Embed Size (px)
Citation preview

NON-TARGET-SITE RESISTANCE TO ALS INHIBITORS IN WATERHEMP
BY
JIAQI GUO
THESIS
Submitted in partial fulfillment of the requirements
for the degree of Master of Science in Crop Sciences
in the Graduate College of the
University of Illinois at Urbana-Champaign, 2014
Urbana, Illinois
Master’s Committee:
Professor Patrick J. Tranel, Chair
Professor Dean E. Riechers
Associate Professor Aaron G. Hager

ii
ABSTRACT
The acetolactate synthase (ALS) enzyme, or acetohydroxyacid synthase (AHAS) enzyme,
is an essential enzyme in branched-chain amino acid biosynthesis, and is the target site of five
families of herbicides referred to as ALS inhibitors. Waterhemp (Amaranthus tuberculatus) is
considered one of the most problematic weeds in the Midwest cropping region. The evolution of
herbicide resistance and multiple resistance mechanisms within the species is one of the major
properties making it difficult to control, and ALS-resistant waterhemp populations have been
found and studied considerably. A waterhemp population (designated MCR) from Illinois with
resistance to HPPD and atrazine was found to segregate for both high and moderate levels of
resistance to ALS inhibitors. Plants in this population with high-level resistance had the
Trp574Leu ALS mutation, which is present in other waterhemp populations resistant to ALS
inhibitors. Plants from the MCR population that showed only moderate levels of resistance to
ALS inhibitors did not have this mutation. Thus, research was conducted to investigate the
resistance mechanism in the waterhemp plants with moderate resistance to ALS-inhibitors.
Plants with moderate resistance were crossed and the resulting progeny where characterized.
Firstly the ALS gene of the progeny was sequenced and in vitro ALS enzyme assays were
conducted, and results indicated that the plants lacked a target-site mutation. Secondly, a series
of greenhouse dose-response experiments were conducted to evaluate the resistance level across
different chemical families of ALS-inhibitors. Thirdly, malathion, a cytochrome P450-inhibiting
pesticide, was incorporated with ALS-inhibitor application to unveil the possible mechanism of
resistance. Based on the results obtained, it was concluded that both target-site-mutation-based
and metabolism-based ALS resistance mediated by cytochrome P450s exist in the original MCR
population.

iii
ACKNOWLEDGEMENTS
I would like to thank Dr. Patrick Tranel who gave me the opportunity to join the weed
science group at the University of Illinois and work on my Masters’ degree, as well as provided
guidance and supported me throughout the process. I am grateful to have Dr. Dean Riechers and
Dr. Aaron Hager as my committee members, whose valuable suggestions and enlightenment
helped my research tremendously.
Also, the help and assistance I received from many people cannot be neglected, including
my lab mates, both past and present, Chenxi Wu, Dr. Chance Riggins, Laura Chatham, Janel
Huffman, Ahmed Sadequ, Jung Eun Song, Stephanie Rousonelos and Edhilvia Josefina, who not
only created a great atmosphere in the lab but became great friends with me as well. Dr. Adam
Davis, Dr. Danman Zheng, Rong Ma, Nicholas Hausman and many others helped me with lab
experiments, greenhouse research and data analysis, which I am obliged greatly. Without their
help, I would not have gotten to this point.
I also want to express my appreciation to all my friends on campus, former and present,
most of whom are just like me and traveled across the Pacific Ocean for study. I am fortunate to
know so many caring and wonderful people at places far from home, and their combined support
is one of the greatest things I received during the past five years and something that I will always
treasure.
And last but not least, my deepest gratitude goes to my mother, Xiaodong Guo and my
father, Tianwen Guo, for their dedication and unconditional love, and letting me be carefree and
sometimes even indulgent in the pursuit of my own goal. Their support is the ultimate source of
my confidence and courage. Their solicitude and understanding for my absence drove me to
work hard on my research and take good care of myself. I will always keep them in my priority.

iv
TABLE OF CONTENTS
CHAPTER 1 ................................................................................................................................................. 1
Introduction ................................................................................................................................................... 1
1.1 History of ALS Inhibitors ................................................................................................................... 1
1.2 Site of Action of ALS Inhibitors ......................................................................................................... 2
1.3 Mode of Action of ALS inhibitors ...................................................................................................... 3
1.4 Resistance to ALS Inhibitors .............................................................................................................. 5
1.5 Waterhemp Biology ............................................................................................................................ 7
1.6 ALS-Inhibitor-Resistant Populations of Waterhemp .......................................................................... 8
1.7 Research Objective ........................................................................................................................... 10
1.8 Attributions ....................................................................................................................................... 10
1.9 Literature Cited ................................................................................................................................. 11
1.10 Figures............................................................................................................................................. 19
CHAPTER 2 ............................................................................................................................................... 21
Confirmation of Non-target-site Resistance to ALS Inhibitors in a Waterhemp Population ...................... 21
2.1 Abstract ............................................................................................................................................. 21
2.2 Introduction ....................................................................................................................................... 22
2.3 Materials and Methods ...................................................................................................................... 23
2.3.1 Full-length ALS Gene Sequencing ............................................................................................ 23
2.3.2 ALS Enzyme In Vitro Assay ...................................................................................................... 26
2.4 Results and Discussion ..................................................................................................................... 29
2.4.1 Full-length ALS Gene Sequencing ............................................................................................ 29
2.4.2 ALS Enzyme In Vitro Assay ...................................................................................................... 30
2.5 Literature Cited ................................................................................................................................. 31
2.6 Tables and Figures ............................................................................................................................ 33
CHAPTER 3 ............................................................................................................................................... 42
Quantifying Herbicide Resistance in a Waterhemp Population with Non-Target-Site Resistance to
ALS-Inhibiting Herbicides .......................................................................................................................... 42
3.1 Abstract ............................................................................................................................................. 42
3.2 Introduction ....................................................................................................................................... 43
3.3 Materials and Methods ...................................................................................................................... 44

v
3.3.1 Greenhouse Dose Response with ALS-inhibiting Herbicide ..................................................... 44
3.3.2 Response of ALS Inhibitors Combined with Malathion ............................................................ 46
3.4 Results and Discussion ..................................................................................................................... 47
3.4.1 Greenhouse Dose Response with ALS-inhibiting Herbicide ..................................................... 47
3.4.2 Response to ALS Inhibitors Combined with Malathion ............................................................ 48
3.5 Literature Cited ................................................................................................................................. 49
3.6 Tables and Figures ............................................................................................................................ 51
CHAPTER 4 ............................................................................................................................................... 65
Concluding Remarks ................................................................................................................................... 65
4.1 Research Conclusions and Implications ........................................................................................... 65
4.2 Literature Cited ................................................................................................................................. 69
4.3 Tables ................................................................................................................................................ 71

1
CHAPTER 1
Introduction
1.1 History of ALS Inhibitors
The acetolactate synthase (ALS) enzyme, also called acetohydroxyacid synthase (AHAS),
is the target site of five families of herbicides, which are all classified as ALS-inhibiting
herbicides, namely sulfonylureas (SUs) (Ray, 1984), imidazolinones (IMIs) (Shaner et al., 1984),
triazolopyrimidines (TPs) (Gerwick et al., 1990), pyrimidinylthiobenzoates (PTBs) (Takahashi et
al., 1991), and sulfonylamino-carbonyl-triazolinones (SCTs) (Babczinski et al., 1992; Chaleff
and Mauvais, 1984; Corbett and Tardif, 2006; Mallory-Smith and Retzinger, Jr., 2003;). The
various chemicals perform similarly in plants by interfering with amino acid biosynthesis,
specifically, by inhibiting the biosynthesis of branched-chain amino acids (BCAAs) (Ray, 1984).
The first commercialized ALS-inhibiting herbicide was chlorsulfuron, a sulfonylurea, in 1982.
At that time triazines were widely used for weed control, and a variety of weed species had
evolved resistance to them. When compared to triazines and other herbicides at that time, which
were used at rates of kilograms per hectare, the dosage that ALS-inhibiting herbicides used was
significantly less (grams per hectare), which reduced the amount of chemicals in agricultural
greatly. And the specific properties of ALS inhibitors, including low mammalian toxicity,
systematic activity, suitable soil residual activity, and a broad spectrum of weed control,
prompted their wide adoption soon after commercialization (Tranel and Wright, 2002). Currently,
more than 50 ALS-inhibiting herbicide active ingredients are commercialized, including 6 IMIs,
5 PTBs, 3 SCT, 33 SUs, and 3 TPs (Heap, 2013a), with modified functional groups from earlier
inventions for the purpose of altering chemical property, formulation, or weed control spectrum.

2
1.2 Site of Action of ALS Inhibitors
ALS is a crucial enzyme in the biosynthesis pathways of three essential branched-chain
amino acids, valine, leucine and isoleucine, in plants and microorganisms. The ALS enzyme
consists of two subunits. The subunit involved with the catalyzing process has a mass of about
60 to 70 kDa, and the subunit for regulation has mass from 9.5 to 54 kDa (Chipman et al., 2005).
The biosynthesis of the branched-chain amino acids consists of two parallel pathways and ALS
catalyzes the first step of the parallel reactions through condensation of two molecules of
pyruvate to yield 2-acetolactate, or condensation of one molecule of pyruvate and one molecule
of 2-ketobutyrate to yield 2-aceto-hydroxybutyrate (Singh, 1999). During the condensation
reactions, thiamine pyrophosphate (TPP), flavin adenine dinucleotide (FAD) and divalent cation
(Mg2+
or Mn2+
) are required as cofactors. ALS firstly binds TTP and the ALS-TTP complex
reacts with one molecule of pyruvate, to decarboxylate the pyruvate and form a hydroxyethyl-
TPP intermediate. The hydroxyethyl-TPP intermediate then reacts with another molecule of
pyruvate or a molecule of 2-ketobutyrate by nucleophilic attack to yield 2-acetolactate or 2-
aceto-hydroxybutyrate (Singh, 1999). FAD is also required in this reaction and is involved in the
structural stabilization of the protein with the flavin group (Schloss et al., 1988). The
condensation products will continue reacting with other enzymes as precursors of BCAAs in the
pathways.
In plants, ALS is encoded in the nucleus, and targeted to chloroplasts by a transit peptide
(Singh, 1999). ALS gene copy numbers vary among different plant species. For instance,
Arabidopsis thaliana has only one gene (Mazur et al., 1987), while six different ALS genes were
identified in Gossypium hirsutum (Grula et al., 1995). Most of the ALS sequences known have
no introns, and sequence similarity among species is rather high with evidence of divergence

3
between monocots and dicots (Singh, 1999; Uchino and Watanabe, 2002). Despite that amino
acid sequence is quite conserved in ALS enzyme within one species (2 inferred amino acid
polymorphisms in 24 common ragweed (Ambrosia artemisiifolia L.) from IL, MN and OH),
rather high intraspecific variability in nucleotide sequence of the ALS gene has been documented
within some weedy species, particularly among outcrossed species (e.g. common ragweed has
more polymorphic nucleotides than common cocklebur (Xanthium strumarium L.)) (Tranel et al.,
2004).
1.3 Mode of Action of ALS inhibitors
Inhibition of the ALS enzyme impedes the synthesis of the BCAAs, and causes plant
death by depletion of leucine, valine, and isoleucine (Tan et al., 2006). This is the key principle
of all the ALS-inhibiting herbicides. Primarily, the inhibiting herbicide irreversibly binds to the
ALS enzyme in susceptible plants, alters the geometry and structure of its active site and
obstructs its catalytic ability, without affecting other enzymes in the BCAA pathway (Duggleby
and Pang, 2000; Muhitch et al., 1987). After ALS inhibition, mitosis greatly decreases and
eventually stops (Rost and Reynolds, 1985). The biosynthetic rate of protein decreases
correspondingly (Ray, 1984). None of the ALS-inhibitors are regarded as substrate analogs or
cofactors in the ALS enzyme. In other words, ALS herbicides are considered to be non-
competitive inhibitors that do not mimic the substrate of the original biosynthesis process
(Duggleby and Pang, 2000). Because the biosynthesis of BCAAs is mainly in shoot meristems
(where the most active cell growth and division occurs) the phenotypic effect of ALS-inhibiting
herbicides manifests mostly in young tissue, showing as necrosis and malformation of the
meristem (García-Garijo et al., 2012). Also, the ALS inhibitors are mostly in weak acid form

4
after uptake by plants, so they can translocate through phloem and accumulate at the meristem
(Bromilow et al., 1990). Plants die gradually as protein biosynthesis and cell division slow down,
and the free amino acid pool depletes (Shaner and Singh, 1993)
Besides the mechanisms of inhibiting BCAA synthesis, other possible effects of ALS-
inhibiting herbicides contributing to plant death were proposed. However, these effects have
been neither proven to have significant inhibition of plant growth nor concluded to cause plant
death. Research has shown that the accumulation of 2-ketobutyrate and 2-aminobutyrate in the
BCAA synthesis pathway were not the cause of phytotoxicity induced by ALS inhibitors (Shaner
and Singh, 1993). Although the accumulation of phytotoxic compounds is a consequence
induced by chlorsulfuron (Rhodes et al., 1987) and 2-ketobutyrate accumulation induced by
chlorsulfuron is followed by the cessation of plant growth (LaRossa et al., 1987), when reducing
2-ketobutyrate and 2-aminobutyrate accumulation by isoleucine supplementation, the plants
could not recover following imazaquin treatment. In addition, when directly increasing 2-
ketobutyrate and 2-aminobutyrate pools by feeding the plant 2-aminobutyrate, there was no
significant effect on plant growth observed, indicating the accumulation of these intermidiates is
not the cause of plant injury (Shaner and Singh, 1993). The reduction of transportation of
carbohydrates and amino acids is suggested to be a side effect of ALS herbicide inhibition,
which co-occurs with limited utilization of carbohydrates and amino acids in plants treated with
imidazolinone herbicides (Bestman et al., 1990; Gaston et al., 2003). Other possible inhibiting
mechanisms, including induction of high rates of alternative oxidase formation, were proposed as
consequences of sulfonylurea and imidazolinone herbicides, however the actual effects regarding
plant death were unclear (Aubert et al., 1997).

5
1.4 Resistance to ALS Inhibitors
The ALS-inhibiting herbicides cause high selection pressure on weeds; as a result, a wide
variety of weeds evolved resistance to ALS inhibitors. Currently, 132 weed species have been
confirmed resistant to ALS inhibitors throughout the world (Heap, 2013c). The increasing
frequency of resistance limits the further development and use of a large group of herbicides
targeting the same enzyme. The vast majority of resistance cases reported were caused by single
point mutations in the ALS gene. The mutations alter the structure of the ALS enzyme, causing it
to be less sensitive to herbicide binding. Since resistance alleles are dominant (although the level
of dominance varies among species) over susceptible alleles (Sebastian et al., 1989; Wright and
Penner, 1998), herbicide selection also acts on heterozygous individuals. Dominance helps
facilitate spreading of the resistance trait in weed species that propagate by outcrossing. The
mutations in the ALS gene occur commonly and spontaneously in nature, and are utilized in
breeding of ALS-tolerant crops (Tan et al., 2005).
In nature, there are 8 amino acids in ALS known to have substitutions that confer
herbicide resistance in weed populations. They are Ala122, Pro197, Ala205, Asp376, Arg377,
Trp574, Ser653 and Gly654 (Tranel et al., 2013). Following intentional laboratory selection, 17
amino acids were identified to confer resistance to ALS inhibition when substituted (Duggleby
and Pang, 2000; Tranel and Wright, 2002). The mutations at each site are not necessarily
restricted to only one amino acid substitution (Guttieri et al., 1995). For example, nine different
amino acids can substitute for Pro197 in different weed populations that are resistant to ALS
inhibitors (Tranel et al., 2013). The various ALS mutations can confer different patterns of cross
resistance among the ALS inhibitor families. For example, whereas substitution of Trp574

6
confers high resistance across all ALS inhibitors, substitution of Pro197 generally confers high
resistance to SUs but little or no resistance to IMIs (Tranel and Wright, 2002).
Another method whereby plants overcome ALS inhibition is enhanced metabolism that
detoxifies the inhibitors rapidly after uptake. Enhanced detoxification is usually used as the basis
of crop tolerance to ALS inhibitors (Powles and Holtum, 1996). The resistance efficacy of
detoxification usually is less than 10-foldand whereas target-site mutation may confer 100-fold
or higher resistance (Powles and Holtum, 1996). However, a less resistant phenomenon does not
suggest a less severe problem. The enhanced metabolism is not necessarily selected by previous
usage of ALS-inhibiting herbicides, but can arise as cross resistance resulting from selection by
herbicides with other modes of action. For instance, acetyl-CoA carboxylase (ACCase)-
inhibitors are herbicides to which weeds have also evolved both target-site and metabolism-
based resistance. Research showed that after four years of treatment with diclofop-methyl (an
ACCase-inhibitor) in the wheat-growing region of Australia, a biotype of rigid ryegrass (Lolium
rigidum Gaudin) evolved resistance to ACCase-inhibiting herbicides, and cross resistance to
some ALS-inhibiting herbicides (Cotterman and Saari, 1992). In that case, the rigid ryegrass was
resistant to ALS inhibitors even when they have never been exposed to such herbicides. Similar
circumstances were found in a population of large crabgrass (Digitaria sanuinalis L.) that has
cross-resistance to ACCase inhibitors and ALS inhibitors (Hidayat and Preston, 2001). This
causes difficulties in weed control since utilizing alternate side-of-action herbicides is a primary
strategy for herbicide-resistance management (Holtum et al., 1991). Blackgrass (Alopecurus
myosuroides Huds), which is one of the most problematic weeds in Western Europe, has been
reported to have biotypes resistant to ALS inhibitors by enhanced detoxification (Kemp et al.,
1990).

7
Besides the examples of rigid ryegrass, large crabgrass and blackgrass, other weeds also
have been reported to have metabolism-based resistance to ALS-inhibitors. A late watergrass
(Echinochloa phyllopogon (Stapf) Koss) biotype in California was reported to have both
insensitive ALS and enhanced detoxification by cytochrome P450 (Fischer et al., 2000). The first
broadleaf weed reported to have metabolism-based resistance to the ALS inhibitor
ethametsulfuron-methyl was wild mustard (Sinapis arvensis L.) (Veldhuis et al., 2000).
1.5 Waterhemp Biology
Waterhemp [Amaranthus tuberculatus (Moq.) Sauer] is a weed species in the
Amaranthaceae family (Sauer, 1955). It is a small-seeded, summer annual broadleaf weed native
to the Midwest. It is a dioecious species, which means it has both male and female plants that
outcross when reproducing (Steckel, 2007). Waterhemp can grow as tall as 2 meters, and it is a
C4 plant with a relatively high photosynthetic rate, which makes it a very competitive weed for
light, water, and nutrients (Horak and Loughin, 2000; Steckel et al., 2003). In addition, the
ability of a single female waterhemp to produce up to 1 million seeds and its prolonged
germination period make it even more problematic (Steckel and Sprague, 2004a). It has severely
impacted crop production by reducing yield when poorly controlled (Steckel and Sprague,
2004b). The challenge of managing waterhemp has become more difficult as the species
continues to evolve resistance to many commonly used herbicides. Currently, 39 cases of
herbicide-resistant waterhemp have been reported, with all but two found in the Midwest United
States (Heap, 2013b). Herbicides to which waterhemp has evolve resistance include ALS
inhibitors, photosystem II (PSII) inhibitors, protoporphyrinogen oxidase (PPO) inhibitors, p-
hydroxyphenylpyruvate dioxygenase (HPPD) inhibitors, synthetic auxins and glyphosate.

8
Individual waterhemp plants can stack multiple resistant mechanisms together (Bell et al., 2013).
The species also has been reported to outcross with other Amaranthus species, posing even
further challenges to weed management (Wetzel et al., 1999).
1.6 ALS-Inhibitor-Resistant Populations of Waterhemp
Waterhemp evolved resistance to ALS inhibitors soon after their introduction. The first
report waterhemp demonstrated resistance to SU and IMI herbicides (Horak and Peterson, 1995;
Hinz and Owen, 1997; Lovell et al., 1996; Sprague et al., 1997). A combination of ALS inhibitor
and photosystem II (PSII) inhibitor, such as atrazine, could provide adequate control of the ALS-
resistant biotypes (Sprague et al., 1997). However, a waterhemp biotype with resistance to both
ALS inhibitors and atrazine was identified soon thereafter (Foes et al., 1998). A population of
waterhemp from Adams County, Illinois, designated ACR, was characterized as containing
resistance to three different herbicide modes of action, including PPO inhibitors, PSII inhibitors
and ALS inhibitors (Patzoldt et al., 2005). In that population, individual plants displayed
resistance to multiple herbicides. Resistance to ALS inhibitors in ACR was 17,000- to 18,000-
fold compared with a susceptible population, and it showed a cross-resistance to both SU and
IMI herbicides. The resistance mechanism of this population was reported to be a Trp574Leu
substitution in ALS (Patzoldt and Tranel, 2007). The specific mutation can be detected by PCR-
RFLP (restriction fragment length polymorphism) using restriction endonucleases to digest PCR
products of the ALS gene. The sequence with mutation can be recognized and cut, and resulting
in a fragment that is shorter than that observed when the mutation is not present (Corbett and
Tardif, 2006).

9
Waterhemp biotypes with resistance to ALS inhibitors have also been identified with
amino acid substitution (either Asn or Thr) of Ser653 (Patzoldt and Tranel, 2007). In these cases,
the biotypes were resistant particularly to IMI but not SU herbicides. Besides mutations caused
by substitution of Trp574 or Ser653, no other ALS mutations conferring resistance have been
reported in waterhemp. Furthermore, there is not yet any report of waterhemp (or any other
Amaranthus species) being resistant to ALS-inhibitors via rapid herbicide detoxification.
A waterhemp biotype from McLean County, IL, designated MCR, which was
characterized to be resistant to HPPD inhibitors and atrazine (Hausman et al., 2011), also showed
resistance to both SU and IMI herbicides. The field where the population was discovered was not
exposed to ALS inhibiting herbicides for the previous seven years but, nevertheless, plants of the
population were resistant to these herbicides. It was also shown that the resistance level varied
within the population, with some plants being largely unaffected by ALS-inhibitors, and others
being significantly injured but able to recover from normally lethal application rates (Figure 1.1).
By detecting gene mutation with the PCR-RFLP method, it was shown that the highly resistant
plants contained the Trp574Leu ALS mutation, whereas the moderately resistant plants did not
have this mutation (N. Hausman and C. Riggins, personal communication, demonstrated in
Figure 1.2). These observations raised the hypothesis that the MCR waterhemp population
possessed a second and non-target-site resistance mechanism for ALS-inhibition. Since data
indicated that the MCR population was resistant to HPPD inhibitors and atrazine by enhanced
detoxification by cytochrome P450s and glutathione-S-transferase activities, respectively (Ma, et
al., 2013), it was suspected that enhanced herbicide detoxification could also play a role in
resistance to ALS inhibitors.

10
1.7 Research Objective
The objective of this research was to investigate the second resistance mechanism for
ALS-inhibitors present in the MCR waterhemp population. Seeds from waterhemp plants
suspected to demonstrate non-target-site ALS resistance were obtained from a cross in the
greenhouse with two plants from the MCR population. Both plants used for the cross survived a
normally lethal rate of an ALS inhibitor, but were found to lack the Trp574Leu ALS mutation by
PCR-RFLP.
To achieve the objective of this research, full length ALS gene sequencing was
performed to determine if other target-site mutations might be present. An in vitro ALS enzyme
assay was performed to compare the ALS enzyme activities in the presence and absence of an
ALS inhibitor. ALS inhibitors were applied to the population under greenhouse condition to
determine cross resistance among five families of ALS-inhibitors. In addition, malathion, an
inhibitor of cytochrome P450s, was applied together with ALS inhibitors to determine if it could
overcome the resistance mechanism.
1.8 Attributions
The original crossing and genetic marker analysis were performed by Nicholas Hausman
and Chance Riggins. Both laboratory experiments and greenhouse trials described in Chapter 2
and Chapter 3 were conducted by me under the instruction of my graduate committee. The
materials presented in the following chapters will be submitted for publication in Weed Science
in collaboration with Chance W. Riggins, Nicholas E. Hausman, Aaron G. Hager, Dean E.
Riechers and Patrick J. Tranel.

11
1.9 Literature Cited
Aubert, S., Bligny, R., Day, D. A., Whelan, J., and Douce, R. (1997). Induction of alternative
oxidase synthesis by herbicides inhibiting branched-chain amino acid synthesis. The
Plant Journal, 11: 649-657.
Babczinski, P., Müller, K.-H., Santel, H.-J., and Schmidt, R. R. (1992). Patent No. EP 0422469
A3. Germany.
Bell, M. S., Hager, A. G., and Tranel, P. J. (2013). Multiple resistance to herbicides from four
site-of-action groups in waterhemp (Amaranthus tuberculatus). Weed Science, 61: 460-
468.
Bestman, H. D., Devine, M. D., and Vander Born, W. H. (1990). Herbicide chlorsulfuron
decreases assimilate transport out of treated leaves of field pennycress (Thlaspi arvense
L.) seedlings. Plant Physiology, 93: 1441-1448.
Bromilow, R. H., Chamberlain, K., and Evans, A. A. (1990). Physicochemical aspects of phloem
translocation of herbicides. Weed Science, 38: 305-314.
Chaleff, R. S., and Mauvais, C. J. (1984). Acetolactate synthase is the site of action of two
sulfonylurea herbicides in higher plants. Science, 224: 1443-1445.
Chipman, D. M., Duggleby, R. G., and Tittmann, K. (2005). Mechanisms of acetohydroxyacid
synthases. Current Opinion in Chemical Biology, 9: 475-481.
Corbett, C.-A. L., and Tardif, F. J. (2006). Detection of resitance to acetolactate synthase
inhibitors in weeds with enphasis on DNA-based techniques: a review. Pest Management
Science, 62: 584-597.

12
Cotterman, J. C., and Saari, L. L. (1992). Rapid metabolic inactivation is the basis for cross-
resistance to chlorsulfuron in diclofop-methyl-resistance rigid ryegrass (Lolium rigidum)
biotype SR4/84. Pesticide Biochemistry and Physiology, 43: 182-192.
Diebold, R. S., McNaughton, K. E., Lee, E. A., and Tardif, F. J. (2003). Multiple resistance to
imaethapyr and atrazine in powell amaranth (Amaranthus powellii). Weed Science, 51:
312-318.
Doyle, J. J., and Doyle, J. L. (1990). Isolation of plant DNA from fresh tissue. Focus, 12: 13-15.
Duggleby, R. G., and Pang, S. S. (2000). Acetohydroxyacid Synthase. Journal of Biochemistry
and Molecular Biology, 33: 1-36.
Fischer, A. J., Bayer, D. E., Carriere, M. D., Ateh, C. M., and Yim, K.-O. (2000). Mechanisms of
resistance to bispyribac-sodium in an Echinochloa phyllopogon accession. Pesticide
Biochemistry and Physiology, 68: 156-165.
Foes, M. J., Tranel, P. J., Wax, L. M., and Stoller, E. W. (1998). A biotype of common
waterhemp (Amaranthus rudis) resistant to triazine and ALS herbicides. Weed Science,
46: 514-520.
García-Garijo, A., Palma, F., Iribarne, C., Lluch, C., and Tejera, N. A. (2012). Alterations
induced by imazamox on acetohydroxyacid synthase activity of common bean
(Phaseolus vulgaris) depend on leaf position. Pesticide Biochemistry and Physiology,
104: 72-76.
Gaston, S., Ribas-Carbo, M., Busquets, S., Berry, J. A., Zabalza, A., and Royuela, M. (2003).
Changes in mitochondral electron partitioning in response to herbicides inhibiting
branched-chain amino acid biosynthesis in soybean. Plant Physiology, 133: 1351-1359.

13
Gerwick, B. C., Subramanian, M. V., Loney-Gallant, V. I., and Chandler, D. P. (1990).
Mechanism of action of the 1,2,4-triazolo[1,5-a]pyrimidines. Pesticide Science, 29: 357-
364.
Grula, J. W., Hudspeth, R. L., Hobbs, S. L., and Anderson, D. M. (1995). Organization,
inheritance and expression of acetohydroxyacid synthase genes in the cotton
allotetraploid Gossypium hirsutum. Plant Molecular Biology, 28: 837-846.
Guttieri, M. J., Vberlein, C. V., and Thill, D. C. (1995). Diverse mutations in the actolactate
synthase gene confer chlorsulfuron resistance in Kochia (Kochia scoparia) biotypes.
Weed Science, 43: 175-178.
Hausman, N. E., Singh, S., Tranel, P. J., Riechers, D. E., Kaundun, S. S., Polge, N. D., Thomas,
D. A., and Hager, A. G. (2011). Resistance to HPPD-inhibiting herbicides in a population
of waterhemp (Amaranthus tuberculatus) from Illinois, United States. Pest Management
Science, 67: 258-261.
Heap, I. (2013a). ALS inhibitors - HRAC GROUP B. Retrieved September 27, 2013, from The
International Surwey of Herbicide Resistant Weeds: http://www.weedscience.com/
Heap, I. (2013b). Herbicide resistant common waterhemp globally (Amaranthus tuberculatus
(syn. rudis)). Retrieved September 27, 2013, from The International Survey of Herbicide
Resistant Weeds: http://www.weedscience.org/
Heap, I. (2013c). Weeds Resistant to ALS inhibitors (B/2). Retrieved September 26, 2013, from
The International Survey of Herbicide Resistant Weeds: www.weedscience.org
Hidayat, I., and Preston, C. (2001). Cross-resistance to imazethapyr in a fluazifop-p-butyl-
resistant population of Digitaria sanguinalis. Pesticide Biochemistry and Physiology, 71:
190-195.

14
Hinz, J. R., and Owen, M. D. (1997). Acetolactate synthase resistance in a common waterhemp
(Amaranthus rudis) population. Weed Technology, 11: 13-18.
Holtum, J. A., Matthews, J. M., Häusler, R. E., Liljegren, D. R., and Powles, S. B. (1991). Cross-
resistance to herbicides in annual ryegrass (Lolium rigidum). Plant Physiology, 97: 1026-
1034.
Horak, M. J., and Loughin, T. M. (2000). Growth analysis of four Amaranthus species. Weed
Science, 48: 347-355.
Horak, M. J., and Peterson, D. E. (1995). Biotypes of Palmer amaranth (Amaranthus palmeri)
and common waterhemp (Amaranthus rudis) are resistant to imazethapyr and
thifensulfuron. Weed Technology, 9: 192-195.
Kemp, M. S., Moss, S. R., and Thomas, T. H. (1990). Herbicide resistance in Alopecurus
myosuroides. In M. B. Green, H. M. LeBaron, and W. K. Moberg, Managing Resistance
to Agrochemicals From Fundamental Research to Practical Strategies (pp. 376-393).
Washington DC: American Chemical Society.
Knezevic, S. Z., Streibig, J. C., and Ritz, C. (2007). Utilizing R software package for dose-
response studies: theconcept and data analysis. Weed Technology, 21: 840-848.
LaRossa, R. A., Van Dyk, T. K., and Smulski, D. R. (1987). Toxic accumulation of α-
ketobutyrate caused by inhibition of the branched-chain amino acid biosynthetic enzyme
acetolactate synthase in Salmonella typhimurium. Journal of Bacteriology, 169: 1372-
1378.
Lovell, S. T., Wax, L. M., Horak, M. J., and Peterson, D. E. (1996). Imidazolinone and
sulfonylurea resistance in a biotype of common waterhemp (Amaranthus rudis). Weed
Science, 44: 789-794.

15
Ma, R., Kaundun, S. S., Tranel, P. J., Riggins, C. W., McGinness, D. L., Hager, A. G., Hawkes,
T., McIndoe, E., Riechers, D. E. (2013). Distinct detoxification mechanisms confer
resistance to mesotrione and atrazine in a population of waterhemp. Plant Physiology,
163: 363-377.
Mallory-Smith, C. A., and Retzinger, E. J., Jr. (2003). Revised classification of herbicides by site
of action for weed resistance manegement strategies. Weed Technology, 17: 605-619.
Mazur, B. J., Chui, C.-F., and Smith, J. K. (1987). Isolation and characterization of plant genes
coding for acetolactate synthase, the target enzyme for two classes of herbicides. Plant
Physiology, 85: 1110-1117.
McNaughton, K. E., Letarte, J., Lee, E. A., and Tardif, F. J. (2005). Mutations in ALS confer
herbicide resistance in redroot pigweed (Amaranthus retroflexus) and powell amaranth
(Amaranthus powellii). Weed Science, 53: 17-22.
Muhitch, M. J., Shaner, D. L., and Stidham, M. A. (1987). Imidazolinones and acetohydroxyacid
synthase from higher plants. Plant Physiology, 83: 451-456.
Ohkawa, H., Tsujii, H., and Ohkawa, Y. (1999). The use of cytochrome P450 genes to introduce
herbicide tolerance in crops: a review. Pesticide Science, 55: 867-874.
Patzoldt, W. L., and Tranel, P. J. (2007). Multiple ALS mutations confer herbicide resistance in
waterhemp (Amaranthus tuberculatus). Weed Science, 55: 421-428.
Patzoldt, W. L., Tranel, P. J., and Hager, A. G. (2005). A waterhemp (Amaranthus tuberculatus)
biotype with multiple resistance across three herbicide sites of action. Weed Science, 53:
30-36.
Powles, S. B., and Holtum, J. A. (1996). Herbicide resistance in plants: biology and biochemistry.
Boca Raton: Lewis Publishers Inc.

16
Ray, T. B. (1984). Site of action of chlorsulfuron-inhibition of valine and isoleucine biosynthesis
in plants. Plant Physiology, 75: 827-831.
Rhodes, D., Hogan, A. L., Deal, L., Jamieson, G. C., and Haworth, P. (1987). Amino acid
metabolism of Lemna minor L. II. responses to chlorsulfuron. Plant Physiology, 84: 775-
780.
Rost, T. L., and Reynolds, T. (1985). Reversal of chlorsulfuron-induced inhibition of mitotic
entry by isoleucine and valine. Plant Physiology, 77: 481-482.
Sauer, J. (1955). Revision of the dioecious amaranths. Madrono, 13: 5-46.
Schloss, J. V., Ciskanik, L. M., and Van Dyk, D. E. (1988). Origin of the herbicide binding site
of acetolactate synthase. Nature, 331: 360-362.
Sebastian, S. A., Fader, G. M., Ulrich, J. F., Forney, D. R., and Chaleff, R. S. (1989).
Semidominant soybean mutation for resistance to sulfonylurea herbicides. Crop Science,
29: 1403-1408.
Shaner, D. L., and Singh, B. K. (1993). Phytotoxicity of acetohydroxyacid synthase inhibitors is
not due to accumulation of 2-ketobutyrate and/or 2-aminobutyrate. Plant Physiology, 103:
1221-1226.
Shaner, D. L., Anderson, P. C., and Stidham, M. A. (1984). Potent inhibitors of
acetohydroxyacid synthase. Plant Physiology, 76: 545-546.
Siminszky, B. (2006). Plant cytochrome P450-mediated herbicide metabolism. Phytochemistry
Reviews, 5: 445-458.
Singh, B. K. (1999). Biosynthesis of VAline, Leucine, and Isoleucine. In B. K. Singh, Plant
Amino Acids: Biochemistry and Biotechnology (pp. 227-247). New York: Marcel Dekker.

17
Sprague, C. L., Stoller, E. W., and Wax, L. M. (1997). Response of an acetolactate synthase
(ALS)-resistant biotype of Amaranthus rudis to selcted ALS-inhibiting and alternative
herbicides. Weed Research, 37: 93-101.
Steckel, L. E. (2007). The dioecious Amaranthus spp.: here to stay. Weed Technol., 21: 567-570.
Steckel, L. E., and Sprague, C. L. (2004a). Common waterhemp (Amaranthus rudis) interference
in corn. Weed Science, 52: 359-364.
Steckel, L. E., and Sprague, C. L. (2004b). Late-season common waterhemp (Amaranthus rudis)
interference in narrow- and wide-row soybean. Weed Technology, 18: 947-952.
Steckel, L. E., Sprague, C. L., Hager, A. G., Simmons, F. W., and Bollero, G. A. (2003). Effects
of shading on common waterhemp (Amaranthus rudis) growth and development. Weed
Science, 51: 898-903.
Takahashi, S., Shigematsu, S., Morita, A., Nezu, M., Claus, J. S., and Williams, C. S. (1991).
KIH-2031, a new herbicide for cotton. Brighton Crop Protection Conference (pp. 57-62).
Farnham, U.K.: Brighton Crop Protection Council.
Tan, S., Evans, R. R., Dahmer, M. L., Singh, B. K., and Shaner, D. L. (2005). Imidazolinone-
tolerant crops: history, current status, and future. Pest Management Science, 61: 246-257.
Tan, S., Evans, R., and Singh, B. (2006). Herbicidal inhibitors of amino acid biosynthesis and
herbicide-tolerant crops. Amino Acids, 30: 195-204.
Tranel, P. J., and Wright, T. R. (2002). Resistance of weeds to ALS-inhibiting herbicides: what
have we learned? Weed Science, 50: 700-712.
Tranel, P. J., Jiang, W., and Wright, T. R. (2004). Intraspecific variability of the acetolactate
synthase gene. Weed Science, 52: 236-241.

18
Tranel, P. J., Wright, T. R., and Heap, I. M. (2013). Mutations in herbicide-resistant weeds to
ALS inhibitors. Retrieved September 29, 2013, from International Survey of Herbicide
Resistant Weeds: http://www.weedscience.org/.
Uchino, A., and Watanabe, H. (2002). Mutations in the actolactate synthase genes of
sulfonylurea-resistant biotypes of Lindernia spp. Weed Biology and Management, 2: 104-
109.
Veldhuis, L. J., Hall, L. M., O'Donovan, J. T., Dyer, W., and Hall, J. C. (2000). Metabolism-
based resistance of a wild mustard (Sinapis arvensis L.) biotype to ethametsulfuron-
methyl. Journal of Agricultural and Food Chemistry, 48: 2986-2990.
Wetzel, D. K., Skinner, D. Z., and Kulakow, P. A. (1999). Transferal of herbicide resistance
traits from Amaranthus palmeri to Amaranthus rudis. Weed Science, 47: 538-543.
Wright, T. R., and Penner, D. (1998). Corn (Zea mays) acetolate synthase sensitivity to four
classes of ALS-inhibiting herbicides. Weed Science, 46: 8-12.

19
1.10 Figures
Figure 1.1 Responses of MCR waterhemp plants after treatment with ALS inhibitors. Herbicide
active ingredients and rates applied are listed below the picture. There were two distinct
phenotypes in the population; some of the plants showed little to no injury and the rest were
moderately injured by the ALS inhibitors but recovered later.

20
Figure 1.2 Demonstration of PCR-RFLP results of Trp574Leu mutation. ALS alleles with
mutation were recognized and cut by MfeI restriction enzyme and show smaller bands on the
agarose gel while the alleles lacking the specific mutation had larger fragments. Plants showing
little to no injury by ALS inhibitors in Figure 1.1 contain homozygous or heterozygous
Trp574Leu mutation, indicated by orange and blue arrows in the photo, respectively. An
example of a plant from Figure 1.1 showing injury but nevertheless surviving, and not containing
the mutation, is indicated by the white arrow.

21
CHAPTER 2
Confirmation of Non-target-site Resistance to ALS Inhibitors in a Waterhemp Population
2.1 Abstract
The acetolactate synthase enzyme is the target site of ALS-inhibiting herbicides, and a
variety of weed species have developed resistance to these herbicides over the past few decades.
The predominant resistance mechanism is gene mutation, with a few distinctive cases of
enhanced herbicide metabolism. Waterhemp [Amaranthus tuberculatus (Moq.) Sauer], a
common weed in Midwest United States has been reported to have ALS resistance. Researches
have confirmed single mutations on waterhemp ALS gene cause high level resistance (Chapter
1). The population found in McLean County, IL (MCR) has been reported resistant to both
HPPD inhibitors and atrazine, and demonstrating two phenotypes after ALS-inhibiting herbicide
application. The plants showing high resistance have Trp574Leu mutation, while plants with the
other phenotype do not contain this specific mutation. In order to confirm the possible resistance
mechanism of the moderate resistant individuals, a progeny (named JG11) was made by crossing
two moderate resistant plants without Trp574Leu mutation to be used in the study. The portion
of the ALS gene encoding the mature ALS protein was sequenced from eight JG11 plants that
survived a normally lethal dose of ALS inhibitor to detect other possible gene mutations that
might confer herbicide resistance. In vitro protein assays were performed to compare the
sensitivity of ALS enzyme of JG11 with that of both resistant and susceptible populations. None
of the deduced amino acid sequences obtained contained resistance-related mutations, and the
protein assay results indicated the JG11 plants did not have insensitive enzyme that confers

22
resistance. Both sets of experiments indicate that a non-target-site resistance mechanism exists in
the JG11 population.
2.2 Introduction
ALS-inhibiting herbicides have selected resistant populations of various weed species.
Most of the resistance cases contain single mutations in ALS gene that make the target site
insensitive to herbicide binding. Waterhemp is a common weed species in the Amaranthaceae
family (Sauer, 1955). It has evolved resistance to herbicides from numerous modes of action
including ALS inhibitors (Heap, 2013b). There are three different target-site amino acids
substitutions found in waterhemp populations resistant to ALS inhibitors: Trp574Leu,
Ser653Asp, and Ser653Thr. Waterhemp populations containing the Trp574Leu mutation have
shown cross resistance to all ALS-inhibitor families, while Ser653Asp and Ser653Thr mutation
confer an IMI-specific resistance pattern (Patzoldt and Tranel, 2007; Tranel and Wright, 2002).
Another resistance mechanism of enhanced metabolism was found in several weed species,
which was discussed in Chapter 1. There is no waterhemp population that has been reported with
enhanced metabolism as a mechanism to overcome ALS-inhibitors; however, a waterhemp
biotype from McLean County, IL, designated MCR, has metabolism-based HPPD and atrazine
resistance (Hausman, et al., 2011; Ma, et al., 2013). Without pre-selecting the population with
ALS-inhibitors in the crop field at least in the past 7 years, this population had shown both high
and moderate resistance to herbicides from the SU and IMI families (Chapter 1.6). Since
Trp574Leu mutation was not found in some of the resistant individuals that nevertheless were
able to survive labeled rates of either SU or IMI herbicides, two such individuals were crossed to
create a population for further analysis. Progeny of this cross were designated as JG11.

23
The objective of the study was to test the hypothesis that the JG11 population does not
contain a target-site-mutation that confers ALS-resistance. To address this objective, ALS gene
encompassing the coding sequence of the mature protein was sequenced to detect any amino acid
substitution that may confer resistance to ALS inhibitors. An in vitro ALS assay was also
conducted to determine if ALS enzyme in the JG11 population was sensitive to an ALS inhibitor.
2.3 Materials and Methods
2.3.1 Full-length ALS Gene Sequencing
2.3.1.1 Plant Material and Greenhouse Cultivation
In addition to JG11 (described in Introduction), two additional waterhemp populations,
ACR and WCS (described in Patzoldt et al., 2005) were used as controls. Population ACR
contains the ALS Trp574Leu substitution and was used as a positive control, while population
WCS is sensitive to ALS inhibitors and was used as a negative control. All seeds were stratified
before planting. Prior to stratification, seeds were soaked 10 min in a 1:1 mixture of commercial
bleach and water, and then rinsed with sterile water. Seeds were stratified in a solution of water
and 0.1% (w/v) agarose for at least one month at 4 C. Seeds were sown in plastic inserts
containing commercial potting mix (LC1 Professional Growing Mix, Sun Gro Horticulture, Inc.,
110 110th Ave. NE, Suite 490, Bellevue, WA 98004). Seedlings were transplanted to individual
inserts with the same potting mix when displaying one to two true leaves. A second transplant,
when the seedlings were about 5 cm tall moved them into 11-cm square pots filled with 3 : 1 : 1 :
1 mixture of commercial potting mix : sand : soil : peat. A slow-release fertilizer (Osmocote 13-
13-13 slow release fertilizer or Osmocote Plus 15-9-12 water soluble fertilizer. The Scotts
Company, 14111 Scottlawn Rd., Marysville, Oh 43041) was mixed in with the growing medium.

24
All plants were grown in a greenhouse at the University of Illinois in Urbana-Champaign campus,
and the environmental conditions were set to 28/22 C day/night temperature and 16 h
photoperiod. Sunlight was supplemented with halide and sodium vapor lights to maintain a
minimum of 313 µmol m-2
sec-1
during the diurnal period.
2.3.1.2 Herbicide Screening
Primisulfuron (Beacon, Syngenta Crop Protection) was applied to JG11 plants about 7
days after the second transplanting when the plants were 10-12 cm tall. The application rates
were 16.9 or 53.3 g a.i. per ha with 1% crop oil concentrate (COC, Herbimax®, Loveland
Products , Inc., P.O. Box 1286, Greeley, Co 80632) and 2.5% ammonium sulfate (AMS, N-Pak
AMS Liquid, Winfield Solutions, LLC, P.O. Box 64589, St. Paul, MN 55164-0589). Herbicide
treatments were performed in a greenhouse spraying chamber (Generation III Research Sprayer.
DeVries Manufacturing, 28081 870th Ave, Hollandale, MN 56045) with a flat-fan nozzle
(TeeJet 80015EVS. TeeJet Technologies, P.O. Box 7900, Wheaton, IL 60187) that delivered
75.7 liter min-1
. The nozzle was about 45 cm above the plant canopy when spraying. WCS plants
of similar size were included in the herbicide application to verify herbicide efficacy.
2.3.1.3 DNA Extraction and Purification
Leaf samples were collected during the second week after herbicide application, at which
time plants from the JG11 population had recovered from the herbicide treatment and were
developing new leaves. Only newly growing small leaflets were collected and put into 1.5 ml
tubes individually. The tubes were kept on ice until DNA extraction.

25
The DNA extraction and purification procedure was modified from a protocol of Doyle
and Doyle (1990) using hexadecyltrimethyl-ammonium bromide (CTAB). Extracted DNA was
resuspended in deionized sterile water and quantified using a Nanodrop 1000 Spectrophotometer
(Thermo Fisher Scientific, Inc., 81 Wyman St., Waltham, MA 02454). The DNA samples were
stored at -20 C until use for sequencing.
2.3.1.4 PCR Amplification and Sequencing
Polymerase chain reaction (PCR) was applied to amplify the ALS sequences. Because the
length of the ALS gene is over 2000 base pairs, several sets of primers were used to amplify
overlapping sections of the sequence (Table 2.1). Each PCR contained 1.0 µL of DNA samples,
0.2 µL GoTaq DNA polymerase (5 U/ µL. Promega Corporation, 2800 Woods Hollow Rd,
Madison, WI 53711), 1.0 µL each of forward and reverse primers (10 µM), 2.0 µL dNTP (2.5
mM), 2.5 µL MgCl2 solution (25 mM), 5 µL 5X GoTaq buffer, and 12.3 µL purified water to
make a total 25 µL volume. The amplification took place in a PTC-100 thermocycler (MJ
Research, Inc., 590 Lincoln Street, Waltham, MA, 02451) programed for an initial denaturation
of 1 min at 95 C, then 36 cycles of denaturation at 94 C for 15 sec, annealing at 50 C for 15 sec,
and extension at 60 C for 4 min. After a final elongation at 60 C for 4 min, the samples were held
at 4 C. PCR products were separated by electrophoreses through a 1% agarose gel (Certified
Molecular Biology Agarose, Bio-Rad Laboratories, 1000 Alfred Nobel Drive, Hercules, CA
94547) and visualized alongside Quick-Load® 100 bp ladder (New England Biolabs, Inc., 240
County Road, Ipswich, MA, 01938-2723) by ethidium bromide-staining/UV light. PCR products
of the expected sizes were purified with a Cyclic Purification Kit (Omega Bio-Tek, Inc., 400
Pinnacle Way Suite, 450 Norcross, GA 30071).

26
Purified DNA fragments were sequenced using an ABI Prism BigDye® TerminatorTM
v3.1 Cycle Sequencing Kit (Applied Biosystems, Inc., 850 Lincoln Centre Drive, Foster City,
CA 94404). The same primers used for PCR were used individually to sequence each DNA
strand. Each reaction contained 2.0 µL DNA template, 1.0 µL BigDye®, 2.0 µL primer, 2.0 µL
5X buffer containing 10 mM MgCl2 and 400 nM Tris base, 5.2 µL 12.5% glycerol, and 1.8 µL
sterile water to make a total of 14 µL reaction volume. The reaction was performed in a PTC-100
thermocycler, with an initial denaturation of 95 C for 3 min followed by 35 cycles of
denaturation of 95 C for 1 min, annealing at 56 C for 1 min, and extension at 72 for 1 min. After
a final extension of 5 min at 72 C the samples were kept at 4 C. Samples were analyzed by the
W.M. Keck Center for Comparative and Functional Genomics High Throughput Sequencing Lab
(University of Illinois Biotechnology Venter, 340 Edward R. Madigan Laboratory, 1201 W.
Gregory Drive, Urbana, IL 61801) using an ABI 3730xl DNA Analyzer. The returned data were
compared and aligned with FinchTV software v1.4 (Geospiza, Inc., 100 West Harrison, North
Tower, Suite #330, Seattle, WA 98119) and ClustalW package in FinchTV.
2.3.2 ALS Enzyme In Vitro Assay
2.3.2.1 Plant Material
Seeds of ACR, WCS and JG11 were stratified and sown as described in 2.3.1.2. Leaf
tissue was collected from plants when they were about 15 to 20 cm tall. New leaves from plant
apexes were selected. Collected leaf samples were put on ice and used for the ALS enzyme
extraction process immediately after collection.

27
2.3.2.2 Protein Extraction and Desalinization
The protein extraction procedure used was modified from Zheng et al. (2005). About 6.0
g of fresh leaf tissue (pooled from 10 to 15 plants) were frozen with liquid nitrogen and grinded
into powder, then homogenized with 35 ml homogenization buffer of 100 mM potassium
phosphate (pH 7.0), 5 mM pyruvate, 5 mM MgCl2, 1 mM thiamine pyrophosphate (TPP), 10 µM
flavin adenine dinucleotide (FAD), 1 mM dithiothreitol (DTT), 10% glycerol (v/v), and 1%
polyvinylpolypyrrolidone (PVPP) (w/v). The homogenate was centrifuged (Beckman J2-HS
Centrifuge. Beckman Coulter, Inc., Diagnostics Division Headquarters, 250 South Kraemer
Boulevard, Brea CA 92821-6232) at 20,000 × g for 20 min at 4 C and the supernatant was
transferred to another centrifuge tube. Ammonium sulfate was added to create a 45% solution
(w/v), which was then gently shaken on ice for 45 min to precipitate protein containing ALS
enzyme. The precipitate was collected by centrifugation at 20,000 × g for 20 min at 4 C. After
discarding the supernatant, the pellet was resuspended in 2.5 ml of resuspension buffer,
consisting of 50 mM potassium phosphate (pH 7.0), 5 mM MgCl2, 1 mM TPP, and 10 µM FAD.
Disposable PD-10 Desalting Columns (GE Healthcare Bio-Sciences, 800 Centennial Avenue,
P.O. Box 1327, Piscataway, NJ 08855-1327) were used at 4 C to purify and desalt the protein.
Protein concentrations of the extracts were determined with a Nanodrop 1000 Spectrophotometer
v3.7.1 using Coomassie Plus (Bradford) Assay Kit (Thermo ScientificTM
PierceTM
. Thermo
Fisher Scientific, 81 Wyman St., Waltham, MA 02454). Protein concentrations ranged from 1 to
1.5 µg/µl. The protein samples were frozen with liquid nitrogen and stored at -80 C until needed.

28
2.3.2.3 ALS Enzymatic Assay
Corning 96-well assay flat clear bottom plates (Corning, Inc., One Riverfront Plaza,
Corning, NY 14831) were used in protein assay. Technical-grade imazethapyr (98.1%, BASF
Corporation, 26 David Drive, Research Triangle Park, NC 27709) was used to determine enzyme
activity under herbicide interference. Each reaction contained 50 µl of protein extract (diluted to
1.0 µg/µl), 50 µl of reaction buffer of 50 mM potassium phosphate buffer (pH 7.0), 100mM
pyruvate, 5 mM MgCl2, 1 mM TPP, 10 µM FAD and 50 µl of herbicide solution. Imazethapyr
was dissolved in 9% dimethylsulfoxide (DMSO, v/v) solution to concentrations ranging from 10-
1 to 10
6 nM in 10-fold increments. For the positive controls and background readings, 50 µl of 9%
DMSO were added as solvent in substitution of herbicide solution. Negative controls without
herbicide were created by adding 25 µl 3.5% H2SO4 before addition of protein extract. The
enzyme assay mixtures were incubated at 37 C for 90 min and terminated by adding 25 µl 3.5%
H2SO4 and incubating 20 min at 60 C. The amount of acetoin formed was determined by a
chromogenic reaction with 100 µl of a mixture of 0.55% (w/v) creatine and 5.5% (w/v) α-
naphthol in 1.375 N NaOH added to each enzyme assay, followed by 40 min incubation at 37 C.
Absorbance was measured by a spectrophotometer at a wavelength of 530 nm.
2.3.2.4 Data Analysis
Two separate extracts were obtained from each population for two independent assays,
and each assay contained three replications of each treatment (treated statistically as subsamples).
The data of absorbance were analyzed using a non-linear regression model with the dose-
response curve package in R software (Knezevic et al, 2007). The equation
{ [ ( ) ( )]}

29
was used to construct the dose-response curves of enzymatic assays. The four-parameter non-
linear logistic model is described as: b is the slope of the curve, c is the lower limit, d is the
upper limit and ED50 is 50% reduction in acetoin accumulation.
2.4 Results and Discussion
2.4.1 Full-length ALS Gene Sequencing
A portion of the ALS gene encompassing the coding region of the mature protein was
sequenced from each of eight JG11 plants that survived primisulfuron at either 16.9 or 53.3 g per
ha. The same herbicide rates were lethal to WCS plants and all the WCS plants sprayed
simultaneously with JG11 were dead when collecting tissue samples for DNA extraction (data
not shown). Additional waterhemp ALS gene sequences from WCS, ACR and an imidazolinone-
resistant population IR-101, described by Paltzoldt and Tranel (2007), were obtained from
GenBank (accessions EF157818, EF157819 and EF157821, respectively) for comparison. The
mature protein sequences were aligned and compared (Figure 2.1). ACR and IR-101 contain
resistance-conferring mutations at W574 and S653, respectively (Patzoldt, et al., 2005; Patzoldt
and Tranel, 2007), but these mutations were not identified in JG11 plants. The majority of
sequence polymorphisms identified (23 out of 29 codons) were synonymous substitutions.
Across all JG11 protein sequences obtained, six nonsynonymous polymorphisms were identified
relative to the WCS population. However, these same polymorphisms were shared with ACR
and/or IR-101 and, therefore, not likely to be associated with resistance. In addition, these
polymorphic amino acid substitutions codons were observed in other weed species as well, in
both resistant and susceptible biotypes, suggesting they are in non-conserved regions of ALS and

30
not associated with herbicide resistance (Diebold, et al., 2003; McNaughton, et al., 2005;
Patzoldt and Tranel, 2007).
2.4.2 ALS Enzyme In Vitro Assay
The reaction catalyzed by ALS in vitro is the condensation of two molecules of pyruvate
to yield one molecule of acetolactate. Since the next enzyme of the BCAA synthesis pathway,
ketoacid reductoisomerase (KARI) is not present in the extracts, the acetolactate accumulates,
and can then be decarboxylated into acetoin, which yields a red color after the chromogenic
reaction (Corbett & Tardif, 2006). Decreasing color intensity, which is the indication of enzyme
inhibition, can be seen with increasing herbicide concentration (
Figure 2.2).
The combination of two runs of enzymatic assay results is shown in Figure 2.3. The
positive (ACR) and negative (WCS) controls behaved as expected with the former being quite
insensitive to herbicide inhibition and the latter greatly inhibited with increasing herbicide
concentration. The ED50 (effective dose causing 50% reduction of action accumulation) for ALS
extracted from WCS was 1642 nM imazethapyr, whereas even the highest dose tested (3.3 × 106
nM imazethapyr) failed to reduce ACR ALS enzyme activity by 50%. The dose response curve
for ALS from JG11 was very similar to that from WCS, and the calculated ED50 value (1559 nM
imazethapyr) was indistinguishable (p=0.9) from that obtained from WCS. Therefore, it was
concluded that ALS in the JG11 population is fully sensitive to ALS herbicides.
The results of both gene sequencing and enzymatic assay provide evidence that the
observed whole plant resistance to ALS-inhibitors in JG11 is not based on an insensitive target

31
site. More research is needed to determine the actual resistance level of the JG11 population as
well as the alternative mechanism of resistance that is occurring in this population.
2.5 Literature Cited
Corbett, C.-A. L., and Tardif, F. J. (2006). Detection of resitance to acetolactate synthase
inhibitors in weeds with enphasis on DNA-based techniques: a review. Pest Management
Science, 62: 584-597.
Diebold, R. S., McNaughton, K. E., Lee, E. A., and Tardif, F. J. (2003). Multiple resistance to
imaethapyr and atrazine in powell amaranth (Amaranthus powellii). Weed Science, 51:
312-318.
Doyle, J. J., and Doyle, J. L. (1990). Isolation of plant DNA from fresh tissue. Focus, 12: 13-15.
Hausman, N. E., Singh, S., Tranel, P. J., Riechers, D. E., Kaundun, S. S., Polge, N. D., Thomas,
D. A., and Hager, A. G. (2011). Resistance to HPPD-inhibiting herbicides in a population
of waterhemp (Amaranthus tuberculatus) from Illinois, United States. Pest Management
Science, 67: 258-261.
Heap, I. (2013b). Herbicide resistant common waterhemp globally (Amaranthus tuberculatus
(syn. rudis)). Retrieved September 27, 2013, from The International Survey of Herbicide
Resistant Weeds: http://www.weedscience.org/
Ma, R., Kaundun, S. S., Tranel, P. J., Riggins, C. W., McGinness, D. L., Hager, A. G., Hawkes,
T., McIndoe, E. and Riechers, D. E. (2013). Distinct detoxification mechanisms confer
resistance to mesotrione and atrazine in a population of waterhemp. Plant Physiology,
163: 363-377.

32
McNaughton, K. E., Letarte, J., Lee, E. A., and Tardif, F. J. (2005). Mutations in ALS confer
herbicide resistance in redroot pigweed (Amaranthus retroflexus) and powell amaranth
(Amaranthus powellii). Weed Science, 53: 17-22.
Patzoldt, W. L., Tranel, P. J., and Hager, A. G. (2005). A waterhemp (Amaranthus tuberculatus)
biotype with multiple resistance across three herbicide sites of action. Weed Science, 53:
30-36.
Patzoldt, W. L., and Tranel, P. J. (2007). Multiple ALS mutations confer herbicide resistance in
waterhemp (Amaranthus tuberculatus). Weed Science, 55: 421-428.
Sauer, J. (1955). Revision of the dioecious amaranths. Madrono, 13: 5-46.
Tranel, P. J., and Wright, T. R. (2002). Resistance of weeds to ALS-inhibiting herbicides: what
have we learned? Weed Science, 50: 700-712.
Zheng, D., Patzoldt, W. L., and Tranel, P. J. (2005). Association of the W574L ALS substitution
with resistance to cloransulam and imazamox in common ragweed (Ambrosia
artemisiifolia). Weed Science, 53: 424-430.

33
2.6 Tables and Figures
Table 2.1 Primers used in ALS gene amplification.
Name Sequence (5’ → 3’) Tm (C) MW
ALS5UTR-F CTT CAA TCT TCA ACA ATG GCG 52.5 6365.2
WHals-F CGC CCT CTT CAA ATC TCA TC 53.3 5947.9
ALSr1 TCA ATC AAA ACA GGT CCA GG 52.7 6119.0
ALSf1 AGC TCT TGA ACG TGA AGG TG 55.1 6197.1
ALS1603-R AAC TCC CAT CCC CAT CAA TGT C 57 6559.3
ALS1530-F TTT GGG GGC TAT GGG GTT TG 57.9 6266.1
AmALS-F2 TCC CGG TTA AAA TCA TGC TC 52.8 6052.0
AmALS-R2 CTA AAC GAG AGA ACG GCC AG 55.4 6169.1
ALS1426-f ACG AAG GGT GAT GCG ATT GT 57 6237.1

34
Table 2.2 Output from statistical analysis of enzymatic assay.
Estimate Std. Error Lower Upper
JG11:50 1558.746 545.220 477.046 2640.45
WCS:50 1641.904 638.409 375.318 2908.49
Estimate Std. Error t-value p-value
JG11/WCS 0.94935 0.49651 -0.10201 0.919

35
Figure 2.1 ALS codon alignments of WCS, ACR, IR-101 and JG11 populations. Amino acid sequences of 8 individual plants of JG11
population are listed. Protein sequence of WCS (S-EF157818) is used as standard for comparison. Positions marked with “X” indicate
the identification of synonymous nucleotide polymorphism, and heterozygous substitutions are in regular font while homozygous
substitutions are in bold font. Nonsynonymous polymorphisms are shown as the amino acids corresponding to each codon, with
heterozygous substitutions in regular font and homozygous substitutions in bold font.
WCS P D E P R K G C D V L V E A L E R E G V T D V F A Y P G G A S M E I H Q A L T R S N I I R N V L P R
ACR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
IR-101 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1104 full . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1105 full . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1106 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1107 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1108 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1109 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1110 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1111 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
WCS H E Q G G V F A A E G Y A R A T G R V G V C I A T S G P G A T N L V S G F A D A L L D S V P L V A I
ACR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
IR-101 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1104 full . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1105 full . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1106 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1107 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1108 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1109 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1110 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1111 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
89 99 109 119 129
139 149 159 169 179

Figure 2.1 (cont.)
36
WCS T G Q V P R R M I G T D A F Q E T P I V E V T R S I T K H N Y L V L D V E D I P R I V K E A F F L A
ACR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
IR-101 . . . . . X . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1104 full . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1105 full . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . . . . .
1106 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . . . . .
1107 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . . . . .
1108 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1109 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1110 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1111 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
WCS N S G R P G P V L I D I P K D I Q Q Q L V V P N W E Q P I K L G G Y L S R L P K P T F S A N E E G L
ACR . . . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
IR-101 . . . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1104 full . . . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1105 full . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . . .
1106 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . . .
1107 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . . .
1108 . . . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1109 . . . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1110 . . . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1111 . . . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
WCS L D Q I V R L V G E S K R P V L Y T G G G C L N S S E E L R K F V K L T G I P V A S T L M G L G A F
ACR . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . E . . . . . . . . . . . . . . . .
IR-101 . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . E . . . . . . . . . . . . . . . .
1104 full . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . E . . . . . . . . . . . . . . . .
1105 full . . . . . . . . . . . . . . . . . . . . . . . . X . . . X . . . . E . . . . . . . . . . . . . . . .
1106 . . . . . . . . . . . . . . . . . . . . . . . . X . . . X . . . . E . . . . . . . . . . . . . . . .
1107 . . . . . . . . . . . . . . . . . . . . . . . . X . . . X . . . . E . . . . . . . . . . . . . . . .
1108 . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . E . . . . . . . . . . . . . . . .
1109 . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . E . . . . . . . . . . . . . . . .
1110 . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . E . . . . . . . . . . . . . . . .
1111 . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . E . . . . . . . . . . . . . . . .
189 199 219 229
249 259 269
209
289 299 309 319 329
239 279

Figure 2.1 (cont.)
37
WCS P C T D D L S L Q M L G M H G T V Y A N Y A V D K A D L L L A F G V R F D D R V T G K L E A F A S R
ACR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . . . . X
IR-101 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . . . . X
1104 full . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . X . . . . . . . . . X
1105 full . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . X . . . . . . . . X X
1106 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . X . . . . . . . . X X
1107 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . X . . . . . . . . X X
1108 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . X . . . . . . . . . X
1109 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . X . . . . . . . . . X
1110 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . X . . . . . . . . . X
1111 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . X . . . . . . . . . X
WCS A K I V H I D I D S A E I G K N K Q P H V S I C G D V K V A L R G L N N I L E S R K G K V K L D F S
ACR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
IR-101 . . . . . . . . . . . . . . . . . . . . . . . . . . I . . . . Q . . . K . . . . . . . . L . . . . .
1104 full . . . . . . . . . . . . . . . . . . . . . . . . . . I . . . . Q . . . K . . . . . . . . L . . . . .
1105 full . . . . . . . . . . . . . . . . . . . . . . . . . . I . . . . Q . . . K . . . . . . . . L . . . . .
1106 . . . . . . . . . . . . . . . . . . . . . . . . . . I . . . . Q . . . K . . . . . . . . L . . . . .
1107 . . . . . . . . . . . . . . . . . . . . . . . . . . I . . . . Q . . . K . . . . . . . . L . . . . .
1108 . . . . . . . . . . . . . . . . . . . . . . . . . . I . . . . Q . . . K . . . . . . . . L . . . . .
1109 . . . . . . . . . . . . . . . . . . . . . . . . . . I . . . . Q . . . K . . . . . . . . L . . . . .
1110 . . . . . . . . . . . . . . . . . . . . . . . . . . I . . . . Q . . . K . . . . . . . . L . . . . .
1111 . . . . . . . . . . . . . . . . . . . . . . . . . . I . . . . Q . . . K . . . . . . . . L . . . . .
WCS N W R E E L N E Q K K K F P L S F K T F G D A I P P Q Y A I Q V L D E L T K G D A I V S T G V G Q H
ACR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . .
IR-101 . . . . X . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . .
1104 full . . . . X . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . X . . . . . . . . . . . . . .
1105 full . . . . X . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . X . . . . . . . . . . . . . .
1106 . . . . X . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . X . . . . . . . . . . . . . .
1107 . . . . X . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . X . . . . . . . . . . . . . .
1108 . . . . X . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . X . . . . . . . . . . . . . .
1109 . . . . X . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . X . . . . . . . . . . . . . .
1110 . . . . X . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . X . . . . . . . . . . . . . .
1111 . . . . X . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . X . . . . . . . . . . . . . .
379
479
339 349 359 369
389 399 409 419 429
439 449 459 469

Figure 2.1 (cont.)
38
WCS Q M W A A Q F Y K Y R N P R Q W L T S G G L G A M G F G L P A A I G A A V A R P D A V V V D I D G D
ACR . . . . . . X . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
IR-101 . . . . . . X . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1104 full . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . X . . . . . . . X
1105 full . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . X . . . . . . . X
1106 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . . X
1107 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . . X
1108 . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . X . . . . . . . X
1109 . . . . . . . . . . . . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . X . . . . . . . X
1110 . . . . . . X . . . . . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . X . . . . . . . X
1111 . . . . . . X . . . . . . . . . . . . . . . . . . . . X . . . . . . . . . . . . . X . . . . . . . X
WCS G S F I M N V Q E L A T I R V E N L P V K I M L L N N Q H L G M V V Q W E D R F Y K A N R A H T Y L
ACR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . L . . . . . . . . . . . . . .
IR-101 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1104 full . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1105 full . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1106 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1107 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1108 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1109 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1110 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1111 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
WCS G N P S N S S E I F P D M L K F A E A C D I P A A R V T K V S D L R A A I Q T M L D T P G P Y L L D
ACR . . X . . . X . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
IR-101 . . . . K . . . . . . . . X . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1104 full . . . . K . . . . . . . . X . . . . . . . . . . . . . X . . . . . . . . . . . . . . . . . . . . . .
1105 full . . . . K . . . . . . . . X . . . . . . . . . . . . . X . . . . . . . . . . . . . . . . . . . . . .
1106 . . X . K . X . . . X . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1107 . . X . K . X . . . X . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1108 . . . . K . . . . . . . . X . . . . . . . . . . . . . X . . . . . . . . . . . . . . . . . . . . . .
1109 . . . . K . . . . . . . . X . . . . . . . . . . . . . X . . . . . . . . . . . . . . . . . . . . . .
1110 . . . . K . . . . . . . . X . . . . . . . . . . . . . X . . . . . . . . . . . . . . . . . . . . . .
1111 . . . . K . . . . . . . . X . . . . . . . . . . . . . X . . . . . . . . . . . . . . . . . . . . . .
569
629
489 499 519 529
549 559
509
539
589
579
599 609 619

Figure 2.1 (cont.)
39
WCS V I V P H Q E H V L P M I P S G A A F K D T I T E G D G R R A Y *
ACR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
IR-101 . . . . . . . . . . . . . . T . . . . . . . . . . . . . . . . . .
1104 full . . . . . . . . . . . . . . X . . . . . X . . . . . . . . . . . .
1105 full . . . . . . . . . . . . . . X . . . . . X . . . . . . . . . . . .
1106 . . . . . . . . . . . . . . X . . . . . X . . . . . . . . . . . .
1107 . . . . . . . . . . . . . . X . . . . . X . . . . . . . . . . . .
1108 . . . . . . . . . . . . . . X . . . . . X . . . . . . . . . . . .
1109 . . . . . . . . . . . . . . X . . . . . X . . . . . . . . . . . .
1110 . . . . . . . . . . . . . . X . . . . . X . . . . . . . . . . . .
1111 . . . . . . . . . . . . . . X . . . . . . . . . . . . . . . . . .
669649 659639

40
1 2 3 4 5 6 7 8 9 10 11 12
A ACR
106
ACR
106
ACR
106
JG11
106
JG11
106
JG11
106
WCS
106
WCS
106
WCS
106
ACR
DMSO
ACR
DMSO
ACR
DMSO
B ACR 105
ACR 105
ACR 105
JG11 105
JG11 105
JG11 105
WCS 105
WCS 105
WCS 105
JG11 DMSO
JG11 DMSO
JG11 DMSO
C ACR
104
ACR
104
ACR
104
JG11
104
JG11
104
JG11
104
WCS
104
WCS
104
WCS
104
WCS
DMSO
WCS
DMSO
WCS
DMSO
D ACR 103
ACR 103
ACR 103
JG11 103
JG11 103
JG11 103
WCS 103
WCS 103
WCS 103
ACR H2SO4
ACR H2SO4
ACR H2SO4
E ACR
102
ACR
102
ACR
102
JG11
102
JG11
102
JG11
102
WCS
102
WCS
102
WCS
102
JG11
H2SO4
JG11
H2SO4
JG11
H2SO4
F ACR
10 ACR
10 ACR
10 JG11
10 JG11
10 JG11
10 WCS
10 WCS
10 WCS
10 WCS H2SO4
WCS H2SO4
WCS H2SO4
G ACR
1
ACR
1
ACR
1
JG11
1
JG11
1
JG11
1
WCS
1
WCS
1
WCS
1
DMSO
DMSO
DMSO
H ACR 0.1
ACR 0.1
ACR 0.1
JG11 0.1
JG11 0.1
JG11 0.1
WCS 0.1
WCS 0.1
WCS 0.1
DMSO
DMSO
DMSO
Figure 2.2 ALS enzymatic assay with imazethapyr as inhibitor and the plate arrangement is
shown below. In the bottom panel, the orange area shows the treatments, the blue area contained
no herbicide and was used as positive control, the green area was used as negative control by
adding H2SO4 before adding enzyme extracts, and the gray area contained only reaction buffer
and DMSO to obtain background readings. The photo was taken after the chromatic reaction, and
the red color indicates the accumulation of acetoin, which reflects ALS activity.

41
Figure 2.3 Dose response curves of enzymatic assay. X-axis is the herbicide concentration (nM)
and Y-axis is ALS activity expressed relative to the no-herbicide control.
Imazethapyr (nM)
En
zym
e a
ctiv
ity (
rela
tiv
e to
no
-her
bic
ide
con
tro
l)

42
CHAPTER 3
Quantifying Herbicide Resistance in a Waterhemp Population with Non-Target-Site Resistance to
ALS-Inhibiting Herbicides
3.1 Abstract
Previous studies reported several ALS-resistance waterhemp biotypes. In all cases,
resistance was found to result from amino acid substitutions that alter the herbicide-binding site
(Chapter 1). In Chapter 2, both gene sequencing and protein assay showed the resistance of JG11,
a progeny derived from the MCR population, is not caused by the same mechanism, because no
mutations in ALS gene that may affect herbicide binding were found, nor was an insensitive
ALS enzyme observed in the JG11 population. In initial trials of herbicide application, the JG11
plants showed minor damage, which is different from the symptom of target-site mediated
resistant waterhemp biotypes. In addition, ALS inhibitors comprise five different chemical
families, so the focus following is to evaluate the actual resistance level of JG11 across all
families of ALS inhibitors. A series of greenhouse dose response experiments were conducted
with seven different ALS-inhibiting herbicides to compare the response of JG11 to that of the
susceptible population, WCS. Results indicated that JG11 was resistant to ALS-inhibiting
herbicides, and the R/S ratios varied from 3 to 90 among different herbicides. In order to
investigate the possible resistance mechanism, a cytochrome P450 inhibitor, malathion, was
incorporated with herbicide application. The combination caused more injury to the JG11
population, which suggested P450 hydroxylation contributes to herbicide detoxification in the
JG11 waterhemp, and is the possible mechanism of ALS resistance.

43
3.2 Introduction
Waterhemp [Amaranthus tuberculatus (Moq.) Sauer] is a common weed species in the
Amaranthaceae family (Sauer, 1955). It has evolved resistance to multiple herbicides with
different modes of action (Heap, 2013b). As described in Chapter 1, MCR (from McLean
County, IL), which has multiple resistance to HPPD inhibitors and atrazine, also showed
resistance to ALS inhibitors even though ALS inhibitors have not been used recently in the crop
field from which MCR was obtained. In preliminary greenhouse trials, this population was found
to contain plants with either high or moderate resistance to the ALS inhibitors chlorimuron and
imazethapyr. After testing individual plants treated by ALS inhibitors with PCR-RFLP
(restriction fragment length polymorphism), a Trp574Leu ALS mutation was found in the highly
resistant plants but not in moderately resistant plants (described in Chapter 1). Evidence provided
by analyzing the full protein-coding sequence and comparing the progeny from moderately
resistant plants with other waterhemp populations suggested that there is a mechanism of
resistance other than mutation in ALS gene in this population (Chapter 2).
There are five different chemical families of herbicides that target the ALS enzyme. But
because of the variation of the chemical structures, these ALS inhibitors may bind slightly
differently to the ALS enzyme, and as a result, mutation at ALS may confer resistance to only
one family of ALS inhibitors, or may confer cross resistance to more than one group of ALS-
inhibiting compounds (Patzoldt and Tranel, 2007). These cross resistance patterns can now be
predicted based on the knowledge we have obtained from numerous ALS mutations previously
characterized (Tranel et al., 2013; Chapter 1). But for a non-target-site resistance mechanism, the
resistance pattern across different families of ALS inhibitors is more difficult to predict.

44
In terms of the mechanism of non-target-site resistance, since MCR population has been
confirmed to have metabolism-based resistance to both HPPD herbicides and atrazine (Ma, et al.,
2013), the former is detoxified by hydroxylation process induced by cytochrome P450s and the
latter is detoxified through glutathione conjugation, it is indicated that this population has a high
potency of enhanced metabolism. In addition, ALS inhibitor detoxified by cytochrome P450s is a
common mechanism of crop tolerance to ALS inhibitors (Siminszky, 2006). The hypothesis
raised is that the ALS resistance in JG11 is caused by enhanced detoxification by cytochrome
P450s.
The objective of the study was to evaluate the resistance level of JG11 across the five
families of ALS inhibitors. To address this objective, JG11 population was treated with at least
one herbicide from each of the five families of ALS-inhibiting herbicides at a range of dosages
and compared with a standard susceptible population. A waterhemp population with the ALS
Trp574Leu mutation was also included for comparison. Also, a second set of experiments
included the combination of an inhibitor of cytochrome P450s, malathion, with herbicide
treatment to test for the involvement of P450s in enhanced herbicide detoxification.
3.3 Materials and Methods
3.3.1 Greenhouse Dose Response with ALS-inhibiting Herbicide
3.3.1.1 Plant Material
Populations and plant culture methods used in this section were the same as described in
Chapter 2.3.1.1.

45
3.3.1.2 Herbicide Dose Response
Uniformly sized waterhemp plants (10 to 12 cm tall) of WCS and JG11 were included in
the dose response experiments, as well as the population with the Trp574Leu mutation, ACR,
which was used as a positive control. Plants were sprayed with a TeeJet flat-fan nozzle (TeeJet
80015EVS. TeeJet Technologies, P.O. Box 7900, Wheaton, IL 60187) in a greenhouse spraying
chamber (Generation III Research Sprayer. DeVries Manufacturing, 28081 870th Ave,
Hollandale, MN 56045) that delivers 185 L per ha at 275 kPa. Multiple ALS-inhibiting
herbicides were used to study the resistance level of the JG11 population. All herbicides used are
listed in Table 3.1, and the rates increased equally spaced on a base 3.16 logarithmic scale. All
the treatment mixtures included COC, (1% v/v) and AMS (2.5% v/v) expect for pyrithiobac
sodium, which was mixed with nonionic surfactant (NIS, 0.25% v/v, Agriliance). Non-treated
controls were treated with water and adjuvant only. Because ACR has a very high level of
resistance to ALS inhibitors, this population was treated only with the highest rates of ALS
inhibitors and not the full range of dosages. After herbicide application, plants were placed on
greenhouse benches in a randomized complete block design. Each treatment had 4 replicates and
the experiment was conducted twice.
At 21 days after treatment (DAT), plant injury was visually evaluated and recorded using
a scale from 0 to 100, with 0 indicating a plants was dead and without any green tissue and 100
indicated no injury observed. Aboveground plant tissue was harvested and dried at 65 C for 4 to
7 days.
Dry weight data (m) and visual rating data (v) were combined to obtain an adjusted dry
weight (y) using the function

46
The adjusted dry weight was converted to a proportion of the average of untreated control plants
from the corresponding population. Data from the two runs of each herbicide were pooled, since
Levene’s test for homogeneity of variance was not significant, and analyzed using a non-linear
regression model with the dose-response curve package in R software (Knezevic et al, 2007).
This analysis is similar to the algorithm described in 2.3.2.4, except that GR50 indicating the
effective dose causing 50% growth reduction in biomass is used instead of ED50.
3.3.2 Response of ALS Inhibitors Combined with Malathion
Waterhemp plants from the WCS and JG11 population were treated with ALS inhibitors
with or without commercial liquid malathion insecticide (50% v/v. Spectracide® Malathion
Insect Spray Concentrate. Spectrum Group, Division of United Industries, P.O.Box 142642, St.
Louis, MO 63114-0642). Waterhemp plants (10-12 cm tall) were firstly treated with malathion at
a rate of 2,000 g a.i. ha-1
, including 0.25% NIS. ALS-inhibiting herbicides were applied to plants
1 h after malathion treatment at rates showed in Table 3.. Herbicides and malathion applications
were made using the spray chamber described in 3.3.1.2. A soil drench of 5 mM malathion
solution (50 mL pot-1
) was applied 2 days after foliar treatment as described in Ma et al. (2013).
At 14 days after treatment (DAT), plant injury was visually evaluated and recorded, and
aboveground plant tissue was harvested and dried at 65 C for 4 to 7 days. Each treatment had 6
replicates and the experiment was conducted twice. All dry weight data were converted as
relative to the average of the corresponding untreated control plants as described in 3.3.1.2. Data
from the two runs were compared using the PROC GLM procedure with SAS 9.3. Treatment
means were compared using PROC ANOVA also with SAS 9.3. The significant differences
among treatments were compared by Duncan’s Multiple Range Test with SAS 9.3.

47
3.4 Results and Discussion
3.4.1 Greenhouse Dose Response with ALS-inhibiting Herbicide
All treatments of JG11 and WCS by ALS inhibitors injured the plants at higher dosages
and some of the plants were dead 21 DAT. Based on visual observation, JG11 was less sensitive
to all seven herbicides across the five chemical families (Figure 3.1). As the results show in
Figure 3.2, it is confirmed that JG11 is resistant to ALS inhibitors relative to the WCS population.
For some herbicides, for example primisulfuron, imazethapyr and cloransulam, the commercially
recommended rate did not control JG11 waterhemp plants under greenhouse conditions. In
addition, although JG11 confers resistance across a broad spectrum of ALS inhibitors, the R/S
ratio varied among herbicides, and did not necessarily relate to the chemical families. This is
different from resistance caused by amino acid substitution, which typically confers resistance to
a group of chemicals from the same family (Tranel and Wright, 2002). For instance, Pro197His
mutation in wild radish (Raphanus raphanistrum) conferred resistance to SUs and TPs, which
have similar structures, but not to IMIs (Yu et al., 2003). In our case of JG11, the greatest
resistance was observed from cloransulam (a TP), with an R/S ratio of more than 90-fold. The
R/S ratio of two SUs ranged from 6 to 11, and those of two IMIs ranged from 9 to 19.
The variation of responses among different herbicides was large, which made us curious
about the mechanism behind it. Since we confirmed the population does not contain mutations
related to herbicide binding, the second most likely resistant mechanism is enhanced
detoxification. Cytochrome P450 hydroxylation is a common detoxification process in crops
tolerant to ALS inhibitors and it hydroxylates the herbicide molecule at the ring structure and
makes it inactive (Ohkawa et al, 1999). There is also a similar process found in other weed
species (Fischer et al., 2000). Moreover, MCR population, the original population from which

48
JG11 was derived, was characterized with P450-mediated resistance to HPPD inhibitors. We
therefore hypothesized that P450s also mediate resistance to ALS inhibitors in the JG11
population. Therefore, we conducted an experiment using malathion, an inhibitor of cytochrome
P450s.
3.4.2 Response to ALS Inhibitors Combined with Malathion
Figure 3.3 and Figure 3.4 show plants 14 DAT with ALS inhibitor with and without
malathion, Herbicide rates were chosen to cause only moderate injury to JG11 plants. Addition
of malathion resulted in injury to JG11 that was similar to that observed when WCS was treated
by herbicide only. Results of the analysis of the dry weight and adjusted dry weight data were
summarized in Table 3.3 and Table 3.4. Except for sulfometuron, all cases show means of JG11
treated only with herbicide were significantly less than those obtained when malathion was
included with the herbicide, and the latter were not significantly different from WCS population
treated with herbicides, with or without malathion present. In sulfometuron treatment, the
herbicide dosage was rather high and caused a severe injury to the JG11 population.
Nevertheless, by adding malathion, the injury became insignificant to WCS population treated
with herbicide only (Figure 3.3).
There are still many aspect of metabolism-based resistance needed to be investigated.
The complexity of cytochrome P450s increases the difficulty of elucidating the genetics and
expression of resistance. However, we can draw the conclusion that cytochrome P450s
contribute to the enhanced detoxification process in JG11, which makes JG11 more resistance to
ALS inhibitors. It also explains the differentiation of resistance level across various ALS-
inhibiting chemicals; some of the chemicals, sulfometuron and imazapyr, are harder to be

49
detoxified through hydroxylation by P450s than the others, like cloransulam and primisulfuron.
This conclusion agrees with the results of the study of HPPD resistance in MCR population,
which is caused by enhanced metabolism (Ma, et al., 2013). MCR is another example of a weed
population with herbicide resistance to ALS inhibitors selected by other modes of action, which
has been observed in other weed species as discussed in Chapter 1.
3.5 Literature Cited
Fischer, A. J., Bayer, D. E., Carriere, M. D., Ateh, C. M., and Yim, K.-O. (2000). Mechanisms of
resistance to bispyribac-sodium in an Echinochloa phyllopogon accession. Pesticide
Biochemistry and Physiology, 68: 156-165.
Heap, I. (2013b, September 27). Herbicide resistant common waterhemp globally (Amaranthus
tuberculatus (syn. rudis)). Retrieved September 27, 2013, from The International Survey
of Herbicide Resistant Weeds: http://www.weedscience.org/
Knezevic, S. Z., Streibig, J. C., and Ritz, C. (2007). Utilizing R software package for dose-
response studies: theconcept and data analysis. Weed Technology, 21: 840-848.
Ma, R., Kaundun, S. S., Tranel, P. J., Riggins, C. W., McGinness, D. L., Hager, A. G., Hawkes,
T., McIndoe, E. and Riechers, D. E. (2013). Distinct detoxification mechanisms confer
resistance to mesotrione and atrazine in a population of waterhemp. Plant Physiology,
163: 363-377.
Ohkawa, H., Tsujii, H., and Ohkawa, Y. (1999). The use of cytochrome P450 genes to introduce
herbicide tolerance in crops: a review. Pesticide Science, 55: 867-874.
Patzoldt, W. L., and Tranel, P. J. (2007). Multiple ALS mutations confer herbicide resistance in
waterhemp (Amaranthus tuberculatus). Weed Science, 55: 421-428.

50
Sauer, J. (1955). Revision of the dioecious amaranths. Madrono, 13: 5-46.
Siminszky, B. (2006). Plant cytochrome P450-mediated herbicide metabolism. Phytochemistry
Reviews, 5: 445-458.
Tranel, P. J., and Wright, T. R. (2002). Resistance of weeds to ALS-inhibiting herbicides: what
have we learned? Weed Science, 50: 700-712.
Tranel, P. J., Wright, T. R., and Heap, I. M. (2013). Mutations in herbicide-resistant weeds to
ALS inhibitors. Retrieved September 29, 2013, from International Survey of Herbicide
Resistant Weeds: http://www.weedscience.org/.

51
3.6 Tables and Figures
Table 3.1 Herbicide used in greenhouse dose response experiments. Recommended application rates are obtained from product labels
as used postemergence.
Trade
name
Active ingredient Chemical family Manufacture Formulation Recommended
application rate
(g a.i. per ha)
Dose response rate
(g a.i. per ha)
Beacon primisulfuron-methyl Sulfonylurea Syngenta 75WG 40 0.4 to 400
Oust sulfometuron-methyl Sulfonylurea DuPont 75WG 105 0.105 to 105
Pursuit imazethapyr Imidazolinone BASF 22.87G 65 6.5 to 6500
Arsenal imazapyr Imidazolinone BASF 26.7G 900 0.009 to 9
Staple pyrithiobac sodium Pyrimidinyl(thio)-
benzoate
DuPont 3.2L 726 1.57 to 1570
Olympus propoxycarbazone Sulfonylamino-
carbonyl-triazolinone
Bayer 70WG 55 0.174 to 174
FirstRate cloransulam-methyl Triazolopyrimidine Dow AgroSciences 84WG 36 3.6 to 3600
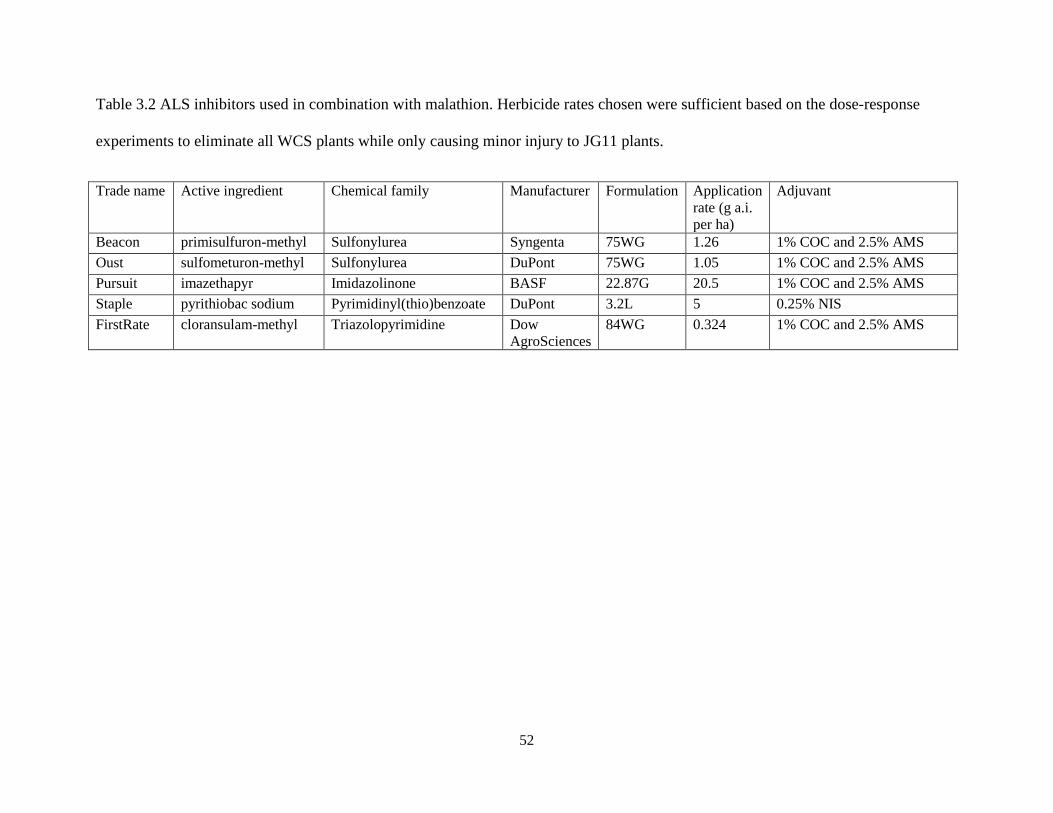
52
Table 3.2 ALS inhibitors used in combination with malathion. Herbicide rates chosen were sufficient based on the dose-response
experiments to eliminate all WCS plants while only causing minor injury to JG11 plants.
Trade name Active ingredient Chemical family Manufacturer Formulation Application
rate (g a.i.
per ha)
Adjuvant
Beacon primisulfuron-methyl Sulfonylurea Syngenta 75WG 1.26 1% COC and 2.5% AMS
Oust sulfometuron-methyl Sulfonylurea DuPont 75WG 1.05 1% COC and 2.5% AMS
Pursuit imazethapyr Imidazolinone BASF 22.87G 20.5 1% COC and 2.5% AMS
Staple pyrithiobac sodium Pyrimidinyl(thio)benzoate DuPont 3.2L 5 0.25% NIS
FirstRate cloransulam-methyl Triazolopyrimidine Dow
AgroSciences
84WG 0.324 1% COC and 2.5% AMS

53
Table 3.3 ANOVA results of malathion incorporated with herbicide treatment. The factors of
malathion, herbicide and biotype all had significant impact on plant growth, which was reflected
by both dry weight and adjusted dry weight.
ANOVA – Dry Weight
Estimate Std. Error t value P value
Intercept 1.05535 0.05020 21.024 <2e-16
Biotype 0.17199 0.02683 6.410 6.04e-10
Herbicide 0.63939 0.04854 13.173 <2e-16
Malathion 0.19429 0.02683 7.241 4.18e-12
ANOVA – Adjusted Dry Weight
Estimate Std. Error t value P value
Intercept 0.97812 0.05109 19.147 <2e-16
Biotype 0.11391 0.02731 4.172 4.02e-5
Herbicide 0.070496 0.04940 14.271 <2e-16
Malathion 0.16947 0.02731 6.206 1.92e-9

54
Table 3.4 Duncan’s Multiple Range Test of malathion-ALS inhibitor combination treatments.
The columns of “trt” are labeled with population and treatment combination. Following the
population of either “JG11” or “WCS”, “OO” stands for untreated control, “MO” stands for
treated with only malathion, “HO” stands for treated with only herbicide, and “HM” stands for
the combination application of both herbicide and malathion. Dry weight means with the same
letter are not significantly different. Responses of both populations under herbicide treatment
with or without malathion are highlighted in bold for comparison.
Primisulfuron
Cloransulam
Duncan's Multiple Range Test Duncan's Multiple Range Test
Duncan Grouping Mean N trt Duncan Grouping Mean N trt
A 1 12 JG11OO A 1 12 JG11OO
A 1 12 WCSOO A 1 12 WCSOO
B 0.62743 12 JG11OM B 0.72206 12 JG11OM
B 0.61042 12 WCSOM C 0.50276 12 JG11HO
C 0.30634 12 JG11HO C 0.45384 12 WCSOM
D 0.15253 12 WCSHO D 0.22604 12 JG11HM
D 0.10212 12 JG11HM D 0.11201 12 WCSHO
D 0.06123 12 WCSHM D 0.06808 12 WCSHM
Sulfometuron Pyrithiobac
Duncan's Multiple Range Test Duncan's Multiple Range Test
Duncan Grouping Mean N trt Duncan Grouping Mean N trt
A 1 12 JG11OO A 1 12 JG11OO
A 1 12 WCSOO A 1 12 WCSOO
B 0.62743 12 JG11OM B 0.62743 12 JG11OM
B 0.61042 12 WCSOM B 0.61042 12 WCSOM
C 0.2464 12 JG11HO C 0.40196 12 JG11HO
CD 0.12747 12 JG11HM D 0.12014 12 JG11HM
D 0.07279 12 WCSHO D 0.08288 12 WCSHO
D 0.03266 12 WCSHM D 0.02635 12 WCSHM
Imazethapyr
Duncan's Multiple Range Test
Duncan Grouping Mean N trt
A 1 12 JG11OO
A 1 12 WCSOO
B 0.62743 12 JG11OM
B 0.61042 12 WCSOM
B 0.59072 12 JG11HO
C 0.16335 12 JG11HM
C 0.09019 12 WCSHO
C 0.05177 12 WCSHM

55
Figure 3.1 Demonstration of dose response to ALS-inhibiting herbicides in the greenhouse. In
each set the photo on top shows the response of JG11 population and the bottom is WCS. The
plant in the far left of each photo is an untreated control, and plants to the right were treated with
increasing herbicide dosages; dosages are listed in Table 3.1.
A. Primisulfuron

Figure 3.1(cont.)
56
B. Cloransulam C. Propoxycarbazone

Figure 3.1 (cont.)
57
D. Sulfometuron E. Pyrithiobac sodium

Figure 3.1 (cont.)
58
F. Imazethapyr E. Imazapyr

59
Primisulfuron Estimate of R/S Std. Error t-value p-value
10.8266 3.7446 2.6242 0.0102
Cloransulam Estimate of R/S Std. Error t-value p-value
90.0264 43.0252 2.0692 0.0397
Figure 3.2 Dose response curves and statistical analyses. JG11/WCS values reflect the R/S ratio
corresponding to each herbicide treatment. The X-axis represents the herbicide dosage (g of a.i.
per ha) on a log-scale, and the Y-axis represents the adjusted dry weight data relative to
untreated control.
Primisulfuron (g a.i./ha)
Cloransulam (g a.i./ha)
Ad
just
ed d
ry w
eigh
t re
lati
ve t
o
un
trea
ted
co
ntr
ol
Ad
just
ed d
ry w
eigh
t re
lati
ve t
o
un
trea
ted
co
ntr
ol

Figure 3.2 (cont.)
60
Propoxycarbazone Estimate of R/S Std. Error t-value p-value
3.1109 1.0172 2.0751 0.0426
Sulfometuron Estimate of R/S Std. Error t-value p-value
5.7662 1.4724 3.2369 0.0016
Pyrithiobac Sodium Estimate of R/S Std. Error t-value p-value
2.83098 0.91997 1.99026 0.0257
Propoxycarbazone (g a.i./ha)
Sulfometuron (g a.i./ha)
Pyrithiobac (g a.i./ha)
Ad
just
ed d
ry w
eigh
t re
lati
ve t
o
un
trea
ted
co
ntr
ol
Ad
just
ed d
ry w
eigh
t re
lati
ve t
o
un
trea
ted
co
ntr
ol
Ad
just
ed d
ry w
eigh
t re
lati
ve t
o
un
trea
ted
co
ntr
ol

Figure 3.2 (cont.)
61
Imazethapyr Estimate of R/S Std. Error t-value p-value
19.4032 5.9427 3.0967 0.0025
Imazapyr Estimate of R/S Std. Error t-value p-value
8.8605 3.4103 2.3050 0.0229
Imazethapyr (g a.i./ha)
Imazethapyr (g a.i./ha)
Ad
just
ed d
ry w
eigh
t re
lati
ve t
o
un
trea
ted
co
ntr
ol
Ad
just
ed d
ry w
eigh
t re
lati
ve t
o
un
trea
ted
co
ntr
ol

62
Figure 3.3 Malathion-ALS inhibitor treatments. “OO” stands for untreated control. “MO” stands
for applied with only malathion. “HO” stands for applied with only herbicide. And “HM” stands
for the combined application of both herbicide and malathion.
JG11 JG11 JG11 JG11 WCS WCS WCS WCS
OO MO HO HM HO HM MO OO
JG11 WCS JG11 WCS JG11 JG11 WCS WCS
OO OO MO MO HO HM HO HM
JG11 JG11 JG11 JG11 WCS WCS WCS WCS
OO MO HO HM HO HM MO OO
JG11 JG11 JG11 JG11 WCS WCS WCS WCS
OO MO HO HM HO HM MO OO
JG11 JG11 JG11 JG11 WCS WCS WCS WCS
OO MO HO HM HO HM MO OO
A. Primisulfuron B. Cloransulam
C. Sulfometuron E. Pyrithiobac sodium
F. Imazethapyr

63
Figure 3.4 Photos showing multiple plants of the JG11 and WCS populations treated with
herbicide or herbicide plus malathion.
A. Primisulfuron B. Cloransulam
C. Sulfometuron E. Pyrithiobac sodium
F. Imazethapyr
JG11
Herbicide
Treatment
WCS
Herbicide
Treatment
JG11
Herbicide
+ Malathion
Treatment
WCS
Herbicide
+ Malathion
Treatment
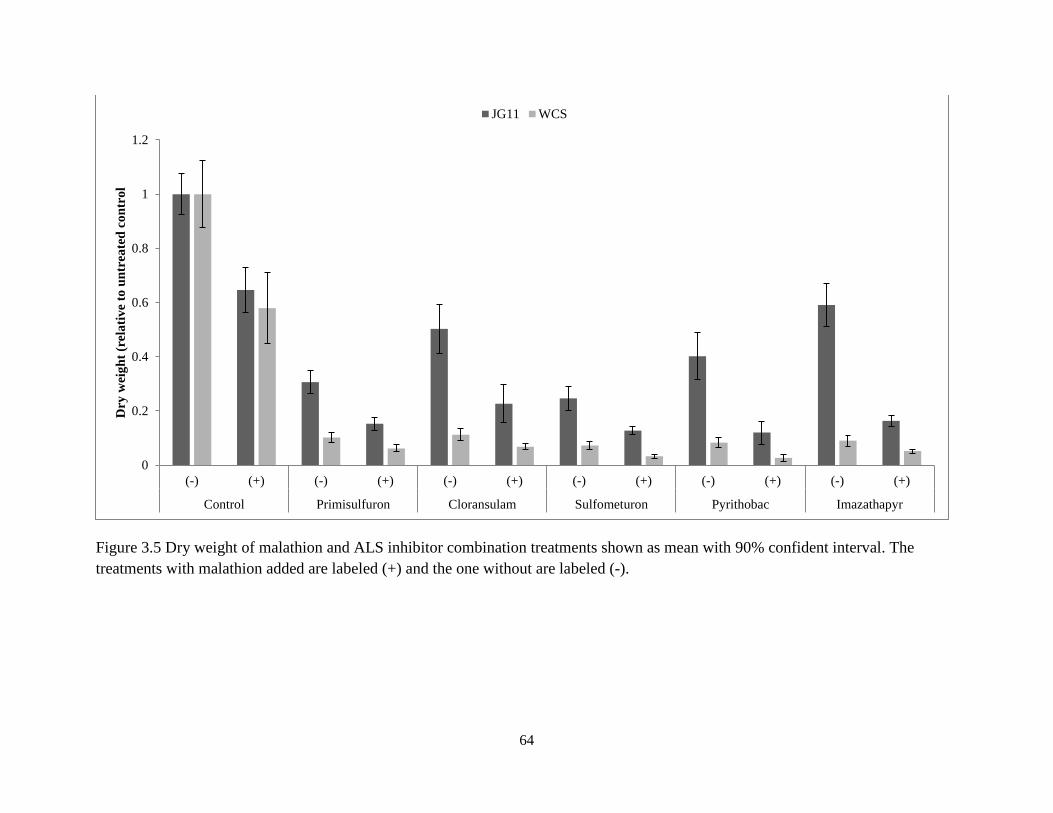
64
Figure 3.5 Dry weight of malathion and ALS inhibitor combination treatments shown as mean with 90% confident interval. The
treatments with malathion added are labeled (+) and the one without are labeled (-).
0
0.2
0.4
0.6
0.8
1
1.2
(-) (+) (-) (+) (-) (+) (-) (+) (-) (+) (-) (+)
Control Primisulfuron Cloransulam Sulfometuron Pyrithobac Imazathapyr
Dry
wei
gh
t (r
ela
tiv
e to
un
trea
ted
co
ntr
ol
JG11 WCS

65
CHAPTER 4
Concluding Remarks
4.1 Research Conclusions and Implications
The acetolactate synthase (ALS) enzyme, or acetohydroxyacid synthase (AHAS) enzyme
is the target site of five families of herbicides. The herbicides inhibit substrate binding to the
enzyme, and stop the biosynthesis of branched-chain amino acids (BCAA). ALS-inhibiting
herbicides have been widely used since they have been developed, but a great variety of weed
species have evolved resistance to them. As one of the most problematic weed in the Midwest
region, waterhemp is notorious for being a highly reproductive, highly competitive, and vigorous
weed species that affects crop yield greatly when not managed effectively (Steckel et al., 2003;
Steckel, 2007). The fact that waterhemp has evolved resistance to multiple herbicide modes of
action narrows the control options and makes the weedy properties of waterhemp more
challenging (Hausman et al., 2011; Patzoldt et al., 2005). Reports of waterhemp being resistant
to 6 modes of action, including ALS inhibitors, have been found across the Midwest (Heap,
2013b).
Previous studies of ALS-resistant waterhemp was mostly focused on amino acid
substitutions in ALS enzyme that leads to an insensitive target site to herbicide binding, and
mutations at two amino acids, Trp574 and Ser653, were found in waterhemp biotypes that confer
high levels of resistance (Patzoldt, et al, 2005; Patzoldt and Tranel, 2007). However, a
waterhemp population, MCR, displayed two phenotypes of resistance to ALS inhibitors in
greenhouse experiments (Chapter 1). The Trp574Leu mutation was found in all highly resistant

66
individuals but it was not found in moderately resistant plants from this population. Since MCR
has been confirmed with metabolism-based resistance to HPPD inhibitors (Hausman, et al., 2011;
Ma, et al., 2013), and enhanced metabolism is how crops tolerate ALS-inhibiting herbicides
(Fonné-Pfister et al., 1990), the hypothesis was raised that there is a non-target-site mechanism
of ALS resistance in MCR waterhemp.
To test the hypothesis, progeny JG11 of two moderately resistant waterhemp plants from
MCR was used in a series of experiments. In Chapter 2, DNA of leaf tissue of JG11 survivors of
herbicide treatment was extracted and the ALS gene was sequenced. Among the 582 codons of
ALS mature protein from 8 different individuals of JG11 plants, none of the known ALS
mutations that affect herbicide binding was found. Twenty-nine other amino acid
polymorphisms in alleles of the 8 individuals were observed, but only 5 of them appeared
consistently. Moreover, these substitutions were detected in sequences of other Amaranthus
species as well, both ALS sensitive and resistant biotypes, inferring those sites are naturally less
conserved, and are unlikely the genetic base of ALS resistance in JG11.
To further evaluate the hypothesis of non-target-site resistance, an in vitro assay was
performed to compare the activity of the ALS enzyme with or without herbicide inhibition. ALS
enzyme from JG11 plants was as sensitive to an ALS herbicide as was that of WCS, a biotype
known to be sensitive to ALS inhibitors. Collectively, results from ALS gene sequencing and the
ALS enzyme assay provide strong evidence that the whole plant resistance of JG11 is not based
on an insensitive target site.
With the confirmation of a non-target-site mechanism of ALS resistance, the actual
resistance pattern across ALS herbicide families was characterized in Chapter 3. Greenhouse
dose response experiment was conducted with 7 different chemicals on JG11 population together

67
with WCS and ACR as controls. Relative to WCS, the majority of R/S ratios ranged from 2 to 10,
except cloransulam, for which an R/S ratio over 50 was obtained. Both visual rating and biomass
data suggested JG11 is moderately resistant to ALS herbicide across all five families, and even
survives commercially suggested rates of application.
An inhibitor of cytochrome P450s, malathion, was incorporated with herbicide treatments
based on the hypothesis that the resistance could be related to enhanced detoxification, which is
similar to the mechanism conferring resistance to HPPD inhibitors in MCR. The experiments
were performed in the greenhouse with five different ALS inhibitors at sub-lethal dosages to
JG11. The addition of malathion significantly increased herbicide activity, to the extent that
biomass was not significantly different from that obtained for WCS receiving the same treatment.
The results suggest detoxification by cytochrome P450s plays an important role in the non-
target-site ALS resistance in JG11. The presence of non-target-site ALS resistance could have
significant impacts on herbicide cross resistance and on weed management for this species.
Despite the fact that there previously was no known metabolism-based resistance of
waterhemp to ALS inhibitors, and the main focus of ALS resistance in waterhemp has been on
gene mutation, the nature of waterhemp’s high reproduction rate and great genetic variability
makes the selection of alternate resistance mechanisms inevitable. In the previous chapters the
hypothesis of a metabolism-based resistance mechanism is supported in the JG11 population and,
by extension, in the original HPPD-resistant MCR population as well. The metabolism-based
resistance in waterhemp or weeds in general impacts weed management in three major aspects.
First of all, the resistant traits are not necessarily selected by ALS inhibitors, but by herbicides of
other modes of action as well, and may confer unpredictable cross resistance patterns. In addition,
the possibility of some ALS mutated resistant populations to possess non-target-site resistance

68
mechanism has not been erased, meaning those populations may not be susceptible to HPPD
inhibitors or other herbicide modes of action. Secondly, comparing to mutated resistant
populations, metabolism-based resistant populations are more sensitive, but still are able to
survive and reproduce after treatment with typical rates of ALS inhibitors. Thirdly, the approach
of reducing metabolism in resistant weeds with cytochrome P450s inhibitors would potentially
damage crops that tolerate herbicide treatments through metabolism.
Future research directions suggested by my findings include further physiological studies
to investigate metabolism, and genetic studies. For instance, although I have hypothesized that
cytochrome P450s are involved in ALS-inhibitor detoxification, due to the complexity of
cytochrome P450s and hydroxylation process, more research is needed (Schuler, 1996).
Radiolabeled herbicide application and liquid scintillation spectrometry can be used. To
investigate herbicide uptake and translocation, radiolabeled herbicide could be applied to young
leaves of JG11 and WCS waterhemp plants, and the radioactive residue in plant tissue can be
quantified and compared at various time points after application. The results would reveal
whether the two populations have significant difference in herbicide uptake and translocation.
Whole plant application with radiolabeled herbicide can be performed on JG11, WCS, and a crop
with enhanced metabolism to detoxify ALS inhibitor, wheat, for example. HPLC could then be
used to identify and quantify the amount of parent herbicide and metabolites as evidence of
enhanced metabolism and confirm the resistance mechanism.
Meanwhile, the genetics and inheritance of the non-target-site ALS resistant trait and its
co-segregation pattern with resistance to HPPD inhibition should be investigated. F1 populations
of JG11 x WCS have been made and can be used as material in future dose response study and
co-segregation study. JG11 plants were selected from the survivors under ALS-inhibiting

69
herbicide treatment applied at 10 to 12 cm high. Herbicide treatment injured and stunted the
plants but the majority recovered and started flowering around 14 DAT. WCS was grown in the
greenhouse with proper moisture and fertilizer supply without any herbicide treatment. When the
inflorescence was formed and was able to be identified, the JG11 males and WCS females were
chosen and put in a pollination bag supported by PVC pipe frame to isolate from the rest of the
plants. The waterhemp plants for crossing were kept in the pollination bag for at least 21 days,
and the female plants baring seeds were harvested and dried in paper bags at room temperature
for another 7 days before cleaning and stratification. The seed lines are described in Table 4.1.
Further crosses to obtain F2 and backcrosses can be created and utilized as well. WCS,
JG11 and progenies of F1, F2 and BC can be screened with different doses of ALS inhibitors and
HPPD inhibitors, and generate dose response curves with biomass data using the function
described in Chapter 3, to characterize the resistance of future progenies. Also, a two-run
screening with ALS and HPPD inhibitor, in comparison with single herbicide treatments and
untreated control can help determine the co-segregation pattern.
4.2 Literature Cited
Fonné-Pfister, R., Gaudin, J., Kreuz, K., Ramsteiner, K., and Ebert, E. (1990). Hydroxylation of
primisulfuron by an inducible cytochrome P450-dependent monooxygenase system from
maize. Pesticide Biochemistry Physiology, 37: 165-173.
Hausman, N. E., Singh, S., Tranel, P. J., Riechers, D. E., Kaundun, S. S., Polge, N. D., Thomas,D.
A., and Hager, A. G. (2011). Resistance to HPPD-inhibiting herbicides in a population of
waterhemp (Amaranthus tuberculatus) from Illinois, United States. Pest Management
Science, 67: 258-261.

70
Heap, I. (2013b). Herbicide resistant common waterhemp globally (Amaranthus tuberculatus
(syn. rudis)). Retrieved September 27, 2013, from The International Survey of Herbicide
Resistant Weeds: http://www.weedscience.org/
Ma, R., Kaundun, S. S., Tranel, P. J., Riggins, C. W., McGinness, D. L., Hager, A. G., Hawkes,
T., McIndoe, E. and Riechers, D. E. (2013). Distinct detoxification mechanisms confer
resistance to mesotrione and atrazine in a population of waterhemp. Plant Physiology,
163: 363-377.
Patzoldt, W. L., and Tranel, P. J. (2007). Multiple ALS mutations confer herbicide resistance in
waterhemp (Amaranthus tuberculatus). Weed Science, 55: 421-428.
Patzoldt, W. L., Tranel, P. J., and Hager, A. G. (2005). A waterhemp (Amaranthus tuberculatus)
biotype with multiple resistance across three herbicide sites of action. Weed Science, 53:
30-36.
Schuler, M. A. (1996) Plant Cytochrome P450 Monooxygenases, Cr. Rev. Plant Science 15:
235-284
Steckel, L. E. (2007). The dioecious Amaranthus spp.: here to stay. Weed Technology, 21: 567-
570.
Steckel, L. E., Sprague, C. L., Hager, A. G., Simmons, F. W., and Bollero, G. A. (2003). Effects
of shading on common waterhemp (Amaranthus rudis) growth and development. Weed
Science, 51: 898-903.

71
4.3 Tables
Table 4.1 Seed lot chart of F1 and JG11 crosses.
No. Date of Collection Parents
JG13 8/5/2013 Female JG11*Male JG11
JG14 8/5/2013 Female JG11*Male JG11
JG15 8/5/2013 Female JG11*Male JG11
JG16 8/5/2013 Female JG11*Male JG11
JG17 8/5/2013 Female JG11*Male JG11
JG18 10/31/2013 Female JG11*Male JG11
JG19 10/31/2013 Female JG11*Male JG11
JG20 10/31/2013 Female WCS*Male JG11
JG21 10/31/2013 Female WCS*Male JG11
JG22 10/31/2013 Female WCS*Male JG11
JG23 1/23/2014 Female JG11*2 Males JG11
JG24 1/23/2014 Female JG11*2 Males JG11
JG25 1/23/2014 Female JG11*2 Males JG11
JG26 1/23/2014 Female JG11*2 Males JG11
JG27 1/23/2014 Female WCS*2 Males JG11
JG28 1/23/2014 Female WCS*2 Males JG11
JG29 1/23/2014 Female WCS*2Males JG11
















![Protease Inhibitors - DNA GDAŃSK€¦ · Tab. 1: Individual Protease Inhibitors Prod.-No. Description M [g/mol] Structure Target Protease Class, Target Enzymes Mechanism Recommen-ded](https://img.dokumen.tips/doc/110x75/5ad6ea197f8b9a5b538bf718/protease-inhibitors-dna-gdansk-tab-1-individual-protease-inhibitors-prod-no.jpg)












