Embed Size (px)
Citation preview

(Ne)riebus Mycobacterium
gyvenimas
2007–2013 m. Žmogiškųjų išteklių pl÷tros veiksmų programos 3 prioriteto „Tyr÷jų geb÷jimų stiprinimas“ VP1-3.1-ŠMM-05-K priemon÷s „MTTP tematinių tinklų, asociacijų veiklos stiprinimas“projektas „Lietuvos biochemikų draugijos potencialo kurti žinių visuomenę didinimas“
(NR. VP1-3.1-ŠMM-05-K-01-022)
gyvenimas
Rolandas Meškys
VU Biochemijos institutas
2011-01-26, 28
Vilnius, Kaunas

Ar žinote, kad jos gramteigiamos, rūgščiai atsparios, aerobin÷s, nejudrios,
storos lazdel÷s formos bakterijos; sukelia vieną labiausiai pasaulyje
paplitusių infekcinių ligų, kuria kasmet suserga apie 9 mln. pasaulio
gyventojų;
sintetina F420 kofaktorių, ergotioneiną (kurio transporto pažaidoms esant,
žmon÷s linkę sirgti reumatoidiniu artritu), diterpenus ir šakotas riebalų
rūgštis, turinčias grandin÷je 40 ir daugiau anglies atomų;
gali alkanus, benzo[a]pireną, dyzelį ar metil tert-butil eterį naudoti kaip anglies gali alkanus, benzo[a]pireną, dyzelį ar metil tert-butil eterį naudoti kaip anglies
ir energijos šaltinį;
PUPilina baltymus bei
gali gyventi, amebose ir makrofaguose? Jei nežinote, tai paskaitos metu bus
trumpai aptarta šių mikroorganizmų biochemin÷ įvairov÷ bei kai kurie jų
metabolizmo ypatumai.

Actinobacteria (High G+C Gram-Positive Bacteria)

High G+C Gram-Positive Bacteria
• Phylum: Actinobacteria
• Highly pleomorphic in morphology
• Most are filamentous
• Some resemble molds by their external • Some resemble molds by their external
asexual spores
• Morphology gives the organism greater
surface-to-volume ratio.

Mycobacterium
• Aerobe
• Non-endospore forming rods.
• Acid-fast
• Have distinctive cell wall, the outermost • Have distinctive cell wall, the outermost
lipopolysaccharide layer is made of
mycolic acids.
• Mycolic acids make bacteria water
resistant.

Mycobacterium tuberculosis
• Causes tuberculosis
• Normally attacks and
grows in lungs but
can attack anywherecan attack anywhere
• Causes symptoms
like cough, chest pain

Mycobacterium leprae, raupsai
• Causes Leprosy A.K.A Hansen’s Disease
– Chronic disease causes infection in skin and peripheral nerves
• Symptoms
– Skin lesions, thickened dermis
• Transmission uncertain but possibly person to person in • Transmission uncertain but possibly person to person in respiratory droplets

Corynebacterium
Pleomorphic
• Shape can alter with age
• Non-motile, rod-shaped, do not branch or
make spores, club shaped.
• Includes animal and plant pathogens • Includes animal and plant pathogens
(Corynebacterium diphtheriae
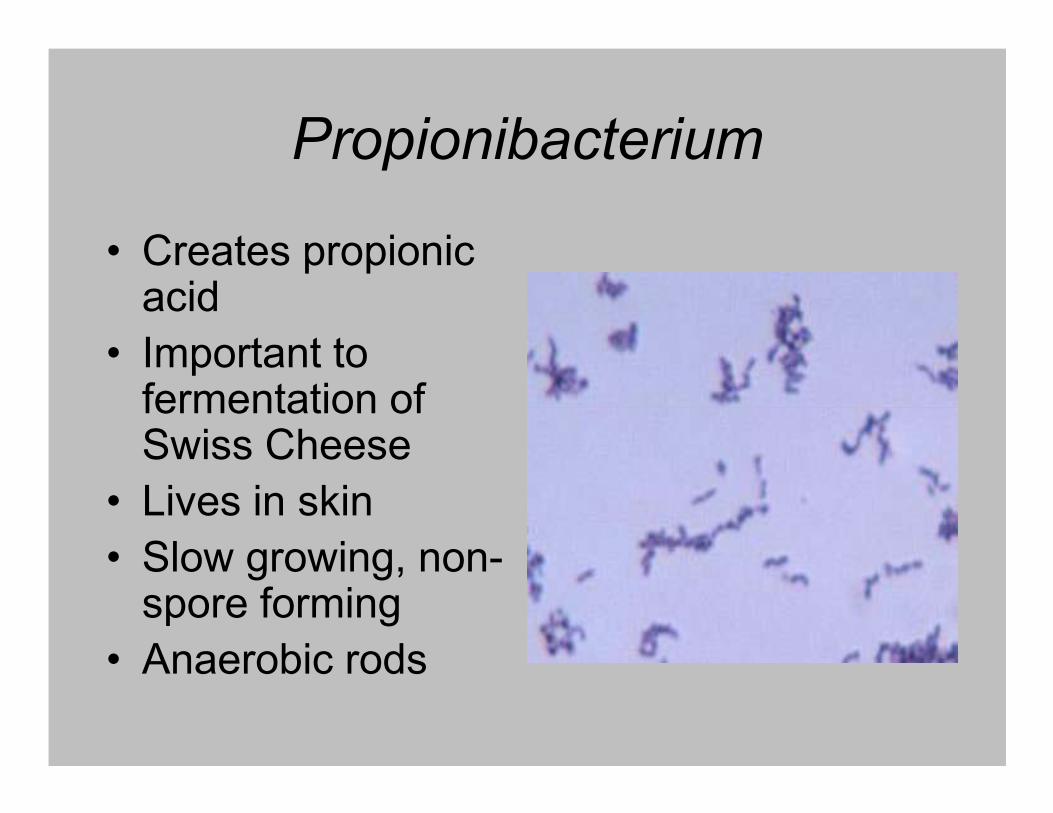
Propionibacterium
• Creates propionic acid
• Important to fermentation of fermentation of Swiss Cheese
• Lives in skin
• Slow growing, non-spore forming
• Anaerobic rods

Propionic acnes
• Causes acne
• Normal inhabitant of skin, but if trapped in
hair follicle becomes acne
• Bacteria releases lipases to digest skin oil • Bacteria releases lipases to digest skin oil
digestive products and bacterial antigens
cause inflammation
• Treatment Topical and oral agents are
used combination of Salicylic acid, Benzyl
peroxide, drying and peeling agents and
antibiotics and Isotretinoin.

Gardnerella
• Pleomorphic
• Facultative Anaerobe
• Stain Gram-negative
• No mycelia• No mycelia
• Gardnerella vaginalis

Frankia
• Filamentous
• Live in soil
• Causes Nitrogen fixation nodules that form
in alder tree rootsin alder tree roots
• Lives in symbiosis with actinorhizal
• Non Pathogenic

Streptomyces
• Best known from actinomycetes
• Found in soil
• Asexual spores
• Produce enzymes to use proteins, polysaccharides cellulose and other organic
• Produce enzymes to use proteins, polysaccharides cellulose and other organic materials in soil
• Produces gaseous compound called geosmin
• Produces most commercial antibiotics like Streptomycin and erythromycin
• Infrequent pathogen

Actinomyces• Facultative anaerobe
• Found in mouth and throat of humans and
animals
• Filamentous
• Non-endospore forming rod shaped• Non-endospore forming rod shaped
• Pathogenic

Nocardia
• Morphologically similar to Actinomyces
• Aerobic
• Filamentous and short rods
• Acid-fast• Acid-fast
• Common in soil

Mycobacterium bovis Bacille Calmette-Guérin
(BCG)• BCG originated with “lait Nocard,” a virulent strain of M. bovis isolated from the milk of a
cow suffering from tuberculous mastitis. Around 1901, this strain was brought to the
Institute Pasteur in Lille, France, and used by Albert Calmette and Camille Guérin for
studies of bovine tuberculosis.
• In order to minimize bacterial clumping and optimize animal infection experiments,
Calmette added ox bile, a detergent, to the glycerol-soaked potato slices on which the M.
bovis was cultured. Within a few months, an isolate with unusual colony morphology
appeared. Moreover, this laboratory-adapted strain exhibited reduced virulence in guinea
pigs. pigs.
• Recognizing the implications of these observations in terms of vaccine development,
Calmette and Guérin continued the serial in vitro passaging of this M. bovis strain for the
next 13 years (1908–1921). During this time, experiments with diverse animal models,
including guinea pigs, rabbits, dogs, cattle, horses, chickens and non-human primates,
established both the safety and efficacy of BCG.
• The first human trial occurred in July 1921. There were no deleterious side effects and,
most importantly, the child did not develop TB even though the infant’s mother had died of
TB shortly after giving birth.

Genealogy of BCG vaccines. Polymorphisms that affect
known virulence genes are highlighted

Detailed structure of a macrophage
showing a typical process of phagocytosis
Meena and Rajni FEBS Journal 277 (2010) 2416

• Key factors of the
survival
mechanisms
involved in the
phagosome
maturation arrest
of Mycobacterium of Mycobacterium
tuberculosis inside
macrophages.
Meena and Rajni FEBS Journal 277 (2010) 2416
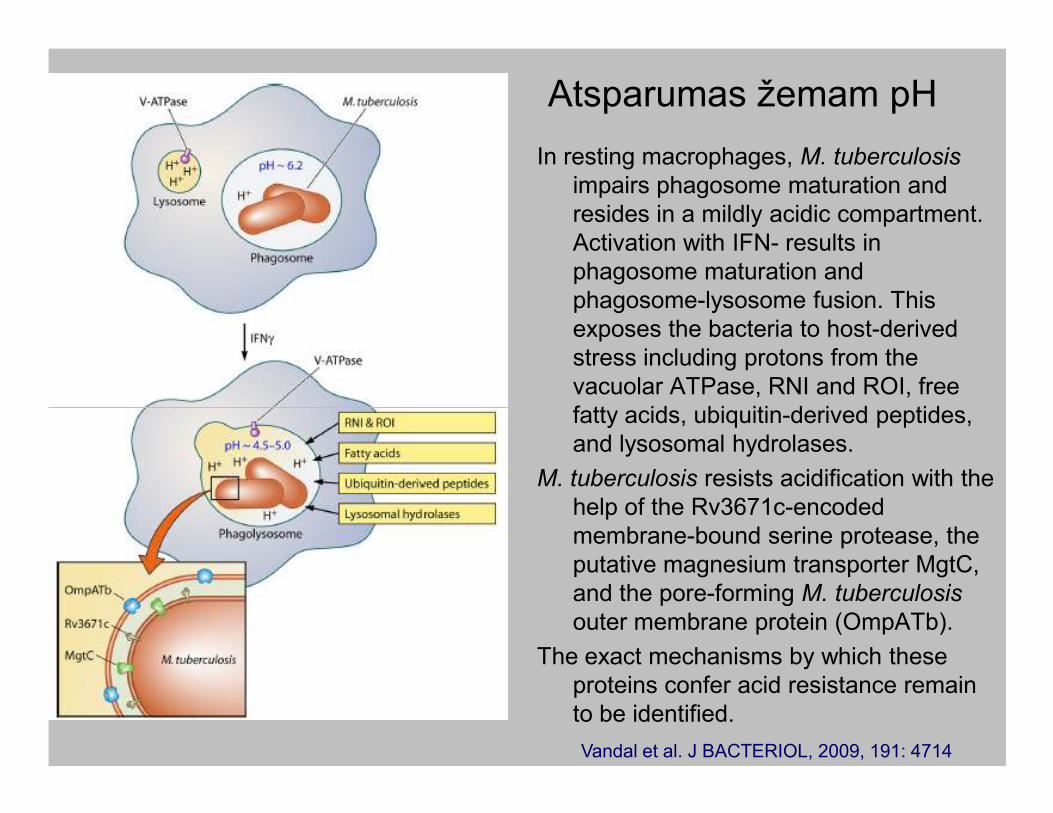
Atsparumas žemam pH
In resting macrophages, M. tuberculosis
impairs phagosome maturation and
resides in a mildly acidic compartment.
Activation with IFN- results in
phagosome maturation and
phagosome-lysosome fusion. This
exposes the bacteria to host-derived
stress including protons from the
vacuolar ATPase, RNI and ROI, free
fatty acids, ubiquitin-derived peptides,fatty acids, ubiquitin-derived peptides,
and lysosomal hydrolases.
M. tuberculosis resists acidification with the
help of the Rv3671c-encoded
membrane-bound serine protease, the
putative magnesium transporter MgtC,
and the pore-forming M. tuberculosis
outer membrane protein (OmpATb).
The exact mechanisms by which these
proteins confer acid resistance remain
to be identified.
Vandal et al. J BACTERIOL, 2009, 191: 4714

Nitrogen uptake, metabolism and control in
M. tuberculosis
Amon et al. J Mol Microbiol Biotechnol 2009;17:20–29

TACO baltymai
• Mycobacterium bovis utilizes a host protein, tryptophan
aspartate-containing coat protein (TACO; also known as
CORO1A or coronin-1), to escape detection by the CORO1A or coronin-1), to escape detection by the
immune system. Upon M. bovis BCG infection, CORO1A
is recruited, in association with tubulin, from the plasma
membrane to the phagosomal membrane to play an
essential role in inhibiting the phagosome–lysosome
fusion, as well as in the survival of bacilli within
macrophages.

Mycobacterium tuberculosis
truncated hemoglobin, trHbN
Fe(II)O2 + NO Fe(III) + NO3-
Lama et al. J Biol Chem 284, 2009 14457–14468
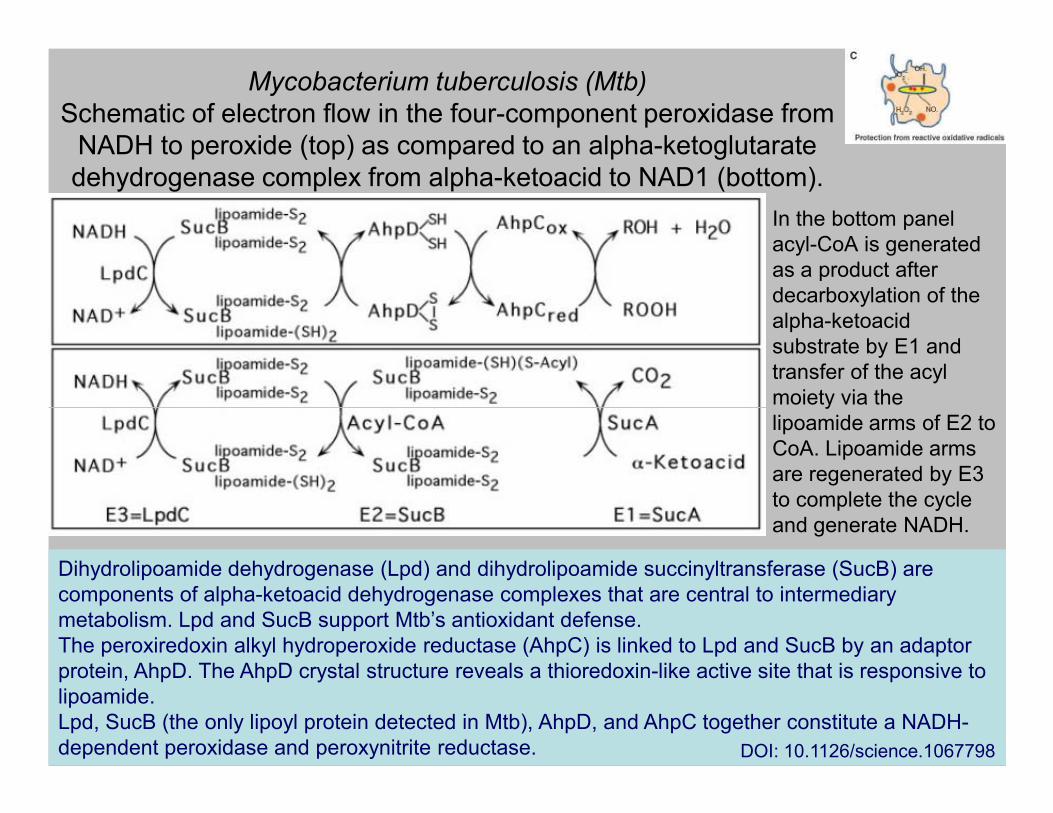
Mycobacterium tuberculosis (Mtb)
Schematic of electron flow in the four-component peroxidase from
NADH to peroxide (top) as compared to an alpha-ketoglutarate
dehydrogenase complex from alpha-ketoacid to NAD1 (bottom).
In the bottom panel
acyl-CoA is generated
as a product after
decarboxylation of the
alpha-ketoacid
substrate by E1 and
transfer of the acyl
moiety via the moiety via the
lipoamide arms of E2 to
CoA. Lipoamide arms
are regenerated by E3
to complete the cycle
and generate NADH.
Dihydrolipoamide dehydrogenase (Lpd) and dihydrolipoamide succinyltransferase (SucB) are
components of alpha-ketoacid dehydrogenase complexes that are central to intermediary
metabolism. Lpd and SucB support Mtb’s antioxidant defense.
The peroxiredoxin alkyl hydroperoxide reductase (AhpC) is linked to Lpd and SucB by an adaptor
protein, AhpD. The AhpD crystal structure reveals a thioredoxin-like active site that is responsive to
lipoamide.
Lpd, SucB (the only lipoyl protein detected in Mtb), AhpD, and AhpC together constitute a NADH-
dependent peroxidase and peroxynitrite reductase. DOI: 10.1126/science.1067798

CO + H2O CO-DH(MCD-HCOO-+ H+/[Fe-S]/FAD) CO2
CO-DH(MCD/[Fe-S]/FADH2)
Oxidation of CO by CO
dehydrogenase
CO-DH(MCD/[Fe-S]/FAD) 2H+ + 2e-
CO + H2O CO
2+ 2H+ + 2e-
Young Min Kim Department of Biology, Yonsei University

Mycobacterial carboxydobacteria
++ +M. vaccae
++ +
-+ +M. tuberculosis H37Ra
++ +Mycobacterium sp. strain JC1
M. parafortuitum
Growthon CO
CO-DHactivity
RubisCOactivityBacteria
+
+
+ +
+ +
M. phlei
M. flavescens
++ +M. peregrinum
++ +M. neoaurum
++ +M. smegmatisMC2
++ +M. gastri
++ +M. vaccae
Park et al., J. Bacteriol. 185:142-147 (2003) Young Min Kim Department of Biology, Yonsei University

CO-DH and NO-DH activities in mycobacterial anda Gram-negative carboxydobacteriaa
BacteriaSpecific activityb
SourceCO-DH NO-DH
Mycobacterium sp. strain JC1 DSM 3803M. tuberculosis H37Ra ATCC 35835
6.09.9
1.52.0
Park et al., BBRC 362:449-453 (2007)
aActivity was determined with cell-free extracts prepared from cells grown at 37oC in SMB-CO.b Nanomoles of INT reduced per milligram of protein per minute.
NO source: sodium nitroprusside (SNP)
M. tuberculosis H37Ra ATCC 35835M. vaccae ATCC 15483O. carboxidovorans OM5 DSM 1227
9.917.518.1
2.03.10.0
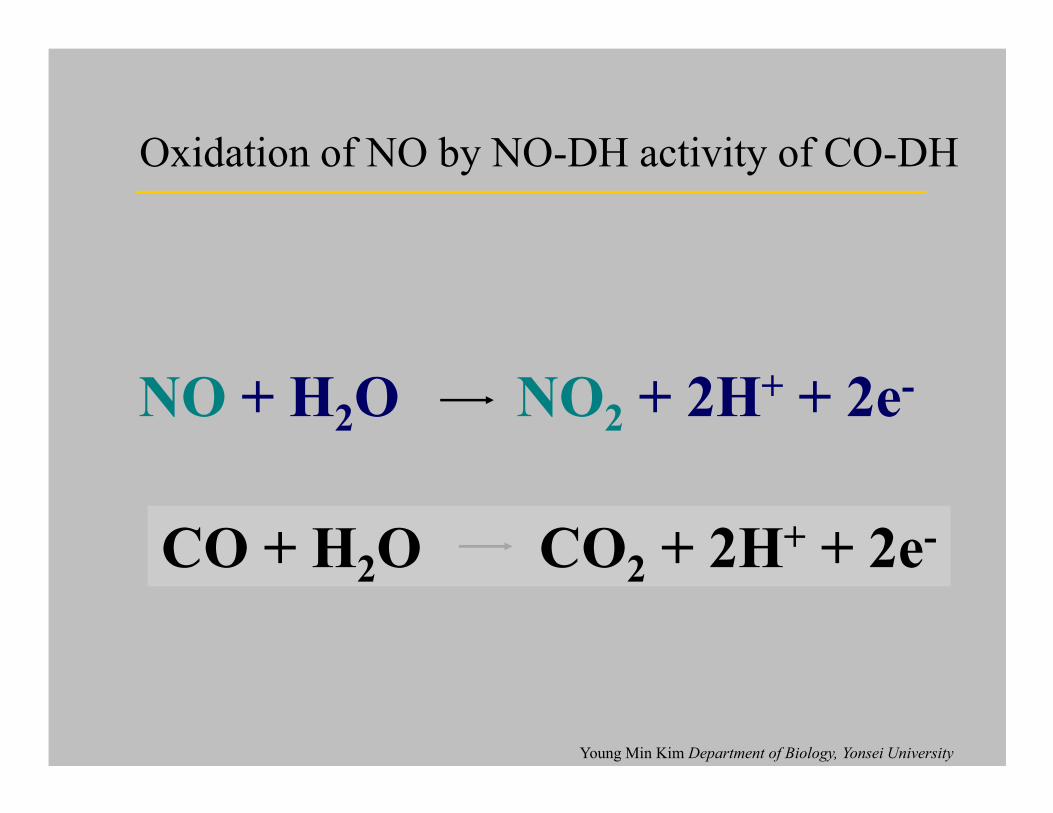
NO + H2O NO
2+ 2H+ + 2e-
Oxidation of NO by NO-DH activity of CO-DH
NO + H2O NO
2+ 2H + 2e
CO + H2O CO
2+ 2H+ + 2e-
Young Min Kim Department of Biology, Yonsei University

PE_PGRS family in
Mycobacterium tuberculosis94-110 aa
170-588 aa
40-1680 aa 7-314 aa
200-3500 aa~ 180 aa
PE
PGRS
PE
PE
Unique sequence
(GGAGGAGGN)n
Proteins whose N-termini contain the characteristic motifs Pro–Glu (PE) or Pro–Pro–
Glu (PPE). A subgroup of the PE proteins contains polymorphic GC-rich
sequences (PGRS), while a subgroup of the PPE proteins contains major
polymorphic tandem repeats (MPTR). The function of most of these proteins
remains unknown.Bottai and Brosch Molecular Microbiology (2009) 73, 325; Tian and Jian-ping. Microbial Pathogenesis 49 (2010) 311
200-3500 aa~ 180 aa
MPTRPPE
200-400 aa
GxxSVPxxWPPE
0-400 aa
Unique PPE
(NXGXGNXG)n

PE_PGRS (M. tuberculosis Rv1818c)
MSFVVTIPEALAAVATDLAGIGSTIGTANAAAAVPTTTVLAAAADEVSAAM
AALFSGHAQAYQALSAQAALFHEQFVRALTAGAGSYAAAEAASAAPLEGVLAALFSGHAQAYQALSAQAALFHEQFVRALTAGAGSYAAAEAASAAPLEGVL
DVINA PALALLGRPLIGNGANGAPGTGANGGDGGILIGNGGAGGSGAAGMP
GGNGGAAGLFGNGGAGGAGGNVASGTAGFGGAGGAGGLLYGAGGAGGAGGR
AGGGVGGIGGAGGAGGNGGLLFGAGGAGGVGGLAADAGDGGAGGDGGLFFG
VGGAGGAGGTGTNVTGGAGGAGGNGGLLFGAGGVGGVGGDGVAFLGTAPGG
PGGAGGAGGLFGVGGAGGAGGIGLVGNGGAGGSGGSALLWGDGGAGGAGGV
GSTTGGAGGAGGNAGLLVGAGGAGGAGALGGGATGVGGAGGNGGTAGLLFG
AGGAGGFGFGGAGGAGGLGGKAGLIGDGGDGGAGGNGTGAKGGDGGAGGGA
ILVGNGGNGGNAGSGTPNGSAGTGGAGGLLGKNGMNGLP
Brennan and Delogu TRENDS in Microbiology Vol.10 No.5 May 2002

PE_PGRS proteins are secreted by a specialized
pathway variously named ESAT-6-, SNM-, ESX-5, or
type VII (T7S) secretion system
Bitter et al. (2009) PLoS Pathog 5(10): e1000507. doi:10.1371/journal.ppat.1000507

Kolonijų morfologija
(A) M. chubuense
(B) M. gilvum
(C) M. obuense.
(D) M. parafortuitum.
(E) M. vaccae.
On the left are shown smooth
and rough colonies grown on and rough colonies grown on
TSA plates.
On the right, the growth on TSB
medium of an isolated
colony taken from TSA is
shown.
The size of the scale bars is 2
mm.
Julian et al. J BACTERIOL 2010, 192: 1751
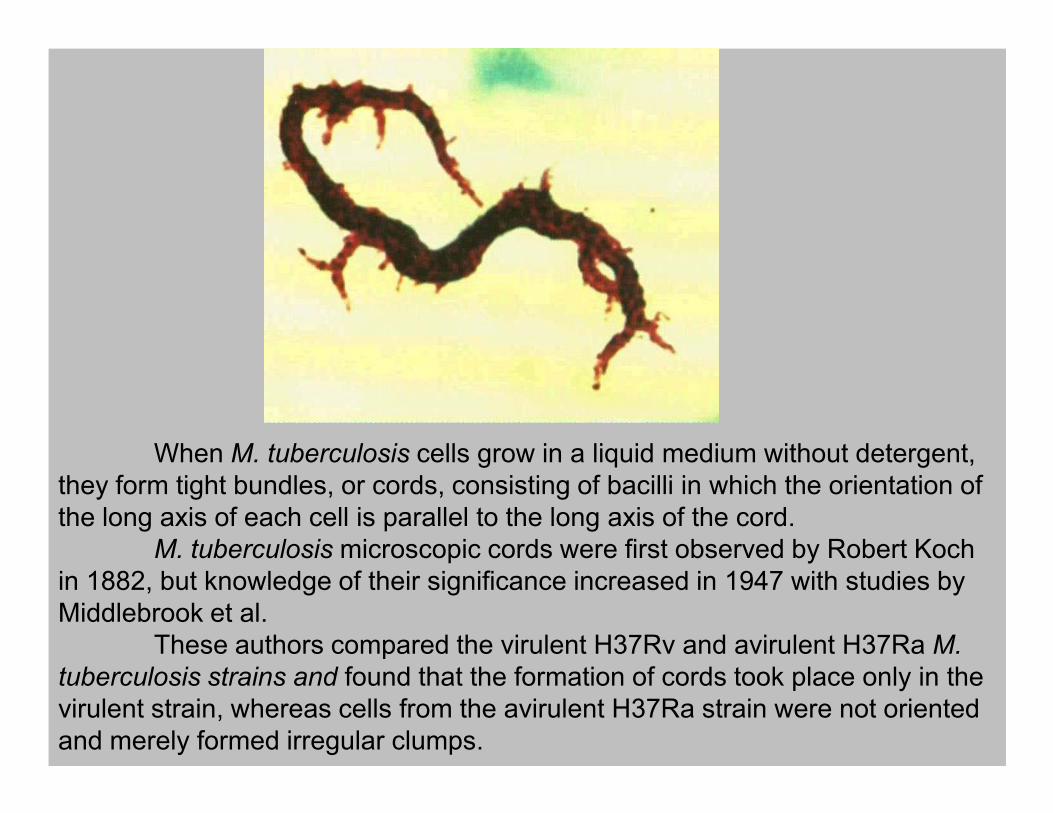
When M. tuberculosis cells grow in a liquid medium without detergent,
they form tight bundles, or cords, consisting of bacilli in which the orientation of
the long axis of each cell is parallel to the long axis of the cord.
M. tuberculosis microscopic cords were first observed by Robert Koch
in 1882, but knowledge of their significance increased in 1947 with studies by
Middlebrook et al.
These authors compared the virulent H37Rv and avirulent H37Ra M.
tuberculosis strains and found that the formation of cords took place only in the
virulent strain, whereas cells from the avirulent H37Ra strain were not oriented
and merely formed irregular clumps.
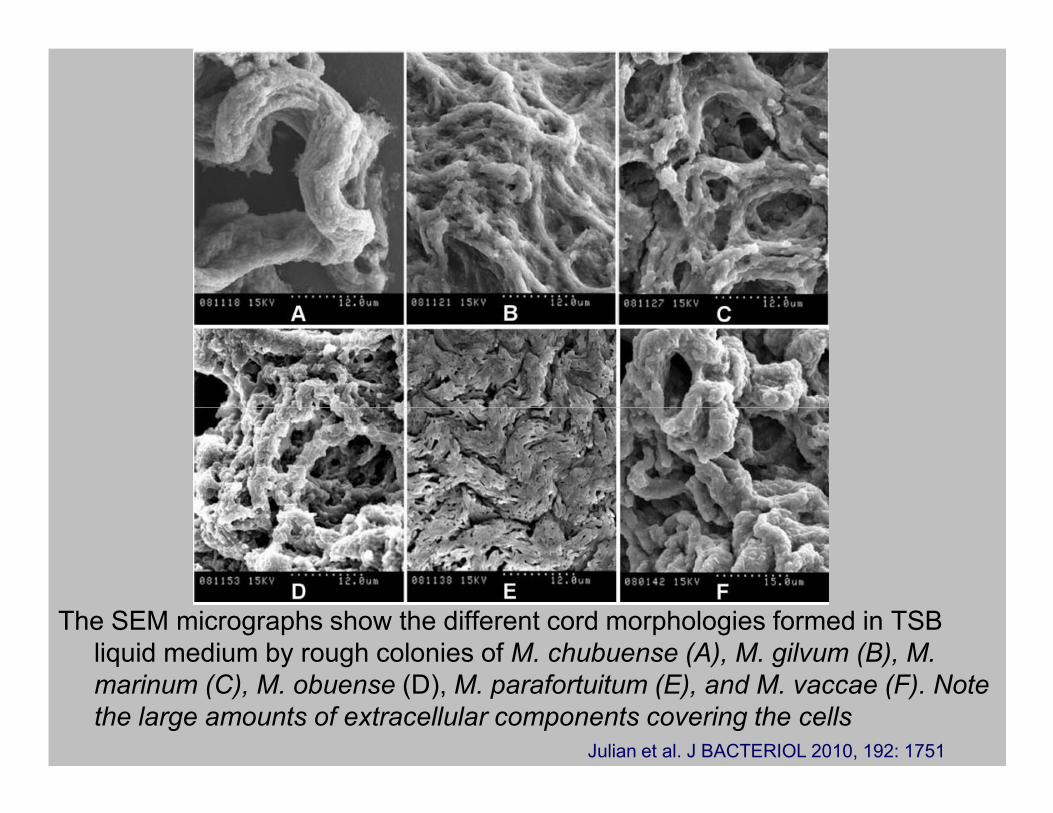
The SEM micrographs show the different cord morphologies formed in TSB
liquid medium by rough colonies of M. chubuense (A), M. gilvum (B), M.
marinum (C), M. obuense (D), M. parafortuitum (E), and M. vaccae (F). Note
the large amounts of extracellular components covering the cells
Julian et al. J BACTERIOL 2010, 192: 1751

• SEM micrographs at high magnification show details of cords formed in
liquid medium by rough colonies of M. chubuense (A), M. gilvum (B), M.
marinum (C), M. obuense (D), M. parafortuitum (E), and M. vaccae (F)
Julian et al. J BACTERIOL 2010, 192: 1751

Trehaloz÷s dimikolatas
In 1953, Bloch isolated a toxic glycolipid from M.
tuberculosis and related it to the virulence of the tubercle
bacillus and to cording.
Bloch named the glycolipid cord factor, and later, it was
identified as trehalose dimycolate (TDM)
Julian et al. J BACTERIOL 2010, 192: 1751
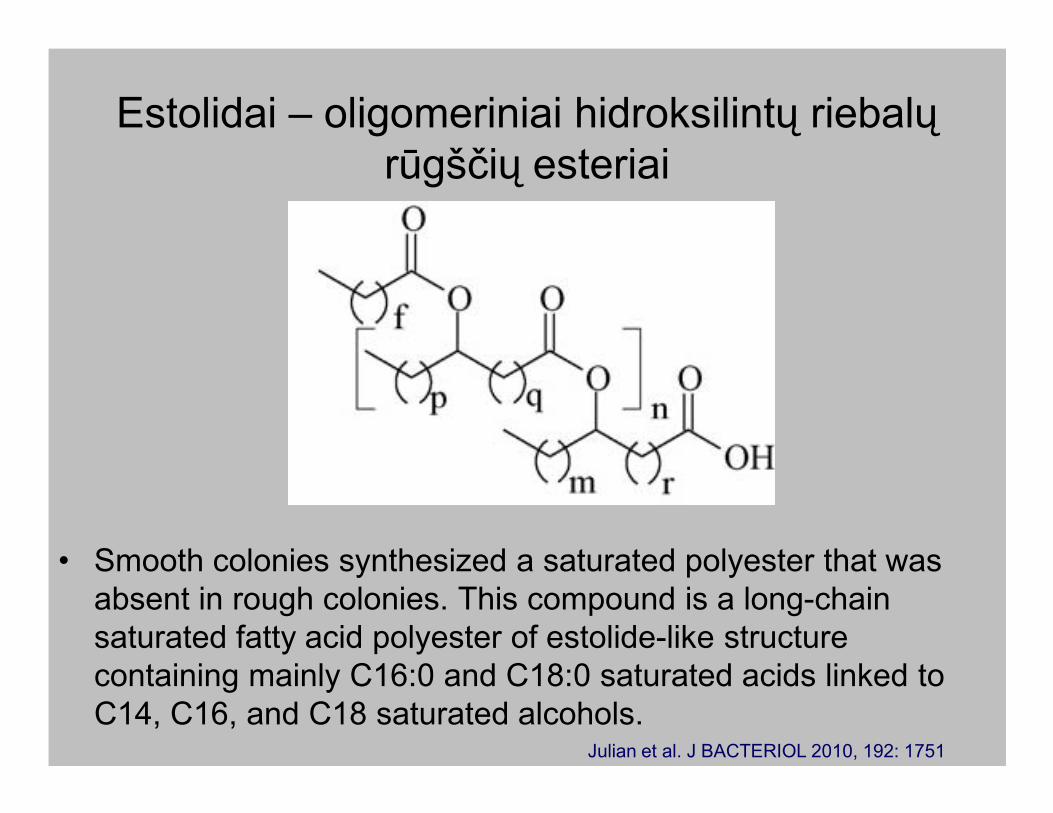
Estolidai – oligomeriniai hidroksilintų riebalų
rūgščių esteriai
• Smooth colonies synthesized a saturated polyester that was
absent in rough colonies. This compound is a long-chain
saturated fatty acid polyester of estolide-like structure
containing mainly C16:0 and C18:0 saturated acids linked to
C14, C16, and C18 saturated alcohols.Julian et al. J BACTERIOL 2010, 192: 1751

Visualization of the capsule in its native state
• Cryo electron micrographs of intact M. tuberculosis (D), M. marinum (E), and
M. bovis BCG (F). The density profile in C’ was obtained from the image.
Arrow heads point to plasma membrane (PM; magenta) and outer
membrane/mycomembrane (MOM; blue). Scale bars: 100 nm.
doi:10.1371/journal.ppat.1000794.g006

The spatial organization of the mycobacterial cell envelope
exhibiting the capsule
doi:10.1371/journal.ppat.1000794.g006

Schematic
representation
of
M. tuberculosis
cell envelope
Three forms of mycolic acids are depicted. α-Mycolates are the most abundant form in M. tuberculosis (orange).
It has 2 cyclopropane rings (triangles) in cis configuration. Oxygenated mycolates (keto- and methoxy-, shown in
red) have one cyclopropane ring each that is in either cis or trans configuration. They are covalently linked to the
arabinogalactan layer, which is linked to the peptidoglycan layer. Other lipid complexes in the cell wall include
acyl glycolipids (including trehalose dimycolate) and other complex free lipids (e.g., phthiocerol
dimycocerosate) as well as sulfolipids. Lipoarabinomannan is shown linked to the plasma membrane via a
phosphodiester bond.
L. W. Riley J. Clin. Invest. (2006) 116:1475

The M. tuberculosis cell wall is composed of peptidoglycan (PG),
arabinogalactan (AG), mycolic acids and lipoglycans such as
lipoarabinomannan (LAM). Other cell wall associated lipids include trehalose
dimycolate (TDM), trehalose monomycolate (TMM), phthiocerol dimycocerosate
(PDIM) and di-acyl trehalose (DAT).
http://biosciences-people.bham.ac.uk/About/staff_profiles_research.asp?ID=119

Chemical structures of the major mycolic acids
of M. tuberculosis and BCG Russia
• Cyclopropane rings and methyl branches are shown and annotated with
the methyltransferase responsible for their synthesis. BCG Pasteur lacks
methoxymycolates due to a mutation in MmaA3Barkan et al. Chem Biol. 2009 16(5): 499

Fatty acid/mycolic acid biosynthesis in mycobacteria
FAS-I is involved in the
synthesis of C16 and C26.
The C16 acyl-CoA product
acts as a substrate for the
synthesis of meromycolic
acids by FAS-II, whereas the
C26 fatty acid constitutes the
alpha-branch of the final
mycolic acid.
MtFabH has been proposed to
be the link between FAS-I and
FAS-II by converting C14-CoA
FAS-II consists of the condensing enzymes KasA and KasB, the keto-reductase MabA, an unidentified
dehydratase, and the enoyl-reductase InhA. Finally, the polyketide synthase Pks13 catalyses the condensation of
the alpha-branch and the meromycolate to produce mycolic acids. Targets for the action of activated isoniazid
(INH), ethionamide (ETH), triclosan (TRC), or thiolactomycin (TLM) are indicated. FAS-II enzymes are labelled in
black, excepted the condensing enzymes, which are indicated in red. The relative contribution of FAS-I and FAS-II
activities in fatty acid/mycolic acid biosynthesis is represented in green and purple respectively.
Bhatt et al. Molecular Microbiology (2007) 64(6), 1442
FAS-II by converting C14-CoA
generated by FAS-I to C16-
AcpM, which is channelled
into the FAS-II cycle ultimately
leading to meromycolates
(C56).

Proposed scheme
for the biosynthesis
of mycolic acids
• Asymmetrical carbons of the
mycolic motif have a R,R
configuration.
• R1-CO, meromycolic chain; • R1-CO, meromycolic chain;
• R2, branch chain.
• In mycobacteria, R1-CO C40-
C60 and R2 C20-C24.
• In corynebacteria, R1-CO
C16-C18 and R2 C14-C16;
• X1, unknown acceptor of the
mycolic alpha-alkyl beta-
ketoacyl chains;
• X2, unknown acceptor of the
mycolic acyl chains.Gavalda et al. J BIOL CHEM 2009, 284: 19255

Scheme of the stepwise activity of
FadD32-Pks13 PKS and its domain
organization.
To simplify, C16 acyl chains were drawn.
FadD32 synthesizes meromycoloyl-AMPs
from the meromycolic acids and ATP (1).
The meromycoloyl chain of these
intermediates is then specifically loaded by
FadD32 onto the P-pant arm of the N-ACP
domain of Pks13 (2). This is an
acyl-AMP/ACP transacylation. The
meromycoloyl chain is then transferred onto
the KS domain (3). The extender unit
carboxyacyl-CoA is specifically loaded onto carboxyacyl-CoA is specifically loaded onto
the AT domain, which catalyzes the covalent
attachment of the carboxyacyl chain to its
active site (1) and its subsequent transfer
specifically onto the C-ACP domain (2). The
KS domain catalyzes the Claisen-type
condensation between the meromycoloyl and
the carboxyacyl chains to produce alpha-
alkyl-ketothioester linked to the C-ACP
domain (3). Then, it is likely that the
thioesterase domain catalyzes the release of
the alpha-alkyl beta-ketoacyl chain and its
transfer onto an unknown acceptor (X1) (4). Gavalda et al. J BIOL CHEM 2009, 284: 19255

Proposed reaction mechanism for ring
formation in cyclopropane synthase
• Four members of the MACS family have been identified, namely CmaA1, • Four members of the MACS family have been identified, namely CmaA1,
CmaA2, MmaA2, and PcaA. These enzymes were found to have different
selectivities.
• CmaA1: cis cyclopropanation at the distal position of α-mycolate,
• CmaA2: trans cyclopropanation at the proximal position of oxygenated
mycolate.
• MmaA2: cis cyclopropanation at the distal position of α-mycolate or at the
proximal position of oxygenated mycolate.
• PcaA: cis cyclopropanation at the proximal position of α-mycolate.
Liao et al. Biochemistry, 2011, DOI: 10.1021/bi101493p

Trehaloz÷s dimikolatas
Julian et al. J BACTERIOL 2010, 192: 1751

Model of trehalose recycling as an accessory
component in mycolic acid processing
• AG, arabinogalactan layer; Ag85, antigen 85 complex; CM, cytoplasmic
membrane; PG, peptidoglycan layer; TMM, trehalose monomycolate;
TDM, trehalose dimycolate.
Kalscheuer et al. Proc Natl Acad Sci U S A. 2010 107: 21761

Phthiocerol dimycocerosates
(DIMs) and glycosylated
phenolphthiocerol
dimycocerosates, also called dimycocerosates, also called
phenolic glycolipids (PGLs)

Structures of the DIM A and DIM B produced by M. tuberculosis and M. bovis BCG and
of the glycosylated phenolphthiocerol dimycocerosates produced by M. tuberculosis
(PGL-tb) and M. bovis BCG (mycoside B). p, p’ = 3–5; n, n’ = 16–18; m1 = 20–22; m2 =
15–17; R = C2H5 or CH3.
R. Simeone et al. FEBS Journal 277 (2010) 2715

Schematic representation of the roles of the FadD proteins encoded by the DIM + PGL locus in the biosynthesis of
DIMs and related compounds in Mycobacterium tuberculosis. R = C2H5 or CH3. KS, ketoacylsynthase; AT,
acyltransferase; DH, dehydratase; ER, enoylreductase; KR, ketoreductase; ACP, acyl carrier protein.
R. Simeone et al. FEBS Journal 277 (2010) 2715

Schematic
representation of
the general
structure of the
mycobacterial
cell wall

Proposed biosynthetic pathway of phosphatidyl-myo-inositol
(PI) and archaetidyl-myo-inositol (AI) in Bacteria and Archaea
(solid arrow), and Eucarya (broken arrow)
1: 1L-myo-Inositol 1-phosphate synthase; 2: Phosphatidyl-myo-inositol phosphate (PIP)
synthase/Archaetidyl-myo-inositol phosphate (AIP) synthase; 3: PIP phosphatase/AIP
phosphatase; 4: 1L-myo-Inositol 1-phosphate phosphatase; 5: Phosphatidyl-myo-inositol
(PI) synthase; 6: Inositol kinase.
Morii et al. J. Biochem. 2010;148(5):593

Schematic representation of LM and LAM biosynthetic pathway in Mtb
First mannosylation occurs at C-2 position of myo-inositol to form PIM1 and second
mannosylation at C-6 position of myo-inositol in PIM1 or AcPIM1. First acylation occurs at C-6
position of Man in PIM1. CDP-DAG, CDP-diacylglycerol; ManT, mannopyranosyltransferase;
AraT, arabinofuranosyltransferase; PPM, polyprenolmonophosphomannose; DPA, decaprenyl-1-
monophosphoryl-arabinose.UMESIRI ET AL.
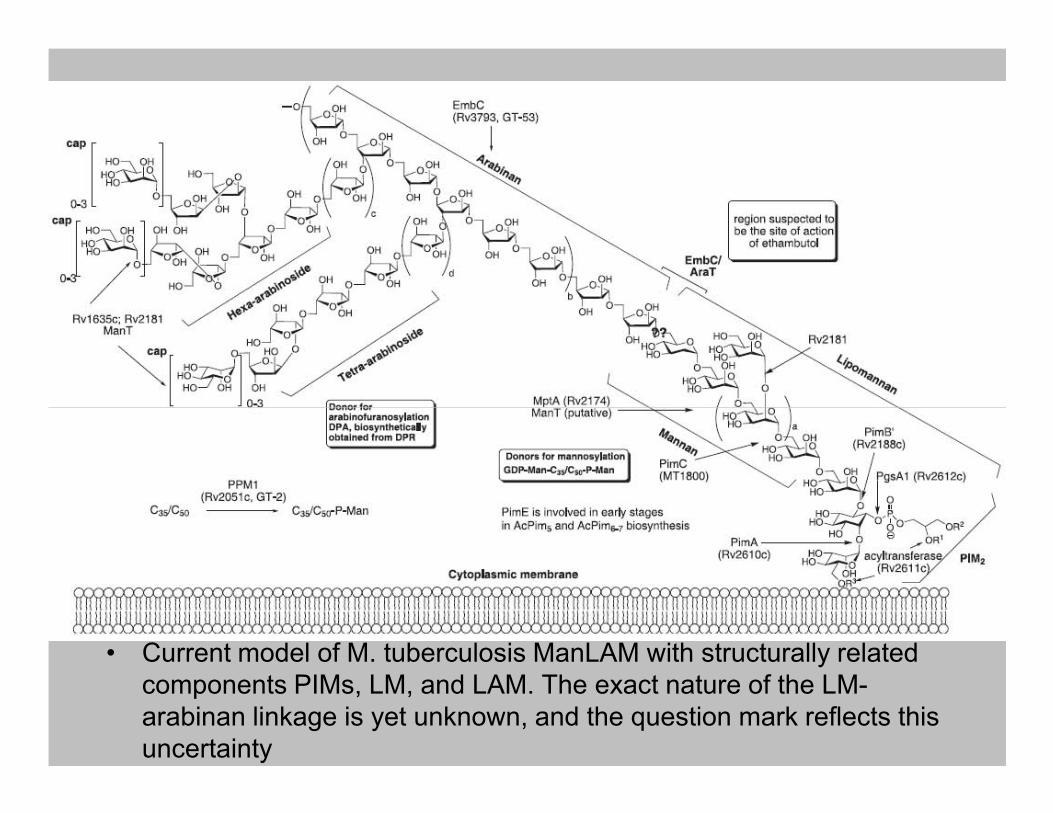
• Current model of M. tuberculosis ManLAM with structurally related
components PIMs, LM, and LAM. The exact nature of the LM-
arabinan linkage is yet unknown, and the question mark reflects this
uncertainty

The M. tuberculosis cell wall is composed of peptidoglycan (PG),
arabinogalactan (AG), mycolic acids and lipoglycans such as
lipoarabinomannan (LAM). Other cell wall associated lipids include trehalose
dimycolate (TDM), trehalose monomycolate (TMM), phthiocerol dimycocerosate
(PDIM) and di-acyl trehalose (DAT).
http://biosciences-people.bham.ac.uk/About/staff_profiles_research.asp?ID=119
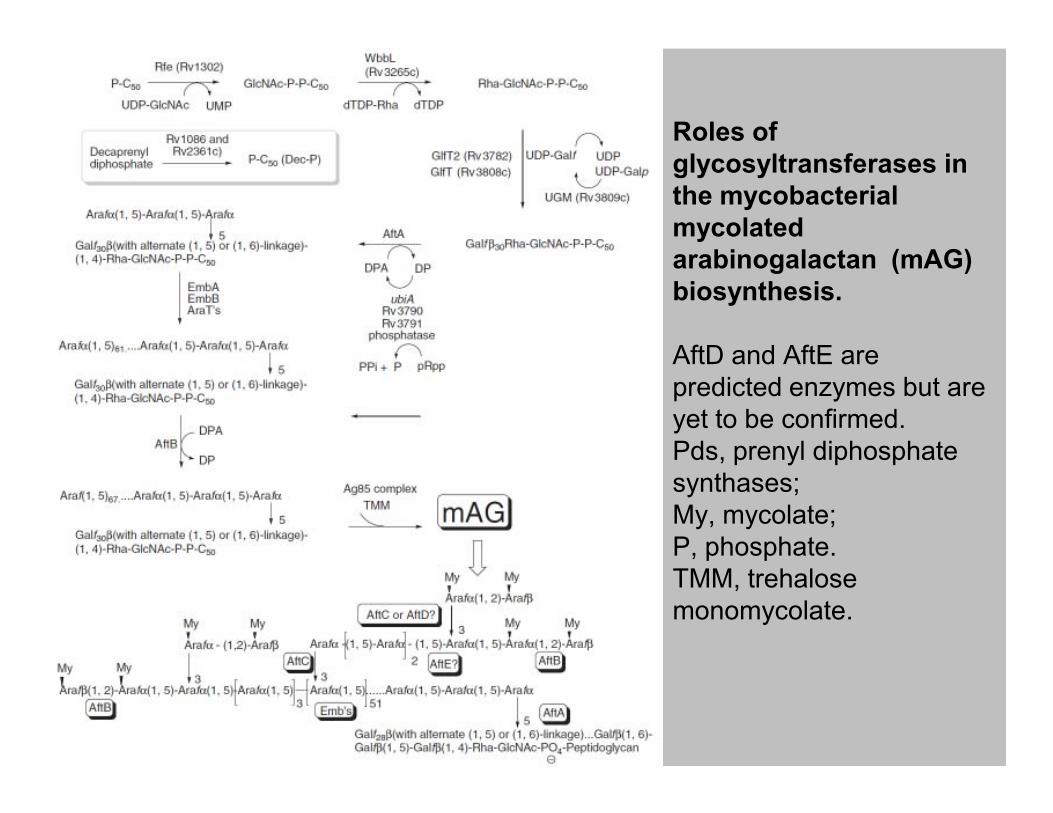
Roles of
glycosyltransferases in
the mycobacterial
mycolated
arabinogalactan (mAG)
biosynthesis.
AftD and AftE are
predicted enzymes but are
yet to be confirmed. yet to be confirmed.
Pds, prenyl diphosphate
synthases;
My, mycolate;
P, phosphate.
TMM, trehalose
monomycolate.

Proposed model for the biosynthesis of the GalN substituent of AG
PG, peptidoglycan; AG, arabinogalactan; PP, polyprenol phosphate; PP-GalNAc,
polyprenylmonophosphoryl-N-acetyl-galactosaminyl; PP-GalN, polyprenyl-monophosphoryl-
galactosaminyl; PPMan, polyprenyl-phosphomannose.
GT-C1 located on the cytosolic side of the plasma membrane is involved in the transfer of a
mannose residue from GDP-α-D-Manp to polyprenyl phosphate; GT-C2 is predicted to be in the
last extra-cytoplasmic loop of the protein and to catalyze the transfer of D-GalN(Ac) onto AG.
http://www.jbc.org/cgi/doi/10.1074/jbc.M110.188110
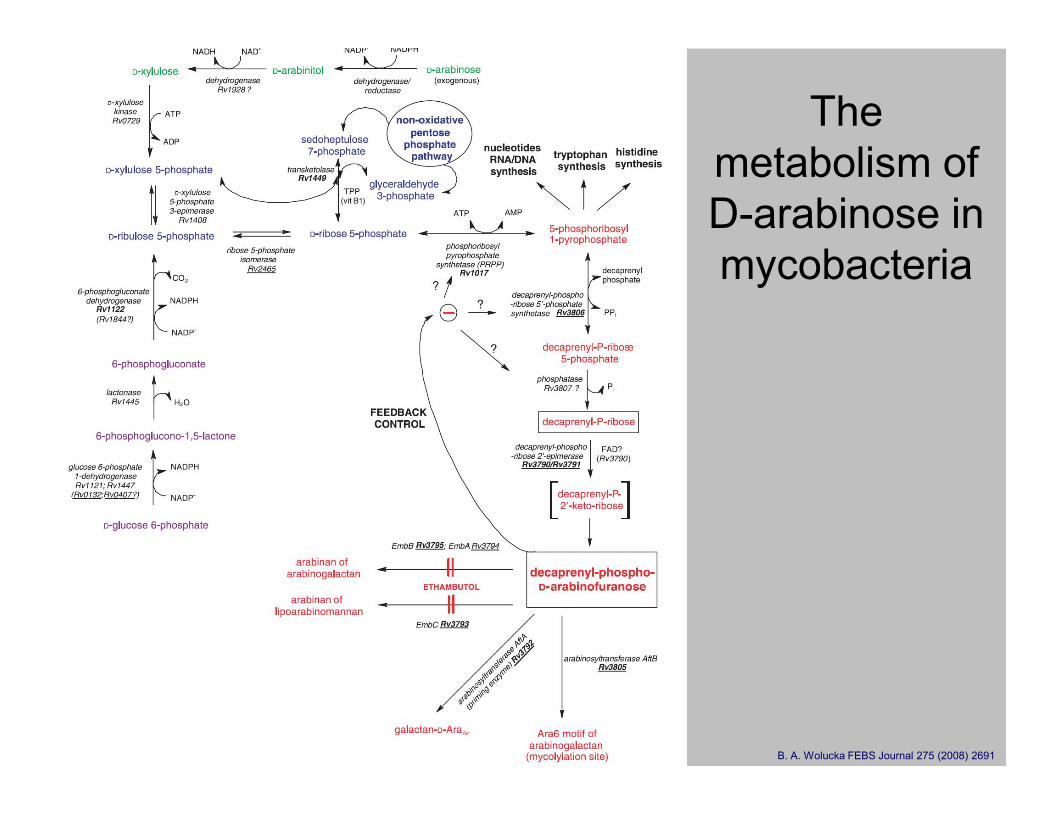
The
metabolism of
D-arabinose in
mycobacteria
B. A. Wolucka FEBS Journal 275 (2008) 2691

Decaprenyl phosphate and decaprenyl-
phospho-monosaccharides of mycobacteria (A) The mycobacterial lipid
carrier C50-decaprenyl
phosphate has a unique
stereoconfiguration and contains
only one trans (E)-isoprene
residue at its omega-end .
(B) Decaprenyl-phospho-
arabinose, the only known D-
arabinose donor for the
synthesis of the cell-wall
arabinogalactan and
lipoarabinomannan in
mycobacteria.
(C) Decaprenyl-phospho-ribose,
the direct precursor of the beta-
D-arabinofuranosylmono-
phosphodecaprenol donor (B)
and the major form of the
naturally occurring decaprenyl-
phospho-sugars of mycobacteria
(D) The mycobacterial
decaprenylphospho-mannose, a
minor componentB. A. Wolucka FEBS Journal 275 (2008) 2691

Diterpenai
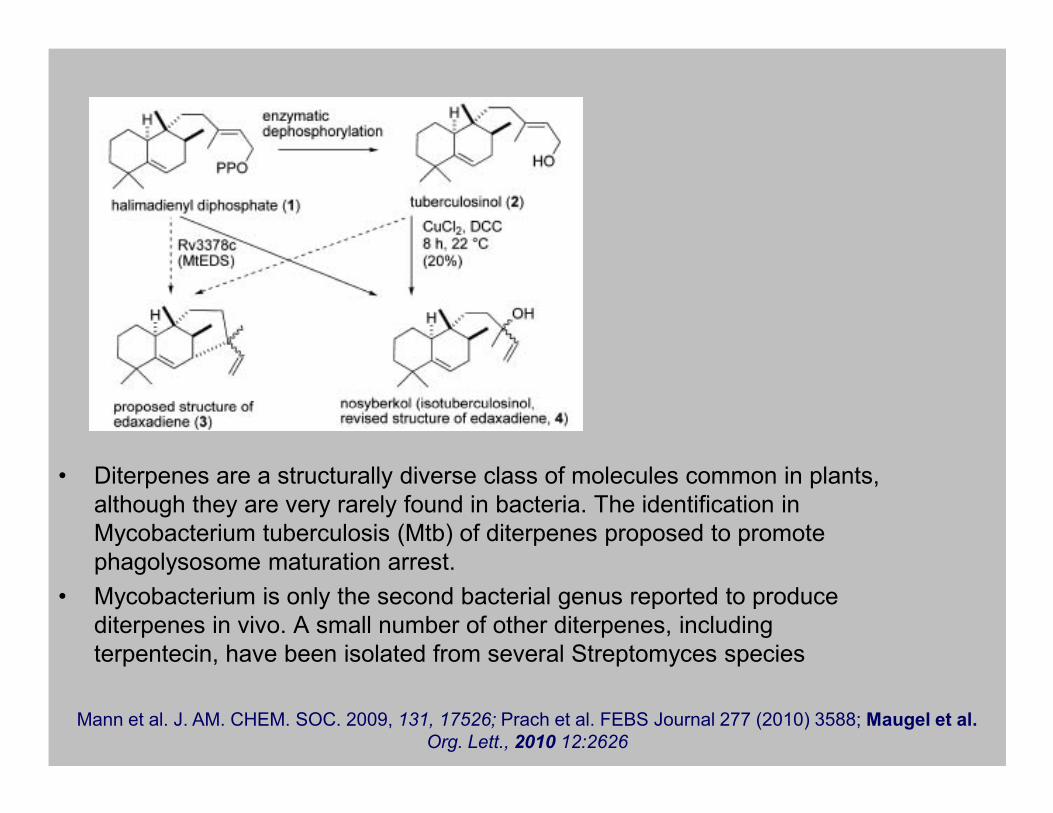
• Diterpenes are a structurally diverse class of molecules common in plants,
although they are very rarely found in bacteria. The identification in
Mycobacterium tuberculosis (Mtb) of diterpenes proposed to promote
phagolysosome maturation arrest.
• Mycobacterium is only the second bacterial genus reported to produce
diterpenes in vivo. A small number of other diterpenes, including
terpentecin, have been isolated from several Streptomyces species
Mann et al. J. AM. CHEM. SOC. 2009, 131, 17526; Prach et al. FEBS Journal 277 (2010) 3588; Maugel et al.
Org. Lett., 2010 12:2626

Anti-aging creams

Top 10 botanical ingredients in
2010 anti-aging creams• Some of the most common botanicals that
are hot off the market are:
• Rosmarinus officinalis, Vitis vinifera (grape
seed extract), Citronellol, Limonene, seed extract), Citronellol, Limonene,
Oenothera biennis (evening primrose),
Glycyrrhiza glabra (licorice extract),
Aframomum angustifolium seed extract,
Diosgenin (wild yam), N6 furfuryladenine
(kinetin), and Ergothioneine. Cronin and Draelos J Cosmet Dermatol. 2010 9(3):218

• Ergothioneine has been the subject of numerous studies
following its isolation from ergot in 1909, greatly following its isolation from ergot in 1909, greatly
stimulated by studies showing that it accumulates in
animal blood and tissue.
• The occurrence, chemistry, and biosynthesis of
ergothioneine have been reviewed, but most studies
have involved eukaryotic systems, and results with
prokaryotes are limited.

Ergothioneine
• Aerobic cells depend on cysteine derivatives to control intracellular redox potential
and metal homeostasis and to fend off electrophilic toxins. In addition to their main
intracellular thiols, such as glutathione and mycothiol, certain fungi and
mycobacteria also produce ergothioneine
• The function ergothioneine plays in microbial cells is not well understood, but
recent findings point to critical functions in human physiology. recent findings point to critical functions in human physiology.
• The human body absorbs ergothioneine from dietary sources and concentrates it in
specific tissues or cells such as liver, kidney, central nervous system, and red blood
cells.
• A cation transporter (OCTN1) with high specificity for ergothioneine is responsible
for this nonuniform distribution, and both OCTN1 hyperactivity and OCTN1
deficiency exert negative effects on human cells.
• Despite these recent discoveries, the precise function of 1 in human tissue is still a
matter of debate.

• The mycobacterial ergothioneine gene cluster codes for aγ-glutamyl cysteine
synthetase (EgtA), an FGE-like protein (EgtB), a glutamine amidotransferase (EgtC),
a methyltransferase (EgtD), and a PLPbinding protein (EgtE).
• Reaction sequence of ergothioneine biosynthesis with hercynine (2), 3, and
hercynylcysteine sulfoxide (4) as intermediates.
• A previously suggested abbreviated pathway in N. crassa is shown in green
• A blastp search for this pair of enzymes suggests that production of 1 is a common
trait among actinobacteria, cyanobacteria, pezizomycotina, and basidiomycota but
also occurs in numerous bacteroidetes and alpha-, beta-, γ-, and δ-proteobacteria
Seebeck J. AM. CHEM. SOC. 2010, 132, 6632

Tricarboxylic acid (TCA) cycle• Aerobic organisms have a tricarboxylic acid (TCA) cycle that is functionally
distinct from those found in anaerobic organisms.
• The aerobic pathogen Mycobacterium tuberculosis lacks detectable alpha-
ketoglutarate (KG) dehydrogenase activity and drives a variant TCA cycle in
which succinyl-CoA is replaced by succinic semialdehyde.
• M. tuberculosis expresses a CoA-dependent KG dehydrogenase activity, albeit
one that is typically found in anaerobic bacteria. Unlike most enzymes of this
family, the M. tuberculosis KG: ferredoxin oxidoreductase (KOR) is extremely
stable under aerobic conditions. stable under aerobic conditions.
• This activity is absent in a mutant strain deleted for genes KOR.
• Interestingly, inhibition of the glyoxylate shunt or exclusion of exogenous fatty
acids alleviates this growth defect, indicating the presence of an alternate
pathway that operates in the absence of beta-oxidation.
• Simultaneous disruption of KOR and the first enzyme of the succinic
semialdehyde pathway (KG decarboxylase; KGD) results in strict dependence
upon the glyoxylate shunt for growth.
• Unlike most organisms M. tuberculosis utilizes two distinct TCA pathways from
KG, one that functions concurrently with beta-oxidation (KOR-dependent), and
one that functions in the absence of beta-oxidation (KGD-dependent).
Baughn et al. PLoS Pathog 5(11): e1000662.

Integrated model of routes and regulation in the M. tuberculosis
TCA cycle
• The glyoxylate cycle (inner cycle), canonical TCA cycle (medial cycle), and variant TCA cycle (outer cycle)
are depicted. Blue lines indicate pathways that are utilized concurrently with beta-oxidation, green lines
indicate pathways that are utilized during growth on carbohydrates as the sole carbon source, and black lines
indicate pathways that are common to both modes of growth. Red lines indicate blocks imposed by 3NP on
isocitrate lyase (ICL), PknG on GarA, and GarA on KGD. The dotted red lines represent the putative blocks
imposed by glyoxylate on SSA dehydrogenase and PknG.

O kur katabolin÷ represija?

• Metabolic adaptation to the host environment is a defining feature of the
pathogenicity of Mycobacterium tuberculosis (Mtb), but we lack biochemical
knowledge of its metabolic networks. Many bacteria use catabolite repression as a
regulatory mechanism to maximize growth by consuming individual carbon
substrates in a preferred sequence and growing with diauxic kinetics.
• Surprisingly, untargeted metabolite profiling of Mtb growing on 13C-labeled carbon • Surprisingly, untargeted metabolite profiling of Mtb growing on 13C-labeled carbon
substrates revealed that Mtb could catabolize multiple carbon sources
simultaneously to achieve enhanced monophasic growth.
• Moreover, when co-catabolizing multiple carbon sources, Mtb differentially
catabolized each carbon source through the glycolytic, pentose phosphate, and/or
tricarboxylic acid pathways to distinct metabolic fates.
• This unusual topologic organization of bacterial intermediary metabolism has not
been previously observed and may subserve the pathogenicity of Mtb.
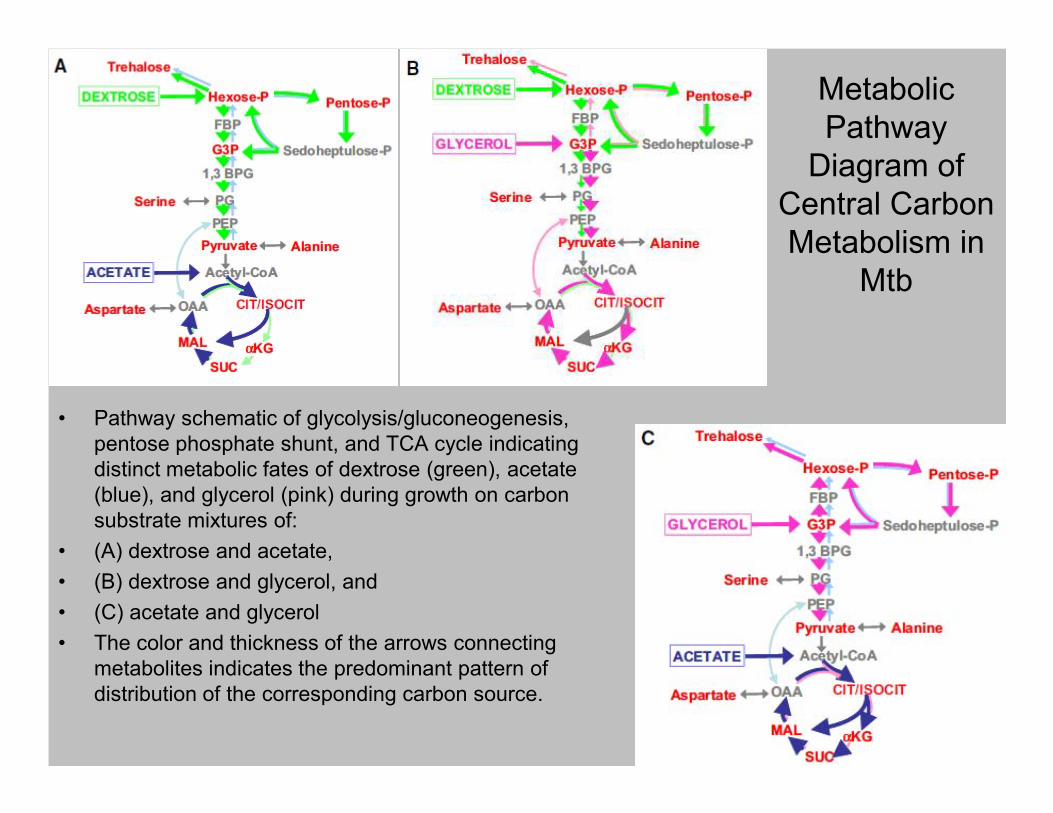
Metabolic
Pathway
Diagram of
Central Carbon
Metabolism in
Mtb
• Pathway schematic of glycolysis/gluconeogenesis, • Pathway schematic of glycolysis/gluconeogenesis,
pentose phosphate shunt, and TCA cycle indicating
distinct metabolic fates of dextrose (green), acetate
(blue), and glycerol (pink) during growth on carbon
substrate mixtures of:
• (A) dextrose and acetate,
• (B) dextrose and glycerol, and
• (C) acetate and glycerol
• The color and thickness of the arrows connecting
metabolites indicates the predominant pattern of
distribution of the corresponding carbon source.

A model for the Eut microcompartment and its
metabolic pathway
• (A) A hypothetical model of the Eut microcompartment emphasizing the construction of a semiregular
polyhedron primarily from hexameric shell proteins.
• (B) A model for the metabolism of ethanolamine in the Eut microcompartment. Ethanolamine is converted
into ethanol, acetyl-phosphate, and acetyl–coenzyme A (CoA). The volatile intermediate, acetaldehyde
(boxed in orange), is consumed before it can escape the protein shell.
• The four homologous shell proteins belonging to the conserved BMC family (EutK, EutL, EutM, and EutS) are
colored in light blue.
• EutBC, ethanolamine ammonialyase; EutD, phosphotransacetylase; EutE, aldehyde dehydrogenase; EutG,
alcohol dehydrogenase. Tanaka, et al. Science 327, 81 (2010);

• The open (left) and closed (right) configurations of EutL
trimers are shown in both ribbon diagram and surface
representations.

Model for the structure and function of the
Pdu MCP
• (A) Organization of the pdu operon. At least 14 pdu genes (colored) encode proteins that are components of
purified Pdu microcompartments (MCPs ). Asterisks indicate genes that encode polypeptides having potential
N-terminal targeting sequences. Seven genes (blue and cyan) encode shell proteins . Homologues of the
bacterial microcompartment (BMC) family of shell proteins are shown in blue.
• (B) Electronmicrograph of purified Pdu MCPs from S. enterica. (Scale bar: 100 nm).
• (C) Model for B12-dependent 1,2-PD degradation by Salmonella. 1,2-PD is metabolized within the MCP
lumen, first to propionaldehyde (PA) and then to propionyl-CoA (PrpCoA). The enzymes that localize to the
MCP interior include coenzyme B12-dependent diol dehydratase (PduCDE) and PduP, as well as
adenosyltransferase (PduO) and a reactivase (PduGH) that are required for the maintenance of diol
dehydratase activity. The proposed function of the Pdu MCP is to sequester propionaldehyde to minimize its
toxicity.

A case of Dop-ing in bacterial
pupylation

Prokaryotic
ubiquitin protein
(PUP)A simplified comparison of the
eukaryotic ubiquitination (A) and
prokaryotic PUPylation (B)
proteasomal degradation
pathways. In eukaryotes, the
backbone C-terminal carboxylate
of ubiquitin is ligated to the target of ubiquitin is ligated to the target
lysine via E1, E2 and E3 ubiquitin
ligases followed by subsequent
proteasomal degradation. In
prokaryotes, the side chain
amine of the C-terminal
glutamine is deamidated by Dop
followed by ligation to the target
lysine by PafA. The AAAATPase
facilitates translocation of
PUPylated protein into the
proteasomal core where the
protein is degraded.

Ir pabaigai apie benziną
• Methyl tert-butyl ether (MTBE) has been incorporated in
reformulated gasoline at concentrations up to 15% (vol/vol) to
replace lead tetraethyl in order to comply with the octane index and
to reduce the polluting emissions in exhaust gases.
• Other oxygenates, such as ethyl tert-butyl ether (ETBE) and tert-
amyl methyl ether (TAME) and their corresponding alcohols, tert-amyl methyl ether (TAME) and their corresponding alcohols, tert-
butyl alcohol (TBA) and tert-amyl alcohol (TAA), can play the same
role regarding the octane index.
• MTBE is the dominant fuel oxygenate, with a worldwide production
capacity of around 25 million tons.
• Because of its widespread use and the high frequency of
underground tank leakage, this compound is now the second most
commonly detected contaminant in urban groundwater in the United
States. In Europe, detectable levels of MTBE in rivers have also
been reported.
DOI: 10.1128/AEM.68.6.2754–2762.2002

Pathway for MTBE degradation in Mycobacterium
austroafricanum IFP 2012
DOI: 10.1128/AEM.68.6.2754–2762.2002





























