Embed Size (px)
Citation preview

Molec. gen. Genet. 147, 45-5l (1976) © by Springer-Verlag 1976
Non Xenopus-like DNA Sequence Organization in the Chironomus tentans Genome
Richard Wells, Hans-Dieter Royer, and Cornelis P. Hollenberg
Max-Planck-Institut fiir Biologie, Abt. Beermann, Spemannstr. 34, Ttibingen, Federal Republic of Germany
Summary. We have examined the sequence organiza- tion of Chironomus tentans DNA by means of optical and hydroxyapatite renaturation kinetics of total DNA fragment sizes of 0.36, 2.6 and 13.5 kilobases (kb) as well as isolated middle repeat DNA at sizes of 0.36 and 13.5 kb. 90 % of the DNA renatured as unique sequences of a genome of 0.20 pg with the balance of DNA renaturing as middle repetitive sequences present on average 90 times per haploid genome. At a DNA fragment length of 13.5 kb, 35 % of the DNA was trapped on the hydroxyapatite as middle repetitive fraction. We concluded C. tentans DNA to have a mean repeat length of about 4.3 kb distributed through out at least 35 % of the genome with an inter repeat spacing of at least 13.5 kb but possibly being distrib- uted throughout the whole genome with an inter repeat spacing of 36 kb. This shows C. tentans DNA organization not to follow the almost ubiquitous Xenopus model but to be similar to the organization of Drosophila melanogaster DNA.
Introduction
It is now well established that the DNA of eukaryotes contains middle repetitive sequences some of which are interspersed with most of the non-repetitive DNA in regular patterns.
In Xenopus laevis, the organism for which the first quantitative estimates were', made by Davidson et al. (1973 a, b), 50 % of the genome consists of repet- itive sequences about 300 nucleotides long inter- spersed between non-repetitive sequences about 700-900 nucleotides long. In another 25% of the genome these 300 nucleotide repetitive sequences are interspersed among non-repetitive sequences longer than 4,000 nucleotides. 25 % of the middle repetitive sequences occurs as longer fragments and 20 % of the
genome consists of unrepetitive D N A without detec- table repetitive sequences being present.
An interspersion pattern basically similar to that of Xenopus DNA has been found in the DNA of a variety of animals belonging to both the Deutero- stomata and Protostomata (see review Davidson et al., 1975). In all cases but one a major fraction of the DNA consisted of non-repetitive sequences less than about 3,000 nucleotides in length, interspersed with repetitive sequences, in majority 300 nucleotides in length. The exception was found by Manning, Schmid and Davidson (1975) in Drosophila melano- gaster, where middle repetitive sequences with an average length 5,600 nucleotides are interspersed within non-repetitive sequences longer than 13,000 nucleotides. This organisation pattern, however, does not seem to exist in all insects. Efstradiatis et al. (1976) have shown that the D N A of the moth Antheraea pernyi has the Xenopus type of sequence interspersion, while the DNA of the honey bee Apis mellifica follows the Drosophila model (E. Davidson, personal com- munication). In view of these unexpected differences within the insects, it was of interest to ask which interspersion pattern the DNA of another species of the group of Diptera, Chironomus tentans, follows.
We have determined the renaturation kinetics of C. tentans at single strand size classes of 0.36, 2.6 and 13.5 kilobases (kb) to answer this question.
Materials and Methods
DNA Preparation
Chironomus tentans larvae (40 g) or adults (20 g) were ground in a cool mortar in 0.15 M NaCI-0.1 M EDTA (pH 8) and further homogenized in a volume of 50 ml in a Dounce homogenizer by five strokes of the loose pestle. Sarkosyl (0.5 ~) and proteinase K (400 ~tg/ml) were added and the lysate was incubated for 1 h at 60 °C. After addition of fresh proteinase K (100 gg/ml) and Sarkosyl to a final concentration of 1%, the mixture was incubated for 20 min
46 R. Wells et al.: DNA Sequence Organization in C. tentans
at 60 °C. 13 ml of 5 M NaC10 4 was added at room temperature and the solution was shaken for 5 min with 1 vol of water saturated phenol and subsequently for 5 min with 1 vol chloroform. The phases were separated by centrifugation for 20 min at 8,000 xg. The chloroform/phenol layer was removed and the remaining upper phase was shaken for 5 rain with 1 vol chloroform. 2 vols ethanol were added to the recovered water phase. Nucleic acids were spooled out, dissolved in 10 ml of 1 x SSC and incubated with pancreatic (20 gg/ml) and T1 (200 U/ml) ribonucleases and ~- amylase (100 gg/ml) for 2 h at 30 °C, 200 gg of proteinase K per ml was added, followed by 3 h incubation at 30 °C. The solution was shaken for 15 rain with 10 ml chloroform. The recovered water phase was overlayered with 2 vols ethanol. DNA was spooled,out and dissolved in 1 x SSC. 1.3 g CsC1 was added per ml. The solution was centrifuged in a Spinco 60 Ti rotor for 42 h at 44,000 rpm. All fractions containing DNA were pooled and diluted with 1 vol H20. DNA was precipitated with 2 vols ethanol and dissolved in SSC. Recovery: 1 mg DNA per 5 g flies; 1 mg DNA per 30 g larvae.
Preparation and Sizing of DNA Fragments
Large DNA fragments were those obtained directly from the DNA preparation. Smaller fragment sizes were obtained by using a Virtis 60 homogenizer as described by Britten, Graham and Neufeld (1974) or by sonication in a Branson sonifier (only used for smallest fragments). Average molecular weights of single strand DNA were determined by sedimentation in 0.9 M NaC1, 0.1 M NaOH (Studier, 1965) in an MSE Centriscan 75 at speeds of 20,000-40,000 rpm.
Reassociation of DNA
All reassociation was performed at 60 °C in 0.12 M sodium phos- phate buffer (PB) (pH 6.9). Prior to the reassociation the DNA was run over Chelex (Bio Rad Labs Chelex 100) in order to remove metal ions.
For optically measured kinetics the DNA was placed in 1 em or 1 mm path length cells of a Gilford 240. The temperature was raised to 100 °C for 5 rain and then lowered to 60 °C. Under these conditions 75 °C is reached within 1 rain from 100 °C and this point was taken as the start time of the run. K 2 values were calculated according to Wetmur and Davidson (1968).
Hydroxyapatite (HAP) kinetics were followed by measuring duplex-region containing DNA attaching to the HAP at 0.12 M PB at 60°C as described by Britten and Kohne (1967). HAP was prepared according to Tiselius, Hjert6n and Levin (1956). 10 ~1 to 25 gl samples of DNA at concentrations of 300 to 700 ~g/ml were placed in 50 ~tl microcaps and sealed. The samples were denatured for 5 min at 100 °C and renatured at 60 °C. A renaturation point was obtained by loading the content of one microcap onto an HAP column and measuring the amount of DNA retained compared to the total DNA, all A 2 6 0 n m values being taken at 98 °C. Bound DNA was eluted from the column by 0.5 M PB. Hyperchromicity was measured in a Gilford 240 and taken as the relative rise in optical density at 260 nm between 60 °C and 98 °C. Cot1/2 values were calculated according to Britten and Kohne (1967) and expressed in M x see. For the calculation of the percentage of repetitive sequences, the apparent percentage was corrected for zero-time binding, assuming a random distribution of fold-back sequences throughout the genome.
Isolation of Repetitive DNA
Repetitive sequences were isolated from DNA of single strand length of 0.36 kb by three steps. (1) Isolation of DNA renaturing
between Cot I 0 and Cot 30.0. This should result in obtaining all the repetitive sequences plus contaminating inverted repeats and a small amount of unique sequences. (2) Removing DNA of Cot 3 x 10 -3, which should result in removing inverted repeats. (3) Re- moving DNA of Cot 30.0 which essentially consists of unique sequences. The renaturation rate compared to the DNA 3.103< Cot<30.0 was 100 times slower. Repetitive sequences from the 13.5 kb were isolated in a similar manner except that the 1st cut contained the fraction of DNA bound within the limits of 10-3< Cot < 5.0.
Results
Characteristics of C. tentans D N A
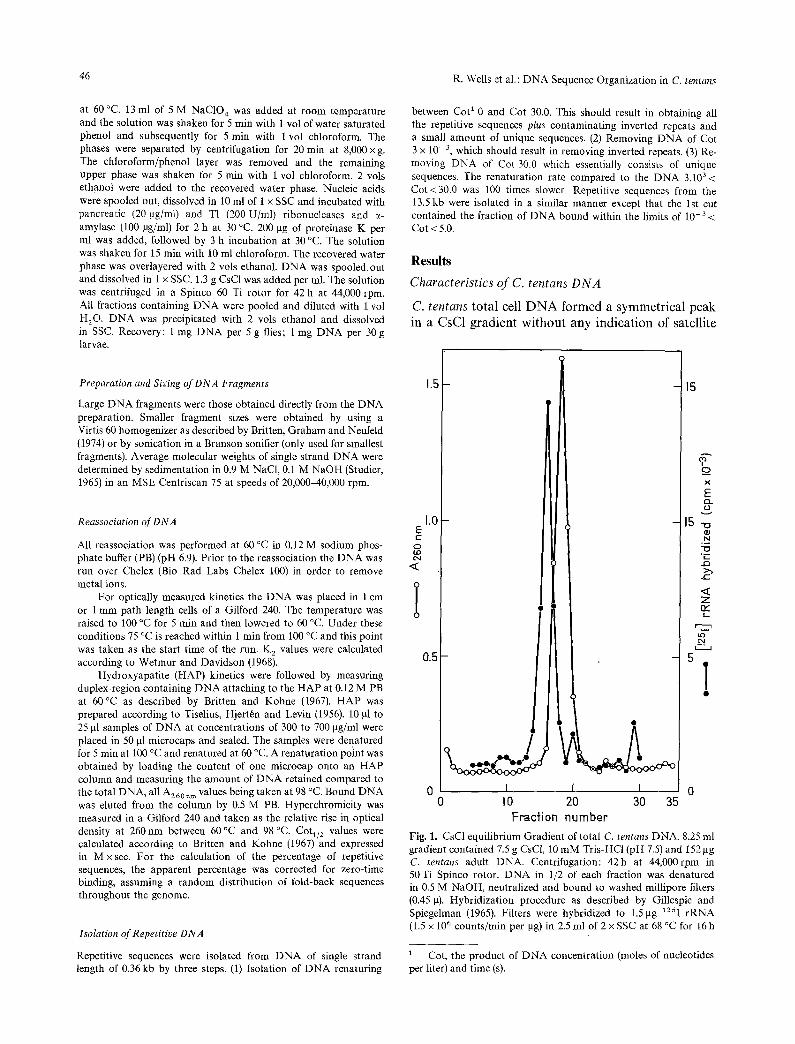
C. tentans total cell DNA formed a symmetrical peak in a CsC1 gradient without any indication of satellite
1.5
I.O E c-
(21
<
0 . 5 -
0 I I I 0 I0 20
Fraction number
- 1 5
13. O
- 1 5 -a N
. m
z.. A21
< z n,- L..
5
I
I 0 30 35
Fig. 1. CsC1 equilibrium Gradient of total C. tentans DNA, 8.25 ml gradient contained 7.5 g CsC1, 10 mM Tris-HCl (pH 7.5) and 152gg C. tentans adult DNA. Centrifugation: 42h at 44,000rpm in 50 Ti Spinco rotor. DNA in 1/2 of each fraction was denatured in 05 M NaOH, neutralized and bound to washed millipore filters (0.45 g). Hybridization procedure as described by Gillespie and Spiegelman (1965). Filters were hybridized to 1.5 gg 125I rRNA (1.5 x 106 counts/rain per gg) in 2.5 ml of 2 x SSC at 68 °C for 16 h
t Cot, the product of DNA concentration (moles of nucleotides per liter) and time (s).
R. Wells et al.: DNA Sequence Organization in C. tentans
1.4
1.3
O
E O
.~ 1.2 2 (D e-I
-1-
1.0 ~l I 70 80 90 IO0
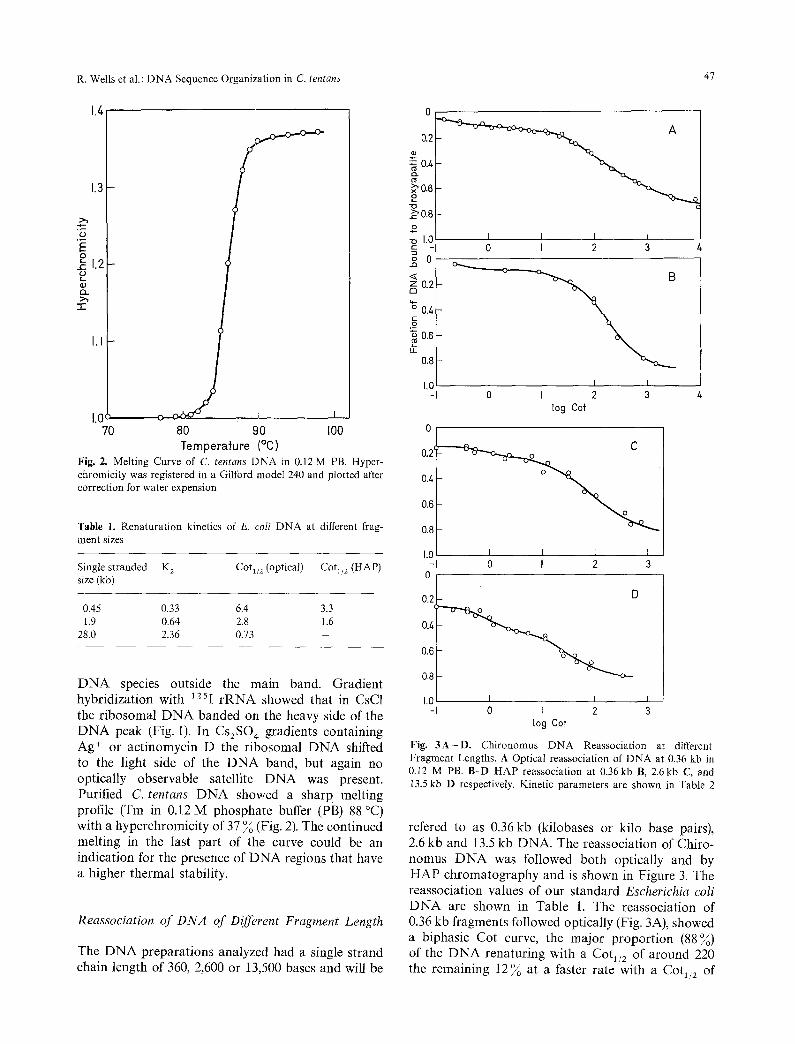
Temperature (%) Fig. 2. Melting Curve of C. tentans DNA in 0.12 M PB. Hyper- chromicity was registered in a Gilford model 240 and plotted after correction for water expension
Table 1. Renaturation kinetics of E. coli DNA at different frag- ment sizes
Single stranded K 2 Cot1/2 (optical) Cot1/2 (HAP) size (kb)
0.45 0.33 6.4 3.3 1.9 0.64 2.8 1.6
28.0 2.36 0.73 -
D N A species outside the main band. Gradient hybridization with 125I r R N A showed that in CsC1 the ribosomal D N A banded on the heavy side of the D N A peak (Fig. 1). In Cs2SO 4 gradients containing Ag ÷ or actinomycin D the ribosomal D N A shifted to the light side of the D N A band, but again no optically observable satellite D N A was present. Purified C. tentans D N A showed a sharp melting profile (Tm in 0.12 M phosphate buffer (PB) 88 °C) with a hyperchromicity of 37 % (Fig. 2). The continued melting in the last part of the curve could be an indication for the presence of D N A regions that have a higher thermal stability.
Reassociation of DNA of Different Fragment Length
The D N A preparations analyzed had a single strand chain length of 360, 2,600 or 13,500 bases and will be
47
o =z o -o c
o
0.2
0.4
0.6
0.8
1.0 -I
0
I I I f 0 I 2 3
.13
<[ Z £3
"s
9 I_ U_
0 . 2 -
0.4-
0.6-
0.8-
1.0 -I
I I I I 0 I 2 3
tog Cot
0 . 2
0.4
0.6
0.8 /
[.c [ I [ I -I 0 I 2 3
0
I D
o o
I I I 0 I 2
log Cot
0.2
0.4
0.6
0.8
1.0 -I
Fig. 3A-D. Chironomus DNA Reassociation at different Fragment Lengths. A Optical reassociation of DNA at 0.36 kb in 0.12 M PB. B-D HAP reassociation at 0.36kb B, 2.6kb C, and 13.5 kb D respectively. Kinetic parameters are shown in Table 2
refered to as 0.36 kb (kilobases or kilo base pairs), 2.6 kb and 13.5 kb DNA. The reassociation of Chiro- nomus D N A was followed both optically and by HAP chromatography and is shown in Figure 3. The reassociation values of our standard Escherichia coli D N A are shown in Table 1. The reassociation of 0.36 kb fragments followed optically (Fig. 3A), showed a biphasic Cot curve, the major proportion (88%) of the D N A renaturing with a Cot1/2 of around 220 the remaining 12% at a faster rate with a Cot1/2 of
48 R. Wells et al.: DNA Sequence Organization in C. tentans
Table 2. Renaturation kinetics of Chironomus DNA
Single stranded COtl/2 (HAP) Cot1/2 Cot1/2 size (kb) 1st component 2nd component
0.36 150 2.0 190 2.6 80 0.5 75
13.5 15 0.9 32
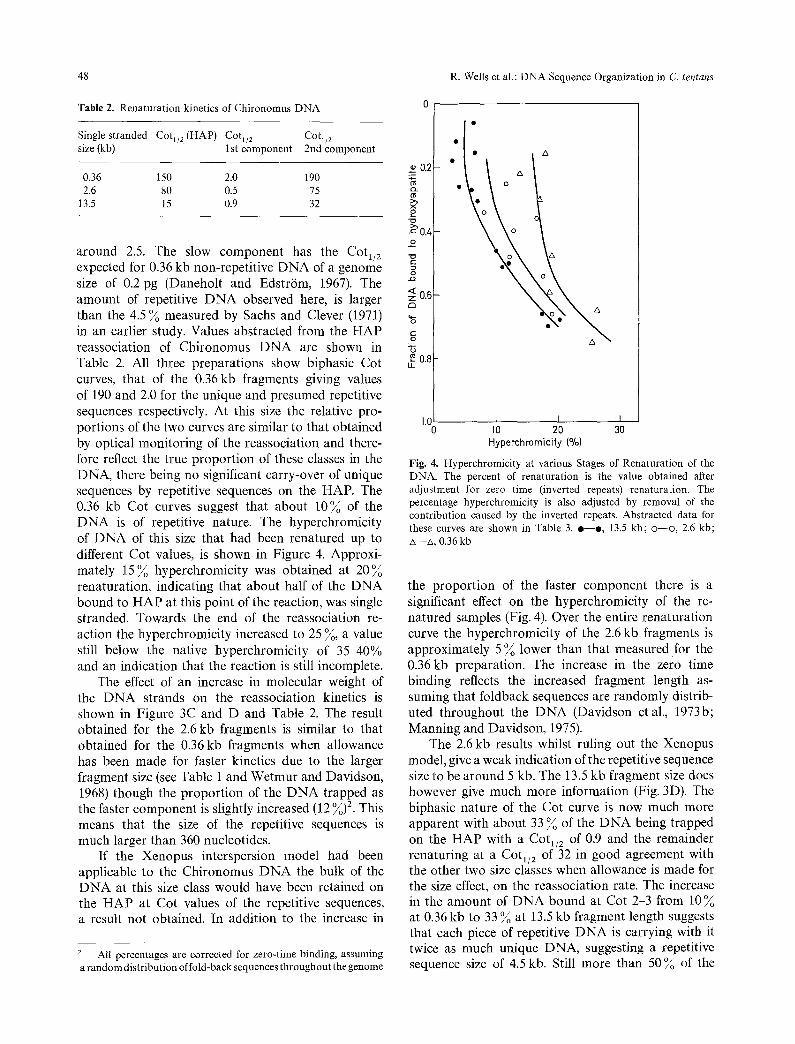
around 2.5. The slow component has the Cot1/2 expected for 0.36 kb non-repetitive DNA of a genome size of 0.2 pg (Daneholt and Edstr/Sm, 1967). The amount of repetitive DNA observed here, is larger than the 4.5 ~o measured by Sachs and Clever (1971) in an earlier study. Values abstracted from the HAP reassociation of Chironomus DNA are shown in Table 2. All three preparations show biphasic Cot curves, that of the 0.36kb fragments giving values of 190 and 2.0 for the unique and presumed repetitive sequences respectively. At this size the relative pro- portions of the two curves are similar to that obtained by optical monitoring of the reassociation and there- fore reflect the true proportion of these classes in the DNA, there being no significant carry-over of unique sequences by repetitive sequences on the HAP. The 0.36 kb Cot curves suggest that about 10~o of the DNA is of repetitive nature. The hyperchromicity of DNA of this size that had been renatured up to different Cot values, is shown in Figure 4. Approxi- mately 15 ~o hyperchromicity was obtained at 20~o renaturation, indicating that about half of the DNA bound to HAP at this point of the reaction, was single stranded. Towards the end of the reassociation re- action the hyperchromicity increased to 25 ~o, a value still below the native hyperchromicity of 35-40% and an indication that the reaction is still incomplete.
The effect of an increase in molecular weight of the DNA strands on the reassociation kinetics is shown in Figure 3C and D and Table 2. The result obtained for the 2.6 kb fragments is similar to that obtained for the 0.36 kb fragments when allowance has been made for faster kinetics due to the larger fragment size (see Table 1 and Wetmur and Davidson, 1968) though the proportion of the DNA trapped as the faster component is slightly increased (12 ~o) 2. This means that the size of the repetitive sequences is much larger than 360 nucleotides.
If the Xenopus interspersion model had been applicable to the Chironomus DNA the bulk of the DNA at this size class would have been retained on the HAP at Cot values of the repetitive sequences, a result not obtained. In addition to the increase in
2 All percentages are corrected for zero-time binding, assuming a random distribution of fold-back sequences throughout the genome
0.2 o~ >.1 x o
~0.~ o
e~
< z 0.6
~6
"6 0.8
IJ_
° o o /x //t
• o A
A
1.0 I I I I0 20 30
Hyperehromicity (%)
Fig. 4. Hyperchromicity at various Stages of Renaturation of the DNA. The percent of renaturation is the value obtained after adjustment for zero time (inverted repeats) renaturation. The percentage hyperchromicity is also adjusted by removal of the contribution caused by the inverted repeats. Abstracted data for these curves are shown in Table 3. e - - o , 13.5 kb; o - -o , 2.6 kb; zx--zx, 0.36 kb
the proportion of the faster component there is a significant effect on the hyperchromicity of the re- natured samples (Fig. 4). Over the entire renaturation curve the hyperchromicity of the 2.6 kb fragments is approximately 5 ~o lower than that measured for the 0.36kb preparation. The increase in the zero time binding reflects the increased fragment length as- suming that foldback sequences are randomly distrib- uted throughout the DNA (Davidson et al., 1973b; Manning and Davidson, 1975).
The 2.6 kb results whilst ruling out the Xenopus model, give a weak indication of the repetitive sequence size to be around 5 kb. The 13.5 kb fragment size does however give much more information (Fig. 3D). The biphasic nature of the Cot curve is now much more apparent with about 33 ~o of the DNA being trapped on the HAP with a Cot1/2 of 0.9 and the remainder renaturing at a Cot1/2 of 32 in good agreement with the other two size classes when allowance is made for the size effect, on the reassociation rate. The increase in the amount of DNA bound at Cot 2-3 from 10 ~o at 0.36 kb to 33 ~ at 13.5 kb fragment length suggests that each piece of repetitive DNA is carrying with it twice as much unique DNA, suggesting a repetitive sequence size of 4.5 kb. Still more than 50~o of the
R. Wells et al.: DNA Sequence Organization in C. tentans
Table 3. Hyperchromicities of renatured samples. Values from Figure 4
Single stranded size (kb)
Percentage renatured a
16 20 24 28
0.36 14.8 15.0 15.2 15.4 2.6 9.2 9.7 10.2 10.9
13.5 4.3 4.6 5.0 5.4
0.36/2.6 1.61 1.55 1.49 1.41 0.36/13.5 3.44 3.26 3.04 2.85
" See legend to Figure 4
DNA renatured at a Cot value of unique DNA sequences. Thus at least 50 ~ of the single copy DNA occurs in stretches of 13.5 kb not interspersed with repetitive sequences.
The remelting data (Fig. 4) show the hyper- chromicity of the renatured 13.5kb samples to be very much lower than that of native DNA. Once again the results are consistent with only a portion of the DNA trapped on the HAP to be in duplex form. Early in the reassociation of the DNA these differences are most significant and values for the reassociation range of 16-28 ~ are shown in Table 3. We find the ratio of hyperchromicities of the 0.36 kb to the 13.5 kb fragments to be in the range of 2.85-3.44 suggesting the double stranded region to be between 3.9 kb and 4.7 kb. We appreciate that this is a very crude approximation, but nevertheless feel that it gives a worthwhile estimate.
Reassociation of Isolated Repetitive DNA
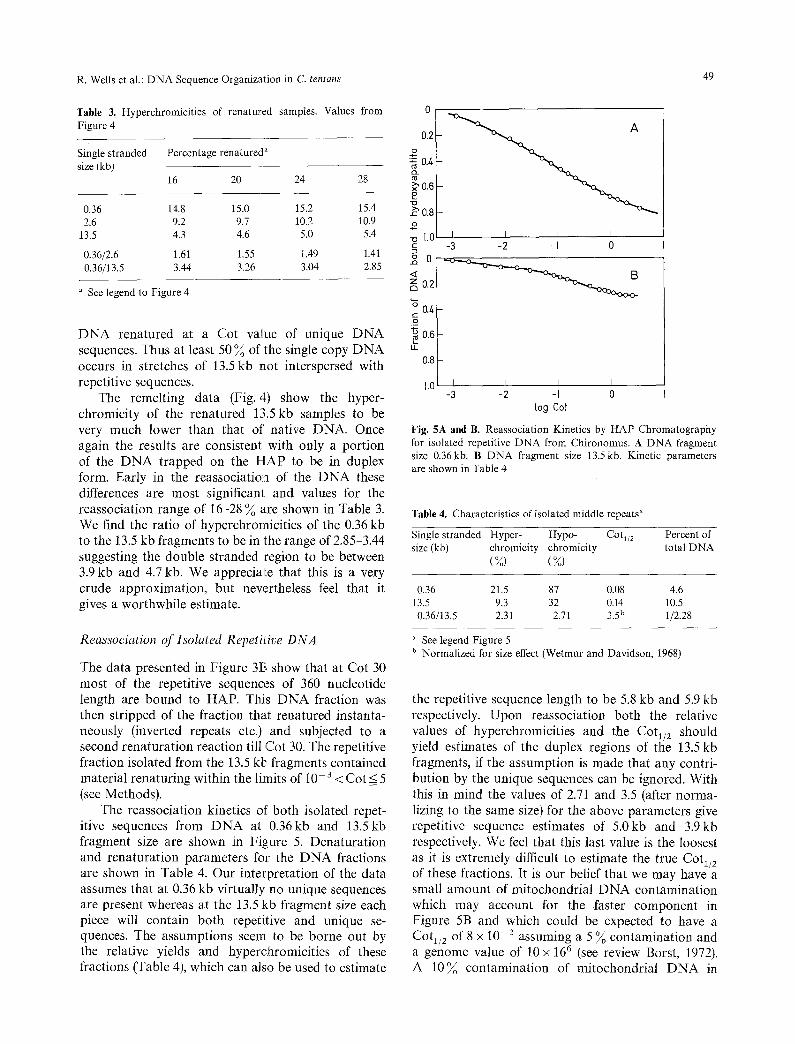
The data presented in Figure 3t3 show that at Cot 30 most of the repetitive sequences of 360 nucleotide length are bound to HAP. This DNA fraction was then stripped of the fraction that renatured instanta- neously (inverted repeats etc.) and subjected to a second renaturation reaction till Cot 30. The repetitive fraction isolated from the 13.5 kb fragments contained material renaturing within the limits of 10- 3 < Cot < 5 (see Methods).
The reassociation kinetics of' both isolated repet- itive sequences from DNA at 0.36kb and 13.5kb fragment size are shown in Figure 5. Denaturation and renaturation parameters for the DNA fractions are shown in Table 4. Our interpretation of the data assumes that at 0.36 kb virtually no unique sequences are present whereas at the 13.5 kb fragment size each piece will contain both repetitive and unique se- quences. The assumptions seem to be borne out by the relative yields and hyperchromicities of these fractions (Table 4), which can also be used to estimate
49
0.2
bo.6~
~ 0.8
~ I . 0 I -2 -I 0 -3
g o A21
z 0.2
46 0.4 g 0.6
L I.L
0.8
1 . 0 I I I I -3 -2 -I 0
lo9 Cot
Fig. 5A and B. Reassociation Kinetics by HAP Chromatography for isolated repetitive DNA from Chironomus. A DNA fragment size 0.36kb. B DNA fragment size 13.5 kb. Kinetic parameters are shown in Table 4
Table 4. Characteristics of isolated middle repeats a
Single stranded Hyper- Hypo- Cot1/2 Percent of size (kb) chromicity chromicity total DNA
(Voo) (%)
0.36 21.5 87 0.08 4.6 13.5 9.3 32 0.14 10.5 0.36/13.5 2.31 2.71 3.5 b 1/2.28
See legend Figure 5 b Normalized for size effect (Wetmur and Davidson, 1968)
the repetitive sequence length to be 5.8 kb and 5.9 kb respectively. Upon reassociation both the relative values of hyperchromicities and the Cot~/2 should yield estimates of the duplex regions of the 13.5 kb fragments, if the assumption is made that any contri- bution by the unique sequences can be ignored. With this in mind the values of 2.71 and 3.5 (after norma- lizing to the same size) for the above parameters give repetitive sequence estimates of 5.0kb and 3.gkb respectively. We feel that this last value is the loosest as it is extremely difficult to estimate the true Cot1/2 of these fractions. It is our belief that we may have a small amount of mitochondrial DNA contamination which may account for the faster component in Figure 5B and which could be expected to have a Cot1/2 of 8 x 10- z assuming a 5 ~ contamination and a genome value of 10 x 166 (see review Borst, 1972). A 10~ contamination of mitochondrial DNA in

50 R. Wells et al.: DNA Sequence Organization in C. tentans
our 0.36 kb fragment size repetitive DNA would have a Cot1/2 value of about 0.3 and not significantly influence the Cot1/2 value of this fraction.
Discussion
We have examined the DNA of C. tentans by means of renaturation kinetics and have found a large proportion (90 %) to be of unique sequences with the remaining 10~o to be middle repeated DNA. The lack of a very fast component in the C. tentans DNA is paralleled by the lack of heterochromatin in this organism (Beermann, 1962).
Internal controls for both HAP and optical renaturation systems were provided by use of E. coli DNA. The possibility of breakdown of our long DNA fragments over the course of renaturation was ruled out by comparing the zero time renaturation of the DNA at the beginning and end of the experiment.
The HAP results with DNA of 2.6 and 13.5 kb in length show that the arrangement of repeated se- quences within the Chironomus DNA is similar to that of Drosophila DNA and is not of the Xenopus type. Analyses of our optical renaturation (Fig. 3A) results at 0.36 kb provide us with an estimate of 12 % for the repeated sequences within the DNA. Our estimates of this fraction from HAP renaturation data are in the order of 10%. At a fragment size of 2.6kb this fraction can be determined to be 12% and at 13.5kb to be about 33%. These HAP ex- periments demonstrate increasingly larger amounts of DNA being trapped on the columns as the DNA size increases and allows us to make an estimate of 4.5 kb for the length of the repeated sequence by a statistical analysis, assuming that the repeated se- quences are at least 13.5 kb apart. This interspersion model is limited to the 33 % of the DNA trapped on HAP at 13.5 kb fragment length, but may apply to most of the genome. Much longer fragment lengths would be necessary to determine the exact proportion of the DNA following this pattern. The hyperchromi- cities of the renatured DNA fractions at 0.36 and 13.5 kb (Table 3) give information on the regions of duplex formation of this DNA and lead to estimates of the length of the repeated sequences to be in the range of 3.9 to 4.7 kb. We prefer the value of 3.9 kb derived from earlier times in the renaturation as the contamination by unique sequences not carrying repeated DNA is minimized.
Results from the isolated middle repeats were analyzed with the assumption that the 0.36 kb fraction represents only repeated sequences and that the 13.5 kb fraction represented this DNA plus unique sequences. We must however take into account the mitochondrial DNA contribution within the iso-
lated middle repetitive fractions that could amount to maximum levels of 10 % and 5 % for the 0.36 and 13.5kb DNA fractions respectively. Ignoring mito- chondrial DNA contribution, we get a range of values of 3.3 to 5.9 kb from the relative hyperchromi- cities and hypochromicities as well as the Cot1/e values and middle repeat content at the two sizes of DNA. If the possible mitochondrial contamination is taken into account the range of values becomes 3.1 to 5.0kb. These values are in good agreement with those obtained from the 13.5 kb HAP experi- ments and the relative hyperchromicities of the 0.36 and 13.5 kb fractions. We therefore propose a value of around 4.3 for the length of the repeated sequences of Chironomus DNA, each sequence being at least 13.5 kb in distance from its neighbouring repeat. As this fraction represents maximally some 12 ~o of the total DNA an upper estimate of 5,000 for the total number of repeated sequences can be put forward. The HAP results however indicate a relative reiteration of around 90 and we therefore propose that the repeated sequences of C. tentans comprise a maximum of 55 families with an average of 90 each. If we assume complete interspersion of these sequences within the total genome this would represent one sequence per 36 kb (DNA of genome 0.2 pg). Our data cannot however rule out the presence of a few percent of the genome with Xenopus-like organisation.
Whilst no positive evidence exists for the function of the repeated DNA within a genome it is interesting to find that two taxonomically close organisms such as D. melanogaster and C. tentans both show non Xenopus-like DNA organisation. It is therefore sur- prising that the more distantly related honeybee (E. Davidson, personal communication) shows the Drosophila pattern whilst the more closely related moth has a Xenopus-like DNA organisation.
Preliminary results with the DNA of Pales ferruginea (Tipulidae) that show a very strong Xenopus- like effect of the fragment length on the Cot1/2 value of the bulk of the DNA, make us think that the inter- spersion of short middle repeated sequences as observed in Cot curves at different DNA fragment lengths, does not reflect an essential aspect of genome organisation but has more to do with the presence and maintenance of an apparent excess of DNA indicated by the striking variation in (haploid) genome size within many groups of closely related organisms. P.ferruginea has about ten times more DNA than C. tentans and D. melanogaster. More data, however, are required to substantiate this notion.
Acknowledgements. We express our gratitude to Drs. B. Hough and E. Davidson who performed computer analysis on our Cot curves and to Professor Dr. W. Beermann for stimulating discussion and support.

R. Wells at al.: DNA Sequence Organization in C. tentans 51
References
Beermann, W.: Riesenchromosomen. Protoplasmatologia VI/D. Wien: Springer 1962
Borst, P.: Mitochondrial nucleic acids. Ann. Rev. Biochem. 41, 333-376 (1972)
Britten. R.J., Graham, D.E., Neufeld, B.R.: Analysis of repeating DNA sequences by reassociation. In: Methods in enzymology. (L. Grossman and K. Moldave, eds.), Vol. 29, part E, pp. 363-418. New York: Academic Press 1974
Britten, R.J., Kohne, D. E.: Nucleotide sequence repetition in DNA. Carnegie Inst. Year Book 65, 78-106 (1967)
Daneholt, B., Edstr/Sm, J.-E.: The content of deoxyribonucleic acid in individual polytene chromosomes of Chironomus tentans. Cytogenetics 6, 350-356 (1967)
Davidson, E.H., Galau, G.A., Angerer, R.C., Britten, R.J.: Com- parative aspects of DNA sequence organization in metazoa. Chromosoma (Bed) 51, 253-259 (1975)
Davidson, E.H., Graham, D.E., Neufeld, B.R., Chamberlin, M.E., Amenson, C.S., Hough, B.R., Britten, R.J.: Arrangement and characterization of repetitive sequence elements in animal DNAs. Cold Spr. Harb. Syrup. quant. Biol. 38, 295-301 (1973b)
Davidson, E. H., Hough, B. R., Amenson, C. S., Britten, R. J.: General interspersion of repetitive with non-repetitive sequence elements in the DNA of Xenopus. J. molec. Biol. 77, 1-23 (1973a)
Efstradiatis, A., Crain, W. R., Britten, R.J., Davidson, E.H., Kafatos, F.C. : DNA sequence organization in the Lepidopteran Antheraea pernyi. Chromosoma (Bed.) in press (1976)
Gillespie, D., Spiegelman, S.: A quantitative assay for DNA-RNA hybrids with DNA immobilized on a membrane. J. molec. Biol, 12, 829-842 (1965)
Manning, J.E., Schmid, C.W., Davidson, N.: Interspersion of re- petitive and non-repetitive DNA sequences in the Drosophila melanogaster genome. Cell 4, 141-155 (1975)
Sachs, R.I., Clever, U.: Unique and repetitive DNA sequences in the genome of Chironomus tentans. Exp. Cell Res. 74, 587-591 (i972)
Studier, F.W.: Sedimentation studies of the size and shape of DNA. J. molec. Biol. 11, 373-390 (1965)
Tiselius, A., Hjert6n, S., Levin, O.: Protein chromatography on calcium phosphate columns. Arch. Biochem. Biophys. 65, 132-155 (1956)
Communicated by E. Bautz
Received April 12, 1976