Embed Size (px)
Citation preview

American Journal of Medical Genetics 117A:255–260 (2003)
Novel Perforin Mutation in a PatientWith Hemophagocytic Lymphohistiocytosisand CD45 Abnormal Splicing
James McCormick,1 Darren R. Flower,1 Stephan Strobel,2 Diana L. Wallace,1 Peter C.L. Beverley,1
and Elma Z. Tchilian1*1The Edward Jenner Institute for Vaccine Research, Compton, Berkshire, United Kingdom2Immunobiology Unit, Institute for Child Health, London, United Kingdom
Hemophagocytic lymphohistiocytosis (HLH)composes a group of rare heterogenous dis-orders characterized by uncontrolled ac-cumulation and infiltration of activated Tlymphocytes and macrophages. Cytotoxic Tand natural killer cell activity is signifi-cantly reduced or absent in these patients.Mutations in the important mediator of lym-phocyte cytotoxicity perforin were identi-fied in a number of HLH individuals. Here wereport a novel missense mutation thr435metin the conserved Ca2þ binding domain ofperforin in a patient with HLH. Prediction ofthe 3-dimensional structure of the thr435metperforin mutant using comparative molecu-lar modeling indicates that the protein’s abi-lity to bind Ca2þ, and therefore its cytolyticfunction, would be strongly compromised. Inaddition, this patient exhibited abnormalCD45 splicing caused by a C77G mutation inthe gene encoding CD45 (PTPRC). Our find-ings suggest a combined role for perforinmutation and abnormal CD45 splicing assignificant contributory factors in the patho-genesis of HLH. � 2003 Wiley-Liss, Inc.
KEY WORDS: perforin; HLH; CD45 spli-cing; PTPRC
INTRODUCTION
Hemophagocytic lymphohistiocytosis (HLH) is a raredisorder characterized by disregulated activation of T
lymphocytes andmacrophages [Arico et al., 2001]. HLHis genetically heterogenous with both familial andsporadic forms described [Janka, 1983; Dreyer et al.,1991; Dufourcq-Lagelouse et al., 1999]. The geneticdefects underlying HLH have only been partiallydefined. Linkage of a disease gene to 9q21.3-22 (HLHtype 1) was identified in four inbred Pakistani familieswith HLH using homozygosity mapping [Ohadi et al.,1999]. Shortly after the identification of a linkage to10q21-22, mutations in the perforin 1 gene (PRF1) wereidentified [Stepp et al., 1999]. Recent reports haveexpanded the pattern of PRF1 mutations associatedwith HLH [Stepp et al., 1999; Ericson et al., 2001,Clementi et al., 2001, Kogawa et al., 2002]. Perforin issecreted by activated cytotoxic T cells and NK cells,inducing cell death by pore formation in target cells[Metkar et al., 2002]. However, perforin mutations ac-count for a still undefined proportion of HLH caseswhich could be in the range of 20–40% [Ericson et al.,2001], while the majority of HLH cases are likely to becaused by defects in as yet unidentified genes with orwithout a environmental contribution.
A patient with hemophagocytic lymphohistiocytosiswith a defect in CD45 splicing has been previouslydescribed [Wagner et al., 1995]. CD45 is an abundanttyrosine phosphatase expressed on all leucocytes[Penninger et al., 2001] and recently abnormal CD45splicing, caused by a C77G polymorphism in exon Aof the gene encoding CD45 (PTPRC ) was associatedwith the development of multiple sclerosis and HIV-1infection in humans [Jacobsen et al., 2000; Tchilianet al., 2001a]. Here we analyzed the perforin statusin this patient and found a novel mis-sense perforinmutation thr435met in exon 3. The mutated threonine435 is conserved between human, mouse and rat andis located in the highly conserved Ca2þ binding domainof perforin. Molecular modeling predicts that thethr435met mutation would affect the Ca2þ bindingof themolecule, and therefore, impair perforin function.These findings suggest that abnormalities in CD45splicing and perforin mutations might be associatedwith HLH and these associations warrant furtherstudies.
*Correspondence to: Elma Z. Tchilian, Edward Jenner Institutefor Vaccine Research, Compton, Berkshire RG20 7NN, UK.E-mail: [email protected]
Received 19 March 2002; Accepted 12 August 2002
DOI 10.1002/ajmg.a.10010
� 2003 Wiley-Liss, Inc.

MATERIALS AND METHODS
Materials
Fresh blood was obtained from the previously des-cribed family W. [Wagner et al., 1995] from the Im-munobiology Unit, Institute for Child Health, London,UK. PBMC were isolated by centrifugation on a Ficoll-Paque (Amersham Pharmacia Biotech, Buckingham-shire, UK) density gradient and genomic DNA wasextracted by standard procedures [Sambrook et al.,1989].
Mutation Analysis of the Perforin Gene
The perforin gene was amplified by PCR from geno-mic DNA as previously described [Stepp et al., 1999;Ericson et al., 2001]. Briefly, exon 2 was amplified usingPF1 forward (50-CCCTTCCATGTGCCCTGATAATC-30)and PF2 reverse (50-AGGAGCCTCCAAGTTTGAG-30)primers. Exon 3 was amplified using PF3 forward(50-ACTGCTCCCGGCCAGGATCATTG-30) and PF8 re-verse (50-GGCTCCCACTGTGAGA-30) primers. Eachreaction mixture contained 0.5 ml of both forward andreverse primers (20 pmol), 0.5 ml dNTP (10mM), 5 ml10�PCR buffer (PE Applied Biosystems, Cheshire,UK), 4 ml MgCl2 (25 mM), and 2.5 U of AmpliTaq Gold(PE Applied Biosystems). The PCR parameters usedwere denaturation at 968C for 15 min followed by35 cycles (968C for 30 sec, 588C for 20 sec and 728C for30 sec) and a final extension at 728C for 2 min. Thepurified PCR products were subjected to direct auto-mated sequencing using an ABI Prism 377 (PE AppliedBiosystems). The sequences obtained were compared tothe published human sequence (PRF1 Genbank acces-sion number M28393).
Sequence analysis of the perforin gene in family W.showed the presence of a previously described silentheterozygous C to T polymorphism at position 900(his300his) [Ericson et al., 2001] and a novel hetero-zygous C to T transition at position 1304 (C1304T) re-sulting in a change of threonine 435 to methionine(thr435met). The presence of the C1304T mutation wasfurther confirmed by restriction digestion since thistransition changes the sequence from CCACGG toCCATGG which can be detected by digestion with NcoI(Gibco Life Technologies, Paisley, UK). Primers PF6forward (50-GTCACCACCCAGGACTGCT-30) and PF8reverse (see above), flanking the C1304Tmutation wereused to generate a 526 bp product and the mutant allelewas detected by the presence of two fragments of 425 bpand 101 bp after digestion with NcoI.
Molecular Modeling
Sequenceanalysis shows that theC-terminal regionofperforin contains a Ca2þ binding C2 domain [Schultzet al., 2000]. The structures of a number of C2 domainshave now been experimentally determined [Rizo andSudhof, 1998] and are comprised of a compact b-sand-wich fold composed of two orthogonally aligned four-stranded b-sheets. The structures of C2 domains fall intoone of two classes—the S and P variants—that are
related by a circular permutation of their folding to-pology. The three-dimensional structures of four repre-sentatives of each of the two variant classes wereextracted from the protein databank [Berman et al.,2000]: 3RPB, 1RSY, 1DSY, and 1DQV for variant S and1E8X, 1RLW, 1D5R, and 1DJX for variant P. TheMALIGN3D function of theMODELLER suite [Sali andBlundell, 1993]wasused to align the four representativethree-dimensional structures for each of the two va-riants. The sequence of the perforin C2 domainwas thenmanually aligned to the resultant multiple alignmentsusing CINEMA [Parry-Smith et al., 1998]. Because ofsignificantly closer sequence similarity in the region oftheCa2þbinding site betweenperforin and theS variantmultiple alignment, the S type C2 domain was used as atemplate for modeling the three dimensional structureof the C2 domain of perforin usingMODELLER. For thesake of simplicity, it is this model which is describedbelow, although similar results were obtained with theP variant model. The model was briefly optimized in asolvent bath using the molecular mechanics programAMBER [Cornell et al., 1995]. The resulting structurewas then analyzed by visual inspection using the mole-cular graphics program SYBYL.
RESULTS
Analysis of Perforin Status in Patient W.
We obtained material from a patient W. with HLH,previously described as exhibitingCD45abnormal splic-ing as characterized by the lack of the single CD45ROþT cell population [Wagner et al., 1995]. No informationwas available on the perforin status of this patient.Patient W. was the third child of healthy unrelatedBritish Caucasian parents [Wagner et al., 1995]. Hepresented at age 3 months with fever, diarrhea, pallor,increasing irritability and marked cervical lympho-adenopathy and hepatosplenomegaly. Laboratory in-vestigations revealed pancytopenia, coagulopathy, andhypertryglyceridemia. The diagnosis of HLH was madefrom the bone marrow aspirate, which showed hemo-phagocytosis. There was a good response to initialtreatment with dexamethasone and etoposide and heunderwent allogeneic bone marrow transplantationfrom his HLA identical brother.
The perforin gene (PRF1) is located on chromosome10(10q21-22) and is organized into three exons, the firstof which is not translated [Lichtenheld and Podack,1989]. We analyzed exons 2 and 3 of the perforin gene inpatient W. and found two mutations in exon 3. The firstis a previously reported silent heterozygous C to Tpolymorphismat position 900 (his300his) [Ericson et al.,2001]. The second is a novel mis-sense heterozygouspoint mutation at nucleotide position 1304 (C1304T)resulting in a change of threonine 435 to methionine(thr435met). The presence of C1304T was analyzed inall family members by sequencing. Furthermore, thismutation introduces a new restriction site for NcoIwhich was used to confirm the genotype of the familymembers (Fig. 1). Restriction analysis revealed that thepatient, his mother and two siblings are heterozygousfor the mutant C1304T (thr435met) allele (Lanes 2 and
256 McCormick et al.

4–6, Fig. 1) while the father and the oldest brother werehomozygous for the normal perforin allele (Lanes 1 and3, Fig. 1). The other perforin polymorphism C900T(his300his) showedan identical pattern of inheritance totheC1304T (thr435met)mutation in the familyW. (datanot shown).
Thr435met Disrupts the Ca2þ BindingActivity of Perforin
The perforin gene (PRF1) encodes a 534 amino acidpolypeptide that is constitutively expressed by naturalkiller cells [Yu et al., 1999], while cytotoxic T-cells ex-press the protein only when stimulated by effector-target cell interactions. This interaction induces thecytotoxic effector cell to release cytoplasmic granulesinto the intercellular space. In the presence of Ca2þ
perforin inserts into the cell membrane and then poly-merises to form poly-perforin pores, each consisting of12-18monomeric units. Pore formation leads to destruc-tion of cells by osmotic lysis and by allowing the entry ofapoptosis inducing granzymes [Tschopp and Nabholz,1990; Lowin et al., 1995; Darmon et al., 1995].
Perforin is composed of two functional domains. Thefirst is a region of about 330 amino acids which is highlyhomologous to regions of the C6, C7, C8 and C9 com-ponents of the complement cascade. This representsthe membrane channel-forming region also seen in themembrane attack complex of complement. The second
smaller Ca2þ binding domain (C2 domain) is a proteinmodule widely used in nature. In perforin, calciumbinding by the C2 domain triggers a profound con-formational change in the overall structure of the C9-homologous region of the molecule. This probably ex-poses an otherwise hidden hydrophobic region of theoriginally hydrophilic protein, leading to aggregationand the formation of cytolytic pores.
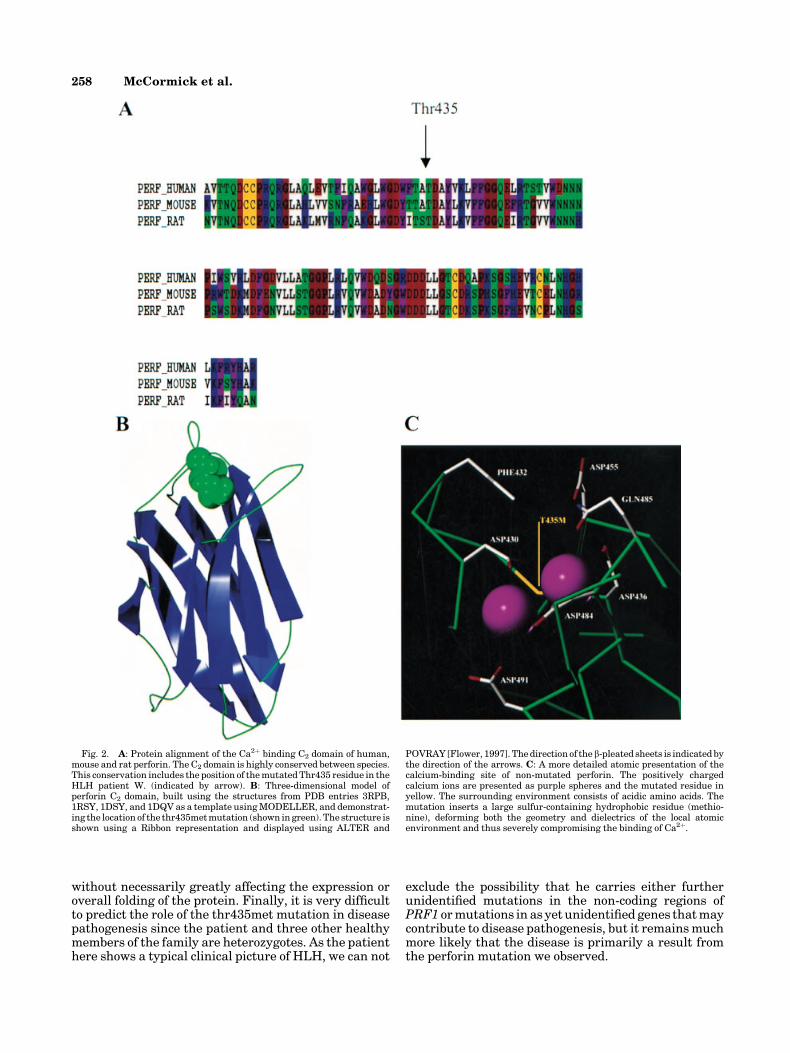
The novel thr435metmutation identified in familyW.is part of the highly conserved C2 domain of perforin.Threonine 435 is conserved between human, mouse andrat (Fig. 2A). Molecular modeling suggests that thethr435met mutation is located within the Ca2þ bindingsite of the C2 domain (Fig. 2B,C). The increased hydro-phobicity and altered polarizability of the methionineresidue is likely to affect severely the carefully con-trolled dielectric environment of this site, which is cru-cial to the tight binding of calcium ions. Moreover, theincreased steric bulk ofmethionine is also likely to causesignificant disruption to the arrangement of amino acidside-chains in this region. The carefully arranged geo-metry of an array of negatively charged acidic aminoacid side chains is an absolute requirement to properlyco-ordinate the positively charged calcium ions withinthe binding site. Both effects of the thr435met mutationwill impinge directly on the ability of the protein to bindCa2þ ions and, through the conformational change thatthis triggers within the protein, will severely compro-mise its cytolytic activity.
DISCUSSION
HLH is a rapidly fatal disease if left untreated. It ischaracterized by the infiltration of activated lympho-cytes and histiocytes (macrophages) into the spleen,liver, lymph nodes, bone marrow and the central ner-vous system. Mutations in perforin were recently iden-tified in anumber ofHLH individuals [Stepp et al., 1999;Ericson et al., 2001, Clementi et al., 2001; Kogawa et al.,2002]. In this study, we have analyzed the perforinstatus in a patient with HLH [Wagner et al., 1995]and identified a novel perforin mis-sense mutationthr435met in exon 3. Threonine 435 is conserved bet-ween human, mouse and rat. Other mis-sense hetero-zygous perforin mutations have been previouslyreported, although not all of them involve conservedamino acids [Stepp et al., 1999; Ericson et al., 2001].Patients harboring thesemutations have been shown todisplay substantially reduced levels of expression ofperforin protein as well as reduced cytolytic activity[Stepp et al., 1999; Kogawa et al., 2002]. This is the firstdescription of thr435met mutation and further studiesof the expression and function of this mis-sense muta-tion will be required to determine how it may affectperforin function. Because of the lack of further clinicalmaterial, we were prevented from undertaking thesestudies. The mutation is, however, located in the highlyconserved Ca2þ binding domain of the perforinmoleculeandmolecularmodeling suggests that thr435met wouldaffect the ability of perforin to undergo conformationalchange andmembrane insertion in the presence ofCa2þ.Such a drasticmutationwould impair perforin function,
Fig. 1. Identification of perforin C1304T transition in family W. A:Family tree indicating the perforin phenotype in each member of family W.The patient with HLH (5) is indicated by asterisks. B: Primers flankingthe mutation were used to amplify a 526 bp fragment. The C to Ttransition introduces an NcoI restriction site which cleaves the mutantPCR product into two fragments of 425 and 101 bp. The patient (lane 5),his mother and two siblings (lanes 2, 4, and 6) are heterozygous for theC1304T mutant perforin allele, while the father and older brother (lanes 1and 3) have only the 526 bp undigested wild type allele. The perforinmutation co-segregated in the family with the C77G mutation in the geneencoding CD45.
A Novel Perforin Mutation in HLH 257
without necessarily greatly affecting the expression oroverall folding of the protein. Finally, it is very difficultto predict the role of the thr435met mutation in diseasepathogenesis since the patient and three other healthymembers of the family are heterozygotes. As the patienthere shows a typical clinical picture of HLH, we can not
exclude the possibility that he carries either furtherunidentified mutations in the non-coding regions ofPRF1 ormutations in as yet unidentified genes thatmaycontribute to disease pathogenesis, but it remainsmuchmore likely that the disease is primarily a result fromthe perforin mutation we observed.
Fig. 2. A: Protein alignment of the Ca2þ binding C2 domain of human,mouse and rat perforin. The C2 domain is highly conserved between species.This conservation includes the position of themutatedThr435 residue in theHLH patient W. (indicated by arrow). B: Three-dimensional model ofperforin C2 domain, built using the structures from PDB entries 3RPB,1RSY, 1DSY, and 1DQV as a template usingMODELLER, and demonstrat-ing the location of the thr435metmutation (shown in green). The structure isshown using a Ribbon representation and displayed using ALTER and
POVRAY [Flower, 1997]. Thedirection of the b-pleated sheets is indicated bythe direction of the arrows. C: A more detailed atomic presentation of thecalcium-binding site of non-mutated perforin. The positively chargedcalcium ions are presented as purple spheres and the mutated residue inyellow. The surrounding environment consists of acidic amino acids. Themutation inserts a large sulfur-containing hydrophobic residue (methio-nine), deforming both the geometry and dielectrics of the local atomicenvironment and thus severely compromising the binding of Ca2þ.
258 McCormick et al.

In this respect, it is interesting that the patientexhibits abnormal CD45 splicing [Wagner et al., 1995].We confirmed that the C77G polymorphism in exon A ofthe gene encoding CD45 is the cause for the observedvariant splicing in patient W., which is in accordancewith our previous studies in which C77G was identi-fied as the common cause of abnormal CD45 splicing[Tchilian et al., 2001b]. Interestingly, the perforinthr435met and the CD45 exon A (C77G) mutations co-segregate together in family W., so that the patienttogether with the mother and two healthy sibs exhibitboth perforin thr435met andCD45C77Gmutations.Wepostulate that the C77G mutation is a factor provokingthe disease along with the perforin mutation. This isplausible since studies with transgenic mice suggestthat expressionof ahighmolecularweightCD45 isoformalone causes immunodeficiency and these mice cannotgenerate any cytotoxic T cell responses or neutralisingantibodies after viral infection [Kozieradzki et al.,1997]. Sporadic HLH cases are often provoked by viralinfection later in childhood [Dreyer et al., 1991] and onemight speculate that individuals with abnormal CD45splicingmay have a higher risk of viral infection. This isfurther supported by our recent findings that abnormalCD45 splicing and C77G polymorphism is associatedwith HIV-1 infection [Tchilian et al., 2001a] suggestingthat individuals with abnormal CD45 splicingmay haveincreased susceptibility to viral infection or impairedanti-viral responses particularly in infancy.
Our data has shown thatPRF1mutations can occur inthe Ca2þ binding domain encoded by exon 3 and, in theabsence of the necessary but unavailable material re-quired for further study, molecular modeling provides apowerful molecular rationale for its impaired function.Although mutations in the genes encoding perforin andCD45 are associated with HLH, the primary cause maybe due to mutations in one or more as yet unidentifiedgenes or, as seemsmore likely, thedisease is the result ofa number of gene mutations, including those identifiedhere, interacting to produce the disease phenotype. Inany case, the identification of these genetic defects mayhave important diagnostic, prognostic, and therapeuticimplications in the screening and future treatment ofHLH.
ACKNOWLEDGMENT
Wethank thepatient family, clinicians, andnurses forhelp with patient samples.
REFERENCES
Arico M, Imashuku S, Clementi R, Hibi S, Teramura T, Danesino C, HaberDA, Nichols KE. 2001. Hemophagocytic lymphohistiocytosis due togermline mutations in SH2D1A, the X-linked lymphoproliferativedisease gene. Blood 97:1131–1133.
Berman HM, Westbrook J, Feng Z, Gilliland G, Bhat TN, Weissig H,Shindyalov XX, Bourne PE. 2000. In the protein data bank. NucleicAcids Res 28:235–242.
Clementi R, zur Stadt U, Savoldi G, Varoitto S, Conter V, De FuscoC, Notarangelo LD, Schneider M, Klersy C, Janka G, Danesino C,Arico M. 2001. Six novel mutations in the PRF1 gene in childrenwith hemophagocytic lymphohistiocytosis. J Med Genet 38:643–646.
Cornell WD, Cieplak P, Bayly CI, Gould IR, Merz KM Jr, Ferguson DM,Spellmeyer DC, Fox T, Caldwell JW, Kollman PA. 1995. A secondgeneration force field for the simulation of proteins and nucleic acids.J Am Chem Soc 117:5179–5197.
Darmon AJ, Nicholson DW, Bleackley RC. 1995. Activation of the apoptoticprotease CPP32 by cytotoxic T-cell-derived granzyme B. Nature377:446–448.
Dreyer ZE, Dowell BL, Chen H, Hawkins E, McClain KL. 1991.Infection-associated hemophagocytic syndrome. Evidence for Epstein-Barr virus gene expression. Am J Pediatr Hematol Oncol 13:476–481.
Dufourcq-Lagelouse R, Jabado N, Le Deist F, Stephan JL, Souillet G,BruinM, Vilmer E, Schneider M, Janka G, Fischer A, de Saint Basile G.1999. Linkage of familial hemophagocytic lymphohistiocytosis to10q21-22 and evidence for heterogeneity. Am J Hum Genet 64:172–179.
EricsonG,FadeelB,Nilsson-ArdnorS, SoderhallC,SamuelssonA, JankaG,SchneiderM, Gurgey A, YalmanN, Revesz T, Egeler R, Jahnukainen K,Storm-Mathiesen I, Haraldsson A, Poole J, de Saint Basile G,Nordenskjold M, Henter J. 2001. Spectrum of perforin gene mutationsin familial hemophagocytic lymphohistiocytosis. Am J Hum Genet68:590–597.
Flower DR. 1997. ALTER: Eclectic management of molecular structuredata. J Mol Graph Model 15:161–169.
Jacobsen M, Schweer D, Ziegler A, Gaber R, Schock S, Schwinzer R,Wonigeit K, Lindert RB, Kantarci O, Schaefer-Klein J, Schipper HI,Oertel WH, Heidenreich F, Weinshenker BG, Sommer N, Hemmer B.2000. A point mutation in PTPRC is associated with the development ofmultiple sclerosis. Nat Genet 26:495–499.
Janka GE. 1983. Familial hemophagocytic lymphohistiocytosis. Eur JPediatr 140:221–230.
KogawaK,LeeSM,Villanueva J,MarmerD, Sumegi J, FilipovichAH. 2002.Perforin expression in cytotoxic lymphocytes from patients withhemophagocytic lymphohistiocytosis and their family members. Blood99:61–66.
Kozieradzki I, Kundig T, Kishihara K, Ong CJ, Chiu D, Wallace VA,Kawai K, Timms E, Ionescu J, Ohashi P, Marth JD, Mak TW,Penninger JM. 1997. T cell development in mice expressing splicevariants of the protein tyrosine phosphatase CD45. J Immunol 158:3130–3139.
LichtenheldMG, Podack ER. 1989. Structure of the human perforin gene. Asimple gene organization with interesting potential regulatorysequences. J Immunol 143:4267–4274.
Lowin B, Peitsch MC, Tschopp J. 1995. Perforin and granzymes:Crucial effector molecules in cytolytic T lymphocyte and naturalkiller cell-mediated cytotoxicity. Curr Top Microbiol Immunol 198:1–24.
Metkar SS, Wang B, Aguilar-Santelises M, Raja SM, Uhlin-Hansen L,Podack E, Trapani JA, Froelich CJ. 2002. Cytotoxic cell granule-mediated apoptosis: Perforin delivers granzyme B-serglycin complexesinto target cells without plasma membrane pore formation. Immunity16:417–428.
Ohadi M, Lalloz MR, Sham P, Zhao J, Dearlove AM, Shiach C, Kinsey S,Rhodes M, Layton DM. 1999. Localization of a gene for familialhemophagocytic lymphohistiocytosis at chromosome 9q21.3-22 byhomozygosity mapping. Am J Hum Genet 64:165–171.
Parry-Smith DJ, Payne AW, Michie AD, Attwood TK. 1998. CINEMA–Anovel colour interactive editor formultiple alignments.Gene221:GC57–GC63.
Penninger JM, Irie-Sasaki J, Sasaki T,Oliveira-Dos-SantosAJ. 2001.CD45:New jobs for an old acquaintance. Nat Immunol 2:389–396.
Rizo J, Sudhof TC. 1998. C2-domains, structure, and function of a universalCa2þ-binding domain. J Biol Chem 273:15879–15882.
Sali A, Blundell TL. 1993. Comparative protein modelling by satisfaction ofspatial restraints. J Mol Biol 234:779–815.
Sambrook J, Fritsch EF, Maniatis T. 1989. Molecular cloning: A labora-tory manual. Cold Spring Harbour: Cold Spring Harbour LaboratoryPress.
Schultz J, Copley RR, Doerks T, Ponting CP, Bork P. 2000. SMART: A web-based tool for the study of genetically mobile domains. Nucleic Acids Res28:231–234.
Stepp SE, Dufourcq-Lagelouse R, Le Deist F, Bhawan S, Certain S,Mathew PA, Henter JI, Bennett M, Fischer A, de Saint Basile G,
A Novel Perforin Mutation in HLH 259

Kumar V. 1999. Perforin gene defects in familial hemophagocyticlymphohistiocytosis. Science 286:1957–1959.
Tchilian EZ, Wallace DL, Dawes R, Imami N, Burton C, Gotch F, BeverleyPC. 2001a. A point mutation in CD45 may be associated with HIV-1infection. AIDS 15:1892–1894.
Tchilian EZ, Wallace DL, Imami N, Liao HX, Burton C, Gotch F, MartinsonJ, Haynes BF, Beverley PC. 2001b. The exon A (C77G) mutation is acommon cause of abnormal CD45 splicing in humans. J Immunol166:6144–6148.
Tschopp J, Nabholz M. 1990. Perforin-mediated target cell lysis by cytolyticT lymphocytes. Annu Rev Immunol 8:279–302.
Wagner R, Morgan G, Strobel S. 1995. A prospective study of CD45isoform expression in hemophagocytic lymphohistiocytosis: An abnor-mal inherited immunophenotype in one family. Clin Exp Immunol 99:216–220.
Yu CR, Ortaldo JR, Curiel RE, Young HA, Anderson SK, Gosselin P. 1999.Role of a STAT binding site in the regulation of the human perforinpromoter. J Immunol 162:2785–2790.
260 McCormick et al.





























