Embed Size (px)
DESCRIPTION
Nucleic Acids: Chemistry & Structure. Andy Howard Introductory Biochemistry 8 October 2009. Nucleic acid chemistry Pyrimidines: C, U, T Purines: A, G Nucleosides & nucleotides Oligo- and polynucleotides. DNA duplexes and helicity DNA sequencing DNA secondary structure: A, B, Z - PowerPoint PPT Presentation
Citation preview

10/08/2009Biochemistry:Nucleic Acids I
Nucleic Acids:Chemistry & Structure
Andy HowardIntroductory Biochemistry
8 October 2009

10/08/2009 Biochemistry:Nucleic Acids I
p. 2 of 57
What we’ll discuss
Nucleic acid chemistry Pyrimidines: C, U, T
Purines: A, G Nucleosides & nucleotides
Oligo- and polynucleotides
DNA duplexes and helicity
DNA sequencing DNA secondary structure: A, B, Z
Folding kinetics

10/08/2009 Biochemistry:Nucleic Acids I
p. 3 of 57
Chemistry Nobel Prize 2009 Structural studies of the ribosome
Venki Ramakrishnan, LMB Cambridge Thomas Steitz, HHMI Yale University Ada Yonath, Weizmann Institute
QuickTime™ and a decompressor
are needed to see this picture.
QuickTime™ and a decompressor
are needed to see this picture.
QuickTime™ and a decompressor
are needed to see this picture.

10/08/2009 Biochemistry:Nucleic Acids I
p. 4 of 57
Pyrimidines Single-ring nucleic acid bases 6-atom ring; always two nitrogens in the ring, meta to one another
Based on pyrimidine, although pyrimidine itself is not a biologically important molecule
Variations depend on oxygens and nitrogens attached to ring carbons
Tautomerization possible Note line of symmetry in pyrimidine structure
N
N
pyrimidine
1
2
3
4
5
6

10/08/2009 Biochemistry:Nucleic Acids I
p. 5 of 57
Uracil and thymine
Uracil is a simple dioxo derivative of pyrimidine: 2,4-dioxopyrimidine
Thymine is 5-methyluracil Uracil is found in RNA; Thymine is found in DNA
We can draw other tautomers where we move the protons to the oxygens
HN
OHN O
uracil
HN
O NH
O
thymine

10/08/2009 Biochemistry:Nucleic Acids I
p. 6 of 57
Tautomers
Lactam and Lactim forms
Getting these right was essential to Watson & Crick’s development of the DNA double helical model
HN
OHN O
uracil - lactam
NH
ONO
uracil - lactimH
HN
O NH
O
thymine - lactam
HN
O N OH
thymine - lactim

10/08/2009 Biochemistry:Nucleic Acids I
p. 7 of 57
Cytosine
This is 2-oxo,4-aminopyrimidine
It’s the other pyrimidine base found in DNA & RNA
Spontaneous deamination (CU) Again, other tautomers can be drawn
N
OHN NH2
cytosine

10/08/2009 Biochemistry:Nucleic Acids I
p. 8 of 57
Cytosine:amino and imino forms Again, this tautomerization needs to be kept in mind
N
OHN NH
cytosine -imino form
N
OHN NH2
cytosine -amino form

10/08/2009 Biochemistry:Nucleic Acids I
p. 9 of 57
Purines Derivatives of purine; again, the root molecule isn’t biologically important
Six-membered ring looks a lot like pyrimidine
Numbering works somewhat differently: note that the glycosidic bonds will be to N9, whereas it’s to N1 in pyrimidines
HN
NN
N
purine
1
2
3
4
56 7
8
9
10/08/2009 Biochemistry:Nucleic Acids I
p. 10 of 57
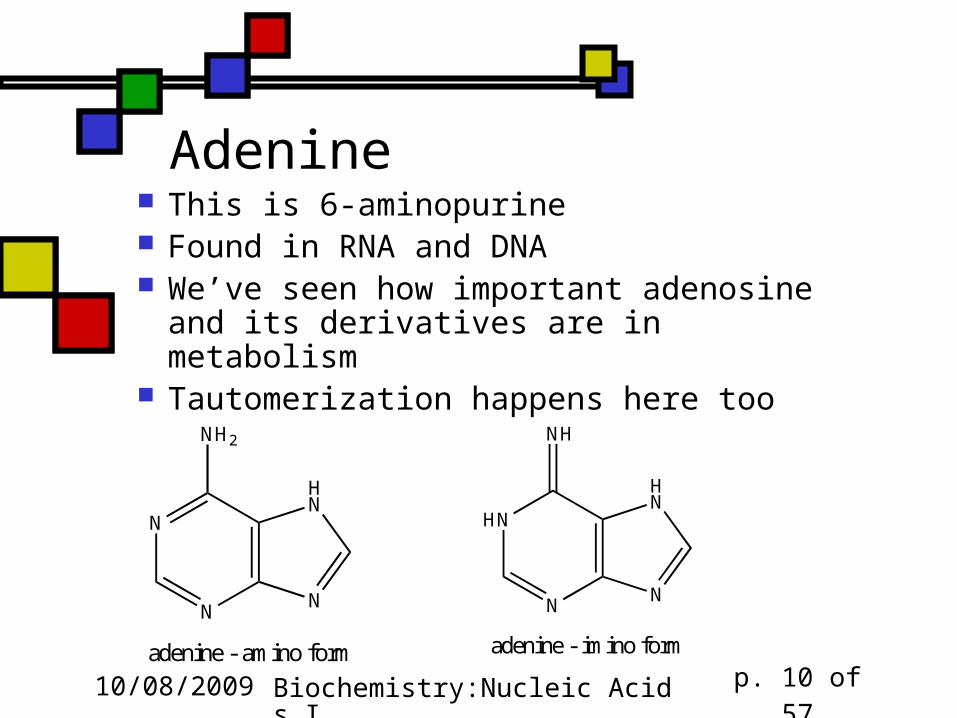
Adenine This is 6-aminopurine Found in RNA and DNA We’ve seen how important adenosine and its derivatives are in metabolism
Tautomerization happens here too
N
N
NH2
N
HN
adenine - amino form
HN
N
NH
N
HN
adenine - imino form
10/08/2009 Biochemistry:Nucleic Acids I
p. 11 of 57
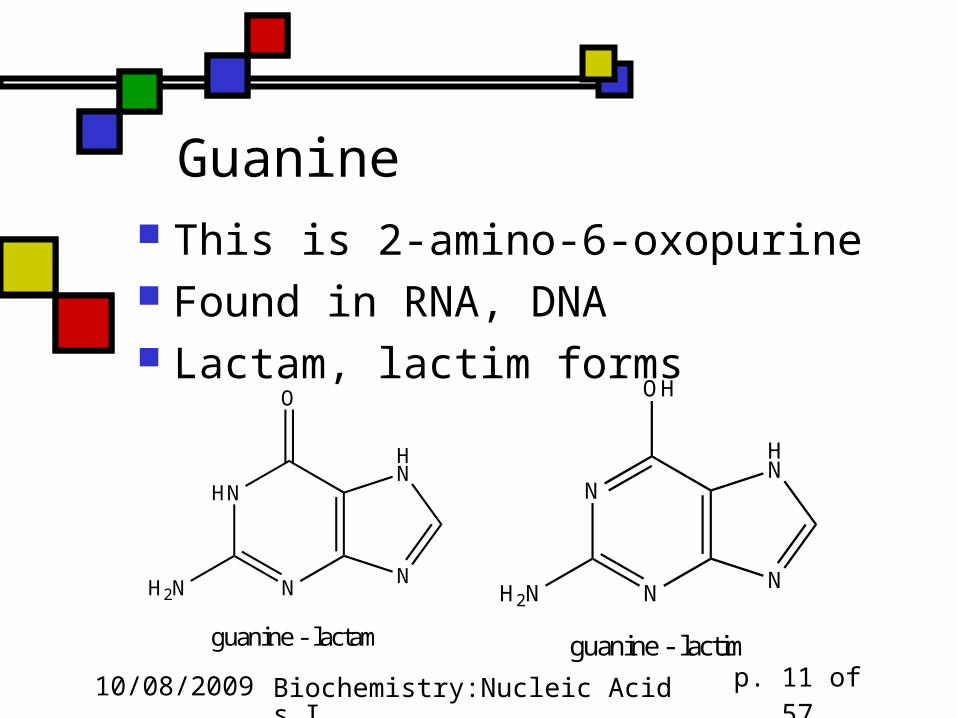
Guanine This is 2-amino-6-oxopurine Found in RNA, DNA Lactam, lactim forms
HN
NNH2N
HN
O
guanine - lactam
HN
NNH2N
N
OH
guanine - lactim

10/08/2009 Biochemistry:Nucleic Acids I
p. 12 of 57
Other natural purines
Hypoxanthine and xanthine are biosynthetic precursors of A & G
Urate is important in nitrogen excretion pathways

10/08/2009 Biochemistry:Nucleic Acids I
p. 13 of 57
Tautomerization and H-bonds Lactam forms predominate at neutral pH
This influences which bases are H-bond donors or acceptors
Amino groups in C, A, G make H-bonds So do ring nitrogens at 3 in pyrimidines and 1 in purines
… and oxygens at 4 in U,T, 2 in C, 6 in G

10/08/2009 Biochemistry:Nucleic Acids I
p. 14 of 57
Nucleosides
As mentioned in ch. 8, these are glycosides of the nucleic acid bases
Sugar is always ribose or deoxyribose
Connected nitrogen is: N1 for pyrimidines (on 6-membered ring) N9 for purines (on 5-membered ring)
NR1R2
OH
HO
O
HO
N-glycoside of ribofuranose

10/08/2009 Biochemistry:Nucleic Acids I
p. 15 of 57
Pyrimidine nucleosides
Drawn here in amino and lactam forms
OH
OHHO
ON
ONH2N
cytidine
OH
OHHO
ON
ONH
O
uridine

10/08/2009 Biochemistry:Nucleic Acids I
p. 16 of 57
Pyrimidine deoxynucleosides
OH
OHH
ON
ONH
O
2'-deoxyuridine
OH
OHH
ON
ONH
O
2'-deoxythymidineOH
OH
ON
ONH2N
deoxycytidine

10/08/2009 Biochemistry:Nucleic Acids I
p. 17 of 57
A tricky nomenclature issue Remember that thymidine and its phosphorylated derivatives ordinarily occur associated with deoxyribose, not ribose
Therefore many people leave off the deoxy- prefix in names of thymidine and its derivatives: it’s usually assumed.

10/08/2009 Biochemistry:Nucleic Acids I
p. 18 of 57
Purine nucleosides
Drawn in amino and lactam forms
OH
HO
HO
O
N
N
NH2
N
N
adenosine
OH
HO
HO
O
N
N
O
HN
H2N N
guanosine

10/08/2009 Biochemistry:Nucleic Acids I
p. 19 of 57
Purine deoxynucleosides
OH
HO
O
N
N
O
HN
H2N N
deoxyguanosine
OH
HO
O
N
N
NH2
N
N
deoxyadenosine

10/08/2009 Biochemistry:Nucleic Acids I
p. 20 of 57
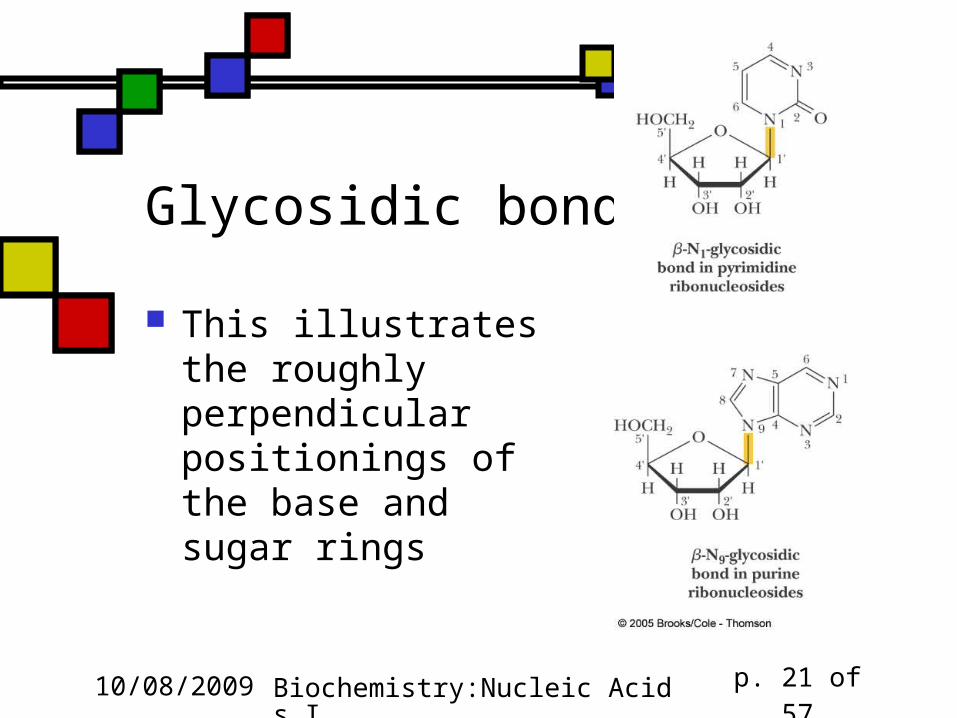
Conformations around the glycosidic bond Rotation of the base around the glycosidic bond is sterically hindered
In the syn conformation there would be some interference between the base and the 2’-hydroxyl of the sugar
Therefore pyrimidines are always anti, and purines are usually anti
Furanose and base rings are roughly perpendicular
10/08/2009 Biochemistry:Nucleic Acids I
p. 21 of 57
Glycosidic bonds
This illustrates the roughly perpendicular positionings of the base and sugar rings

10/08/2009 Biochemistry:Nucleic Acids I
p. 22 of 57
Solubility of nucleosides and lability of glycosidic linkages The sugar makes nucleosides more soluble than the free bases
Nucleosides are generally stable to basic hydrolysis at the glycosidic bond
Acid hydrolysis: Purines: glycosidic bond fairly readily hydrolyzed
Pyrimidines: resistant to acid hydrolysis

10/08/2009 Biochemistry:Nucleic Acids I
p. 23 of 57
Chirality in nucleic acids Bases themselves are achiral Four asymmetric centers in ribofuranose, counting the glycosidic bond.
Three in deoxyribofuranose Glycosidic bond is one of those 4 or 3.
Same for nucleotides:phosphates don’t add asymmetries

10/08/2009 Biochemistry:Nucleic Acids I
p. 24 of 57
Mono-phosphorylated nucleosides
We have specialized names for the 5’-phospho derivatives of the nucleosides, i.e. the nucleoside monophosphates:
They are nucleotides Adenosine 5’-monophosphate = AMP = adenylate
GMP = guanylate CMP = cytidylate UMP = uridylate
P
O
O-
O-O
HO
HO
O
N
N
NH2
N
N
adenylate

10/08/2009 Biochemistry:Nucleic Acids I
p. 25 of 57
pKa’s for base N’s and PO4’sNucleotide
pKa base-N
pK1 of PO4
pK2 of PO4
5’-AMP 3.8(N-1) 0.9 6.1
5’-GMP 9.4 (N-1)
0.7 6.1
2.4 (N-7)
5’-CMP 4.5 (N-3)
0.8 6.3
5’-UMP 9.5 (N-3)
1.0 6.4

10/08/2009 Biochemistry:Nucleic Acids I
p. 26 of 57
UV absorbance These aromatic rings absorb around 260

10/08/2009 Biochemistry:Nucleic Acids I
p. 27 of 57
Deoxynucleotides Similar nomenclature dAMP = deoxyadenylate
dGMP = deoxyguanylate
dCMP = deoxycytidylate
dTTP (= TTP) = deoxythymidylate = thymidylate
P
O
O-
O-O
HO
O
N
N
O
HN
H2N N
deoxyguanylate

10/08/2009 Biochemistry:Nucleic Acids I
p. 28 of 57
Di and triphosphates Phosphoanhydride bonds link second and perhaps third phosphates to the 5’-OH on the ribose moiety
OHHO
O
N
O
N
H2NP
O
O
O-O-O
P
O
O-O
P
O
OH
cytidine triphosphate
Mg2+

10/08/2009 Biochemistry:Nucleic Acids I
p. 29 of 57
Cyclic phospho-diesters
3’ and 5’ hydroxyls are both involvedin -O-P-O bonds
cAMP and cGMP are the important ones(see earlier in the course!)

10/08/2009 Biochemistry:Nucleic Acids I
p. 30 of 57
Oligomers and Polymers
Monomers are nucleotides or deoxynucleotides
Linkages are phosphodiester linkages between 3’ of one ribose and 5’ of the next ribose
It’s logical to start from the 5’ end for synthetic reasons

10/08/2009 Biochemistry:Nucleic Acids I
p. 31 of 57
Typical DNA dinucleotide
Various notations: this is pdApdCp Leave out the p’s if there’s a lot of them!
P
O
-O O-
O
O
NN
O
HN
NH2
N P
O
-O
O
O
O
ON
O N NH2
P
O--O
O

10/08/2009 Biochemistry:Nucleic Acids I
p. 32 of 57
DNA structure
Many years of careful experimental work enabled fabrication of double-helical model of double-stranded DNA
Explained [A]=[T], [C]=[G]
Specific H-bonds stabilize double-helical structure: see fig. 10.20

10/08/2009 Biochemistry:Nucleic Acids I
p. 33 of 57
What does double-stranded DNA really look like? Picture on previous slide emphasizes only the H-bond interactions; it ignores the orientation of the sugars, which are actually tilted relative to the helix axis
Planes of the bases are almost perpendicular to the helical axes on both sides of the double helix

10/08/2009 Biochemistry:Nucleic Acids I
p. 34 of 57
Sizes (cf fig. 10.20, 11.7)
Diameter of the double helix: 2.37nm Length along one full turn:10.4 base pairs = pitch = 3.40nm
Distance between stacked base pairs = rise = 0.33 nm
Major groove is wider and shallower;minor groove is narrower and deeper

10/08/2009 Biochemistry:Nucleic Acids I
p. 35 of 57
What stabilizes this?
Variety of stabilizing interactions Stacking of base pairs Hydrogen bonding between base pairs
Hydrophobic effects (burying bases, which are less polar)
Charge-charge interactions:phosphates with Mg2+ and cationic proteins
Courtesy dnareplication.info

10/08/2009 Biochemistry:Nucleic Acids I
p. 36 of 57
How close to instability is it? Pretty close. Heating DNA makes it melt: fig. 11.14
pH > 10 separates strands too The more GC pairs, the harder it is to melt DNA thermally Weaker stacking interactions in A-T One more H-bond per GC than per AT

10/08/2009 Biochemistry:Nucleic Acids I
p. 37 of 57
iClicker quiz, 1st question
1. What positions of a pair of aromatic rings leads to stabilizing interactions? (a) Parallel to one another (b) Perpendicular to one another (c) At a 45º angle to one another (d) Both (a) and (b) (e) All three: (a), (b), and ( c)

10/08/2009 Biochemistry:Nucleic Acids I
p. 38 of 57
iClicker question 2
2. Which has the highest molecular mass among the compounds listed? (a) cytidylate (b) thymidylate (c) adenylate (d) adenosine triphosphate (e) they’re all the same MW

10/08/2009 Biochemistry:Nucleic Acids I
p. 39 of 57
Base composition for DNA
As noted, [A]=[T], [C]=[G] because of base pairing
[A]/[C] etc. not governed by base pairing Can vary considerably (table 10.3) E.coli : [A], [C] about equal Mycobacterium tuberculosis: [C] > 2*[A] Mammals: [C] < 0.74*[A]

10/08/2009 Biochemistry:Nucleic Acids I
p. 40 of 57
Supercoiling Refers to levels of organization of DNA beyond the immediate double-helix
We describe circular DNA as relaxed if the closed double helix could lie flat
It’s underwound or overwound if the ends are broken, twisted, and rejoined.
Supercoils restore 10.4 bp/turn relation upon rejoining

10/08/2009 Biochemistry:Nucleic Acids I
p. 41 of 57
Supercoiling and flat DNA
Diagram courtesy SIU Carbondale

10/08/2009 Biochemistry:Nucleic Acids I
p. 42 of 57
Sanger dideoxy method Incorporates DNA replication as an analytical tool for determining sequence
Uses short primer that attaches to the 3’ end of the ssDNA, after which a specially engineered DNA polymerase
Each vial includes one dideoxyXTP and 3 ordinary dXTPs; the dideoxyXTP will be incorporated but will halt synthesis because the 3’ position is blocked.
See figs. 11.3 & 11.4 for how these are read out

10/08/2009 Biochemistry:Nucleic Acids I
p. 43 of 57
Automating dideoxy sequencing Laser fluorescence detection allows for primer identification in real time
An automated sequencing machine can handle 4500 bases/hour
That’s one of the technologies that has made large-scale sequencing projects like the human genome project possible

10/08/2009 Biochemistry:Nucleic Acids I
p. 44 of 57
DNA secondary structures
If double-stranded DNA were simply a straight-legged ladder: Base pairs would be 0.6 nm apart Watson-Crick base-pairs have very uniform dimensions because the H-bonds are fixed lengths
But water could get to the apolar bases So, in fact, the ladder gets twisted into a helix.
The most common helix is B-DNA, but there are others. B-DNA’s properties include: Sugar-sugar distance is still 0.6 nm Helix repeats itself every 3.4 nm, i.e. 10 bp

10/08/2009 Biochemistry:Nucleic Acids I
p. 45 of 57
Properties of B-DNA Spacing between base-pairs along helix axis = 0.34 nm
10 base-pairs per full turn So: 3.4 nm per full turn is pitch length
Major and minor grooves, as discussed earlier
Base-pair plane is almost perpendicular to helix axis

10/08/2009 Biochemistry:Nucleic Acids I
p. 46 of 57
Major groove in B-DNA
H-bond between adenine NH2 and thymine ring C=O
H-bond between cytosine amine and guanine ring C=O
Wide, not very deep

10/08/2009 Biochemistry:Nucleic Acids I
p. 47 of 57
Minor groove in B-DNA
H-bond between adenine ring N and thymine ring NH
H-bond between guanine amine and cytosine ring C=O
Narrow but deep

10/08/2009 Biochemistry:Nucleic Acids I
p. 48 of 57
Cartoon of AT pair in B-DNA

10/08/2009 Biochemistry:Nucleic Acids I
p. 49 of 57
Cartoon of CG pair in B-DNA

10/08/2009 Biochemistry:Nucleic Acids I
p. 50 of 57
What holds duplex B-DNA together? H-bonds (but just barely)
Electrostatics: Mg2+ –PO4-2
van der Waals interactions - interactions in bases Solvent exclusion
Recognize role of grooves in defining DNA-protein interactions

10/08/2009 Biochemistry:Nucleic Acids I
p. 51 of 57
Helical twist (fig. 11.9a)
Rotation about the backbone axis
Successive base-pairs rotated with respect to each other by ~ 32º

10/08/2009 Biochemistry:Nucleic Acids I
p. 52 of 57
Propeller twist
Improves overlap of hydrophobic surfaces
Makes it harder for water to contact the less hydrophilic parts of the molecule

10/08/2009 Biochemistry:Nucleic Acids I
p. 53 of 57
A-DNA (figs. 11.10)
In low humidity this forms naturally Not likely in cellular duplex DNA, but it does form in duplex RNA and DNA-RNA hybrids because the 2’-OH gets in the way of B-RNA
Broader 2.46 nm per full turn 11 bp to complete a turn
Base-pairs are not perpendicular to helix axis:tilted 19º from perpendicular

10/08/2009 Biochemistry:Nucleic Acids I
p. 54 of 57
Z-DNA (figs. 11.10)
Forms in alternating Py-Pu sequences and occasionally in PyPuPuPyPyPu, especially if C’s are methylated
Left-handed helix rather than right
Bases zigzag across the groove

10/08/2009 Biochemistry:Nucleic Acids I
p. 55 of 57
Getting from B to Z
Can be accomplished without breaking bonds
… even though purines have their glycosidic bonds flipped (anti -> syn) and the pyrimidines are flipped altogether!

10/08/2009 Biochemistry:Nucleic Acids I
p. 56 of 57
DNA is dynamic Don’t think of these diagrams as static
The H-bonds stretch and the torsions allow some rotations, so the ropes can form roughly spherical shapes when not constrained by histones
Shape is sequence-dependent, which influences protein-DNA interactions

10/08/2009 Biochemistry:Nucleic Acids I
p. 57 of 57
What does DNA do? Serve as the storehouse and the propagator of genetic information:That means that it’s made up of genes Some code for mRNAs that code for protein Others code for other types of RNA Genes contain non-coding segments (introns)
But it also contains stretches that are not parts of genes at all and are serving controlling or structural roles
Avoid the term junk DNA!





























