Embed Size (px)
Citation preview

1
UNIVERSIDADE FEDERAL DE MINAS GERAIS
INSTITUTO DE CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA,
CONSERVAÇÃO E MANEJO DA VIDA SILVESTRE
ANA CAROLINA VIEIRA PIRES
DISTRIBUIÇÃO DE BORBOLETAS
NECTARÍVORAS AO LONGO DO GRADIENTE
ALTITUDINAL DE UMA MONTANHA
TROPICAL: PADRÕES E MECANISMOS
Belo Horizonte
2014

2
ANA CAROLINA VIEIRA PIRES
DISTRIBUIÇÃO DE BORBOLETAS AO LONGO DO GRADIENTE
ALTITUDINAL DE UMA MONTANHA TROPICAL: PADRÕES E
MECANISMOS
Dissertação apresentada ao Programa de Pós-
Graduação em Ecologia, Conservação e Manejo da
Vida Silvestre do Instituto de Ciências Biológicas da
Universidade Federal de Minas Gerais, como
requisito para obtenção do título de Mestre em
Ecologia.
Orientador: Geraldo Wilson Fernandes
Belo Horizonte
2014

3
“A natureza não faz milagres, faz revelações.”
“A natureza são duas.
Uma,
tal qual se sabe a si mesma.
Outra, a que vemos. Mas vemos?
Ou é a ilusão das coisas?”
(Carlos Drummond de Andrade)

4
AGRADECIMENTOS
Nunca pensei que trabalharia com borboletas, até entrar no LEEB. E de repente
vi que muita coisa na minha vida estava relacionada a essas pequenas criaturas. E só
tenho a agradecer ao meu orientador Geraldo Wilson Fernandes, à Marina Beirão e
Yumi Oki por terem aberto as portas e me dado essa oportunidade de trabalho e
aprendizado.
Agradeço aos meus pais, Ricardo e Christiane, pelo apoio durante essa fase. Aos
meus irmãos, Rapha e Flávia, pelo companheirismo de sempre, até me levando e
buscando na faculdade com minhas tralhas.
Um “muito obrigada” mais que especial às pessoas que passaram os perrengues
comigo durante as coletas. À Fabiola, meu braço direito nesse projeto, que foi em quase
todas as campanhas. À Irene, Geanne, Isabela, Milton, Oripe, Jéssica e Vanessa, equipe
Lepidoptera em ação! E àqueles que foram com a maior boa vontade do mundo para
ajudar: Beul, Ju, Vanessa, Luiza, Caio e Warlei.
Agradeço às pessoas que fizeram parte de mais de uma etapa desse trabalho.
Renato por me apoiar sempre, me ajudar em campo e com as minhas tabelas. Marina
por me ajudar na fase crítica da estatística e da escrita. Daniel que me ajudou com
algumas análises e fez comentários muito pertinentes, sempre me empolgando com o
trabalho.
E não me esquecendo das pessoas queridas do LEEB: Thaíse, Leandra, Yumi,
Marcel, Fernando, Cotonete e Ana Carolina. Agradeço pelas conversas e cafés que
fizeram o dia a dia muito melhor.
Aos amigos de sempre, que foram também essenciais, Carla pela versão em
inglês do resumo, Gudryan pela paciência e capricho em fazer os gráficos, Camila pelas
conversas sobre a vida, Daniel que me ajudou diversas vezes nas minhas dúvidas
estatísticas. Ao professor Fred que me ajudou em dúvidas sobre os modelos.
Agradeço também aos profissionais que me ajudaram na identificação dos
espécimes: prof. Dr. Olaf H. H. Mielke, Msc Diego Rodrigo Dolibaina e Dr. Fernando
Dias. E à todos os integrantes do Laboratório de Estudos de Lepidoptera Neotropical da
UFPR pela ótima recepção.
Aos professores Programa de Pós-Graduação em Ecologia, Manejo e Vida
Silvestre pelo conhecimento transmitido. Aos funcionários da secretaria, Fred e Cris,
por estarem sempre prontos a auxiliar nos tramites burocráticos e trabalhando

5
eficientemente agilizando documentos e divulgando informações. Aos funcionários do
setor de transporte, superintendente Vinícius, motoristas Seu Luiz, Seu Messias, Marlon
e , pela atenção e prestatividade no agendamento dos carros e no campo.
À equipe do Parque Nacional da Serra do Cipó pela disponibilização do
alojamento e logística para a realização do trabalho. E aos demais parceiros do PELD:
ao Rogério da Pousada Serra Morena que sempre esteve de braços abertos para nos
receber em sua propriedade, à Reserva Vellozia pelo alojamento e autorização para
trabalhar dentro de sua área, à Cedro Cachoeira Indústria Têxtil, Pousada Pedra do
Elefante pela disponibilização de sua área para o projeto.
Ao CNPq pela concessão da bolsa de mestrado e pelo financiamento dos
projetos “Bioindicadores para avaliação do efeito de mudanças climáticas globais em
montanhas tropicais” (PELD - Processo 403781/2012-4) e do Programa de Pesquisa em
Biodiversidade (PPBIO - Processo 457519/2012-6). À Fapemig pelo financiamento do
projeto “Diagnóstico biológico das alterações climáticas globais em montanhas
tropicais” (Edital Universal - Proc. APQ-02158-10).
O empenho e a ajuda de todos foram essenciais para a realização desse trabalho.
O companheirismo e a amizade foram essenciais para que eu continuasse nessa jornada.
À todos, muito obrigada.

6
Artigo formatado segundo as normas da revista Diversity and Distributions
(Tabelas e figuras foram colocadas ao longo do texto para facilitar a compreensão)
DISTRIBUIÇÃO DE BORBOLETAS AO LONGO DO GRADIENTE ALTITUDINAL
DE UMA MONTANHA TROPICAL: PADRÕES E MECANISMOS

7
RESUMO
Objetivo
A influência de forças geofísicas sobre a distribuição das espécies é uma das questões
mais antigas da ecologia e ambientes montanhosos são o cenário ideal para testá-las. O
objetivo desse estudo foi fornecer um panorama de distribuição espacial de borboletas
nectarívoras em uma montanha tropical, bem como indicar os mecanismos responsáveis
pela variação da diversidade ao longo do gradiente altitudinal.
Local
Serra do Cipó, Minas Gerais, Brasil.
Métodos
Ao longo do gradiente de altitude foram demarcadas sete áreas, distribuídos entre 800 e
1400m de altitude. As borboletas encontradas durante o período de amostragem foram
capturadas com rede entomológica. Foram realizadas oito campanhas em dois anos de
estudo. Para verificar a variação da diversidade e composição de borboletas ao longo do
gradiente foram construídos GLMs e também analisados através da co-inércia. O
turnover de espécies do gradiente foi analisado através da partição multiplicativa da
diversidade e a identificação de espécies indicadoras foi feita utilizando a análise de
IndVal.
Resultados
A diversidade de borboletas decresceu conforme o aumento da altitude e a temperatura
é o principal fator que determinam dessa variação. A composição de espécies mudou
com a altitude, porém foi similar em altitudes intermediárias. Há pouca troca de
espécies num mesmo local, não havendo uma relação clara entre o turnover de espécies
e a altitude.
Principais conclusões
Esse estudo mostrou a importância da temperatura sendo o principal fator que influencia
a variação da diversidade de borboletas. Frente a um cenário de mudanças climáticas, é
possível que várias espécies desta região possam entrar em alguma categoria de
vulnerabilidade ou mesmo tornar-se extintas em um futuro próximo. Esse estudo é o
primeiro passo para se entender os processos por trás da distribuição de espécies em
ambientes tropicais montanhosos.
Palavras-chave
Campo rupestre, diversidade de espécies, gradiente altitudinal, padrões biogeográficos

8
ABSTRACT
ALTITUDINAL BUTTERFLY DISTRIBUTION IN A TROPICAL MOUNTAIN: PATTERNS AND
MECHANISMS
Aim
The influence of geophysics upon the distribution of species is one of oldest
questionings inside ecology, and mountainous environments are the ideal scenery to test
this influence. The aim of this study was to provide an overview on the space-time
distribution of the nectarivorous butterflies in a tropical mountain, as well as indicate
the mechanisms responsible for the diversity’s variation along the altitudinal gradient.
Location
Serra do Cipó, Minas Gerais, Brazil
Methods
Data for this study was obtained by diving the altitudinal gradient into seven areas
distributed between 800 and 1400m of height. The butterflies found during the sampling
period were catch with entomological nets. Eight journeys were made in the period of
two years’ time. General Linear Model (GLM) and co-inertia analysis were used in
order to verify the butterflies’ diversity and composition variation. The turnover of
species’gradient was analysed through multiplicative partition and the identification of
species through the IndVal analysis.
Results
The diversity of butterflies diminished according to the increasement of hight and
temperature is the main element behind this variation. The species’ composition
changed with the altitude, but in the intermediate heights they were similar. There is a
small trade of species at the same place, however there is no clear relation between the
species ‘turnover and height.
Main conclusions
On the basis of the results of this research, it can be seen how important temperature is,
being the main influencer in the variation of the butterflies’ diversity. Facing a scenery
of climate change, it is possible to see that a great number of species from this areas
could be endangered or even become extinct in a near future. This study is the first step
to understand the processes behind the species’ distribution in tropical-mountainous
environments.
Keywords

9
Altitudinal gradient, biogeographic patterns, diversity, rupestrian grasslands, species
distribution

10
INTRODUÇÃO
Ambientes montanhosos proporcionam à ciência um cenário singular onde as
mudanças na paisagem ao longo do gradiente altitudinal fornecem ao pesquisador um
experimento natural para o teste de uma das questões mais antigas da ecologia: o da
influência das forças geofísicas sobre a distribuição das espécies (Fernandes & Price
1988, Körner 2007). Nessas regiões as espécies conseguem se deslocar entre ambientes
de climas distintos em distâncias relativamente curtas (Körner 2004, Wilson et al.
2007), favorecendo os estudos observacionais e experimentais. Além disto, as
montanhas são extremamente importantes devido à superfície ocupada (ca. 25% de toda
superfície terrestre) (Kapos et al. 2000) e pelo fato de que mais da metade de toda a
população humana é suprida por recursos provenientes de áreas montanhosas cuja
integridade funcional é mantida pela flora, fauna e microrganismos especializados,
aninhados em uma grande variedade de microhabitats (Körner 2004).
Embora amplamente aceito que a diversidade de plantas e animais é maior em
terras baixas do que em topos de morro (e.g. Pianka 1966, Fernandes & Price 1991), o
decréscimo na diversidade com o aumento da altitude não é necessariamente
monotônico. Após fazer uma revisão utilizando 97 trabalhos, englobando estudos com
plantas, vertebrados e invertebrados, Rahbek (1995) concluiu que o padrão de
distribuição mais encontrado era em formato de corcova, ou seja, a maior diversidade
está em elevações intermediárias (Janzen et al. 1976, Wolda 1987, Rahbek 1995).
Porém esse padrão é dependente do taxon e do ambiente estudado (Grytnes & McCain
2007). Devido à relação entre produtividade, temperatura e pluviosidade, organismos
que habitam ambientes montanhosos áridos ou sazonalmente secos podem apresentar
pico de diversidade em altitudes intermediárias, enquanto que a diversidade em
ambientes montanhosos úmidos tende a decrescer monotonicamente com o aumento da
elevação (Grytnes & McCain 2007, McCain 2007).
Montanhas tropicais abrigam uma fauna diversa, com grande variedade de
espécies co-existindo dentro de zonas climáticas ou tipos de habitats. Nestas montanhas
há uma alta mudança (turnover) de espécies entre essas zonas (Jankowski et al. 2009).
A diversidade beta (β) descreve a variação na composição de espécies observadas ao
comparar unidades de amostragem (Cáceres et al. 2012). Quantificar a diversidade β ao
longo de gradientes de altitude pode ajudar na criação de estratégias para proteção da
diversidade biológica, uma vez que, se a diversidade β é alta, o incremento de pequenas
áreas pode ajudar na conservação de várias espécies, preservando os mecanismos que

11
geram essa diversidade (Jankowski et al. 2009). É esperado que a diversidade β diminua
com o aumento da altitude, já que tanto a riqueza local (α) quanto a regional (γ)
decrescem (Kraft et al. 2011) e também porque o habitat tende a ficar menos
heterogêneo em altitudes mais elevadas; consequentemente conduzindo a uma
diminuição na diversidade de espécies (Tews et al. 2004, Stein et al. 2014).
As borboletas são extremamente sensíveis às mudanças na composição e
estrutura da vegetação além de estarem envolvidas em muitas interações ecológicas
dentro dos ecossistemas (Sawchik et al. 2003). Desempenham um papel fundamental na
polinização em grandes altitudes, chegando a superar a polinização por abelhas em
zonas mais elevadas (Arroyo et al. 1982) e representam cerca de 75% de todos os
visitantes de flores da família Asteraceae (Mani & Saravanan 1999), uma das famílias
mais comuns nos ambientes campestres montanhosos do Brasil (veja Giulietti et al.
1987).
A temperatura, umidade, pluviosidade e velocidade do vento são fatores
climáticos que, reconhecidamente, afetam a comunidade de Lepidoptera (Brown &
Freitas 2000, Brehm et al 2003, Stefanescu et al 2004, Bhardwaj et al. 2012).A
sazonalidade climática e a variação espacial e temporal dos recursos alimentares são
fatores cruciais que determinam a diversidade, composição e dispersão de comunidades
de lepidópteros (Hill 1992, Darrault & Schlindwein 2002, Abrahamczik et al. 2011,
Baguette et al. 2011). As borboletas são insetos amplamente distribuídos no globo e
representadas por seis subfamílias, que possuem características bem distintas umas das
outras, seja no habitat onde ocorrem, no ciclo de vida, no comportamento ou
especificidade de plantas hospedeiras (Brown Jr 1992, Ferrer-Paris et al. 2013). Por
possuírem relações tão estreitas com o habitat em que vivem, podem ser considerados
bons indicadores ambientais, servindo como ferramenta para estudos de conservação
(New et al. 1995).
A Cadeia do Espinhaço compreende um grupo de serras no sudeste brasileiro
cuja extensão total é de 1100 km e possui altitudes entre 800 e 1700 m. Localizada na
parte sul da Cadeia do Espinhaço encontra-se a Serra do Cipó, onde predominam os
campos rupestres. Devido a particularidades edáficas, topográficas e climáticas, a flora
desse ecossistema apresenta várias peculiaridades, como os altos graus de convergência
morfológica, diversidade e endemismo (Negreiros et al. 2014, Echternacht et al. 2011).
Essa formação vegetal montanhosa única está ameaçada (Costa et al. 1998, Ministério
do Meio Ambiente 1999) devido à expansão urbana e agropecuária, queimadas, plantio

12
de eucalipto, retirada de plantas ornamentais, mineração e construção e pavimentação
de estradas (Barbosa et al. 2010, Ribeiro & Freitas 2010, Negreiros et al. 2011).
Entre os animais conhecidos da Serra do Cipó apenas 32,6% (i.e. 251 espécies)
são invertebrados, o que claramente ressalta a carência de levantamento de espécies
desse grupo (Madeira et al. 2008). Entre a fauna de Lepidoptera ameaçada, a única
espécie que se encontra nos domínios da Serra do Cipó é Nirodia belphegor
(Riodinidae) (Brown 1993, Machado et al. 2008, Nascimento & Campos 2011),
refletindo a escassez de estudos específicos sobre o grupo nesta região.
Uma das hipóteses mais citadas para explicar os padrões encontrados em
gradientes altitudinais é a que considera os fatores climáticos responsáveis pela variação
da diversidade. Em relação ao grupo estudado, a tolerância climática pode restringir a
quantidade de espécies que podem sobreviver em diferentes elevações (Grytness &
McCain 2007). No presente trabalho buscamos entender o padrão de distribuição
espacial das espécies de borboletas nectarívoras em um ambiente montanhoso tropical,
testando a hipótese de que fatores ambientais (bióticos e abióticos) relacionados com o
gradiente altitudinal são responsáveis pela variação da diversidade, a riqueza,
abundância e composição de borboletas nectarívoras. Também testamos a hipótese de a
comunidade de borboletas tende a ficar mais semelhante entre as trilhas nas altitudes
maiores do que em baixas altitudes.
MÉTODOS
Área de estudo
As amostragens foram realizadas em áreas de cerrado e campo rupestre na Serra
do Cipó, localizada na porção sul da Cadeia do Espinhaço entre as coordenadas 19°15’
e 19°30’ S e 43°55’ W (Figura 1). A Serra do Cipó apresenta um clima do tipo Cwb de
Köppen (clima mesotérmico com verões brandos e estação chuvosa no verão), com
temperaturas médias entre 17,4 e 19,8ºC. A precipitação anual da região está em torno
de 1500 mm, com um período chuvoso de novembro a janeiro, um período de transição
pós-chuva de fevereiro a abril, uma época seca de maio a setembro e um período de
transição pós-seca apenas em outubro (Madeira & Fernandes 1999). O solo da Serra do
Cipó é arenoso, raso, com baixa capacidade de retenção de água, extremamente
oligotrófico e com grande concentração de alumínio (Negreiros et al. 2008, 2011). As
áreas de estudo localizam-se no Parque Nacional da Serra do Cipó e APA Morro da
Pedreira, no município de Cardeal Mota, Minas Gerais.

13
Figura 1 – Mapa da Serra do Cipó, MG. Os pontos brancos representam os locais onde
estão instaladas as torres meteorológicas (mapa produzido por F. Carvalho).
Desenho amostral e dados climáticos
Ao longo do gradiente de altitude na Serra do Cipó foram instaladas torres de
monitoramento meteorológico equipadas com o data-logger Onset HOBO® U30. As
torres foram distribuídas entre 800 e 1400m de altitude, uma a cada 100 m de altitude e
distanciadas geograficamente em pelo menos 2km (Figura 1). Os seguintes parâmetros
climáticos foram extraídos: temperatura e umidade do ar, velocidade do vento e
radiação. Para as análises foram utilizadas as médias de cada uma dessas variáveis dos
meses amostrados.
Amostragem
Para a coleta de borboletas foram demarcados, em cada uma das sete altitudes,
três transectos de 250m espaçados em 500m, compreendendo assim uma unidade
amostral. Cada transecto foi percorrido por dois amostradores durante duas horas e os
indivíduos encontrados foram capturados com rede entomológica. Foram estabelecidos
três horários para a captura, iniciando duas horas após o nascer do sol (às 8:00 h) e
terminando até duas horas antes do pôr do sol (às 16:00 h). As coletas aconteceram nos

14
meses de janeiro, abril, julho e outubro de 2012 e janeiro, maio, agosto e outubro de
2013, contemplando assim períodos distintos do ano. Apenas borboletas nectarívoras
foram utilizadas nesse estudo, excluindo todas as espécies frugívoras coletadas.
Os indivíduos capturados foram retirados individualmente, imediatamente
sacrificados e colocados em envelopes de papel vegetal. Cada envelope conteve um
espécime, sendo registrada a data, hora da coleta e ponto amostral. No laboratório os
indivíduos foram montados com as asas esticadas, fixados com alfinete entomológico e
posteriormente identificados até o menor nível taxonômico possível com auxílio de
guias de campo e quando necessários foram enviados a especialistas. Os dados de
diversidade vegetal foram extraídos de Mota et al. 2014. As espécies de lepidópteros
foram identificadas por O. H. H. Mielke e M. Duarte.
Análises estatísticas
Todas as análises foram feitas utilizando o software R (R Development Core
Team 2013), com exceção apenas da análise de cluster, onde foi utilizado o software
Past. Devido à natureza dos dados, as análises dos modelos lineares generalizados
foram feitas utilizando as distribuições de Poisson e Quasipoisson.
Para determinar a representatividade da amostragem calculamos uma curva de
acumulação de espécies. A coleta estratificada em três transectos em cada um dos
pontos amostrais permitiu comparar a riqueza de espécies e abundância das borboletas
entre altitudes e com os fatores ambientais, com o uso de modelos lineares
generalizados (GLM). Para construção dos GLMs utilizamos a riqueza e a abundância
de borboletas acumulada por ponto e como variáveis explicativas utilizamos a altitude,
os parâmetros abióticos fornecidos pelas torres meteorológicas (temperatura, umidade,
velocidade do vento, radiação solar) e variáveis bióticas obtidas de Mota et al. 2014
(riqueza, abundância e altura das plantas). Todas as variáveis foram testadas no GLM,
mas as que não tiveram significância estatística na variação da riqueza ou da abundância
de borboletas foram retiradas uma a uma, a fim de simplificar o modelo, tornando-o
mais parcimonioso.
Foi feita uma análise de co-inérica para determinar a relação entre as variáveis
ambientais (bióticas e abióticas) e a comunidade de borboletas. Essa análise é utilizada
para determinar se existe uma co-estrutura entre duas matrizes de dados, analisando
simultaneamente as matrizes. A máxima covariância entre as matrizes significa a
máxima correlação e simultaneamente o máximo desvio padrão entre os escores

15
ambientais e os escores faunísticos (Dolédec & Chessel 1994). A análise de co-inércia
realizada para esse estudo foi baseada em uma PCA centrada utilizando a incidência das
espécies de borboletas (127 espécies) e uma PCA (sete variáveis ambientais) em sete
locais (sete pontos de altitude). Foi feito um teste de Monte-Carlo para apurar a
significância da co-inércia. Foi utilizado o pacote ade4 (Oksanen et al. 2013)para a
realização dessa análise.
Para testar a heterogeneidade da comunidade foi feita a partição multiplicativa
da diversidade, uma abordagem mais apropriada para o estudo em questão, gerando um
valor de β independente do valor de α (Baselga 2010). Jost (2007) propôs a utilização
dos números equivalentes para o cálculo dos componentes de diversidade e demonstrou
que seriam mais intuitivos e refletiriam a verdadeira diversidade, enquanto os índices
clássicos não. Assim, utilizando os números equivalentes a partir da frequência das
espécies, a diversidade beta foi calculada segundo a fórmula de Whittaker (1960). Para
verificar se a comunidade era heterogênea ou homogênea, diferindo do acaso, foram
criados modelos nulos. Para acessar a significância de cada componente da diversidade,
a frequência de cada espécie foi aleatorizada dentro de cada altitude, preservando assim
o seu tamanho. Esse processo foi repetido 10000 vezes para obter a distribuição nula de
cada estimativa de diversidade em cada local. Cada conjunto de dados randomizados foi
particionado em seus componentes, e a significância estatística (a probabilidade de obter
um valor maior ou menor que a diversidade observada) de cada componente foi obtida a
partir da proporção de valores nulos maiores ou menores que o valor observado. Essa
análise foi realizada através do procedimento “multipart” do pacote vegan (Oksanen et
al. 2013).
RESULTADOS
A comunidade de borboletas
Foram registrados 976 indivíduos de borboletas nectarívoras pertencentes a 127
espécies e treze subfamílias durante os dois anos de estudo. Seis famílias de borboletas
nectarívoras diurnas foram amostradas, sendo identificadas 47 espécies de Hesperiidae,
33 de Lycaenidae, 14 de Nymphalidae, 5 de Papilionidae, 12 de Pieridae e 16 de
Riodinidae (Tabela 1). A espécie mais comum foi Eurema elathea (Pieridae) com 125
indivíduos coletados, seguida por Hemiargus hanno (Lycaenidae) com 122 indivíduos
coletados e Pyrisitia nise (Pieridae) com 75 indivíduos coletados. Estes indivíduos
juntos somaram 33% do total das borboletas amostradas. Foram registradas 52 (40,9%)

16
espécies com apenas um indivíduo (singleton). Apenas quatro espécies ocorreram em
todas as sete áreas: Hemiargus hanno, Junonia evarete (Nymphalidae), Eurema elathea
e Phoebis sennae (Pieridae). Uma espécie nova, Pyrrhopyge sp.nv. (Hesperiidae), foi
encontrada na altitude de 800m, dentro do Parque Nacional da Serra do Cipó (O. H. H.
Mielke, com. pess.).
Tabela 1 – Espécies de borboletas nectarívoras, sua distribuição e frequência no
gradiente altitudinal da Serra do Cipó, Brasil. A letra S representa a quantidade de
espécies de cada nível taxonômico.
Altitude (m)
800 900 1000 1100 1200 1300 1400 TotalHesperiidae (S= 47)
Eudaminae (S= 14) Chioides catillus 1 4 5 1 11Cogia azila 3 3Cogia calchas 2 2 1 1 6Cogia hassan evansi 4 3 2 3 7 1 20Epargyreus sp. 1 1 2Phocides polybius 2 2Typhedanus undulatus 1 1 2Udranomia spitzi 4 4 1 4 1 14Urbanus carmelita barra 2 2Urbanus cindra 4 2 2 3 11Urbanus dorantes 1 1 2Urbanus evenus 3 2 5Urbanus procne 1 1 2Urbanus simplicius 1 1
Hesperiinae (S= 19) Chalcone briquenydan 1 1 2Copaeodes jean favor 1 1 2Hylephila phyleus 3 2 5Hylephila sp. 1 1Lerema caraca 1 1Lerema veadeira 1 1Panoquina bola 2 1 1 4Panoquina peraea 7 7Polites vibex catilina 1 1 2Pompeius dares 1 1Quasimellana mielkei 1 1 5 7Synale metella 1 1Thespieus homochromus 5 5Vehilius inca 1 1 2

17
Vettius lucretius 1 1Vidius nostra nostra 1 2 3Vidius similis 1 1 2Wallengrenia premnas 1 1Xeniades chalestra 1 1
Pyrginae (S= 14) Chiomara basigutta 5 3 3 4 9 3 27Elbella hegesipe 2 2Gesta gesta 1 1Gesta heteropterus 1 1Gorgythion sp. 3 1 4Heliopetes arsalte 1 1 4 2 8Heliopetes macaira orbigera 1 3 1 1 1 7Heliopetes omrina 5 1 4 3 1 2 16Pyrgus orcus 12 3 6 7 2 30Pyrrhopyge sp. nv. 1 1Sophista latifasciata 1 2 3Timochreon doria 1 1Viola violella 2 2 4Zopyrion evenor 4 1 5
Lycaenidae (S= 33) Polyommatinae (S= 2)
Hemiargus hanno 61 23 13 21 2 1 1 122Leptotes cassius 5 15 3 6 2 1 32
Theclinae (S= 31) Arcas ducalis 1 1Cyanophrys herodotus 1 1Evenus regalis 1 1Pseudolycaena marsyas 1 1Strymon cestri 1 1Strymon mulucha 1 1Theritas triquetra 1 1Theclinae 1 1 1Theclinae 2 1 1 2Theclinae 3 1 1Theclinae 4 1 1Theclinae 5 1 1Theclinae 6 1 1Theclinae 7 1 1Theclinae 8 2 3 5Theclinae 9 1 1 2Theclinae 10 4 5 9Theclinae 11 1 2 3Theclinae 12 3 2 2 1 2 10Theclinae 13 1 1 2Theclinae 14 1 3 3

18
Theclinae 15 1 1Theclinae 16 1 1 2 4Theclinae 17 2 2Theclinae 18 1 1Theclinae 19 1 1Theclinae 20 1 1Theclinae 21 1 1Theclinae 22 1 1 2Theclinae 23 2 2Theclinae 24 2 2
Nymphalidae (S= 14) Danainae (S= 2)
Danaus gilippus 1 3 5 9Danaus plexippus 1 1
Heliconiinae (S= 3) Heliconius besckei 1 1Heliconius erato 2 1 1 3Heliconius ethilla 1 1
Ithomiinae (S= 1) Placidina euryanassa 1 1
Nymphalinae (S= 8) Anartia jatrophae 2 2Eresia lansdorfi 1 1Euptoieta hegesia 2 1 3Junonia evarete 7 3 6 12 5 8 2 43Tegosa claudina 1 1 2Vanessa braziliensis 11 11Vanessa myrinna 5 2 7Vanessa virginiensis 1 1
Papilionidae (S= 5) Papilioninae (S= 5)
Battus polydamas 1 1Heraclides thoas 1 1Parides anchises 1 1Parides bunichus 6 17 3 3 8 36Parides neophilus 1 1
Pieridae (S= 12) Coliadinae (S= 8)
Anteos clorinde 1 1Aphrissa statira 1 1Eurema albula 1 1Eurema elathea 73 30 6 12 2 1 1 125Eurema phiale 2 1 3Phoebis sennae 7 4 2 6 6 1 2 28Pyrisitia leuce 1 2 1 1 5Pyrisitia nise 11 12 10 35 4 3 63

19
Pierinae (S= 4) Ascia monuste 1 1Cunizza hirlanda 1 1Glutophrissa drusilla 2 1 3Hesperocharis anguitia 1 1 2
Riodinidae (S= 16) Riodininae (S= 15)
Anteros lectabilis 1 1Ariconias glaphyra 8 5 4 11 28Aricoris cf pasquita 1 1Aricoris propitia 20 9 11 1 2 5 48Aricoris tutana 4 8 12Baeotis johannae 3 5 8Chalodeta theodora 1 1Dachetola azora 2 1 1 4Emesis diogenia 5 5 1 2 13Lemonias stalachtioides 2 2 4Lyropteryx terpsichore 2 1 3Nirodia belphegor 3 3Stichelia bocchoris 5 5Synargis galena 1 1Synargis axenus 7 7Synargis calyce 1 1
Total de indivíduos 285 207 114 158 80 88 45 977Total de espécies 51 59 36 35 34 29 25 127
Apesar do esforço amostral de 336 horas-rede, a curva de acumulação de
espécies ainda não indica uma tendência à assíntota. Portanto, novas espécies devem ser
adicionadas à lista de borboletas nectarívoras da Serra do Cipó com a realização de mais
amostragens (Figura 2).
Figura 2 – Curva de acumulação de espécies de borboletas nectarívoras da Serra do
Cipó, MG. A área em cinza representa o desvio padrão dos valores médios.

20
Variação no gradiente altitudinal
Foram encontrados 24 estudos que analisaram a distribuição de borboletas em
gradientes altitudinais, a partir de levantamento bibliográfico nas plataformas Web os
Science e Google Scholar, utilizando as palavras-chaves “butterfly”, “altitudinal
gradient” e “elevational gradient”. Esses estudos parecem não apontar uma resposta
única, mas os padrões mais comuns são o decréscimo da riqueza com a altitude ou a
maior riqueza em altitudes intermediárias (Tabela 2).
Tabela 2 – Estudos utilizando borboletas como modelos em gradientes altitudinais. Na
coluna sobre os padrões: Baixo platô = riqueza não varia nas baixas altitudes, mas
decresce nas altitudes maiores; Extremos = riqueza menor em altitudes intermediárias;
Intermediário = riqueza maior em altitudes intermediárias, Negativo = riqueza decresce
com aumento da altitude; Nenhum = nenhuma variação na riqueza ao longo do
gradiente; Positivo = riqueza aumenta com a altitude.
País Taxon Gradiente Padrão Referência
México Papilionidae, Pieridae e Nymphalidae 100 - 3000 m Baixo platô Molina-Martinez et al. 2013
Indonésia Borboletas 1600 - 2080 m Extremos Tati-Subahar et al. 2007
Espanha Borboletas 1110 - 2000 m Intermediário Sanchez-Rodriguez & Baz 1995
Espanha Borboletas 520 - 1330 m Intermediário Gutiérrez 1997 Estados Unidos Borboletas 1917 - 3272 m Intermediário Fleishman et al. 1998
Ilhas Comores Borboletas endêmicas (Papilionidae, Pieridae e Nymphalidae)
0 - 1400 m Intermediário Lewis et al 1998
Venezuela Satyrinae (Nymphalidae) 2250 - 3025 m Intermediário Pyrcz & Wojtusiak 2002
México Borboletas 1900 - 3800 m Intermediário Luna-Reyes & Llorente-Bousquets 2004
Espanha Borboletas 620 - 2040 m Intermediário Wilson et al. 2007 Espanha Borboletas 596 - 2260 m Intermediário Illán et al. 2010 Israel Borboletas 500 - 2200 m Intermediário Levanoni et al. 2011 Espanha Borboletas 0 - 1930 m Intermediário Stefanescu et al. 2011 Itália Borboletas 0 - 1500 m Intermediário Dainese & Poldini 2012 Chile Borboletas 3000 - 4850 Intermediário Despland et al. 2012 Alemanha Borboletas e mariposas 340 - 750 m Intermediário Wagner et al. 2013
México Borboletas 300 - 2450 m Negativo Vargas-Fernández et al. 1992
Ilhas Comores
Borboletas não endêmicas (Papilionidae, Pieridae e Nymphalidae)
0 - 1400 m Negativo Lewis et al. 1998
Colombia Borboletas 1900 - 3000 m Negativo Camero & Calderon 2007

21
Índia Borboletas 1780 - 3600 m Negativo Chettri 2010 Áustria Borboletas 2050 - 3200 m Negativo Schnepf 2010 Himalaia Borboletas 900 - 3500 m Negativo Bhardwaj et al. 2012 Brasil Hesperiidae 900 - 1800 m Negativo Carneiro et al. 2014 Chile Borboletas 2200 - 3600 m Nenhum Arroyo et al. 1982
Equador Satyrinae (Nymphalidae) 1600 - 2600 m Positivo Pyrcz et al. 2009
No presente estudo, a riqueza de espécies de borboletas nectarívoras diminuiu
monotonicamente com o aumento da altitude (p<0,001; Figura 3). A abundância de
indivíduos seguiu o mesmo padrão, com menor quantidade de borboletas registradas nas
maiores elevações (p<0,001; Figura 3).
Figura 3 – Riqueza (círculo preenchido e linha contínua) e abundância (círculo aberto e
linha tracejada) de borboletas distribuídas no gradiente altitudinal na Serra do Cipó,
MG.
Metade das famílias apresentaram um padrão de decréscimo da riqueza com a
altitude [Hesperiidae (p=0,007; Figura 4a), Lycaenidae (p=0,017, Figura 4b) e
Papilionidae (p=0,009; Figura 4d)]. A riqueza dessas famílias somadas representa
aproximadamente 67% da riqueza total da comunidade. As demais famílias não
apresentaram variação da riqueza de espécies com a altitude: Nymphalidae (p=0,35;
Figura 4c), Pieridae (p=0,14; Figura 4e) e Riodinidae (p=0,10; Figura 4f). Não foi
encontrada nenhuma espécie pertencente à família Papilionidae no ponto mais elevado
(1400m de altitude). Em relação à abundância de indivíduos, houve um decréscimo com
o aumento da altitude nas famílias Hesperiidae (p<0,001; Figura 4a), Lycaenidae

22
(p<0,001; Figura 4b), Pieridae (p=0,01; Figura 4e) e Riodinidae (p=0,001; Figura 4f). A
quantidade de indivíduos dessas famílias somadas representa aproximadamente 87% da
abundância total da comunidade. Não apresentaram variação na abundância conforme o
aumento da altitude as famílias Nymphalidae (p=0,96; Figura 4c) e Papilionidae
(p=0,16; Figura 4d).
Figura 4 – Variação na riqueza de espécies (círculos preenchidos e linha contínua) e
abundância de indivíduos (círculo vazio e linha tracejada) de famílias de borboletas
nectarívoras [a) Hesperiidae, b) Lycaenidae, c) Nymphalidae, d) Papilionidae, e)
Pieridae e f) Riodinidae] em cada altitude amostrada na Serra do Cipó, MG.

23
Hesperiidae foi a família mais representativa nesse estudo. Foram capturados
indivíduos pertencentes a três subfamílias, sendo Hesperiinae a mais rica, com 19
espécies, seguida de Eudaminae e Pyrginae, com 14 espécies cada uma. Em relação à
distribuição dessas subfamílias ao longo do gradiente altitudinal, não foi observada o
decréscimo da riqueza de nenhuma delas separadamente. Porém, a abundância de
Eudamiinae (p=0,01) e Pyrginae (p<0,001) decresceu com a elevação.
Dados climáticos
De acordo com os dados obtidos pelas estações meteorológicas instaladas na
Serra do Cipó pode-se observar que o ano de 2012 teve temperaturas ligeiramente
menores que o ano de 2013, com um período de frio de abril a agosto. A umidade
relativa também foi menor no ano de 2012 (Figura 5). Comparando com a média de dez
anos de dados climáticos da área de estudo feita por Madeira e Fernandes (1999), o ano
de 2012 pode ser considerado atípico, já o ano de 2013 seguiu o padrão geral, com
temperaturas mais amenas e menor umidade relativa durante a época seca (junho a
outubro).
Figura 5– Variação temporal da temperatura média (quadrado com linha contínua),
temperatura mínima (círculo com linha pontilhada), temperatura máxima (triângulo com

24
linha tracejada) e umidade (barras) ao longo de dois anos de estudo na Serra do Cipó,
MG.
Houve uma queda na temperatura e aumento da umidade ao longo do gradiente
altitudinal (Figura 6a). A velocidade do vento aumentou a partir de 1200 m de altitude,
enquanto que a radiação solar só diminuiu na altitude de 1400 m (Figura 6b).
Figura 6– a) Temperatura média (quadrado com linha contínua), temperatura mínima
(círculo com linha pontilhada), temperatura máxima (triângulo com linha tracejada),
umidade (barras) e b) velocidade do vento (quadrado com linha contínua) e radiação
solar (círculo com linha tracejada) ao longo de dois anos de estudo na Serra do Cipó,
MG.
a
b
25
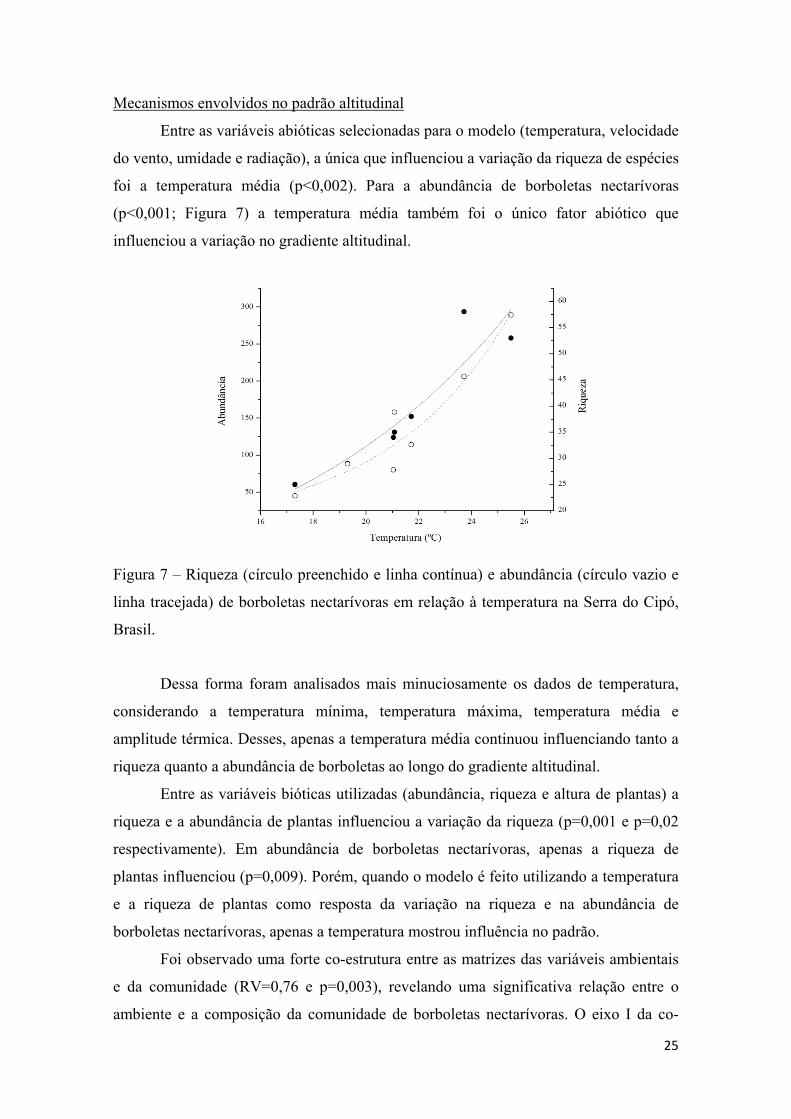
Mecanismos envolvidos no padrão altitudinal
Entre as variáveis abióticas selecionadas para o modelo (temperatura, velocidade
do vento, umidade e radiação), a única que influenciou a variação da riqueza de espécies
foi a temperatura média (p<0,002). Para a abundância de borboletas nectarívoras
(p<0,001; Figura 7) a temperatura média também foi o único fator abiótico que
influenciou a variação no gradiente altitudinal.
Figura 7 – Riqueza (círculo preenchido e linha contínua) e abundância (círculo vazio e
linha tracejada) de borboletas nectarívoras em relação à temperatura na Serra do Cipó,
Brasil.
Dessa forma foram analisados mais minuciosamente os dados de temperatura,
considerando a temperatura mínima, temperatura máxima, temperatura média e
amplitude térmica. Desses, apenas a temperatura média continuou influenciando tanto a
riqueza quanto a abundância de borboletas ao longo do gradiente altitudinal.
Entre as variáveis bióticas utilizadas (abundância, riqueza e altura de plantas) a
riqueza e a abundância de plantas influenciou a variação da riqueza (p=0,001 e p=0,02
respectivamente). Em abundância de borboletas nectarívoras, apenas a riqueza de
plantas influenciou (p=0,009). Porém, quando o modelo é feito utilizando a temperatura
e a riqueza de plantas como resposta da variação na riqueza e na abundância de
borboletas nectarívoras, apenas a temperatura mostrou influência no padrão.
Foi observado uma forte co-estrutura entre as matrizes das variáveis ambientais
e da comunidade (RV=0,76 e p=0,003), revelando uma significativa relação entre o
ambiente e a composição da comunidade de borboletas nectarívoras. O eixo I da co-

26
inércia explicou 89,9% da inércia total. Do lado positivo do eixo I da co-inércia estão as
áreas relativamente mais quentes e secas com maior radiação, riqueza de plantas e
plantas mais altas, e menor velocidade do vento (Figura 8a). Desse lado do eixo estão
espécies como Ariconias glaphyra, Chiomara basigutta, Aricoris propitia, Emesis
diogenia, Pyrisitia nise, Baeotis johannae, Phoebis sennae, Parides bunichus, Leptotes
cassius, Hemiargus hanno, Eurema elathea, Pyrgus orcus, Udranomia spitzi, Urbanus
cindra, Viola violella, Gorgythion sp. e Euptoieta hegesia (Figura 8b). Em contraste, do
lado negativo estão as áreas mais frias e úmidas, com maior velocidade do vento, menor
radiação, menor riqueza de plantas e plantas mais baixas (Figura 8a). As espécies que se
posicionaram desse lado do eixo I foram Vanessa braziliensis, Cogia azila, Stichelia
bocchoris, Lyropteryx terpsichore, Cogia hassan evansi, Quasimellana mielkei, Nirodia
belphegor, Panoquina peraea e Thespieus homochromus, (Figura 8b). As demais
espécies tiveram pouca relação com o eixo I da co-inércia.
a

27
b

28
Figura 8 – Co-estrutura entre comunidade de borboletas nectarívoras e as variáveis
ambientais ao longo de um gradiente altitudinal na Serra do Cipó, MG. Coordenadas no
eixo I da análise de co-inércia da: a) variáveis ambientais e b) espécies de borboletas
nectarívoras. Em a, Umid= umidade, Vento = velocidade do vento, Apl = abundância de
plantas, Rad = radiação, Rpl = riqueza de plantas, Altp = altura das plantas e Temp =
temperatura.
A correlação entre o eixo I da co-inércia e a altitude foi significativa (r = 0,94; p
= 0,001) indicando uma gradual mudança da comunidade de borboletas, bem como das
variáveis ambientais que a estruturam, com a altitude (Figura 9). Além disso, estes
dados reforçam a indicação de que a composição de espécies muda conforme o aumento
da altitude, porém as altitudes intermediárias (de 1000 a 1200 m) apresentaram
coordenadas semelhantes, indicando que tanto as comunidades de espécies quanto as
características ambientais desses locais são semelhantes.
Figura 9 – Correlação de Pearson entre o eixo I da co-inércia e o gradiente de altitude da
Serra do Cipó, MG.
Analisando cada ponto ao longo do gradiente, o componente αobservada foi sempre
maior que o α esperado pelo modelo nulo, enquanto que a diversidade βobservada foi
sempre menor que a esperada, ou seja, a troca de espécie é baixa entre as trilhas em

29
cada altitude. A única exceção foi em 1400m de altitude, onde não houve diferença
entre as diversidades αobservada e αesperada e βobservada e βesperada (Tabela 3).
Tabela 3 – Partição multiplicativa da diversidade de borboletas nectarívoras no
gradiente altitudinal da Serra do Cipó, Brasil.
Altitude Componente da diversidade Observado Esperado p
800m α 28.00 22.41 <0,01 β 1.61 2.01 <0,01 γ 45.03 45.03 900m α 29.00 24.05 <0,01 β 1.79 2.16 <0,01 γ 51.92 1000m α 18.00 15.24 <0,01 β 1.88 2.23 <0,01 γ 33.90 1100m α 20.00 15.77 <0,01 β 1.56 1.98 <0,01 γ 31.13 1200m α 16.33 13.84 <0,01 β 1.88 2.22 <0,01 γ 30.71 1300m α 14.67 12.16 <0,01 β 1.76 2.13 <0,01 γ 25.85 1400m α 10.00 9.19 N.S. β 2.34 2.55 N.S. γ 23.40
A diversidade αobservada (p=0,001), αesperada (p=0,002), e a diversidade γ (p=0,005)
decrescerem com o aumento da altitude (Figura 10 a). Porém nem a diversidade βobservada
(p=0,12) nem a diversidade βesperada (p=0,12) seguiram o mesmo padrão (Figura 10b).
Apesar disso, o β foi maior no ponto mais elevado (β=2,34) do que em altitudes menos
elevadas (β=1,61 para 800m e β=1,79 para 900m). O local que apresentou o menor β foi
o ponto localizado a 1100m de altitude (β=1,56).

30
Figura 10 – a) Diversidade αobservada (círculo fechado e linha tracejada), αesperada (círculo
aberto e linha tracejada) e γ (triângulo e linha contínua) e b) diversidade βobservada
(círculo fechado) e βesperada (círculo aberto) ao longo do gradiente altitudinal da Serra do
Cipó, MG.
DISCUSSÃO
A quantidade de espécies encontradas nesse estudo é similar a outros
levantamentos realizados em ambientes campestres brasileiros (Iserhard et al. 2010,

31
Dolibaina et al. 2011, Carneiro et al. 2014). Dentre as espécies coletadas, 61 (48%) são
consideradas típicas de ambientes abertos ou já foram registradas em outros ambientes
campestres (Callaghan 1982, Mielke & Casagrande 1991, Brown 1993, Hall 2002,
Marchiori & Romanowski 2006, Mielke et al. 2008, Iserhard et al. 2010, Dolibaina et
al. 2011, Kaminski et al. 2012, Kaminski & Carvalho-Filho 2012).
O padrão de distribuição das espécies de borboletas nectarívoras ao longo do
gradiente altitudinal na Serra do Cipó seguiu o padrão negativo, ou seja, houve um
decréscimo monotônico na riqueza e abundância com o aumento da altitude. Esse
resultado corrobora outros estudos com lepidópteros em outras regiões do globo
(Vargas-Fernandez 1992, Camero & Calderon 2007, Schnepf 2010, Carneiro et al.
2014). A maior riqueza da vegetação em altitudes menores pode ser indicador de maior
quantidade de recursos e nichos encontrados em altitudes mais baixas (e.g., Brown &
Freitas 2000, García-Barros 2000, Mota et al. 2014). A temperatura também aumenta
em altitudes inferiores, influenciando positivamente na quantidade de espécies de
borboletas (Molina-Martinez et al. 2013). Porém, analisando separadamente as famílias
de borboletas, foram observadas diferentes respostas à altitude, onde apenas três
(Hesperiidae, Lycaenidae e Papilionidae) mostraram variação de riqueza ao longo do
gradiente. A maioria das espécies da família Lycaenidae tem tamanho corporal pequeno
(New 1993) e como o tamanho corporal das borboletas está negativamente relacionado
com a temperatura (García Barros 2000) é possível que uma resposta ao estresse
ambiental nas altas altitudes seja a diminuição da riqueza dessa família. A família
Hesperiidae apresenta uma alta taxa de endemismo nesses locais. De fato, no presente
estudo das 47 espécies coletadas, 34 (72%) são comumente encontradas em campos ou
áreas abertas (Mielke & Casagrande 1991, Mielke et al. 2008, Dolibaina et al. 2011,
Carneiro 2012). A família Nymphalidae (14 espécies, 11% da riqueza total) foi a única
que não apresentou variação nem da abundância nem da riqueza de espécies ao longo do
gradiente. A espécie Junonia evarete foi encontrada em todas as altitudes, indicando ser
altamente tolerante à variação da temperatura experimentada no gradiente altitudinal da
Serra do Cipó. Além disso, Vanessa braziliensis, outra representante da família
Nymphalidae, foi coletada apenas na altitude mais elevada, com um comportamento
típico de hilltopping (Shields 1967). Assim, a distribuição dessa família parece ser
influenciada por outros fatores, além da altitude testada nesse estudo.
A temperatura é um fator preponderante na vida dos insetos, regulando e
atuando diretamente na sua fisiologia, metabolismo, ciclo de vida, comportamento,

32
desenvolvimento e sobrevivência (Speight et al. 2008). A temperatura influencia a
distribuição de vários grupos de invertebrados em gradientes altitudinais (Jacobsen et al.
1997, Bale et al. 2002, Sanders et al. 2007), incluindo borboletas (Molina-Martinez
2013). Nesse estudo, esse foi o fator que mais influenciou a estruturação da comunidade
de borboletas nectarívoras da Serra do Cipó. Assim, a temperatura parece ser o principal
determinante ambiental, direcionando a diversidade e definindo padrões de ocorrência
de borboletas nessas montanhas do sudeste brasileiro.
Apesar da extensa documentação sobre a intrínseca relação entre borboletas e a
vegetação (Gilbert 1971, Chew 1975, Smiley 1978, Dennis et al. 2004, Ferrer-Paris et
al. 2013), no presente estudo ela parece atuar de forma secundária na diversidade de
borboletas nectarívoras. Como a diversidade florística também decresce no gradiente
altitudinal da Serra do Cipó (Mota et al. 2014) a temperatura também é um filtro
ambiental para a comunidade de plantas. Dessa forma, a diversidade de plantas estaria
indiretamente associada à diversidade de borboletas nectarívoras, pois ambas seriam
afetadas pela mesma pressão ambiental. No entanto, estudos com maior detalhamento
das interações entre essas borboletas e as plantas hospedeiras podem fornecer
informações mais precisas e revelar interessantes padrões ecológicos.
A fauna de Lepidoptera das montanhas neotropicais é pouco conhecida, o que
dificulta o reconhecimento e a determinação do status de ameaça e endemismo desse
componente da biodiversidade. O presente estudo corrobora esta afirmativa, pois os
registros evidenciam que algumas espécies de borboletas podem estar ameaçadas,
enquanto outras tem pequenas áreas de distribuição, sendo portanto especialmente
vulneráveis. Por exemplo, Arcas ducalis (Lycaenidae) encontrada neste estudo é uma
espécie restrita a topos de morro de vegetação preservada do sul e sudeste brasileiro
(Nicolay 1971, Brown Jr. 1992, 1993), que, apesar de não figurar na lista de espécies
ameaçadas do estado de Minas Gerais (Casagrande et al. 1998) está presente nas listas
dos estados de São Paulo (Brown Jr & Freitas 1999), Paraná (Casagrande & Mielke
1995) e Rio de Janeiro (Otero et al. 2000). Outro caso é o da espécie Nirodia belphegor
(Riodinidae), com três indivíduos coletados em apenas um local a 1300 m de altitude.
Essa espécie consta na lista da fauna ameaçada de extinção do Brasil (Machado et al.
2008) e é classificada como EN (ameaçada) pela IUCN. Embora conste que ambas as
espécies foram categorizadas como ameaçadas principalmente devido à degradação e
modificação do habitat, este estudo acrescentou novos pontos de ocorrência para as
mesmas. Há, portanto, a necessidade de estudos mais detalhados com essas espécies

33
ameaçadas assim como a aplicação de políticas eficazes para a preservação dos habitats
onde ocorrem (Brown Jr 1993, Freitas & Marini-Filho 2011). Um aspecto de grande
relevância é o aprofundamento do conhecimento sobre as plantas hospedeiras e história
natural das espécies de lepidópteros da região incluindo estudos populacionais nesses
ambientes já ameaçados de campo rupestre (veja Barbosa et al. 2010, Fernandes &
Barbosa 2013).
A análise de co-inércia indicou que espécies pouco estudadas, como Lyropteryx
terspichore (Riodinidae), Cogia azila e Quasimellana mielkei (Hesperiidae) tiveram
uma forte especificidade para habitats campestres mais úmidos, frios e com ventos mais
fortes, os quais ocorrem predominantemente nas altitudes mais elevadas. Essas espécies
e outras que apresentaram forte relação com esse eixo do gráfico são espécies típicas de
ambientes montanhosos (Burns 1994, Carneiro 2012). Em contraposição, espécies com
maior afinidade a ambientes mais quentes, secos e com menos vento, que ocorrem nas
terras mais baixas, possuem sua biologia muito mais estudada. Dessas, muitas são
espécies amplamente distribuídas, que ocorrem tanto em ambientes abertos quanto em
matas fechadas ou mesmo ambientes ruderais, como Hemiargus hanno, Leptotes
cassius (Lycaenidae), Pyrisitia nise, Eurema elathea (Pieridae) e Ariconias glaphyra
(Riodinidae) (Brown Jr. 1992). Estes dados evidenciam mais uma vez a importância dos
ambientes campestres montanhosos para a conservação de espécies e interações
ecológicas.
Apesar de existirem grandes eventos de migração em algumas espécies (Urquhar
1960, Baker 1969), algumas borboletas são sedentárias ou possuem pouca
movimentação, estando boa parte da vida restritas a locais onde ocorre sua planta
hospedeira (Erlich & Raven 1964, Beirão et al. 2012) ou devido a fatores
comportamentais (Erlich 1961). As borboletas também são muito sensíveis às mudanças
ambientais, o que pode ser uma barreira para seu deslocamento no gradiente amostrado,
tornando as comunidades mais homogêneas em cada faixa altitudinal. Embora as
diversidades α e γ tenham decrescido ao longo do gradiente, a diversidade β não
decresceu, ou seja, não houve uma tendência à homogeneização da comunidade,
diferindo de outros estudos (Kraft et al 2011, Mori et al 2011). A maior diversidade β
encontrada a 1400m de altitude pode ser devido à presença de capões, que são ilhas com
formações vegetacionais similares à Mata Atlântica (Giulietti & Pirani 1997), que são
capazes de abrigar uma comunidade de borboletas completamente distinta daquela
encontrada nas áreas campestres. Nestes capões de mata podem ocorrer espécies

34
restritas a esta altitude como A. ducalis e Evenus regalis coletadas nessa faixa
altitudinal.
De acordo com os dados obtidos ao longo de dois anos de pesquisa, observou-se
que houve uma redução média de temperatura de 0,85ºC a cada 100m de elevação, valor
um pouco maior que o padrão de 0,6ºC. Uma das respostas biológicas esperadas para
espécies de montanha frente ao aquecimento global é o deslocamento vertical das
espécies (Parmesan et al 1999, Wilson et al 2007, Colwell et al 2008). Dessa forma,
espécies que estão distribuídas em altitudes inferiores podem se deslocar para altitudes
mais elevadas, enquanto aquelas espécies restritas a topos de montanha serão excluídas
(Parmesan & Yohe 2003) ocorrendo também empobrecimento da diversidade em baixas
altitudes (Wilson et al 2007, Colwell et al 2008). Topos de morro podem ser o habitat
de espécies endêmicas ou de localização restrita, como na espécie A. ducalis,
encontrada apenas à 1400 m de altitude. Além dessa, foram encontradas oito espécies
exclusivas localmente a 1200 m de altitude (incluindo a ameaçada N. belphegor), oito
espécies exclusivas a 1300 m de altitude e 11 espécies exclusivas do local a 1400 m de
altitude. Frente um cenário de mudanças climáticas, é possível que várias espécies desta
região possam vir a figurar em alguma categoria de vulnerabilidade ou mesmo tornar-se
extintas, não obstante a possibilidade de migração para altitudes superiores nesta cadeia
de montanhas. Mas independentemente dos fatos e evidências do aumento de eventos
relacionados a mudanças climáticas na região (veja Fernandes et al. 2011), estudos
sobre a historia natural das espécies de borboletas e suas interações ecológicas são
fundamentais para subsidiar planos de conservação das espécies e habitats nestas
diversas montanhas tropicais. Embora seja difícil prever com exatidão o que irá
acontecer com a comunidade de borboletas nectarívoras da Serra do Cipó diante das
ameaças, seja pela mudança no uso da terra ou pelas mudanças climáticas, o presente
estudo representa o passo inicial na busca de padrões e respostas, estimulando trabalhos
que elucidem como as interações entre as borboletas e o ambiente podem ser afetadas
em diferentes escalas futuramente e fornecendo embasamento para estratégias de
conservação dos campos rupestres.
Baseado nos resultados desse estudo, evidenciamos o papel fundamental dos
ambientes montanhosos na biodiversidade de borboletas. O mosaico formado pela
presença de regiões de matas e de campo em elevadas altitudes na Serra do Cipó revela
uma alta importância biológica desse local, destacando a importância da conservação de
áreas elevadas.

35
AGRADECIMENTOS
Agradecemos ao Diego Dolibaina e Fernando Dias pela identificação de
espécies. Ao Parque Nacional da Serra do Cipó, Pousada Serra Morena, Cedro
Cachoeira Indústria Têxtil, Reserva Vellozia e Pousada Pedra do Elefante pelo apoio
logístico. Também ao CNPq (Proc. 403781/2012-4 e Proc. 457519/2012-6) e à Fapemig
(Proc. APQ-02158-10) pelos financiamentos de bolsa e projetos.
REFERÊNCIAS BIBLIOGRÁFICAS
Abrahamczyk, S.; Kluge, J.; Gareca, Y.; Reichle, S.; Kessler, M. (2011) The influence
of climatic seasonality on the diversity of different tropical pollinator groups.
PlosOne, 6: 1 – 9.
Anderson M. J. (2001) A new method for non-parametric multivariate analysis of
variance. Austral Ecology, 26: 32 – 46.
Arroyo, M.T.K.; Primack, R.; Armesto, J. (1982) Community studies in pollination
ecology in the high temperate Andes of central Chile. I pollination mechanisms
and altitudinal variation. American Journal of Botany, 69: 82 – 97.
Baguette, M.; Clobert, J.; Schtickzelle, N. (2011) Metapopulation dynamics of the bog
fritillary butterfly: experimental changes in habitat quality induced negative
density-dependent dispersal. Ecography, 34: 170 – 176.
Bale, J., Masters, G., Hodkinson, I., Awmack, C., Bezemer, T., Brown, V., Butterfield,
J., Buse, A., Coulson, J., Farrar, J., Good, J.E.G., Harrignton, R., Hartley, R.,
Jones, T.H., Lindroth, L.R., Press, M.C., Symrnioudis, I., Watt, A.D., Whittaker,
J.B. (2002) Herbivory in global climate change research: direct effects of rising
temperature on insect herbivores. Global Change Biology, 8: 1 – 16.
Barbosa, N.P.U.; Fernandes, G.W.; Carneiro, M.A.A.; Júnior, L.A.C. (2010)
Distribution of non-native invasive species and soil properties in proximity to
paved roads and unpaved roads in a quartzitic mountainous grassland of
southeastern Brazil (rupestrian fields). Biological Invasions, 12: 3745 – 3755.
Baselga, A. (2010) Multiplicative partition of true diversity yields independent alpha
and beta components; additive partition does not. Ecology, 91: 1974 – 1981.
Beirão, M.V.; Campos-Neto, F.C.; Pimenta, I.A.; Freitas, A.V.L. (2012 Population
biology and natural history of Parides burchellanus (Papilionidae: Papilioninae:

36
Troidini), an endangered Brazilian butterfly. Annals of the Entomological
Society of America, 105: 36 – 43.
Bhardwaj, M.; Uniyal, V.P.; Sanyal, A.K.; Singh, A.P. (2012) Butterfly communities
along an elevational gradient in the Tons valley, Western Himalayas:
Implications of rapid assessment for insect conservation. Journal of Asia-Pacific
Entomology, 15: 207 – 217.
Brehm, G.; Homeier, J.; Fiedler, K. (2003) Beta diversity of geometrid moths
(Lepidoptera: Geometridae) in an Andean montane rainforest. Diversity and
Distributions, 9: 351 – 366.
Brown Jr, K.S. (1992) Borboletas da Serra do Japi: diversidade, habitats, recursos
alimentares e variação temporal. História Natural da Serra do Japi: ecologia e
preservação de uma área florestal no Sudeste do Brasil. (ed by: Morellato,
L.P.C. ) pp.142 – 187. Editora Unicamp.
Brown Jr, K.S. (1993) South America: Selected species. Conservation Biology of
Lycaenidae (Butterfly). New, T.R.(ed.), IUCN.
Brown Jr, K.S. & Freitas, A.V.L. (1999) Lepidoptera. Biodiversidade do Estado de São
Paulo, Brasil: síntese do conhecimento ao final do século XX – invertebrados
terrestres. Joly, C.A. & Bicudo, C.E.M.(eds.). FAPESP. 279 p.
Brown Jr, K.S. & Freitas, A.V.L. (2000) Atlantic forest butterflies: indicators for
landscape conservation. Biotropica, 32: 934 – 956.
Burns, J.M. (1994). Genitalia at the generic level: Atrytone restricted, Anatrytone
resurrected, new genus Quasimellana-and yes! We have no Mellanas
(Hesperiidae). Journal of Lepidopterists Society, 48: 273 – 337.
Callaghan, C.J. (1982) A study of isolating mechanisms among Neotropical butterflies
of the subfamily Riodininae. Journal of Research on the Lepidoptera, 21: 159 –
176.
Camero, E., Calderón, A.M. (2007) Comunidad de mariposas diurnas
(Lepidoptera:Rhopalocera) em un gradiente altitudinal del canon del rio
Combeima-Tollma, Colombia. Acta Biológica Colombiana, 12: 95 – 110.
Carneiro, E. (2012) Padrões de diversidade e distribuição de Hesperiidae (Lepidoptera)
na Serra do Mar paranaense, Paraná, Brasil, e as relações macroecológicas
das áreas campestres da América do Sul segundo sua composição. Thesis.
Universidade Federal do Paraná, UFPR. 156 p.

37
Carneiro, E.; Mielke, O.H.H.; Casagrande, M.M.; Fiedler, K. (2014) Skipper richness
(Hesperiidae) along elevational gradients in Brazilian Atlantic Forest.
Neotropical Entomology, 43: 27 – 38.
Casagrande, M.M. & Mielke, O.H.H. (1995) Borboletas ameaçadas de extinção no
Paraná. Lista vermelha de animais ameaçados de extinção no Estado do
Paraná. Secretaria de Estado do Meio Ambiente.
Casagrande, M.M.; Mielke, O.H.H.; Brown Jr, K.S. (1998) Borboletas (Lepidoptera)
ameaçadas de extinção em Minas Gerais, Brasil. Revista Brasileira de Zoologia,
15: 241 – 259.
Chettri, N. (2010) Cross-taxon congruence in a trekking corridor of Sikkim Himalayas:
Surrogate analysis for conservation planning. Journal for Nature Conservation,
18: 75 – 88.
Chew, F.S. (1975) Coevolution of pierid butterflies and their cruciferous food plants. I
The relative quality of available resources. Oecologia, 20: 117 – 127.
Colwell, R.K.; Brehm, G.; Cardelús, C.L.; Gilman, A.C.; Longino, J.T. (2008) Global
warming, elevational range shifts, and lowland biotic attrition in the wet tropics.
Science, 322: 258 – 261.
Costa, C.M.R.; Hermann, G.C.S.; Martins, C.S.; Lins, L.V.; Lamas, I.R. (1998)
Biodiversidade em Minas Gerais: um altas para sua conservação. Fundação
Biodiversitas.
Dainese, M., Poldini, L. (2012). Plant and animal diversity in a region of the Southern
Alps: the role of environmental and spatial processes. Landscape Ecology, 27:
417 – 431.
Darrault, R.O. &Schlindwein, C. (2002) Esfingídeos (Lepidoptera, Sphingidae) no
Tabuleiro Paraibano, nordeste do Brasil: abundância, riqueza e relação com
plantas esfingófilas. Revista Brasileira de Zoologia, 19: 429 – 443.
Dennis, R.L.H.; Hodgson, J.G.; Grenyer, R. Shreeve1, T.G.; Roy, D.B. (2004) Host
plants and butterfly biology. Do host-plant strategies drive butterfly status?
Ecological Entomology, 29: 12 – 26.
Despland, E., Humire, R., Martín, S.S. (2012) Species richness and phenology of
butterflies along an altitude gradient in the desert of northern Chile. Arctic,
Antarctic, and Alpine Research, 44: 423 – 431.

38
DeVries, P.J. (1985) Hostplant records and natural history on Costa Rican butterflies
(Papilionidae, Pieridae e Nymphalidae). The Journal of Research on the
Lepidoptera, 24: 290 – 333.
Dolédec, S. & Chessel, D. (1994) Co-inertia analysis: an alternative method for
studying species-environment relationships. Freshwater Biology, 31: 277 – 294.
Dolibaina, D.R.; Mielke, O.H.H.; Casagrande, M.M. (2011) Borboletas (Papilionoidea e
Hesperioidea) de Guarapuava e arredores, Paraná, Brasil: um inventário com
base em 63 anos de registros. Biota Neotropica, 11: 341 – 354.
Dufrêne, M., Legendre, P. (1997) Species assemblages and indicator species: the need
for a Xexible asymmet-rical approach. Ecological Monographs, 67: 345 – 366.
Echternacht, L., Trovó, M., Oliveira, C.T., Pirani, J.R. (2011) Areas of endemism in the
Espinhaço Range in Minas Gerais, Brazil. Flora, 206: 782–791.
Erlich, P.R. (1961) Intrinsic barriers to dispersal in checkerspot butterfly. Science, 134:
108 – 109.
Erlich, P.R. & Raven, P.H. (1964) Butterflies and plants: a study in coevolution.
Evolution, 18: 586 – 608.
Fernandes, G.W. & Price, P. (1988) Biogeographical gradients in galling species
richness. Oecologia, 76: 161 – 167.
Fernandes, G.W. & Price, P. (1991) Comparisons of tropical and temperate galling
species richness: the roles of environmental harshness and plant nutrient status.
Plant-animal interactions: evolutionary ecology in tropical and temperate
regions. P. W. Price; T.M. Lewinsohn; G.W. Fernandes; W W Benson. (org.).
Wiley & Sons, New York.
Fernandes, G. W., Oki, Y., Sanchez-Azofeifa, A., Faccion, G., & Amaro-Arruda, H. C.
(2011). Hail impact on leaves and endophytes of the endemic threatened
Coccoloba cereifera (Polygonaceae). Plant Ecology, 212: 1687-1697.
Fernandes , G.W. & Barbosa N.P.U. (2013) Bombas relógio que ameaçam a Natureza
em Minas Gerais. Scientific American Brasil, 60 – 61.
Ferrer-Paris, J.R.; Sánchez-Mercado, A.; Viloria, A.L.; Donaldson, J. (2013)
Congruence and diversity of butterfly-host plant associations at higher
taxonomic levels. PlosOne, 8: 1 – 15.
Fleishman, E., Austin, G.T., Wiess, A.D. (1998) An empirical test of Rapoport’s rule:
elevational gradients in montane butterfly communities. Ecology, 79: 2482 –
2493.

39
Freitas, A.V.L. & Marini-Filho, O.J. (2011) Plano de ação nacional para a
conservação dos lepidópteros. ICMBio. 124 p.
García-Barros, E. (2000) Body size, egg size, and their interspecific relationships with
ecological and life history traits in butterflies (Lepidoptera: Papilionoidea,
Hesperioidea). Biological Journal of the Linnean Society, 70: 251 – 284.
Gilbert, L.E. (1971) Butterfly-plant coevolution: has Passiflora adenopoda won the
selectional race with heliconiine butterflies? Science, 172: 585 – 586.
Giulietti, A.M.; Menezes, N.L.; Pirani, J.R.; Meguro, M.; Wanderley, M.G.L. (1987)
Flora da Serra do Cipó, Minas Gerais: caracterização e lista das espécies.
Boletim de Botânica da Universidade de São Paulo, 9: 1 – 151.
Giulietti, A.M.; Pirani, J.R.; Harley, R.M. (1997) Espinhaço range region: eastern
Brazil. Centre of plants diversity: a guide and strategy for their conservation.
Davis, S.D.; Herrera-MacBryde, V.H.H.; Villa-Lobos. J.; Hamilton, A.C.(eds.).
World Conservation Union, Cambridge.
Gutiérrez, D. (1997) Importance of historical factors on species richness and
composition of butterfly assemblages (Lepidoptera: Rhopalocera) in a northern
Iberian mountain range. Journal of Biogeography, 24: 77 – 88.
Grytness, J.A. & McCain, C.M. (2007) Elevational trends in biodiversity. Encyclopedia
of Biodiversity. Levin, S. (ed.). Elsevier, Amsterdam.
Hall, J.P.W. (2002) A review of Chalodeta Stichel with a revision of the chelonis group
(Lepidoptera: Riodinidae). Proceedings of the Entomological Society of
Washington, 104: 376 – 389.
Hill, C.J. (1992) Temporal changes in abundance of two Lycaenidae butterflies
(Lycaenidae) in relation to adult food resource. Journal of the Lepidopterists’
Society, 46: 173 – 181.
IBGE. (2012) Manual técnico da vegetação brasileira, 271 p . Instituto Brasileiro de
Geografia e Estatística. Rio de Janeiro, RJ.
Illán, J.G., Gutiérrez, D., Wilson, R. J. (2010). Fine-scale determinants of butterfly
species richness and composition in a mountain region. Journal of
Biogeography, 37: 1706 – 1720.
IPCC - Intergovernmental Panel on Climate Change. (2002) Climate Change and
Biodiversity. IPCC Technical Paper V. H. Gitay, A. Suárez, R.T. Watson & D.J.
Dokken (eds).

40
Iserhard, C.A.; Quadros, M.T.; Romanowski, H.P.; Mendonça Jr, M.S. (2010)
Borboletas (Lepidoptera: Papilionoidea e Hesperioidea) ocorrentes em diferentes
ambientes na Floresta Ombrófila Mista e nos Campos de Cima da Serra do Rio
Grande do Sul, Brasil. Biota Neotropica, 10: 309 – 320.
Jacobsen, D., Schultz, R., Encalada, A. (1997) Structure and diversity of stream
invertebrate assemblages: the influence of temperature with altitude and latitude.
Freshwater Biology, 38: 247 – 261.
Jankowski, J.E.; Ciecka, A.L.; Meyer, N.Y.; Rabenold, K.N. (2009) Beta diversity
along environmental gradients: implications of habitat specialization in tropical
montane landscapes. Journal of Animal Ecology, 78: 315 – 327.
Janzen, D.H.; Ataroff, M.; Fariñas, M.; Reyes, S.; Rincon, N.; Soler, A.; Soriano, P.;
Vera, M. (1976) Changes in the arthropod community along an elevational
transect in the Venezuelan Andes. Biotropica, 8: 193 – 203.
Jost, L. (2007) Partitioning diversity into independent alpha and beta components.
Ecology, 88: 2427 – 2439.
Kaminski, L.A. & Carvalho-Filho, F.S. (2012) Life history of Aricoris propitia
(Lepidoptera: Riodinidae) - a myrmecophilous butterfly obligately associated
with fire ants. Psyche, 1 – 10.
Kaminski, L.A.; Barbosa, E.P.; Freitas, A.V.L. (2012) Immature stages of the
neotropical mistletoe butterfly Cunizza hirlanda planasia Fruhstorfer (Pieridae:
Anthocharidini). Journal of Lepidopterists’ Society, 66: 143 – 146.
Kapos, V.; Rhind, J.; Edwards, M.; Price, M.F.; Ravilious, C. (2000) Developing a map
of the world's mountain forests. Forests in sustainable mountain development: a
state of knowledge, Price, M.F. & Butt, N. (eds.). CABI Publishing.
Kitching, R.L.; Orr, A.G.; Thalib, L.; Mitchell, H.; Hopkins, M.S.; Graham, A.W.
(2000) Moth assemblages as indicators of environmental quality in remnants of
upland Australian rain forest. Journal of Applied Ecology, 37: 284 – 297.
Kraft, N.J.B.; Comita, L.S.; Chase, J.M.; Sanders, N.J.; Swenson, N.G.; Crist, T.O.;
Stegen, J.C.; Vellend, M.; Boyle, B.; Anderson, M.J.; Cornell, H.V.; Davies,
K.F.; Freestone, A.L.; Inouye, B.D.; Harrison, S.P.; Myers, J.A. (2011)
Disentangling the drivers of β diversity along latitudinal and elevational
gradients. Science, 333: 1755 – 1758.
Körner, C. (2004) Mountain biodiversity, its causes and function. Ambio special report,
13: 11 – 17.

41
Körner, C. (2007) The use of ‘altitude’ in ecological research. Trends in Ecology and
Evolution, 22 : 569 – 574.
Levanoni, O.; Levin, N.; Pe’er, G.; Turbé, A.; Kark, S. (2011) Can we predict butterfly
diversity along an elevation gradient from space? Ecography, 34: 372 – 383.
Lewis, O.T.; Wilson, R.J.; Harper, M.C. (1998) Endemic butterflies of Grande Comore:
habitat preferences and conservation priorities. Biological Conservation, 85: 113
– 121.
Luna-Reyes, M., Llorente-Bousquets, J. (2004). Papilionoidea (Lepidoptera:
Rhopalocera) de la Sierra Nevada, México. Acta Zoologica Mexicana, 20: 79 –
102.
Machado, A.B.M; Drummond, G.M. &Paglia, A.P. (2008) Livro Vermelho da Fauna
Brasileira Ameaçada de Extinção. Volume I. Brasília, DF: Ministério do Meio
Ambiente.
Madeira, J. A. & Fernandes, G. W. Reproductive phenology of sympatric species of
Chamaecrista (Leguminosae) in Serra do Cipó, Brazil. Journal of Tropical
Ecology, 15: 463 – 479.
Madeira, J. A.; Ribeiro, K.T.; Oliveira, M.J.R.; Nascimento, J.S.; Paiva, C.L. (2008)
Distribuição espacial do esforço de pesquisa biológica na Serra do Cipó, Minas
Gerais: subsídios ao manejo das unidades de conservação da região.
Megadiversidade,4: 233 – 247.
Mani, M.S. &Saravanan, J.M. (1999) Pollination ecology and evolution in Compositae
(Asteraceae). Science Publishers, New Hampshire.
Marchiori, M.O. & Romanowski, H.P. (2006) Borboletas (Lepidoptera, Papilionoidea e
Hesperioidea) do Parque Estadualdo Espinilho e entorno, Rio Grande do Sul,
Brasil. Revista Brasileira de Zoologia, 23: 1029 – 1037.
McCain, C.M. (2007) Could temperature and water availability drive elevational species
richness patterns? A global case study for bats. Global Ecology and
Biogeography, 16: 1 – 13.
Mielke, O.H.H. & Casagrande, M.M. (1991) Papillonoidea e Hesperioidea
(Lepidoptera) do Parque Estadual do Morro do Diabo, Teodoro Sampaio, São
Paulo, Brasil. Revista Brasileira de Zoologia, 14: 967 – 1001.
Mielke, O.H.H.; Emery, E.O.; Pinheiro, C.E.G. (2008) As borboletas Hesperiidae
(Lepidoptera, Hesperioidea) do Distrito Federal, Brasil. Revista Brasileira de
Entomologia, 52: 283 – 288.

42
MMA - Ministério do Meio Ambiente (1999) Ações prioritárias para a conservação da
biodiversidade do cerrado e pantanal. Ministério do Meio Ambiente, Funatura,
Conservation - International, Fundação Biodiversitas e Universidade de Brasília.
Molina-Martinez, A., León-Cortés, J.L., Regan, H.M. (2013). Climatic and geometric
constraints as driving factors of butterfly species richness along a Neotropical
elevational gradient. Journal of Insect Conservation, 17: 1169 – 1180.
Mori, A.S., Shiono, T., Koide, D., Kitagawa, R., Ota, A.T., Mizumachi, E. (2013)
Community assembly processes shape an altitudinal gradient of forest
biodiversity. Global Ecology and Biogeography, 22: 878 – 888.
Morrone, J.J. (2006) Biogeographic areas and transition zones of :Latin America and
the Caribbean islands based on panbiogeographic and cladistic analyses of the
entomofauna. Annual Reviwe of Entomology, 51: 467 – 494.
Mota, G. S. (2012) Variação na estrutura, na composição florística e nas formas de
vida ao longo de um gradiente altitudinal na Cadeia do Espinhaço. Dissertação.
Universidade Estadual de Montes Claros, UNIMONTES, MG. 97 p.
Nascimento, J.L. & Campos, I.B. (2011) Atlas da Fauna Brasileira Ameaçada de
Extinção em Unidades de Conservação Federais. Instituto Chico Mendes de
Conservação da Biodiversidade, Brasília.
Negreiros, D. ; Moraes, M. L. B. ; Fernandes, G. W. (2008) Caracterização da
fertilidade dos solos de quatro leguminosas de campos rupestres, Serra do Cipó,
MG, Brasil. Journal of Plant Nutrition and Soil Science, 8: 30 – 39.
Negreiros, D.; Fernandes, G.W.; Silveira, F.A.O.; Chalub, C. (2009) Seedling growth
and biomass allocation of endemic and threatened shrubs of rupestrian
fields.Acta Oecologica, 35: 301 – 310.
Negreiros, D.; Fernandes, G.W.; Berbara, R.L.L.; Rodarte, L.H.O.; Barbosa, N.P.U.
(2011) Caracterização físico-química de solos quartzíticos degradados e áreas
adjacentes de campo rupestre na Serra do Cipó, MG, Brasil. Neotropical Biology
and Conservation, 6: 156 – 161.
Negreiros, D., Le-Stradic, S., Fernandes, G.W., Rennó, H.C. (2014). CSR analysis of
plant functional types in highly diverse tropical grasslands of harsh
environments. Plant Ecology, 215: 379 – 388.
New, T.R. (1993) Conservation Biology of Lycaenidae (Butterflies). International Union
for Conservation of Nature and Natural Resources, Chicago.

43
New, T.R., Pyle, R.M., Thomas, J.A., Thomas, C.D., Hammond, P.C. (1995). Butterfly
conservation management. Annual Review of Entomology, 40: 57 – 83.
Nicolay, S.S. (1971) A review of the genus Arcas with descriptions of new species
(Lycaenidae, Strymonini). Journal of the Lepidopterist’s Society, 25: 87 – 108.
Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., O'Hara, R. B.,
Simpson, G. L., Solymos, P., Stevens, M. H. H., Wagner, H. (2013). vegan:
Community Ecology Package. R package version 2.0-10.
Otero, L.S.; Brown Jr., K.S.; Mielke, O.H.H.; Monteiro, R.F.; Costa, J.M.; Macedo,
M.V.; Maciel, N.C.; Becker, J.; Salgado, N.G.; Santos, S.B.; Moya, G.E.;
Almeida, J.M.; Duarte, M. (2000) Invertebrados terrestres. A fauna ameaçada
de extinção do estado do Rio de Janeiro. Bergallo, H.G.; Rocha, C.F.D.; Alves,
M.A.S.; Vansluys, M.(eds.) Editora da Universidade do Estado do Rio de
Janeiro.
Parmesan, C., Ryrholm, N., Stefanescu, C., Hillk, J.K., Thomas, C.D., Descimon, H.,
Huntleyk, B., Kaila, L., Kullberg, J., Tammaru, T., Tennent, W.J., Thomas, J.A.,
Warren, M. (1999) Poleward shifts in geographical ranges of butterfly species
associated with regional warming. Nature, 399: 579 – 583.
Parmesan, C.; Yohe, G.A. (2003) Globally coherent fingerprint of climate impacts
across natural systems. Nature, 421: 37 – 42.
Pianka, E.R. (1966). Latitudinal gradients in species diversity: a review of concepts. The
American Naturalist, 910: 33 – 46.
Pyrcz T.W., Wojtusiak, J. (2002) The vertical distribution of pronophiline butterflies
(Nymphalidae, Satyrinae) along a elevational transect in Monte Zerpa
(Cordillera de Mérida, Venezuela) with remarks on their diversity and parapatric
distribution. Global Ecology and Biogeography, 11: 211 – 221.
Pyrcz, T.W.; Wojtusiak, J.; Garlacz, R. (2009) Diversity and distribution patterns of
Pronophilina butterflies (Lepidoptera: Nymphalidae: Satyrinae) along an
altitudinal transect in North-Western Ecuador. Neotropical Entomology, 38: 716
– 726.
R Core Team (2013). R: A language and environment for statistical computing. R
Foundation for Statistical Computing, Vienna, Austria. URL http://www.R-
project.org/.

44
Raguso, R.A. & Llorente-Bousquets, J. (1990) The butterflies (Lepidoptera) of the
Tuxtlas Mts., Veracruz, Mexico, revisited: species-richness and habitat
disturbance. Journal of Research on the Lepidoptera, 29: 105 – 133.
Rahbek, C. (1995) The elevational gradient of species richness: a uniform pattern?
Ecography, 18: 200 – 205.
Rapini, A.; Ribeiro, P.L.; Lambert, S.; Pirani, J.B. (2008) A flora dos campos rupestres
da Cadeia do Espinhaço. Megadiversidade, 4: 16 – 24.
Ribeiro, K.T.; L. Freitas. (2010) Impactos potenciais das alterações no Código Florestal
sobre a vegetação de campos rupestres e campos de altitude. Biota Neotropica,
10: 239 – 246.
Rodrigues, J.J.S., Brown Jr, K.S., Ruszczyk, A. (1993) Resources and conservation of
neotropical butterflies in urban forest fragments. Biological Conservation, 64: 3
– 9.
Salgado, L.O. 1983. Pragas das brássicas, características e métodos de controle. Informe
Agropecuário, 98: 43 – 47.
Sanchez-Rodriguez, J.F. & Baz, A. (1995) The effects of elevation on the butterfly
communities of a Mediterranean mountain, Sierra de Javalambre, central Spain.
Journal of Lepidopterists’ Society, 49: 192 – 207.
Sanders, N.J., Lessard, J.P., Fitzpatrick, M.C., Dunn, R.R. (2007) Temperature, but not
productivity or geometry, predicts elevational diversity gradients in ants across
spatial grains. Global Ecology and Biogeography, 16: 640 – 649.
Sawchik, J., Dufrêne, M., Lebrun, P. (2003) Estimation of habitat quality based on plant
community, and effects of isolation in a network of butterfly habitat patches.
Acta Oecologica, 24: 25 – 33.
Schnepf, J. (2010) Diversity of diurnal Lepidoptera across an elevational gradient
above the timberline: a case study from the Austrian Alps. Diplomarbeit.
UniversitatWein. 39 p.
Shapiro, A.M. & Shapiro, A.R. (1973) The ecological associations of the butterflies of
Staten Island (Richmond County, New York). Journal of Research on the
Lepidoptera, 12: 65 – 128.
Shields, O. 1967. Hilltopping. An ecological study of summit congregation behavior of
butterflies on a southern California hill. Journal of Research on Lepidoptera, 6:
69 – 178.

45
Smiley, J. (1978) Plant chemistry and the evolution of host specificity: new evidence
from Heliconius and Passiflora. Science, 201: 745 – 747.
Speight, M.R., Hunter, M.D., Watt, A.D. (2008) Ecology of Insects Concepts and
Aplications, 2nd edn. Wiley-Blackwell, Oxford.
Stefanescu, C., Herrando, S., Páramo, F. (2004) Butterfly species richness in the north-
west Mediterranean Basin: the role of natural and human-induced factors.
Journal of Biogeography, 31: 905 – 915.
Stefanescu, C., Carnicer, J., Peñuelas, J. (2011) Determinants of species richness in
generalist and specialist Mediterranean butterflies: the negative synergistic
forces of climate and habitat change. Ecography, 34: 353 – 363.
Suurkuukka, H., Meissner, K.K., Muotka, T. (2012) Species turnover in lake littorals:
spatial and temporal variation of benthic macroinvertebrate diversity and
community composition. Diversity and Distributions, 18: 931 – 941.
Tati-Subahar, S.S., Amasya, A.F., Choesin, D.N. (2007) Butterfly (Lepidoptera:
Rhopalocera) distribution along an altitudinal gradient on mount Tangkuban
Parahu, west Java, Indonesia. The Raffles Bulletin of Zoology, 55: 175 – 178.
Tews, J., Brose, U., Grimm, V., Tielborger, K., Wichmann, M.C., Schwager, M.,
Jeltsch, F. (2004) Animal species diversity driven by habitat
heterogeneity/diversity: the importance of keystone structures. Journal of
Biogeography, 31: 79 – 92.
Urquhart, F. A. (1960). The Monarch Butterfly. University of Toronto Press, Toronto.
van Halder, I., Barbaro, L., Corcket, E., Jactel, H. (2008) Importance of semi-natural
habitats for the conservation of butterfly communities in landscapes dominated
by pine plantations. Biodiversity Conservation, 17: 1149 – 1169.
Vargas-Fernandez, I., Llorent-Bousquets, J., Luis-Martinez, A. (1992) Listado
lepidopterofaunistico de la Sierra de Atoyac de Alvarez en el estado del
Guerrero: notas acerca de su distribuicion local y estacional (Rhopalocera:
Papilionoidea). Folia Entomologica Mexicana, 86: 41 – 178.
Wagner, K.D., Krauss, J., Steffan-Dewenter, I. (2013). Butterfly diversity and historical
land cover change along an altitudinal gradient. Journal of Insect Conservation,
17: 1039 – 1046.
Whittaker, R.H. (1960) Vegetation of the Siskiyou Mountains, Oregon and California.
Ecological Monographs, 30:279 – 338.

46
Wilson, R.J., Gutiérrez, D., Gutiérrez, J., Monserrat, V.J. (2007) An elevational shift in
butterfly species richness and composition accompanying recent climate change.
Global Change Biology, 13: 1873 – 1887.
Wolda, H. (1987) Altitude, habitat and tropical insect diversity. Biological Journal of
the Linnean Society, 30: 313 – 323.





























