Embed Size (px)
Citation preview

Current Biology Vol 22 No 12R496
conditioning or test trials. Furthermore,the authors had participants reportif they saw a face on each trial;in fact participants had to makea two-alternative forced-choicedecision whether the face was maleor female — participants’ decisionswere just below chance. After thisdiscrimination task participantshad to rate the confidence of theirchoice — confidence ratings wereno higher on correct trials thanincorrect trials.
The conditioning or learning effectsoutside of awareness reported byRaio et al. [3] display some distinctcharacteristics that differentiate themfrom learning with awareness. Thelearning effects appeared very rapidlyand subsequently diminished veryrapidly. Unlike normal learning theseeffects faded during furtherconditioning, whereas typically in thiskind of conditioning experiment thelearning would continue beforestabilising. Such brevity in associativelearning dynamics is clearly distinctfrom typical conditioning effects,which often last for days. Might thislearning outside of awareness betapping into a categorically differentlearning mechanism, or perhapsa subset of normal learning processes?This is an interesting idea that iscompatible with the data in the newstudy [3].
Raio et al. [3] did include a fully visiblecondition, which showed a verydifferent temporal learning profile. Intheir visible condition, however, boththe learning and test-trials were bothvisible, while in the unaware conditionboth the training and test-trials wereinvisible. Hence, we do not havea conscious measure of conditioningoutside of awareness, only anunconscious one. To help clarify theunderlying mechanism what is neededis a third condition in which only thetest trials are visible while the trainingtrials remain suppressed fromawareness. Such an experiment wouldhelp tease apart the nature of thisunconscious learning.
Previous claims of unconsciousconditioning have been criticised ona number of methodological groundssuch as trial sequence artifacts, failureto assess participant hypotheses, andinsensitivity to partial awareness [2]. Infact, some researchers have gone sofar as to argue that all conditioninginvolves cognitive representation andhence conscious awareness [12].
Others maintain that conditioning iscarried out by a separately evolvedspecialised system [13,14]. Willcontinuous flash suppression finallyprovide the experimental tool toresolve this long-standing debate?Watch this space!
Associative learning is thought toform the backbone of the mechanismsof many psychological disorders andtheir treatments [15,16]. Manybehavioural interventions forpsychological disorders rely oncounterconditioning or extinction-likeapproaches, such as cognitivebehavioural therapy. Does this newpaper by Raio et al. [3] shed light on anynew clinical treatment possibilities?Potentially yes, if mechanisms ofassociative learning can operatewithout awareness, it is possible toimagine a future non-intrusivetreatment option that might be run onpatients without their consciousinvolvement. However, the brief lifetimeof the effects in the new papermight limit any potential clinicalapplications.
References1. Carew, T.J., Walters, E.T., and Kandel, E.R.
(1981). Classical conditioning in a simplewithdrawal reflex in Aplysia californica. J.Neurosci. 1, 1426–1437.
2. Lovibond, P.F., and Shanks, D.R. (2002). Therole of awareness in Pavlovian conditioning:empirical evidence and theoreticalimplications. J. Exp. Psychol. Anim. Behav.Proc. 28, 3–26.
3. Raio, C., Carmel, D., Carrasco, M., andPhelps, E.A. (2012). Nonconscious fear isquickly acquired but swiftly forgotten. Curr.Biol. 22, R477–R479.
4. Tsuchiya, N., and Koch, C. (2005). Continuousflash suppression reduces negativeafterimages. Nat. Neurosci. 8, 1096–1101.
5. Tsuchiya, N., Koch, C., Gilroy, L.A., andBlake, R. (2006). Depth of interocularsuppression associated with continuous flashsuppression, flash suppression, and binocularrivalry. J. Vis.n 6, 1068–1078.
6. Stein, T., Hebart, M.N., and Sterzer, P. (2011).Breaking continuous flash suppression: a newmeasure of Uunconscious processing duringinterocular suppression? Front. Hum. Neurosci.5, 167.
7. Pearson, J., and Clifford, C.W. (2005).Suppressed patterns alter vision duringbinocular rivalry. Curr. Biol. 15, 2142–2148.
8. Blake, R., Tadin, D., Sobel, K.V., Raissian, T.A.,and Chong, S.C. (2006). Strength of early visualadaptation depends on visual awareness. Proc.Natl. Acad. Sci. USA 103, 4783–4788.
9. Pasley, B.N., Mayes, L.C., and Schultz, R.T.(2004). Subcortical discrimination ofunperceived objects during binocular rivalry.Neuron 42, 163–172.
10. Almeida, J., Mahon, B.Z., Nakayama, K., andCaramazza, A. (2008). Unconscious processingdissociates along categorical lines. Proc. Natl.Acad. Sci. USA 105, 15214–15218.
11. Jiang, Y., Costello, P., Fang, F., Huang, M., andHe, S. (2006). A gender- and sexualorientation-dependent spatial attentional effectof invisible images. Proc. Natl. Acad. Sci. USA103, 17048–17052.
12. Mitchell, C.J., De Houwer, J., andLovibond, P.F. (2009). The propositional natureof human associative learning. Behav. BrainSci. 32, 183–198, discussion 198–246.
13. Clark, R.E., and Squire, L.R. (1998). Classicalconditioning and brain systems: the role ofawareness. Science 280, 77–81.
14. Ohman, A., and Mineka, S. (2001). Fears,phobias, and preparedness: toward an evolvedmodule of fear and fear learning. Psychol. Rev.108, 483–522.
15. Rauch, S.L., Shin, L.M., and Phelps, E.A. (2006).Neurocircuitry models of posttraumatic stressdisorder and extinction: human neuroimagingresearch–past, present, and future. Biol. Psych.60, 376–382.
16. Milad, M.R., Rauch, S.L., Pitman, R.K., andQuirk, G.J. (2006). Fear extinction in rats:implications for human brain imaging andanxiety disorders. Biol. Psychol. 73, 61–71.
School of Psychology, The University ofNew South Wales, Sydney, Australia.E-mail: [email protected]
DOI: 10.1016/j.cub.2012.04.042
Nuclear Positioning: Dynein Neededfor Microtubule Shrinkage-CoupledMovement
Nuclear movement often requires interactions between the cell cortex andmicrotubules. A new study has revealed a novel protein interaction linkingmicrotubule plus-ends with the cortex and a role for dynein in microtubuleshrinkage-coupled movement.
Xin Xiang
Proper positioning of nuclei andmitoticspindles is crucial for the normalgrowth and development of many
eukaryotic organisms [1]. Unlike othercellular organelles that move alongmicrotubule tracks, nuclei/spindlesmove in response to either pushingor pulling force on the microtubules

DispatchR497
whose minus-ends are physicallylinked with the nuclei/spindles.Pushing or pulling force is oftengenerated through interactions ofgrowing (polymerizing) or shrinking(depolymerizing) microtubuleplus-ends with the cell cortex,but the mechanistic details ofthese interactions are not clear. Amicrotubule’s ‘end-on’ interactionwith the cortex can be non-productiveif it is simply followed by catastrophe(the switch from growth to shrinkage)and detachment from the cortex.Cortex-trigged catastrophe isa commonly observed phenomenonlikely caused by the ‘barrier effect’based on diminished on-rate ofGTP–tubulin subunits [2]. However,incorporation of GTP–tubulin subunitsto the plus ends at the cortex cangenerate pushing force for nuclearmovement [3–5]. It is not clearwhether additional linker proteins arerequired for the pushing mechanismin vivo. For generation of microtubuleshrinkage coupled pulling force, a linkbetween the shrinking plus-end andthe cortex is strictly required but itsidentity remains to be elucidated.Importantly, a new study by tenHoopen et al. [6], reported in thisissue of Current Biology, hasidentified a novel physical interactionbetween Bim1 (budding yeasthomolog of EB1, a microtubuleplus-end-tracking protein) at themicrotubule plus-end and Bud6 atthe cortex, which is required forgrowth/shrinkage-coupled nuclearmotility. Furthermore, this study hasrevealed a new in vivo roleof cytoplasmic dynein, a minus-end-directed microtubule motor, inmicrotubule shrinkage-couplednuclear movement.
Nuclear/spindle positioning inbudding yeast utilizes severalmechanisms operating at different cellcycle stages [4,5,7]. During G1/S, theold spindle pole body (SPB) emanatescytoplasmic microtubules that probethe cortex of the small bud. Bud6, anactin-interacting protein localized atthe bud cortex captures microtubulesto cause movements of the SPB andits attached nucleus [6]. Previously,a microtubule plus-end depolymerase,Kip3 (kinesin-8), was found to play animportant role in nuclear positioning[8]. Loss of Kip3 causes excessivemicrotubule growth at the cortex,which causes the nucleus to bepushed back to the mother cell [8].
Interestingly, the new study foundthat Bud6 is required for theexcessive microtubule growth inthe absence of Kip3, indicating thatthe plus-ends captured by Bud6 arepermitted to grow at the cortex togenerate pushing force. Themicrotubule plus-end-tracking proteinBim1 is also required for theBud6-dependent capturemechanism. Rather than beingindirectly involved by enhancinga microtubule’s searching ability, Bim1directly interacts with Bud6, and thisinteraction is crucial for the plus-end–cortex link required for SPBmovements. Importantly, microtubuleshrinkage-coupled SPB movementsdepend on dynein [6]. Dynein isknown to pull the anaphase spindleinto the bud neck [4,5] usinga ‘microtubule-sliding’ mechanismrather than a ‘capture/shrinkage’mechanism [4]. During anaphase, thedynein tail binds to its cortical anchor,Num1, and the motor heads walkalong an engaged astral microtubuletowards its minus-end, causing themicrotubule to slide along the budcortex [4,7]. Interestingly, Num1 is notrequired for SPB movement at G1/S,which uses the capture/shrinkagemechanism [6]. During SPB movementtoward the cortex, dynein isaccumulated at the shrinking end incontact with the cortex, and thisaccumulation appears to be crucial forshrinkage-coupled SPB movement [6].Thus, dynein seems to playa Num1-independent role in tetheringthe shrinking plus-end to pull theminus-end-attached SPB toward thecortex.
Remarkably, a direct role of dyneinin tethering the dynamic microtubuleplus-ends has been demonstrated bytwo recent studies in minimal in vitrosystems using either purified yeastdynein attached to microfabricatedbarriers [9] or purified bovine braindynein bound to optical trap-controlledbeads [10]. The requirement of ATPindicates that dynein’s ability to walk isimportant for its tethering function[9,10]. Most relevant to the new study[6] is that barrier-anchored yeastdynein is sufficient for generatingsignificant pulling force on thecaptured microtubule [9]. Dynein isunlikely to function as a plus-enddepolymerase [9]. However, the abilityof the anchored dynein to pull bywalking may cause the plus-end to hitthe barrier, and the particular
geometry of the interaction mayfacilitate catastrophe due to thebarrier effect [2,9]. In vivo, moreplayers are involved. The Bud6–Bim1interaction is required for the initialcapture of growing microtubules [6],and it remains to be dissected whetherit is continuously required aftershrinkage is initiated. The plus-enddepolymerase Kip3 contributes to thecapture/shrinkage mechanism byenhancing catastrophe [6,8]. Aplus-end depolymerase can in theorybe a major player in the capture/shrinkage mechanism if itsconnections with the cortex and theplus-end are maintained duringshrinkage. Indeed, Kar3 (kinesin-14),a minus-end-directed kinesin witha plus-end depolymerase activity [11],is critical for dynein-independentnuclear movement towards theyeast mating protrusion, and itsaccumulation persists at theplus-end–cortex junction duringshrinkage [12]. However, theaccumulation of Kip3 disappearedfrom the shrinking end contacting thecortex in the most recent study [6],and thus Kip3 is unlikely to continue itsdepolymerizing function, especiallygiven the cooperative mechanism ofKip3 action [13]. In contrast, dyneinaccumulation persists at theshrinking end contacting the cortex[6], and dynein seems to be a keyplayer in the capture/shrinkagemechanism [6]. However, the identityof dynein’s cortical anchor andthe mechanism of anchoring areunclear.As mentioned earlier, a link
between the shrinking end and thecortex is essential for pulling-forcegeneration. If a plus-end-trackingprotein is required as a structuralcomponent of the link, it mustinteract not only with the growing endbut also with the shrinking end.Although EB1 and its homologs inother systems only track the growingends, Bim1 tracks both the growingand shrinking ends [6,14]. Themechanism behind this phenomenonremains a total mystery. Dynein andCLIP-170 (a microtubule plus-endtracking protein) homologs in thefilamentous fungus Aspergillusnidulans and budding yeast are alsoable to track both growing andshrinking ends [15–19], and theaccumulation at the shrinking ends iscorrelated with the requirement ofplus-end-directed kinesins [17–19]. In

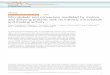
Figure 1. Two models of dynein–cortex interactions during microtubule shrinkage-couplednuclear movement.
The microtubule plus-end faces the cortex and the minus-end is linked to the nucleus. Corti-cally anchored dynein, with its minus-end-directed motor activity that pulls the microtubuletoward the cortex, enhances catastrophe due to the barrier effect [9]. Microtubule shrinkingvelocity is relatively low, and the shrinking end is not detached from dynein during shrinkage[9]. In the stable association model, a cortical anchor protein (black circle) associates with thesame dynein molecule at two different time points during shrinkage. In the dynamic associa-tion model, the cortical interaction with a dynein molecule (red) at the first time point is lost butquickly replaced by a new interaction with a different dynein molecule (brown) at the secondtime point. For simplicity, only one dynein molecule is depicted. In the cell, however, dyneinmolecules accumulate at the shrinking end, allowing multiple consecutive interactions tomaintain the link between the shrinking end and the cortex.
Current Biology Vol 22 No 12R498
the budding yeast and the dimorphicfungus Ustilago maydis, analyses offluorescence recovery afterphotobleaching of Bik1 (yeastCLIP-170 homolog) and dyneinrevealed that these proteins are highlydynamic at the plus ends, evenwhen the ends are stable [18,20]. It islikely that dynein’s shrinking-endaccumulation described in the newpaper [6] is also dynamic, with newmolecules constantly arriving toreplenish the population, as theprevious members are lost with theold end. Thus, two distinct modelscan be envisioned for dynein–cortexinteraction during plus-endshrinkage-coupled nuclear movement(Figure 1). The ‘stable association’model involves a stable associationbetween a dynein molecule and itscortical anchor, similar to the situationin the in vitro system [9]. For thismodel, while the growing-endaccumulation of dynein may facilitatedelivery of dynein to its corticalanchor [7,16], the shrinking-end
accumulation would seemunnecessary. In the ‘dynamicassociation’ model, old interactionsare constantly being broken andreplaced by new interactions betweenthe cortex and newly arrived dyneinmolecules at the new end. Theaccumulation of dynein at the shrinkingend would allow multiple interactionsthat are unlikely to be broken alltogether at any particular time point,and such a collective effort of theplus-end molecules benefits corticaltethering of the shrinking end. Inaddition, the Bud6–Bim1 interactioncould possibly serve as an additionaltether, which may also involvedynamic interactions of multiple Bim1molecules at the shrinking end withmultiple Bud6 molecules at theBud6-marked cortical site. Finally,one interesting possibility notexcluded is that Bim1 indirectlyinteracts with the dynein tail viaseveral other proteins at the shrinkingend, thereby linking dynein to Bud6 atthe cortex.
References1. Morris, N.R. (2000). Nuclear migration. From
fungi to the mammalian brain. J. Cell Biol. 148,1097–1101.
2. Janson, M.E., de Dood, M.E., and Dogterom, M.(2003). Dynamic instability of microtubulesis regulated by force. J. Cell Biol. 161,1029–1034.
3. Tran, P.T., Marsh, L., Doye, V., Inoue, S., andChang, F. (2001). A mechanism for nuclearpositioning in fission yeast based onmicrotubule pushing. J. Cell Biol. 153, 397–411.
4. Adames, N.R., and Cooper, J.A. (2000).Microtubule interactions with the cell cortexcausing nuclear movements in Saccharomycescerevisiae. J. Cell Biol. 149, 863–874.
5. Yeh, E., Yang, C., Chin, E., Maddox, P.,Salmon, E.D., Lew, D.J., and Bloom, K. (2000).Dynamic positioning of mitotic spindlesin yeast: role of microtubule motors andcortical determinants. Mol. Biol. Cell 11,3949–3961.
6. ten Hoopen, R., Cepeda-Garcı́a, C.,Fernandez-Arruti, R., Juanes, M.A.,Delgehyr, N., and Segal, M. (2012). Mechanismfor astral microtubule capture by corticalBud6p priming spindle polarity in S. cerevisiae.Curr. Biol. 22, 1075–1083.
7. Markus, S.M., and Lee, W.L. (2011).Microtubule-dependent path to the cellcortex for cytoplasmic dynein in mitoticspindle orientation. BioArchitecture 1, 1–7.
8. Gupta, M.L., Jr., Carvalho, P., Roof, D.M., andPellman, D. (2006). Plus end-specificdepolymerase activity of Kip3, a kinesin-8protein, explains its role in positioning theyeast mitotic spindle. Nat. Cell Biol. 8,913–923.
9. Laan, L., Pavin, N., Husson, J., Romet-Lemonne, G., van Duijn, M., Lopez, M.P.,Vale, R.D., Julicher, F., Reck-Peterson, S.L.,and Dogterom, M. (2012). Cortical dyneincontrols microtubule dynamics to generatepulling forces that position microtubule asters.Cell 148, 502–514.
10. Hendricks, A.G., Lazarus, J.E., Perlson, E.,Gardner, M.K., Odde, D.J., Goldman, Y.E., andHolzbaur, E.L. (2012). Dynein tethers andstabilizes dynamic microtubule plus ends. Curr.Biol. 22, 632–637.
11. Sproul, L.R., Anderson, D.J., Mackey, A.T.,Saunders, W.S., and Gilbert, S.P. (2005). Cik1targets the minus-end kinesin depolymerasekar3 to microtubule plus ends. Curr. Biol. 15,1420–1427.
12. Maddox, P.S., Stemple, J.K., Satterwhite, L.,Salmon, E.D., and Bloom, K. (2003). The minusend-directed motor Kar3 is required forcoupling dynamic microtubule plus ends to thecortical shmoo tip in budding yeast. Curr. Biol.13, 1423–1428.
13. Varga, V., Leduc, C., Bormuth, V., Diez, S., andHoward, J. (2009). Kinesin-8 motors actcooperatively to mediate length-dependentmicrotubule depolymerization. Cell 138,1174–1183.
14. Wolyniak, M.J., Blake-Hodek, K., Kosco, K.,Hwang, E., You, L., and Huffaker, T.C. (2006).The regulation of microtubule dynamics inSaccharomyces cerevisiae by three interactingplus-end tracking proteins. Mol. Biol. Cell 17,2789–2798.
15. Han, G., Liu, B., Zhang, J., Zuo, W.,Morris, N.R., and Xiang, X. (2001). TheAspergillus cytoplasmic dynein heavy chainand NUDF localize to microtubule ends andaffect microtubule dynamics. Curr. Biol. 11,719–724.
16. Sheeman, B., Carvalho, P., Sagot, I., Geiser, J.,Kho, D., Hoyt, M.A., and Pellman, D. (2003).Determinants of S. cerevisiae dyneinlocalization and activation: implications for themechanism of spindle positioning. Curr. Biol.13, 364–372.
17. Zhang, J., Li, S., Fischer, R., and Xiang, X.(2003). Accumulation of cytoplasmic dyneinand dynactin at microtubule plus ends inAspergillus nidulans is kinesin dependent. Mol.Biol. Cell 14, 1479–1488.

DispatchR499
18. Carvalho, P., Gupta, M.L., Jr., Hoyt, M.A., andPellman, D. (2004). Cell cycle control ofkinesin-mediated transport of Bik1 (CLIP-170)regulates microtubule stability and dyneinactivation. Dev. Cell 6, 815–829.
19. Efimov, V.P., Zhang, J., and Xiang, X. (2006).CLIP-170 homologue and NUDE playoverlapping roles in NUDF localization in
Aspergillus nidulans. Mol. Biol. Cell 17,2021–2034.
20. Schuster, M., Kilaru, S., Ashwin, P., Lin, C.,Severs, N.J., and Steinberg, G. (2011).Controlled and stochastic retentionconcentrates dynein at microtubule ends tokeep endosomes on track. EMBO J. 30,652–664.
Department of Biochemistry and MolecularBiology, Uniformed Services University of theHealth Sciences, Bethesda, MD 20814, USA.E-mail: [email protected]
DOI: 10.1016/j.cub.2012.04.044
Single-Molecule Imaging:A Collagenase Pauses beforeEmbarking on a Killing Spree
Single-molecule tracking provides new insights into how an ATP-independentendo-proteolytic machine digests collagen fibrils during their remodeling.
Gwangrog Lee1 and Taekjip Ha2,*
The extracellular matrix isa well-organized macromolecularplatform that specifies the mechanicalproperties of connective tissues tomaintain the cell shapes. Collagen isa major element of the extracellularmatrix and is themost abundant proteinin human tissues. Somewhat like the artof knitting, collagen is weaved intoprotein strings to form collagen fibrils,which then form a lattice (Figure 1A),which are highly resistant toproteolytic degradation. Over time,however, this highly stable scaffoldmust undergo remodeling duringpathophysiological processes, suchas wound healing, tumor progression,metastatic invasion, and hostdefense mechanisms [1]. Matrixmetalloproteases (MMPs) are theendopeptidases in charge of degradingcollagen fibrils, hence called‘collagenases’, and their activitiesmustbe tightly regulated. Although it is nowknown that other types of processiveproteases, e.g. ClpXP, use chemicalenergy derived from ATP hydrolysis tomechanically unfold protein structuresbefore digestion [2,3], it has been apuzzle how MMPs can help remodelstableorganizationsof collagenwithoutusing additional energy sources.
Studying native collagen fibrils isdifficult using traditional enzymologytools because the extended substrateis insoluble and heterogeneous. In2004, a new approach of fluorescencecorrelation spectroscopy thatexamines molecular diffusion ona sub-micron scale was applied tothe study of an MMP subtype, MMP1,
and led to the proposal of a Brownianratchetmodel;MMP1diffuses on type 1collagen but its Brownian motion isbiased through a ‘burnt bridge’ effectcaused by collagen proteolysis [4]. But,because of the difficulty in handingnative collagen samples and thetechnical limitations of averaging overmanymolecules, the earlier study couldnot address how MMPs initiate andcarry out the degradation of the nativesubstrate. Now, in this issue of CurrentBiology, Sarkar et al. [5] report the useof single-molecule fluorescenceimaging to shed new light on theseissues and provide a major leap in ourunderstanding of the multiple phasesof native collagen degradation.Fluorescently labeled MMP1 proteinswere added to native collagen fibrilsimmobilized on the sample cell surfaceand the motion of single MMP1molecules on the fibrils was monitoredin real time through total internalreflection fluorescence microscopy.
As anticipated, the authors foundthat MMP1 diffuses on the collagenfibrils. But direct imaging allowedthem to show that the motion isone-dimensional (1D), occurring alongthe collagen fibril, but not across fibrils,raising the possibility that MMP1uses 1D diffusional search to findthe cleavage sites on the 3D collagenlattice. Interestingly, MMP1’s 1Ddiffusion was not continuous but waspunctuated by pauses. In fact, MMP1spent w90% of the time in pausedstates with only w10% of time spenttransiting between different pausingsites. As a result, these pausesdominate the overall diffusiontimescales. One class of pauses
followed a single exponentialdistribution of their lifetimes andoccupied no special positions on thefibrils. The second class of pauseswas longer in duration and had adistinct lag phase before escaping thepaused state. Furthermore, statisticalanalysis showed that multiplesequential steps are necessary beforethe escape (Figure 1B). The molecularorigin of these class II pauses thatexhibit the lag phase is as yet unknownbut these pauses are reminiscent ofthe activity of nucleic acid enzymesthat can accumulate elastic energythrough in multiple irreversiblereactions before transitioning to asubsequent phase [6–8]. Furthermore,these class II pauses occur at periodiclocations (see below).As the enzyme escapes the class II
pause site, it shows a so-called‘ballistic’ behavior with a distinct biasin its initial motion along one fibrildirection. This biased random walkwas not observed with an active sitemutant of MMP1, suggesting that thedirectional bias is related to theendopeptidase activity of MMP1.Furthermore, the ballistic behaviorwas observed at 37�C but not at 25�C.These observations led to an intriguingpossibility that thermally induced localunfolding of collagen may allow MMP1to initiate the collagenolysis. Uponinitiation, cleavage reaction would biasthe diffusion by burning the bridgebehind so that subsequent diffusionappears ballistic along the collagenfibril. To obtain quantitative detailsof the collagenolysis the authorsperformed modeling and simulations.They found that only 5% of class IIpauses result in the actual initiationof cleavage but this killing rampageis highly processive and, on average,15 consecutive cleavage events resultfrom one initiation event. MMP1spends w90% of its time at pausingstates due to the inaccessibility ofthe cleavage sites, but once the firstcleavage occurs, subsequentcleavages progress rapidly as the