GlcNAcylated-β-catenin was determined by twoand detection of β-catenin specifically with anti-β-cateninatenin antibody and detection of O-GlcNAcylated proteint of three separate experiments. (B) Comparison of the level ofactions fromCaP cell lines (DU-145 and LNCaP) with that fromods as described above. Nuclear and cytosolic extracts wereas described in Experimental procedures. There is a muchpared to the PNT1A cells. A similar pattern is also seen in theGlcNAcylated form of the protein is higher in both the cytosollevels of themodified protein in the cytosol and nucleus of ther enrichment and the expression of β-tubulin is used as then of the nuclear fraction. Figure is a representative experimenttenin/TCF transcriptional activities of PNT1A to that of the tworase reporter assay. A mutated TCF-luciferase reporteractivity. TOPFlash and FOPFlash luciferase activity waswas significantly higher in the two CaP lines compared tolevels of total β-catenin in the cells and is inversely related toificantly different to normal prostate cells PNT1A. Each bar is

2777E X P E R I M E N T A L C E L L R E S E A R C H 3 1 4 ( 2 0 0 8 ) 2 7 7 4 – 2 7 8 7
with the experimental TCF promoter/luciferase reporter gene(TOPFLASH). The TOPFlash-luciferase reporter construct spe-cifically measures β-catenin/TCF regulated transcriptionalfunction and is comprised of a multimeric synthetic β-catenin/TCF-4 binding site upstream of a Thymidine Kinase(TK) minimal promoter and a luciferase open reading frame[16]. A mutated TCF-luciferase reporter construct (pFOPFlash)served as a negative control for TOPFlash activity. A controlreporter pRL-TK Renilla luciferase (Promega) was co-trans-fected in each sample to serve as an internal control fortransfection efficiency. The β-catenin-independent MCPT1-luciferase reporter construct was used as a negative control tomonitor any non-specific effects of the treatments. Transfec-tions of cells were done with the use of ExGen500 reagent(Fermentas Life Sciences) according to the manufacturer'sguidelines. Reporter activity was measured using a lumin-ometer (Tropix TR717).
QRT-PCR analysis of Cyclin D1 and VEGFA mRNA expression
DU-145 cells were allowed to reach 70% confluency in DMEM/10% FBS media. Cells were treated with PUGNAc (100 μM) asdescribed above for the various time intervals. Total RNA wasisolated from DU-145 cells using the Trizol reagent (Invitro-gen Life Technologies) according to the manufacturer'sinstructions and quantified spectrophotometrically. 1 μg oftotal RNA was reverse-transcribed using iSCRIPT cDNAsynthesis kit (Bio-Rad) according to the manufacturer'sprotocol. Real time quantification of Cyclin D1 and VEGFAgene expressions were assessed using the iQ SYBR GreenSupermix Kit (Bio-Rad). Samples were amplified with aprecycling hold at 94 °C for 5 min, followed by 35 cycles ofdenaturation at 94 °C for 1 min, annealing at 54 °C for 2 min,and extension at 72 °C for 3 min. Primers used for Cyclin D1were 5′ – CTGGCCATGAACTACCTGGA – 3′ (sense) and 5′ –GTCACACTTGATCACTCTGG – 3′ (anti-sense) and for VEGFAwere 5′ – GCAGAATCATCACGAAGTGG – 3′ (sense) and 5′ –GCATGGTGATGTTGGACTCC – 3′ (anti-sense). For externalcontrol, the glucose-6-phosphate dehydrogenase (G6PDH)gene was amplified using oligonucleotides 5′ – ACCTGG-TGCTCAGTGTAGCC – 3′ (sense) and 5′ – CAATGACCCCTT-CATTGACC – 3′ (anti-sense). Quantitative PCR (QPCR) wasperformed on the cDNA in triplicates using the MiniOpticonReal-Time PCR Detection System (Bio-Rad). The amount ofmRNA was calculated using the comparative Ct method andis expressed as 2−ΔΔCt.
Treatment of cells with Wnt conditioned media
Mouse L cells that were stably transfected withWnt3a plasmid(LWnt3a), Wnt5a plasmid (LWnt5a) and the parental L cellswere purchased from the American Type Culture Collection(ATCC). The cell lines were cultured in DMEM containing 10%FBS. Control conditioned media (CM) and Wnt3a or Wnt5a CMwere prepared from confluent cultures according to manufac-turer's guidelines (ATCC). Briefly, CM was collected, centri-fuged and sterilized by filtration through 0.2-μm filters.Conditioned media were mixed with an equal volume ofDMEM (1:2 dilution) and used for treatment of DU145 cells forthe indicated time periods. Treatment of DU145 cells with CM
from the L cells served as control for the treatments withLWnt3a and LWnt5a CM.
Results
Nuclear levels and transcriptional function of β-catenin isinversely related to its O-GlcNAcylation
Constitutive transcription of target genes mediated byincreased cellular, and more importantly, nuclear levels ofβ-catenin is associated with the genesis and progression ofmany cancers [2–4,17,18]. Intracellular levels of β-catenin areregulated by cycles of phosphorylation and dephosphoryla-tion. However, β-catenin is required to localize to thenucleus to fulfill its transcriptional role and it is still unclearas to how this process is mediated. Recently, β-catenin wasreported to be modified by O-GlcNAcylation [8]. While thisobservation is novel in itself the greater question of thefunctional significance of this modification remainsunknown. In this study we asked whether O-GlcNAcylationof β-catenin plays a role in regulating the protein's nuclearlocalization and hence transcriptional activity. To addressthis question, we used prostate cells as models and firstcompared the O-GlcNAcylation of β-catenin in a normalimmortalized prostate cell line, PNT1A, to that in two CaPcell lines (DU-145, LNCaP) (Fig. 1A). Our results indicate thatwhile there are high levels of O-GlcNAcylated β-catenin inthe PNT1A cell line, the CaP cell lines exhibit low levels of O-GlcNAcylated-β-catenin (Fig. 1A). In contrast, the totalcellular level of β-catenin is higher in the CaP cells comparedto the PNT1A cells (Fig. 1A). More importantly, we notedgreater total nuclear levels of β-catenin in the CaP cell linescompared to the PNT1A cells (Fig. 1B). All the cell linesexhibited lower amounts of the O-GlcNAcylated form of β-catenin in the nucleus than in the cytoplasm (Fig. 1B). Itshould be pointed out that although the relative quantity ofO-GlcNAcylated β-catenin in the PNT1A cells appear greaterthan the total cellular β-catenin (Fig. 1A), this is likely due tothe concentrating effect of WGA precipitation of cell lysatesrequired for the isolation of the O-GlcNAcylated proteinsprior to the immunoblot for β-catenin. The alternate methodof immunoprecipitation with anti-β-catenin antibody priorto immunoblotting with WGA-HRP will result in a similarconcentrating effect. In comparison, determination of totalβ-catenin involves direct immunoblotting of the dilute celllysates without any prior precipitation.
We then measured TOPFlash-luciferase reporter activity inthe cell lines to determine their transcriptional status. Incorrelation with the observed total and nuclear levels of β-catenin (Figs. 1A and B), TOPFLASH activity was significantlyhigher in the two CaP cell lines compared to the PNT1A cells(Fig. 1C). TOPFLASH activity was inversely correlated to the O-GlcNAcylated β-catenin levels in all the cell lines (Figs. 1A–C).These results indicate a reciprocal relationship between thenuclear levels/transcriptional activity of β-catenin and thelevels of the O-GlcNAc modified protein. This is suggestive,that O-GlcNAcylation of β-catenin is a mechanism thatnegatively regulates the localization and accumulation of theprotein in the nucleus.

2778 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 4 ( 2 0 0 8 ) 2 7 7 4 – 2 7 8 7
Nuclear localization of β-catenin is regulated by itsO-GlcNAcylation
In order to further explore this paradigm, we next treated DU-145 cells with the drug PUGNAc. PUGNAc is a GlcNAc analoguethat potently inhibits O-GlcNAcase in cells [19], thus prevent-ing the cycling of O-GlcNAc on proteins, leading to globallyelevated levels of this modification. PUGNAc has been shownto induceO-GlcNAcylation of a variety of proteins [19]. Optimaltreatment conditions required that PUGNAc treatment of DU-145 cells be done in the absence of serum in order to prevent
serum factors from interfering with the activity of the drug.Thuswe carried out experimentswhere cellswere treatedwithPUGNAc in the presence of different concentrations of serum(FBS) for various time periods in order to determine theconditions for optimal efficiencyof PUGNAc treatmentwithoutcompromising the viability of the cells (data not shown).Further, a dose response of PUGNAC treatment of DU-145 cellsunder these treatment conditions was done and a workingconcentration of 100 μM was determined as optimal (data notshown). As a result of our findings from these experiments,DU-145 cells grown in the presence of full serum (10%FBS)were

Table 1 – Levels of β-catenin from nuclear and cytosolicfractions of DU-145 cells treated with PUGNAc
Hours ofPUGNAcTreatment
O.D. Nuclearβ-cateninng/mL
O.D. Cytosolicβ-cateninng/mL
0 8689.02 11.94+/−0.77 8755.74 12.07+/−0.468 7443.14 9.56+/−0.95 10,778.12 15.94+/−2.58 ⁎14 7081.55 8.87+/−1.95 ⁎ 11,705.63 17.72+/−1.13 ⁎18 6245.35 7.27+/−1.85 ⁎ 12,603.78 19.44+/−2.15 ⁎22 5402.61 5.66+/−7.71 14,086.62 22.27+/−3.69
Densitometry from Western blot analysis of a diluted β-cateninstandard protein produced a standard curve from which amountsof nuclear and cytosolic β-catenin could be interpolated. Theamounts of nuclear β-catenin decreased approximately 50% from0 h to 22 h of PUGNAc treatment. While the amount of β-catenin inthe cytosol increased to slightly less than two-fold from 0 to 22 h ofPUGNAc treatment. Values are means of three independentexperiments and error bars are the standard error of the mean.⁎ pb0.05 compared to 0 h control (Two-tailed T-test).
2779E X P E R I M E N T A L C E L L R E S E A R C H 3 1 4 ( 2 0 0 8 ) 2 7 7 4 – 2 7 8 7
incubated in media containing 1% serum for 18 h prior totreatment with PUGNAc (100 μM) for various times points (asshown in figures) in the absence of serum.
Treatment of DU-145 cells with PUGNAc resulted in a time-dependent increase in the cellular levels of O-GlcNAcylated β-catenin (Fig. 2A). Subcellular increase in levelsofO-GlcNAcylatedβ-catenin was evident only in the cytosolic pool of β-cateninwithout any change to the level of O-GlcNAc β-catenin in thenucleus (Fig. 2B). Importantly, as the cytosolic levels of O-GlcNAcylated β-catenin increased with PUGNAc treatment weobserved a time-dependent decrease in nuclear levels of β-catenin and an equal associated increase in the cytosolic level ofthe protein (Fig. 2B). Interestingly, the total cellular levels of β-catenin remained constant throughout the treatment. This isfurther clarified in Table 1, which shows themean values of thedensitometric analysis of β-catenin from nuclear and cytosolicfractions of DU-145 cells treated with PUGNAc. The resultsclearly indicate that the decrease in nuclear β-catenin levels(50% from 0 h to 22 h of PUGNAc treatment) is almost equivalentto the increase in cytosolic β-catenin levels (to slightly less than
Fig. 2 – Nuclear localization of β-catenin is regulated by its O-Gl(100 μM) for various time points results in a progressive increaseO-GlcNAcylated-β-catenin was determined by two methods: 1. pdetection of β-catenin specifically with anti-β-catenin antibody aantibody and detection of O-GlcNAcylated protein specifically wiunchanged by PUGNAc treatment. Figure is a representative expDU-145 cells for various time points with PUGNAc (100 μM) resuβ-catenin, which is accompanied by progressive increase in cytocauses a time-dependent increase in the level of O-GlcNAcylatedO-GlcNAcylated-β-catenin was determined by two methods as dfraction is unaffected by PUGNAc treatment. Lamin B is used asβ-tubulin is used as the cytosolic marker that indicates the levelrepresentative experiment of three separate experiments. (C) Bre12 and 18 h. Results show a decrease in the nuclear levels ofβ-catcytosol. O-GlcNAcylated β-catenin levels increased in the cytosotreatment. These results are similar to those seen upon treatmenThis indicates that the effect of O-GlcNAcylation on nuclear localFigure is a representative experiment of three separate experim
two-fold from0to22hof PUGNAc treatment). Similar alterationsin the nuclear and cytosolic levels of β-cateninwere observed inLNCaP cells (data not shown). Further, the breast cancer cell lineMCF-7 exhibited a similar pattern of changes in nuclear andcytosolic levels of β-catenin upon PUGNAc treatment, albeit atdifferent time points (Fig. 2C). These results further underscorethe negative regulatory effect of O-GlcNAcylation on nuclearaccumulationofβ-catenin. Further, the results also indicate thatthe effect of O-GlcNAcylation upon nuclear localization of β-catenin is not cell specific or cancer specific.
O-GlcNAcylation of β-catenin does not alter its stability in cells
Our results in Figs. 2B, C indicate that the attenuation ofnuclear levels of β-catenin with increasingO-GlcNAcylation ofβ-catenin was associated with an increase in the cytosoliclevels of the protein. This can conceivably be caused by either aredistribution of the protein from the nuclear to the cytosoliccompartments or due to O-GlcNAcylation-associated stabili-zation of the protein. Indeed, studies have shown that O-GlcNAcylation of proteins slow down their proteolytic degra-dation [20,21]. Thusweexamined the effect ofO-GlcNAcylationon the stability of β-catenin by pulse-chase analysis of DU-145cells treated with PUGNAc (Fig. 3). We observed that the rate ofdegradation of β-catenin was unchanged in cells treated withPUGNAc compared to cells untreated with PUGNAc indicatingthat the rate of degradation of β-catenin is not affected by O-GlcNAcylation. Together the results in Figs. 2 and 3 and Table 1suggest that the observed increase in cytosolicβ-catenin levelsthat occurs in parallel with the decrease in nuclear β-cateninupon PUGNAc treatment is likely the result of a redistributionof the protein from the nuclear to the cytosolic compartmentsand not due to alterations in the stability of the protein.
Nuclear localization of Sp1 is directly related to itsO-GlcNAcylation
It is arguable that the negative effect of increasing cellular O-GlcNAcylation upon nuclear β-catenin levels is the result ofglobalalterations innormal import via thenuclearporeproteins,
cNAcylation. (A) Treatment of DU-145 cells with PUGNAcin the cellular levels of O-GlcNAcylated β-catenin.recipitation of O-GlcNAcylated proteins with WGA andnd 2. immunoprecipitation of β-catenin with anti-β-cateninth WGA-HRP. The level of total cellular β-catenin remainseriment of three separate experiments. (B) Treatment oflts in a time-dependent decrease in the nuclear levels ofsolic level of β-catenin. In parallel, treatment with PUGNAcβ-catenin in the cytosolic compartment of the cells.escribed above. O-GlcNAcylation of β-catenin in the nucleara marker for nuclear enrichment and the expression ofof cytosolic contamination of the nuclear fraction. Figure is aast cancer MCF-7 cells were treated with PUGNAc (100 μM) foreninwith a corresponding increase in level ofβ-catenin in thel, whereas the nuclear fraction was unaffected by PUGNAct of DU-145 cells with PUGNAc, albeit at different time points.ization of β-catenin is not cell line or cancer specific.ents.
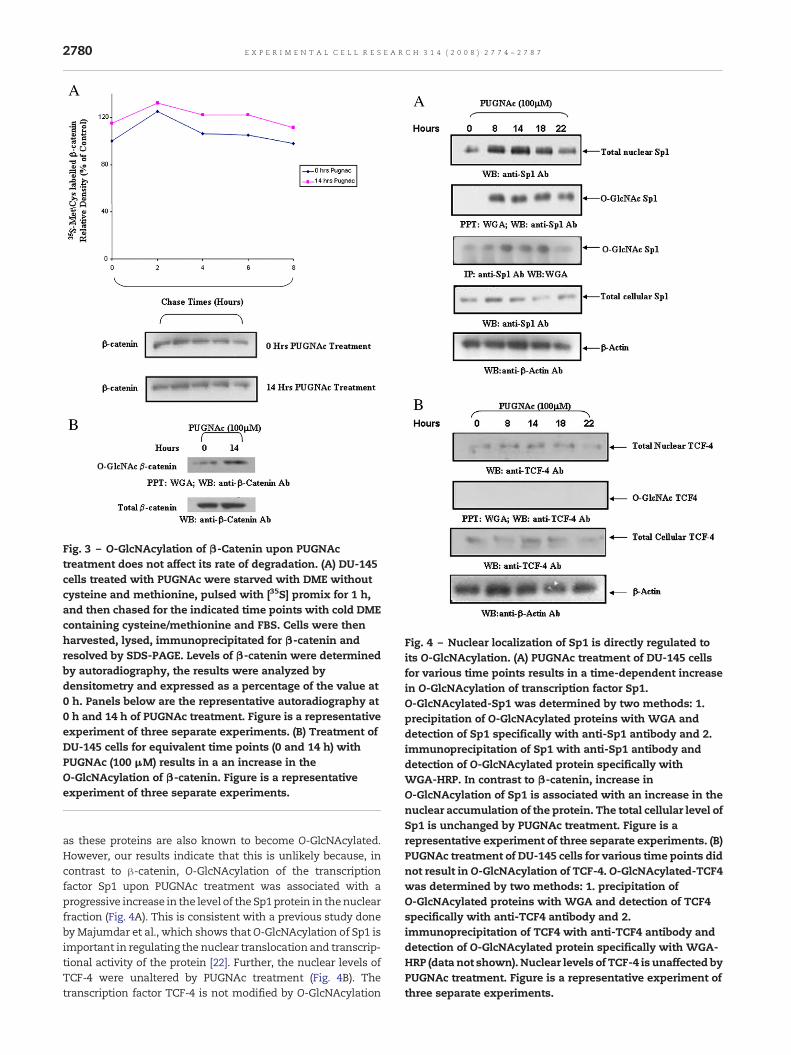
Fig. 3 – O-GlcNAcylation of β-Catenin upon PUGNActreatment does not affect its rate of degradation. (A) DU-145cells treated with PUGNAc were starved with DME withoutcysteine and methionine, pulsed with [35S] promix for 1 h,and then chased for the indicated time points with cold DMEcontaining cysteine/methionine and FBS. Cells were thenharvested, lysed, immunoprecipitated for β-catenin andresolved by SDS-PAGE. Levels of β-catenin were determinedby autoradiography, the results were analyzed bydensitometry and expressed as a percentage of the value at0 h. Panels below are the representative autoradiography at0 h and 14 h of PUGNAc treatment. Figure is a representativeexperiment of three separate experiments. (B) Treatment ofDU-145 cells for equivalent time points (0 and 14 h) withPUGNAc (100 μM) results in a an increase in theO-GlcNAcylation of β-catenin. Figure is a representativeexperiment of three separate experiments.
Fig. 4 – Nuclear localization of Sp1 is directly regulated toits O-GlcNAcylation. (A) PUGNAc treatment of DU-145 cellsfor various time points results in a time-dependent increasein O-GlcNAcylation of transcription factor Sp1.O-GlcNAcylated-Sp1 was determined by two methods: 1.precipitation of O-GlcNAcylated proteins with WGA anddetection of Sp1 specifically with anti-Sp1 antibody and 2.immunoprecipitation of Sp1 with anti-Sp1 antibody anddetection of O-GlcNAcylated protein specifically withWGA-HRP. In contrast to β-catenin, increase inO-GlcNAcylation of Sp1 is associated with an increase in thenuclear accumulation of the protein. The total cellular level ofSp1 is unchanged by PUGNAc treatment. Figure is arepresentative experiment of three separate experiments. (B)PUGNAc treatment of DU-145 cells for various time points didnot result in O-GlcNAcylation of TCF-4. O-GlcNAcylated-TCF4was determined by two methods: 1. precipitation ofO-GlcNAcylated proteins with WGA and detection of TCF4specifically with anti-TCF4 antibody and 2.immunoprecipitation of TCF4 with anti-TCF4 antibody anddetection of O-GlcNAcylated protein specifically with WGA-HRP (data not shown). Nuclear levels of TCF-4 is unaffected byPUGNAc treatment. Figure is a representative experiment ofthree separate experiments.
2780 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 4 ( 2 0 0 8 ) 2 7 7 4 – 2 7 8 7
as these proteins are also known to become O-GlcNAcylated.However, our results indicate that this is unlikely because, incontrast to β-catenin, O-GlcNAcylation of the transcriptionfactor Sp1 upon PUGNAc treatment was associated with aprogressive increase in the level of the Sp1protein in thenuclearfraction (Fig. 4A). This is consistent with a previous study doneby Majumdar et al., which shows thatO-GlcNAcylation of Sp1 isimportant in regulating the nuclear translocation and transcrip-tional activity of the protein [22]. Further, the nuclear levels ofTCF-4 were unaltered by PUGNAc treatment (Fig. 4B). Thetranscription factor TCF-4 is not modified by O-GlcNAcylation

2781E X P E R I M E N T A L C E L L R E S E A R C H 3 1 4 ( 2 0 0 8 ) 2 7 7 4 – 2 7 8 7
as demonstrated by the absence of any bands when either O-GlcNAcylated proteins were precipitated with WGA and Wes-tern blotted with TCF-4 (Fig. 4B) or TCF4 was immunoprecipi-tated with anti-TCF4 antibody and Western blotted with WGA-HRP for the detection ofO-GlcNAc (data not shown). These datasuggest that the effect of PUGNAc treatment, and hence O-GlcNAcylation, on β-catenin is quite specific.
O-GlcNAcylation of β-catenin results in decreasedβ-catenin/TCF transcriptional activity
Given that nuclear β-catenin complexed to TCF induces thetranscription of genes containing a TCF/DNA binding site in
Fig. 5 – O-GlcNAcylation ofβ-catenin results in decreasedβ-catenactivities (TOPFlash-luciferase reporter) in response to the varioutransfected with 0.5μg of TOPFlash reporter using EXGEN-500 tradramatically and significantly reduced in a time-dependent mannegative control for TOPFlash activity. There is a parallel increasthat was used for the TOPFlash/FOPFlash reporter assays. (B) ThTOPFlash-luciferase activity is quite specific as PUGNAc treatmeβ-catenin-independentMCPT-1-luciferase reporter construct. *p<is the average of three separate experiments done in triplicate. (nuclear lysates from DU-145 cells treated without or with PUGNAassociation (complex formation) between TCF-4 and β-catenin wCo-immunoprecipitation of APC with β-catenin using nuclear lysand 14 h. The results indicate that the nuclear association (compincreased cellular O-GlcNAcylation of β-catenin. C and D are eac
their promoter region, we next focused on investigatingwhether the decrease in nuclear β-catenin observed uponPUGNAc treatment is reflected in β-catenin-mediated transac-tivation in the cell. We examined this by transfecting theTOPFlash-luciferase reporter construct into DU-145 cells andsubsequently treating the transfected cells with PUGNAc forvarious time intervals. The results presented in Fig. 5A showthat as the level of O-GlcNAcylated β-catenin increases uponPUGNAc treatment it is accompanied by a correspondingdramatic temporal decrease in theTOPFlash-luciferaseactivity.
In order to rule out the possibility that alterations inTOPFlash-luciferase activity upon PUGNAc treatment is theresult of non-specific global cellular alterations, we carried out
in/TCF transcriptional activity. (A) The relativeβ-catenin/TCFs times of PUGNAc treatment. DU-145 cells were transientlynsfection reagent. Figure shows thatβ-catenin/TCF activity isner with PUGNAc treatment. FOPFlash activity served as ae in O-GlcNAcylation of β-catenin at the same time pointse effect of PUGNAc upon the β-catenin/TCF-dependentnt had no effect upon the luciferase activity of the0.05 significantly different to untreated DU-145 cells. Each barC) Co-immunoprecipitation of TCF-4 with β-catenin usingc for 8 and 14 h. The results indicate a decrease in theith increased cellular O-GlcNAcylation of β-catenin. (D)ates from DU-145 cells treated without or with PUGNAc for 8lex formation) between APC and β-catenin is not altered withh representative experiments of three separate experiments.

2782 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 4 ( 2 0 0 8 ) 2 7 7 4 – 2 7 8 7
parallel studies with the MCPT-1-luciferase reporter constructwhich is a β-catenin-independent luciferase reporter con-struct. The results in Fig. 5B indicate that the activity of theMCPT-1-luciferase reporter construct was unaffected byPUGNAc treatment underscoring the specificity of the rela-tionship between PUGNAc treatment, O-GlcNAcylation of β-catenin and changes in TOPFlash-luciferase activity.
The data thus far supports a model where O-GlcNAcylationof β-catenin plays a role in limiting its nuclear accumulation,which subsequently results in a functional response of adecrease in its transcriptional/transactivation activity. How-ever, increased or decreased levels of β-catenin in the nucleus,while crucial in determining its nuclear availability fortransactivation is not the only factor involved. It is mandatoryfor β-catenin to interact and form a complex with members oftheTCF family of proteins in order to initiate transcriptionasβ-catenin itself is unable to bindDNAandpromotes transcriptionof target genes via its binding to TCFproteins. TCF-4 is the formof TCFmost abundantly found inmammalian cells. Therefore,we next tested the effect of PUGNAc treatment of DU-145 cellsupon the interaction of β-catenin with TCF-4. The results inFig. 5C shows decreased coimmunoprecipitation of TCF-4withβ-catenin upon PUGNAc treatment indicating an attenuationin the formation of the functional transcription complexbetween the two proteins. This is likely a reflection of thelower levels ofβ-catenin available in thenucleusuponPUGNAc
Fig. 6 – Alterations in the mRNA expression of Cyclin D1 and VEexpressions of (A) Cyclin D1 (upper panel) and (B) VEGFA (upper pinhibited in a time-dependent manner when DU-145 cells are tre(lower panel) and (B) VEGFA (lower panel) are observed when ceequivalent time periods. *p<0.05 significantly different to untreaexperiments done in triplicate.
treatment. Since the nuclear levels of O-GlcNAcylated β-catenin does not change with PUGNAc treatment the interac-tion between β-catenin and TCF-4 in the formation of the β-catenin/TCF-4 complexmay ormay not be associated with theO-GlcNAcylation of the latter. However, since decreases in β-catenin/TCF-4 complex is observed in the absence of anyalteration in O-GlcNAcylation of nuclear β-catenin, one mightspeculate that β-catenin in it O-GlcNAcylated form does notbind TCF and formation of the β-catenin/TCF-4 transcriptioncomplex is likely dependent upon the availability of the non-modified form of the protein. Further studies are required tounderstand the full implications of O-GlcNAcylation upon theinteraction of β-cateninwith TCF-4 and the other co-activatorsof the transcription complex such as Bcl-9.
Interestingly, in contrast to its interaction with TCF-4, theinteraction of β-catenin with the protein AdenomatousPolyposis Coli (APC) was unaltered in DU145 cells treatedwith and without PUGNAc (Fig. 5D). APC is known to interactwith β-catenin and assist the latter's export out of the nucleus.
O-GlcNAcylation of β-catenin results in attenuation of themRNA expressions of Cyclin D1 and VEGFA
β-catenin regulates Cyclin D1 [2] and VEGFA [23,24] expres-sions at the transcriptional level through its interaction withTCF-4. Thus, we examined whether the mRNA expression of
GFA in DU-145 cells treated with PUGNAc. The mRNAanel), determined by QRT-PCR analysis, are both profoundlyated with PUGNAc. No change in mRNA levels of (A) cyclin D1lls are maintained in serum free media without PUGNAc forted DU-145 cells. Each bar is the average of three separate

Fig. 7 – PUGNAc treatment results in a dramatic decrease in the protein expression of (A) Cyclin D1 (upper panel) and (B) VEGFA(upper panel). Treatment of cells for equivalent time periods in serum free media in the absence of PUGNAc had no effect upon(A) Cyclin D1 (lower panel) or (B) VEGFA (lower panel) protein expression. This indicates that the alterations observed areassociatedwith theO-GlcNAcylation caused by PUGNAc treatment and not due to the conditions of serumdeprivation. Figure isa representative experiment of three separate experiments.
2783E X P E R I M E N T A L C E L L R E S E A R C H 3 1 4 ( 2 0 0 8 ) 2 7 7 4 – 2 7 8 7
Cyclin D1 and VEGFA are altered by the treatment of DU145cells with or without PUGNAc. Quantitative RT-PCR analysis oftotal RNA from PUGNAc-treated and untreated DU-145 cellswere used to determine the effect of O-GlcNAcylation uponCyclin D1 (Fig. 6A-upper panel) and VEGFA (Fig. 6B-upperpanel) mRNA levels. Our data indicate that treatment withPUGNAc results in a decrease in themRNA expressions of bothCyclin D1 (5-fold) and VEGFA (2-fold). These results are in
Fig. 8 – O-GlcNAcylation of β-cateninis regulated by Wnt3a. DU1expressing L cells (LWnt3a CM), Wnt5a expressing L cells (LWnt5Treatment of DU-145 cells with LWnt3a CM results in a progressLWnt3a CM treatment of DU145 cells for the same time periods cO-GlcNAcylated β-catenin. Treatment of DU145 cells with LWnt5aor O-GlcNAcylated β-catenin levels. Figure is a representative ex
agreement with our results in Fig. 5. Indeed, the attenuation inmRNA expressions of the target genes is likely the directconsequence of the decreased formation of the β-catenin/TCF-4 complexes resulting in a decreased β-catenin/TCF-4transcriptional activity upon PUGNAc treatment. However,since PUGNAc treatment is carried out in the absence of serum(see Experimental procedures) we further determinedwhetherthe decrease in Cyclin D1 mRNA (Fig. 6A-lower panel) and
45 cells were incubated with conditioned media from Wnt3aa CM) or parental L cells (L CM) for the indicated time periods.ive increase in the cellular levels of β-catenin. In contrast,auses a time-dependent decrease in the cellular levels ofCM or the control L CM has no effect upon cellular β-cateninperiment of three separate experiments.

Fig. 9 – Schematic representation of mechanismsregulating β-catenin. The cellular level of β-catenin is verytightly regulated by phosphorylation. Phosphorylation ofβ-catenin by glycogen synthase kinase-3beta (GSK-3β)results in its degradation. Prevention of phosphorylationstabilizes the protein and results in an increase of itslevels inside cells. In this study we show that β-catenin
2784 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 4 ( 2 0 0 8 ) 2 7 7 4 – 2 7 8 7
VEGFA mRNA (Fig. 6B-lower panel) was a result of serumdeprivation. However, treatment of DU-145 cells with medialacking serum for equivalent time periods in the absence ofPUGNAc did not cause any reduction in themRNA expressionsof Cyclin D1 or VEGFA. Interestingly, in parallel with themRNAexpressions, PUGNAc treatment also resulted in a time-dependent decrease of both factors at the protein level (Figs.7A and B-upper panels) underscoring a functionally relevantend point to the observed alterations. Treatment of DU-145cells with media lacking serum for equivalent time periods inthe absence of PUGNAc did not cause any reduction in theprotein expressions of Cyclin D1 (Fig. 7A-lower panel) orVEGFA (Fig. 7B-lower panel).
O-GlcNAcylation of β-catenin is regulated by Wnt3A but notWnt5A
Wnt proteins comprise a large family of glycoproteins that areinvolved in the regulation of gene transcription, cell adhesionand cell polarity [25,26]. Many of the effects ofWnt proteins areexerted by regulating the expression of target genes throughthe stabilization and nuclear localization of β-catenin (Cano-nicalWnt signaling pathway) [27,28]. Since our results indicatethat nuclear localization of β-catenin is altered by its O-GlcNAcylation, we examined whether the O-GlcNAcylation ofβ-catenin is affected by Wnt stimulation. Since specific Wntproteins signal via either the Wnt/β-catenin (canonical) or theWnt/Ca+ (non-canonical) signaling pathways [25,26], wedetermined the effect of both pathways uponO-GlcNAcylationof β-catenin.Wnt3a is a prototypical canonical Wnt andWnt5ais a prototypical non-canonical Wnt. Thus stimulation ofDU145 cells with CM from Wnt3a expressing L cells (LWnt3aCM) was used to study the regulation by the canonical Wntsignaling pathway and stimulation with CM from Wnt5aexpressing L cells (LWnt5a CM) was used to determineregulation by the canonical Wnt signaling pathway. CM fromL cells served as the overall control as these cells do notsecrete anyWnt factors. Incubation of DU145 cells with LWnt3aCM resulted in the stabilization and increase in the cellularlevels of β-catenin (Fig. 8). In contrast, cellular levels of O-GlcNAcmodified β-catenin decreased dramatically by incuba-tion of DU145 cells with LWnt3a CM in a time-dependentmanner (Fig. 8). No alterations in cellular total β-catenin levelsor O-GlcNAcylated β-catenin levels were observed whenDU145 cells were incubated with L Wnt5a CM or the control LCM (Fig. 8). The fact that O-GlcNAcylation of β-catenin isaltered only by theWnt3a containing CM and not by theWnt5acontaining CM suggests that this modification of β-catenin isspecifically regulated by the canonicalWnt signaling pathway.Although these results support a canonical Wnt regulatorycomponent upon the O-GlcNAcylation of β-catenin, furtherinvestigations are required to fully appreciate themechanisticdetails of this regulation.
is also modified by O-linked beta-N-acetylglucosamine(O-GlcNAcylation). O-GlcNAc modification does not preventβ-catenin from interacting with E-cadherin (8). Our dataindicate that O-GlcNAcylation negatively regulates thenuclear localization ofβ-catenin resulting in a decrease in thenuclear levels of the protein and thereby its transcriptionalactivity. OGT = O-GlcNAc Transferase.
Discussion
Many molecules rely on specific peptide sequences to localizeto given membrane-bound compartments such as nuclearlocalization/export sequences, NLS or NES respectively, [29–
31] and cell membrane localization signals [32]. β-catenin doesnot contain a nuclear localization sequence [7] and little isknown about how nuclear levels of the protein is regulated.
In this study we demonstrate a novel mechanism involvingO-GlcNAcylation that tightly and inversely regulates thenuclearlocalization of β-catenin andhence its activity as a transcriptionfactor (Fig. 9). Data presented here demonstrate that O-GlcNAcylated-β-catenin is inversely related to the transcrip-tional status of the cell such that there is minimal O-GlcNAcylation of β-catenin in cancer cellswhere transcriptionalactivity is high and a significant level of O-GlcNAcylated β-catenin in normal cells where transcriptional activity is low.Further, increasing the levels of O-GlcNAcylated β-catenin incancer cells results in a loss of nuclear β-catenin and aconcomitant increase in its cytosolic levels. We propose thatO-GlcNAcylation limits β-catenin within the compartment(nuclear or cytosolic) in which it is O-GlcNAcylated by yetundefined mechanisms. To this end, O-GlcNAcylation ofproteins is known to occur independently in the cytosol andnucleus by separate cytosolic and nuclear pools of O-GlcNActransferase [33]. Given that in normal cells the bulk of β-cateninis in the cytosol, and we report that most of it is in an O-

2785E X P E R I M E N T A L C E L L R E S E A R C H 3 1 4 ( 2 0 0 8 ) 2 7 7 4 – 2 7 8 7
GlcNAcylated form, then O-GlcNAcylation may be a regulatorymechanismemployedby cells to restrictβ-catenin to the cytosoland compromise its availability in the nucleus for transcrip-tional function. In cancer cells, as well as in other malignancieswhere this modification is suppressed, this control is lostresulting in increased shuttling of β-catenin, increased nuclearaccumulation and increased transcription of its targets. Inter-estingly, previous studies have shown that O-GlcNAcylationplays a role in regulating nuclear transport of proteins [34] andthere is evidence that suggest that O-GlcNAcylation mayrepresent an alternative nuclear transport signal.
Although the precise mechanism/s by which O-GlcNAcyla-tion regulates nuclear trafficking is not clear at present, ourresults indicate that the nuclear localization of three differenttranscription factors (β-catenin, Sp1 and TCf-4) are affectedspecifically and differentially by PUGNAc treatment. Therelative specificity of the changes in the patterns of nuclearlocalization of the three transcription factors negates thepossibility that the alterations observed are the result of aglobal change in nuclear trafficking induced by O-GlcNAcyla-tion of nuclear pore proteins. Rather, the observed alterationsin the subcellular localization of β-catenin upon PUGNActreatment are quite specific in nature and are associated withits modification by O-GlcNAcylation.
β-catenin is known to continuously shuttle between thecytosolic and nuclear compartments [7]. It is possible that O-GlcNAcylation may alter the tertiary conformation of β-catenin such that the protein's shuttling between the cytosolicand nuclear compartments becomes compromised thuseffectively restricting the protein to the compartment inwhich it was modified. Increased level of O-GlcNAcylated β-catenin in the cytosolwould therefore restrict the protein fromleaving the cytosol and enter the nucleus. However, as there isno change in the level of O-GlcNAcylated β-catenin in thenucleus with PUGNAc treatment the export of β-catenin out ofthe nucleus continues. The result is an emptying of β-cateninfrom the nucleus and an increase of its levels in the cytosol.
Putative O-GlcNAcylation-related changes in β-cateninwhich may be responsible for the observed alterations are asfollows: O-GlcNAcylation may negatively modulate β-cate-nin's interactions with nuclear pore proteins involved in itsimport to the nucleus. β-catenin is known to enter the nucleusindependent of any known nucleo-cytoplasmic binding part-ners [35] by interacting directly with components of thenuclear pore complex [36–38]. This possibility is beinginvestigated at present. Alternatively, O-GlcNAcylation mayselectively enhance β-catenin's interaction with cytoplasmic[39,40] or nuclear [41–44] binding partners resulting in itssequestration in the respective compartment. In this regard itis interesting to note that the interaction of β-catenin withTCF-4 (the major molecular interaction that effectivelysequesters β-catenin in the nucleus) is decreased withPUGNAc treatment (Fig. 5C). On the other hand, the interactionof β-catenin with the APC (an interaction that will effectivelyremove β-catenin from the nucleus) is unchanged by PUGNActreatment (Fig. 5D). The net effect of O-GlcNAcylation on thesetwo interactions would favor a decrease of β-catenin from thenucleus and an increase in its levels in the cytoplasm. This issupported by our observations in Fig. 2B. However, theseinteractions have to be studied in greater detail and the
interaction of β-catenin with other binding partners needs tobe evaluated. These objectives are being investigated in ourongoing studies at present.
The cellular biological consequence of increased levels ofβ-catenin in the nucleus that is relevant to disease progres-sion is an increase in its transcription/transactivation. Thusalterations in nuclear levels of β-catenin are of biologicalimportance only when there is a parallel effect upon itstransactivation function. Consistent with this paradigm, wedemonstrate a decrease in TOPFlash-luciferase activity andthe levels of nuclear β-catenin/TCF-4 transcription complexesin DU-145 CaP cells treated with PUGNAc. The O-GlcNAcyla-tion-associated decrease in transcriptional activity of β-catenin/TCF-4 is further underscored by the attenuation ofmRNA transcript of Cyclin D1 and VEGF A, two TCF-4/β-catenin target genes, with PUGNAc treatment. To this end, theactivities of several transcription factors are known to bemodified by O-GlcNAcylation [45,46].
In conclusion,we propose thatwhile suppressed phosphor-ylation of β-catenin increases its cytoplasmic levels, sup-pressed O-GlcNAcylation increases its nuclear level. The twopost-translational modifications will therefore have to bealtered in synchrony to regulate the transcriptional functionof this protein. Interestingly, our results show that thecanonical Wnt signaling pathway, which is well known toregulate the phosphorylation of β-catenin, also appears toexert a regulatory role upon its O-GlcNAcylation. Control ofcellular β-catenin in cancerswhere there aremutations in APCor β-catenin presents an almost non-surmountable task. Inlight of this, O-GlcNAcylation by virtue of its function inregulating the localization of this protein in the nucleusprovides an additional level of control to the transcriptionalactivity of this protein.
Future directions
The sites of O-GlcNAcylation of β-catenin are unknown atpresent. TheO-GlcNAcmodification, like all post-translationalmodifications, are substoichiometric and often labile, makingtheir identification difficult [47]. The lability of the glycosidiclinkage makes evaluation of O-GlcNAc using mass spectro-metry difficult [47]. Thus we are presently involved in thelengthy process of determining the sites of O-GlcNAcylationby sequential site directed mutagenesis of putative O-GlcNA-cylation sites. Identification of the O-GlcNAcylation sites willallow the generation of O-GlcNAc-site-specific antibodies inthe same manner as the phosphor-specific antibodies. Thiswill subsequently greatly expand our scope in understandingthe site-specific function of this modification in β-catenin, atranscription factor whose cellular levels and subcellulardistribution is intimately involved in promoting the progres-sion of many cancers.
Acknowledgments
The work reported in this study was supported by grants fromThe Canadian Institute for Health Research (to SP) and theCancer Research Society (to SP).

2786 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 4 ( 2 0 0 8 ) 2 7 7 4 – 2 7 8 7
R E F E R E N C E S
[1] S.Y. Lin, W. Xia, J.C. Wang, K.Y. Kwong, B. Spohn, Y. Wen, R.G.Pestell, M.C. Hung, b-catenin, a novel prognostic marker forbreast cancer: its roles in cyclin D1 expression and cancerprogression, Proc. Natl. Acad. Sci. USA 97 (2000) 4262–4266.
[2] O. Tetsu, F. McCormick, b-catenin regulates expression ofcyclin D1 in colon carcinoma cells, Nature 398 (1999) 422–426.
[3] T.C. He, A.B. Sparks, C. Rago, H. Hermeking, L. Zawe, L.T.da Costa, P.J. Morin, B. Vogelstein, K.W. Kinzler, Identificationof c-myc as a target of the APC pathway, Science 281 (1998)1509–1512.
[4] H.C. Crawford, B.M. Fingleton, L.A. Rudolph-Owen, K.J. Goss,B. Rubinfeld, P. Polakis, L.M. Matrisian, Themetalloproteinasematrilysin is a target of b-catenin transactivation in intestinaltumors, Oncogene 18 (1999) 2883–2891.
[5] B. Mann, M. Gelos, A. Siedow, M.L. Hanski, A. Gratchev,M. Ilyas, W.F. Bodmer, M.P. Moyer, E.O. Riecken, H.J. Buhr,C. Hanski, Target genes of b-catenin-T cellfactor/lymphoid-enhancer-factor signalling in human coloncarcinoma, Proc. Natl. Acad. Sci. USA. 96 (1999) 1603–1608.
[6] H. Aberle, A. Bauer, J. Stappert, A. Kispert, R. Kemler,b-catenin is a target for the ubiquitin–proteasome pathway,EMBO J. 16 (1997) 3797–3804.
[7] H. Clevers, Wnt\beta-catenin signalling in development anddisease, Cell 127 (2006) 469–480.
[8] W. Zhu, B. Leber, D.W. Andrews, Cytoplasmic O-glycosylationprevents cell surface transport of E-cadherinduring apoptosis,EMBO J. 20 (2001) 5999–6007.
[9] K. Vosseller, L. Wells, G.W. Hart, NucleocytoplasmicO-glycosylation: O-GlcNAc and functional proteomics,Biochimie 83 (2001) 575–581.
[10] L. Wells, K. Vosseller, G.W. Hart, Glycosylation ofnucleocytoplasmic proteins: signal transduction andO-GlcNAc, Science 291 (2001) 2376-23788.
[11] Y. Gao, L. Wells, F.I. Comer, G.J. Parker, G.W. Hart, DynamicO-glycosylation of nuclear and cytosolic proteins: cloningand characterization of a neutral, cytosolic beta-N-acetylglucosaminidase from human brain, J. Biol. Chem. 276(2001) 9838–9845.
[12] L.K. Kreppel, M.A. Blomberg, G.W. Hart, Dynamicglycosylation of nuclear and cytosolic proteins. Cloning andcharacterization of a unique O-GlcNAc transferase withmultiple tetratricopeptide repeats, J. Biol.Chem. 272 (1997)9308–9315.
[13] C.S. Arnold, G.V. Johnson, R.N. Cole, D.L. Dong, M. Lee, G.W.Hart, The microtubule-associated protein tau is extensivelymodified with O-linked N-acetylglucosamine, J. Biol. Chem.271 (1996) 28741–28744.
[14] F.I. Comer, G.W. Hart, Reciprocity between O-GlcNAc andO-phosphate on the carboxyl terminal domain of RNApolymerase II, Biochemistry 40 (2001) 7845–7852.
[15] S. Persad, A.A. Troussard, T.R. McPhee, D.J. Mulholland, S.Dedhar, Tumor suppressor PTEN inhibits nuclear accumulationof b-catenin and T cell/lymphoid enhancer factor1-mediatedtranscriptional activation, J. Cell. Biol. 153 (2001) 1161–1174.
[16] V. Korinek, N. Barker, P.J. Morin, D. vanWiechen, R. deWeger,K.W. Kinzler, B. Vogelstein, H. Clevers, Constitutivetranscriptional activation by a b-catenin-Tcf complex inAPC −/− colon carcinoma, Science 275 (1997) 1784–1787.
[17] J.S. Michaelson, P. Leder, b-catenin is a downstreameffector of wnt-mediated tumorigenesis in mammary gland,Oncogene 20 (2001) 5093–5099.
[18] J.P. Morin, b-catenin signalling in cancer, Bioessay 21 (1999)1021–1030.
[19] R.S. Haltiwanger, K. Grove, G.A. Philipsberg, Modulation ofO-linked N-acetylglucosamine levels on nuclear andcytoplasmic proteins in vivo using the peptide O-GlcNAc-
beta-N-acetylglucosaminidase inhibitor O-(2-acetamido-2-deoxy-D-glucopyranosylidene) amino-N-phenylcarbamate,J. Biol. Chem. 273 (1998) 3611–3617.
[20] G.W. Hart, M.P. Housley, C. Slawson, Cycling of O-linkedb-N-acetylyglucosamine on nucleocytoplasmic proteins,Nature 446 (2007) 1017–1022.
[21] R.S. Haltiwanger, M.A. Blomberg, G.W. Hart, Glycosylation ofnucleocytoplasmic proteins. Purification and characterizationof a uridine diphospho-N-acetylglucoseamne: polypeptideb-N-acetylglucosaminyltransferase, J. Biol. Chem. 267 (1992)9005–9013.
[22] G. Majumdar, A. Harrington, J. Hungerford, A.Martinez-Hernandez, I.C. Gerling, R. Raghow, S. Solomon,Insulin dynamically regulates calmodulin gene expressionby sequential O-glycosylation and phosphorylation of Sp1and its subcellular compartmentalization in liver cells, J. Biol.Chem. 281 (2006) 3642–3650.
[23] X. Zhang, J.P. Gaspard, D.C. Chung, Regulation of vascularendothelial growth factor by the Wnt and K-ras pathways incolonic neoplasia, Cancer Res. 61 (2001) 6050–6054.
[24] V. Easwaran, S.H. Lee, L. Inge, L. Guo, C. Goldbeck, E. Garrett,M. Wiesmann, P.D. Garcia, J.H. Fuller, V. Chan, F. Randazzo, R.Gundel, R.S. Warren, J. Escobedo, S.L. Aukerman, R.N. Taylor,W.J. Fantl, β-catenin regulates vascular endothelial growthfactor expression in colon cancer, Cancer Res. 63 (2003)3145–3153.
[25] J.R. Miller, A.M. Hocking, J.D. Brown, R.T. Moon, Mechanismand function of signal transduction by Wnt/beta-catenin andWnt/Ca+ pathways, Oncogene 18 (1999) 7860–7872.
[26] R.H. Giles, J.H. van Es, H. Clevers, Caught up in a Wnt storm:Wnt signalling in cancer, Biochim. Biophys. Acta 1653 (2003)1–24.
[27] P.J. Morin, A.B. Sparks, V. Korinek, N. Barker, H. Clevers,B. Vogelstein, K.W. Kinzler, Activation of beta-catenin-Tcfsignalling in colon cancer by mutations in beta-catenin orAPC, Science 275 (1997) 1787–1790.
[28] B. Rubinfeld, P. Robbins, M. El-Gamil, I. Albert, E. Porfiri,P. Polakis, Stabilization of beta-catenin by genetic defects inmelanoma cell lines, Science 275 (1997) 1790–1792.
[29] E. Conti, E. Izaurralde, Nucleocytoplasmic transport entersthe atomic age, Curr. Opin. Cell Biol. 13 (2001) 310–319.
[30] U. Kutay, S. Guttinger, Leucine-rich nuclear-export signals:born to be weak, Trends Cell Biol. 15 (2005) 121–124.
[31] B. Lee, A.E. Cansizoglu, K.E. Suel, T.H. Louis, Z. Zhang, Y.M.Chook, Rules for nuclear localization sequence recognition bykaryopherin beta 2, Cell 126 (2006) 543–558.
[32] S. Shikano, B. Cobiltz, H. Sun, M. Li, Genetic isolation oftransport signals directing cell surface expression, Nat. CellBiol. 7 (2005) 985–992.
[33] D.M. Snow, G.W. Hart, Nuclear and cytoplasmic glycosylation,Int. Rev. Cytol. 181 (1998) 43–74.
[34] C. Guinez, W. Morelle, J.C. Michalski, T. Lefebvre, O-GlcNAcglycosylation: a signal for nuclear transport of cytosolicproteins? Int. J. Biochem. Cell Biol. 37 (2004) 765–774.
[35] N. Funayama, F. Fagotto, P. McCrea, B.M. Gumbiner,Embryonic axis induction by armadillo repeat domain ofb-catenin: evidence for intracellular signalling, J. Cell. Biol.128 (1995) 9–968.
[36] M. Koike, S. Kose, M. Furuta, N. Taniguchi, F. Yokoya, Y.Yoneda, N. Imamoto, b-catenin shows an overlappingsequence requirement but distinct molecular interactions forits bidirectional passage through nuclear pores, J. Biol. Chem.279 (2004) 34038–34047.
[37] F. Fagotto, U. Gluck, B.M. Gumbiner, Nuclear localizationsignal-independent and importin/karyopherin-independentnuclear import of b-catenin, Curr. Biol. 8 (1998) 181–190.
[38] E.K. Suh, B.M. Gumbiner, Translocation of b-catenin into thenucleus independent of FG-rich nucleoporins, Exp. Cell Res.290 (2003) 447–456.

2787E X P E R I M E N T A L C E L L R E S E A R C H 3 1 4 ( 2 0 0 8 ) 2 7 7 4 – 2 7 8 7
[39] A.I. Barth, A.L. Pollack, Y. Altschuler, K.E. Mostov, W.J. Nelson,NH2-terminal deletion of β-catenin results in stablec-localization of mutant b-catenin with adenomatouspolyposis coli protein and altered MDCK cell adhesion, J. CellBiol. 136 (1997) 693–706.
[40] N.S. Tolwinski, E. Weischaus, Armadillo nuclear import isregulated by cytoplasmic anchor axin and nuclear anchordTCF/Pan, Development 128 (2001) 2107–2117.
[41] J. Behrens, J.P. von Kries, M. Kuhl, L. Bruhn, D. Wedlich, R.Grosschedl, W. Birchmeier, Functional interaction of β-catenin with transcription factor LEF-1, Nature 382 (1996)638–642.
[42] O. Huber, R. Korn, J. McLaughlin, M. Ohsugi, B.G. Herrmann, R.Kemler, Nuclear localization of β-catenin by interaction withtranscription factor LEF-1, Mech. Dev. 59 (1996) 3–10.
[43] T. Kramps, O. Peter, E. Bunner, D. Nellen, B. Froesch, S.Chatterjee, M. Murone, S. Zullig, K. Basler, Wnt/Wingless
signalling requires BCL9/Legless-mediated recruitment ofPygopus to the nuclear β-catenin-TCF complex, Cell 109 (2002)47–60.
[44] F. Townsley, A. Cliffeand, M. Bienz, Pygopus and Leglesstarget Armadillo/b-catenin to the nucleus to enable itstranscriptional co-activator function, Nat. Cell Biol. 6 (2004)626–633.
[45] F.I. Comer, G.W. Hart, O-GlcNAc and the control of geneexpression, Biochim. Biophys. Acta. 1473 (1999) 161–171.
[46] L. Wells, G.W. Hart, O-GlcNAc turns twenty: functionalimplications for post-translational modifications of nuclearand cytosolic proteins with a sugar, FEBS Lett. 546 (2003)154–158.
[47] L. Wells, K. Vosseller, R.N. Cole, J.M. Cronshaw, M.J. Matunis,G.W. Hart, Mapping sites of O-GlcNAc modification usingaffinity tags for serine and threonine post-translationalmodifications, Mol. Cell. Proteom. 1 (2002) 791–804.





























