Embed Size (px)
Citation preview

Reviews�POSTSCREEN
Drug Discovery Today � Volume 19, Number 8 �August 2014 REVIEWS
Novel immunological targets inrheumatic diseases: clues from currenttherapies
Fulvio D’Acquisto, Lorenza Rattazzi,Giuseppa Piras and Maria Letteria Galuppo
William Harvey Research Institute, Barts and the London School of Medicine, Queen Mary University of London, Charterhouse Square, London EC1 M 6BQ, UK
Many years have elapsed since the discovery of immunomodulators as effective therapeutics for the
treatment of rheumatic diseases, and we are still learning about their various mechanisms of action.
Here, we provide a concise overview of the most recent discoveries in this field of research, focusing in
particular on signaling pathways targeted by therapeutics currently used in the clinic. We highlight
areas of investigation that could potentially be explored for the development of new classes of
antirheumatic drugs.
IntroductionNow more than ever, immunology is a very exciting and fast-
evolving area of research [1]. The availability of sophisticated
technologies, together with a wide array of information obtained
from comparative biology, has provided scientists with a deeper
understanding of the complexity of adaptive and innate immune
responses. If we consider just one class of immune cells, the T
helper (Th) effector cells, it is easy to see how drastic these changes
have been. The ‘old’ and simplified Th1/Th2 duo representing the
yin and yang of the immune response has now been replaced with
spider diagrams showing naive T cells in the middle and a number
of ramifications leading to Th17, Th22, Th granulocyte macro-
phage colony-stimulating factor (GM-CSF) and Treg, to name the
latest cells reported in the literature [2]. This ever-expanding
universe of T cell subsets, as originally termed by Mosmann and
Sad [3], has served as inspiration for many scientists who have
pushed the boundaries and set themselves the task of discovering
similar subsets of B cells [4], dendritic cells [5], macrophages and
monocytes [6], to keep the list short.
Thus, it seems that the immune system has become as plastic as
the nervous system [7] and, as such, one can barely afford to focus
attention on one cell subset without considering the implications
that this might have on the others. Not surprisingly, the expansion
in so many universes has left scientists (and we the authors of this
review) with the very difficult challenge of singling out specific cell
Corresponding author: D’Acquisto, F. ([email protected])
1359-6446/06/$ - see front matter � 2014 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.drudis.
targets or signaling pathways as the main focus for drug discovery.
Having to face this choice, we decided to focus our attention
deliberately on immunological mechanisms that are currently
targeted by clinically used therapeutics. Two reasons guided us
in this decision: (i) this would be of interest to the readers of Drug
Discovery Today; (ii) the awareness that the mechanisms of action
of long-used and well-known drugs have changed in light of these
discoveries. One such example is aspirin: born as a simple inhibitor
of prostaglandin production [8], it is now considered a transcrip-
tional regulator [9] as well as a tolerogenic agent for dendritic cells
and a modulator of T cell differentiation [10].
Proliferation and survival pathwaysThe lifespan of immune cells is governed by a wide-range of
signaling pathways that are cell-type-specific but that ultimately
lead to two main endpoints: apoptosis and/or necrosis. Whereas
apoptosis is considered an anti-inflammatory response leading to
the resolution of inflammation, necrosis initiates a cascade of
reactions that cause tissue damage and malfunction [11].
Several disease-modifying antirheumatic drugs (DMARDs) have
been shown to influence immune cell proliferation and apoptosis.
Sulfasalazine and its metabolite 5-acetyl salicylic acid have been
shown to exert a proapoptotic effect on vascular smooth muscle
cells [12] and to accelerate immunocomplex-induced neutrophil
apoptosis [13]. Both these effects occurred at concentrations
(20–100 mM), considered too low to inhibit nuclear factor (NF)kB
activation and found in serum of patients taking a standard oral
2014.06.021 www.drugdiscoverytoday.com 1155
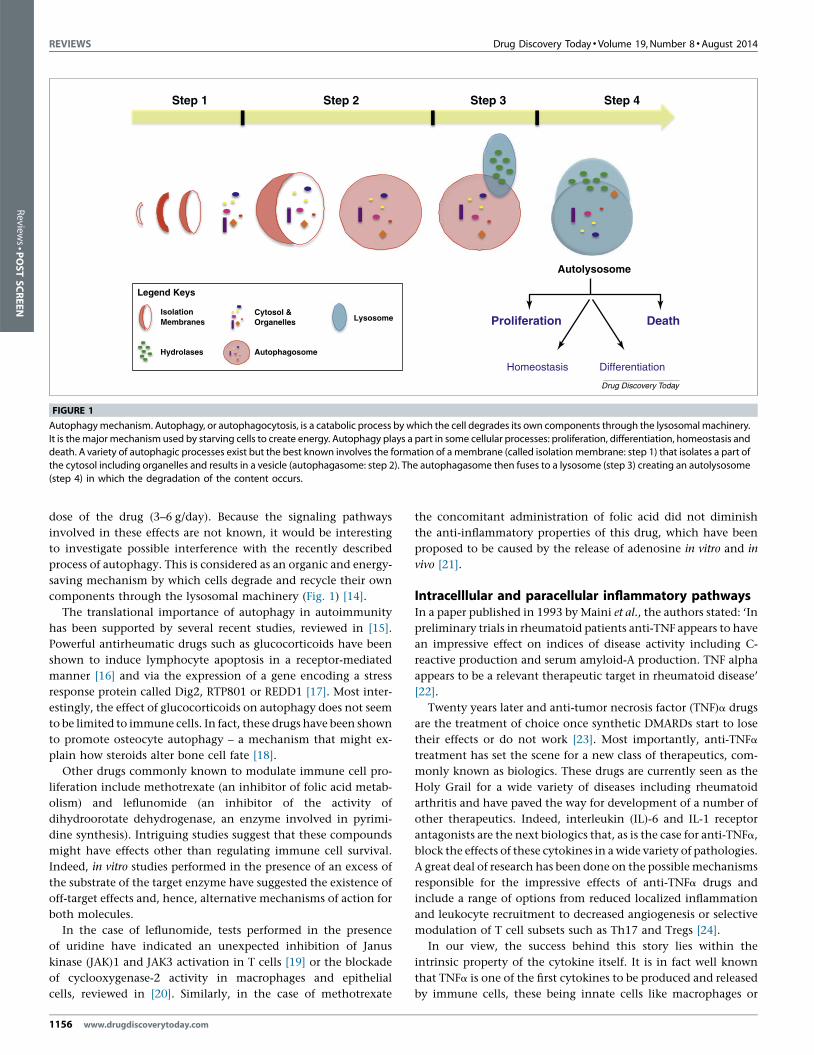
REVIEWS Drug Discovery Today � Volume 19, Number 8 �August 2014
Legend Keys
Isolation Membranes
Cytosol &Organelles Lysosome
Hydrolases Autophagosome
Step 1 Step 2 Step 3 Step 4
Autolysosome
Proliferation Death
Homeostasis Differentiation
Drug Discovery Today
FIGURE 1
Autophagy mechanism. Autophagy, or autophagocytosis, is a catabolic process by which the cell degrades its own components through the lysosomal machinery.
It is the major mechanism used by starving cells to create energy. Autophagy plays a part in some cellular processes: proliferation, differentiation, homeostasis anddeath. A variety of autophagic processes exist but the best known involves the formation of a membrane (called isolation membrane: step 1) that isolates a part of
the cytosol including organelles and results in a vesicle (autophagasome: step 2). The autophagasome then fuses to a lysosome (step 3) creating an autolysosome
(step 4) in which the degradation of the content occurs.
Review
s�P
OSTSCREEN
dose of the drug (3–6 g/day). Because the signaling pathways
involved in these effects are not known, it would be interesting
to investigate possible interference with the recently described
process of autophagy. This is considered as an organic and energy-
saving mechanism by which cells degrade and recycle their own
components through the lysosomal machinery (Fig. 1) [14].
The translational importance of autophagy in autoimmunity
has been supported by several recent studies, reviewed in [15].
Powerful antirheumatic drugs such as glucocorticoids have been
shown to induce lymphocyte apoptosis in a receptor-mediated
manner [16] and via the expression of a gene encoding a stress
response protein called Dig2, RTP801 or REDD1 [17]. Most inter-
estingly, the effect of glucocorticoids on autophagy does not seem
to be limited to immune cells. In fact, these drugs have been shown
to promote osteocyte autophagy – a mechanism that might ex-
plain how steroids alter bone cell fate [18].
Other drugs commonly known to modulate immune cell pro-
liferation include methotrexate (an inhibitor of folic acid metab-
olism) and leflunomide (an inhibitor of the activity of
dihydroorotate dehydrogenase, an enzyme involved in pyrimi-
dine synthesis). Intriguing studies suggest that these compounds
might have effects other than regulating immune cell survival.
Indeed, in vitro studies performed in the presence of an excess of
the substrate of the target enzyme have suggested the existence of
off-target effects and, hence, alternative mechanisms of action for
both molecules.
In the case of leflunomide, tests performed in the presence
of uridine have indicated an unexpected inhibition of Janus
kinase (JAK)1 and JAK3 activation in T cells [19] or the blockade
of cyclooxygenase-2 activity in macrophages and epithelial
cells, reviewed in [20]. Similarly, in the case of methotrexate
1156 www.drugdiscoverytoday.com
the concomitant administration of folic acid did not diminish
the anti-inflammatory properties of this drug, which have been
proposed to be caused by the release of adenosine in vitro and in
vivo [21].
Intracelllular and paracellular inflammatory pathwaysIn a paper published in 1993 by Maini et al., the authors stated: ‘In
preliminary trials in rheumatoid patients anti-TNF appears to have
an impressive effect on indices of disease activity including C-
reactive production and serum amyloid-A production. TNF alpha
appears to be a relevant therapeutic target in rheumatoid disease’
[22].
Twenty years later and anti-tumor necrosis factor (TNF)a drugs
are the treatment of choice once synthetic DMARDs start to lose
their effects or do not work [23]. Most importantly, anti-TNFa
treatment has set the scene for a new class of therapeutics, com-
monly known as biologics. These drugs are currently seen as the
Holy Grail for a wide variety of diseases including rheumatoid
arthritis and have paved the way for development of a number of
other therapeutics. Indeed, interleukin (IL)-6 and IL-1 receptor
antagonists are the next biologics that, as is the case for anti-TNFa,
block the effects of these cytokines in a wide variety of pathologies.
A great deal of research has been done on the possible mechanisms
responsible for the impressive effects of anti-TNFa drugs and
include a range of options from reduced localized inflammation
and leukocyte recruitment to decreased angiogenesis or selective
modulation of T cell subsets such as Th17 and Tregs [24].
In our view, the success behind this story lies within the
intrinsic property of the cytokine itself. It is in fact well known
that TNFa is one of the first cytokines to be produced and released
by immune cells, these being innate cells like macrophages or

Drug Discovery Today � Volume 19, Number 8 �August 2014 REVIEWS
LPS
TLR4
(Step 2) P
Cytoplasm c-Rel p65
(Step1)
P P P
p50 p65 26S Proteasome
c-Rel p65
26S Proteasome
p50 p65
c-Rel p65
NFκB
NFκB
NFκBNFκB
NFκB
(c) (Step 3)
(Step 4)
IκBβ
IκBβ
IκBβ
IκBβ
IκBα
IκBα
p50 p65
c-Rel p65
Nucleus TNFα
TNFα
↑↑
↑↑↑↑
κB2 site of the TNF-α promoter
Drug Discovery Today
FIGURE 2
Control of tumor necrosis factor (TNF)a expression by nuclear factor (NF)kB. Activation of cells by lipopolysaccharide (LPS) induces the phosphorylation of IkBa
and IkBb via two different kinetics: IkBa is the first to be phosphorylated and releases activated p50/p65 heterodimer (step 1); IkBb phosphorylation occurs withslower kinetics and leads to the activation of c-Rel/p65 (step 2). The first ‘pool’ of activated NFkB (p50/p65) translocates first to the nucleus and induces the
transcription of various genes including IkBa, IkBb and TNFa (step 3). The newly synthesized IkBa loops back and recaptures the p50/p65 heterodimer. Activated
c-Rel/p65 specifically binds the kB2 site of the TNFa promoter and forms a complex with the hypophosphorylated and newly synthesized form of IkBb (step 4). In
this way, IkBb makes c-Rel/p65 unresponsive to IkBa and contributes to the long-lasting expression of inflammatory genes such as TNFa.
Reviews�POSTSCREEN
members of the adaptive arm of the immune system like T and B
cells. Such a simple observation might not be so trivial if one
considers the rheumatic inflammatory cytokine cascade to be like
a domino effect where TNFa is just the first element.
A recent interesting study addressed the question of what could
be the mechanism behind the very early production of TNFa
compared with other cytokines. Using an elegant combination
of genetically modified mice and molecular tools, Rao et al. have
shown that activated NFkB complexes contribute to the initial
expression of TNFa. Then, a newly synthesized hypophosphory-
lated form of IkBb facilitates the formation of IkBb/p65/c-Rel
complexes, which selectively bind to the second of the four kB
sites (kB2) in the TNFa promoter, augmenting transcription
(Fig. 2). Consistent with these observations, IkBb-deficient mice
displayed delayed onset, lower incidence and decreased severity of
collagen-induced arthritis [25].
A deeper understanding of the mechanisms regulating IL-1
production has also been recently described. The biologic anakinra
exploits the ability of the IL-1 receptor antagonist (IL-1Ra) to
interfere with the formation of a tripartite complex composed
of IL-1, IL-1R type 1 and IL-1R accessory protein (IL-1acp) [26].
Another way to stop the activity of IL-1 could be achieved by
interfering with the process leading to the release of biologically
active IL-1b. Indeed, contrary to IL-1a, IL-1b must be proteolyti-
cally cleaved from its 31 kDa precursor form to its 17 kDa fragment
to be activated. The inflammasome is a multiprotein complex that
www.drugdiscoverytoday.com 1157

REVIEWS Drug Discovery Today � Volume 19, Number 8 �August 2014
Legend Key s
LRR repeats NACHTdomain
PYD domain CARD domain FIIND
Apoptoticstimuli
IL-1 β IL- 18
NALP1
Active CA SP1
K+ eff lux
Proteolytic
cleava ge
membrane
perturbation NALP3 Pro-IL-1 β
Pro-IL-18
Active CA SP1
Proteolytic
cleava ge
Drug Discovery Today
ASC
ASC
FIGURE 3
NACHT leucine-rich repeat protein (NALP)1 and NALP3 inflammasomes and caspase-1 signaling cascade. The oligomerization and activation of NALP1 and 3 occur
following signals (i.e. K+ efflux and membrane perturbation) triggered by apoptotic stimuli. During their activation, NALP1 and 3 inflammasomes interact with and
activate the apoptosis-associated speck-like protein containing a caspase recruitment domain (CARD) (ASC) through pyrin domain (PYD)–PYD interactions. TheCARD of ASC interacts with the CARD domain of pro-caspase-1, thus giving the active form of caspase-1 by proteolytic cleavage. Caspase-1 then activates the
precursors of interleukin (IL)-1b and IL-18, thus leading to the release of IL-1b and IL-18.
Review
s�P
OSTSCREEN
mediates the cleavage and activation of caspase-1, leading to
maturation and release of IL-1b and IL-18 [27]. Cytoplasmic
receptors of the NACHT leucine-rich repeat (LRR) and (pyrin
domain) PYD domain-containing protein (NLRP) and Nod-like
receptor (NLR) family are crucial components of the inflamma-
some and interact with the adaptor protein apoptosis-associated
speck-like protein containing a caspase recruitment domain
(CARD) (ASC), which recruits the precursor form of caspase-1
(Fig. 3).
The design of small molecules such as specific inhibitors of
inflammasome components might represent a useful alternative
to targeting of the IL-1 signaling pathway. Such an approach
would provide a number of advantages compared with current
biologic therapies. One of the most obvious pros is that synthetic
small molecules, like the DMARDs described before, are not gen-
erally immunogenic and hence not limited in their action by
counter-response of the patient’s immune system to these foreign
molecules. The increasing number of studies reporting the pres-
ence of anti-infliximab and anti-adalimumab antibodies in ‘resis-
tant rheumatoid arthritis patients’ has acted as an alarm bell,
pushing scientists and clinicians in this direction [28].
1158 www.drugdiscoverytoday.com
Exciting news in this regard has recently emerged. Atsttrin (an
antagonist of TNF/TNFR signaling via targeting TNF receptors) is a
small molecule including parts of three granulin domains from
progranulin (PGRN). PGRN was a receptor–orphan polyfunctional
protein until Liu and colleagues discovered that TNFR1 and TNFR2
bind to this protein (Fig. 4). Administration of human recombi-
nant PGRN to mice subjected to collagen-induced arthritis re-
duced disease progression and inhibited synovitis and cartilage
destruction [29]. In the same mouse model, atsttrin decreased
disease severity and delayed onset and progression better than
recombinant human (rh)PGRN; in a collagen–antibody-induced
arthritis mouse model, atsttrin was more effective at treating
inflammation than rhPGRN and etanercept, a soluble TNF recep-
tor. Atsttrin is well absorbed via intraperitoneal administration
and shows high stability and a long half-life (about five days). The
biological functions of PGRN go beyond inflammatory rheumatic
diseases and include other autoimmune pathologies. Patients
suffering from type 2 diabetes have higher levels of PGRN in
the serum compared with healthy subjects [30]. This is consistent
with the view of PGRN as a key adipokine mediating high-fat
diet (HFD)-induced insulin resistance, adipocyte hypertrophy and

Drug Discovery Today � Volume 19, Number 8 �August 2014 REVIEWS
Legend Keys
TNF-α Infliximab Prog ranulin or Atstt rin
(a) (b) (c)
TNF-α TNF-α PGRN PGRN
TNFR1 TNFR2 TNFR1 TNFR2 TNFR1 TNFR2
Pro-I nfl ammati on No signal Anti-I nfl ammati on
Rheumato id Arthritis Rh eumato id Arthritis
Drug Discovery Today
FIGURE 4
Modulation of tumor necrosis factor (TNF)a inflammatory pathway by progranulin. Binding of TNFa to its receptors: TNFR1 and TNFR2, causes their trimerization
and initiates a cascade of events leading to inflammation and autoimmune diseases like rheumatoid arthritis (a). Anti-TNFa therapies such as infliximab block thebinding of TNFa to its receptors and their subsequent oligomerization (b). Progranulin and its derived peptide atsttrin bind to TNFR1 and TNFR2 and act as
physiological antagonists of TNFa signaling (c).
Reviews�POSTSCREEN
obesity [31]. PGRN, like many other members of the adipokine
family such as leptin and adiponectin, is a representative of a new
class of mediators that are implicated in inflammation and meta-
bolic diseases [32]. This new and exciting area of research will
undoubtedly receive further attention in the future because it
remains unclear whether these mediators are harmful or protective
factors [33,34].
Cell–cell interaction and crosstalkIt takes two to tango not only in life but also in autoimmunity. The
realization of this very simple concept has prompted scientists to
investigate key molecules in the crosstalk between T cells and
antigen-presenting cells (APCs) as a potential target for therapy.
One of the best examples in this context is abatacept: a cytotoxic T
lymphocyte antigen (CTLA)-4–IgG fusion protein that modulates
CD28-mediated T cell co-stimulation and is efficacious in the
treatment of rheumatoid arthritis and many other autoimmune
diseases. Elegant recent studies have shown that abatacept sup-
presses antigen-specific T and B cell responses in vivo by suppres-
sing the acquisition of CXCR5+ICOS+ T follicular helper cell
phenotype by antigen-specific T cells despite normal B cell clonal
expansion [35]. Other studies suggested that abatacept reduced the
production of TNFa and IL-12p70 by macrophages [36].
Intriguing evidence has suggested that CTLA-4 expressed on
activated T cells can capture its ligands from opposing APCs by a
process of transendocytosis. T cells actively internalize the CTLA-
4/CD86 complexes through endocytosis and rapidly degrade
CD86 molecules in the cytoplasm thus resulting in impaired
co-stimulation via CD28 [37]. A similar ‘stealing’ mechanism
has been suggested recently for the B-cell-depleting antibody
rituximab (a chimeric anti-CD20 monoclonal antibody). The
wide therapeutic efficacy of this antibody lies in its ability to
cause B cell depletion through mechanisms such as complement-
mediated cytotoxicity and antibody-dependent cellular cytotox-
icity. However, recent studies in the cancer field have hypothe-
sized that rituximab might promote monocyte-mediated shaving
of the CD20/RTX complex from the B cell surface. This shaving
mechanism was the result of active protease activity because
EDTA and phenylmethylsulfonyl fluoride (PMSF) were able to
mediate partial inhibition [38]. Further studies are needed to
verify that such mechanisms occur in rheumatoid arthritis
patients.
www.drugdiscoverytoday.com 1159

REVIEWS Drug Discovery Today � Volume 19, Number 8 �August 2014
Review
s�P
OSTSCREEN
Further research is also needed to identify the mechanisms of
action of tocilizumab, a humanized antibody to IL-6-receptor-a
(IL6R-a). This biologic is supposed to block the multiple effects of
IL-6 in the crosstalk between T cells and APCs. However, despite
on-target suppression of IL-6R signaling in human monocyte-
derived dendritic cells and T cells, there are no data showing an
effect on dendritic cell maturation and/or activation, alloreactive
T cell proliferation, Treg expansion or allogeneic Th1/Th17
responses in vitro [39]. Thus, it seems that tocilizumab in analogy
to metathrexate and lefluonamide might have several other off-
target effects that are yet to be discovered.
Concluding remarks and future perspectivesThe modulation of the immune system in rheumatic disorders
has provided us with an extremely wide-range of opportunities
1160 www.drugdiscoverytoday.com
for drug discovery. We hope that this short review has shed some
light on the current views of the mechanism of action of effec-
tive immunotherapeutics. Whether these novel findings will
support the design of new evidence-based drugs is difficult to
say. Indeed, we firmly think that this is by no means a compre-
hensive account of the myriad of possible ways by which these
drugs continue to improve the life of many people. Conversely,
we hope that these investigations will inspire clinicians and
basic scientists to think ‘outside the box’ and look further
and further.
AcknowledgmentsWe would like to thank Dr Dianne Cooper and Professor Mauro
Perretti for carefully reading the manuscript.
References
1 Medzhitov, R. et al. (2011) Highlights of 10 years of immunology in Nature Reviews
Immunology. Nat Rev Immunol 11, 693–702
2 Annunziato, F. and Romagnani, S. (2009) Heterogeneity of human effector CD4+ T
cells. Arthritis Res Ther 11, 257
3 Mosmann, T.R. and Sad, S. (1996) The expanding universe of T-cell subsets: Th1,
Th2 and more. Immunol Today 17, 138–146
4 Mauri, C. and Bosma, A. (2012) Immune regulatory function of B cell. Annu Rev
Immunol 30, 221–241
5 Adema, G.J. (2009) Dendritic cells from bench to bedside and back. Immunol Lett
122, 128–130
6 Geissmann, F. et al. (2010) Development of monocytes, macrophages, and dendritic
cells. Science 327, 656–661
7 Lee, Y.K. et al. (2009) Developmental plasticity of Th17 and Treg cells. Curr Opin
Immunol 21, 274–280
8 Vane, J.R. (1971) Inhibition of prostaglandin synthesis as a mechanism of action for
aspirin-like drugs. Nat New Biol 231, 232–235
9 Kopp, E. and Ghosh, S. (1994) Inhibition of NF-kappa B by sodium salicylate and
aspirin. Science 265, 956–959
10 Hussain, M. et al. (2011) Aspirin and immune system. Int Immunopharmacol 12, 10–
20
11 Hotchkiss, R.S. et al. (2009) Cell death. N Engl J Med 361, 1570–1583
12 Kim, J.Y. et al. (2009) Sulfasalazine induces haem oxygenase-1 via ROS-dependent
Nrf2 signalling, leading to control of neointimal hyperplasia. Cardiovasc Res 82,
550–560
13 Bertolotto, M. et al. (2009) Sulphasalazine accelerates apoptosis in neutrophils
exposed to immune complex: role of caspase pathway. Clin Exp Pharmacol Physiol
36, 1132–1135
14 Levine, B. et al. (2011) Autophagy in immunity and inflammation. Nature 469, 323–
335
15 Pierdominici, M. et al. (2012) Role of autophagy in immunity and autoimmunity,
with a special focus on systemic lupus erythematosus. FASEB J 26, 1400–1412
16 Swerdlow, S. et al. (2008) Apoptosis inhibition by Bcl-2 gives way to autophagy in
glucocorticoid-treated lymphocytes. Autophagy 4, 612–620
17 Molitoris, J.K. et al. (2011) Glucocorticoid elevation of dexamethasone-induced
gene 2 (Dig2/RTP801/REDD1) protein mediates autophagy in lymphocytes. J Biol
Chem 286, 30181–30189
18 Xia, X. et al. (2010) Glucocorticoid-induced autophagy in osteocytes. J Bone Miner
Res 25, 2479–2488
19 Gonzalez-Alvaro, I. et al. (2009) Inhibition of tumour necrosis factor and IL-17
production by leflunomide involves the JAK/STAT pathway. Ann Rheum Dis 68,
1644–1650
20 Claussen, M.C. and Korn, T. (2012) Immune mechanisms of new therapeutic
strategies in MS – teriflunomide. Clin Immunol 142, 49–56
21 Chan, E.S. and Cronstein, B.N. (2010) Methotrexate – how does it really work? Nat
Rev Rheumatol 6, 175–178
22 Maini, R.N. et al. (1993) TNF-alpha in rheumatoid arthritis and prospects of anti-
TNF therapy. Clin Exp Rheumatol 11 (Suppl 8), 173–175
23 Deighton, C. et al. (2009) Management of rheumatoid arthritis: summary of NICE
guidance. Br Med J 338, b702
24 Feldmann, M. et al. (2010) What have we learnt from targeted anti-TNF therapy?
Ann Rheum Dis 69 (Suppl 1), i97–i99
25 Rao, P. et al. (2010) IkappaBbeta acts to inhibit and activate gene expression during
the inflammatory response. Nature 466, 1115–1119
26 Gabay, C. et al. (2010) IL-1 pathways in inflammation and human diseases. Nat Rev
Rheumatol 6, 232–241
27 Mitroulis, I. et al. (2013) Targeting IL-1beta in disease; the expanding role of NLRP3
inflammasome. Eur J Intern Med 21, 157–163
28 Emi Aikawa, N. et al. (2013) Immunogenicity of anti-TNF-alpha agents in
autoimmune diseases. Clin Rev Allergy Immunol 38, 82–89
29 Tang, W. et al. (2011) The growth factor progranulin binds to TNF receptors and is
therapeutic against inflammatory arthritis in mice. Science 332, 478–484
30 Youn, B.S. et al. (2009) Serum progranulin concentrations may be associated with
macrophage infiltration into omental adipose tissue. Diabetes 58, 627–636
31 Matsubara, T. et al. (2012) PGRN is a key adipokine mediating high fat diet-induced
insulin resistance and obesity through IL-6 in adipose tissue. Cell Metab 15, 38–50
32 Scotece, M. et al. (2011) Beyond fat mass: exploring the role of adipokines in
rheumatic diseases. Sci World J 11, 1932–1947
33 Ebina, K. et al. (2009) Serum adiponectin concentrations correlate with severity of
rheumatoid arthritis evaluated by extent of joint destruction. Clin Rheumatol 28,
445–451
34 Otero, M. et al. (2006) Changes in plasma levels of fat-derived hormones
adiponectin, leptin, resistin and visfatin in patients with rheumatoid arthritis. Ann
Rheum Dis 65, 1198–1201
35 Platt, A.M. et al. (2010) Abatacept limits breach of self-tolerance in a murine model
of arthritis via effects on the generation of T follicular helper cells. J Immunol 185,
1558–1567
36 Wenink, M.H. et al. (2012) Abatacept modulates proinflammatory macrophage
responses upon cytokine-activated T cell and Toll-like receptor ligand stimulation.
Ann Rheum Dis 71, 80–83
37 Qureshi, O.S. et al. (2011) Trans-endocytosis of CD80 and CD86: a molecular basis
for the cell-extrinsic function of CTLA-4. Science 332, 600–603
38 Pedersen, A.E. et al. (2011) Monocytes mediate shaving of B-cell-bound anti-CD20
antibodies. Immunology 133, 239–245
39 Betts, B.C. et al. (2011) Anti-IL6-receptor-alpha (tocilizumab) does not inhibit
human monocyte-derived dendritic cell maturation or alloreactive T-cell response.
Blood 118, 5340–5343





























