Embed Size (px)
Citation preview

Nematodes associated with phytodetritus in salt marshes. Colonization patterns and species- and diversity related impacts on
decomposition processes
Nematoden geassocieerd met fytodetritus in schorren. Kolonisatiepatronen en soort- en diversiteitsafhankelijke invloed op
afbraakprocessen
Ilse De Mesel
Proefschrift voorgelegd tot het behalen van de graad van Doctor in de Wetenschappen (Biologie)
Promotor: Prof. Dr. Magda Vincx Co-promotor: Prof. Dr. Ir. Jean Swings
Dr. Tom Moens

Table of contents
Acknowledgements – Dankwoord … … … … … … … … ...… … … … … … … … … … … i
Summary… … … … … … … … … … … … … … ..… … … … … … … … … … … ...… … … .iii
Samenvatting … … … … … … … … … … … … … … … .… … … … … … … … .… … … ...vii
Chapter 1: General Introduction and Aims… … … ...… … … … … … … … … … … … 1
Introduction… … … … … … … … … … … … … … … … … … … … … … … … … … … … … 3 Topics and outline of the thesis… … … … … … … … … … … … … … … … … … … … .… .6
Chapter 2: Influence of bacterivorous nematodes on the decomposition of cordgrass … … … … … … … … … … … … … … … … … … … … … … … … … … … … … .9
De Mesel I, Derycke S, Swings J, Vincx M, Moens T (2003) Influence of bacterivorous nematodes on the decomposition of cordgrass. Journal of
Experimental Marine Biology and Ecology 296: 227-242
Abstract… … … … … … … … … … … … … … … … … … … … … … … … … … … … … … 11 Introduction… … … … … … … … … … … … … … … … … … … … … … … … … … … … ..13 Materials and Methods… … … … … … … … … … … … … … … … … … … … … ...… … ..14
Experimental set-up… … … … … … … … … … … … … … … … … … … … … ...14 Sampling… … … … … … … … … … … … … … … … … … … … … … .… … … ...15 Weight loss… … … … … … … … … … … … … … … … … … … … … … … … ....16 C/N-analysis… … … … … … … … … … … … … … … … … … … … … … ..… … 16 Nematodes… … … … … … … … … … … … … … … … … … … … … … … ..… ...17 Microbial activity… … … … … … … … … … … … … … … … … … … … … … ..17 Statistical analysis… … … … … … … … … … … … … ...… … … … … … … ......17
Results… … … … … … … … … … … … … … … … … … … … … … … … … … … … … … ..18 Decomposition… … … … … … … … … … ...… … … … … … … … … … … … … 18 Nematodes… … … … … … … … … … … … … … … … … … … ...… ...… … … ...21 Microbial activity… … … … … … … … … … … … … … … … … … … … ...… ...22
Discussion… … … … … … … … … … … … … … … … … … … … … … … … … … … … … 24 Decomposition process… … … … … … … … … … … … … … … … … ..… … … 24 Microbial activity… … … … … … … … … … … … … … … … … … … ..… … … 25
Nematodes and their influence on the microbial activity and decomposition process… … … … … … … … … … … … … ...… … … … … … … … … … … … ....25 Conclusions… … … … … … … … … … … … … … … … … … … … … … … … ...28
Acknowledgements… … … … … … … … … … … … … … … … … … … … … … … ...… ...29

Chapter 3: Top-down impact of bacterivorous nematodes on the bacterial
community structure: a microcosm experiment… … … … … … … ...… … … … … ...31
De Mesel I, Derycke S, Moens T, Van der Gucht K, Vincx M, Swings J (in press) Top-down impact of bacterivorous nematodes on the bacterial community
structure: a microcosm experiment. Environmental Microbiology
Abstract… … … … … … … … … … … ..… … … … … … … … … … … … … … … … … … ..33 Introduction… … … … … … … … … … … … … … … … … … … … … … … … … … ..… … 35 Materials and Methods… … … … … … … … … … … … … … … … … … … … ...… … … ..37
Experimental set-up and sampling… … … … … … … … … … … … … .… … ...37 DNA-extraction and – purification… … … … … … … … … … … … … … … … .38 PCR and DGGE… … … … … … … … … … … … … … … … … … … … … … … 38 Sequencing of excised DGGE bands… … … … … … … … … … … … … … … .39 Bacterial diversity… … … … … … … … … … … … … … … … … … … … ...… ..39 Statistical analysis of the DGGE fingerprints… ...… … … … … … … … … … 40 Nucleotide sequence accession numbers… … … … … … … … … … … … … ...40
Results and Results and Discussion… … … … … … … … .… … … … … … … … … … … 41 Effects on bacterial community composition… … … … … … … … … … … … .41 Effects on bacterial diversity… … … … … … … … … … … … … … … … … … .48 Conclusions… … … … … … … … … … … … … … … … … … … … … .… … … ..51
Acknowledgements… … … … … … … … … … … … … … … … … … … … … … … … … ..52
Chapter 4: Effect of nematode diversity on organic matter decomposition and the associated microbial community: a microcosm study… … … ..… … … … … … 53
De Mesel I, Derycke S, Swings J, Vincx M, Moens T
Abstract… … … … … … … … … … … … … … … … … … … … … … … … … … … … … ....55 Introduction… … … … … … … … … … … … … … … … … … … … … … … … … … … … ..57 Materials and Methods… … … … … … … … … … … … … … … … … … … … … … ...… ..58
Experimental set-up… … … … … … … … … … … … … … … … … … … … … ...58 Decomposition rate… … … … … … … … … … … … … … … … … … … … .… ..59 Nematodes… … … … … … … … … … … … … … … … … … … … … … … ...… ..60 Microbial activity… … … … … … … … … … … … … … … … … … … … … … ..60 Bacterial diversity… … … … … … … … … … … … … … … … … … ...… … … ..60 Statistical analysis… … … ...… … … … … … … … … … … … … … … … … ......61
Results… … … … … … … … … … … … … … … … … … … … … … … … … … … … … ......62 Nematodes… … … … … … … … … … … … … … … … … … … … … … … … .....62 Microbial activity… … … … … … … … … … … … … … … … … … … … … ......63 Bacterial diversity… … … … … … … … … … … … … … … … … … ...… … … ..67 Decomposition rate… … … … … … … … … … … … … … … … … … … … … ...69
Discussion… … … … … … … … … … … … … … … … … … … … … … … … … … … … … 73 Development of nematode populations… … … … … … … … … … … … … … .73 Effects of nematodes on microbial activity and bacterial diversity… … … ..75 Impacts on the decomposition rates… … … … … … … … … … … … … … … ..76 Conclusions… … … … … … … … … … … … … … … … … … … … … … … … ...78
Acknowledgements………………………………………………………………..…..79

Chapter 5: Development of a nematode community on decaying organic matter (Spartina anglica) under contrasting conditions.....................................................81
De Mesel I, Vanaverbeke J, Vincx M, Moens T (submitted) Development of a
nematode community on decomposing organic matter (Spartina anglica) under contrasting conditions
Abstract… … … … … … … … … … … … … … … … … … … … … … … … … … … … … … 83 Introduction… … … … … … … … … … … … … … … … … … … … … … … … … … … … ..85 Materials and Methods… … … … … … … … … … … … … … … … … … … … … … … … .86
Experimental set-up… … … … … … … … … … … … … … … … … … … … … ..86 Sampling… … … … … … … … … … … … … … … … … … … … … … … … ..… .87 Nematode community… … … … … … … … … … … … … … … … … … … … ... 87 Statistical analysis… … … … … … … … … … … ...… … … … … … … … … .… 88
Results… … … … … … … … … … … … … … … … … … … … … … … … … … … … … … ..89 Nematode densities… … … … … … … … … … … … … … … … … … … … ...… 89 Nematode community… … … … … … … … … … … … … … … … … … … … ... 91
Discussion… … … … … … … … … … … … … … … … … … … … … … … … … … … … … 95 Acknowledgements… … … … … … … … … … … … … … … … … … … … … … … … … 100
Conclusions… … … … … … … … … … … … ..… … … … … … … … … … … … … … … .101 Species manipulation experiment… … … … … … … … … … … … … … … ...103 Colonization experiment… … … … … … … … … … … … … … … … … … … .108
Appendix A...… … … … … … … … … … … … … … … … … … … … … … … … … … ....111
Cited literature… … … … … … … … … … ..… … … … … … … … … … … ..… … … … .123

Dankwoord
i
Dankwoord
Het moment is gekomen om een aantal mensen te bedanken die me de voorbije jaren
met raad en daad hebben bijgestaan. Graag wil ik beginnen bij mijn promotor Prof.
Dr. Magda Vincx en mijn co-promotor Prof. Dr. Ir. Jean Swings. Zonder jullie steun
zou van dit doctoraat geen sprake geweest zijn! Jullie hebben mij indertijd de kans
gegeven om een doctoraatsbeurs aan te vragen en zijn altijd achter mij blijven staan.
Magda, bedankt om me de vrijheid te laten ‘mijn ding’ te doen! Jean, ik ben enorm
blij dat ik op het Labo voor Microbiologie heb mogen kennis maken met de
microbiële onderzoekswereld. Jouw enthousiasme heeft me vaak deugd gedaan!
Dr. Tom Moens heeft me indertijd warm gemaakt voor dit onderzoeksproject, en daar
ben ik nog steeds heel erg blij om. De discussies rond mijn manuscripten hebben
zeker de kwaliteit van dit proefschrift omhoog gehaald.
Bedankt Prof. Dr. Ann Vanreusel voor de hulp bij mijn statistische problemen en voor
het nalezen van een aantal van mijn manuscripten, maar ook voor de babbels
tussendoor en voor het aangename gezelschap op de staalnamecampagne met de
Polarstern. Ik vond het enorm fijn dat ik van jou en Magda de kans heb gekregen
ervaring op te doen tijdens de campagnes met de Discovery en Polarstern.
Dirk Vangansbeke en Annick Van Kenhove wil ik graag bedanken voor de vele
chemische analyses, Bernard Timmerman voor de hulp bij de staalnames. Annick,
Bernard en Bart Beuselinck hebben ook heel wat staaltjes voor mij opgespoeld en
nematoden in preparaat gebracht, waarvoor mijn dank! Bedankt Gunther Van
Ryckegem voor de ergosterolanalyses.
De mensen van het oranje labo op de microbiologie, Dr. Katleen Van der Gucht, Nele
Vloemans en Sylvie Cousin verdienen hier zeker een plaatsje! Eerst en vooral omdat ik
altijd van jullie labo heb mogen gebruik maken. Nele, jij hebt me indertijd de DGGE
techniek en alles daarrond aangeleerd. Ik kon altijd op je hulp rekenen als ik weer
eens een overklaarbaar probleem had. Katleen, bedankt voor de hulp bij de
verwerking van de DGGE analyses en de suggesties bij het manuscript!
Zonder Margo Cnockaert was ik waarschijnlijk nog altijd op zoek naar die ene pipet,
dat bepaald product, dat specifieke protocol.... Margo, heel erg bedankt voor alle hulp,
tips, je luisterend oor! Het was niet altijd evident om mijn draai te vinden op het labo,
maar ik heb altijd op jou kunnen rekenen!
Verder wil ik ook iedereen van de microbiologie bedanken die me op de één of andere
manier heeft geholpen of gesteund. Ik denk hierbij speciaal aan Renata Coopman voor

Dankwoord
ii
de hulp bij de sequenties, Anne Willems voor het overleg tussen de labo’s, Evie De
Brandt voor de laatste DGGE, en natuurlijk ook Liesbeth Masco (eten? 12h?).
Ook een woordje van dank voor Prof. Dr. Jacques Vanfleteren omdat ik ettelijke
keren van zijn labo heb mogen gebruik maken. Bij Annemie De Vreese en Andy
Vierstraete kon ik altijd terecht met vragen en problemen. Jullie tips hebben mij
dikwijls een flinke stap vooruit geholpen!
In de beginjaren kon ik rekenen op Kris Hostens om alle computerproblemen op te
lossen, de laatste jaren komt Guy Desmet de wormen van mijn computer verjagen en
andere depannages uitvoeren! Ook Tim Deprez en Tom Gheskiere hebben mij al vaak
met raad en daad bijgestaan.
Ik heb het geluk gehad drie fijne thesisstudentes te mogen begeleiden: bedankt Sofie
Derycke, Annelies Phillips en Phaedra Van Brussel voor jullie enthousiasme!
Bedankt Marleen De Troch voor het lezen van een aantal van mijn manuscripten. Dr.
Peter Herman wil ik graag bedanken voor de hulp met statistische problemen en het
doornemen van één van mijn manuscripten.
Een speciaal woordje van dank ook voor de vele mensen van de mariene die
misschien niet rechtstreeks bij mijn onderzoek betrokken waren, maar die wel altijd
klaar stonden met goeie raad, bemoedigende woordjes, .... Ik denk hierbij vooral aan
Maaike Steyaert, Jan Vanaverbeke, Sofie Derous, Thomas Remerie, Jan Wittoeck,
Nancy Fockedey, Sofie Derycke, Maarten Raes, Henni Hampel, Isolde De Grem,
Annick Verween (ook voor het uitlenen van je wrak... euh fiets natuurlijk!), ....
Eigenlijk verdienen alle ‘Marbiollers’ van de voorbije 4,5 jaar hier een plaatsje als
dank voor de goede sfeer al die tijd!
Graag wil ik nog een aantal mensen buiten ‘den unief’ bedanken om mij af en toe
eens de doctoraatsdruk te laten vergeten. Bedankt Liesbeth Desmyter, Nathalie Hardy,
Tim en Sofie, Ann en Daf, Peter en An, Frederik, kleine Lukja....
Tot slot een speciaal woordje van dank voor mijn ouders voor alle steun de voorbije
jaren.... en natuurlijk ook voor Peter.
De financiële steun voor dit doctoraatsonderzoek werd verschaft door ‘Instituut voor
de aanmoediging van Innovatie door Wetenschap en Technologie in Vlaanderen’
(IWT).

Summary
iii
Summary Decaying phytodetritus deposited on the sediment surface in salt marshes becomes
quickly colonised by nematodes. Both taxonomic and functional diversity within these
communities are generally low; they are often dominated by the bacterivore, deposit
feeding Monhysterida and Rhabditida. Bacterial grazers are assumed to stimulate the
decomposition rate of organic matter through interactions with the bacterial
community. It is however not clear whether the co-occurrence of these functionally
similar species is of importance in the functioning of the ecosystem. One of the most
accepted hypotheses suggests that species within a functional group are redundant and
thus perform the same role within an ecosystem. The general aims of a first
experiment were to test whether bacterivorous nematodes affect (i) the decomposition
of organic matter and (ii) the activity of the associated microbial community, and to
examine whether these effects (iii) differed between functionally similar species or
(iv) were diversity dependent. (v) Species specific effects on the bacterial community
structure were also analysed.
The influence of four bacterivorous nematode species (Diplolaimelloides meyli,
Diplolaimelloides oschei, Diplolaimella dievengatensis, Panagrolaimus paetzoldi) on
the decomposition of cordgrass (Spartina anglica) and its associated microbial
community was investigated using laboratory microcosm experiments. Cordgrass
leaves were incubated on a sediment surface with a natural mixture of bacteria
originating from sediment, cordgrass detritus and habitat water. Nematodes were
applied separately or in combinations of two or three species. This experimental set-
up allowed testing for species-specific and/or diversity-dependent effects.
In contrast with our expectations, no stimulation of the decomposition process nor of
the microbial community was observed in the presence of the nematodes. In absence
of nematodes, a more extensive growth of fungi, important in the early stages of
cordgrass decay, might have caused higher decomposition rates. Fungal growth
seemed to be hampered by the nematodes, but whether this was a direct or indirect
effect, could not be distinguished. Furthermore, obvious differences were found
between nematode treatments. Panagrolaimus paetzoldi reached much higher
densities than the other species, both in the monospecific and in the combination
treatments. Within the communities containing exclusively monhysterid nematode
species, inhibitory inter-specific interactions were observed, probably due to chemical

Summary
iv
interference. Panagrolaimus paetzoldi caused a decrease in microbial activity. This
was most likely a result of overgrazing. Remarkably, this low microbial activity did
not result in a slow-down of the decomposition process compared to the other
treatments with nematodes. We suggest that P. paetzoldi might be able to directly
assimilate detrital compounds. Also within the treatments containing exclusively
monhysterid nematode species, differences in microbial activity could not be directly
linked to decomposition rates. The fact that fungal and bacterial activity could not be
measured completely separately complicates the establishment of any such link, but
differences in the efficiency of the bacterial communities in each treatment should
also be considered.
Not only impact on bacterial activity, but also on bacterial community composition
was analysed in presence of bacterivorous nematodes. The bacterial community
structure in the monospecific treatments and the control was analysed by means of
Denaturing Gradient Gel Electrophoresis of the 16S rRNA genes. Multi Dimensional
Scaling showed grouping of the bacterial communities per treatment. Analysis of
Similarities indicated that the differences between treatments were significantly larger
than differences within treatments. Our results demonstrate that nematodes can have a
significant structuring top-down influence on the ‘pool’ of bacteria growing on the
detritus, even at the low densities as obtained by the monhysterid species, and that
these impacts are species-specific. Differences in bacterial community composition
between the treatments with monhysterids (D. meyli, D. oschei and D. dievengatensis)
can be explained by species-specific food preferences. Panagrolaimus paetzoldi on
the other hand feeds unselectively, and thus affects the bacterial community
differently. Part of the consumed bacteria are not digested and consequently excreted
again into the environment. Not palatability, but rather cell wall properties would then
determine whether bacteria are digested. Bacterial diversity was assessed from the
DGGE-profiles in the monospecific and control treatments, as well as in the
combination treatments. A top-down effect of the nematodes on the diversity of the
bacterial community was mainly evident under high grazing pressure, i.e. in the
presence of P. paetzoldi, but also with increasing nematode diversity a decrease in
bacterial diversity was observed.
Our results do not support the idea that bacterivorous nematodes are functionally
redundant. The response of P. paetzoldi in the system was profoundly different from
that of the monhysterid nematode species (D. meyli, D. oschei and D. dievengatenis),

Summary
v
yielding considerably higher densities, which resulted in clear impacts on the bacterial
community. As for the monhysterid species, differences in microbial activity and
decomposition rate were found in the presence of different species or species
combinations, without any clear species-specific or diversity-dependent link. This
suggests an idiosyncratic relationship between bacterivorous nematode diversity and
ecosystem functioning.
In a second experiment, the development of the nematode community on decaying
cordgrass detritus in litterbags was studied in the laboratory and in the field. The aims
were to test whether (i) detritus availability could impact the distribution of the
nematode community in an undisturbed laboratory environment, (ii) this distribution
could be affected by natural disturbances, mainly related to tidal currents, in the field
and (iii) the contrasting environmental conditions in the field and in the laboratory
would alter the nematode community structure. Nematodes showed a clear preference
for the detritus under controlled conditions in the laboratory, and the nematode
assemblage on the cordgrass differed considerably from that in the surrounding
sediments. By contrast, in the field, nematodes were rather distributed at random,
probably through passive transport with the tides, showing that active habitat
colonization was of minor importance in the hydrodynamically impacted system.
We assumed that the contrasting incubation conditions in the laboratory and in the
field would affect the structural and functional aspects of the nematode community. In
the laboratory, incubations were performed in the dark, preventing growth of
microalgae, which is an important food source for epistrate feeders and deposit
feeders. Epistrate feeders were however equally abundant in the field and in the
laboratory, indicating that other food sources, probably bacteria, can be consumed.
Deposit feeders, dominating field samples, were mainly replaced by microvores in the
laboratory; still some species were able to adapt to a principally bacterial diet.
Nematode diversity was not affected by resource availability, nor by the daily tidal
disturbance.

Samenvatting
vii
Samenvatting In schorren wordt rottend fytodetritus dat op het sedimentoppervlak terecht komt snel
gekoloniseerd door nematoden. Deze nematodengemeenschappen zijn over het
algemeen gekenmerkt door zowel een lage taxonomische als functionele diversiteit.
Ze worden vaak gedomineerd door de bacterivore, ‘deposit-feeding’ Monhysterida en
Rhabditida. Bacteriële grazers worden verondersteld het afbraakproces van detritus te
stimuleren via interacties met de bacteriegemeenschap. Het is echter niet duidelijk of
het samen voorkomen van deze functioneel erg gelijkende soorten van belang is voor
het functioneren van het systeem. Een vaak gestelde hypothese is dat soorten binnen
een functionele groep dezelfde rol zouden vervullen in een ecosysteem. De algemene
doelstellingen van een eerste experiment waren na te gaan of bacterivore nematoden
(i) de afbraak van organisch materiaal en (ii) de activiteit van de geassocieerde
microbiële gemeenschap beïnvloeden, en te onderzoeken of deze impact (iii)
verschilde tussen functioneel gelijkende soorten of (iv) diversiteitsafhankelijk was.
(v) Ook soortspecifieke effecten op de bacteriële gemeenschapsstructuur werden
geanalyseerd.
Met behulp van microcosmosexperimenten werd de invloed onderzocht van vier
bacterivore nematodensoorten (Diplolaimelloides meyli, Diplolaimelloides oschei,
Diplolaimella dievengatensis en Panagrolaimus paetzoldi) op de afbraak van slijkgras
(Spartina anglica) en op de geassocieerde bacteriële gemeenschap. Slijkgrasbladeren
werden geïncubeerd op een sedimentoppervlak en geënt met een natuurlijk bacterieel
mengsel. Nematoden werden ofwel afzonderlijk toegevoegd, ofwel in combinaties
van twee of drie soorten. Deze experimentele opzet liet toe om soortspecifieke en/of
diversiteitsafhankelijke effecten te achterhalen.
In tegenstelling tot de verwachtingen werd er geen stimulatie van het afbraakproces,
noch van de microbiële gemeenschap waargenomen in de aanwezigheid van
nematoden. In afwezigheid van nematoden zou een sterkere groei van fungi
verantwoordelijk kunnen zijn voor de hogere afbraaksnelheden. Fungi zijn belangrijk
in de eerste stadia van het decompositieproces van slijkgras. De groei van fungi leek
gehinderd in aanwezigheid van nematoden, maar of dit een gevolg was van directe of
indirecte interacties kon niet onderscheiden worden. Ook tussen de experimentele
reeksen met verschillende nematodensoorten en -combinaties werden belangrijke
verschillen gevonden. Panagrolaimus paetzoldi haalde veel hogere densiteiten dan de

Samenvatting
viii
andere soorten, zowel in de monospecifieke als in de combinatiereeksen. Binnen de
gemeenschappen met enkel monhysteride nematodensoorten werden negatieve
interspecifieke interacties waargenomen, die vermoedelijk veroorzaakt werden door
chemische interferentie. In aanwezigheid van P. paetzoldi werd de microbiële
activiteit onderdrukt, waarschijnlijk als gevolg van overbegrazing. Vreemd genoeg
resulteerde deze lagere microbiële activiteit niet in een vertraging van het
decompositieproces in vergelijking met andere nematodenreeksen. We
veronderstellen dat P. paetzoldi in staat is om rechtstreeks componenten van het
organische materiaal te assimileren. Ook binnen de reeksen waarin enkel
monhysteriden groeiden kon geen verband gelegd worden tussen microbiële activiteit
en afbraaksnelheid. Dit zou kunnen liggen aan het feit dat de activiteit van fungi en
bacteriën niet volledig afzonderlijk kon bepaald worden, maar ook verschillende
groeiefficiënties van de bacteriële gemeenschap vormen een mogelijke verklaring.
Naast de impact van bacterivore nematoden op de bacteriële activiteit, werd ook de
invloed op de bacteriële gemeenschapssamenstelling onderzocht. De bacteriële
gemeenschapsstructuur in de monospecifieke reeksen en de controle werd
geanalyseerd aan de hand van ‘Denaturing Gradient Gel Electrophoresis’ van de 16S
rRNA genen. ‘Multi Dimensional Scaling’ toonde een groepering van de bacteriële
gemeenschappen per reeks. ‘Analysis of Similarities’ bevestigde dat de verschillen
tussen de reeksen significant groter waren dan de verschillen binnen de reeksen. Onze
resultaten tonen aan dat nematoden zelfs bij lage densiteiten, zoals deze van de
monhysteriden, een aanzienlijke structurerende ‘top-down’ impact kunnen hebben op
de ‘pool’ van bacteriën die groeien op detritus, en dat deze invloed soortspecifiek is.
Verschillen in bacteriële gemeenschapsstructuur in de reeksen met de monhysteride
nematodensoorten (D. meyli, D. oschei en D. dievengatensis) kunnen verklaard
worden door soortspecifieke voedselpreferenties. Panagrolaimus paetzoldi
daarentegen voedt zich niet selectief en scheidt een deel van de opgenomen bacteriën
onverteerd terug uit; deze soort heeft daardoor een erg verschillende impact op de
bacteriële gemeenschap. In dit geval zou niet zozeer ‘smaak’, maar eerder
eigenschappen van de celwand bepalen of bacteriën al dan niet worden verteerd. Aan
de hand van de DGGE-profielen werd ook de bacteriële diversiteit bepaald in de
monospecifieke reeksen, in de controle, en in de combinatiereeksen. Een top-down
effect van de nematoden op de diversiteit van de bacteriële gemeenschappen werd

Samenvatting
ix
duidelijk onder hoge graasdruk, zoals in aanwezigheid van P. paetzoldi, maar ook met
toenemende nematodendiversiteit nam de bacteriële diversiteit af.
De hypothese dat veel bacterivore nematodensoorten redundant zouden zijn wordt
niet ondersteund door onze resultaten. De respons van P. paetzoldi verschilde
fundamenteel van die van de monhysteride nematodensoorten (D. meyli, D. oschei en
D. dievengatensis). Panagrolaimus paetzoldi bereikte veel hogere densiteiten, wat
resulteerde in een duidelijke impact op de bacteriële gemeenschap. Binnen de reeksen
met monhysteride soorten werden verschillen gevonden in microbiële activiteit en
afbraaksnelheden in aanwezigheid van verschillende soorten of soortencombinaties,
maar zonder duidelijk soort- of diversiteitsafhankelijk verband. Dit suggereert een
idiosyncratisch verband tussen diversiteit van bacterivore nematoden en het
functioneren van het systeem.
In een tweede experiment werd de ontwikkeling van nematodengemeenschappen op
rottend slijkgras bestudeerd met behulp van ‘litterbag’- incubaties, enerzijds onder
gecontroleerde omstandigheden in het laboratorium, en anderzijds in het veld, waar
incubaties onderhevig waren aan fluctuaties, vooral geassocieerd met de
getijdenwerking. Het doel van het experiment was na te gaan of (i) de aanwezigheid
van detritus een invloed heeft op de verspreiding van nematoden in een onverstoord
milieu, (ii) deze verspreiding kon beïnvloed worden door de natuurlijke verstoringen
in het veld, en of (iii) de verschillende omgevingsfactoren in het veld en het labo een
impact hebben op de structuur van de nematodengemeenschap.
Onder gecontroleerde omstandigheden in het labo toonden nematoden een duidelijke
voorkeur voor het detritus, en de nematodengemeenschap op het slijkgras verschilde
fundamenteel van die in het omringende sediment. In het veld daarentegen waren de
nematoden eerder ad random verspreid, waarschijnlijk ten gevolge van passief
transport met de getijden, wat aantoont dat actieve kolonisatie hier van minder belang
was.
We verwachtten dat de contrasterende incubatie-omstandigheden (laboratorium-veld)
de structurele en functionele aspecten van de nematodengemeenschap zouden
beïnvloeden. In het labo werden de incubaties uitgevoerd in het donker, hetgeen de
groei van microalgen, een belangrijke voedselbron voor ‘epistrate feeders’ en
‘deposit-feeders’, verhinderde. ‘Epistrate feeders’ waren echter in gelijke mate
aanwezig in het veld en in het laboratorium, wat aantoont dat ze ook andere
voedselbronnen kunnen benutten, vermoedelijk bacteriën. ‘Deposit-feeders’ waren

Samenvatting
x
dominant aanwezig in de veldstalen, en werden in het laboratorium voornamelijk
vervangen door microvoren; toch konden enkele soorten zich aanpassen aan een
hoofdzakelijk bacterieel dieet. De nematodendiversiteit werd niet beïnvloed door
voedselbeschikbaarheid, noch door de (voornamelijk) tidale verstoringen.

Chapter 1
General Introduction and Aims

General Introduction and Aims
3
Introduction Salt marshes are among the most productive ecosystems of the world (Odum, 1971).
They occur along many coasts and estuaries in temperate regions. The annual primary
production can be up to 4 kg C per m², most of it produced by grasses (Spartina spp.)
which generally dominate these ecosystems (Howarth, 1993). The net export of
organic matter to coastal waters is usually limited, especially in the Westerscheldt
Estuary (Hemminga et al. 1992, 1993), and only a very small fraction of the
production is permanently accreted in marsh sediments. Most of the organic matter
produced in marshes is decomposed in situ, fueling intense rates of microbial
processes (Howarth, 1993). Fungi and bacteria are the principal competitors for
organic substrates. Fungi are mostly aerobic decomposers and dominate the
aboveground decomposition processes of plant detritus (Newell et al., 1989). Once the
detritus is deposited on the sediment surface, or becomes mixed with the sediment,
bacteria take over their function (Howarth, 1993). The benthic bacterial community
can be affected by the activity of their grazers, mainly protozoa and nematodes.
Free-living nematodes are generally the dominant metazoan meiobenthic taxon in
marine and estuarine sediments. Their densities vary between 105 and 107 individuals
per m² (Heip et al., 1985). Despite their numerical dominance, the function and
trophic position of nematodes in benthic food webs has long been poorly understood.
Meanwhile it has been shown that they play an important role in energy fluxes in the
benthos. On the one hand they are food sources for macrofauna and epi- and
hyperbenthic organisms (Gee, 1989; Coull, 1990; Hamerlynck and Vanreusel, 1993).
On the other hand, a wide spectrum of particulate organic food sources, e.g. bacteria,
diatoms, ciliates, flagellates or other meiofaunal organisms, can be consumed by
nematodes. Based on morphological aspects of their buccal cavity, an assessment was
made of the feeding preferences of free-living marine nematodes, leading to the
subdivision in trophic groups (Wieser, 1953). This classification has been refined by
incorporating observations on the gut contents (e.g. Perkins, 1958; Hopper and
Meyers, 1967; vanThun, 1968; Deutsch, 1978) and on the feeding habits of a range of
nematode species (Romeyn and Bouwman, 1983; Jensen, 1987; Moens and Vincx,
1997).
Not only the densities of nematodes are high in marine sediments, their communities
are also characterised by high diversities (Heip et al., 1985). The question can be

General Introduction and Aims
4
raised why so many (functionally) similar species occur in marine and estuarine
benthic sites. In general, several, often contrasting, hypotheses about the biodiversity-
ecosystem functioning relationship have been formulated (Schläpfer et al., 1999)1.
The extremes range from the (modified) null hypothesis, predicting no effect of
changing species diversity on the functioning of ecosystems, to the rivet hypothesis
(Ehrlich and Ehrlich, 1981), describing an enhancement of processes each time a
species is added to the system. Over the last few years, the redundancy hypothesis has
received a lot of attention. According to this model, the effect of altering species
diversity will decrease at higher diversity levels (Walker, 1992). The underlying idea
is that with increasing diversity, the function of a disappearing species is more likely
to be taken over by other species of the same functional level. In contrast with the
other theories, the idiosyncratic model predicts that altering species diversity will
affect ecosystem functioning, but in an unpredictable manner (Lawton, 1994).
Empirical studies on the biodiversity-function link generally focus on vegetations in
terrestrial systems in which production and stability of the ecosystems are studied
(a.o. Ewel et al., 1991; Givnish, 1994; Silvertown et al., 1994; Tilman and Downing,
1994, Collins, 1995; Tilman et al., 1996). The lack of straightforward results in these
studies demonstrates that a generally applicable hypothesis probably does not exist
and emphasises the necessity to incorporate other ecosystems and other functions into
this discussion.
In many ecosystems, primary production enters the decomposition cycle as plant litter
(Wardle et al., 1997). As mentioned before, this also goes for detritus in salt marshes
along the Westerscheldt (Hemminga 1992, 1993). When deposited on the sediment
surface, decaying phytodetritus becomes colonised by nematodes. These communities
are generally characterised by a lower taxonomic and functional diversity compared to
those of the surrounding sediments, with typically the bacterivore deposit-feeding
Monhysterida and Rhabditida occurring in these spots (Bouwman, 1984; Warwick,
1987; Alkemade et al., 1993). The main reason for this microhabitat preference may
be related to food preferences (Trotter and Webster, 1984; Moens et al., 1999).
However, an evaluation of the relative importance of abiotic disturbances, mainly
1 see also introduction Chapter 4

General Introduction and Aims
5
related to tidal action, in structuring intertidal nematode communities in salt marshes
has not yet been made2.
Bacterivorous nematodes are assumed to affect decomposition of organic matter
through interactions with heterotrophic bacteria3. A number of mechanisms have been
proposed by which they could stimulate a bacterial community. Grazing for instance
would enhance bacterial turn-over (e.g. Tenore et al., 1977; Abrams and Mitchell,
1980; Tietjen, 1980; Findlay and Tenore, 1982; Anderson et al., 1983; Rieper-
Kirchner, 1989; Alkemade et al., 1992a, b; Beare et al., 1992). Excretion of N-rich
compounds has been found to be important in stimulating bacterial activity in N-
limited systems (Anderson et al., 1983). Bacterial growth might be facilitated by the
Figure 1: Overview of the fate of primary production in salt marshes along the Westerscheldt.
Dotted lines represent the interactions studied in this thesis.
secretion of mucus trails (Riemann and Schrage, 1978), and through their movements,
nematodes are thought to enhance diffusion of nutrients and oxygen in the sediment
(Cullen, 1973; Alkemade et al., 1992a; Aller and Aller, 1992). Whether nematodes
can also affect bacterial community structure, as observed under protist grazing 2 see also introduction Chapter 5 3 see also introduction Chapter 2

General Introduction and Aims
6
pressure (Van Hannen et al., 1999), needs to be investigated4. Experimental data
indicate that nematodes are able to distinguish between, and preferentially consume
different bacterial strains when offered in pure cultures (Moens et al., 1999), but
whether they maintain the ability of selecting bacteria from mixed communities is
unknown.
Bacterivorous nematode communities associated with decaying phytodetritus in salt
marshes are very well suited to test for diversity-related impacts on ecosystem
functioning. Many of the species typically associated with phytodetritus can easily be
cultivated and manipulated under laboratory conditions (Moens and Vincx, 1998).
Additionally, their small body sizes (<1mm) and limited dispersal capacities allows
studying their impact on ecosystem processes at a small spatial scale, in the order of
centimetres (Coleman, 1983, Moore et al., 1996; Verhoef, 1996; Mikola and Setälä,
1998). In specific, it is easily feasible to compose artificial nematode communities
with different species compositions and diversity levels under controlled conditions,
in order to evaluate the importance of diversity related or species-specific impacts on
bacterial communities and decomposition rates.
Topics and outline of this thesis
The first topic of this thesis concentrates on the influence of free-living nematodes
on the decomposition of salt marsh phyto-detritus and its associated bacterial
community. This was studied using microcosm experiments in which cordgrass
detritus was incubated on a sediment surface. The microcosms were inoculated with a
natural mixture of bacteria, originating from the sediment, fresh and decaying
cordgrass, habitat water and nematode cultures. Three monhysterid nematode species
(Diplolaimelloides meyli, Diplolaimelloides oschei, Diplolaimella dievengatensis) and
one rhabditid (Panagrolaimus paetzoldi) commonly found on decaying cordgrass
detritus in the Paulina salt marsh (Westerscheldt Estuary) (Moens, pers. comm.), were
added to the microcosms, either separately, or in combination with one or two of the
other species. The general aims of this experiment were to test whether nematodes
affect (i) the decomposition process and (ii) the activity of the microbial community,
4 see also introduction Chapter 3

General Introduction and Aims
7
and to examine whether these influences are (iii) species-specific or (v) rather
diversity-dependent. (iv) Also the bacterial community structure in the presence of the
four nematodes species was studied. These results are discussed in chapter 2, chapter
3 and chapter 4 of this thesis.
In Chapter 2 species-specific influences on the decomposition process and on the
activity of the associated microbial community are described, using the results of the
monospecific treatments and the controls. It was expected that the nematodes would
enhance the decomposition process through stimulation of the bacterial community
and, taking the redundancy hypothesis as basic assumption, that this effect would not
be species-specific. This chapter has been published as De Mesel, I, Derycke, S,
Swings, J, Vincx, M, and Moens, T (2003) Influence of bacterivorous nematodes on
the decomposition of cordgrass. J. Exp. Mar. Biol. Ecol. 296: 227-242.
In Chapter 3 species-specific effects of the different nematode species on the
composition and diversity of the bacterial community developing on the detritus was
examined using Denaturing Gradient Gel Electrophoresis. Such impacts have already
been demonstrated under protist grazing activities (Van Hannen et al., 1999; Rønn et
al., 2002). This chapter has been published as De Mesel, I, Derycke, S, Moens, T, Van
der Gucht, K, Vincx, M, and Swings, J (2004) Top-down impact of bacterivorous
nematodes on the bacterial community structure: a microcosm study. Env. Microbiol.
6(7): 733-744.
In Chapter 4 we examined whether the species-specific effects on the decomposition
process and the associated microbial community observed in the monospecific
treatments (Chapter 2) persisted in the combination treatments, and/or whether the
impact was diversity related. To do so, information on microbial activity, bacterial
diversity, and on the decomposition rate in the monospecific treatments and the
combination treatments with two and three species were integrated.
A second topic of this thesis concerns the structuring force of active dispersion
and food availability on a nematode community and the impact of abiotic
disturbances on this distribution in a salt marsh habitat. Chapter 5 describes an
experiment in which cordgrass and controls (plastic strips) in litterbags were
incubated on salt marsh sediment in the laboratory under controlled conditions and in
the field where they were flooded twice daily. Microhabitat preference for phyto-
detritus in a salt marsh habitat has been described by several authors (Bouwman et al.,
1984; Warwick, 1987). Others however have described a strong passive dispersion of

General Introduction and Aims
8
meiofauna in general and nematodes in specific in intertidal habitats through tidal
currents (e.g. Sherman and Coull, 1980). We tested whether (i) detritus availability
could impact the distribution of the nematode community in an undisturbed laboratory
environment, (ii) this distribution could be affected by natural disturbances in the
field, and (iii) the contrasting environmental conditions in the field and in the
laboratory would alter the nematode community structure.
This chapter has been submitted for publication as De Mesel, I, Vanaverbeke, J,
Vincx, M, and Moens, T. (submitted) Development of a nematode community on
decaying organic matter under contrasting conditions.

Published as:
De Mesel, I, Derycke, S, Swings, J, Vincx, M, Moens, T (2003) Influence of
bacterivorous nematodes on the decomposition of cordgrass. Journal of Experimental
Marine Biology and Ecology 296: 227-242
Chapter 2
Influence of bacterivorous nematodes on the decomposition of cordgrass

Chapter 2: Species-specific effects on decomposition 11
Abstract
The influence of bacterivorous nematodes (Diplolaimelloides meyli,
Diplolaimelloides oschei, Diplolaimella dievengatensis, Panagrolaimus paetzoldi) on
the decomposition of a macrophyte (Spartina anglica) in an aquatic environment was
investigated by using laboratory microcosm experiments. Several earlier studies have
shown enhancement of the decomposition process in the presence of nematodes, but
nematode species-specific effects were never tested. In this study four bacterivorous
nematode species were applied separately to microcosms to investigate such species-
specific influences.
No stimulation of the decomposition process nor of the microbial community was
observed in the presence of the nematodes, both were highest in the absence of
nematodes. However, clear differences were found between nematode treatments.
Panagrolaimus paetzoldi reached much higher numbers than the other species,
causing a decrease in microbial activity, probably due to (over)grazing. Remarkably
this low microbial activity did not result in a slow-down of the decomposition process
compared to the other nematode treatments, raising the question whether P. paetzoldi
might be able to directly assimilate detrital compounds. Other nematode species
reached much lower densities, but nevertheless an influence on the decomposition
process was observed. However, this experiment does not support the view that
bacterivorous nematodes enhance decomposition rates.
The experimental results show that in nematode communities the use of functional
groups is inadequate for biodiversity studies. The four nematode species used in this
study belong to the same functional group, but are clearly not functionally redundant
since they all have a different influence on the cordgrass decomposition. This suggests
that the relationship between nematode species diversity and ecosystem functioning
may be idiosyncratic.

Chapter 2: Species-specific effects on decomposition 13
Introduction
Nematodes occur in all types of soils and sediments and are often very abundant,
especially in marine and brackish sediments (Heip et al., 1985). They are classified in
trophic or functional groups based on the morphology of their buccal cavity (Wieser,
1953). Nematodes within a functional group are thought to perform the same or very
similar function(s) in ecosystems. This view has recently been questioned by Mikola
and Setälä (1998) based on experimental studies in a soil food web in which species-
specific effects, rather than functional group effects, were observed.
Bacterivorous nematodes are deposit feeders sensu Jensen (1987) which mainly
appear in spots with high microbial activity. They are thought to enhance
decomposition processes through stimulation of the microbial community (Riemann
and Schrage, 1978; Abrams and Mitchell, 1980; Anderson et al., 1981; Findlay and
Tenore, 1982). Three main ways of stimulation have been proposed: (i) bioturbation
resulting in a higher diffusion of oxygen and nutrients (Abrams and Mitchell, 1980;
Aller and Aller, 1992; Alkemade et al., 1992b), (ii) secretion of nutrient rich
compounds such as mucus (Riemann and Schrage, 1978; Riemann and Helmke, 2002)
and (iii) grazing, which implies keeping the bacterial community active and
remineralising nutrients (a.o. Abrams and Mitchell, 1980; Ingham et al., 1985).
The role of nematodes in decomposition processes has mainly been studied in
terrestrial ecosystems (e.g. Abrams and Mitchell, 1980; Anderson et al., 1981; Ingham
et al., 1985). In marine environments similar studies have been scarce.
The lower parts of salt marshes are often dominated by a cordgrass vegetation. Most
of the cordgrass litter is decomposed in situ on the sediment surface, still attached to
the plant (Groenendijk, 1984; Newell et al., 1985; Buth and Voesenek, 1988). As a
consequence laboratory microcosm experiments studying this decomposition process
tend to be artificial. However, manipulative lab experiments are a necessary tool to
study (aspects of) specific processes, such as litter decomposition. In some studies
litter was oven dried, crushed and mixed with sediments (Findlay and Tenore, 1982;
Alkemade et al., 1992b). Alkemade et al. (1992a) incubated cordgrass fragments on
agar which tends to stimulate an explosive growth of bacteria and bacterivorous
nematodes (pers. observation). In a study of Lillebø et al. (1999) leaves were
submerged during the whole experiment. All have shown a stimulation of the

Chapter 2: Species-specific effects on decomposition
14
decomposition of the cordgrass leaves by bacterivorous nematodes, and these results
are at the basis of the general idea that - much as in many terrestrial systems -
nematodes enhance the decomposition of phytodetritus in aquatic systems.
Our study differed from previous studies in that leaves were incubated on water
saturated sediment obtained from the habitat. The aim of this study was to investigate
whether bacterivorous nematodes commonly found on cordgrass detritus indeed
stimulate the decomposition process. Furthermore species-specific influences were
investigated.
The effect of four bacterivorous nematode species was studied (Diplolaimelloides
meyli, Diplolaimelloides oschei, Diplolaimella dievengatensis and Panagrolaimus
paetzoldi). Based on the above mentioned evidence for a stimulatory effect we
assumed that (1) all four nematode species used would stimulate this decomposition
process and that (2) this effect would not be species-specific since they all belong to
the same functional group.
Materials and Methods Experimental set-up
Decomposition of cordgrass litter was studied in laboratory microcosms. These
microcosms consisted of cordgrass fragments on sediment, inoculated with a
microbial suspension and – except in the control treatment – nematodes in sterile petri
dishes. The microcosms were incubated in the dark at 20 ± 2°C.
Sediment, cordgrass, microbiota and nematodes were all collected in the Paulina
saltmarsh (ca. 60 ha) (Westerscheldt Estuary, The Netherlands) in September, 2000.
The sediment was washed with tap water over a 1 mm sieve and the fraction that was
retrieved on a 38 µm sieve was dried overnight at 180°C. 15 ± 0.5 g sediment was
added to sterile petri dishes (∅ 90 mm) and rehydrated with 6.5 ± 1 ml sterile
artificial sea water (ASW, Dietrich and Kalle, 1957) with a salinity of 20. Extra
nutrients were added as a 1/1000 (vol/vol) Killian solution (von Thun, 1966) in
artificial sea water.
Green cordgrass leaves (Spartina anglica) were cut from standing green plants. They
were washed successively with ethanol (70%) , sterile destilled water, ethanol, sterile

Chapter 2: Species-specific effects on decomposition 15
destilled water, ethanol and (twice) ASW (Alkemade et al., 1992a). Leaves were air-
dried and 1.9 ± 0.2 g leaves were incubated on the surface of the sediment.
A natural microbial inoculum was added to the petri dishes. The inoculum was
obtained by mixing habitat water, sediment, decomposing and fresh cordgrass leaves
and agar from the nematode cultures (see below). This mixture was filtered over
Whatman GF/C filters to remove flagellates and other small eukaryotes. Every petri
dish was inoculated with 2 ml of this suspension. The microcosms were incubated in
the dark for 24h at 20 ± 2°C before nematodes were added.
Four nematode species, which are common on decomposing cordgrass in the Paulina
salt marsh (Moens, unpublished), were used in our experiment: the monhysterids
Diplolaimelloides meyli, Diplolaimella dievengatensis and Diplolaimelloides oschei
and the rhabditid Panagrolaimus paetzoldi. Agnotobiotic cultures of all species
growing together with unidentified bacteria are available in the Biology Department.
Each microcosm, except for the controls, received 36 individuals of a single nematode
species (Table 1). Nematodes were handpicked with the tip of a fine needle.
Treatment code Nematode species
A Diplolaimelloides meyli
B Diplolaimelloides oschei
C Panagrolaimus paetzoldi
D Diplolaimella dievengatensis
S (no nematodes)
Table 1: Overview of the treatments
Sampling
Samples were taken after 10, 20, 30, 40, 50, 65 and 75 days. On each sampling
occasion three microcosms from every treatment were destructively sampled.
Bacteria were removed from the cordgrass leaves through rinsing with sterile ASW.
After sub-sampling the rinse water to measure respiration rate and enzymatic activity
of the microbial community (see below), nematodes which still adhered to the leaves
were removed with ASW and MgCl2 (8%). Nematodes were preserved in a
formaldehyde solution (final concentration: 4%).

Chapter 2: Species-specific effects on decomposition
16
The same procedure as at the start of the experiment was used to clean the cordgrass
leaves: washing with ethanol and sterile destilled water (2X), again ethanol and finally
twice sterile ASW. They were then preserved at –20°C for C- and N- analyses.
The sediment was preserved on 4% formaldehyde solution for quantification of the
nematodes.
Weight loss
At every sampling event the cordgrass fragments were dried to the air and weighed.
These values were converted to dry weight according to the equation (De Mesel,
unpublished):
dry weight = 0.2084*EXP(0.6945*weightair dried)
Weight is expressed as the percentage of the initial dry weight of the fragment still
present in the microcosm at the moment of sampling.
weight = (dry weightsampling/dry weightstart) *100
C/N-analysis
The C- and N-concentrations, expressed as percentages, of the organic matter were
determined with a Carlo-Erba element analyser type NA-1500. Plant fragments were
crushed manually using liquid nitrogen. 3 to 5 mg of the litter was used for analysis.
Carbon and nitrogen content are expressed as the percentage of the initial value still
present at the time of sampling.
C-content = startstart
samplingsampling
dry weight*/100)(%Cdry weight*)100/(%C * 100
N-content = startstart
samplingsampling
dry weight*/100)(%Ndry weight*)100/(%N * 100

Chapter 2: Species-specific effects on decomposition 17
Nematodes
Nematodes were extracted from the sediment by centrifugation with Ludox (Heip et
al., 1985). Nematodes were stained with Rose Bengal and counted under a dissecting
microscope.
Microbial activity
Total aerobic activity was measured using the Strathkelvin 928 6-channel Dissolved
Oxygen System. 1 ml of the suspension that was rinsed off the cordgrass leaves (see
above) was transferred to each measurement unit. The oxygen concentration was
measured at 20°C during approximately 30 minutes. Respiration rates (µmol O2 h-1)
were calculated from the slope of the regression of oxygen concentration versus time.
The EnzChek® Protease Assay Kit E6638 for green fluorescence (Molecular Probes)
was used to measure the proteolytic activity of the microbial community.
Fluorescently labelled casein served as substrate. It is hydrolysed by metallo-, serine,
acid and sulphydryl proteases secreted by the micro-organisms, thus releasing the
otherwise quenched fluorescent labels into the medium. 100 µl of the suspension
obtained by rinsing the cordgrass leaves was mixed with an equal volume of the
casein containing solution. After an incubation of 30 minutes in the dark the
fluorescence was read on a Victor multilabel counter using excitation and emission
wavelengths of 505 and 513 nm respectively. Readings were performed at 25°C.
Statistical analysis
Univariate 2-way analysis of variance (ANOVA) was used to test for differences of
remaining weight, C-content, N-content, respiration rate and enzymatic activity
between treatments, times and treatment x time. Respiration rate was log-transformed
and enzymatic activity square-root transformed to meet the required assumptions; for
the other effects no transformation was needed. When ANOVA indicated significant
differences the Tukey HSD test was used for pairwise post hoc comparisons.
All tests were done with the STATISTICA software.

Chapter 2: Species-specific effects on decomposition
18
Results Decomposition
Weight loss
A two-way ANOVA showed significant differences for the effects ‘time’, ‘treatment’
but not for the combined effect ‘time’ x ‘treatment’ (Table 2). For the effect
‘treatment’ the Tukey HSD post-hoc test indicated a significantly lower weight loss in
the presence of D. dievengatensis (treatment D) than in the control treatment S
(p<0.01) or in the presence of D. meyli (treatment A) (p<0.001). The graph (fig. 1)
shows that most of the differences between treatments occurred during the first 50
days of the experiment although no significant differences were found for the
combined effect. By the end of the experiment (day 75) weight losses were similar for
all treatments.
Figure 1: Remaining weight of the cordgrass leaves expressed as percentage of the initial
values (average of 3 replicates ± 1 standard error)
0 10 20 30 40 50 65 75
Days
25
50
75
100
% w
eigh
t rem
aini
ng
D. meyliD. oscheiP. paetzoldiD. dievengatensiscontrol
0 10 20 30 40 50 65 75
Days
25
50
75
100
% w
eigh
t rem
aini
ng
25
50
75
100
% w
eigh
t rem
aini
ng
D. meyliD. oscheiP. paetzoldiD. dievengatensiscontrol
D. meyliD. oscheiP. paetzoldiD. dievengatensiscontrol

Chapter 2: Species-specific effects on decomposition 19
Table 2: Results of the two-way ANOVA for the effect ‘weight’ (ns: not
significant, **:p<0.01, ***:p<0.001)
Carbon loss
Carbon content in the cordgrass detritus showed a similar pattern as for weight (fig. 2)
in every treatment. The two-way ANOVA showed significant differences for the
effects ‘time’ and ‘treatment’ and for the combined effect ‘time’ x ‘treatment’ (table
3).
Figure 2: C-content of the cordgrass leaves expressed as percentage of the initial values
(average of 3 replicates ± 1 standard error)
Effect DF Effect DF Error F p-level
Time 6 70 112.8143 ***
Treatment 4 70 4.5950 **
Time x Treatment 24 70 1.4714 ns
0 10 20 30 40 50 65 75
Days
25
50
75
100
% C
rem
aini
ng
D. meyliD. oscheiP. paetzoldiD. dievengatensisControl
0 10 20 30 40 50 65 75
Days
25
50
75
100
% C
rem
aini
ng
0 10 20 30 40 50 65 75
Days
25
50
75
100
% C
rem
aini
ng
25
50
75
100
% C
rem
aini
ng
D. meyliD. oscheiP. paetzoldiD. dievengatensisControl
D. meyliD. oscheiP. paetzoldiD. dievengatensisControl

Chapter 2: Species-specific effects on decomposition
20
As for the variable weight, C- content of the cordgrass differed significantly between
treatment D (D. dievengatensis) and on the one hand the control treatment S (p<0.01)
and on the other hand treatment A (D. meyli) (p<0.01). In addition the latter two
differed significantly from treatments B (D. oschei) (both p<0.05) and C (P.
paetzoldi) (p<0.01 and p<0.05 respectively). The post hoc test for the combined effect
showed an unique significant difference between treatment C (P. paetzoldi) and the
control treatment S (p<0.05) after 40 days.
Table 3: Results of the two-way ANOVA for the effect ‘Carbon content’
(*:p<0.05, ***:p<0.001)
Nitrogen loss
The N content of the cordgrass fragments, expressed as a percentage of the original
value, is shown in figure 3. The results of the two-way ANOVA are given in table 4.
As shown by the Tukey HSD post hoc test the remaining nitrogen content of the
detritus was significantly higher in treatment D (D. dievengatensis) than in all other
treatments (p<0.001) and significantly lower in control treatment S than in treatments
Table 4: Results of the two-way ANOVA for the effect ‘N-content‘
(***:p<0.001)
A (D. meyli) and C (P. paetzoldi) (both p<0.001). From day 10 until day 40 the
nitrogen content of the cordgrass leaves differed significantly between treatments D
(D. dievengatensis) and control treatment S (day 10, day 20 and day 30: p<0.001; day
40: p<0.01); significant differences between treatment D and treatment B (D. oschei)
Effect DF Effect DF Error F p-level
Time 6 70 126.8398 ***
Treatment 4 70 7.6499 ***
Time x Treatment 24 70 1.9895 *
Effect DF Effect DF Error F p-level
Time 6 70 62.10779 ***
Treatment 4 70 33.94058 ***
Time x Treatment 24 70 3.17894 ***

Chapter 2: Species-specific effects on decomposition 21
were found from day 20 until day 40 and again on day 65 (day 20: p<0.01, day 30:
p<0.001, day 40: p<0.01 and day 65: p<0.05).
Figure 3: N-content of the cordgrass leaves expressed as percentage of the initial values
(average of 3 replicates ± 1 standard error)
Nematodes
Figure 4: Number of nematodes per microcosm (log scale) (average of 3 replicates ± 1
standard error)
0
25
50
75
100
0 10 20 30 40 50 65 75
% N
rem
aini
ng
D. meyliD. oscheiP. paetzoldiD. dievengatensisControl
0
25
50
75
100
0
25
50
75
100
0 10 20 30 40 50 65 75
% N
rem
aini
ng
D. meyliD. oscheiP. paetzoldiD. dievengatensisControl
Days
10 20 30 40 50 65 75
Days
1
10
100
1000
10000
100000
Num
ber o
f nem
atod
es (l
og)
t
D. meyliD. OscheiP. paetzoldiD. dievengatensis
10 20 30 40 50 65 75
Days
1
10
100
1000
10000
100000
Num
ber o
f nem
atod
es (l
og)
t
D. meyliD. OscheiP. paetzoldiD. dievengatensis

Chapter 2: Species-specific effects on decomposition
22
From day 30 onwards, P. paetzoldi (treatment C) reached densities 10 to 103 times
higher than those of other nematodes (fig. 4). Densities up to 125000 individuals per
microcosm were recorded.
The differences between the abundances of D. meyli (treatment A), D. oschei
(treatment B) and D. dievengatensis (treatment D) were smaller, D. meyli reaching the
lowest nematode numbers on the average. High variation between replicates was
observed.
Microbial activity
Respiration rate
Aerobic activity of the microbial community, represented by the respiration rate, was
high at the start of the experiment in all treatments (fig. 5A); it steadily decreased and
stabilised from 40 days onwards at lower respiration rates. Significant differences
were indicated by two-way ANOVA for the effects ‘time’, ‘treatment’ and ‘time’ x
‘treatment’. Tukey HSD post-hoc test showed significantly lower respiration rates in
the presence of P. paetzoldi (treatment C) than in the control (treatment S) (p<0.001).
Figure 5: Respiration rate of the microbial community (average of 3 replicates ± 1 standard
error)
In all treatments, respiration rates on days 10 and 20 differed significantly with those
from 40 days onwards (all p<0.001); within treatment C (P. paetzoldi) differences
00.10.20.30.40.50.60.70.80.9
1
10 20 30 40 50 65 75
Days
Res
pira
tion
rate
(µm
ol O
2h-1
)
D. oscheiP. paetzoldiD. dievengatensiscontrol
D. meyli
00.10.20.30.40.50.60.70.80.9
1
00.10.20.30.40.50.60.70.80.9
1
10 20 30 40 50 65 75
Days
Res
pira
tion
rate
(µm
ol O
2h-1
)
D. oscheiP. paetzoldiD. dievengatensiscontrol
D. meyliD. oscheiP. paetzoldiD. dievengatensiscontrol
D. meyli

Chapter 2: Species-specific effects on decomposition 23
were already significant between day 10 and day 30 (p<0.001), indicating a faster
decrease in respiration rate in this treatment.
DF Effect DF Error F p-level
Time 6 70 139,8768 ***
Treatment 4 70 8,085291 ***
Time x Treatment 24 70 1,849047 *
Table 5: Results of the two-way ANOVA for the effect ‘respiration rate’
(*:p<0.05, ***:p<0.001)
Enzymatic activity
Enzymatic activity, which is a measure of aerobic as well as anaerobic microbial
activity, increased at the beginning of the experiment and decreased at the end in most
treatments except in treatment C (P. paetzoldi). In the latter, enzymatic activity was
low during the whole experiment. The Tukey HSD test following two-way ANOVA
(table 6) confirmed that enzymatic activity in the presence of P. paetzoldi was
significantly lower than in other treatments (all p<0.001).
Figure 6: Enzymatic activity of the microbial community (average of 3 replicates ± 1 standard
error)
10 20 30 40 50 65 75
Days
0
5000
10000
15000
20000
25000
Enzy
mat
ic a
ctiv
ity (c
ount
s min
-1)
10 20 30 40 50 65 75
Days
0
5000
10000
15000
20000
25000
Enzy
mat
ic a
ctiv
ity (c
ount
s min
-1)
D. oschei
P. paetzoldi D. dievengatensis control
D. meyli

Chapter 2: Species-specific effects on decomposition
24
DF Effect DF Error F p-level
Time 5 59 7,639149 ***
Treatment 4 59 19,04666 ***
Time x Treatment 20 59 3,603706 ***
Table 6: Results of the two-way ANOVA for the effect ‘enzymatic activity’
(***:p<0.001)
Discussion Decomposition process
The major differences in the decomposition process between treatments were
observed during the first 40 days. At the end of the experiment, after 75 days, the
decomposition process in all treatments was in a similar phase as shown by the weight
loss and the elemental analyses. At that time presumably all easily degradable
compounds were decomposed and mainly high molecular weight compounds, such as
cellulose and lignin, were left over (Valiela, 1995). The differences in the
decomposition process between treatments the first 40 days indicate that the final,
comparable situation is reached at different speeds, depending on the presence or
absence and the identity of the nematode species. This demonstrates the importance of
the duration and the sampling interval in these studies: short time experiments or
unique samplings at the end of a long experiment can cause misleading conclusions.
The high nitrogen loss from the cordgrass leaves in all treatments during the first 10
days is a consequence of the leaching of nutrient rich compounds, mainly proteins
(Valiela, 1995), and was probably enhanced by the cleaning of the leaves prior to the
experiment.
The chemical dynamics in the cordgrass leaves are considered to be best suited to
express the decomposition process. In the absence of macrofauna no detritus
fragments are grazed away and the decomposition of the detritus is a result of the
selective removal of organic compounds by the microbiota. Detailed information
about the selective use of organic compounds would be lost when looking only at
weight loss, especially when they make up only small amounts of the total weight,
like nitrogen (2.4% of total weight at the start of the experiment, data not shown).

Chapter 2: Species-specific effects on decomposition 25
Microbial activity
Bacteria and fungi are the principal actors in decomposition processes (Valiela, 1995).
Although microbial activity was not measured in situ, respiration rate and enzymatic
activity give a good indication of the microbial activity in the microcosms and thus
enable us to compare between treatments.
The high initial aerobic activity in each treatment can partly be explained by the
leaching of easily degradable compounds from the detritus which are immediately
broken down by the microbial community. The aerobic activity declined during the
first part of the experiment and stabilised from 40 days onwards at lower respiration
rates. The major part of the respiration measured during the decomposition of
cordgrass is considered to be ‘non-bacterial’, most probably fungal (Padgett et al.,
1985). The increase of the total microbial activity, indicated by the enzymatic activity,
while the aerobic activity, represented by the respiration rate, is decreasing, suggests a
shift from aerobic to anaerobic activity in the microbial community. Since aquatic
fungi in general are aerobic (Rheinheimer, 1992), it can be assumed that the
enzymatic activity represents mainly bacterial activity. These findings suggest that
initially the fungal activity dominates and is then taken over by bacterial activity.
Nematodes and their influence on the microbial activity and
decomposition process
The four nematode species used in this experiment (Diplolaimelloides meyli,
Diplolaimelloides oschei, Diplolaimella dievengatensis and Panagrolaimus paetzoldi)
belong to the same functional group, the deposit feeders (Jensen, 1987) and are
assumed to feed mainly on bacteria (Moens and Vincx, 1997). Panagrolaimidae are
enrichment opportunists (Bongers et al., 1995): they are the first to colonise organic
deposits (Bouwman et al., 1984; Warwick, 1987; Bongers et al., 1991). Monhysterids,
to which the three other nematode species belong, are general opportunists and appear
in an early stage at locations with high organic input (Warwick, 1987; Alkemade et
al., 1993). They are typical ‘Aufwuchs species’ indicating their occurrence in
association with decaying organic matter but not, or to a much lesser extent, in the
surrounding sediments (Bouwman et al., 1984).

Chapter 2: Species-specific effects on decomposition
26
A faster initial increase of P. paetzoldi densities was expected since their generation
time is shorter than of the monhysterids (Moens and Vincx, 1998). To our surprise the
numbers of the monhysterids remained rather low throughout the experiment while
high densities of P. paetzoldi were reached and sustained. It is not likely that this
discrepancy is due to a lack of suitable food sources for the monhysterids. Cultivation
experiments of bacterivorous nematodes on different strains of bacteria, either
arbitrarily chosen or isolated from the natural habitat, have shown that bacterivorous
nematodes can grow and reproduce on a wide range of bacteria (pers. observations,
unpublished; Sohlenius, 1968; Venette and Ferris, 1998). Since the cordgrass
fragments were inoculated with a mixture of bacteria isolated from their natural
habitat the required diet must have been available for each nematode species.
An alternative explanation for the low monhysterid numbers might be found in the
presence of high ammonium concentrations building up in the microcosms due to
intense microbial activity. Although both panagrolaimids and monhysterids occur in
polluted sediments, monhysterids in general show a more sensitive reaction to
chemical compounds in lab experiments then do Panagrolaimidae (pers.
observations). These low numbers of the monhysterids may have influenced our
results. In situ nematode densities are in general one order of magnitude higher.
Moreover, other experiments, such as those by Alkemade et al. (1992a;b), in which
results contrasted with ours, obtained considerably higher nematode numbers.
However, Freckman (1988) noted that nematode biomass should not necessarily be
high to have an influence on the decomposition process. Since differences in
decomposition between treatments, and especially with the control treatment, were
found (see below) we assume that the numbers reached in all treatments were high
enough to have some influence on the decomposition process.
In analogy to many terrestrial soil systems, benthic bacterivorous nematodes are
thought to enhance decomposition processes by stimulating the activity of the
microbial, especially the bacterial, community (Findlay and Tenore, 1982; Alkemade
et al., 1992a,b). This was not confirmed by our experiments. Microbial activity was
amongst the highest in the absence of nematodes, resulting in a higher decomposition
rate. Observations in the course of the experiments suggested that the fungal growth
in the presence of the nematodes was lower compared to the control treatment. This is
confirmed by the lower respiration rate, mainly originating from fungal activity (see
above), in the presence of nematodes. Growth inhibition of fungal mycelia in the

Chapter 2: Species-specific effects on decomposition 27
presence of bacterivorous nematodes has already been reported in commercial
mushroom cultures (Kaufman et al., 1983; Grewal, 1991; Grewal and Richardson,
1991; Grewal and Hand, 1992). Since fungi are known to be important in the early
stages of decomposition of cordgrass (Newell et al., 1989; Valiela, 1995) this could be
a reason for the initially slower decomposition process in the presence of nematodes.
The contrast with the former studies, in which generally enhancement of the
decomposition process by nematodes is reported, may partly be explained by
differences in the experimental set up (Findlay and Tenore, 1982; Alkemade et al.,
1992a,b; Lillebø et al., 1999). When crushed cordgrass leaves were mixed with salt
marsh sediments bioturbation was found to be the most important factor for
stimulating the microbial community and the decomposition process (Alkemade et al.,
1992b). Through the movement of nematodes diffusion of oxygen and nutrients is
stimulated, resulting in a higher microbial activity and higher decomposition rate.
This factor is probably of minor importance on intact cordgrass fragments, as in our
experiment, or in the salt marshes where leaves decompose attached to the plant. Only
in late stages of the decomposition process the detritus becomes mixed with the
sediment and bioturbation is likely to play a role.
Grazing by bacterivorous nematodes on the bacteria has also been reported as a source
of stimulation (a.o. Abrams and Mitchell, 1980). Grazing can have two important
consequences. Firstly the bacterial community is kept in the active growth phase,
implying a higher demand of nutrients by the bacteria and thus resulting in a higher
decomposition rate of the organic matter. Secondly, when feeding on bacteria, an
excess of nutrients is consumed, because of the higher C/N ratio of nematodes
compared to bacteria (Anderson et al., 1983; Ferris et al., 1997). After excretion, this
nitrogen is again available for the bacterial community or can bind to the cordgrass
detritus (White and Howes, 1994; Valiela, 1995). It can be questioned whether the
recycled nitrogen can be of importance for the activity of bacteria in eutrophic
systems such as salt marshes. We assume that this can only enhance microbial activity
and consequently the decomposition process in N-limited systems, such as many
terrestrial soil systems.
Several elements indicate that nematode grazing had an important influence on the
microbial activity in our experiment. Firstly the low bacterial activity in the presence
of P. paetzoldi is most probably due to overgrazing since high feeding rates on
bacteria have been observed for this species (Moens, unpubl) and the decrease in

Chapter 2: Species-specific effects on decomposition
28
activity occurred when nematode densities reached over 50000 individuals per
microcosm. Secondly the absorption of excreted N to the detritus may partly explain
the high nitrogen content in the cordgrass leaves in the presence of the nematodes.
This could mask the selective removal of nitrogen rich compounds from the detritus
by the microbial community.
Many marine and brackish-water nematodes are known to secrete mucus which is
rapidly colonised by high bacterial densities (Riemann and Schrage, 1978; Nehring et
al., 1990; Jensen, 1996). Recently Riemann and Helmke (2002) suggested a
commensalistic relationship between the mucus-secreting nematodes and the
associated bacteria. Nematodes are believed to discharge hydrolytic enzymes in the
mucus which break down, alone or together with the bacterial enzymes, chemical
compounds. The hydrolysed compounds, such as sugars, could then be directly
consumed by the nematodes, thus yielding extra nutrients directly from the detritus. A
better ability of P. paetzoldi to secrete the necessary enzymes and subsequently
assimilate the organic compounds could explain (i) why the decomposition at the end
of the experiment in their presence is similar to other treatments despite the reduced
microbial activity and (ii) why they can sustain high densities while their expected
food source, bacteria, is being repressed. This trait of P. paetzoldi might facilitate
their growth in sites with high organic input (Bongers et al., 1995).
Bacteria growing on the mucus tracks could be nematode specific (Riemann and
Schrage, 1978; Warwick, 1981; Jensen, 1996), implying a potentially different
influence of the bacterial community on the decomposition process in the presence of
each nematode species. The results of a bacterial community analysis from this
experiment are presented in a next paper1 (De Mesel et al., 2004).
Conclusions
Despite the lack of stimulation by the nematodes of the decomposition process and of
the activity of the associated microbial community, differences were found dependent
on the identity of the nematode species present. Panagrolaimus paetzoldi was able to
reach and sustain higher densities even though the bacterial community was
repressed, suggesting a different life strategy compared to the three monhysterid
1 see Chapter 3

Chapter 2: Species-specific effects on decomposition 29
nematode species. Within the closely related monhysterids differences in population
development and in influences on the decomposition process were seen. Our results
show that the use of ‘functional groups’ is rather inadequate when studying the
relationship between nematodes and ecosystem processes. Integrating information on
life strategies might be more reliable. These findings also imply that the idea of
species redundancy, assuming that species within a functional group have the same or
a very similar influence on an ecosystem process, might not be applicable in
nematode communities. It seems that the influence of every species is, or can be,
different, agreeing with the conclusions of Mikola and Setälä (1998), that
relationships between nematode species diversity and ecosystem functioning may be
largely idiosyncratic.
Finally, a comparison of our results with those of previous studies highlights the
influence of experimental conditions on the obtained results. As a consequence
caution is required when extrapolating results to a natural situation. However, the
value of controlled experiments to generate hypotheses about in situ processes should
not be underestimated since many processes can not be studied in the field due to
practical problems.
Acknowledgements
We thank Annick Van Kenhove for the chemical analyses and Wim Bert for
identification of one of the nematode species. Prof. Dr. J. Vanfleteren is
acknowledged for the use of the Victor Multilabel counter. Jan Vanaverbeke and two
anonymous referees provided constructive comments on the manuscript. The first and
second author acknowledge grants from the Flemish Institute for the Promotion of
Scientific-Technological Research (I.W.T.). The last author is a postdoctoral fellow
with the Flemish Fund for Scientific Research. Further financial support was obtained
from Ghent University in BOF- projects 1205398 and 011060002.

Published as:
De Mesel, I, Derycke, S, Moens, T, Van der Gucht, K, Vincx, M, Swings, J (2004)
Top-down impact of bacterivorous nematodes on the bacterial community structure: a
microcosm study. Environmental Microbiology 6(7): 733-744
Chapter 3
Top-down impact of bacterivorous nematodes on the bacterial community structure: a microcosm experiment

Chapter 3: Impact of nematodes on a bacterial community
33
Abstract
The influence of bacterivorous nematodes (Diplolaimelloides meyli,
Diplolaimelloides oschei, Diplolaimella dievengatensis, Panagrolaimus paetzoldi) on
the development of a bacterial community growing on decaying cordgrass detritus
was studied in laboratory microcosm experiments. Cordgrass leaves were incubated
on a sediment surface with a natural bacterial mixture containing bacteria from
sediment, cordgrass detritus and habitat water. The four nematode species were
applied separately to the microcosms; controls remained without nematodes. Samples
were taken seven times over a 65-day period. The bacterial community structure was
analysed by means of DGGE of the 16S rRNA genes. Multi Dimensional Scaling
showed grouping of the samples per treatment. Analysis of Similarities indicated that
the differences between treatments were significantly larger than differences within
treatments. Our results suggest that nematodes can have a significant structuring top-
down influence on the ‘pool’ of bacteria growing on the detritus, even at low
densities. Dissimilarities were similar between all treatments. Differences in bacterial
community composition within the treatments with monhysterids (D. meyli, D. oschei
and D. dievengatensis) can be explained by species-specific food preferences.
Panagrolaimus paetzoldi on the other hand feeds unselectively, and thus affects the
bacterial community differently. A top-down effect of the nematodes on the diversity
of the bacterial community was only evident under high grazing pressure, i.e. in the
presence of P. paetzoldi.

Chapter 3: Impact of nematodes on a bacterial community
35
Introduction
Heterotrophic bacteria and fungi are an important link in the functioning of many
ecosystems, particularly for the mineralization of nutrients (Valiela, 1995). An
increase in nutrient mineralization has been reported in the presence of grazers of
bacteria, mainly nematodes and protists (Johannes, 1965; Ferris et al., 1997;
Bonkowski et al., 2000). Grazers can affect bacterial communities at different levels.
They are able to influence bacterial activity, either stimulatory (Findlay and Tenore,
1982; Alkemade et al., 1992a,b) or inhibitory (De Mesel et al., 2003). This can be a
direct effect of grazing (Ingham et al., 1985), but bioturbation (Abrams and Mitchell,
1980; Alkemade et al. 1992b) and secretion of mucus trails (Riemann and Schrage,
1978; Jensen, 1996) by nematodes can also be important. Grazing impacts on the
bacterial community structure have also been reported. Protists can cause a size shift
by grazing the medium-sized part of the community (Hahn and Höfle, 2001). Van
Hannen et al. (1999) were the first to demonstrate changes in the bacterial species
composition under protist grazing pressure, while Rønn et al. (2002) found that these
changes were protist-species specific. The structuring influence of nematodes,
however, has not yet been studied well. Only Griffiths et al. (1999) reported bacterial
shifts under nematode grazing using phospholipid fatty acid analysis, community
level physiological profiling (using BIOLOG plates) and DNA fingerprinting
(Denaturing Gradient Gel Electrophorsis). The shifts in the bacterial community
composition would presumably be the result of selective grazing. For example, large
and filamentous bacterial cells can escape uptake by some nematodes due to their
small buccal cavity (Tietjen, 1980). Additionally, experimental data suggest that some
nematode species can distinguish between and preferentially consume different strains
of bacteria, the preference differing between even closely related nematode species
(Tietjen and Lee, 1977; Moens et al., 1999).
The aims of the present paper were: (i) to investigate the potential effect of
bacterivorous nematodes on the bacterial community composition and diversity and
(ii) to assess whether any such effects are nematode-species specific. Cordgrass
(Spartina alterniflora) detritus was used as a substrate for the bacterial community.
Cordgrass is often the dominant vegetation in the lower parts of salt marshes and
decomposes mainly in situ, on the sediment surface (Groenendijk, 1984; Newell et al.,

Chapter 3: Impact of nematodes on a bacterial community
36
1985; Buth and Voesenek, 1988). High densities of bacterivorous nematodes occur on
the detritus (Montagna and Ruber, 1980; Reice and Stiven, 1983; Buth and de Wolf,
1985; Hemminga and Buth, 1991).
Four bacterivore nematode species commonly found on decaying cordgrass leaves
(Diplolaimelloides meyli, Diplolaimelloides oschei, Diplolaimella dievengatensis and
Panagrolaimus paetzoldi) were inoculated separately in microcosms containing
cordgrass detritus and a natural bacterial inoculum; controls without nematodes were
incubated in parallel. P. paetzoldi (Rhabditida) is an extreme coloniser, appearing
promptly at sites with high organic input where they can quickly reach high densities.
The three other species belong to the Monhysterida and are also characteristic for
organically enriched sites. Rhabditida and Monhysterida generally dominate the
nematode community on decaying phytodetritus in salt marshes (Warwick, 1987).
The choice of the three species within a single family (Monhysteridae) allows testing
for different effects on the bacterial community between closely related grazer
species, with a similar life strategy.
At seven sampling occasions over a period of 65 days, the bacterial communities were
analysed by means of Denaturing Gradient Gel Electrophoresis (DGGE) of the 16S
rRNA gene. DGGE allows a fast screening of complex bacterial communities, without
going into detail on community assemblage or functional aspects of the bacterial
community, which was beyond the scope of this study. The use of DGGE was first
introduced in ecological studies by Muyzer et al. (1993), and has since proven to be
very useful in analysing bacterial communities originating from a large number of
habitats (Muyzer, 1999). Recently, this technique has been applied successfully in a
number of experimental studies (Degans et al., 2002; Massana and Jürgens, 2003;
Zöllner et al., 2003).

Chapter 3: Impact of nematodes on a bacterial community
37
Materials and Methods Experimental set-up and sampling
The experimental set-up has been described in detail in De Mesel et al. (2003).
Briefly, green cordgrass leaves (Spartina anglica) collected in the Paulina Salt Marsh
(Westerscheldt, The Netherlands) were cleaned with ethanol (70%) and artificial sea
water (ASW) (Dietrich and Kalle, 1957), put in petri dishes (∅ 90 mm) on cleaned
sediment saturated with ASW with a salinity of 20, and inoculated with a microbial
inoculum. This inoculum consisted of bacteria from fresh and decomposing cordgrass
leaves, sediment and habitat water and from the nematode cultures. This mixture was
filtered over Whatman GF/C filters to remove flagellates and other small eukaryotes.
All petri dishes were inoculated with 2 ml of this suspension. After an incubation of
24h, nematodes were added to the microcosms.
Four nematode species, commonly found on decaying cordgrass in the Paulina Salt
Marsh (Moens, unpublished) were selected: Diplolaimelloides meyli,
Diplolaimelloides oschei, Diplolaimella dievengatensis and Panagrolaimus paetzoldi.
These species are available in monospecific cultures in the Marine Biology Section.
Each microcosm, except for the controls, received 36 individuals of a single nematode
species.
Microcosms were incubated in the dark at 20°C.
Three replicate microcosms per treatment were destructively sampled after each of the
following incubation times: 10, 20, 30, 40, 50, 55 and 65 days. However, for the
following days and treatments, only two replicates were successfully analysed: D.
meyli: day 55; D. oschei: day 30, 50 and 65; P. paetzoldi: day 40; control: day 40 and
55. Detritus fragments were homogenized in sterile ASW with a salinity of 20 to
collect the bacterial community. 4 ml of this suspension was centrifuged at 13000
rpm. After removing the supernatant the pellet was preserved at –80°C.

Chapter 3: Impact of nematodes on a bacterial community
38
DNA-extraction and -purification
DNA was extracted as described by Zwart et al. (1998) using the bead-beating method
concomitant with phenol extraction and ethanol precipitation. The DNA was purified
on a Wizard column (Promega, Madison, WI, USA) according to the manufacturer’s
recommendations.
PCR and DGGE
The amplification of DNA fragments and the DGGE analysis were performed as
described by Van der Gucht et al. (2001). DNA fragments of about 250 bp were
amplified using primers for the V3-region: F357-GC primer (5’-
CGCCCGCCGCGCCCCGCGCCCGGCCCGCCGCCCCCGCCCCCCTACGGGAG
GCAGCAG-3’) and the R518 primer (5’-ATTACCGCGGCTGCTGG-3’) which are
specific for most bacteria (Muyzer et al., 1993). The temperature cycling started with
a preincubation of 5 minutes at 94°C. 20 cycles were performed consisting of
denaturation at 94°C for 1 minute, annealing at 65°C for 1 minute, with the
temperature decreasing 0.5°C every cycle until a temperature of 56°C was reached,
and primer extension at 72°C for 1 minute. During the next 5 cycles the annealing
temperature stayed at 55°C for 1 minute. Finally temperature was held at 72°C for 10
minutes.
DNA concentrations were checked visually by analysing 5µl of the product on 1%
agarose gel, staining with ethidium bromide, and comparison with a molecular weight
marker (Smart ladder, Eurogenetic). 35µl of the samples were loaded on the DGGE
gels. The linear gradient on the DGGE-gels increased from 40% at the top to 75% at
the bottom. To compare the banding patterns between gels a marker was loaded and
some experimental samples were run on different gels. Additionally the information
on the DNA sequence of a number of bands was used to make a reliable alignment of
the gels.
The software package Bionumerics 5.1 (Applied Maths BVBA, Kortrijk, Belgium)
was used to analyse the banding patterns. The Bionumerics software measures an
optical density profile through each lane (corresponding to a single sample), identifies
band positions, and calculates the percent contribution of the intensity of each band to

Chapter 3: Impact of nematodes on a bacterial community
39
the total intensity of the lane. This procedure yielded a matrix with the relative
intensity of each band in all samples.
Sequencing of excised DGGE bands
The procedure used was based on Van der Gucht et al. (2001), but with some small
modifications. Individual bands were excised from the DGGE-gel and put in a sterile
recipient to which 50 µl 1X TE was added. DNA could diffuse form the acrylamide
gel to the TE-buffer during incubation overnight at 4°C. 5µl of the buffer was used for
amplification (PCR mix and temperature cycle as described above) and the position of
the band was checked on a DGGE gel. DNA from the bands was then amplified with
the F357GC primer (5’-CGCCCGCCGCGCCCCGCGCCCGGCCCGCCGCCCCCG
CCCCCCTACGGGAGGCAGCAG-3’) and the R518+ primer (5’-GCGTTCTTCAT
CGTTGCGAGATTACCGCGGCTGCTGG-3’) using the temperature cycling as
described by Van der Gucht et al. (2001), followed by a sequencing reaction with a
Big Dye TM terminator ready reaction kit (Applied Biosystems, Foster City, CA,
USA) and the Stef1Tex primer (5’-GCGTTCTTCATCGTTGCGAG -3’). The
temperature cycling was as described by Zwart et al. (1998). Sequencing reactions
were analysed on an Applied Biosystem 3100 genetic analyser.
Bands were identified by comparing the partial 16S rRNA gene sequences with
GenBank and EMBL sequences with BLAST (see table 2) (Altschul et al., 1990).
Bacterial diversity
The Shannon Wiener Index (H’) was used to express bacterial diversity and was
calculated as explained by Magurran (1988), based on the relative intensities of the
bands:
H’ = ∑=
=
−ni
iii pp
1ln
n represents the number of bands in the sample and pi the relative intensity of the ith
band. The index indicates how the intensity of the bands is distributed within a
sample.

Chapter 3: Impact of nematodes on a bacterial community
40
Statistical analysis of the DGGE fingerprints
The banding profiles of the DGGE gels were analysed by means of Multi
Dimensional Scaling (MDS) using the PRIMER software package and based on the
arcsin squareroot transformations of the relative band intensities. The Bray-Curtis
index was chosen as similarity coefficient. MDS analysis places samples in a multi
dimensional space, based on the similarities between them. This multi-dimensional
space can be represented by a 2D ordination plot, with the stress value indicating how
faithfull the high-dimensional relationships between the samples are represented in
the 2D plot. This plot is very easy to interpret as similar samples are placed close
together, and distance between samples increases as similarity decreases (Clarke,
1993; Clarke and Gorley, 2001; Clarke and Warwick, 2001).
Analysis of similarities (ANOSIM) is a non-parametric permutation procedure, based
on the similarity matrix underlying the ordination of the samples. ANOSIM was used
to test for significant differences between treatments (Clarke, 1993; Clarke and
Gorley, 2001; Clarke and Warwick, 2001).
Similarity percentage analysis (SIMPER) was used to weight the contribution of each
band to the similarity/dissimilarity within/between treatments and to seek for those
bands which were important during the succession in each treatment (Clarke and
Gorley, 2001; Clarke and Warwick, 2001).
Univariate 2-way analysis of variance (ANOVA) and the Tukey HSD post hoc test
were used to test for differences in the Shannon-Wiener Index (H’) between
treatments, times and treatment x time (Statistica software).
Nucleotide sequence accession numbers
All partial sequences have been deposited in the EMBL database under accession nos
AY510428-AY510447

Chapter 3: Impact of nematodes on a bacterial community
41
Results and Discussion Effects on bacterial community composition
The bacterial community composition was analysed by means of Multi Dimensional
Scaling (MDS) and Analysis of Similarities (ANOSIM), both non-parametric
multivariate tests for changes in community structure (Clarke, 1993; Clarke and
Warwick, 2001). The use of either relative band intensities, or of a reduced data
matrix based on presence/absence of bands, to analyse community structure has been
discussed and questioned before (Schauer et al., 2000). Both are commonly used,
depending on the type of dataset studied. We believe that the use of relative band
intensities in this study was justified because bands could clearly be seen fading away
or appearing and intensifying through the succession. Using a binary dataset would
strongly influence our analysis by overestimating very weak, sometimes uncertain
bands. Moreover, Muylaert et al. (2002) showed that the use of relative band
intensities is better suited for ordination analysis than a binary data matrix.
Ordinations have been found very useful in ecological research in general and for
processing community analyses by means of DGGE in specific (Fromin et al., 2002).
Figure 1: MDS ordination plot based on the relative band intensities of the DGGE profiles,
with the letter referring to the treatments and the number to the sampling day.
D. meyli (A)
D. oschei (B)
P. paetzoldi (C)
D. dievengatensis (D)
control (S)
A10
A10
A10A20
A20
A20
A30
A30
A30
A40 A40A40
A50
A50
A50A55
A55
A65
A65
A65
B10
B10
B10
B20
B20
B20
B30 B30
B40B40
B40B50
B50
B55
B55B55
B65
B65
C10
C10C10 C20
C20 C20
C30 C30
C30
C40
C40
C50
C50
C50C55
C55
C55
C65
C65
C65
D10
D10
D10D20
D20
D20D30
D30D30 D40
D40D40
D50D50
D50D55D55
D55D65
D65
D65
S10
S10
S10
S20S20 S20
S30S30
S30
S40
S40
S50
S50
S50
S55
S65
S65
S65
Stress: 0.25
D. meyli (A)
D. oschei (B)
P. paetzoldi (C)
D. dievengatensis (D)
control (S)
D. meyli (A)
D. oschei (B)
P. paetzoldi (C)
D. dievengatensis (D)
control (S)
A10
A10
A10A20
A20
A20
A30
A30
A30
A40 A40A40
A50
A50
A50A55
A55
A65
A65
A65
B10
B10
B10
B20
B20
B20
B30 B30
B40B40
B40B50
B50
B55
B55B55
B65
B65
C10
C10C10 C20
C20 C20
C30 C30
C30
C40
C40
C50
C50
C50C55
C55
C55
C65
C65
C65
D10
D10
D10D20
D20
D20D30
D30D30 D40
D40D40
D50D50
D50D55D55
D55D65
D65
D65
S10
S10
S10
S20S20 S20
S30S30
S30
S40
S40
S50
S50
S50
S55
S65
S65
S65
Stress: 0.25

Chapter 3: Impact of nematodes on a bacterial community
42
The MDS analysis on all samples from each sampling event showed a clustering per
treatment (fig. 1). A pronounced time aspect was observed in the presence of
nematodes, but was less clear in the control. Although the stress value, indicating how
faithful the high-dimensional relationship among the samples is represented in the 2D
plot, was rather high, ANOSIM indicated that the variation in DGGE profiles was
higher between samples from different treatments than between samples within one
treatment (table 1). Similar observations have been made under protist grazing
pressure in terrestrial soil systems (Rønn et al., 2002). The pairwise comparisons
further showed that dissimilarities between all treatments were similar.
Table 1: Results of the ANOSIM routine to analyse the differences between treatments.
A first explanation for the differences between treatments may be related to
differences in feeding habits between the nematode species used here.
Panagrolaimidae are typical enrichment opportunists (Bongers et al., 1995). Direct
observations indicate they non-selectively consume food particles at high rates as they
show a constant pumping activity of the oesophagus (Moens, unpublished). Since the
passage through the gut is very fast, it is plausible that not all ingested food particles
are digested, implying that some viable cells are again released. The subset of
bacterial cells that maintains viability after passage through the gut is assumed to be
protected mainly by their cell wall. Within the Monhysteridae however, bacterial
selection would rather occur at the level of food ingestion. Moens et al. (1999) carried
out a number of experiments with the monhysterid species used in our study, showing
they can select between bacterial strains. Chemotaxic responses in which ‘taste’ and
‘smell’ of the prey would be of importance may determine this selection (Moens et
Comparison R Statistic
Significance level (%)
D. meyli vs D. oschei 0.727 0.1 D. meyli vs P. paetzoldi 0.838 0.1 D. meyli vs D. dievengatensis 0.839 0.1 D. meyli vs control 0.769 0.1 D. oschei vs P. paetzoldi 0.689 0.1 D. oschei vs D. dievengatensis 0.637 0.1 D. oschei vs control 0.774 0.1 P. paetzoldi vs D. dievengatensis 0.448 0.1 P. paetzoldi vs control 0.602 0.1 D. dievengatensis vs control 0.652 0.1

Chapter 3: Impact of nematodes on a bacterial community
43
al., 1999 and references therein). This implies that chemical cues emanating from the
bacteria rather than bacterial cell wall structure influence their palatability and uptake
by monhysterid nematodes. The nematode-species-specific preference for different
bacterial strains, observed by Moens et al. (1999), may have caused the development
of different bacterial communities in the presence of each monhysterid nematode
species.
The structure of the bacterial community may also have been affected by some other
nematode traits that have been found important in stimulating bacterial activity.
Bioturbation by the nematodes enhances diffusion of nutrients and oxygen through
sediments (Abrams and Mitchell, 1980; Alkemade et al., 1992b; Aller and Aller,
1992) and could thus affect the bacterial community structure. However, we assume
that bioturbation is mainly of importance when organic matter is mixed in the
sediment, and not, as in our experiment, when larger organic fragments decompose on
the sediment surface (De Mesel et al., 2003). Nematodes can secrete mucus trails
which are characterised by a dense growth of bacteria (Riemann and Schrage, 1978;
Jensen, 1996). These bacteria could be nematode-species-specific, but research on
their identity is, to our knowledge, non-existent. However, this kind of ‘cultivation’ of
bacteria by the nematodes is presumably of minor importance, since most bands can
be found in two or more treatments, but with other relative intensities and/or different
frequencies.
Sequencing of DGGE-bands provided some taxonomic information on the bacteria.
The DNA fragments we cut out of the DGGE gel were 200bp at most; therefore
information at the species level could not be obtained. The identification is restricted
to the genus or higher taxonomic level (see table 2)
Most of the sequences showed high similarities (>95%) with species commonly found
in marine and brackish environments. Members of the genus Clostridium are often
found in marine sediments and on decaying vegetation and are characteristic for
anaerobic environments. They have the ability to break down a large number of
organic compounds (Hippe et al., 1992). Cytophaga species are marine cellulose
degraders (Reichenbach, 1992), while Neptunomonas species are marine bacteria
breaking down aromatic hydrocarbons (Hedlund et al., 1999). Oceanospirillum
species are often associated with particulate plant matter in marine sites (Pot et al.,
1992). One Arcobacter species (A. nitrofigilis) has been described associated with

Chapter 3: Impact of nematodes on a bacterial community
44
Table 2: Sequence similarities of the excised DGGE bands; for the position on the DGGE
fingerprints, see fig. 2. Unidentified species 7 to 9 only showed very low similarity with
sequences of the database.
Spartina roots in salt marshes (McClung et al., 1983). Sulfurospirillum and
Bdellovibrio species occur in sediments of marine and freshwater systems (Ruby,
1992; Kersters et al., 2003). Unidentified species 1 belongs to the CFB-group
bacteria. Unidentified species 2 to 6 showed high similarities with other partial
Band no.
Band name Closest relative Percentage similarity
Accession no.
1 Clostridium sp.1 Clostridium algidixylanolyticum
99% AY510428
2 Clostridium sp.2 Clostridium lentocellum
95% AY510429
3 Clostridium sp.3 Clostridium sp. 97% AY510430 4 Unidentified 1 Uncultured CFB-
group bacterium from ocean floor
96% AY510431
5 Unidentified 2 Uncultured bacterium clone s3 form marine sediment
97% AY510432
6 Unidentified 3 Uncultured eubacterium clone F10.2
96% AY510433
7 Unidentified 4 Marine bacterium SCRIPPS_739
100% AY510434
8 Sulfurospirillum sp. Sulfurospirillum sp. 96% AY510435 9 Cytophaga sp. Cytophaga sp. 100% AY510436 10 Unidentified 5 Uncultured gamma
proteobacterium clone CD3B12
97% AY510437
11 Arcobacter sp. Arcobacter sp. 98% AY510438 12 Clostridium related Uncultured bacterium
RSb40 100% AY510439
13 Oceanospirillum sp.
Oceanospirillum pusillum
99% AY510440
14 Unidentified 6 Uncultured bacterium clone ZB131
98% AY510441
15 Bdellovibrio sp. Bdellovibrio sp. 98% AY510442 16 Neptunomonas sp. Neptunomonas
naphthovorans 96% AY510443
17 Anaerofilum sp. Anaerofilum pentosovorans
99% AY510444
18 Unidentified 7 no match AY510445 19 Unidentified 8 no match AY510446 20 Unidentified 9 no match AY510447

Chapter 3: Impact of nematodes on a bacterial community
45
sequences in the database of which no taxonomic information was available; no match
with available sequences (<<95%) was obtained for Unidentified species 7 to 9 (table
2).
Ā ⎯Si ⎯Si /SD ⎯Si % ∑ ⎯Si % Diplolaimelloides meyli (average similarity: 47.56) Clostridium sp.2 0.23 6.78 1.67 14.25 14.25 Arcobacter sp. 0.19 4.87 1.27 10.24 24.49 Unknown 3 0.11 3.53 2.45 7.42 31.92 Unidentified 1 0.14 3.44 1.28 7.23 39.15 Clostridium sp.1 0.1 2.8 1.22 5.88 45.02 Diplolaimelloides oschei (average similarity: 47.54) Clostridium sp.2 0.25 8.62 2.3 18.13 18.13 Unknown 8 0.19 6.41 2.18 13.49 31.61 Arcobacter sp. 0.19 5.44 1.32 11.45 43.06 Unidentified 1 0.11 3.3 2.1 6.95 50.01 Clostridium sp.1 0.12 3.06 1.23 6.43 56.44 Panagrolaimus paetzoldi (average similarity: 42.48) Clostridium sp.2 0.28 10.57 3.66 24.88 24.88 Unidentified 5 0.12 3.66 1.24 8.62 33.49 Clostridium sp.1 0.12 3.02 1.04 7.12 40.61 Anaerofilum sp. 0.1 2.41 0.92 5.67 46.28 Arcobacter sp. 0.14 2.29 0.51 5.38 51.66 Diplolaimella dievengatensis (average similarity: 52.74) Clostridium sp.2 0.28 10.52 3.29 19.95 19.95 Arcobacter sp. 0.27 9.23 2.45 17.5 37.45 Unidentified 7 0.13 4.35 1.92 8.05 45.7 Oceanospirillum sp. 0.13 3.54 1.2 6.71 52.41 Clostridium sp.1 0.12 3.27 1.44 6.2 58.6 control (average similarity: 48.83) Clostridium sp.2 0.28 9.53 2.57 19.52 19.52 Arcobacter sp. 0.21 6.72 2.36 13.77 33.29 Unknown 5 0.15 4.06 1.23 8.32 41.61 Oceanospirillum sp. 0.14 3.96 1.64 8.11 49.72 Clostridium sp.1 0.11 3.27 1.34 6.7 56.42
Table 3: Results of the Simper analysis giving the similarities within treatments. Five species
which contribute most to the similarity are listed. Āi: average abundance of the i th species
over all samples of the treatment; ⎯Si: contribution of the i th species to the total similartity;
⎯Si/SD: the value of the i th species as a discriminating species; ⎯Si%: percentage contribution
of the i th species to the total similarity; ∑ Si %: cumulative contribution to the total similarity.
Similarity percentages (SIMPER) analysis was used to look for those bacterial species
which contributed most to the average similarity within treatments and average
dissimilarity between treatments (Clarke and Gorley, 2001) (table 3 and 4; see table 2
for sequence information of the excised bands). Clostridium sp.1 and sp.2 and
Arcobacter sp. were prominent members of the bacterial community in each
treatment. Density differences in Arcobacter sp. generally explained a substantial part
of the dissimilarity between treatments. One unknown species occurred only in the

Chapter 3: Impact of nematodes on a bacterial community
46
presence of D. oschei and explained most of the dissimilarity with other treatments.
For the control, the presence of Anaerofilum sp. accounted for much of the
dissimilarity with the nematode treatments.
Table 4: Results of the Simper analysis giving the dissimilarities between treatments. Three
species which contribute most to the dissimilarity are listed.⎯δi: contribution of the i th species
to the total dissimilartity;⎯δi/SD: the value of the i th species as a discriminating species; ⎯δ%:
percentage contribution of the i th species to the total dissimilarity; ∑⎯δi%: cumulative
contribution to the total dissimilarity.
Ā ⎯δi/SD ⎯δi% ∑⎯δi% D. meyli vs D. oschei (average dissimilarity 68.27) Unknown 8 3.93 2.72 5.75 5.75 Arcobacter sp. 2.74 1.38 4.02 9.77 Unknown 18 2.45 1.12 3.58 13.35 D. meyli vs P. paetzoldi (average dissimilarity: 75.46) Arcobacter sp. 3.54 1.44 4.69 4.69 Unidentified 1 2.82 1.55 3.74 8.43 Unidentified 5 2.59 1.75 3.44 11.87 D. meyli vs D. dievengatensis (average dissimilarity: 68.91) Arcobacter sp. 2.96 1.41 4.29 4.29 Unknown 1 2.37 1.11 3.43 7.72 Unknown 14 2.34 1.64 3.4 11.12 D. meyli vs control (average dissimilarity: 68.96) Anaerofilum sp.2 2.95 1.46 4.27 4.27 Unidentified 7 2.75 1.55 3.99 8.27 Arcobacter sp. 2.56 1.43 3.71 11.97 D. oschei vs P. paetzoldi (average dissimilarity: 70.59) Unknown 8 3.88 2.41 5.5 5.5 Arcobacter sp. 3.53 1.46 5 10.51 Unidentified 5 2.59 1.72 3.67 14.18 D. oschei vs D. dievengatensis (average dissimilarity: 61.80) Unknown 8 3.64 2.27 5.89 5.89 Arcobacter sp. 2.78 1.31 4.5 10.39 Unknown 18 2.4 1.07 3.88 14.28 D. oschei vs control (average dissimilarity; 67.39) Unknown 8 3.91 2.6 5.8 5.8 Anaerofilum sp.2 2.9 1.43 4.3 10.1 Arcobacter sp. 2.39 1.34 3.55 13.65 P. paetzoldi vs D. dievengatensis (average dissimilarity: 61.61) Arcobacter sp. 4.11 1.49 6.67 6.67 Unidentified 7 2.78 2.12 4.51 11.18 Unidentified 2 2.57 1 4.18 15.36 P. paetzoldi vs control (average dissimilarity: 67.37) Arcobacter sp. 3.56 1.67 5.28 5.28 Anaerofilum sp.2 2.67 1.33 3.96 9.24 Unidentified 5 2.64 1.74 3.92 13.17 D. dievengatensis vs control (average dissimilarity: 61.20) Anaerofilum sp.2 2.71 1.36 4.43 4.43 Unidentified 7 2.71 2.12 4.43 8.86 Arcobacter sp. 2.43 1.31 3.97 12.83

Chapter 3: Impact of nematodes on a bacterial community
47
Figure 2: DGGE fingerprints with indications of the band classes. Numbers refer to table 2.
1
2
4
67
11
13
12
1617
10 days20 days30 days40 days50 days55 days65 days
20
1
2
4
67
11
13
12
1617
10 days20 days30 days40 days50 days55 days65 days
20
1
211
9
158
13
17
10 days20 days30 days40 days50 days55 days65 days
Diplolaimelloides meyli
Diplolaimelloides oschei
2
1
11
133
14
4 10 days20 days30 days40 days50 days55 days65 days
Panagrolaimus paetzoldi
11
1
2
4
510
6
10 days20 days30 days40 days50 days55 days65 days
Diplolaimella dievengatensis
11
1
2
4
610
13 10 days20 days30 days40 days50 days55 days65 days
Control
1918
18
18
1
211
9
158
13
17
10 days20 days30 days40 days50 days55 days65 days
1
211
9
158
13
17
10 days20 days30 days40 days50 days55 days65 days
Diplolaimelloides meyli
Diplolaimelloides oschei
2
1
11
133
14
4 10 days20 days30 days40 days50 days55 days65 days
Panagrolaimus paetzoldi
11
1
2
4
510
6
10 days20 days30 days40 days50 days55 days65 days
Diplolaimella dievengatensis
11
1
2
4
610
13 10 days20 days30 days40 days50 days55 days65 days
Control
1918
18
18
1
2
4
67
11
13
12
1617
10 days20 days30 days40 days50 days55 days65 days
20
1
2
4
67
11
13
12
1617
10 days20 days30 days40 days50 days55 days65 days
20
1
2
4
67
11
13
12
1617
10 days20 days30 days40 days50 days55 days65 days
20
1
2
4
67
11
13
12
1617
10 days20 days30 days40 days50 days55 days65 days
20
1
211
9
158
13
17
10 days20 days30 days40 days50 days55 days65 days
1
211
9
158
13
17
10 days20 days30 days40 days50 days55 days65 days
Diplolaimelloides meyli
Diplolaimelloides oschei
2
1
11
133
14
4 10 days20 days30 days40 days50 days55 days65 days
Panagrolaimus paetzoldi
11
1
2
4
510
6
10 days20 days30 days40 days50 days55 days65 days
Diplolaimella dievengatensis
11
1
2
4
610
13 10 days20 days30 days40 days50 days55 days65 days
Control
1918
18
18
1
211
9
158
13
17
10 days20 days30 days40 days50 days55 days65 days
1
211
9
158
13
17
10 days20 days30 days40 days50 days55 days65 days
Diplolaimelloides meyli
Diplolaimelloides oschei
2
1
11
133
14
4 10 days20 days30 days40 days50 days55 days65 days
Panagrolaimus paetzoldi
11
1
2
4
510
6
10 days20 days30 days40 days50 days55 days65 days
Diplolaimella dievengatensis
11
1
2
4
610
13 10 days20 days30 days40 days50 days55 days65 days
Control
1918
18
18

Chapter 3: Impact of nematodes on a bacterial community
48
In all treatments a clear shift of bands with time could be observed (fig. 2), which, for
the nematode treatments, was also reflected in the MDS plot (fig. 1). Simper analysis
(results not shown) of the effect of time within each treatment was performed to
determine those bands which were most important in determining the succession. The
dynamics of Arcobacter sp. were important in all treatments. In the presence of D.
meyli, the appearance of Sulfurospirillum sp. from day 50 onwards and the dynamics
of four unknown species determined the major differences in time. In the treatment
with D. oschei the decreasing intensity of Clostridium sp.2, the disappearance of
Unidentified sp. 6 and the appearance of two unknown species were of major
importance. In the presence of P. paetzoldi the succession was primarily driven by
dynamics in Clostridium sp.1, Unidentified sp.2 and three unknown species. The
succession in the D. dievengatensis treatment was mainly determined by the
appearance of Oceanospirillum sp. after 20 days and the variations of Unidentified
sp.5 and two unknown species. In the control treatment the decreasing intensity of
Clostridium sp.2 and the variation of Unidentified sp. 4 and two unknown species
through time were the most important dynamics in the succession.
Effects on bacterial diversity
It was assumed that each band on the DGGE gel represents one ‘Operational
Taxonomic Unit’ (OTU). However, some shortcomings of the DGGE technique
should be considered when interpreting the results. Species that make up less than 1%
of the total bacterial community will not be represented on a DGGE-gel (Muyzer et
al., 1993). The presence of multiple heterogeneous rRNA operons might result in
multiple bands from one bacterial species (Nübel et al., 1996) and co-migration of
sequences might also bias the banding profile (Vallaeys et al., 1997). Therefore, the
diversity index calculated from the DGGE banding patterns should not be interpreted
as an absolute measure, but rather as an indication of bacterial diversity (Eichner et
al., 1999). Nübel et al. (1999) showed that the number of bands and the Shannon-
Wiener diversity index can be used as an estimate of the bacterial diversity. Since all
samples in our experiment were treated in the same way, it is expected that bias
operated uniformly on all samples allowing proper comparison of the results (Fromin
et al., 2002; Schauer et al., 2000).

Chapter 3: Impact of nematodes on a bacterial community
49
Bacterial diversity is represented by the Shannon-Wiener Index (fig. 3), which
integrates information on the number of bands and on the band-intensities. The
number of bands (data not shown) showed a similar pattern through time as the
Shannon-Wiener Index.
Figure 3: Shannon-Wiener Index (H’) during the cause of the experiment (average of 3 values
± 1 standard error).
Figure 3 shows that diversity reached a peak after 40 days in the controls and after 50
days in the presence of the monhysterids. Diversity was rather constant in the P.
paetzoldi treatment. A Tukey HSD pairwise post-hoc test only showed significant
differences within treatments in the presence of D. oschei, in which the index on day
10 was significantly lower than on day 50 (p<0.01) and day 55 (p<0.05), and in the D.
dievengatensis treatment where the index on day 50 was significantly higher than on
day 20 (p<0.01) and day 30 (p<0.05). No significant differences between treatments
could be found at any sampling occasion.
The generally lower bacterial diversity from 30 days onwards in the presence of P.
paetzoldi compared to the treatments with Monhysteridae could be ascribed to the
significantly higher densities of the former species. From that moment onwards, P.
paetzoldi reached densities up to 105 individuals per microcosm, while the
monhysterid nematode species (D. meyli, D. oschei, D. dievengatensis) attained
densities of a few thousand individuals (De Mesel et al., 2003). The high densities of
P. paetzoldi could have led to overgrazing of the bacterial community. This effect will
have been intensified by the unselective feeding habit of this species, enabling it to
eliminate a large proportion of the bacterial community. A high grazing pressure in
10 20 30 40 50 55 65Days
1.7
1.9
2.1
2.3
2.5
2.7
2.9
3.1
Shan
non-
Wie
ner I
ndex
(H')
D. meyliD. oscheiP. paetzoldiD. dievengatensiscontrol
10 20 30 40 50 55 65Days
1.7
1.9
2.1
2.3
2.5
2.7
2.9
3.1
Shan
non-
Wie
ner I
ndex
(H')
D. meyliD. oscheiP. paetzoldiD. dievengatensiscontrol
D. meyliD. oscheiP. paetzoldiD. dievengatensiscontrol

Chapter 3: Impact of nematodes on a bacterial community
50
the presence of P. paetzoldi was confirmed by measurements of respiration and
proteolytic activity of the microbial community (fig. 4a) (De Mesel et al., 2003). A
reduction of diversity at the bacterial level in the presence of high protist densities has
been observed before (van Hannen et al., 1999). Most information on the relationship
between grazer densities and prey diversity, however, originates from studies using
herbivores and plants as a model (Valiela, 1995 and references therein; Hillebrand,
2003). These studies confirm low diversity under high grazing pressure, and suggest
that highest prey diversity develops under intermediate grazing pressure. Applying
this to our experiment, the highest bacterial diversity would be expected in the
presence of the monhysterid species. This however was not unambiguously confirmed
by our findings. Only on day 50 and day 55 was bacterial diversity higher in all
monhysterid treatments versus the control; with none of these differences being
statistically significant. We assume that the monhysterid densities were not high
enough to reach an ‘intermediate disturbance’ level since the numbers of monhysterid
nematodes obtained in our experiments were situated at the lower end of the range
found in field conditions. Furthermore Herman and Vranken (1988) showed that
monhysterids only affected a small subset of the standing stock of bacteria, even at
higher densities than those reached in this experiment. Additionally we found that in
these treatments diversity was linked to nutritional value (N-content) of the leaves
(fig. 4b), rather than to nematode densities. The Shannon-Wiener Index was lowest at
the beginning and at the end of the experiment, and reached a peak around 50 days.
Initially, few opportunistic bacterial species will have benefited from the availability
of high concentrations of labile compounds. As these easily degradable compounds
became depleted, opportunists became replaced by a more diverse community of
decomposers. Towards the end of the experiment, the detritus mainly consisted of
refractory substances, which can only be broken down by few specialised species.
Similar dynamics in bacterial diversity were seen in the control, however, at the end
of the experiment an unexpected increase of bacterial diversity occurred. This
coincides with a peak of nitrogen content in the organic matter (De Mesel et al.,
2003), which may be a result of the binding of nitrogen-rich compounds to the
cordgrass detritus (White and Howes, 1994).

Chapter 3: Impact of nematodes on a bacterial community
51
A
B
Figure 4: A. Bacterial activity expressed as enzymatic activity (average of 3 samples ± 1
standard error) B. N-content of the cordgrass leaves, expressed as percentage of the initial
values (average of 3 replicates ± 1 standard error). The high loss the first 10 days is probably
due to leaching of nutrient rich compounds (De Mesel et al., 2003).
Conclusion
Our results suggest that the bacterial community composition in the microcosms was
shaped by a combination of bottom-up and top-down effects: the same bacterial
species appeared in different treatments, indicating that the types of bacteria occurring
were primarily determined by the substrate. However the composition and the relative
importance of different members of this ‘pool’ of bacteria was severely modified by
the grazing activities of the bacterivorous nematodes, even at relatively low densities.
A nematode effect on bacterial diversity was only evident under high grazing
pressure, as in the presence of P. paetzoldi.
0 0 10 20 30 40 50 55 65Days
5000
10000
15000
20000
25000
Enzy
mat
ic a
ctiv
ity (c
ount
s min
-1)
0 0 10 20 30 40 50 55 65Days
5000
10000
15000
20000
25000
Enzy
mat
ic a
ctiv
ity (c
ount
s min
-1)
0 10 20 30 40 50 55 65Days
0
2
4
6
8
10
12
mg
N
0 10 20 30 40 50 55 65Days
0
2
4
6
8
10
12
mg
N
0
2
4
6
8
10
12
mg
N
D.meyliD. oscheiP. paetzoldiD. dievengatensiscontrol
D.meyliD. oscheiP. paetzoldiD. dievengatensiscontrol

Chapter 3: Impact of nematodes on a bacterial community
52
Acknowledgements
We thank Nele Vloemans for the help with the DGGE analyses. The first and second
author acknowledge grants from the Flemish Institute for the Promotion of Scientific-
Technological Research (I.W.T.). Tom Moens is a postdoctoral fellow with the
Flemish Fund for Scientific Research. Further financial support was obtained from
Ghent University in BOF- projects 1205398 and 011060002.

De Mesel, I, Derycke, S, Swings, J, Vincx, M, Moens, T
Chapter 4
Effect of nematode diversity on organic matter decomposition and the associated microbial community: a
microcosm study

Chapter 4: Diversity related impacts on decomposition
55
Abstract
Diversity related and species-specific impacts of bacterivorous nematodes on the
decomposition rate of cordgrass detritus (Spartina anglica) and on the associated
microbial community were investigated using laboratory microcosm experiments.
Four bacterivorous nematode species (Diplolaimelloides meyli, Diplolaimelloides
oschei, Diplolaimella dievengatensis and Panagrolaimus paetzoldi) were added either
separately or in combinations of two or three species to the microcosms.
In contrast with previous reports, no stimulation of the decomposition process was
observed in the presence of nematodes. In absence of nematodes, a more extensive
growth of fungi, important in the early stages of cordgrass decay, might have caused
the higher decomposition rates. Fungal growth seemed to be hampered in the
presence of nematodes, but whether this was a consequence of direct or indirect
interactions (through facilitation of bacterial growth) could not be distinguished.
Still, clear differences in process rates were found between nematode treatments.
Panagrolaimus paetzoldi, reaching considerably higher densities than the other
nematode species, suppressed bacterial activity and diversity, probably due to
overgrazing. This was however not translated into a slower decomposition process.
Within treatments exclusively containing monhysterid nematode species (D. meyli, D.
oschei and D. dievengatensis), differences in microbial activity and decomposition
rates were found, but again no link was observed between both. The lack of
completely separate measurements of bacterial and fungal activity, different C-use
efficiencies of bacterial communities or direct assimilation of detrital compounds by
(some) nematode species may have complicated the establishment of any such link.
Our data did not support any hypothesis predicting enhancement of process rates with
an (initial) increase in numbers of nematode species, nor for redundancy among the
studied species. We rather obtained support for an idiosyncratic diversity model as
differences in the effect of bacterivore nematode species and species combinations
could not be predicted at the start of the experiment. This could be explained by the
unpredictable interactions between nematode species. Within the treatments
containing only monhysterid species, densities were much lower in the combination
treatments compared to the monospecific treatments, indicating interspecific
inhibitory interactions, probably of a chemical nature.

Chapter 4: Diversity related impacts on decomposition
57
Introduction
The role of biodiversity in the functioning of ecosystems has been discussed
extensively over the past few years. Many studies have concentrated on terrestrial
systems and emphasised on the link between plant diversity and productivity. These
results have led to a number of hypotheses, which generally describe a predictable
consequence of altering biodiversity (Schläpfer et al., 1999). The extremes range from
no effect of diversity as predicted by the (modified) null hypothesis, to an
improvement of the ecosystem functioning each time a species is added to the system,
referred to as the rivet hypothesis (Ehrlich and Ehrlich, 1981). Of particular interest is
the redundancy hypothesis (Walker, 1992), which states that it is of major importance
that all functions within an ecosystem are fulfilled; species within a functional group
are thought to be redundant. This does not exclude however, that ecosystem stability
could be better assured when each functional group comprises several equivalent
species (Walker, 1995) as their different responses to environmental factors can allow
persistence of the ecosystem function after disturbance. In contrast to the previous
hypotheses, the idiosyncratic model suggests that altering biodiversity generally
affects ecosystem functioning, but in an unpredictable manner (Lawton, 1994). This
results from a lack of proper understanding of the complex and often subtle
interactions between species within and across functional groups.
Fewer studies up to now have investigated the importance of diversity in other
ecosystem processes, such as decomposition. Decomposition is mainly a microbial
process and can be affected by higher trophic levels. Bacterial grazers, such as
protozoa and nematodes, have often been found to stimulate bacterial activity and
subsequently decomposition processes (e.g. Riemann and Schrage, 1978; Findlay and
Tenore, 1982; Alkemade 1992a, b, 1993).
Here we present the results of a microcosm study where four estuarine bacterivorous
nematode species from the same functional group have been used as a model to study
within-functional-group redundancy. The decomposition of cordgrass, often the
dominant vegetation in salt marshes (Howarth, 1993), was the ecosystem function of
interest. The effect of Diplolaimelloides meyli, Diplolaimelloides oschei,
Diplolaimella dievengatensis and Panagrolaimus paetzoldi, all commonly found on
decaying cordgrass in the Paulina Salt Marsh (Westerscheldt) (Moens, pers. comm.),

Chapter 4: Diversity related impacts on decomposition
58
was investigated. Analysis of the monospecific treatments has already shown some
species-specific effects, mainly on the bacterial community structure (De Mesel et al.,
2004) and activity (De Mesel et al., 2003), and to a lesser extent also on the
decomposition rate (De Mesel et al., 2003). Here we focus on the effects of pooling
two or three nematode species. We tested (1) whether species-specific effects
observed in the monospecific treatments were maintained in treatments with species
combinations and (2) whether there was any diversity-dependent effect on (i) the
decomposition rate, (ii) microbial activity, and (iii) bacterial diversity.
Materials and Methods Experimental set-up
The experimental set-up has been described in detail in De Mesel et al. (2003).
Briefly, green cordgrass leaves (Spartina anglica) were collected in the Paulina Salt
Marsh (Westerscheldt, The Netherlands). In the laboratory, they were cleaned with
ethanol (70%) and artificial sea water (ASW, Dietrich and Kalle, 1957). The leaves
were cut in fragments, and 1.9 g ± 0.2 g (wet weight) was put in petri dishes (∅ 90
mm) on sterile sediment saturated with ASW with a salinity of 20. A microbial
inoculum consisting of bacteria from fresh and decomposing cordgrass leaves,
sediment and habitat water and from the nematode cultures was then added. This
microbial inoculum was first filtered over Whatman GF/C filters to remove flagellates
and other small eukaryotes. All petri dishes were inoculated with 2 ml of this
suspension. After a 24 h incubation, nematodes were added to the microcosms.
Four nematode species, commonly found on decaying cordgrass in the Paulina Salt
Marsh (Moens, pers. comm.) were selected for this experiment: Diplolaimelloides
meyli, Diplolaimelloides oschei, Diplolaimella dievengatensis and Panagrolaimus
paetzoldi. The former three belong to the Monhysteridae, the latter one to the
Panagrolaimidae. Together with the Rhabditidae, these nematode families are
typically associated with decomposing plant detritus in aquatic sediments (Warwick,
1987). All species were applied separately to the microcosms (De Mesel et al., 2003),
and in two- and three- species combinations. Each time, a total of 36 individuals were

Chapter 4: Diversity related impacts on decomposition
59
inoculated in the microcosms (2 x 18 in treatments with 2 species; 3 x 12 in
treatments with 3 species). An overview of the treatments is given in table 1.
Microcosms were incubated in the dark at 20 ± 2°C.
Three replicate microcosms per treatment were destructively sampled after each of the
following incubation times: 10, 20, 30, 40, 50, 65 and 75 days. Microbial activity
(respiration rate and enzymatic activity) and decomposition parameters (weight,
carbon- and nitrogen- content) were measured at each sampling; nematode densities
and bacterial diversity were analysed on day 30, 50 and 65.
Table 1: Overview of the treatments
Decomposition rate
Percentage C and N in the cordgrass leaves were determined with a Carlo-Erba
element analyser type NA-1500 and converted to absolute amounts; C- and N-
contents of the detritus were expressed in mg. Similar patterns were obtained when
decomposition parameters are expressed as percentages remaining of the initial values
(cfr. De Mesel et al., 2003), however using absolute weight units instead of
percentages, allows proper transformation in order to fulfil the requirements for the
statistical analysis (see below).
Treatment code Nematode species
A Diplolaimelloides meyli
B Diplolaimelloides oschei
C Panagrolaimus paetzoldi
D Diplolaimella dievengatensis
E D. meyli, D. oschei
G D. meyli, D. dievengatensis
I D. oschei, D. dievengatensis
J P. paetzoldi, D. dievengatensis
L D. meyli, D. oschei, D. dievengatensis
N D. meyli, P. paetzoldi, D. dievengatensis
S (no nematodes)

Chapter 4: Diversity related impacts on decomposition
60
Nematodes
Nematodes were extracted from the sediment by centrifugation with Ludox (Heip et
al., 1985). After staining with Rose Bengal they were counted and, in the combination
treatments, identified under a dissecting microscope.
Microbial activity
The microbial community was rinsed off the cordgrass leaves with ASW in order to
measure their activity. Respiration rate of the microbial community was measured
with a Strathkelvin 928 6-channel Dissolved Oxygen System on one millilitre sub-
samples. The EnzCheck® Protease Assay Kit E6638 for green fluorescence
(Molecular Probes) was used to measure the proteolytic activity of the microbial
community. Both protocols are described in detail in De Mesel et al. (2003).
Bacterial diversity
An assessment of bacterial diversity was made based on DGGE (Denaturing Gradient
Gel Electrophoresis) banding patterns of the V3 region of the 16S rDNA genes. The
protocols used for DNA-extraction and -purification, amplification of the fragments
and the DGGE analysis have been described in detail in De Mesel et al. (2004). The
banding patterns were analysed with the software package Bionumerics 5.1 (Applied
Maths BVBA, Kortrijk, Belgium). An optical density profile through each lane
(corresponding to a single sample) identifies band positions, and calculates the
percent contribution of the intensity of each band to the total intensity of the lane. In
analogy with the results presented in De Mesel et al. (2004), bacterial diversity is
expressed as the Shannon Wiener Index (H’), based on the relative band intensities
(Magurran, 1988):
H’=∑=
n
iii pp
1ln
n represents the number of bands in the sample and pi the relative intensity of the ith
band.

Chapter 4: Diversity related impacts on decomposition
61
Statistical analysis
To evaluate the importance of nematode diversity on the decomposition rate (N- and
C-content and weight), microbial activity (respiration rate), and bacterial diversity, a
nested analysis of variance (ANOVA), in which treatments were nested within
diversity levels (i.e. level 0, 1, 2, 3 referring to 0, 1, 2, 3 nematode species
respectively), was performed by means of the GLM module in the Statistica software
package. The time factor was also integrated into the analysis. The output of the
nested ANOVA provided information on the effect of diversity, on the effect of time
and indicated if differences were found between treatments nested within a diversity
level. Due to the unbalanced design of the experiment, the combined effect ‘diversity
x time’ could not be tested with the Statistica software. When ANOVA indicated
significant differences, the Tukey HSD test was used for pairwise post hoc
comparison. Weight and respiration rate were fourth root transformed, N-content
square root transformed and C-content log-transformed to meet the required
assumptions; no transformation was required for bacterial diversity. The data on
enzymatic activity did not fulfil the assumptions for nested ANOVA, even after
transformation. Due to the unbalanced design of the experiment, no non-parametric
alternative could be applied for a similar analysis.
Species- or combination-specific effects were tested by means of a two-way ANOVA
in which the effect treatment, time and the combined effect of treatment and time was
tested. When the overall test was significant, Tukey HSD post hoc test was used for
pairwise comparison. C-content, weight, and respiration rate were log transformed
and proteolytic activity square root transformed to meet the required assumption. A
transformation of N-content was not necessary.
While counting nematodes, it became obvious that monhysterid densities reached in
combination treatments were lower than in monospecific treatments (see results).
Whether these differences were significant was tested by means of two-way ANOVA,
including the effects time and treatment in the analysis. This test was run on total
nematode densities and species-specific densities in treatments containing exclusively
monhysterid species. Again, Tukey HSD post hoc test was used for pairwise
comparison. Densities were log-transformed to meet the required assumptions.

Chapter 4: Diversity related impacts on decomposition
62
Results Nematodes
Nematode densities in all treatments with Panagrolaimus paetzoldi were 10 to 103
times higher than in other treatments (fig. 1). In the combination treatments this was
solely due to the high numbers of P. paetzoldi; the monhysterids made up only a
minor part of the total nematode numbers. In general, high variation between
replicates was present.
Nematode densities clearly differed between treatments containing only monhysterid
nematode species. Due to the considerably higher densities of P. paetzoldi, these
differences were not revealed in any statistical test when all treatments were included.
Therefore, a two-way ANOVA on species-specific densities in treatments containing
only monhysterid nematodes was carried out (fig. 2). It showed that for both D. oschei
and D. dievengatensis densities were significantly higher in the monospecific
treatments compared to the combination treatments (for all pairwise comparisons
p<0.01); within combination treatments, densities of both species were similar. For D.
1 nematode species
1
10
100
1000
10000
100000
30 50 65
ABCD
2 nematode species
1
10
100
1000
10000
100000
30 50 65
EGIJ
3 nematode species
1
10
100
1000
10000
100000
30 50 65days
LN
daysdays
1 nematode species
1
10
100
1000
10000
100000
30 50 65
ABCD
ABCD
ABCD
2 nematode species
1
10
100
1000
10000
100000
30 50 65
EGIJ
EGIJ
EGIJ
3 nematode species
1
10
100
1000
10000
100000
30 50 65days
LNLN
daysdays
Figure 1: Total nematode densities per microcosm in the monospecific treatments and the combination treatments (log scale) (average of 3 replicates ± 1 standard error)

Chapter 4: Diversity related impacts on decomposition
63
Figure 2: Nematode densities per microcosm in treatments containing exclusively
monhysterid species (average of 3 replicates ± 1 standard error)
meyli, densities in the monospecific treatments were only significantly higher
compared to the 3-species combination (p<0.01). Also the total numbers of nematodes
were generally lower in the combination treatments, but differences with the
monospecific treatments were not always significant due to high standard errors (table
2).
Table 2: Results of the Tukey HSD post hoc test
following two-way ANOVA for total nematode densities
in treatments containing exclusively monhysterid species
(ns: not significant; *:p<0.05; **:p<0.01)
Microbial activity
Respiration rate
For respiration rate, the Tukey HSD post hoc test following nested ANOVA (table 3)
showed that the difference between the control treatment (diversity level = 0) and all
dversity levels was significant (p<0.05; p<0.01; p<0.05) (fig. 3A). Within diversity
A B D E G I L A * ns ns ns ns ns B * ns ** ** ns ns D ns ns * ns ns ns E ns ** * ns ns ns G ns ** ns ns ns ns I ns ns ns ns ns ns L ns ns ns ns ns ns
D. meyliD. oscheiD. dievengatensis
0
200
400
600
800
1000
1200
A B D E G I L30 days 50 days 65 days
A B D E G I L A B D E G I L
Num
ber o
f nem
atod
esD. meyliD. oscheiD. dievengatensis
0
200
400
600
800
1000
1200
A B D E G I L30 days 50 days 65 days
A B D E G I L A B D E G I L
D. meyliD. oscheiD. dievengatensis
D. meyliD. oscheiD. dievengatensis
0
200
400
600
800
1000
1200
0
200
400
600
800
1000
1200
A B D E G I LA B D E G I L30 days 50 days 65 days
A B D E G I LA B D E G I L A B D E G I LA B D E G I L
Num
ber o
f nem
atod
es

Chapter 4: Diversity related impacts on decomposition
64
A
B
Figure 3: (A) Average respiration rate (± standard error) per diversity level at each sampling
occasion; (B) Average respiration rate (± standard error) per treatment over the whole
experiment
levels, for the monospecific treatments a significant difference between treatments A
and C (p<0.01), and for the 2-species combinations between treatment G and I
(p<0.05) were found. Only the significant differences between the control and the
two-species level were largely confirmed (significant differences for 3 combinations
out of 4) by Tukey HSD test (table 5) following two-way ANOVA (table 4).
DF MS F p-level
Diversity 3 0.016483127 5.863176 ***
Treat(Div)a 7 0.012415832 4.416408 ***
Time 6 0.894397227 318.144 ***
Error 214 0.002811297
Table 3: Results of the nested ANOVA for respiration rate (***:p<0.001)
(aTreatment nested within diversity)
Days
0
0.2
0.4
0.6
0.8
1
10 20 30 40 50 65 75
Res
pira
tion
rate
(µm
ol O
2h-1
)
0 species1 species2 species3 species
Days
0
0.2
0.4
0.6
0.8
1
10 20 30 40 50 65 7510 20 30 40 50 65 75
Res
pira
tion
rate
(µm
ol O
2h-1
)
0 species1 species2 species3 species
0.1
0.2
0.3
0.4
S A B C D E G I J L N
Res
pira
tion
rate
(µm
ol O
2h-1
)
0.1
0.2
0.3
0.4
S A B C D E G I J L NS A B C D E G I J L N
Res
pira
tion
rate
(µm
ol O
2h-1
)

Chapter 4: Diversity related impacts on decomposition
65
Figure 3B gives an indication on the overall respiration rate in each treatment over the
whole experiment. Details on temporal changes per treatment are represented in the
appendix.
DF Effect DF error F p –level
Time 6 154 428.8421 ***
Treatment 10 154 7.632465 ***
Time x Treatment 60 154 2.828222 ***
Table 4: Results of the two-way ANOVA for respiration rate (***:p<0.001)
Table 5: Results of the Tukey HSD post hoc test following the two-way
ANOVA for respiration rate (ns: not significant; *:p<0.05; **:p<0.01;
***:p<0.001)
Proteolytic activity
Within the nematode treatments, the overall trend is a slight increase in enzymatic
activity with increasing nematode diversity, while the activity in the control is situated
in between the other treatments (fig. 4A). Nested ANOVA, to test for the significance
Table 6: Results of the two-way ANOVA for proteolytic activity
(***:p<0.001)
A B C D S E G I J L N A ns *** ns ns ns 1 *** *** *** ns B ns ns ns ns ns ns ns ns ns ns C *** ns ns *** ns ** ns ns ns * D ns ns ns ns ns ns * ns ns ns S ns ns *** ns ** ns *** *** *** ns E ns ns ns ns ** ns ns ns ns ns G ns ns *** ns ns ns *** ns ns ns I *** ns ns * *** ns *** ns ns ** J ** ns ns ns *** ns ns ns ns ns L ** ns ns ns *** ns ns ns ns ns N ns ns * ns ns ns ns ** ns ns
DF Effect DF Error F p-level
Time 5 132 22.63869 ***
Treatment 10 132 12.27109 ***
Time x Treatment 50 132 4.66061 ***

Chapter 4: Diversity related impacts on decomposition
66
of the differences at the diversity level, could not be run as the assumptions were not
fulfilled.
Within each diversity level, enzymatic activity is reduced when P. paetzoldi is present
(treatments C, J and N)(fig. 4B). This is largely confirmed by the post hoc test (table
7) following two-way ANOVA (table 6).
In the appendix, a detailed representation of the proteolytic activity in each treatment
throughout the experiment is given.
A
B
Figure 4: (A) Average proteolytic activity (± standard error) per diversity level at each
sampling occasion; (B) Average proteolytic activity (± standard error) per treatment over the
whole experiment
0
5000
10000
15000
20000
25000
30000
Days
Enzy
mat
ic a
ctiv
ity (c
ount
s min
-1)
10 20 30 40 50 65 750
5000
10000
15000
20000
25000
30000
Days
Enzy
mat
ic a
ctiv
ity (c
ount
s min
-1)
10 20 30 40 50 65 75
5000
70009000
11000
1300015000
1700019000
S A B C D E G I J L NEnzy
mat
ic a
ctiv
ity (c
ount
s min
-1)
5000
70009000
11000
1300015000
1700019000
5000
70009000
11000
1300015000
1700019000
S A B C D E G I J L NS A B C D E G I J L NEnzy
mat
ic a
ctiv
ity (c
ount
s min
-1)
0 species1 species2 species3 species
0 species1 species2 species3 species

Chapter 4: Diversity related impacts on decomposition
67
A B C D S E G I J L N A ns *** ns ns ns ns ** ns * ns B ns *** ns ns ns ns ns * ns ns C *** *** *** *** *** *** *** ns *** *** D ns ns *** ns ns ns ns ** ns ns S ns ns *** ns ns ns ns *** ns ns E 1 ns *** ns ns ns * ns * ns G ns ns *** ns ns ns ns ns ns ns I ** ns *** ns ns * ns *** ns ** J ns * ns ** *** ns ns *** *** ns L * ns *** ns ns ns ns ns *** * N ns ns *** ns ns ns ns ** ns *
Table 7: Results of the Tukey HSD post hoc test following the two-
way ANOVA for proteolytic activity (ns: not significant; *:p<0.05;
**:p<0.01; ***:p<0.001)
Bacterial diversity
A
B
Figure 5: (A) Average bacterial diversity (± standard error) per nematode diversity level at
each sampling occasion; (B) Average bacterial diversity (± standard error) in each treatment
over the whole experiment
1,5
1,7
1,9
2,1
2,3
2,5
2,7
2,9
30 50 65days
Shan
non-
Wie
ner I
ndex
(H’)
1,5
1,7
1,9
2,1
2,3
2,5
2,7
2,9
1,5
1,7
1,9
2,1
2,3
2,5
2,7
2,9
30 50 65days
Shan
non-
Wie
ner I
ndex
(H’)
1,5
1,7
1,9
2,12,3
2,5
2,7
2,9
S A B C D E I J L N
Shan
non-
Wie
ner I
ndex
(H’)
1,5
1,7
1,9
2,12,3
2,5
2,7
2,9
1,5
1,7
1,9
2,12,3
2,5
2,7
2,9
S A B C D E I J L NS A B C D E I J L N
Shan
non-
Wie
ner I
ndex
(H’)
0 species1 species2 species3 species
0 species1 species2 species3 species

Chapter 4: Diversity related impacts on decomposition
68
A generally lower bacterial diversity in the combination treatments than in the
monospecific treatments and, to a lesser extent, the control was found (fig. 5A).
Tukey HSD post-hoc test following nested ANOVA indicated only the differences
between diversity level 1 and the 2- and 3- species combinations as statistically
significant (respectively p<0.01 and p<0.001). Although the overall nested ANOVA
showed a significant difference between treatments within diversity levels (table 8),
the Tukey HSD post hoc test did not reveal any pairwise differences. Within diversity
levels, however, the treatments containing P. paetzoldi (treatments C, J and N)
generally have a lower bacterial diversity (fig. 5B). For the 3-species combination,
and to a lesser extent the 2-species combination, the lower bacterial diversity in
presence of P. paetzoldi compared to other treatments is confirmed by the post hoc
comparison following two-way ANOVA (tables 9 and 10). For details on bacterial
diversity within each treatment and an overview of the DGGE-banding patterns, see
appendix. Information on bacterial diversity in treatment G (combination treatment
with D. meyli and D. dievengatensis) is not available.
DF MS F p
Diversity 3 0.716504474 10.31198 ***
Treat(Div)a 6 0.32048179 4.612395 ***
Time 2 0.217112495 3.124697 ns
Error 70 0.069482731
Table 8: Results of the nested ANOVA for bacterial diversity (ns: not
significant; ***:p<0.001) (a Treatment nested within diversity)
DF Effect DF error F p-level
Time 2 60 4.537769 *
Treatment 9 60 10.63394 ***
Time x Treatment 18 60 2.598584 **
Table 9: Results of the two-way ANOVA for bacterial diversity (*:p<0.05;
**:p<0.01; ***:p<0.001)

Chapter 4: Diversity related impacts on decomposition
69
Table 10: Results of the Tukey HSD post hoc test following the two-
way ANOVA for bacterial diversity (ns: not significant; *:p<0.05;
**:p<0.01; ***:p<0.001)
Decomposition rate
C-content
A
B
Figure 6: (A) Average C-content (mg) (± standard error) in each treatment over the whole
experiment; (B) Average C-content (mg) (± standard error) per diversity level
A B C D S E I J L N A ns ns ns ns * ns *** ns *** B ns ns ns ** ** ns *** ns *** C ns ns ns ns ns ns ns ns ** D ns ns ns ** ns ns ** ns *** S ns ** ns ** ns ns ** ns *** E * ** ns ns ns ns ns ns ns I ns ns ns ns ns ns ns ns ** J *** *** ns ** ** ns ns ns ns L ns ns ns ns ns ns ns ns ** N *** *** * *** *** ns ** ns **
100
150
200
250
300
350
10 20 30 40 50 65 75100
150
200
250
300
350
100
150
200
250
300
350
10 20 30 40 50 65 7510 20 30 40 50 65 75
150160170180190200210220230
S A B C D E G I J L N
mg
C
150160170180190200210220230
150160170180190200210220230
S A B C D E G I J L NS A B C D E G I J L N
mg
C
0 species1 species2 species3 species
0 species1 species2 species3 species
days
mg
C

Chapter 4: Diversity related impacts on decomposition
70
Figure 6A shows the average C-content in each treatment over the whole experiment.
A slower decomposition process will be represented by a higher average C-content.
An overview of the temporal C-dynamics in each treatment is given in the appendix.
A generally slower C-loss in the treatments with the 2-species combinations
(treatments E, G, I, J) was seen (fig. 6A and 6B). According to the post hoc test
following nested ANOVA (overall test in table 11), the difference between the 2-
species level and the other diversity levels were significant (all p< 0.001); also a low
significant difference between diversity level 0 (controls) and the 1-species diversity
level was found (p<0.05). Only the differences between the 2-species combinations
and the control were clearly confirmed by the two-way ANOVA (table 12 and 13).
The results of weight are very similar to those for C-content (see also De Mesel et al.,
2003) and are therefore not presented here.
Table 11: Results of the nested ANOVA for C-content (*:p<0.05;
***:p<0.001) (a Treatment nested within diversity)
DF Effect DF Error F p-level
Time 6 154 204.0253 ***
Treatment 10 154 8.360312 ***
Time x treatment 60 154 1.501799 *
Table 12: Results of the two-way ANOVA for C-content (*:p<0.05;
***:p<0.001)
DF MS F p
Diversity 3 0.054855677 19.30672 ***
Treat(Div)a 7 0.006239223 2.195925 *
Time 6 0.508193444 178.8611 ***
Error 214 0.002841274

Chapter 4: Diversity related impacts on decomposition
71
Table 13: Results of the Tukey HSD post hoc test following two-way
ANOVA for C-content (ns: not significant; *:p<0.05; ***:p<0.001)
N-content
As for C-content, higher average N-content in figure 7A agrees with slower
decomposition rates. Details on N-content per treatment are represented in the
appendix.
Differences for N-content between the control treatment and nematode treatments
were found to be significant by both nested ANOVA (significance level with diversity
DF MS F p-level
Diversity 3 13.91021609 5.929644 ***
Treat(Div)a 7 21.03011271 8.964712 ***
Time 6 96.11691385 40.9727 ***
Error 214 2.345877222
Table 14: Results of the nested ANOVA for N-content (***:p<0.001)
(aTreatment nested within diversity)
DF Effect DF Error F p-level
Time 6 154 77.37736 ***
Treatment 10 154 15.21042 ***
Time x Treatment 60 154 4.16902 ***
Table 15: Results of the two-way ANOVA for N-content (***:p<0.001)
A B C D S E G I J L N A ns ns ns ns ns * * ns ns ns B ns ns ns ns ns ns ns ns ns ns C ns ns ns ns ns ns ns ns ns ns D ns ns ns *** ns ns ns ns ns *** S ns ns ns *** *** *** *** *** * ns E ns ns ns ns *** ns ns ns ns *** G * ns ns ns *** ns ns ns ns *** I * ns ns ns *** ns ns ns ns *** J ns ns ns ns *** ns ns ns ns *** L ns ns ns ns * ns ns ns ns * N ns ns ns *** ns *** *** *** *** *

Chapter 4: Diversity related impacts on decomposition
72
A
B
Figure 7: (A) Average N-content (mg) (± standard error) in each treatment over the whole
experiment; (B) Average N-content (mg) (± standard error) per diversity level
Table 16: Results of the Tukey HSD post hoc test following two-way ANOVA
for N-content (ns: not significant; *:p<0.05; **:p<0.01; ***:p<0.001)
A B C D S E G I J L N A ns ns *** ** ns ns ns ns * ns B ns ns *** ns ns ns *** *** *** ns C ns ns *** ns ns ns ns ns *** ns D *** *** *** *** *** *** * * ns *** S ** ns ns *** ns ns *** *** *** ns E ns ns ns *** ns ns ns ns ** ns G ns ns ns *** ns ns ** ** *** ns I ns *** ns * *** ns ** ns ns ns J ns *** ns * *** ns ** ns ns ns L * *** *** ns *** ** *** ns ns *** N ns ns ns *** ns ns ns ns ns ***
0
2
4
6
8
10
12
10 20 30 40 50 65 75
mg
N
0
2
4
6
8
10
12
0
2
4
6
8
10
12
10 20 30 40 50 65 7510 20 30 40 50 65 75
mg
N
10
20
30
40
50
S A B C D E G I J L N
mg
N
10
20
30
40
50
10
20
30
40
50
S A B C D E G I J L NS A B C D E G I J L N
mg
N
0 species1 species2 species3 species
0 species1 species2 species3 species

Chapter 4: Diversity related impacts on decomposition
73
levels respectively p<0.05; p<0.05 and p<0.001) and two-way ANOVA (table 15 and
16) (fig. 7B). The significant treatment effect pointed out by nested ANOVA (table
14) was due to significant differences between monospecific treatments D and,
respectively, A (p<0.01), B (p<0.001) and C (p<0.01), and between 3-species
treatments L and N (p<0.05). These significant differences were confirmed by two-
way ANOVA (table 16).
Discussion Development of nematode populations
All four nematode species used in this experiment belong to the deposit feeders
(Wieser, 1953) and are assumed to feed mainly on bacteria (Moens and Vincx, 1997).
Despite their similar functional position, some ecological differences between the
Panagrolaimidae and Monhysteridae have been described. Panagrolaimidae are
considered extreme colonizers and enrichment opportunist, quickly colonizing
organically enriched sites with high microbial activity and disappearing as microbial
activity decreases (Bongers and Bongers, 1998). Monhysteridae also occur in sites
with high resource availability, however they can persist under food-poor conditions;
as such they belong to the general opportunists (Bongers et al., 1995). Their
generation time is slightly longer in comparison with Panagrolaimidae. Due to these
differences, P. paetzoldi was expected to reach peak densities at the start of the
experiment, while the monhysterids were thought to have a slower increase in
numbers, but to persist throughout the experiment. This was however not confirmed
by our observations: densities of P. paetzoldi were much higher during the course of
the experiment, both in the monospecific treatments and in the combination
treatments. Another remarkable observation was that monhysterid densities were
considerably lower in the combination treatments compared to the monospecific
treatments. One reason could have been that the densities of individual species in the
initial inoculum of the combination treatments were lower than in the monospecific
treatments: 36 individuals in the monospecific treatments, compared to 18 and 12 in,
respectively, the 2- and 3- species combinations. This might have caused some delay
in the population development of each species; however this discrepancy should
vanish over subsequent generations, as was the case for P. paetzoldi. Moreover, an

Chapter 4: Diversity related impacts on decomposition
74
additional experiment, with larger nematode inocula, confirmed inhibitory
interactions between two of the monhysterids used here, D. meyli and D.
dievengatensis (pers. observation, unpublished). In a similar experimental set-up as
used in this study, a number of petri dishes were inoculated with either D. meyli or D.
dievengatensis, or stayed without nematodes (controls). After an incubation of 7 days,
half of the petri dishes with D. meyli or D. dievengatensis were inoculated with the
other species, the other half were further incubated without manipulation.
Simultaneously, half of the controls received D. meyli, the other half D.
dievengatensis. Two weeks later, nematode densities were counted and compared. A
lower number of either species was consistently found in the combination treatments
when compared to single-species treatments, with the differences mostly being
significant.
Similar observations have been reported by Ilieva-Makulec (2001), who studied the
life strategy of two terrestrial bacteria-feeding nematodes, Acrobeloides nanus and
Dolichorhabditis dolichura. They explained this mutual inhibition by food shortage in
the combination treatments. Nematode densities in the monospecific treatments show,
however, that, even when all species require the same bacterial diet, higher densities
can be sustained with the resources available. Additionally, a food-selection
experiment (Moens et al., 1999) and analysis of the bacterial community in the
monospecific treatments (De Mesel et al., 2004) indicated different feeding
preferences between species, which should allow them to share the food sources. We
also found similar aerobic activity and even higher proteolytic activity in the
combination treatments compared with the monospecific treatments, which does not
at all indicate a suppression of the bacterial community.
Information on inhibition within benthic fauna is in general very scarce, while
facilitative interactions between species have been reported more often (e.g. David,
1996; Levin et al., 2001; Cardinale et al., 2002). Chandler and Fleeger (1987) were
the first to study interactions within meiofauna. They conducted experiments with
copepods in estuarine sediments and found both inhibition and facilitation between
species. They suggest that the observed inhibitory interactions were due to mucus
trails secreted by one of the copepod species, which not only altered sediment
structure, but might also have evoked a chemical repulsion. Many free living marine
nematodes also secrete mucus tracks (Riemann and Schrage, 1978, Riemann and
Helmke, 2002). It seems unlikely that any structural modification of the sediment

Chapter 4: Diversity related impacts on decomposition
75
could have caused the mutual suppression in population development because (i)
species-specific densities would probably be too low to affect the sediment structure
of the total microcosm and (ii) one would expect one species (the ‘modifier’) to
dominate the system and suppress the other species. This was obviously not the case
in our study: a clear mutual inhibition was observed. The idea that some kind of
chemical interference between species occurred can not be refuted. Chemical cues
could emanate from the mucus tracks, or from other glandular secretions. Nematodes
are susceptible to chemical signals (Huettel, 1986) since they are important in the
reproduction (Bone, 1982) or for tracing food sources (Moens et al., 1999). Up till
now, patchy food distribution (Trotter and Webster, 1984; Moens et al., 1999) was
assumed to be the main cause of the heterogeneous nematode distribution in marine
sediments, but chemical interaction might enhance this phenomenon. Accumulation of
chemicals in the closed system in this experiment could have prevented growth of
nematodes in the whole microcosm.
Effect of nematodes on microbial activity and bacterial diversity
The activity of the microbial community was measured by the respiration rate and the
proteolytic activity. The highest respiration rate was observed in the control. This was
statistically confirmed by analysis at both diversity and treatment level. Respiration
was assumed to mainly represent fungal activity (Padgett et al., 1985; De Mesel et al.,
2003). However, some interference of bacterial activity in the respiration
measurements will have occurred, especially at the start of the experiment. Still, the
high respiration rates in the control coincide with observations of stronger growth of
fungi. In terrestrial systems an inhibition of fungal growth in the presence of
nematodes has been described, either as a consequence of direct effects or through
interactions with bacteria (Kaufman, 1983; Grewal, 1991; Grewal and Richardson,
1991; Grewal and Hand, 1992). Similar interactions in marine environments have not
yet been studied; however we believe they might have occurred in our microcosms.
Nematodes might have facilitated bacterial communities, for instance through
secretion of mucus trails (Riemann and Schrage, 1978) or any other metabolites.
Antagonistic interactions between bacteria and fungi during decomposition of aquatic
plant litter have been described (Wohl and McArthur, 2001); with the reverse effect of

Chapter 4: Diversity related impacts on decomposition
76
bacteria on fungi being more pronounced (Mille-Lindblom and Tranvik, 2003).
Facilitation of the bacterial community in the nematode treatments might thus have
antagonized the development of fungi. Fungi are important in the first stages of
cordgrass decay and their suppression could have slowed down the decomposition
process (see below).
Proteolytic activity would mainly represent anaerobic metabolism, since it increases
while respiration rate decreases. Aquatic fungi are generally aerobic (Rheinheimer,
1992), suggesting that proteolytic activity provides information on anaerobic bacterial
activity; however some interference of fungi can not be excluded. Proteolytic activity
was considerably lower in the presence of P. paetzoldi than in the other treatments
within the same diversity level. Their densities were much higher than those of the
monhysterids, so this reduction can be ascribed to overgrazing of the bacterial
community. The same applies to bacterial diversity: the high grazing pressure in the
presence of P. paetzoldi reduced diversity within the bacterial community (see also
De Mesel et al., 2004). While, in general, a positive trend in the relationship between
number of nematode species and enzymatic activity was seen, the link with bacterial
diversity was negative. Lack of correlation between bacterial diversity and bacterial
activity has been reported before (Troussellier et al., 2002).
Impacts on the decomposition rates
Despite the low nematode densities in the absence of P. paetzoldi, and the dominance
of the latter species when present, we did find a diversity-related impact on the C-loss
of the cordgrass detritus. As explained before (De Mesel et al., 2003), in the
monospecific treatments a slower decomposition rate was observed compared to the
control. An even more severe suppression of C-loss occurred when two nematode
species were pooled, but at the 3-species diversity level C-loss was again comparable
to the monospecific treatments, or even higher as for the treatment with the
combination P. paetzoldi, D. dievengatensis and D. meyli.
Nitrogen content of the cordgrass leaves was generally lower in the control treatment.
This could be due to the presence of fungi in the controls (see above), which are
important during the early stages of decomposition. On the other hand, nematodes
consume nitrogen in excess when feeding on bacteria (Anderson et al., 1983; Ferris et

Chapter 4: Diversity related impacts on decomposition
77
al., 1997). After excretion, this nitrogen can either be used by the bacteria, or can bind
to the detritus (White and Howes, 1994; Valiela, 1995), masking any effect of
microbial activity on the removal of nitrogen. Significant differences in nitrogen
content of the cordgrass leaves between nematode treatments were found, however no
link with diversity nor with specific species or species combinations can be made. For
instance, when D. meyli and D. dievengatensis were applied separately to the
microcosms, N-content was significantly higher than in the control treatment, but
when these species were pooled, the difference with the control was less pronounced;
adding the third monhysterid (D. oschei) again yielded significant difference with the
control.
No link between bacterial diversity and decomposition rates was found. According to
O’Donnell et al. (2001), the central problem in microbial ecology is to understand the
link between genetic diversity and community structure, and between community
structure and function. Studies on functional implications of altering microbial
diversity in soil systems is still scarce, however, a degree of redundancy is expected
(Griffiths et al., 2000; Nannipieri et al., 2003).
Within treatments a positive correlation between respiration rate and both C-content
and, to a lesser extent, N-content was found; links with proteolytic activity were less
clear. This indicates that fungi would largely depend on the cordgrass substrate, and
thus would be mainly responsible for the decomposition of the detritus. This is not
unusual, since fungi can penetrate solid substrates and digest them from within
(Newell, 1996); bacteria on the other hand might have a better ability to utilize
dissolved organic compounds (Wohl and McArthur, 2001; Mille-Lindblom and
Tranvik, 2003). These organic compounds leached into the sediment at the start of the
experiment (Valiela, 1995). This does however not exclude that bacteria might have
impacted the decomposition rate to some extent, but their effect could be obscured by
the fungi.
Differences in respiration rates between treatments did not necessarily result in
(predictable) differences in elemental losses; the same goes for proteolytic activity.
Riemann and Helmke (2002) suggested that nematodes and bacteria growing on their
mucus tracks, ‘share’ enzymes to break down organic matter. This way, nematodes
could directly assimilate organic compounds from the detritus. On the other hand,
differences in C-use efficiencies between bacterial communities might also have
affected our results (Degens, 1998 and references therein).

Chapter 4: Diversity related impacts on decomposition
78
Conclusions
The main aim of this paper was to evaluate the importance of nematode diversity or
nematode species in affecting the microbial activity and the decomposition rate.
Our data did not support any hypothesis predicting enhancement of process rates with
an (initial) increase in species-numbers (e.g. rivet hypothesis, optimum hypothesis,
modified null hypothesis). A key mechanism in enhancing ecosystem function would
be interspecific facilitation (Loreau and Hector, 2001; Mulder et al., 2001; Fridley,
2001; Cardinale et al., 2002). Facilitation was not at all observed in our experiments;
instead we found clear evidence of inhibitory interaction between the monhysterid
species. Redundancy among the monhysterids seems unlikely as it was not
consistently confirmed for all parameters. N-loss for instance, was very different
between monhysterid treatments. Differences between the monhysterids and P.
paetzoldi were obvious. The extremely high densities of P. paetzoldi resulted in a
reduction in bacterial activity and diversity; however this was not translated into a
slower decomposition rate. Also within the treatments exclusively containing
monhysterid nematode species no clear link between microbial activity and
decomposition rate was found. The lack of completely separate measurements of
bacterial and fungal activity might have complicated the establishment of any such
link. Additionally, different C-use efficiencies of the bacterial communities, and the
possibility that (some) nematode species could directly use organic detrital
compounds, should be considered.
Our results rather support the idiosyncratic diversity model, which describes an
unpredictable effect of changing species composition or species diversity on
ecosystem processes. However, in concordance with Mikola and Setälä (1998), who
also found an idiosyncratic response of microbivore nematode diversity in a terrestrial
soil system, we should note that our experiments only contained a small number of
species. It is possible that idiosyncratic effects are mainly visible in low diversity
systems. Still, differences between bacterivore nematode species and species
combinations could not be predicted at the start of the experiment, indicating that no a
priori redundancy within the bacterivores can be expected. Additionally, several other
experimental studies, using other benthic systems, confirm this idiosyncratic theory
(e.g. Emmerson et al., 2001; Downing and Leibold, 2002).

Chapter 4: Diversity related impacts on decomposition
79
Acknowledgements
Annick Van Kenhove is acknowledged for the chemical analyses and Prof. Dr.
Jacques Vanfleteren for the use of the Victor Multilabel counter. We thank Prof. Dr.
Ann Vanreusel, Dr. Peter Herman and Dr. Marleen De Troch for the valuable
comments on the manuscript. The first and second author acknowledge grants from
the Flemish Institute for the Promotion of Scientific-Technological Research (I.W.T.).
The last author is a postdoctoral fellow with the Flemish Fund for Scientific Research.
Further financial support was obtained from Ghent University in BOF- projects
1205398 and 011060002.

Submitted for publication as:
De Mesel, I, Vanaverbeke, J, Vincx, M, Moens, T (submitted) Development of a
nematode community on decomposing organic matter (Spartina anglica) under
contrasting conditions.
Chapter 5
Development of a nematode community on decomposing organic matter (Spartina anglica) under contrasting
conditions

Chapter 5: Colonization of cordgrass detritus under contrasting conditions
83
Abstract
Cordgrass detritus has often been described as a microhabitat sustaining high densities
of nematodes. We performed litterbag experiments in which the development of the
nematode community on decaying cordgrass detritus was studied under stable
conditions in the laboratory and in the field, where incubations were subject to
environmental fluctuations, mainly associated with the tidal currents.
Nematodes showed a clear preference for the detritus under controlled conditions in
the lab, and the nematode assemblage on the cordgrass differed considerably from the
surrounding sediments. By contrast, in the field nematodes were rather distributed at
random, probably through passive transport with the tides, showing that active habitat
colonization was of minor importance in the hydrodynamically impacted system.
We assumed that the contrasting incubation conditions in the lab and the field would
affect the structural and functional aspects of the nematode community. In the lab,
incubations were performed in the dark, preventing growth of microalgae, an
important food source for mainly epistrate feeders and deposit feeders. Epistrate
feeders were however equally abundant in the field and in the lab, indicating that
other food sources, probably bacteria, can be consumed. Deposit feeders, dominating
field samples, were mainly replaced by microvores in the lab; still some species were
able to adapt to a principally bacterial diet. Nematode diversity was not affected by
resource availability, nor by the daily hydrodynamical disturbance.

Chapter 5: Colonization of cordgrass detritus under contrasting conditions
85
Introduction
Marine sediments are generally characterized by high densities of meiofauna. In both
subtidal and intertidal habitats, their distribution can be very patchy (Findlay 1981,
1982). At large scales (m-km) this patchiness is due to gradients in physical factors,
for example salinity, tidal exposure, sediment composition (Findlay, 1981; Coull,
1988). The small scale heterogeneous distribution would mainly be caused by biotic
interactions such as competition, either inter- or intraspecific (Heip, 1975, 1976;
Service and Bell, 1987), facilitation (Heip and Engels, 1977; Fleeger and Gee, 1986;
Chandler and Fleeger, 1987), the presence of biogenic structures (Bell et al., 1978;
Sun et al., 1993; Wetzel et al., 1995; Fenchel, 1996) and the heterogeneous
distribution of their food sources (Buzas, 1969; Boucher and Chamroux, 1976). In a
laboratory experiment, Lee et al. (1977) offered a number of micro-algae to a
meiofaunal inoculum in a so called ‘cafeteria’-design. He reported a selective
attraction of different taxa towards each of the food sources. A similar set-up was
used to seek for the response of a limited number of nematode species towards a
selection of bacterial and diatomaceous food items (Trotter and Webster, 1984;
Moens et al., 1999). These experiments suggested that nematodes can perceive and
move towards their preferred food source. Differences in food availability could also
cause spatial differences and temporal dynamics in the trophic composition of the
nematode community (Danovaro and Gambi, 2002)
In the dynamic environment of intertidal salt marshes, however, small-scale spatial
distribution of meiofauna may not only be influenced by biotic interactions but also
by abiotic factors. Meiofauna generally have limited active dispersal capacities
(Fegley, 1987 and references therein) and can be passively dispersed by tidal currents
(Bell and Sherman, 1980; Sherman and Coull, 1982; Fleeger et al., 1984; Bertelsen,
1998; Powers 1998; Commito and Tita, 2002). Wave action may thus facilitate patch
colonization (Ullberg and Ólafsson, 2003), but, at the same time hamper the
establishment of ‘stable’ populations.
The present study aimed to assess the relative importance of biotic and abiotic forces
in structuring nematode communities colonizing patches of freshly deposited
cordgrass detritus. Cordgrass (Spartina spp) is often the dominant vegetation of the
lower parts of tidal marshes in temperate areas and its detritus can support high

Chapter 5: Colonization of cordgrass detritus under contrasting conditions
86
nematode densities (Reice and Stiven, 1983; Alkemade et al., 1993). These detrital
microhabitats are often characterized by a high relative abundance of characteristic
taxa, mainly Monhysteridae (Bouwman et al., 1984; Warwick 1987; Alkemade et al.,
1993). They may stimulate the decomposition rate of the organic matter (Abrams and
Mitchell, 1980; Anderson et al., 1981; Findlay and Tenore, 1982; Alkemade et al.,
1992 a, b, 1993), although this has been questioned recently (De Mesel et al., 2003).
Cordgrass detritus in litterbags was placed on salt marsh sediment both under
controlled laboratory conditions and in a field site. Samples were taken monthly
during a three-month period. We tested whether (i) detritus availability could impact
the distribution of the nematode community in an undisturbed laboratory
environment, (ii) this distribution could be affected by natural disturbances in the
field, and (iii) the contrasting environmental conditions in the field and in the lab
would alter the nematode community structure.
Materials and Methods
Experimental set up
Litterbag incubations were performed simultaneously in a salt marsh (Paulina Salt
Marsh, Westerschelde, The Netherlands) and under controlled conditions in the
laboratory. The litterbags (10 cm * 12 cm) were made of gauze with a mesh size of
125µm, allowing free movement of micro-and meiofauna. One set of bags was filled
with cordgrass detritus, the other with plastic strips which served as controls.
Green cordgrass leaves were collected in the salt marsh (1st October 2001). They were
thoroughly rinsed with fresh water and dried to the air to remove the attached fauna.
The leaves were cut in fragments of approximately 10 cm and 4.1 ± 0.1g was put in
each litterbag. The control litterbags contained plastic strips with a similar surface as
the cordgrass fragments.
In the salt marsh 4 experimental fields with a surface of approximately 4 m2 each
were selected at the edges of Spartina-patches in the middle of the Spartina
dominated zone, which was flooded twice a day. In each field, 30 litterbags, 15 of
which containing detritus and 15 controls were incubated. The litterbags were
separated approximately 30 cm from each other, preventing mutual influence.

Chapter 5: Colonization of cordgrass detritus under contrasting conditions
87
In the laboratory, plastic dishes (13 cm * 17 cm * 15 cm) were filled with sediment
originating from the Paulina Salt Marsh and incubated in the dark at 12°C for one
week allowing the system to stabilize. Afterwards, half of the dishes received a
detritus litterbag, the others control litterbags. A suspension of sediment from the salt
marsh in artificial sea water (Dietrich and Kalle, 1957) was added when the litterbags
were placed on the dishes and after every sampling in order to provide a fresh fauna
inoculum. Each time a few hundred nematodes were applied to all dishes. Our results
showed however that these inocula did not considerably affect the composition of the
nematode communities. The sediment was water saturated throughout the experiment
and a constant salinity was maintained.
Litterbags were incubated at 12°C in the dark.
Sampling
Samples were taken monthly over a 3-month period (November 2001, December
2001 and January 2002). At each sampling occasion one detritus litterbag and one
control were removed at random from each experimental field in the salt marsh,
resulting in four replicates per sampling. Simultaneously, the upper cm of the
sediment in each field was sampled with Perspex cores (∅ 3.6 cm).
In the laboratory, 4 dishes with cordgrass and 4 with controls were sampled every
month, on the same days as the field samplings. From every dish, sediment
surrounding the litterbags was collected as described above.
Nematodes were rinsed of the cordgrass leaves and plastic strips with tap water. The
rinse water and the sediment were preserved on a 4% buffered formaldehyde solution.
Cordgrass fragments were dried, weighed, an preserved at -20°C for C- and N-
analysis, which was performed with a Carlo-Erba elemental analyzer type NA-1500.
Fungal biomass in the cordgrass leaves was assessed by ergosterol measurements
(Gessner and Newell, 1997)
Nematode community
Nematodes were extracted from the sediment by centrifugation with Ludox (Heip et
al., 1985). Nematodes from the sediment, the detritus and the controls were stained

Chapter 5: Colonization of cordgrass detritus under contrasting conditions
88
with Rose Bengal and counted under a dissecting microscope. At each sampling
occasion 200 nematodes were picked randomly from two replicates from each
treatment (sediment, detritus, controls) and mounted on Cobb slides for identification
to species level. The contribution of each species to the assemblage was expressed as
a relative abundance in order to facilitate comparison of the assemblages from the
detritus, the controls (both ind./litterbag) and the sediment (ind/10cm²). For the
functional analysis of the community, nematodes were grouped in 6 feeding guilds:
microvores, deposit-feeders, ciliate feeders, epigrowth feeders, facultative predators
and predators (Moens and Vincx, 1997). This system is based on the classification of
Wieser (1953) but expanded with information obtained from observations.
Diversity was expressed with the Shannon-Wiener Index (H’) (Shannon and Weaver,
1949):
H’= ∑=
n
iii pp
1ln
n represents the number of species in the sample and pi the relative abundance of the
ith species.
Statistical analysis
Univariate two- or three-way analysis of variance (ANOVA) was used to test for
differences of nematode densities, feeding types and diversity between sites (field vs
laboratory), sampling dates, treatments (detritus vs controls vs sediment) and
combinations of these factors. Fourth root transformation on nematode densities was
performed to meet the required assumptions. When ANOVA indicated significant
differences the Tukey HSD test was used for pairwise post hoc comparison. These
tests were done with the Statistica software.
Nematode community structure was analysed by means of Multidimensional Scaling
(MDS) using the PRIMER software package. The MDS was based on the Bray-Curtis
similarity index, calculated from the relative densities. Analysis of Similarities
(ANOSIM) was applied to detect differences between sites and differences between
treatments within sites. Similarity percentage analysis (SIMPER) was used to weigh
the contribution of each species to similarity/dissimilarity between sites and
treatments.
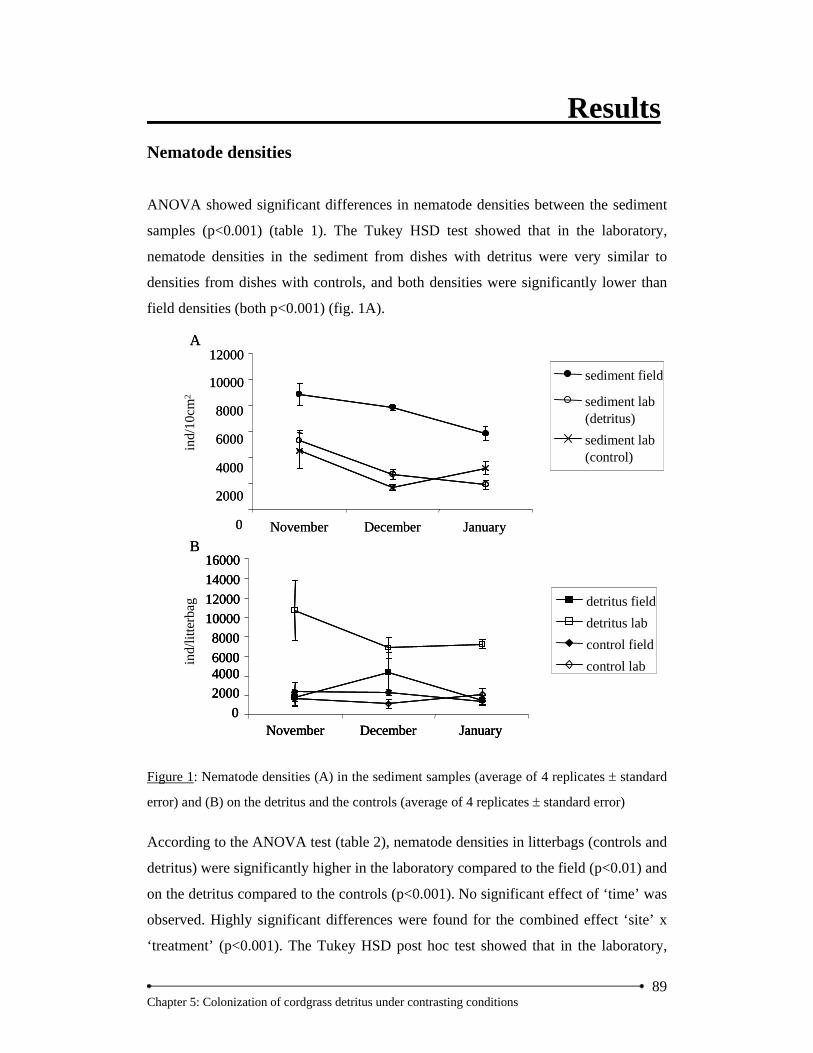
Chapter 5: Colonization of cordgrass detritus under contrasting conditions
89
Results
Nematode densities
ANOVA showed significant differences in nematode densities between the sediment
samples (p<0.001) (table 1). The Tukey HSD test showed that in the laboratory,
nematode densities in the sediment from dishes with detritus were very similar to
densities from dishes with controls, and both densities were significantly lower than
field densities (both p<0.001) (fig. 1A).
Figure 1: Nematode densities (A) in the sediment samples (average of 4 replicates ± standard
error) and (B) on the detritus and the controls (average of 4 replicates ± standard error)
According to the ANOVA test (table 2), nematode densities in litterbags (controls and
detritus) were significantly higher in the laboratory compared to the field (p<0.01) and
on the detritus compared to the controls (p<0.001). No significant effect of ‘time’ was
observed. Highly significant differences were found for the combined effect ‘site’ x
‘treatment’ (p<0.001). The Tukey HSD post hoc test showed that in the laboratory,
0
2000
4000
6000
8000
10000
12000
November December January
sediment field
sediment lab(detritus)sediment lab(control)
02000400060008000
10000120001400016000
November December January
detritus fielddetritus labcontrol fieldcontrol lab
A
B0
2000
4000
6000
8000
10000
12000
November December January
sediment field
sediment lab(detritus)sediment lab(control)
sediment field
sediment lab(detritus)sediment lab(control)
02000400060008000
10000120001400016000
November December January0
2000400060008000
10000120001400016000
November December January
detritus fielddetritus labcontrol fieldcontrol lab
detritus fielddetritus labcontrol fieldcontrol lab
A
B
ind/
10cm
2in
d/lit
terb
ag

Chapter 5: Colonization of cordgrass detritus under contrasting conditions
90
nematode densities were significantly higher on detritus compared to the controls
(p<0.001) and compared to both the detritus and the controls in the field (both
p<0.001) (fig. 1B). In the field, densities on the controls and the detritus were similar.
Table 1: Results of the two-way ANOVA for nematode densities in the
sediment (ns: not significant, ***:p<0.001) a type: sediment core from (1) the field, (2) control dishes in the lab and (3)
dishes with detritus in the lab
Table 2: Results of the three-way ANOVA for nematode densities on the detritus
and the controls (ns: not significant, **:p <0.01, ***:p<0.001) a treatment: (1) detritus and (2) control
Effect DF effect DF error F p-level
Time 2 27 12.55228 ***
Typea 2 27 40.21584 ***
Time x Type 4 27 1.96637 ns
Effect DF effect DF error F p-level
Site 1 36 10.17099 **
Time 2 36 0.12346 ns
Treatmenta 1 36 18.45973 ***
Site x Time 2 36 1.84033 ns
Site x Treatment 1 36 16.6994 ***
Time x Treatment 2 36 0.20101 ns
Time x Treatment x Site 2 36 0.32266 ns

Chapter 5: Colonization of cordgrass detritus under contrasting conditions
91
Nematode community Nematode assemblage
In the MDS plot based on the nematode species assemblages, a separation of the field
and lab samples was clear (fig. 2). ANOSIM indicated that the difference between
sites was significant (p<0.001). By means of SIMPER analysis (results not shown)
Daptonema biggi and Tripyloides marinus were identified as characteristic species for
the field samples. Leptolaimus limicolus, Anoplostoma viviparum and Axonolaimus
typicus were important members of the communities in the laboratory.
Metachromadora remanei and Ptycholaimellus ponticus were abundant at both sites.
Differences between sites were mainly determined by D. biggi and L. limicolus.
Figure 2: MDS ordination plot based on the nematode species assemblages
ANOSIM on the samples of the laboratory showed a highly significant difference
between nematode community composition on the detritus and in the sediment
(p<0.01); the difference between controls and the sediment was low significant
(p<0.05)(table 3). SIMPER analysis showed that L. limicolus, who dominated the
communities on the detritus, explained most of the variation between the detritus and
the sediment community. Ptycholaimellus ponticus was the most characteristic
species on the controls and M. remanei and A. typicus in the cores. Anoplostoma
viviparum was a prominent member of all communities in the lab (fig. 3A).
sediment field
sediment lab
detritus fieldcontrol field
detritus labcontrol lab
nov
nov
novnov
novnov
decdec
dec
dec
dec
dec
dec
jan jan
janjan
jan jan
jan
nov
nov
nov
nov
dec
janjan
decnov
dec
jandec nov
jandec
jan
Stress: 0.15detritus fieldcontrol field
detritus labcontrol lab
nov
nov
novnov
novnov
decdec
dec
dec
dec
dec
dec
jan jan
janjan
jan jan
jan
nov
nov
nov
nov
dec
janjan
decnov
dec
jandec nov
jandec
jan
Stress: 0.15
sediment field
sediment lab
detritus fieldcontrol field
detritus labcontrol lab
nov
nov
novnov
novnov
decdec
dec
dec
dec
dec
dec
jan jan
janjan
jan jan
jan
nov
nov
nov
nov
dec
janjan
decnov
dec
jandec nov
jandec
jan
Stress: 0.15detritus fieldcontrol field
detritus labcontrol lab
nov
nov
novnov
novnov
decdec
dec
dec
dec
dec
dec
jan jan
janjan
jan jan
jan
nov
nov
nov
nov
dec
janjan
decnov
dec
jandec nov
jandec
jan
Stress: 0.15

Chapter 5: Colonization of cordgrass detritus under contrasting conditions
92
A
B
Figure 3: Composition of the nematode community on detritus, controls and in the sediment.
The 5 dominant species in (A) the laboratory and (B) the field samples are represented.
For the field samples ANOSIM showed no significant differences between treatments.
All treatments were characterized by the presence of D. biggi; other species, such as
P. ponticus, M. remanei, Desmolaimus zeelandicus and Tripyloides marinus were also
characteristic for all treatments (fig 3B).
Table 3: Results of the ANOSIM routine to analyze differences in
nematode assemblages between detritus, controls and sediment in the
laboratory (ns: not significant, *:p<0.05, **:p<0.01)
R-statistic p-level
detritus vs control 0.107 ns
detritus vs sediment 0.381 **
control vs sediment 0.228 *
0%
20%
40%
60%
80%
100%
detritus control sediment
Daptonema biggiPtycholaimellus ponticus
Metachromadora remaneiTripyloides marinusDesmolaimus zeelandicusothers
0%
20%
40%
60%
80%
100%
detritus control sediment0%
20%
40%
60%
80%
100%
detritus control sediment
Daptonema biggiPtycholaimellus ponticus
Metachromadora remaneiTripyloides marinusDesmolaimus zeelandicusothers
0%
20%
40%
60%
80%
100%
detritus control sediment
Leptolaimus limicolusPtycholaimellus ponticusMetachromadora remaneiAnoplostoma viviparumAxonolaimus typicusothers
0%
20%
40%
60%
80%
100%
detritus control sediment
Leptolaimus limicolusPtycholaimellus ponticusMetachromadora remaneiAnoplostoma viviparumAxonolaimus typicusothers

Chapter 5: Colonization of cordgrass detritus under contrasting conditions
93
Functional community structure
Table 4: Results of the three-way ANOVA for relative abundances of microvores (MV),
deposit-feeders (DF), epistrate feeders (EF), ciliate feeders (CF), predators (P) and facultative
predators (FP) (ns: not significant, *:p<0.05, **:p<0.01, ***:p<0.001)
An overview of the results of the three-way ANOVA is represented in table 4. The
proportion of microvores, ciliate feeders, predators and facultative predators was
significantly higher in the lab compared to the field (p<0.001, p<0.05; p<0.05;
p<0.001 respectively), while the opposite was found for deposit feeders (p<0.001)
(fig. 4). Similar percentages of epigrowth feeders were found at both sites. No
significant differences were found between treatments within one site.
MV DF CF EF P FP
DF effect DF error
1 18
1 18
1 18
1 18
1 18
1 18
F 64,62328 32,48896 4,88594 0,15354 5,002934 34,50508 Site
p-level *** *** * ns * *** DF effect DF error
2 18
2 18
2 18
2 18
2 18
2 18
F 3,84566 6,39942 1,297919 11,21461 0,432588 0,33567 Tim
e
p-level * ** ns *** ns ns DF effect DF error
2 18
2 18
2 18
2 18
2 18
2 18
F 2,19227 1,80725 0,533884 2,09577 0,325256 1,14379
Trea
tmen
ta
p-level ns ns ns ns ns ns
DF effect DF error
2 18
2 18
2 18
2 18
2 18
2 18
F 4,19017 2,11368 0,134558 0,56834 0,618706 0,27625 Site
x
Tim
e
p-level * ns ns ns ns ns DF effect DF error
2 18
2 18
2 18
2 18
2 18
2 18
F 1,08413 6,11429 0,118275 1,93536 0,846145 0,77165 Site
x
Trea
tmen
t
p-level ns ** ns ns ns ns
DF effect DF error
4 18
4 18
4 18
4 18
4 18
4 18
F 1,14245 1,63149 2,527191 0,43745 0,081061 1,76394 Tim
e x
Trea
tmen
t
p-level ns ns ns ns ns ns DF effect DF error
4 18
4 18
4 18
4 18
4 18
4 18
F 0,85902 1,0576 1,243217 1,3911 0,610571 1,51127 Site
x
Tim
e x
Tr
eatm
ent
p-level ns ns ns ns ns ns

Chapter 5: Colonization of cordgrass detritus under contrasting conditions
94
Figure 4: Trophic community composition in the lab and in the field. MV: microvores, DF:
deposit feeders, EF: epistrate feeders, CF: ciliate feeders, P: predators, FP: facultative
predators
Diversity
In the laboratory, a total of 49 species were counted; in the field 53. Diversity was
rather similar in the field and in the lab, as shown by the Shannon-Wiener Index (fig.
5). ANOVA indicated significant differences for the combined effect ‘time’ x
‘treatment’ (table 5), but the Tukey HSD post hoc test did not reveal any significant
pairwise differences.
Table 5: Results of the three-way ANOVA for the Shannon-Wiener Indices (ns: not
significant, *: p < 0.05). a Treatment: (1) detritus, (2) controls and (3) sediment
DF effect DF error F p-level
Site 1 18 2.58911 ns
Time 2 18 2.582196 ns
Treatmenta 2 18 0.23249 ns
Site x Time 2 18 2.927248 ns
Site x Treatment 2 18 0.380592 ns
Time x Treatment 4 18 3.611206 *
Site x Time x Treatments 4 18 0.43276 ns
0%10%20%30%40%50%60%70%80%90%
100%
field lab
FPPEFCFDFMV
0%10%20%30%40%50%60%70%80%90%
100%
0%10%20%30%40%50%60%70%80%90%
100%
field lab
FPPEFCFDFMV
FPPEFCFDFMV

Chapter 5: Colonization of cordgrass detritus under contrasting conditions
95
Figure 5: Shannon-Wiener Indices in (A) field and (B) laboratory samples (average of 2
replicates ± 1 standard error)
Discussion
Deposits of decomposing salt marsh vegetation have been described as
(micro)habitats supporting very high densities of nematodes (Montagna and Ruber,
1980; Reice and Stiven, 1983; Buth and de Wolf, 1985; Hemminga and Buth, 1991;
Alkemade et al., 1993, 1994). We observed a similar pattern in the laboratory, where
the total nematode community showed a clear preference for the detritus. Spartina
litter incubated in the field, however, supported only moderate nematode densities,
comparable to the controls.
The available resources did not seem to affect the total nematode community in the
field. Here, detritus was solely used as free space. Similar patterns have been
described for macrophyte detritus in intertidal systems (Reice and Stiven, 1983; Buth
and de Wolf, 1985). Nematodes however can show species-specific responses, which
are masked when considering them as one taxon (Heip et al., 1985; Warwick et al.,
1988; Alkemade et al., 1993; Ólafsson, 1992).
A
B1
1.21.41.61.8
22.22.42.62.8
3
November December January
detrituscontrolsediment
11.21.41.61.8
22.22.42.62.8
3
detrituscontrolsediment
November December January
A
B1
1.21.41.61.8
22.22.42.62.8
3
11.21.41.61.8
22.22.42.62.8
3
November December January
detrituscontrolsediment
11.21.41.61.8
22.22.42.62.8
3
11.21.41.61.8
22.22.42.62.8
3
detrituscontrolsediment
November December January

Chapter 5: Colonization of cordgrass detritus under contrasting conditions
96
Spartina patches in salt marshes are often characterised by a number of (dominant)
species which are much less prominently present in the underlying and surrounding
sediments (Bouwman et al., 1984; Warwick, 1987). Again, only under controlled
laboratory conditions, a similar picture was obtained: the nematode assemblage on the
decaying cordgrass detritus differed considerably from that in the surrounding
sediments. A dominance of Leptolaimus limicolus on the detritus and of
Metachromadora remanei and Axonolaimus typicus in the sediment, and the
appearance of Anoplostoma viviparum explained most of this dissimilarity.
Remarkably, the nematode assemblage on the controls showed a low significant
difference with the sediments, mainly due to the high densities of Ptycholaimellus
ponticus. This species is very common in muddy intertidal sediments, especially in
the upper few mm, below which it is almost absent (Coull, 1988; Steyaert et al.,
2003). Therefore we assume that the dominance of P. ponticus on the controls is due
to a contamination by the upper sediment layer attaching to the plastic strips. In the
field, no preference for any microhabitat was observed; all treatments were dominated
by Daptonema biggi and also other species reaching high densities, such as P.
ponticus and Tripyloides marinus, were characteristic for all treatments. These
observations indicate the overriding importance of the dynamic environmental
conditions in shaping the nematode communities in the field. The physical
(hydrodynamical) disturbance regime in the field site, which is flooded twice a day,
appears to hamper the (species-specific) attraction towards Spartina, and the
community build-up in the resource-rich habitat. A strong dispersal capacity of tidal
currents on the nematode community has been described before (Sherman and Coull,
1980; Armonies, 1988). Alkemade et al. (1993) similarly reported that the majority of
nematodes colonizing Spartina litter in another small Westerschelde marsh belonged
to typical sediment species.
In the same study however correlations between species-specific densities and the
decomposition rate of the detritus were reported. Nematodes are assumed to affect
decomposition processes through interaction with the bacterial community developing
on the detritus (Riemann and Schrage, 1978; Abrams and Mitchell, 1980; Alkemade
et al., 1992a,b, 1993; De Mesel et al., 2003). In our study, a positive correlation
between the decomposition rate of Spartina and densities of Axonolaimus typicus was
found only in the field samples. The relevance of this correlation can be questioned as
A. typicus makes up only a very small proportion of the total nematode community

Chapter 5: Colonization of cordgrass detritus under contrasting conditions
97
and a similar correlation was not found in the laboratory, where A. typicus reached
much higher densities. In the lab, decomposition rate was rather linked to fungal
biomass, although the correlation was not significant (fig. 6A and B). Fungi are
important in the early stages of decomposition of cordgrass (Newell et al., 1989;
Valiela, 1995) and their biomass was significantly higher in the lab compared to the
field. Since there is no obvious trophic link between the free-living nematodes and
fungi, no nematode effect on the decomposition rate was observed.
Figure 6: Decomposition rate expressed as g C loss per day (bars) (average of 4 replicates ±
standard error) and fungal biomass in g (line) (average of 2 replicates ± standard error) in (A)
the laboratory and (B) the field.
The high hydrodynamic impacts at the field site studied may also explain why we did
not find the typical nematode species, mainly Monhysteridae, that are often reported
in high abundances on decomposing Spartina detritus (Bouwman et al., 1984;
Warwick, 1987; Alkemade et al., 1994). Several species of the genera
Diplolaimelloides, Diplolaimella, (Geo)monhystera and Thalassomonhystera are
common on phytodetritus within dense Spartina vegetation a little higher up in the
November December JanuaryDec
ompo
sitio
n ra
te (g
C d
ay-1
)
00.0050.01
0.0150.02
0.0250.03
0.0350.04
0.045
00.020.040.060.080.10.120.140.16 Fungal biom
ass (g)
A
B
Dec
ompo
sitio
n ra
te (g
C d
ay-1
)
00.0050.01
0.0150.02
0.0250.03
0.0350.04
0.045
00.020.040.060.080.10.120.140.16
Fungal biomass (g)
November December January
November December JanuaryDec
ompo
sitio
n ra
te (g
C d
ay-1
)
00.0050.01
0.0150.02
0.0250.03
0.0350.04
0.045
00.0050.01
0.0150.02
0.0250.03
0.0350.04
0.045
00.020.040.060.080.10.120.140.16
00.020.040.060.080.10.120.140.16 Fungal biom
ass (g)
A
B
Dec
ompo
sitio
n ra
te (g
C d
ay-1
)
00.0050.01
0.0150.02
0.0250.03
0.0350.04
0.045
00.020.040.060.080.10.120.140.16
Fungal biomass (g)
November December JanuaryDec
ompo
sitio
n ra
te (g
C d
ay-1
)
00.0050.01
0.0150.02
0.0250.03
0.0350.04
0.045
00.0050.01
0.0150.02
0.0250.03
0.0350.04
0.045
00.020.040.060.080.10.120.140.16
00.020.040.060.080.10.120.140.16
Fungal biomass (g)
November December January

Chapter 5: Colonization of cordgrass detritus under contrasting conditions
98
marsh (Moens unpubl.). These habitats are presumably more sheltered and tidal
currents are either reduced, reducing resuspension of the nematodes in the water
column, and thus their passive distribution (Armonies, 1988), or they are not flooded
daily. The latter situation can be compared with the laboratory experiment in which
active migration of the nematodes can determine their micro-distribution. The lack of
these species in the inoculum explains why they were not found in the lab samples.
The stable laboratory conditions resulted in the development of different nematode
communities, with other species dominating the assemblages, namely Leptolaimus
limicolus in the lab and Daptonema biggi in the field. In addition to these differences
in species composition, we had also expected different community attributes in the
field versus the laboratory. First of all, the resource characteristics in field and lab
settings show important differences. While we added the same amount of organic
matter in lab and field, lab incubations were performed in the dark and hence did not
allow a significant primary production by microphytobenthos. Diatoms and other
microalgae are an important food source for a variety of nematodes, mainly belonging
to the deposit and epigrowth feeders (Wieser, 1953; Moens and Vincx, 1997). Due to
the absence of microphytobenthos, food items in the laboratory were probably
dominated by bacteria, especially associated with the decaying organic matter.
Bacteria serve as food sources for microvores, deposit feeders and, to a lesser extent,
facultative predators (Moens and Vincx, 1997). We expected to find a reflection of the
food availability in the trophic structure of the nematode communities; however these
assumptions were only partly confirmed. High densities of deposit feeders were found
in the field, which is common in muddy sediments. Next to the high densities of
Daptonema biggi, also Daptonema oxycerca, Desmolaimus zeelandicus and
Praeacanthonchus punctatus occurred in considerable densities in all treatments. In
the laboratory, however, the proportion of deposit feeders was significantly lower than
in the field. Only Axonolaimus typicus and, to a lesser extent, D. zeelandicus were
present in all treatments. This indicates some flexibility within the trophic group of
the deposit-feeders: some species did survive on a mainly bacterial diet, while others,
such as Daptonema species and P. punctatus, clearly required microphytobenthos as a
food source. In the lab, deposit feeders seemed to be partly replaced by microvores,
with Leptolaimus limicolus reaching high densities in all microhabitats, especially on
the detritus, and also Halalaimus minusculus being prominently present in the
sediment and on controls. These species have very small buccal cavities through

Chapter 5: Colonization of cordgrass detritus under contrasting conditions
99
which particles no larger than bacteria can pass. The life strategy of microvores is thus
adapted to bacteria as major food source, and therefore they might be competitively
stronger than deposit feeders when prey is dominated by bacteria. Remarkably, the
expected lack of epigrowth feeders in the lab due to the absence of their presumed
food source was not observed; they made up about 25% of the total nematode
community, similar to the field. High numbers of mainly M. remanei and P. ponticus
were found, but also Microlaimus globiceps and Microlaimus nanus were quite
abundant. This suggests a trophic plasticity of the epigrowth feeders. Bacteria are the
most plausible alternative food source for these species although this has been
doubted in the past (Tietjen and Lee, 1973; Jennings and Deutch, 1975; Deutsch,
1978). A recent study, however, showed a positive correlation between epigrowth
feeders and dividing bacterial cells, rather than with chloropigment concentrations
(Danovaro and Gambi, 2002). Due to their well developed buccal armature, M.
remanei and P. ponticus, have been assumed to predate on other nematode species
(Wieser, 1953), however this seems unlikely since no such observations have yet been
done. Overall densities of predators and facultative predators were rather low both in
the field and the lab, but still they made up a significantly larger proportion of the
total community in the lab in comparison with the field samples. The stable conditions
in the lab will have allowed (facultative) predators, which generally have long
reproduction times, to establish a population.
Community complexity and diversity among sites and treatments were remarkably
similar. One could have expected the resource characteristics to affect community
diversity, resulting in differences between litter treatments, controls and sediment.
The daily currents can be regarded as disturbances (Garstecki and Wickham, 2003)
generating and maintaining higher diversity compared to the stable lab conditions
(Connell, 1978). However, when disturbance is too strong, this can result in a negative
effect on diversity (Connell, 1978). Since the distribution of the nematodes already
illustrated that the tidal currents might have severely affected the establishment of
their community, we assume that disturbance was indeed too intense and prevented
the development of a more diverse nematode community.
To conclude, we found that nematodes show a clear (species-specific) habitat
preference when only biotic factors play part. In the unstable, intertidal system
however, this active dispersion is of minor importance and nematodes are distributed
at random by the tidal currents. These findings are in agreement with Hogue (1982),

Chapter 5: Colonization of cordgrass detritus under contrasting conditions
100
who noticed that biotic factors tend to aggregate meiofauna, while abiotic factors tend
to disperse them.
Both structural and functional characteristics of the nematode community structure
were clearly affected by the thoroughly different environmental conditions in the lab
and in the field, while diversity was similar at both sites. The trophic structure of the
nematode communities reflected to a certain extent food availability in the two sites,
but our data also showed that species-specific flexibility in food uptake within trophic
guilds is present, especially for the epigrowth feeders and to a lesser extent for the
deposit feeders.
Acknowledgements
We greatly thank Gunter Van Ryckegem for the ergosterol analyses. The first author
acknowledges a grant from the Flemish Institute for the Promotion of Scientific-
Technological Research (IWT). The last author is a postdoctoral fellow with the
Flemish Fund for Scientific Research. Further financial support was obtained from
Ghent University in BOF projects 1205398 and 011060002.

Conclusions

Conclusions
103
Conclusions Species manipulation experiment
Microcosm studies in ecological research
Both experiments in this thesis are (partly) based on microcosm incubations under
controlled laboratory conditions. Microcosm experiments have been widely used in
ecological studies during the last few decades (Ives et al., 1996) to unravel
phenomena observed in the field or to generate hypotheses about the behavior and
functioning of real ecosystems (Drake et al., 1996). They are a generally accepted
research tool in soil ecology (Moore et al., 1996). While in other ecological branches
the limitations of scale can be a problem, this is generally much less the case in soil
systems. Organisms of interest are mostly small so that their actual habitat size can be
approached. Some scientists have criticized the (over)simplification of natural
systems in laboratory set-ups (Carpenter, 1996; Kampichler et al., 2001); however this
can also be considered as the strength of this approach (Lawton, 1996). Natural
ecosystems are very complex, and consist of a large number of interacting processes.
Understanding individual processes seems essential to get insight into the whole
ecosystem. Integrating these observations into field studies could then lead to a better
insight into functions and processes (Carpenter, 1996; Moore et al., 1996).
Development of nematode populations
Some findings in microcosm experiments could simply never have been detected in
field studies. This applies for instance for the interference interactions between the
monhysterid nematode species. Remarkably, despite the competition, they manage
to establish and maintain a rather stable community, though with low (overall)
densities. Based on traditional competition models one would expect a competitively
inferior species to be excluded from the community under the stable conditions of this
laboratory experiment.
Food partitioning is one of the processes which may allow coexistence of
bacterivorous nematode species in field sediments (Grewal and Wright, 1992; Moens
et al., 1999) that could have occurred in our systems as well, as shown by the
differences in the bacterial community structure in the monospecific treatments. Still,

Conclusions
104
this does not provide an explanation for the lower (overall) densities. Based on our
results, we believe that chemical interference should be considered. In our closed
laboratory set-up, accumulation of chemicals could have hampered general growth; in
field sediments this might protect monospecific aggregates from intrusion of other
species. Aggregation is thought to be a way of minimizing interspecific competition,
thus of enhancing the coexistence of highly diverse communities with functionally
similar species (Ettema, 1998). It would be very interesting to further examine these
kinds of interactions in order to formulate a satisfactory explanation on their nature.
Another peculiar observation in our microcosms was the extremely high densities of
Panagrolaimus paetzoldi. As they are extreme colonizers, they generally occur in
sites with freshly deposited organic matter where bacterial activity is high. We
observed however that P. paetzoldi can maintain high densities even when bacterial
activity was suppressed. We assume they might be able to directly assimilate
detrital compounds, for instance through enzyme sharing with the bacterial
community (Riemann and Helmke, 2002). In any case, this species was better adapted
to utilize the available resources than the monhysterid nematodes.
Effect of bacterivorous nematodes on decomposition rates and microbial activity
The idea for studying the effect of bacterivorous nematode diversity on decomposition
processes originated from studies describing a stimulation of decay rates in the
presence of bacterivores (e.g. Abrams and Mitchell, 1980; Findlay and Tenore, 1982;
Anderson et al., 1983; Alkemade et al., 1992 a, b). The outcome of our experiment
was thus rather surprising. No stimulation of the decomposition rate, nor of the
microbial community was observed in the presence of nematodes. We assume that
the impact of bioturbation, generally considered one of the main pathways of
stimulation (Alkemade et al., 1992b) was minor in our microcosms since fragments of
cordgrass leaves were incubated on a sediment surface, as they generally decompose
in situ, instead of mixed with the sediment.
The question then still remains why the decomposition process was slowed down
in the presence of nematodes. Based on our results, we propose a hypothesis which
could explain the different rates in decomposition in the control compared to the
nematode treatments (see also schematic overview in figure 1). We have indications
(observations and data on the respiration rate) that the microbial community in the
controls was dominated by fungi. Fungi are important in the early stages of the

Conclusions
105
decomposition process of cordgrass. In the presence of the nematodes, fungal growth
seemed to be suppressed. This could have had two different causes: either nematodes
directly antagonized the fungi (cfr Kaufman, 1983), or they interacted by favoring the
Figure 1: Schematic representation of the hypothesis on the decomposition process in
absence and presence of nematodes. The dotted lines illustrate N-regeneration by the
nematodes and its subsequent adsorption to the detritus. (DOM: Dissolved Organic Matter,
POM: Particulate Organic Matter)
bacterial community, for instance by facilitating bacterial growth on mucus tracks
(Riemann and Schrage, 1978), which could in turn interfere with fungal growth
(Mille-Lindblom and Tranvik, 2003; Wohl and McArthur, 2001). Both steps in the
latter pathway have already been established in aquatic systems. Bacteria, thus partly
replacing fungi in the nematode treatments, can more easily use dissolved organic
compounds, rather than particulate substrates (Mille-Lindblom and Tranvik, 2003).
Leaching of organic compounds from detritus is generally the first step in a
decomposition process (Valiela, 1995) and was probably enhanced in our experiment
by the treatment of the cordgrass leaves prior to the incubation. Leaching was mainly
confirmed by the high N- loss in all treatments during the first 10 days of the
in ABSENCE of NEMATODES in PRESENCE of NEMATODES
Fungi
Fungi
Bacteria
Bacteria
Nematodes
---
-
+-
C N
DOM
POM
C N
DOM
N
in ABSENCE of NEMATODES in PRESENCE of NEMATODES
Fungi
Fungi
Bacteria
Bacteria
Nematodes
-----
-
+-
C N
DOM
POM
C N
DOM
N

Conclusions
106
experiment. Bacteria will thus have first degraded this dissolved organic matter,
before utilizing the more inaccessible compounds from the detritus, resulting in the
slower decomposition rate observed in the presence of nematodes in our experiments.
By contrast, fungi have the ability to penetrate solid organic substrates and digest
them from within (Newell, 1996). This could have caused the fast initial
decomposition rates in the controls. Within the nematode treatments, different
characteristics of the bacterial communities (e.g. composition, efficiency,….),
interactions with the nematodes (e.g. enzyme sharing,….) and interference of fungi
and bacteria in the microbial activity measurements probably obscured clear
relationships between microbial activity and decomposition rates in the nematode
treatments.
Impact of bacterivorous nematodes on the bacterial community composition
Bacterial communities in our microcosms were shaped by a combination of bottom-up
and top-down impacts. The types of bacteria were primarily determined by the
substrate as the same bacterial species occurred in different treatments. However,
clear species specific top-down impacts of the bacterivorous nematodes on the
pool of bacteria were evident, even at relatively low nematode densities as in the
presence of the monhysterids. We assume that each species grazed on a particular part
of the bacterial community, as has already been suggested by some controlled
laboratory experiments (Grewal and Wright, 1992; Moens et al., 1999). The effect of
nematode diversity on the composition of the bacterial community was not analyzed,
but we did check the impact on bacterial diversity. First, a lower bacterial diversity
in the presence of two- and three- species combinations was noted. This might be
the summed effect of selective grazing pressure of the individual nematode species,
suppressing their preferred food sources below the detection limit of DGGE.
Secondly, within each nematode diversity level, bacterial diversity was
considerably lower when Panagrolaimus paetzoldi was present. This is most likely
due to the high densities of this species, resulting in overgrazing of the bacterial
community, rather than to characteristics of the species itself.
Does species diversity matter in bacterivorous nematode communities?
One of the most discussed theories on the relationship between species diversity and
the functioning of ecosystems predicts redundancy among functionally similar species

Conclusions
107
(Walker, 1992; Schläpfer et al., 1999). Based on this hypothesis, we expected the
bacterivorous nematode species used in our experiment to perform the same function
and no or a minor impact of diversity on the decomposition process and the associated
microbial community. This was however not confirmed, as our results suggest an
idiosyncratic response of the system upon changing diversity or species
composition. It can of course be argued that diversity levels in our experiment were
low compared to natural systems. Idiosyncratic effects of changing community
composition may become apparent mainly in experiments with low numbers of
species. It would thus be interesting to integrate higher diversity levels, or even
natural nematode communities, into a next phase of this study. Still, the effect of
species or species combinations on the decomposition process in our microcosms
could not have been predicted at the start of the experiment. In other words,
redundancy of nematodes within the same functional group, or positive effects when
adding species to a system can not be assumed a priori. Mikola and Setälä (1998)
came to similar conclusions when testing the effect of bacterivorous and fungivorous
nematode species on decomposition rates in a terrestrial system.
As for the effect on bacterial diversity, the species-specific effect of Panagrolaimus
paetzoldi on the microbial activity will have been due to their high densities, which
resulted in overgrazing, rather than being caused by intrinsic properties of the species.
Suggestions for future research
As explained in the introductory paragraph, it is important to link microcosm
experiments to field studies in order to validate their significance and to understand
the functioning of the ecosystem. This step could however not be established in the
framework of this thesis. Before doing so, it is recommended to further expand our
knowledge under laboratory experiments. As mentioned above, it would be interesting
to integrate higher diversity levels and natural nematode communities into the study.
A next step might then be to simulate tidal floods, temperature fluctuations, day
and night rhythm, etc in experiments with manipulated species composition and
natural nematode assemblages in order to test their impact on the observations under
controlled conditions (interspecific interactions, the relationship between nematodes
and bacteria, effects of presence of nematodes on decomposition rates). However,
before doing so, a field study might be required to assess under which tidal regime the
nematode species from our experiment occur. Our colonization experiment has for

Conclusions
108
instance shown that these species are absent in the lower parts of the salt marsh. They
presumably prefer sites higher up the marsh which are flooded only occasionally
(Alkemade et al., 1993; Moens, pers. comm.). Another observation in the field
samples of the colonization experiment was that fungal growth was considerably less
compared to laboratory samples; this could thus indicate that the bacterial community
is favored under fluctuating conditions. The impact of the nematodes on the
bacterial community and on the decomposition rate may be altered in a bacteria-
dominated microbial community. It might therefore also be interesting to integrate a
bacterial based food web into the laboratory incubations, using a different substrate,
so that the impact of bacterivorous nematodes in both soil systems can be compared.
Integrating species manipulation in field experiments may be difficult to achieve. Due
to the open character of the system, and the supply of nematodes from surrounding
sediments with the tidal action, excluding species from an experimental field seems
impossible. Including natural communities in the laboratory experiments could
however enable an evaluation of the processes observed under controlled conditions.
Colonization experiment
Nematode distribution
Under controlled laboratory conditions nematodes from salt marsh sediments have
the ability to actively colonize and exploit a resource-rich microhabitat, such as
freshly deposited cordgrass detritus, but they can not take advantage of this ability
in a physically disturbed field environment. This is most probably due to the daily
tidal disturbance which suspends the nematodes and distributes them over the
sediments surface.
The species composition in our samples showed that the occurrence of mainly
Monhysterida and Rhabditida on phytodetritus may not be as universal as previously
thought, and is presumably restricted to higher, more sheltered parts of intertidal areas
which are not flooded daily (Alkemade et al., 1993; Moens, pers. comm.). These more
stable conditions could favor their growth and might allow active migration of the
nematode to determine their micro-distribution, similar to our laboratory incubations
in this experiment.

Conclusions
109
Functional aspects of the nematode community
Some functional aspects of the nematode community in the laboratory samples
deviated from our expectations. Epigrowth feeders were assumed to be hampered due
to growth inhibition of their supposed food sources, namely microphytobenthos,
under dark laboratory conditions. However, their proportion of the total community
did not differ from field samples. Hence, they must be able to utilize other resources,
presumably bacteria. Since mostly the same species occurred at both sites, this
indicates that the functional position of a species might differ according to food
availability.
The main food items for deposit feeders have been identified as bacteria and diatoms.
While some deposit-feeders adapted very well to a bacterial diet in the laboratory (e.g.
Axonolaimus typicus), others remained highly dependent on the presence of diatoms
as a food source (e.g. Daptonema biggi). These observations point to substantial
variation in diet between nematode species within functional groups.
These observations on both the epistrate feeders and deposit feeders confirm the
above formulated conclusions drawn from the species manipulation experiment
in that the current trophic classification of free living marine nematodes is
inadequate to make predictions on the function of the nematode species in an
ecosystem.
Diversity of the nematode community
Several mechanisms have been proposed by which nematode diversity in soils could
be maintained (Ettema, 1998). Physical disturbance is one of them and could be an
important process in maintaining high diversity of nematodes with similar resource
needs in intertidal environments, such as salt marshes. However, equally high
nematode diversity under controlled laboratory conditions and in field samples
was found. In other words, disturbance does not seem to be essential in maintaining
these high diversities in the studied system; biotic interactions obviously led to a
stable coexistence of comparable numbers of (functionally similar) species.
Resource partitioning might explain the coexistence of the nematode species in our
model system. Still, stronger competition in the laboratory could be expected as the
spectrum of available food items was restricted: most species depend on bacteria for

Conclusions
110
their growth since the development of the other main food source, microphytobenthos,
was hampered. However, this did not result in a reduction of nematode diversity. In
accordance with conclusions of the species manipulation experiment, chemical
interactions supporting coexistence of highly diverse communities could be
considered. Accumulations of chemicals might have been less severe than in the
species manipulation experiments, as the scale of the microcosms was considerably
larger, but nonetheless it might be part of the reason for the lower densities in the
sediment in the laboratory than in the field.

Appendix
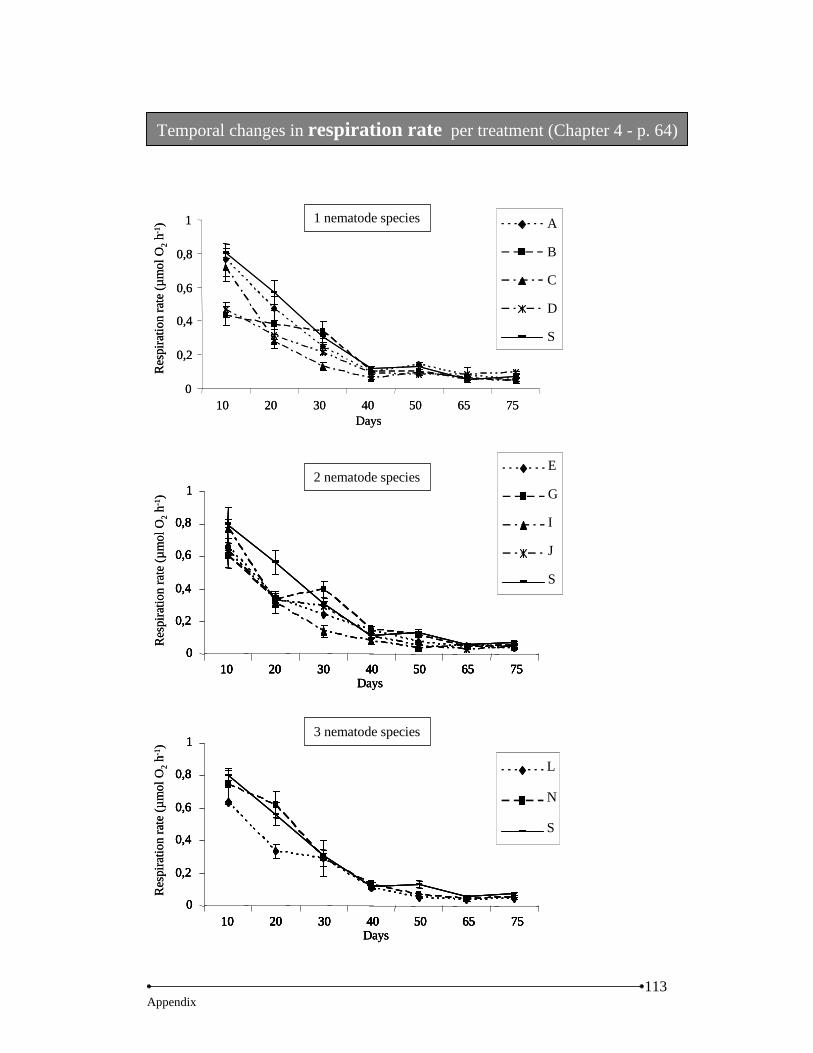
Appendix
113
10 20 30 40 50 65 750
0,2
0,4
0,6
0,8
1
Res
pira
tion
rate
(µm
ol O
2h-1
)
Days10 20 30 40 50 65 75
0
0,2
0,4
0,6
0,8
1
Res
pira
tion
rate
(µm
ol O
2h-1
)
Days
Res
pira
tion
rate
(µm
ol O
2h-1
)
0
0,2
0,4
0,6
0,8
1
10 20 30 40 50 65 75Days
Res
pira
tion
rate
(µm
ol O
2h-1
)
0
0,2
0,4
0,6
0,8
1
10 20 30 40 50 65 75Days
0
0,2
0,4
0,6
0,8
1
10 20 30 40 50 65 750
0,2
0,4
0,6
0,8
1
10 20 30 40 50 65 7510 20 30 40 50 65 75Days
Res
pira
tion
rate
(µm
ol O
2h-1
)
0
0,2
0,4
0,6
0,8
1
10 20 30 40 50 65 75Days
Res
pira
tion
rate
(µm
ol O
2h-1
)
0
0,2
0,4
0,6
0,8
1
10 20 30 40 50 65 75Days
0
0,2
0,4
0,6
0,8
1
10 20 30 40 50 65 750
0,2
0,4
0,6
0,8
1
10 20 30 40 50 65 7510 20 30 40 50 65 75Days
A
B
C
D
S
A
B
C
D
S
E
G
I
J
S
E
G
I
J
S
L
N
S
L
N
S
1 nematode species
2 nematode species
3 nematode species
Temporal changes in respiration rate per treatment (Chapter 4 - p. 64)
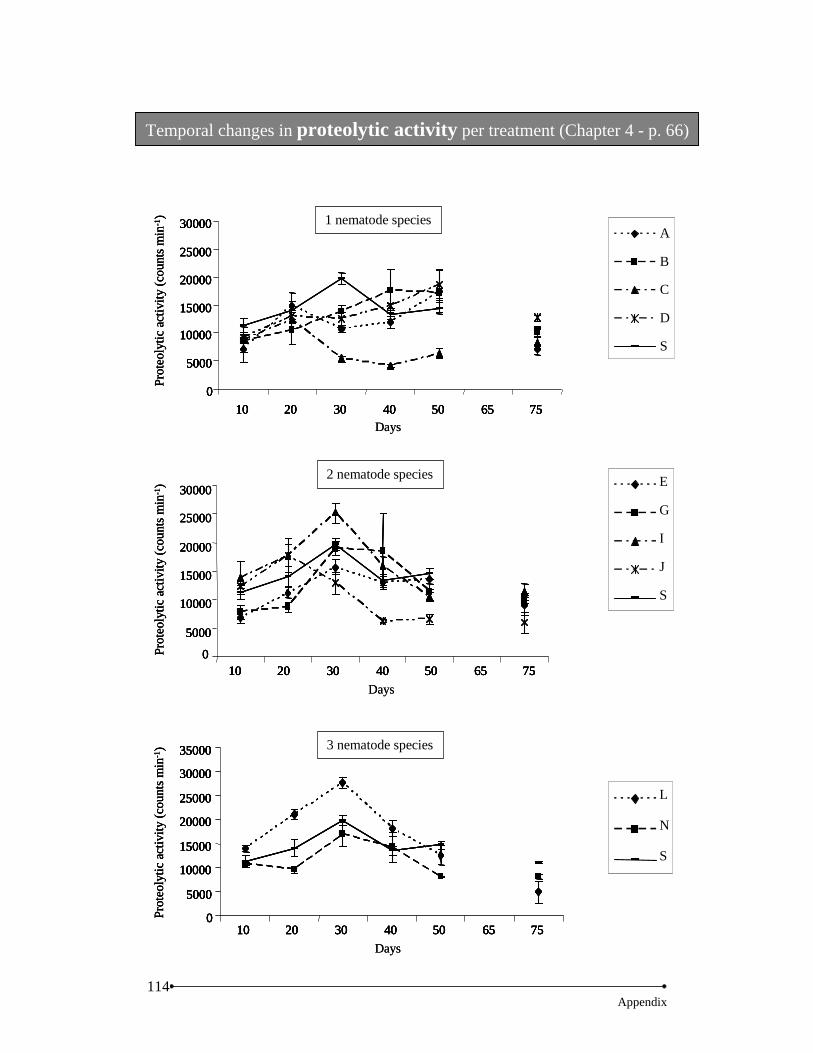
Appendix
114
0
5000
10000
15000
20000
25000
30000
10 20 30 40 50 65 75
Prot
eoly
tic a
ctiv
ity (c
ount
s min
-1)
Days
0
5000
10000
15000
20000
25000
30000
10 20 30 40 50 65 75
Prot
eoly
tic a
ctiv
ity (c
ount
s min
-1)
0
5000
10000
15000
20000
25000
30000
10 20 30 40 50 65 750
5000
10000
15000
20000
25000
30000
0
5000
10000
15000
20000
25000
30000
10 20 30 40 50 65 7510 20 30 40 50 65 75
Prot
eoly
tic a
ctiv
ity (c
ount
s min
-1)
Days
0
5000
10000
15000
20000
25000
30000
10 20 30 40 50 65 75
Prot
eoly
tic a
ctiv
ity (c
ount
s min
-1)
Days
0
5000
10000
15000
20000
25000
30000
10 20 30 40 50 65 75
Prot
eoly
tic a
ctiv
ity (c
ount
s min
-1)
0
5000
10000
15000
20000
25000
30000
10 20 30 40 50 65 750
5000
10000
15000
20000
25000
30000
10 20 30 40 50 65 7510 20 30 40 50 65 75
Prot
eoly
tic a
ctiv
ity (c
ount
s min
-1)
Days
0
5000
10000
15000
20000
25000
30000
35000
10 20 30 40 50 65 75
Prot
eoly
tic a
ctiv
ity (c
ount
s min
-1)
Days
0
5000
10000
15000
20000
25000
30000
35000
10 20 30 40 50 65 75
Prot
eoly
tic a
ctiv
ity (c
ount
s min
-1)
0
5000
10000
15000
20000
25000
30000
35000
10 20 30 40 50 65 750
5000
10000
15000
20000
25000
30000
35000
0
5000
10000
15000
20000
25000
30000
35000
10 20 30 40 50 65 7510 20 30 40 50 65 75
Prot
eoly
tic a
ctiv
ity (c
ount
s min
-1)
Days
A
B
C
D
S
A
B
C
D
S
E
G
I
J
S
E
G
I
J
S
L
N
S
L
N
S
1 nematode species
2 nematode species
3 nematode species
Temporal changes in proteolytic activity per treatment (Chapter 4 - p. 66)

Appendix
115
Shan
non-
Wie
ner I
ndex
(H’)
1,51,7
1,92,1
2,32,5
2,72,9
3,1
30 50 65Days
Shan
non-
Wie
ner I
ndex
(H’)
1,51,7
1,92,1
2,32,5
2,72,9
3,1
30 50 65Days
1,5
1,7
1,9
2,1
2,3
2,5
2,7
2,9
3,1
30 50 65Days
Shan
non-
Wie
ner I
ndex
(H’)
1,5
1,7
1,9
2,1
2,3
2,5
2,7
2,9
3,1
30 50 651,5
1,7
1,9
2,1
2,3
2,5
2,7
2,9
3,1
30 50 65Days
Shan
non-
Wie
ner I
ndex
(H’)
1,51,7
1,92,12,3
2,52,7
2,93,1
30 50 65
Shan
non-
Wie
ner I
ndex
(H’)
Days
1,51,7
1,92,12,3
2,52,7
2,93,1
30 50 65
Shan
non-
Wie
ner I
ndex
(H’)
Days
1 nematode species
2 nematode species
3 nematode species
A
B
C
D
S
A
B
C
D
S
E
G
I
J
S
E
G
I
J
S
L
N
S
L
N
S
Temporal changes in bacterial diversity per treatment (Chapter 4 - p. 68)

Appendix
116
100
150
200
250
300
350
400
10 20 30 40 50 65 75
mg
C
Days
100
150
200
250
300
350
400
10 20 30 40 50 65 75100
150
200
250
300
350
400
10 20 30 40 50 65 75
mg
C
Days
100
150
200
250
300
350
400
10 20 30 40 50 65 75
mg
C
Days
100
150
200
250
300
350
400
10 20 30 40 50 65 75100
150
200
250
300
350
400
10 20 30 40 50 65 75
mg
C
Days
100
150
200
250
300
350
400
10 20 30 40 50 65 75
mg
C
Days
100
150
200
250
300
350
400
10 20 30 40 50 65 75100
150
200
250
300
350
400
10 20 30 40 50 65 75
mg
C
Days
1 nematode species
2 nematode species
3 nematode species
A
B
C
D
S
A
B
C
D
S
E
G
I
J
S
E
G
I
J
S
L
N
S
L
N
S
Temporal changes in C-content per treatment (Chapter 4 - p. 70)

Appendix
117
mg
N
0
2
4
6
8
10
12
Days10 20 30 40 50 65 75
mg
N
0
2
4
6
8
10
12
Days10 20 30 40 50 65 7510 20 30 40 50 65 75
0
2
4
6
8
10
12
10 20 30 40 50 65 75
mg
N
Days
0
2
4
6
8
10
12
10 20 30 40 50 65 7510 20 30 40 50 65 75
mg
N
Days
0
2
4
6
8
10
12
10 20 30 40 50 65 75
mg
N
Days
0
2
4
6
8
10
12
10 20 30 40 50 65 750
2
4
6
8
10
12
10 20 30 40 50 65 75
mg
N
Days
A
B
C
D
S
A
B
C
D
S
E
G
I
J
S
E
G
I
J
S
L
N
S
L
N
S
1 nematode species
2 nematode species
3 nematode species
Temporal changes in N-content per treatment (Chapter 4 - p. 71)

Appendix
118
Overview DGGE-profiles (Chapter 4 - p. 68)
D. meyli
D. oschei
P. paetzoldi
D. dievengatensis
Control
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days
D. meyli
D. oschei
P. paetzoldi
D. dievengatensis
Control
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days

Appendix
119
D. meyli + D. oschei
P. paetzoldi + D. dievengatensis
D. oschei + D. dievengatensis
D. meyli + D. oschei + D. dievengatensis
D. meyli + P. paetzoldi + D. dievengatensis
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days
D. meyli + D. oschei
P. paetzoldi + D. dievengatensis
D. oschei + D. dievengatensis
D. meyli + D. oschei + D. dievengatensis
D. meyli + P. paetzoldi + D. dievengatensis
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days
30 days50 days65 days
Overview DGGE-profiles (Chapter 4 - p. 68)

Cited literature

Cited literature
123
Cited literature
Abrams BI, Mitchell MJ (1980) Role of nematode-bacterial interactions in
heterotrophic systems with emphasis on sewage sludge decomposition. Oikos 35: 404-
410
Alkemade R, Wielemaker A, Hemminga MA (1992a) Stimulation of Spartina anglica
leaves by the bacterivorous marine nematode Diplolaimelloides bruciei. J Exp Mar
Biol Ecol 159: 267-278
Alkemade R, Wielemaker A, de Jong SA, Sandee AJJ (1992b) Experimental evidence
for the role of bioturbation by the marine nematode Diplolaimella dievengatensis in
stimulating the mineralization of Spartina anglica detritus. Mar Ecol Prog Ser 90:
149-155
Alkemade R, Wielemaker A, Hemminga MA (1993) Correlation between nematode
abundance and decomposition rate of Spartina anglica leaves. Mar Ecol Prog Ser 99:
293-300
Alkemade R, Wielemaker A, Herman PMJ, Hemminga MA (1994) The population
dynamics of Diplolaimelloïdes bruciei, a nematode associated with the salt marsh
plant Spartina anglica. Mar Ecol Prog Ser 105: 277-284
Aller RC, Aller JY (1992) Meiofauna and solute transport in marine muds. Limnol
Oceanogr 37: 1018-1033
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment
search tool. J Mol Biol 215: 403-410
Anderson RV, Coleman DC, Cole CV, Elliott ET (1981) Effect of the nematodes
Acrobeloides sp. and Mesodiplogaster lheritieri on substrate utilization and nitrogen
and phosphorus mineralization in soil. Ecology 62: 549-555
Anderson RV, Gould WD, Woods LE, Cambardella C, Ingham RE, Coleman DC
(1983) Organic and inorganic nitrogenous losses by microbivorous nematodes in soil.
Oikos 40: 75-80

Cited literature
124
Armonies W (1988) Hydrodynamic factors affecting behaviour of intertidal
meiobenthos. Ophelia 28: 183-193
Beare MH, Parmelee RW, Hendrix PF, Cheng W, Coleman DC, Crossley DA (1992)
Microbial and faunal interactions and effects on litter nitrogen and decomposition in
agroecosystems. Ecol Monogr 62: 569-591
Bell SS, Sherman KM (1980) A field investigation of meiofauna dispersal: tidal
resuspension and implications. Mar Ecol Prog Ser 3: 245-249
Bell SS, Watzin MC, Coull BC (1978) Biogenic structure and its effect on the spatial
heterogeneity of meiofauna in a salt marsh. J Exp Mar Biol Ecol 35: 99-107
Bertelsen RD (1998) Active and passive settling by marine benthic nematodes. Diss
Abstr Int B Sci Eng 58: 3425
Bone LW (1982) Reproductive chemical communication of helminthes. II.
Aschelminthes. Internat J Invert Reprod 5: 311-321
Bongers T, Bongers M (1998) Functional diversity of nematodes. Appl Soil Ecol 10:
239-251
Bongers T, Alkemade R, Yeates GW (1991) Interpretation of disturbance-induced
maturity decrease in marine nematode assemblages by means of the Maturity Index.
Mar Ecol Prog Ser 76: 135-142
Bongers T, de Goede RGM, Korthals GW, Yeates GW (1995) Proposed changes of c-
p classification for nematodes. Russ J Nematol 3: 61-62
Bonkowski M, Griffiths B, Scrimgeour C (2000) Substrate heterogeneity and
microfauna in soil organic ‘hotspots’ as determinants of nitrogen capture and growth
of ryegrass. Appl Soil Ecol 14: 37-53
Boucher G, Chamroux S (1976) Bacteria and meiofauna in an experimental sand
ecosystem. I. Material and preliminary results. J Exp Mar Biol Ecol 24: 237-249

Cited literature
125
Bouwman LA, Romeyn K, Kremer DR, Van Es FB (1984) Occurrence and feeding
biology of some nematode species in estuarine aufwuchscommunities. Cah Biol Mar
25: 287-303
Buth GJC, de Wolf L (1985) Decomposition of Spartina anglica, Elytrigia pungens
and Halimione portulacoides in a Dutch salt marsh in association with faunal and
habitat influences. Vegetatio 62: 337-355
Buth GJC, Voesenek LACJ (1988) Respiration of standing and fallen plant litter in a
Dutch salt marsh. In Vegetation structure in relation to carbon and nutrient economy.
Verhoeven JTA, Heil GW, Werger MJ (eds) S.P.B. Academic Publishing, The Hague:
51-60
Buzas MA (1969) Foraminiferal species densities and environmental variables in an
estuary. Limnol Oceanogr 14: 411-422
Cardinale BJ, Palmer MA, Collins SL (2002) Species diversity enhances ecosystem
functioning through interspecific facilitation. Nature 415: 426-429
Carpenter SR (1996) Microcosm experiments have limited relevance for community
and ecosystem ecology. Ecology 77: 677-680
Chandler GT, Fleeger JW (1987) Facilitative and inhibitory interactions among
estuarine meiobenthic harpacticoid copepods. Ecology 68: 1906-1919
Clarke KR (1993) Non-parametric multivariate analyses of change in community
structure. Aus J Ecol Syst 3: 169-192
Clarke KR, Gorley RN (2001) PRIMER v5: User Manual/Tutorial. PRIMER-E,
Plymouth
Clarke KR, Warwick RM (2001) Change in marine communities: an approach to
statistical analysis and interpretation, 2nd edition. PRIMER-E, Plymouth
Coldeira MC, Ryel RJ, Lawton JH, Pereira JS (2001) Mechanisms of positive
biodiversity-production relationships: insights provided by delta C-13 analysis in
experimental Mediterranean grassland plots. Ecol Letters 4: 439-443

Cited literature
126
Coleman DC, Reid CPP, Cole CV (1983) Biological strategies of nutrient cycling in
soil systems. In Advances in ecological research. Volume 13. Macfayden A, Ford ED
(eds) Academic Press, New York, USA
Collins S (1995) The measurement of stability in grasslands. Trends Ecol Evol 10: 95-
96
Commito JA, Tita G (2002) Differential dispersal rates in an intertidal meiofauna
assemblage. J Exp Mar Biol Ecol 268: 237-256
Connell JH (1978). Diversity in tropical rain forests and coral reefs. Science 199:
1302-1311
Coull BC (1988) Ecology of the marine meiofauna. In Introduction to the study of
meiofauna. Higgins RP, Thiel H (eds) Smithsonian Institution Press, Washington DC,
London: 18-38
Coull BC (1990) Are members of the meiofauna food for higher trophic levels? Trans
Am Microsc Soc 109: 233-246
Cullen DJ (1973) Bioturbation of superficial marine sediments by interstitial
meiobenthos. Nature 242: 323-324
Danovaro R, Gambi C (2002) Biodiversity and trophic structure of nematode
assemblages in seagrass systems: evidence for a coupling with changes in food
availability. Mar Biol 141: 667-677
David AR (1996) Association among ascidians: facilitation of recruitment in Pyura
spinifera. Mar Biol 126: 34-41
Degans H, Zöllner E, Van der Gucht K, De Meester L, Jürgens K (2002) Rapid
Daphnia-mediated changes in microbial community structure: an experimental study.
FEMS Microbiol Ecol 42: 137-149
Degens BP (1998) Decreases in microbial functional diversity do not result in
corresponding changes in decomposition under different moisture conditions. Soil
Biol Biochem 30: 1989-2000

Cited literature
127
De Mesel I, Derycke, S, Swings, J, Vincx, M, Moens, T (2003) Influence of
bacterivorous nematodes on the decomposition of cordgrass. J Exp Mar Biol Ecol
296: 227-242
De Mesel I, Derycke S, Moens T, Van der Gucht K, Vincx M, Swings J (2004) Top-
down impact of bacterivorous nematodes on the bacterial community structure: a
microcosm study. Env Microbiol (in press)
Deutsch A (1978) Gut structure and digestive physiology of two marine nematodes,
Chromadorina germanica, Bütschli, 1874, and Diplolaimella sp. Biological Bulletin.
Marine Biological Laboratory, Woods Hole 155: 317-335
Dietrich G, Kalle K (1957) Allgemeine Meereskunde. Eine Einführung in die
Ozeanographie. Gebrüder Borntraeger, Berlin, Nikolassee
Downing AL, Leibold MA (2002) Ecosystem consequences of species richness and
composition in pond food webs. Nature 416: 837-841
Drake JA, Huxel GR, Hewitt CL (1996) Microcosms as models for generating and
testing community theory. Ecology 77: 670-677
Ehrlich PR, Ehrlich AH (1981) Extinction. The causes and consequences of the
disappearance of species. Random House, New York
Eichner CA, Erb RW, Timmis KN, Wagner-Döbler I (1999) Thermal gradient gel
electrophoresis analysis of bioprotection from polluted shocks in the activated sludge
microbial community. Appl Environ Microbiol 65: 102-109
Emmerson MC, Solan M, Emes C, Paterson DM, Rafaelli D (2001) Consistent
patterns and the idiosyncratic effects of biodiversity in marine ecosystems. Nature
411: 73-77
Ettema CH (1998) Soil nematode diversity: species coexistence and ecosystem
function. J Nematol 30: 159-169
Ewel JJ, Mazzarino MJ, Berish CW (1991). Tropical soil fertility changes under
monocultures and successional communities of different structure. Ecol Appl 1: 289-
302

Cited literature
128
Fegley SR (1987) Experimental variation of near-bottom current speeds and its effects
on depth distribution of sand-living meiofauna. Mar Biol 95: 183-191
Fenchel T (1996) Worm burrow and oxic microniches in marine sediments. 2.
Distribution patterns of ciliated protozoa. Mar Biol 127: 297-301
Ferris H, Venette RC, Lau SS (1997) Population energetics of bacterial-feeding
nematodes: carbon and nitrogen budgets. Soil Biol Biochem 29: 1183-1194
Findlay SEG (1981) Small-scale spatial distribution of meiofauna on a mud –and
sandflat. Est Coast Shelf Sci 12: 471-484
Findlay SEG (1982) Influence of sampling scale on apparent distribution of
meiofauna on a sandflat. Estuaries 5: 322-324
Findlay S, Tenore KR (1982) Effect of a free-living marine nematode (Diplolaimella
chitwoodi) on detrital carbon mineralization. Mar Ecol Prog Ser 8: 161-166
Fleeger JW, Gee JM (1986) Does interference competition determine the vertical
distribution of meiobenthic copepods? J Exp Mar Biol Ecol 95:173-181
Fleeger JW, Chandler GT, Fritzhugh GR, Phillips FE (1984) Effects of tidal currents
on meiofauna densities in vegetated salt marsh sediments. Mar Ecol Prog Ser 19: 49-
53
Freckman DW (1988) Bacterivorous nematodes and organic-matter decomposition.
Agr Ecosyst Environ 24 : 195-217
Fridley JD (2001) The influence of species diversity on ecosystem productivity: How,
where, and why? Oikos 93: 514-526
Fromin N, Hamelin J, Tarnawski S, Roesti D, Jourdain-Miserez K, Forestier N,
Teyssier-Cuvelle S, Gillet F, Aragno M, Rossi P (2002) Statistical analysis of
denaturing gel electrophoresis (DGE) fingerprinting patterns. Environ Microbiol 4:
634-643

Cited literature
129
Garstecki T, Wickham SA (2003) The response of benthic rhizopods to sediment
disturbance does not support the intermediate disturbance hypothesis. Oikos 103: 528-
536
Gee JM (1989) An ecological and economic review of meiofauna as food for fish.
Zool J Linnean Soc 96: 243-261
Gessner MO, Newell SY (1997) Bulk quantitative methods for the examination of
eukaryotic organoosmotrophs in plant litter. In Manual of environmental
microbiology. Hurst CJ, Knudsen GR, McInerney MJ, Stetzenbach LD, Walter MV
(eds) ASM press, Washington DC: 295-308
Givnish TJ (1994) Does diversity beget stability? Nature 371: 113-114
Green C (1971) Mating and host finding behaviour of plant nematodes. In Plant
parasitic nematodes. Vol. II. Zuckerman BM, Rohde RA (eds) Academic Press, New
York: 247-266
Grewal PS (1991) Relative contribution of nematodes (Caenorhabditis elegans) and
bacteria towards the disruption of flushing patterns and losses in yield and quality of
mushrooms (Agaricus bisporus). Ann Appl Biol 119: 438-499
Grewal PS, Hand P (1992) Effects of bacteria isolated from a saprophagous rhabditid
nematode Caenorhabditis elegans on mycelial growth of Agaricus bisoporus. J App
Bacteriol 72: 173-179
Grewal PS, Richardson PN (1991) Effects of Caenorhabditis elegans (Nematoda:
Rhabditidae) on yield and quality of the cultivated mushroom Agaricus bisporus. Ann
App Biol 118: 381-394
Grewal PS, Wright DJ (1992) Migration of Caenorhabditis elegans (Nematoda:
Rhabditidae) larvae towards bacteria and the nature of the bacterial stimulus. Fund
Appl Nematol 15: 159-166
Griffiths BS, Bonkowski M, Dobson G, Caul S (1999) Changes in soil microbial
community structure in the presence of microbial-feeding nematodes and protozoa.
Pedobiologia 43: 297-304

Cited literature
130
Griffiths BS, Ritz K, Bardgett RD, Cook R, Christensen S, Ekelund F, Sorensen SJ,
Baath E, Bloem J, de Ruiter PC, Dolfing J, Nicolardot B (2000) Ecosystem response o
pasture soil communities to fumigation induced microbial diversity reductions: an
examination of the biodiversity-ecosystem function relationship. Oikos 90: 279-294
Groenendijk AM (1984) Primary production of four dominant salt-marsh angiosperms
in the SW Netherlands. Vegetatio 57: 143-152
Hahn MW, Höfle MG (2001) Grazing of protozoa and its effect on populations of
aquatic bacteria. FEMS Microbiol Ecol 35: 113-121
Hamerlynck O, Vanreusel A (1993) Mesacanthion diplechma (Nematoda:
Thoracostomopsidae), a link to higher trophic levels? J Mar Biol Ass UK 73: 453-456
Hedlund BP, Geiselbrecht AD, Bair T, Staley JT (1999) Polycyclic aromatic
hydrocarbon degradation by a new marine bacterium, Neptunomonas naphthovorans
gen. nov., sp. nov. Appl Environ Microbiol 65:251-259
Heip C (1975) On the significance of aggregation in some benthic marine
invertebrates. In Proceedings of the ninth European Marine Biology Symposium.
Barnes H (Ed) Aberdeen University Press, Aberdeen: 527-538
Heip C (1976) The spatial pattern of Cyprideis torosa (Jones, 1850) (Crustacea:
Ostracoda). J mar biol Ass UK 56:179-189
Heip C, Engels P (1977). Spatial segregation in copepod species from a brackish
water habitat. J exp mar Biol Ecol 26: 77-96
Heip C, Vincx M, Vranken G (1985) The ecology of marine nematodes. Oceanogr
Mar Biol Annu Rev London 23: 399-489
Hemminga MA, Buth GJC (1991) Decomposition in salt marsh ecosystems of the SW
Netherlands: the effects of biotic and abiotic factors. Vegetatio 92: 73-83
Hemminga MA, Klap VA, van Soelen J, Boon JJ (1993) Effect of salt marsh inundation
on estuarine particulate organic matter characteristics. Mar Ecol Prog Ser 99: 153-161

Cited literature
131
Hemminga MA, Klap VA, van Soelen J, de Leeuw J, Boon JJ (1992) Shifts in seston
characteristics after inundation of a European coastal salt marsh. Limnol Oceanogr 37:
1559-1564
Herman PMJ, Vranken G (1988) Studies of the life-history and energetics of marine
and brackish-water nematodes. II. Production, respiration and food uptake by
Monhystera disjuncta. Oecologia 77: 457-463
Hillebrand H (2003) Opposing effects of grazing and nutrients on diversity. Oikos
100: 592-600
Hippe H, Andreesen JR, Gottschalk G (1992) The genus Clostridium non-medical. In
The Prokaryotes. A Handbook on the Biology of Bacteria: Ecophysiology, Isolation,
Identification, Applications. Second edition. Balows A, Trüper HG, Dworkin M,
Harder W, Schleifer K-H (eds) Springer Verlag, New York: 1804-1866
Hogue EW (1982) Sediment disturbance and the spatial distributions of shallow water
meiobenthic nematodes on the open Oregon Coast. J Mar Res 40: 551-573
Hopper BE, Meyers SP (1967) Population studies on benthic nematodes within a
subtropical seagrass community. Mar Biol 1: 85-96
Howarth RW (1993) Microbial processes in salt-march sediments. In Aquatic
Microbiology. An Ecological Approach. Ford TE (ed). Blackwell Scientific
Publications, Boston: 239-259
Huettel RN (1986) Chemical communicators in nematodes. J Nematol 18: 3-8
Ilieva-Makulec K (2001) A comparative study of the life strategies of two bacterial-
feeding nematodes under laboratory conditions. II. Influence of the initial food level
on the population dynamics of Acrobeloides nanus (De Man 1880) Anderson 1968
and Dolichorhabditis dolichura (Schneider 1866) Andrássy 1983. Pol J Ecol 49: 123-
135
Ingham RE, Trofymow JA, Ingham ER, Coleman DC (1985) Interactions of bacteria,
fungi, and their nematode grazers; effects on nutrient cycling and plant growth. Ecol
Monogr 55: 119-140

Cited literature
132
Ives AR, Foufopoulos J, Klopfer ED, Klug JL, Palmer TM (1996) Bottle or big-scale
studies: how do we do ecology? Ecology 77: 681-685
Jennings JB, Deutsch A (1975) Occurrence and possible adaptive significance of b-
glucuronidase and arylamidase (Leucine aminopeptidase) activity in two species of
marine nematodes. Comparative Biochem Physiol 52A: 611-614
Jensen P (1987) Feeding ecology of free-living aquatic nematodes. Mar Ecol Prog Ser
35: 187-196
Jensen P (1996) Burrows of marine nematodes as centres for microbial growth.
Nematologica 42: 320-329
Johannes RE (1965) Influence of marine protozoa on nutrient regeneration. Limnol
Oceanogr 10: 434-442
Kampichler C, Bruckner A, Kandeler E (2001) Use of enclosed model systems in soil
ecology: a bias towards laboratory research. Soil Biol Biochem 33: 269-275
Kaufman TD, Bloom JR, Lukezic FL (1983) Effect of an extract from saprozoic
nematode-infested compost on the mycelial growth of Agaricus brunnescens. J
Nematol 15: 567-571
Kersters K, De Vos P, Gillis M, Swings J, Vandamme P, Stackebrandt E (2003)
Introduction to the Proteobacteria. http://141.150.157.117:8080/prokPUB/index.htm
Lawton JH (1994) What do species do in ecosystems? Oikos 71: 367-374
Lawton JH (1996) The ecotron facility at Silwood Park: the value of “big bottle”
experiments. Ecology 77:665-669
Lee JJ, Tietjen JH, Mastropaolo C, Rubin H (1977) Food quality and the
heterogeneous spatial distribution of meiofauna. Helgoländer Meeresunter 30: 272-
282

Cited literature
133
Levin LA, Boesch DF, Covich A, Dahm C, Erseus C, Ewel KC, Kneib RT, Moldenke
A, Palmer MA, Snelgrove P, Strayer D, Weslawski JM (2001) The function of marine
critical transition zones and the importance of sediment biodiversity. Ecosystems 4:
430-451
Lillebø AI, Flindt MR, Pardal MÂ, Marques JC (1999) The effect of macrofauna,
meiofauna and microfauna on the degradation of Spartina maritima detritus from a
salt marsh area. Acta Oecol 20: 249-258
Loreau M, Hector A (2001) Partitioning selection and complementarity in biodiversity
experiments. Nature 412: 72-76
Magurran AE (1988) Ecological diversity and its measurement. Princeton University
Press, Princeton, NJ
Massana R, Jürgens K (2003) Composition and population dynamics of planktonic
bacteria and bacterivorous flagellates in seawater chemostat cultures. Aquat Microbiol
Ecol 32: 11-22
McClung CR, Patrjiquin DG, Davis RE (1983) Campylobacter nitrofigilis sp. nov., a
nitrogen-fixing bacterium associated with roots of Spartina alterniflora Loisel. Int J
Syst Bacteriol 33: 605-612
Mikola J, Setälä H (1998) Relating species diversity to ecosystem functioning:
mechanistic backgrounds and experimental approach with a decomposer food web.
Oikos 83: 180-194
Mille-Lindblom C, Tranvik LJ (2003) Antagonism between bacteria and fungi on
decomposing aquatic plant litter. Microb Ecol 45: 173-182
Moens T, Vincx M (1997) Observations on the feeding ecology of estuarine
nematodes. J Mar Biol Assoc UK 77: 211-227
Moens T, Vincx M (1998) On the cultivation of free-living marine and estuarine
nematodes. Helgoländer Meeresunters 52: 115-139
Moens T, Vierstraete A, Vincx M (1996) Life strategies in two bacterivorous marine
nematodes: preliminary results. Mar Ecol 17: 509-518

Cited literature
134
Moens T, Verbeeck L, de Maeyer A, Swings J, Vincx M (1999) Selective attraction of
marine bacterivorous nematodes to their bacterial food. Mar Ecol Prog Ser 176: 165-
178
Montagna PA, Ruber E (1980) Decomposition of Spartina alterniflora in different
seasons and habitats of a northern Massachusetts salt marsh, and a comparison with
other Atlantic regions. Estuaries 3: 61-64
Moore JC, de Ruiter PC, Hunt HW, Coleman DC, Freckman DW (1996) Microcosms
and soil ecology: critical linkages between field studies and modelling food webs.
Ecology 77: 694-705
Mulder CPH, Uliassi DD, Doak DF (2001) Physical stress and diversity-productivity
relationships: The role of positive interactions. Proc Nat Acad Sci US 98: 6704-6708
Muylaert K, Van der Gucht K, Vloemans N, De Meester L, Gillis M, Vyverman W
(2002) Relationship between bacterial community composition and bottom-up versus
top-down variables in four eutrophic shallow lakes. Appl Environ Microbiol 68: 4740-
4750
Muyzer G (1999) DGGE/TGGE a method for identifying genes from natural
ecosystems. Curr Opin Microbiol 2: 317-322
Muyzer G, de Waal EC, Uitterlinden AG (1993) Profiling of complex microbial
populations by Denaturing Gradient Gel Electrophoresis analysis of polymerase chain
reaction-amplified genes coding for 16S rRNA. Appl Environ Microbiol 59: 695-700
Nannipieri P, Ascher J, Ceccherini MT, Landi L, Pietramellara G, Renella G (2003)
Microbial diversity and soil functions. Eur J Soil Sci 54: 655-670
Nehring S, Jensen P, Lorenzen S (1990) Tube-dwelling nematodes: tube construction
and possible ecological effects on sediment-water interfaces. Mar Ecol Prog Ser 64:
123-128
Newell SY (1996) Established and potential impacts of eukaryotic mycelial
decomposers in marine/terrestrial ecotones. J Exp Mar Biol Ecol 200: 187-206

Cited literature
135
Newell SY, Fallon RD, Miller JD (1989) Decomposition and microbial dynamics for
standing, naturally positioned leaves of the salt-marsh grass Spartina alterniflora.
Mar Biol 101: 471-481
Newell SY, Fallon RD, Cal Rodriguez RM, Groene LC (1985) Influence of rain, tidal
wetting and relative humidity on release of carbon dioxide by standing-dead salt-
marsh plants. Oecologia 68: 73-79
Nübel U, Garcia-Pichel F, Kühl M, Muyzer G (1999) Quantifying microbial diversity:
morphotypes, 16S rRNA genes, and carotenoids of oxygenic phototrophs in microbial
mats. App Env Microbiol 65: 422-430
Nübel U, Engelen B, Felske A, Snaidr J, Wieshuber A, Amann RI, Ludwig W,
Backhaus H (1996) Sequence heterogeneities of genes encoding 16S rRNAs in
Paenibacillus polymyxa detected by temperature gradient gel electrophoresis. J
Bacteriol 178: 5636-5643
O’Donnell AG, Seasman M, Macrae A, Waite I, Davies JT (2001) Plants and
fertilizers as drivers of change in microbial community structure and function in soils.
Plant and Soil 232: 135-145
Odum EP (1971) Fundamentals of ecology. Saunders College Publishing,
Philadelphia
Ólafsson E (1992) Small-scale spatial distribution of marine meiobenthos: the effects
of decaying macrofauna. Oecologia 90 : 37-42
Padgett DE, Courtney TH, Sizemore RK (1985) A technique for distinguishing
between bacterial and non-bacterial respiration in decomposing Spartina alterniflora.
Hydrobiologia 112: 113-119
Perkins EJ (1958) The food relationships of the microbenthos with particular
reference to that found at Whitstable, Kent. Ann Mag Nat Hist 13: 64-77

Cited literature
136
Pot B, Gillis M, De Ley J (1992) The genus Oceanospirillum. In The Prokaryotes. A
Handbook on the Biology of Bacteria: Ecophysiology, Isolation, Identification,
Applications. Second edition. Balows A, Trüper HG, Dworkin M, Harder W, Schleifer
K-H (eds) Springer Verlag, New York: 3230-3236
Powers SP (1998) Recruitment of soft-bottom benthos (benthic invertebrates,
encrusting community, infaunal community. Diss Abstr Int B Sci Eng 58: 5760
Reice SE, Stiven AE (1983) Environmental patchiness, litter decomposition and
associated faunal patterns in a Spartina alterniflora marsh. Est Coast Shelf Sci 16:
559-571
Reichenbach H (1992) The order Cytophagales. In The Prokaryotes. A Handbook on
the Biology of Bacteria: Ecophysiology, Isolation, Identification, Applications.
Second edition. Balows A, Trüper HG, Dworkin M, Harder W, Schleifer K-H (eds)
Springer Verlag, New York: 3631-3675
Rheinheimer G (1992) Aquatic Microbiology, 4rd ed. John Wiley & Sons, UK
Riemann F, Helmke E (2002) Symbiotic relations of sediment-agglutinating
nematodes and bacteria in detrital habitats: the enzyme-sharing concept. Mar Ecol 23:
93-113
Riemann F, Schrage M (1978) The mucus-trap hypothesis on feeding of aquatic
nematodes and implications for biodegradation and sediment texture. Oecologia 34:
75-88
Rieper-Kirchner M (1989) Microbial degradation of North Sea macroalgae: field and
laboratory studies. Bot Mar 32: 241-252
Romeyn K, Bouwman LA (1983) Food selection and consumption by estuarine
nematodes. Hydrobiol Bull 17: 103-109
Rønn R, McCaig AE, Griffiths BS, Prosser JI (2002) Impact of protozoan grazing on
bacterial community structure in soil microcosms. Appl Environ Microbiol 68: 6094-
6105

Cited literature
137
Ruby EG (1992) The genus Bdellovibrio. In The Prokaryotes. A Handbook on the
Biology of Bacteria: Ecophysiology, Isolation, Identification, Applications. Second
edition. Balows A, Trüper HG, Dworkin M, Harder W, Schleifer K-H (eds) Springer
Verlag, New York: 3400-3415
Schauer M, Massana R, Pedrós-Alió C (2000) Spatial differences in bacterioplankton
composition along the Catalan coast (NW Mediterranean) assessed by molecular
fingerprinting. FEMS Microbiol Ecol 33: 51-59
Schläpfer F, Schmid B, Seidl I (1999) Expert estimates about effects of biodiversity on
ecosystem processes and services. Oikos 84: 346-352
Service SK, Bell SB (1987) Density-influenced active dispersal of harpacticoid
copepods. J Exp Mar Biol Ecol 46: 49-62
Shannon CE, Weaver W (1949) The mathematical theory of communication.
University of Illinois Press, Urbana
Sherman KM, Coull BC (1980) The response of meiofauna to sediment disturbance. J
Exp Mar Biol Ecol 46: 59-71
Silvertown J, Dodd ME, McConway K, Potts J, Crawley M (1994) Rainfall, biomass
variation, and community composition in the Park Grass Experiment. Ecology 75:
2430-2437
Sohlenius B (1968) Influence of microorganisms and temperature upon some
rhabditid nematodes. Pedobiologia 8: 137-145
Steyaert M, Vanaverbeke J, Vanreusel A, Barranguet C, Lucas C, Vincx M (2003)
The importance of fine-scale, vertical profiles in characterising nematode community
structure. Estuar Coast Shelf Sci 58: 353-366
Sun B, Fleeger JW, Carney RS (1993) Sediment microtopography and the small-scale
spatial distribution of meiofauna. J Exp Mar Biol Ecol 167: 73-90
Tenore KR, Tietjen JH, Lee JJ (1977) Effect of meiofauna on incorporation of aged
eelgrass, Zostera marina, detritus by the polychaete Nephthys incisa. J Fish Res Bd
Canada 34: 563-567

Cited literature
138
Tietjen JH (1980) Microbial-meiofaunal interrelationships: a review. In Aquatic
Microbial Ecology. Colwell RR, Foster J (eds) University of Maryland Press,
Maryland: 335-338
Tietjen JH, Lee JJ (1973) Life-history and feeding habits of the marine nematode
Chromadora macrolaimoids Steiner. Oecologia 12: 303-314
Tietjen JH, Lee JJ (1977) Feeding behaviour of marine nematodes. In Ecology of
marine benthos. Coull BC (ed) Univ. South Carolina Press, Columbia: 22-36
Tilman D, Downing JA (1994) Biodiversity and stability in grasslands. Nature 367:
363-365
Tilman D, Wedin D, Knops J (1996) Productivity and sustainability influenced by
biodiversity in grassland ecosystems. Nature 379: 718-720
Trotter DB, Webster JM (1984) Feeding preferences and seasonality of free-living
marine nematodes inhabiting the kelp Macrocystis integrifolia. Mar Ecol Prog Ser 14:
151-157
Troussellier M, Schafer H, Batailler N, Bernard L, Courties C, Lebaron P, Muyzer G,
Servais P, Vives-Rego J (2002) Bacterial activity and genetic richness along an
estuarine gradient (Rhone River plume, France) Aquat Microb Ecol 28: 13-14
Ullberg J, Ólafsson E (2003) Free-living marine nematodes actively choose habitat
when descending from the water column. Mar Ecol Prog Ser 260: 141-149
Valaeys T, Topp E, Muyzer G, Macheret V, Laguerre G, Rigaud A, Soulas G (1997)
Evaluation of denaturing gradient gel electrophoresis in the detection of 16S rDNA
sequence variation in rhizobia and methanotrophs. FEMS Microbiol Ecol 24: 279-285
Valiela I (1995) Marine Ecological Processes. Springer-Verlag, New York: 305-324
Van der Gucht K, Sabbe K, De Meester L, Vloemans N, Zwart G, Gillis M,
Vyverman W (2001) Contrasting bacterioplankton community composition and
seasonal dynamics in two neighbouring hypertrophic freshwater lakes. Environ
Microbiol 3: 680-690

Cited literature
139
Van Hannen EJ, Veninga M, Bloem J, Gons HJ, Laanbroek HJ (1999) Genetic
changes in the bacterial community structure associated with protistan grazers. Arch
Hydrobiol 145: 25-38
Venette RC, Ferris H (1998) Influence of bacterial type and density on population
growth of bacterial-feeding nematodes. Soil Biol Biochem 30: 949-960
Verhoef HA (1996) The role of soil microcosms in the study of ecosystem processes.
Ecology 77: 685-690
von Thun W (1966) Eine Methode zur Kultivierung der Mikrofauna. Veröffn Inst
Meeresforsch Bremerh 2: 277-280
von Thun W (1968) Autökolgische Untersuchungen an freilebenden Nematoden des
Brackwassers. PhD thesis, Universität Kiel
Walker BH (1992) Biodiversity and ecological redundancy. Conserv Biol 6: 18-23
Walker BH (1995) Conserving biological diversity through ecosystem resilience.
Conserv Biol 9: 747- 752
Wardle DA, Bonner KI, Nicholson KS (1997) Biodiversity and plant litter:
experimental evidence which does not support the view that enhanced species
richness improves ecosystem function. Oikos 79: 249-258
Warwick RM (1981) Survival strategies of meiofauna. In Feeding and survival
strategies of estuarine organisms. Jones NV, Wolff WJ (eds) Plenum Press, New
York: 39-52
Warwick RM (1987) Meiofauna: their role in marine detrital systems. Detritus and
microbial ecology in aquaculture. ICLARM conference proceedings 14: 282-295
Warwick RM, Carr MR, Clarke KR, Gee JM, Green RH (1988) A mesocosm
experiment on the effects of hydrocarbon and copper pollution on a sublittoral soft-
sediment meiobenthic community. Mar Ecol Prog Ser 46: 181-191

Cited literature
140
Wetzel MA, Jensen P, Giere O (1995) Oxygen/sulfide regime and nematode fauna
associated with Arenicola marina burrows: new insights in the thiobios case. Mar Biol
124: 301-312
White DS, Howes BL (1994) Nitrogen incorporation into decomposing litter of
Spartina alterniflora. Limnol Oceanogr 39: 133-140
Wieser W (1953) Die Beziehung zwischen Mundhöhlengestalt, Ernährungsweise und
Vorkommen bei freilebenden marinen Nematoden. Eine ökologische-morphologische
Studie. Ark Zoo. 4: 439-484
Wohl DL, McArthur JV (2001) Aquatic actinomycete-fungal interactions and their
effects on organic matter decomposition: a microcosm study. Microb Ecol 42: 446-
457
Zöllner E, Santer B, Boersma M, Hoppe HG, Jürgens K (2003) Cascading predation
effects of Daphnia and copepods on microbial food web components. Freshwater
Biol 48: 2174-2193
Zwart G, Huismans R, van Agterveld MP, Van de Peer Y, De Rijk P, Eenhoorn H,
Muyzer G, van Hannen EJ, Gons HJ, Laanbroek HJ (1998) Divergent members of the
bacterial division Verrucomicrobiales in a temperate freshwater lake. FEMS
Microbiol Ecol 25: 159-169





























