Embed Size (px)
Citation preview

Journal of Antimicrobial Chemotherapy (2003) 51, 865–885DOI: 10.1093/jac/dkg156Advance Access publication 13 March 2003
865. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
© 2003 The British Society for Antimicrobial Chemotherapy
Natural antimicrobial susceptibilities of strains of ‘unusual’ Serratia species: S. ficaria, S. fonticola, S. odorifera, S. plymuthica and
S. rubidaea
Ingo Stock1*, Sonja Burak1, Kimberley Jane Sherwood1, Thomas Grüger2 and Bernd Wiedemann1
1Institut für Medizinische Mikrobiologie und Immunologie, Pharmazeutische Mikrobiologie, Meckenheimer Allee 168, University of Bonn, D-53115 Bonn; 2Institut für Pharmazeutische Biologie und
Mikrobiologie, Bundesstrasse 45, University of Hamburg, D-20146 Hamburg, Germany
Received 2 April 2002; returned 8 August 2002; revised 30 December 2002; accepted 13 January 2003
The natural susceptibility to 71 antibiotics of 104 Serratia strains of Serratia ficaria (n = 15), Serratiafonticola (n = 18), Serratia odorifera (n = 16), Serratia plymuthica (n = 32) and Serratia rubidaea(n = 23) was examined. MICs were determined using a microdilution procedure in IsoSensitestbroth for all the strains and in cation-adjusted Mueller–Hinton broth for some strains. Withfew exceptions, all species tested were uniformly naturally resistant to penicillin G, oxacillin,cefazolin, cefuroxime, all tested macrolides, lincosamides, streptogramins, glycopeptides,fusidic acid and rifampicin, and naturally sensitive to several aminoglycosides, piperacillin,piperacillin/tazobactam, carbapenems, some cephalosporins, fluoroquinolones and folate-pathway inhibitors. Major species-related differences in natural susceptibility affecting clinicalassessment criteria were seen with tetracyclines, some aminoglycosides, aminopenicillins,ticarcillin, cefaclor, loracarbef, cefoxitin, pipemidic acid, chloramphenicol, nitrofurantoin andfosfomycin. Differences in susceptibility dependent on the medium were seen with macrolides,tetracycline, fosfomycin and some β-lactams. The natural antibiotic susceptibility patternssuggest novel species-specific mechanisms of antibiotic resistance. Uncharacterized species-specific aminoglycoside-modifying enzymes and multidrug efflux systems affecting tetra-cyclines, quinolones and chloramphenicol are probably responsible for some of the phenotypesobserved. The natural amoxicillin sensitivity of several strains of some species combined withnatural resistance to some narrow-spectrum cephalosporins indicate the expression of naturallyoccurring β-lactamases with unique substrate profiles. β-Lactamases of representative strainsof each species were characterized phenotypically and genotypically. It was shown that allspecies expressed naturally occurring AmpC β-lactamases and, with respect to S. fonticola,also a species-specific class A β-lactamase. Inducibility of these enzymes was shown in allspecies with the exception of S. rubidaea and four of five strains of S. plymuthica.
Introduction
Within the Enterobacteriaceae, strains of the genus Serratiaare frequently encountered in human nosocomial infections.Apart from Serratia marcescens and the Serratia liquefacienscomplex (S. liquefaciens, Serratia proteamaculans, Serratiagrimesii), which are regarded as causing the majority ofhuman Serratia infections, there is little information about theremaining Serratia spp. (‘unusual’ serratiae), including their
susceptibility patterns to antimicrobial agents or underlyingmechanisms of resistance. Studies dealing with natural anti-microbial susceptibilities of these species have not beenpublished. The main goal of the present study was to compiledata on the natural susceptibility to a wide range of antimicro-bial agents of all known ‘unusual’ Serratia spp. that havebeen described as causing human disease. These data can beapplied to validate forthcoming susceptibility data of theseserratiae as well as facilitating their identification. Identifi-
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
*Corresponding author. Tel: +49-228-732114; Fax: +49-228-735267; E-mail: [email protected]
Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022

I. Stock et al.
866
cation of ‘unusual’ serratiae is of particular interest, because areliable species assignment of these strains requires carbonsource utilization tests that are not included in commoncommercial identification systems.1,2 According to the data inthe literature2 and our experience, it can be suggested that therelative paucity of reported cases of ‘unusual’ Serratia infec-tions is largely due to the misidentification of these strains bycommercial systems. A further aim of the present study was toanalyse the natural susceptibility patterns to β-lactam anti-biotics with respect to the underlying mechanisms of resist-ance. Since natural β-lactam resistance in Enterobacteriaceaespecies is predominantly attributed to the expression ofchromosomally encoded β-lactamases,3 it was likely thatspecies-specific β-lactamases contribute to the β-lactamsusceptibility patterns observed. With one exception, studiesdealing with the β-lactamases of ‘unusual’ Serratia spp. havenot been performed.
‘Unusual’ Serratia spp. include six species, namely Ser-ratia ficaria, Serratia fonticola, Serratia odorifera, Serratiaplymuthica, Serratia rubidaea and Serratia entomophila.With the exception of the latter, all serratiae have been isolatedfrom human clinical specimens,1,4 and virulence-associatedproperties have also been found in Serratia spp. other thanS. marcescens or members of the S. liquefaciens group.5–8
S. ficaria was first characterized in 1979 as an important partof the fig tree ecosystem.9 Although it had been isolated fromclinical samples in several instances2,10–17 its definite role asa human pathogen was only established in two studies.2,10
S. ficaria predominantly causes gallbladder empyemas andsepsis in areas that cultivate fig trees and its frequency to bringabout biliary infections in Mediterranean France has beenestimated to be ∼0.7%.2 In contrast to S. ficaria, S. fonticola iswidely distributed in nature and has been predominantlyisolated from water,1,18 but also from soil and sewage,18 mol-luscs,19 birds20 and clinical samples, mainly from wounds andthe respiratory tract.21,22 The natural reservoir of S. odoriferais unknown; most strains have been isolated from clinicalspecimens and food.23 S. odorifera was described in 1978 byGrimont et al.23 and includes two biovars (biovars 1 and 2).24
Cook & Lopez24 described S. odorifera biovar 1 as an‘emerging pathogen’, because several strains have beenshown to cause sepsis with clinically significant morbidityand high mortality in neonates and patients with underlyingconditions.24–29 In contrast to biovar 1, human infections dueto S. odorifera biovar 2 have only been reported in rareinstances.30 S. plymuthica has frequently been isolated fromthe rhizosphere of various plants,31,32 but it is likely that itsreservoir also comprises water and small mammals.1 Clinic-ally, it is regarded as a significant pathogen,31 and a variety ofinfections including osteomyelitis, peritonitis, pneumonia,sepsis and wound infections have been attributed to this micro-organism.32–39 The natural reservoir of S. rubidaea, consistingof the subspecies S. rubidaea subsp. burdigalensis, S. rubi-
daea subsp. rubidaea and S. rubidaea subsp. colindalensis,1
is predominantly plants, in particular coconuts,1,40,41 whereit can attain high cell densities.41 Human infection due toS. rubidaea is regarded to be associated with the consumptionof contaminated coconuts or vegetable salads, as severalstudies have shown a high degree of salads contaminated withS. rubidaea.1 S. rubidaea has been shown to cause sepsis andother infections in outpatients and hospitalizated patients.1,42
In a recent study, S. rubidaea subsp. burdigalensis was shownto be an invasive pathogen.43 S. entomophila causes a fatal dis-ease in a New Zealand grass grub species.44 It was disregardedin our examinations because it has been isolated from eitherinsects or the environment, but not from clinical sources oranimals other than insects.44
Materials and methods
Bacterial strains
A total of 104 serratiae comprising S. ficaria (n = 15), S. fonti-cola (n = 18), S. odorifera (n = 16), S. plymuthica (n = 32) andS. rubidaea (n = 23) were examined. The strains had beenisolated from human clinical specimens, animals, plants andfrom environmental sources. An overview of the origins ofthe strains tested is shown in Table 1. S. ficaria ATCC 33105T,S. fonticola ATCC 29844T, S. odorifera ATCC 33077T,S. plymuthica ATCC 183T, S. rubidaea ATCC 27593T andEscherichia coli ATCC 25922 (DSM 1103) served asreference and control strains for antibiotic susceptibility test-ing. The type strains of S. ficaria, S. fonticola, S. odorifera,S. plymuthica and S. rubidaea as well as S. marcescens CCUG 6,S. liquefaciens CCUG 5159 and Salmonella enterica subsp.enterica ATCC 13311T (DSM 5569) served as reference andcontrol strains for β-lactamase activity tests. S. marcescensCCUG 6 and S. liquefaciens CCUG 5159 were kindly pro-vided by E. Falsen (Göteborg, Sweden); E. coli DSM 1103and S. enterica DSM 5569 were derived from the GermanCulture Collection of Microoganisms and Cell Cultures(Braunschweig, Germany).
Identification
Identification to the genus level was performed using a com-mercial identification system for Enterobacteriaceae (Micro-naut-E; Merlin-Diagnostika, Bornheim, Germany), accordingto the instructions of the manufacturer. To assure a reliablespecies assignment, additional conventional carbon utiliza-tion tests were performed according to the recommendationsof Grimont & Grimont,1 using adonitol (ADO), D-arabitol(D-ARA), L-arabitol (L-ARA), dulcitol (DUL) and palatinose(PAL; all Fluka Chemie, Buchs, Switzerland) as carbonsources. Carbon utilization tests were performed as describedrecently, using an incubation temperature of 37°C.45
Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022
Antimicrobial susceptibility of unusual Serratia spp.
867
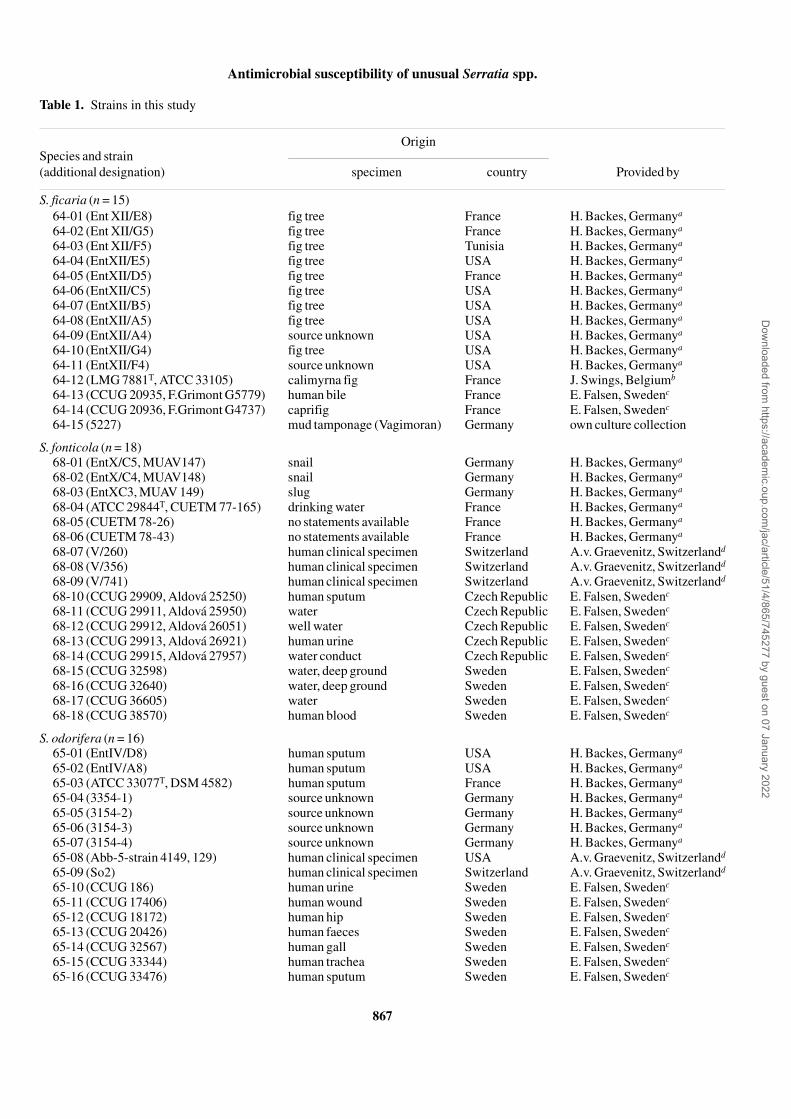
Table 1. Strains in this study
Species and strain (additional designation)
Origin
specimen country Provided by
S. ficaria (n = 15)64-01 (Ent XII/E8) fig tree France H. Backes, Germanya
64-02 (Ent XII/G5) fig tree France H. Backes, Germanya
64-03 (Ent XII/F5) fig tree Tunisia H. Backes, Germanya
64-04 (EntXII/E5) fig tree USA H. Backes, Germanya
64-05 (EntXII/D5) fig tree France H. Backes, Germanya
64-06 (EntXII/C5) fig tree USA H. Backes, Germanya
64-07 (EntXII/B5) fig tree USA H. Backes, Germanya
64-08 (EntXII/A5) fig tree USA H. Backes, Germanya
64-09 (EntXII/A4) source unknown USA H. Backes, Germanya
64-10 (EntXII/G4) fig tree USA H. Backes, Germanya
64-11 (EntXII/F4) source unknown USA H. Backes, Germanya
64-12 (LMG 7881T, ATCC 33105) calimyrna fig France J. Swings, Belgiumb
64-13 (CCUG 20935, F.Grimont G5779) human bile France E. Falsen, Swedenc
64-14 (CCUG 20936, F.Grimont G4737) caprifig France E. Falsen, Swedenc
64-15 (5227) mud tamponage (Vagimoran) Germany own culture collection
S. fonticola (n = 18)68-01 (EntX/C5, MUAV147) snail Germany H. Backes, Germanya
68-02 (EntX/C4, MUAV148) snail Germany H. Backes, Germanya
68-03 (EntXC3, MUAV 149) slug Germany H. Backes, Germanya
68-04 (ATCC 29844T, CUETM 77-165) drinking water France H. Backes, Germanya
68-05 (CUETM 78-26) no statements available France H. Backes, Germanya
68-06 (CUETM 78-43) no statements available France H. Backes, Germanya
68-07 (V/260) human clinical specimen Switzerland A.v. Graevenitz, Switzerlandd
68-08 (V/356) human clinical specimen Switzerland A.v. Graevenitz, Switzerlandd
68-09 (V/741) human clinical specimen Switzerland A.v. Graevenitz, Switzerlandd
68-10 (CCUG 29909, Aldová 25250) human sputum Czech Republic E. Falsen, Swedenc
68-11 (CCUG 29911, Aldová 25950) water Czech Republic E. Falsen, Swedenc
68-12 (CCUG 29912, Aldová 26051) well water Czech Republic E. Falsen, Swedenc
68-13 (CCUG 29913, Aldová 26921) human urine Czech Republic E. Falsen, Swedenc
68-14 (CCUG 29915, Aldová 27957) water conduct Czech Republic E. Falsen, Swedenc
68-15 (CCUG 32598) water, deep ground Sweden E. Falsen, Swedenc
68-16 (CCUG 32640) water, deep ground Sweden E. Falsen, Swedenc
68-17 (CCUG 36605) water Sweden E. Falsen, Swedenc
68-18 (CCUG 38570) human blood Sweden E. Falsen, Swedenc
S. odorifera (n = 16)65-01 (EntIV/D8) human sputum USA H. Backes, Germanya
65-02 (EntIV/A8) human sputum USA H. Backes, Germanya
65-03 (ATCC 33077T, DSM 4582) human sputum France H. Backes, Germanya
65-04 (3354-1) source unknown Germany H. Backes, Germanya
65-05 (3154-2) source unknown Germany H. Backes, Germanya
65-06 (3154-3) source unknown Germany H. Backes, Germanya
65-07 (3154-4) source unknown Germany H. Backes, Germanya
65-08 (Abb-5-strain 4149, 129) human clinical specimen USA A.v. Graevenitz, Switzerlandd
65-09 (So2) human clinical specimen Switzerland A.v. Graevenitz, Switzerlandd
65-10 (CCUG 186) human urine Sweden E. Falsen, Swedenc
65-11 (CCUG 17406) human wound Sweden E. Falsen, Swedenc
65-12 (CCUG 18172) human hip Sweden E. Falsen, Swedenc
65-13 (CCUG 20426) human faeces Sweden E. Falsen, Swedenc
65-14 (CCUG 32567) human gall Sweden E. Falsen, Swedenc
65-15 (CCUG 33344) human trachea Sweden E. Falsen, Swedenc
65-16 (CCUG 33476) human sputum Sweden E. Falsen, Swedenc
Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022

I. Stock et al.
868
Table 1. (Continued)
Species and strain (additional designation)
Origin
specimen country Provided by
S. plymuthica (n = 32)67-01 (EntX/A9) potato (rhizosphere) Germany H. Backes, Germanya
67-02 (EntXIII/B2) water Germany H. Backes, Germanya
67-03 (EntXIII/C2) water Germany H. Backes, Germanya
67-04 (EntXIII/C2) human clinical specimen Germany H. Backes, Germanya
67-05 (EntXIII/F2) human clinical specimen Germany H. Backes, Germanya
67-06 (EntXIII/G2) human clinical specimen Germany H. Backes, Germanya
67-07 (EntXIII/H2) human clinical specimen Germany H. Backes, Germanya
67-08 (EntXIII/A3) water Germany H. Backes, Germanya
67-09 (EntXIII/B3) water Germany H. Backes, Germanya
67-10 (EntXIII/C3) plant USA H. Backes, Germanya
67-11 (EntXIII/E3) rape seed Germany H. Backes, Germanya
67-13 (EntXIII/G3) maize USA H. Backes, Germanya
67-14 (Sp1-509/81) human clinical specimen Switzerland A.v. Graevenitz, Switzerlandd
67-15 (Sp2-22722/83) human clinical specimen Switzerland A.v. Graevenitz, Switzerlandd
67-16 (Sp3-22313/83) human clinical specimen Switzerland A.v. Graevenitz, Switzerlandd
67-17 (Sp4-V12-2747) human clinical specimen Switzerland A.v. Graevenitz, Switzerlandd
67-18 (Sp5-V10-6513/87) human clinical specimen Switzerland A.v. Graevenitz, Switzerlandd
67-19 (LMG 6823) Cichorium intybus Belgium J. Swings, Belgiumb
67-20 (LMG 6824) Cichorium intybus Belgium J. Swings, Belgiumb
67-21 (LMG 6825) Cichorium intybus Belgium J. Swings, Belgiumb
67-22 (LMG 6826) Cichorium intybus Belgium J. Swings, Belgiumb
67-23 (LMG 6827) Cichorium intybus Belgium J. Swings, Belgiumb
67-24 (ATCC 183T, LMG 7886) water France J. Swings, Belgiumb
67-25 (CCUG 26380) human eye (conjunctiva) Sweden E. Falsen, Swedenc
67-26 (CCUG 29919, Aldová 27048) human faeces Czech Republic E. Falsen, Swedenc
67-27 (CCUG 29920, Aldová 27450) human sputum Czech Republic E. Falsen, Swedenc
67-28 (CCUG 44368) human clinical specimen Sweden E. Falsen, Swedenc
67-29 (72-08, 333/74, M3026-3) source unknown Germany H. Backes, Germanya
67-30 (72-09, 9493, M3026-4) source unknown Germany H. Backes, Germanya
67-31 (72-19, II-9-37, M3026-13) human clinical specimen Germany own culture collection67-32 (5228) mud tamponage (Vagimoran) Germany own culture collection67-33 (Vbs-6) human clinical specimen Germany own culture collection
S. rubidaea (n = 23)66-01 (EntIX/B9) vegetable salad USA H. Backes, Germanya
66-02 (EntV/A3) coconut USA H. Backes, Germanya
66-03 (EntVIII/A9) vegetable USA H. Backes, Germanya
66-04 (EntVIII/H8) human clinical specimen USA H. Backes, Germanya
66-05 (EntVIII/G8) human clinical specimen USA H. Backes, Germanya
66-06 (EntVIII/B9) coconut USA H. Backes, Germanya
66-07 (EntVIII/C9) coconut USA H. Backes, Germanya
66-10 (Sr3 4142, 142 Abbott) human clinical specimen USA A.v. Graevenitz, Switzerlandd
66-11 (Sr4 4179, 143 Abbott) human clinical specimen USA A.v. Graevenitz, Switzerlandd
66-12 (Sr5 4180, 144 Abbott) human clinical specimen USA A.v. Graevenitz, Switzerlandd
66-13 (ATCC 27593T, LMG 5019) no statements available France J. Swings, Belgiumb
66-14 (CCUG 29738) human wound Sweden E. Falsen, Swedenc
66-15 (CCUG 29921, Aldová 25674) human urine Czech Republic E. Falsen, Swedenc
66-16 (CCUG 29922, Aldová 25873) human necrotic tissue Czech Republic E. Falsen, Swedenc
66-17 (CCUG 29923, Aldová 26444) unguentum Czech Republic E. Falsen, Swedenc
66-18 (CCUG 29924, Aldová 27564) larynx sputum (pneumonia) Czech Republic E. Falsen, Swedenc
66-19 (CCUG 35289) human sputum (pneumonia) Sweden E. Falsen, Swedenc
66-20 (5229) water Germany own culture collection66-21 (Vbs-1) human clinical specimen Germany own culture collection
Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022

Antimicrobial susceptibility of unusual Serratia spp.
869
Antibiotics and antibiotic susceptibility testing
Antibiotic susceptibility tests were performed according tothe German standard (DIN), applying a microdilutionprocedure in IsoSensitest broth (Oxoid, Basingstoke, UK) forall the strains and in cation-adjusted Mueller–Hinton broth(CAMHB; Difco Laboratories, Detroit, MI, USA) for sevenrepresentative strains each (seven strains/species). The MICswere determined with a photometer for microtitre plates (seeabove) after inoculation of antibiotic-containing microtitreplates (Merlin-Diagnostika) with 100 µL of appropriate bac-terial suspension (3 × 105 to 7 × 105 cfu/mL) and incubationfor 22 h at 36 ± 1°C. All antibiotics were kindly provided byMerlin-Diagnostika who produced the antibiotic-containingplates.
Evaluation of natural antibiotic susceptibility
Plotting the MIC of a particular antibiotic for one speciesagainst the number of strains found with the respective MICusually results in a bimodal distribution. Generally, one peakwith relatively low MICs represents the natural populationand one peak with higher MICs represents the strains withacquired (secondary) resistance. Analysis of the MIC distri-bution of all strains of one species for each antibiotic per-mitted determination of the biological thresholds, which limitthe natural population at high MICs but not those strains withsecondary resistance. Whether the MICs of the natural popu-lation were above or below the breakpoints of the standards,which assess the clinical susceptibility, was investigated.When the natural population was sensitive or intermediateaccording to the cited standard, it was described as naturallysensitive or naturally intermediate, respectively. When thenatural population was clinically resistant, it was described asnaturally resistant. The method has been described in detailpreviously.46,47 In this study, breakpoints according to theGerman standard (DIN) were applied.48 For antibiotics forwhich DIN clinical assessment criteria do not exist, break-points according to French,49 Swedish50 or NCCLS criteria,
valid for Enterobacteriaceae,51 Neisseria gonorrhoeae52 andstaphylococci,53 were employed. Breakpoints for apramycin,ribostamycin, lividomycin A and biapenem were used aspublished recently.54
β-Lactamase activity and induction assay
β-Lactamase activities and induction of five representativestrains of each ‘unusual’ Serratia sp. and of S. marcescensCCUG 6, S. liquefaciens CCUG 5159 (positive controls) andS. enterica ATCC 13311 (negative control) were tested asfollows: an overnight (18 h) culture grown on IsoSensitestagar (Oxoid) at 37°C was used to prepare a saline suspensionwith an OD640 of 0.25 (Hitachi 150–20 Spectralphotometer;Colora, Germany). The suspension (1 mL) was added to eachof two 100 mL Erlenmeyer flasks, each containing 24 mL ofIsoSensitest broth, which had been pre-warmed to 37°C.These cultures were incubated at 37°C in an incubator shaker(New Brunswick Scientific Co., Inc., Edison, NJ, USA) withshaking at 100 rpm until the growth had achieved an OD640 of0.2 ± 0.02. Imipenem was then added to one of the cultures to afinal concentration of 0.5 mg/L. No inducer was added to theother culture, which served as a control. Incubation of bothcultures was allowed to continue in an incubator shaker withshaking for 2 h. Two millilitres from each culture was with-drawn and used to measure the absorbance and for a viablecell count by spread plates; a further 15 mL was centrifuged(15 min, 4000g) and the pellet was resuspended in 5 mL of0.1 M phosphate buffer, pH 7.0. After re-centrifugation(15 min, 4000g), the pellet was resuspended in 1.5 mL ofphosphate buffer and frozen overnight. After defrosting atroom temperature, sonication on ice (Sonifier B12; Danbury,Schwäbish Gmünd, Germany) and centrifugation for 20 minat 20000g yielded a crude supernatant for β-lactamase assays.β-Lactamase activity was quantified as described by Peteret al.55 with nitrocefin as the substrate.56 The protein contentof each sample was determined by the method of Lowryet al.,57 with bovine serum albumin as the standard.
Table 1. (Continued)
aMerlin-Diagnostika, Bornheim, Germany.bBelgian Coordinated Collections of Microorganisms, University of Gent, Belgium.cDepartment of Clinical Bacteriology, Culture Collection, University of Göteborg, Sweden.dInstitut für Medizinische Mikrobiologie, University of Zürich, Switzerland.
Species and strain (additional designation)
Origin
specimen country Provided by
66-22 (Vbs-2) human clinical specimen Germany own culture collection66-23 (Vbs-3) human clinical specimen Germany own culture collection66-24 (Vbs-4) human clinical specimen Germany own culture collection66-25 (Vbs-5) human clinical specimen Germany own culture collection
Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022

I. Stock et al.
870
β-Lactamase detection by SDS–PAGE
The separation of β-lactamases by SDS–PAGE enablesclassification (Ambler β-lactamase categorization) and anestimation of the molecular weight of the enzymes expressed.β-Lactamase crude extracts of the type strains of S. ficaria,S. fonticola, S. odorifera, S. plymuthica and S. rubidaea wereseparated in 13% acrylamide, 0.35% bisacrylamide, 0.1%SDS gels (Roth, Karlsruhe, Germany). After electrophoresisand 1 h incubation in 2.5% Triton X-100 (Fluka, Steinheim,Germany), β-lactamase bands were visualized by stainingwith 1 mM nitrocefin solution. β-Lactamases with knownmolecular weights [AmpC enzymes from S. marcescensCCUG 6 (mol. wt 37.0 kDa) and Enterobacter cloacae 11-14(mol. wt 39.2 kDa), class A β-lactamase TEM-1 from E. coliJM 83 pBR322 (mol. wt 28.9 kDa)] were used for com-parison.
AmpC gene detection by PCR
The type strains of S. ficaria, S. fonticola, S. odorifera,S. plymuthica and S. rubidaea and S. marcescens CCUG 6were examined by PCR for the presence of ampC. Degener-ated ampC primer pairs used for the detection of genes encod-ing chromosomally and plasmid-encoded AmpC enzymes inEnterobacteriaceae species were applied58 (and K. J. Sherwood,unpublished results). The primer pair used to amplify a frag-ment of ampC was P1 (GGATTCCGGGTATGGCSGTNGC)and P4 (TCCCAGCCTARYCCCTGRTACAT). PCR wasconducted under standard conditions; template DNA wasobtained as described by Nakajima et al.,59 but without initialboiling and final centrifugation. PCR products were visualizedon 1% agarose (Roth) gels.
Results
Identification
The Micronaut-E identification system allowed a secureidentification of all submitted Serratia strains to the genuslevel. Although the database of the Micronaut-E systemincludes all species tested in this study with one exception(S. fonticola), it is not capable of identifying ‘unusual’ serra-tiae to the species level (data not shown). Additional carbonutilization tests allowed a reliable species identification of allstrains. The characteristic phenotypic patterns were: ADO+,D-ARA+, L-ARA+, DUL–, PAL+ for S. ficaria; ADO+,D-ARA+, L-ARA+, DUL+, PAL+ for S. fonticola; ADO+, D-ARA–, L-ARA+, DUL–, PAL– for S. odorifera; ADO–,D-ARA–, L-ARA–, DUL–, PAL+ for S. plymuthica; andADO+, D-ARA+, L-ARA–, DUL–, PAL+ for S. rubidaea.The patterns were identical to those published by Grimont &Grimont.1
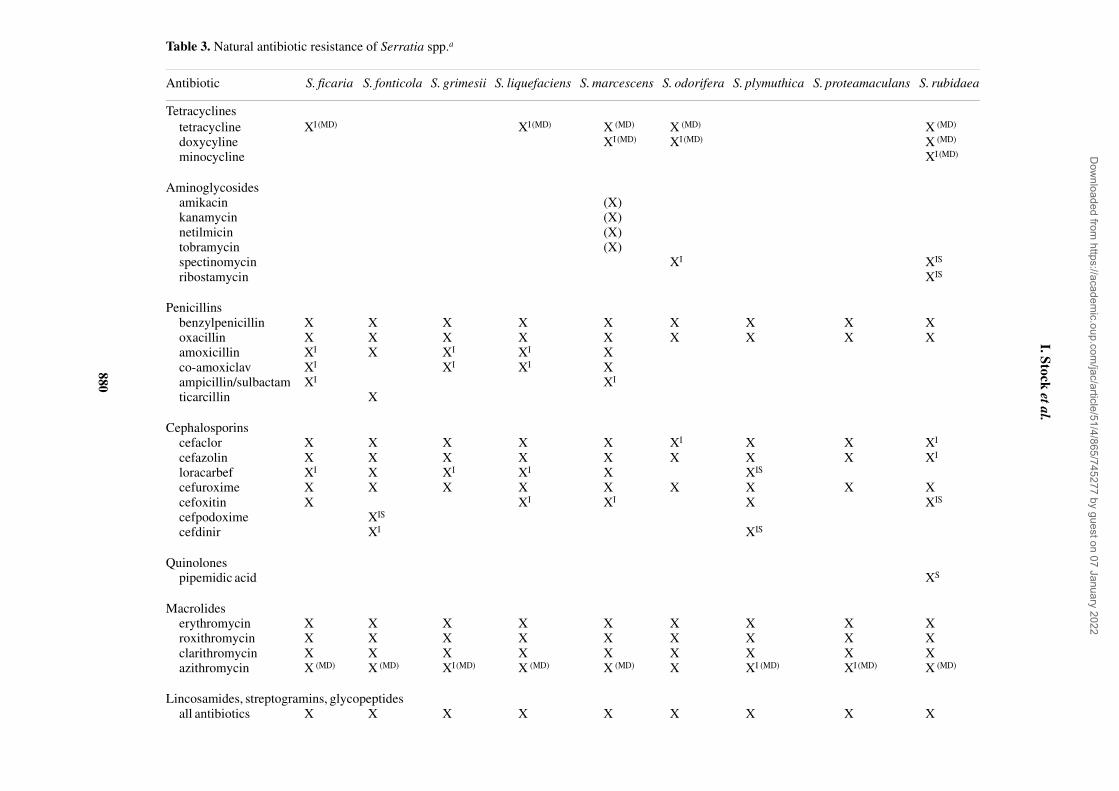
Antibiotic susceptibility, natural sensitivity and resistance
The MIC distributions for all the Serratia strains tested arepresented in Table 2. The natural antimicrobial resistances ofthe tested serratiae as well as a comparison with the respectivedata of S. marcescens and members of the S. liquefaciensgroup taken from our recent study4 are summarized in Table 3.With few exceptions, species-related differences in naturalsusceptibility were seen in each antibiotic group but not alldifferences affected clinical assessment criteria (Tables 2and 3). Despite differences in susceptibility to some agents,all species tested were uniformally naturally resistant to peni-cillin G, oxacillin, cefazolin (some strains of S. rubidaea werenaturally intermediate), cefuroxime, all tested macrolides(some strains of S. plymuthica were naturally intermediateto azithromycin), lincosamides, streptogramins, glycopep-tides, fusidic acid and rifampicin (Table 2). Uniform naturalsensitivity was seen to several aminoglycosides (amikacin,gentamicin, netilmicin, tobramycin, neomycin, apramycinand lividomycin A), piperacillin, piperacillin/tazobactam,some cephalosporins (cefixime, cefoperazone, cefotaxime,ceftibuten, ceftriaxone, ceftazidime and cefepime), carba-penems, aztreonam, all fluoroquinolones (but not pipemidicacid) and to the antifolates. Major species-related differencesin natural susceptibility affecting clinical assessment criteriawere seen with tetracyclines, some aminoglycosides(streptomycin, kanamycin, spectinomycin and ribostamycin),aminopenicillins, ticarcillin, cefaclor, loracarbef, cefoxitin,pipemidic acid, chloramphenicol, nitrofurantoin and fosfo-mycin. S. ficaria was naturally intermediate and resistant totetracyclines, aminopenicillins (in the presence and absenceof β-lactamase inhibitors) and loracarbef, and uniformallynaturally resistant to cefaclor and cefoxitin (concomitant withhigh-level cefazolin and cefuroxime resistance). S. fonticolawas unique in being naturally sensitive and intermediateto tetracyline, and naturally resistant to ticarcillin and amoxi-cillin (but sensitive or intermediate to aminopenicillins inthe presence of β-lactamase inhibitors). S. odorifera andS. rubidaea were the species least susceptible to quinolones,resulting in several S. rubidaea strains with natural resistanceto pipemidic acid and a few strains with intermediate suscep-tibility to some fluoroquinolones (Table 2). Both species werealso the least susceptible to tetracyclines, chloramphenicol,streptomycin and spectinomycin, resulting in natural resist-ance to tetracycline and chloramphenicol for all the strainsand natural resistance to spectinomycin for some strains of therespective natural populations. In addition, S. rubidaea wasthe species least susceptible to ribostamycin and antifolates.S. plymuthica was naturally intermediate to tetracycline andwas the species most susceptible to quinolones, macrolidesand lincosamides. Interestingly, strains of S. odorifera,S. plymuthica and S. rubidaea that were sensitive to amoxi-cillin were naturally resistant or intermediate to cefazolin andcefuroxime (Table 2).
Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022
Antimicrobial susceptibility of unusual Serratia spp.
871
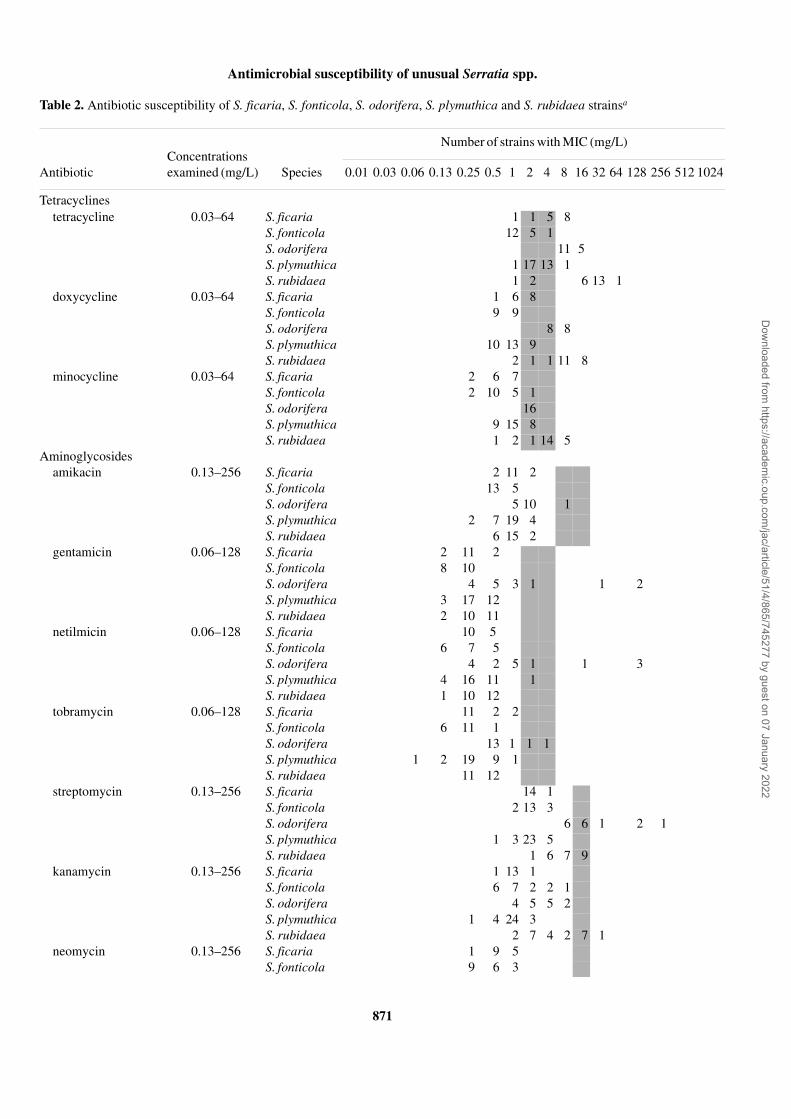
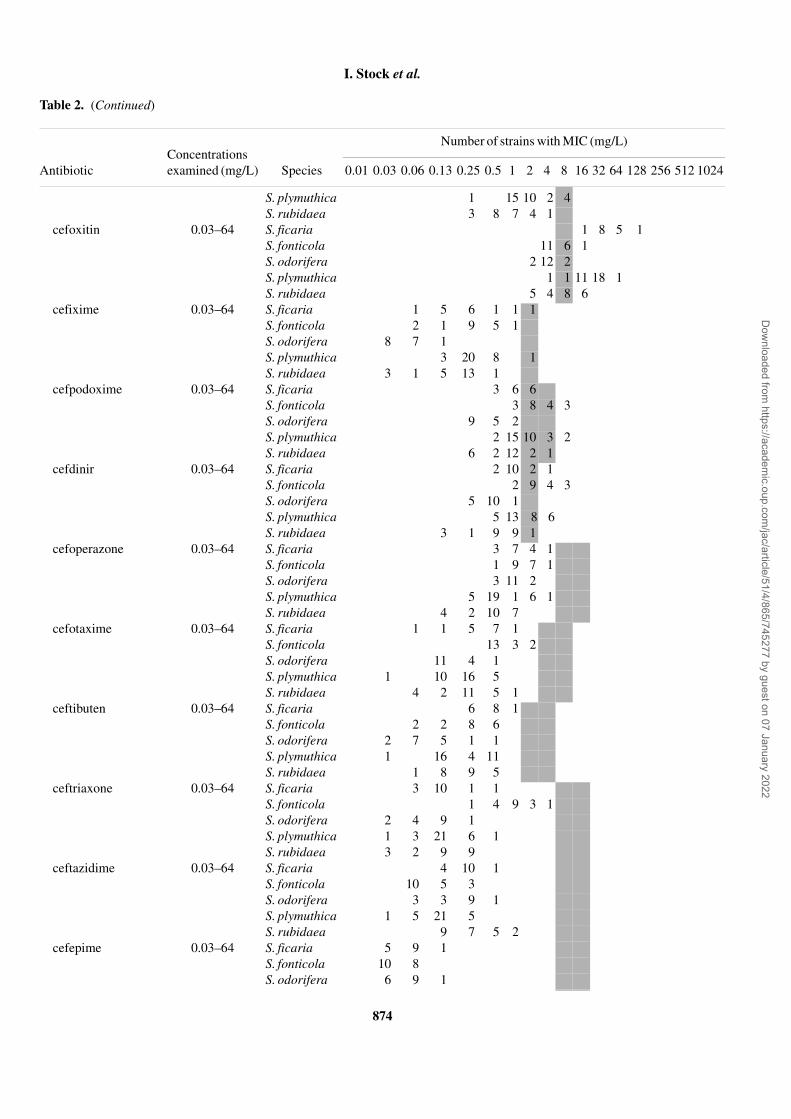
Table 2. Antibiotic susceptibility of S. ficaria, S. fonticola, S. odorifera, S. plymuthica and S. rubidaea strainsa
Concentrations examined (mg/L)
Number of strains with MIC (mg/L)
Antibiotic Species 0.01 0.03 0.06 0.13 0.25 0.5 1 2 4 8 16 32 64 128 256 512 1024
Tetracyclinestetracycline 0.03–64 S. ficaria 1 1 5 8
S. fonticola 12 5 1S. odorifera 11 5S. plymuthica 1 17 13 1S. rubidaea 1 2 6 13 1
doxycycline 0.03–64 S. ficaria 1 6 8S. fonticola 9 9S. odorifera 8 8S. plymuthica 10 13 9S. rubidaea 2 1 1 11 8
minocycline 0.03–64 S. ficaria 2 6 7S. fonticola 2 10 5 1S. odorifera 16S. plymuthica 9 15 8S. rubidaea 1 2 1 14 5
Aminoglycosidesamikacin 0.13–256 S. ficaria 2 11 2
S. fonticola 13 5S. odorifera 5 10 1S. plymuthica 2 7 19 4S. rubidaea 6 15 2
gentamicin 0.06–128 S. ficaria 2 11 2S. fonticola 8 10S. odorifera 4 5 3 1 1 2S. plymuthica 3 17 12S. rubidaea 2 10 11
netilmicin 0.06–128 S. ficaria 10 5S. fonticola 6 7 5S. odorifera 4 2 5 1 1 3S. plymuthica 4 16 11 1S. rubidaea 1 10 12
tobramycin 0.06–128 S. ficaria 11 2 2S. fonticola 6 11 1S. odorifera 13 1 1 1S. plymuthica 1 2 19 9 1S. rubidaea 11 12
streptomycin 0.13–256 S. ficaria 14 1S. fonticola 2 13 3S. odorifera 6 6 1 2 1S. plymuthica 1 3 23 5S. rubidaea 1 6 7 9
kanamycin 0.13–256 S. ficaria 1 13 1S. fonticola 6 7 2 2 1S. odorifera 4 5 5 2S. plymuthica 1 4 24 3S. rubidaea 2 7 4 2 7 1
neomycin 0.13–256 S. ficaria 1 9 5S. fonticola 9 6 3
Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022
I. Stock et al.
872
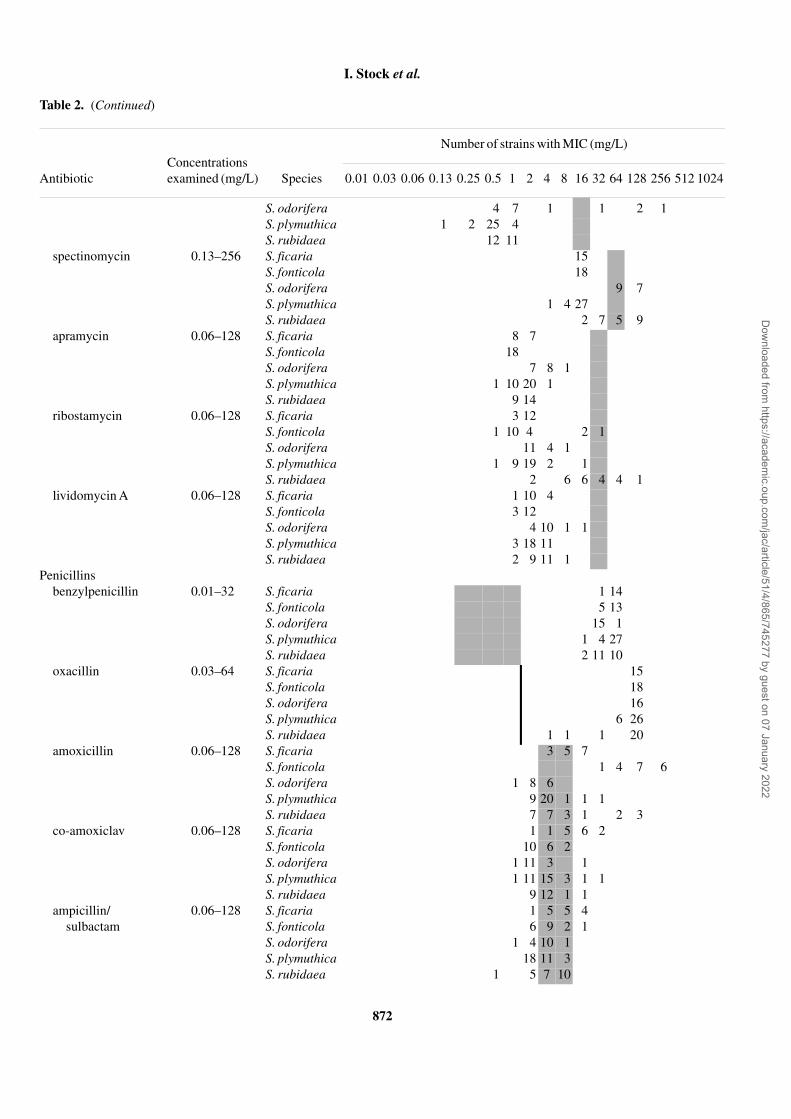
Table 2. (Continued)
Concentrations examined (mg/L)
Number of strains with MIC (mg/L)
Antibiotic Species 0.01 0.03 0.06 0.13 0.25 0.5 1 2 4 8 16 32 64 128 256 512 1024
S. odorifera 4 7 1 1 2 1S. plymuthica 1 2 25 4S. rubidaea 12 11
spectinomycin 0.13–256 S. ficaria 15S. fonticola 18S. odorifera 9 7S. plymuthica 1 4 27S. rubidaea 2 7 5 9
apramycin 0.06–128 S. ficaria 8 7S. fonticola 18S. odorifera 7 8 1S. plymuthica 1 10 20 1S. rubidaea 9 14
ribostamycin 0.06–128 S. ficaria 3 12S. fonticola 1 10 4 2 1S. odorifera 11 4 1S. plymuthica 1 9 19 2 1S. rubidaea 2 6 6 4 4 1
lividomycin A 0.06–128 S. ficaria 1 10 4S. fonticola 3 12S. odorifera 4 10 1 1S. plymuthica 3 18 11S. rubidaea 2 9 11 1
Penicillinsbenzylpenicillin 0.01–32 S. ficaria 1 14
S. fonticola 5 13S. odorifera 15 1S. plymuthica 1 4 27S. rubidaea 2 11 10
oxacillin 0.03–64 S. ficaria 15S. fonticola 18S. odorifera 16S. plymuthica 6 26S. rubidaea 1 1 1 20
amoxicillin 0.06–128 S. ficaria 3 5 7S. fonticola 1 4 7 6S. odorifera 1 8 6S. plymuthica 9 20 1 1 1S. rubidaea 7 7 3 1 2 3
co-amoxiclav 0.06–128 S. ficaria 1 1 5 6 2S. fonticola 10 6 2S. odorifera 1 11 3 1S. plymuthica 1 11 15 3 1 1S. rubidaea 9 12 1 1
ampicillin/sulbactam
0.06–128 S. ficaria 1 5 5 4S. fonticola 6 9 2 1S. odorifera 1 4 10 1S. plymuthica 18 11 3S. rubidaea 1 5 7 10
Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022
Antimicrobial susceptibility of unusual Serratia spp.
873
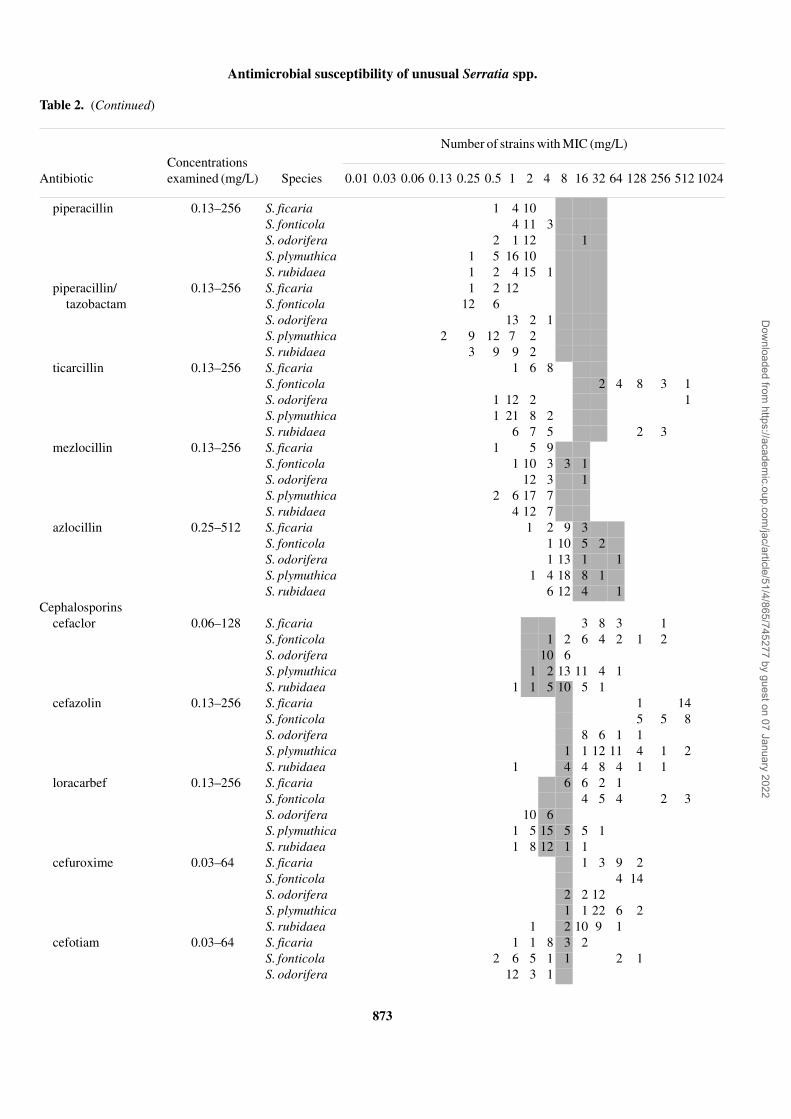
Table 2. (Continued)
Concentrations examined (mg/L)
Number of strains with MIC (mg/L)
Antibiotic Species 0.01 0.03 0.06 0.13 0.25 0.5 1 2 4 8 16 32 64 128 256 512 1024
piperacillin 0.13–256 S. ficaria 1 4 10S. fonticola 4 11 3S. odorifera 2 1 12 1S. plymuthica 1 5 16 10S. rubidaea 1 2 4 15 1
piperacillin/tazobactam
0.13–256 S. ficaria 1 2 12S. fonticola 12 6S. odorifera 13 2 1S. plymuthica 2 9 12 7 2S. rubidaea 3 9 9 2
ticarcillin 0.13–256 S. ficaria 1 6 8S. fonticola 2 4 8 3 1S. odorifera 1 12 2 1S. plymuthica 1 21 8 2S. rubidaea 6 7 5 2 3
mezlocillin 0.13–256 S. ficaria 1 5 9S. fonticola 1 10 3 3 1S. odorifera 12 3 1S. plymuthica 2 6 17 7S. rubidaea 4 12 7
azlocillin 0.25–512 S. ficaria 1 2 9 3S. fonticola 1 10 5 2S. odorifera 1 13 1 1S. plymuthica 1 4 18 8 1S. rubidaea 6 12 4 1
Cephalosporinscefaclor 0.06–128 S. ficaria 3 8 3 1
S. fonticola 1 2 6 4 2 1 2S. odorifera 10 6S. plymuthica 1 2 13 11 4 1S. rubidaea 1 1 5 10 5 1
cefazolin 0.13–256 S. ficaria 1 14S. fonticola 5 5 8S. odorifera 8 6 1 1S. plymuthica 1 1 12 11 4 1 2S. rubidaea 1 4 4 8 4 1 1
loracarbef 0.13–256 S. ficaria 6 6 2 1S. fonticola 4 5 4 2 3S. odorifera 10 6S. plymuthica 1 5 15 5 5 1S. rubidaea 1 8 12 1 1
cefuroxime 0.03–64 S. ficaria 1 3 9 2S. fonticola 4 14S. odorifera 2 2 12S. plymuthica 1 1 22 6 2S. rubidaea 1 2 10 9 1
cefotiam 0.03–64 S. ficaria 1 1 8 3 2S. fonticola 2 6 5 1 1 2 1S. odorifera 12 3 1
Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022
I. Stock et al.
874
Table 2. (Continued)
Concentrations examined (mg/L)
Number of strains with MIC (mg/L)
Antibiotic Species 0.01 0.03 0.06 0.13 0.25 0.5 1 2 4 8 16 32 64 128 256 512 1024
S. plymuthica 1 15 10 2 4S. rubidaea 3 8 7 4 1
cefoxitin 0.03–64 S. ficaria 1 8 5 1S. fonticola 11 6 1S. odorifera 2 12 2S. plymuthica 1 1 11 18 1S. rubidaea 5 4 8 6
cefixime 0.03–64 S. ficaria 1 5 6 1 1 1S. fonticola 2 1 9 5 1S. odorifera 8 7 1S. plymuthica 3 20 8 1S. rubidaea 3 1 5 13 1
cefpodoxime 0.03–64 S. ficaria 3 6 6S. fonticola 3 8 4 3S. odorifera 9 5 2S. plymuthica 2 15 10 3 2S. rubidaea 6 2 12 2 1
cefdinir 0.03–64 S. ficaria 2 10 2 1S. fonticola 2 9 4 3S. odorifera 5 10 1S. plymuthica 5 13 8 6S. rubidaea 3 1 9 9 1
cefoperazone 0.03–64 S. ficaria 3 7 4 1S. fonticola 1 9 7 1S. odorifera 3 11 2S. plymuthica 5 19 1 6 1S. rubidaea 4 2 10 7
cefotaxime 0.03–64 S. ficaria 1 1 5 7 1S. fonticola 13 3 2S. odorifera 11 4 1S. plymuthica 1 10 16 5S. rubidaea 4 2 11 5 1
ceftibuten 0.03–64 S. ficaria 6 8 1S. fonticola 2 2 8 6S. odorifera 2 7 5 1 1S. plymuthica 1 16 4 11S. rubidaea 1 8 9 5
ceftriaxone 0.03–64 S. ficaria 3 10 1 1S. fonticola 1 4 9 3 1S. odorifera 2 4 9 1S. plymuthica 1 3 21 6 1S. rubidaea 3 2 9 9
ceftazidime 0.03–64 S. ficaria 4 10 1S. fonticola 10 5 3S. odorifera 3 3 9 1S. plymuthica 1 5 21 5S. rubidaea 9 7 5 2
cefepime 0.03–64 S. ficaria 5 9 1S. fonticola 10 8S. odorifera 6 9 1
Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022
Antimicrobial susceptibility of unusual Serratia spp.
875
Table 2. (Continued)
Concentrations examined (mg/L)
Number of strains with MIC (mg/L)
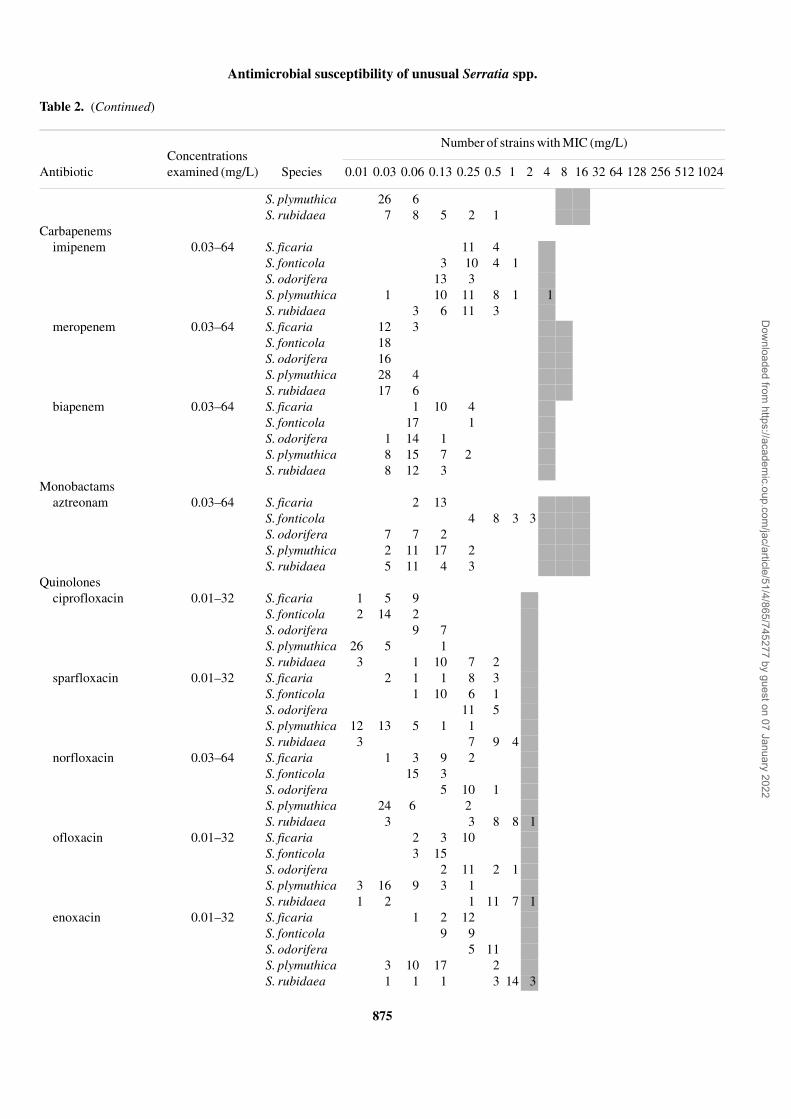
Antibiotic Species 0.01 0.03 0.06 0.13 0.25 0.5 1 2 4 8 16 32 64 128 256 512 1024
S. plymuthica 26 6S. rubidaea 7 8 5 2 1
Carbapenemsimipenem 0.03–64 S. ficaria 11 4
S. fonticola 3 10 4 1S. odorifera 13 3S. plymuthica 1 10 11 8 1 1S. rubidaea 3 6 11 3
meropenem 0.03–64 S. ficaria 12 3S. fonticola 18S. odorifera 16S. plymuthica 28 4S. rubidaea 17 6
biapenem 0.03–64 S. ficaria 1 10 4S. fonticola 17 1S. odorifera 1 14 1S. plymuthica 8 15 7 2S. rubidaea 8 12 3
Monobactamsaztreonam 0.03–64 S. ficaria 2 13
S. fonticola 4 8 3 3S. odorifera 7 7 2S. plymuthica 2 11 17 2S. rubidaea 5 11 4 3
Quinolonesciprofloxacin 0.01–32 S. ficaria 1 5 9
S. fonticola 2 14 2S. odorifera 9 7S. plymuthica 26 5 1S. rubidaea 3 1 10 7 2
sparfloxacin 0.01–32 S. ficaria 2 1 1 8 3S. fonticola 1 10 6 1S. odorifera 11 5S. plymuthica 12 13 5 1 1S. rubidaea 3 7 9 4
norfloxacin 0.03–64 S. ficaria 1 3 9 2S. fonticola 15 3S. odorifera 5 10 1S. plymuthica 24 6 2S. rubidaea 3 3 8 8 1
ofloxacin 0.01–32 S. ficaria 2 3 10S. fonticola 3 15S. odorifera 2 11 2 1S. plymuthica 3 16 9 3 1S. rubidaea 1 2 1 11 7 1
enoxacin 0.01–32 S. ficaria 1 2 12S. fonticola 9 9S. odorifera 5 11S. plymuthica 3 10 17 2S. rubidaea 1 1 1 3 14 3
Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022
I. Stock et al.
876
Table 2. (Continued)
Concentrations examined (mg/L)
Number of strains with MIC (mg/L)
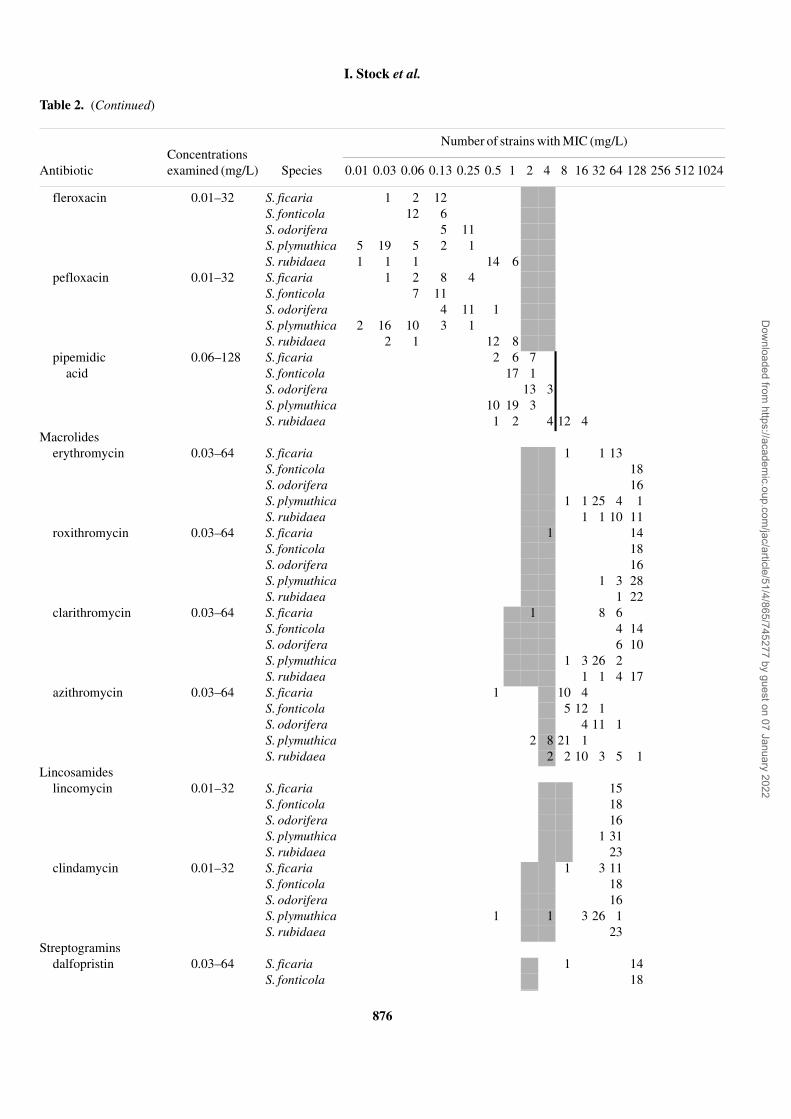
Antibiotic Species 0.01 0.03 0.06 0.13 0.25 0.5 1 2 4 8 16 32 64 128 256 512 1024
fleroxacin 0.01–32 S. ficaria 1 2 12S. fonticola 12 6S. odorifera 5 11S. plymuthica 5 19 5 2 1S. rubidaea 1 1 1 14 6
pefloxacin 0.01–32 S. ficaria 1 2 8 4S. fonticola 7 11S. odorifera 4 11 1S. plymuthica 2 16 10 3 1S. rubidaea 2 1 12 8
pipemidic acid
0.06–128 S. ficaria 2 6 7S. fonticola 17 1S. odorifera 13 3S. plymuthica 10 19 3S. rubidaea 1 2 4 12 4
Macrolideserythromycin 0.03–64 S. ficaria 1 1 13
S. fonticola 18S. odorifera 16S. plymuthica 1 1 25 4 1S. rubidaea 1 1 10 11
roxithromycin 0.03–64 S. ficaria 1 14S. fonticola 18S. odorifera 16S. plymuthica 1 3 28S. rubidaea 1 22
clarithromycin 0.03–64 S. ficaria 1 8 6S. fonticola 4 14S. odorifera 6 10S. plymuthica 1 3 26 2S. rubidaea 1 1 4 17
azithromycin 0.03–64 S. ficaria 1 10 4S. fonticola 5 12 1S. odorifera 4 11 1S. plymuthica 2 8 21 1S. rubidaea 2 2 10 3 5 1
Lincosamideslincomycin 0.01–32 S. ficaria 15
S. fonticola 18S. odorifera 16S. plymuthica 1 31S. rubidaea 23
clindamycin 0.01–32 S. ficaria 1 3 11S. fonticola 18S. odorifera 16S. plymuthica 1 1 3 26 1S. rubidaea 23
Streptograminsdalfopristin 0.03–64 S. ficaria 1 14
S. fonticola 18
Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022

Antimicrobial susceptibility of unusual Serratia spp.
877
Table 2. (Continued)
Concentrations examined (mg/L)
Number of strains with MIC (mg/L)
Antibiotic Species 0.01 0.03 0.06 0.13 0.25 0.5 1 2 4 8 16 32 64 128 256 512 1024
S. odorifera 16S. plymuthica 1 1 30S. rubidaea 23
quinupristin 0.03–64 S. ficaria 15S. fonticola 18S. odorifera 16S. plymuthica 1 31S. rubidaea 23
dalfopristin/quinupristin
0.03–64 S. ficaria 1 1 13S. fonticola 18S. odorifera 16S. plymuthica 1 1 1 29S. rubidaea 23
Antifolatessulfamethoxazole 0.25–512 S. ficaria 2 3 5 2 3
S. fonticola 2 6 7 2 1S. odorifera 5 2 4 5S. plymuthica 1 1 2 4 5 10 5 3 1S. rubidaea 1 5 2 7 1 7
trimethoprim 0.03–64 S. ficaria 2 2 2 2 6 1S. fonticola 14 4S. odorifera 4 7 5S. plymuthica 7 19 5 1S. rubidaea 1 1 1 4 6 7 3
trimethoprim/sulfamethoxazole
0.13–256 S. ficaria 5 9 1S. fonticola 6 11 1S. odorifera 3 9 4S. plymuthica 3 15 13 1S. rubidaea 3 4 6 8 2
Glycopeptidesteicoplanin 0.06–128 S. ficaria 15
S. fonticola 18S. odorifera 16S. plymuthica 1 31S. rubidaea 23
vancomycin 0.03–64 S. ficaria 15S. fonticola 18S. odorifera 16S. plymuthica 1 31S. rubidaea 23
Other antibioticschloramphenicol 0.06–128 S. ficaria 1 4 7 3
S. fonticola 12 6S. odorifera 15 1S. plymuthica 5 10 12 4 1S. rubidaea 1 2 10 10
nitrofurantoin 0.13–256 S. ficaria 2 3 9 1S. fonticola 6 12S. odorifera 5 5 4 2
Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022

I. Stock et al.
878
Medium dependency
For most of the antibiotics tested, there were either no or onlyminor differences in susceptibility depending on the medium(one doubling dilution step at the maximum). For all thespecies, the MICs of macrolides and tetracyclines weregenerally one to two doubling dilution steps and the MICs offosfomycin one to four (depending on the species and the indi-vidual strain) doubling dilutions higher in IsoSensitest broththan in CAMHB (data not shown). This led to an altered clin-ical assessment of the respective natural populations of somespecies to some of these antibiotics (Tables 2 and 3). Forexample, in CAMHB all Serratia spp. tested were naturallysusceptible to fosfomycin. Apart from fosfomycin, species-related medium dependencies in susceptibility testing werefound with some β-lactams, in particular certain cephalosporins(data not shown). In IsoSensitest broth, some amoxicillin-sensitive and -intermediate strains of S. ficaria, S. plymuthicaand S. rubidaea were generally more susceptible to cefaclor,cefazolin and loracarbef (one to two doubling dilution steps)than in CAMHB (the MICs of aminopenicillins were similarin both media.) The MICs of cefixime, cefdinir, ceftibuten,cefotiam, cefuroxime and ceftriaxone for some amoxicillin-sensitive or -intermediate strains of S. rubidaea were also one
to two doubling dilution steps higher in CAMHB than in Iso-Sensitest broth.
Quality assurance
The MICs of all antibiotics were reproducible for S. ficariaATCC 33105T, S. fonticola ATCC 29844T, S. odoriferaATCC 33077T, S. plymuthica ATCC 183T, S. rubidaea ATCC27593T and E. coli ATCC 25922. For certain β-lactams theMIC ranges were broader for reference strains of Serratiathan for E. coli ATCC 25922. With few exceptions, the MICsfor E. coli ATCC 25922 in IsoSensitest broth and CAMHBwere within the control limits for susceptibility testingaccording to DIN and NCCLS criteria (data not shown).
β-Lactamase activity and induction assay
All serratiae examined produced β-lactamase(s). Theβ-lactamases of S. ficaria, S. fonticola and S. odorifera strainswere inducible, whereas β-lactamase activities in S. rubidaeaand in four of five strains of S. plymuthica after induction weresimilar to those measured without an inducer. Ranges ofspecific β-lactamase activity (mmol/L·mg) were as follows:0.17–0.54 (not induced) versus 1.6–19 (induced) for S. ficaria
Table 2. (Continued)
aThe number of strains with the corresponding MIC is cited. Strains in the column for the lowest concentration of the antibiotic (Cmin) have MICs ≤ this lowestconcentration (MIC = Cmin → MIC ≤ Cmin). MICs higher than the highest concentration tested were assigned to twice the highest concentration which was tested.MICs in shaded areas indicate the clinically intermediate area according to the standard applied. A thick black line indicates the breakpoint between clinicallysusceptible and clinically resistant strains, if the ‘intermediate’ does not exist.
Concentrations examined (mg/L)
Number of strains with MIC (mg/L)
Antibiotic Species 0.01 0.03 0.06 0.13 0.25 0.5 1 2 4 8 16 32 64 128 256 512 1024
S. plymuthica 1 2 5 23 1S. rubidaea 7 11 4 1
rifampicin 0.01–32 S. ficaria 2 9 4S. fonticola 4 2 1 11S. odorifera 3 13S. plymuthica 1 3 16 12S. rubidaea 1 3 6 2 11
fosfomycin 0.13–256 S. ficaria 2 2 6 5S. fonticola 4 7 7S. odorifera 1 6 7 2S. plymuthica 1 2 5 4 2 2 4 8 1 1 2S. rubidaea 1 9 3 1 4 3 1 1
fusidic acid 0.01–32 S. ficaria 1 1 13S. fonticola 18S. odorifera 16S. plymuthica 1 2 29S. rubidaea 1 22
Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022

Antimicrobial susceptibility of unusual Serratia spp.
879
[induction factor (IF) 6–>100]; 0.55–2.34 versus 2.9–45 forS. fonticola (IF 4–32); 0.78–1.0 versus 5.0–8.5 for S. odo-rifera (IF 6–9); 0.66–2.0 versus 0.62–2.8 for S. rubidaea(IF 0.9–1.4); 1.0–1.2 versus 1.2–1.6 for four of five strainsof S. plymuthica (IF 1.2–1.4); and 0.87 versus 2.4 (IF 3) forS. plymuthica LMG 6823. As expected, β-lactamases ofS. marcescens CCUG 6 and S. liquefaciens CCUG 5159 wereinducible [specific activities were 0.34 versus 48 mmol/L·mg(IF > 100) and 1.5 versus 4.4 mmol/L·mg (IF 3), respectively];S. enterica ATCC 13311 expressed no β-lactamase.
β-Lactamase detection by SDS–PAGE
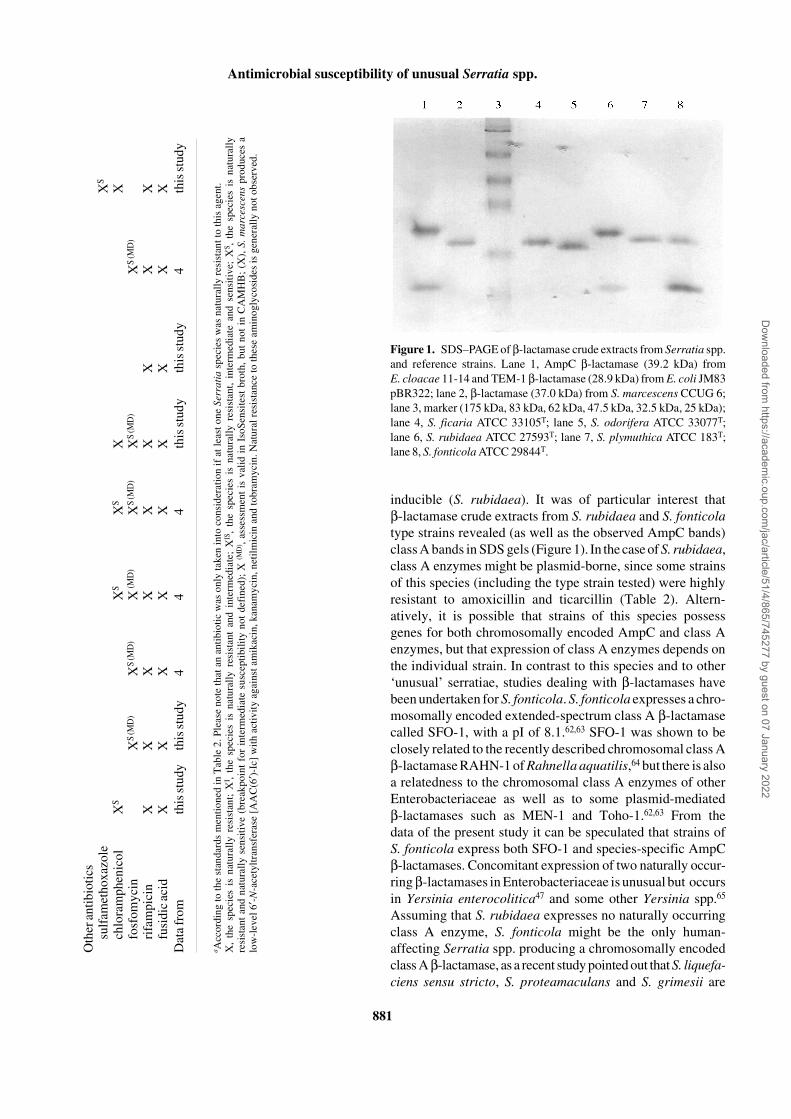
The β-lactamases of the type strains of all tested Serratia spp.showed distinct bands estimating molecular weights between37 and 40 kDa, characteristic for the expression of AmpCenzymes (Figure 1). In addition, a further β-lactamase bandestimating a mol. wt <30 kDa, indicating the expression of aclass A enzyme, was found in S. fonticola ATCC 29844T andS. rubidaea ATCC 27593T.
AmpC gene detection by PCR
PCR amplification products for ampC were obtained for allthe strains tested (data not shown). PCR fragments ranged insize from 750 to 800 bp, as was to be expected based on previ-ous experiments with AmpC genes.58
Discussion
Although Serratia spp. other than S. marcescens were firstdescribed several decades ago, they have barely beenexamined in many respects. The main reason for this lack ofdata in regard to pathogenicity, antimicrobial susceptibilityand mechanisms of antibiotic resistance might be the leadingrole of S. marcescens in human infections caused by Serratiaspp.1 However, the occurrence of all Serratia spp. in clinicalspecimens, the recently described virulence-associated pro-perties in Serratia strains other than S. marcescens, theincreasing number of documented infections as well as themisidentification or non-identifiability of these bacteria bycommercial systems strongly suggest their implication inhuman disease.
Natural antibiotic susceptibility patterns of this studyshowed that ‘unusual’ Serratia spp. are naturally resistant tonumerous, in part structurally unrelated antimicrobial agents,indicating a range of different mechanisms of resistance.Because some phenotypic resistance patterns depended onthe species, it is obvious that the respective underlying mech-anisms must be species specific.
Natural susceptibility patterns to β-lactams indicated thatall Serratia spp. examined are likely to produce chromo-somally encoded β-lactamases, which are specific for thespecies. Generally, phenotypic analysis of MIC susceptibility
patterns of appropriate key β-lactams gives evidence aboutthe β-lactamase type expressed, which in Enterobactericeaebelongs to either class A or class C (AmpC). Examinationsconcerning chromosomally encoded β-lactamases of Serra-tia spp. have been focused on S. marcescens, which producesa well-characterized AmpC β-lactamase.60,61 In S. marcescensand many other Enterobacteriaceae, chromosomal AmpCexpression is reflected by resistance or decreased suscep-tibilities to amoxicillin, the same susceptibilities to amino-penicillins in the presence of clavulanic acid as to amoxicillinalone, natural resistances to narrow-spectrum cephalosporins(e.g. cefaclor, loracarbef and cefazolin) and natural ticarcillinsensitivity.3,4 Surprisingly, in this study it was shown thatnumerous strains of S. odorifera, S. plymuthica and S. rubidaeawere naturally sensitive to amoxicillin, amoxicillin/clavulanate and ticarcillin but resistant to narrow-spectrumcephalosporins including cefuroxime (Tables 2 and 3). To ourknowledge, this phenotypic pattern is unique among Entero-bacteriaceae species and points, together with species-relateddifferences in susceptibility to ceftibuten and cefoxitin, tothe expression of unusual species-specific AmpC enzymes.Sensitivity to amoxicillin but resistance to narrow-spectrumcephalosporins has only been reported for some strains ofS. plymuthica,31,32 but research in the underlying mechanismswas not performed either for S. odorifera, S. plymuthica orS. rubidaea. In contrast to the β-lactamases of these species,the enzyme of S. ficaria seems to be a more common AmpCβ-lactamase, indicated by the natural resistance of severalstrains to amoxicillin and amoxicillin/clavulanate, and thespecies-associated high-level resistance to cefazolin andcefoxitin (Table 2). Studies dealing with the naturallyoccurring β-lactamase of S. ficaria have not been published.The broad variety of different β-lactamases within ‘unusual’serratiae is completed by S. fonticola. With respect to Serratiaspp., the unique β-lactam susceptibility pattern of S. fonticola,showing natural resistance to amoxicillin, ticarcillin andseveral cephalosporins, but natural sensitivity to amoxicillin/clavulanate, points to the expression of a chromosomallyencoded class A β-lactamase with an enhanced cephalo-sporinase activity.
To give an approach to the molecular basis of the suscep-tibility patterns, the β-lactamases of representative strains ofeach species were characterized phenotypically and geno-typically. PCR and SDS–PAGE revealed that the type strainsof the Serratia species examined possess ampC and alsoexpress these enzymes (Figure 1). Species specificity of theseenzymes was not only reflected by MICs, but also by differentapparent molecular weights of the β-lactamase bands in theSDS gels (Figure 1), and by the data obtained from the β-lactamase activity and induction assays. From these data itcan be concluded that each Serratia sp. examined expressesits own naturally occurring AmpC β-lactamase, which mightbe inducible (S. ficaria, S. fonticola, S. odorifera) or not
Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022
I. Stock et al.
880
Table 3. Natural antibiotic resistance of Serratia spp.a
Antibiotic S. ficaria S. fonticola S. grimesii S. liquefaciens S. marcescens S. odorifera S. plymuthica S. proteamaculans S. rubidaea
Tetracyclinestetracycline XI (MD) XI (MD) X (MD) X (MD) X (MD)
doxycyline XI (MD) XI (MD) X (MD)
minocycline XI (MD)
Aminoglycosides amikacin (X)kanamycin (X)netilmicin (X)tobramycin (X)spectinomycin XI XIS
ribostamycin XIS
Penicillinsbenzylpenicillin X X X X X X X X Xoxacillin X X X X X X X X Xamoxicillin XI X XI XI Xco-amoxiclav XI XI XI Xampicillin/sulbactam XI XI
ticarcillin X
Cephalosporinscefaclor X X X X X XI X X XI
cefazolin X X X X X X X X XI
loracarbef XI X XI XI X XIS
cefuroxime X X X X X X X X Xcefoxitin X XI XI X XIS
cefpodoxime XIS
cefdinir XI XIS
Quinolonespipemidic acid XS
Macrolideserythromycin X X X X X X X X Xroxithromycin X X X X X X X X Xclarithromycin X X X X X X X X Xazithromycin X (MD) X (MD) XI (MD) X (MD) X (MD) X XI (MD) XI (MD) X (MD)
Lincosamides, streptogramins, glycopeptidesall antibiotics X X X X X X X X X
Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022
Antimicrobial susceptibility of unusual Serratia spp.
881
inducible (S. rubidaea). It was of particular interest thatβ-lactamase crude extracts from S. rubidaea and S. fonticolatype strains revealed (as well as the observed AmpC bands)class A bands in SDS gels (Figure 1). In the case of S. rubidaea,class A enzymes might be plasmid-borne, since some strainsof this species (including the type strain tested) were highlyresistant to amoxicillin and ticarcillin (Table 2). Altern-atively, it is possible that strains of this species possessgenes for both chromosomally encoded AmpC and class Aenzymes, but that expression of class A enzymes depends onthe individual strain. In contrast to this species and to other‘unusual’ serratiae, studies dealing with β-lactamases havebeen undertaken for S. fonticola. S. fonticola expresses a chro-mosomally encoded extended-spectrum class A β-lactamasecalled SFO-1, with a pI of 8.1.62,63 SFO-1 was shown to beclosely related to the recently described chromosomal class Aβ-lactamase RAHN-1 of Rahnella aquatilis,64 but there is alsoa relatedness to the chromosomal class A enzymes of otherEnterobacteriaceae as well as to some plasmid-mediatedβ-lactamases such as MEN-1 and Toho-1.62,63 From thedata of the present study it can be speculated that strains ofS. fonticola express both SFO-1 and species-specific AmpCβ-lactamases. Concomitant expression of two naturally occur-ring β-lactamases in Enterobacteriaceae is unusual but occursin Yersinia enterocolitica47 and some other Yersinia spp.65
Assuming that S. rubidaea expresses no naturally occurringclass A enzyme, S. fonticola might be the only human-affecting Serratia spp. producing a chromosomally encodedclass A β-lactamase, as a recent study pointed out that S. liquefa-ciens sensu stricto, S. proteamaculans and S. grimesii are
a Acc
ordi
ng to
the
stan
dard
s m
enti
oned
in T
able
2. P
leas
e no
te th
at a
n an
tibi
otic
was
onl
y ta
ken
into
con
side
rati
on if
at l
east
one
Ser
rati
a sp
ecie
s w
as n
atur
ally
res
ista
nt to
this
age
nt.
X,
the
spec
ies
is n
atur
ally
res
ista
nt;
XI ,
the
spec
ies
is n
atur
ally
res
ista
nt a
nd i
nter
med
iate
; X
IS,
the
spec
ies
is n
atur
ally
res
ista
nt,
inte
rmed
iate
and
sen
siti
ve;
XS ,
the
spe
cies
is
natu
rall
yre
sist
ant
and
natu
rall
y se
nsit
ive
(bre
akpo
int
for
inte
rmed
iate
sus
cept
ibil
ity
not
defi
ned)
; X
(MD
) , as
sess
men
t is
val
id i
n Is
oSen
site
st b
roth
, bu
t no
t in
CA
MH
B;
(X),
S.
mar
cesc
ens
prod
uces
alo
w-l
evel
6′ - N
-ace
tylt
rans
fera
se [
AA
C(6
′ )-Ic
] w
ith
acti
vity
aga
inst
am
ikac
in, k
anam
ycin
, net
ilm
icin
and
tobr
amyc
in. N
atur
al r
esis
tanc
e to
thes
e am
inog
lyco
side
s is
gen
eral
ly n
ot o
bser
ved.
Oth
er a
ntib
ioti
cssu
lfam
etho
xazo
leX
S
chlo
ram
phen
icol
XS
XS
XS
XX
fosf
omyc
inX
S (M
D)
XS
(MD
)X
(MD
)X
S (M
D)
XS
(MD
)X
S (M
D)
rifa
mpi
cin
XX
XX
XX
XX
Xfu
sidi
c ac
idX
XX
XX
XX
XX
Dat
a fr
omth
is st
udy
this
stud
y4
44
this
stud
yth
is st
udy
4th
is st
udy
Figure 1. SDS–PAGE of β-lactamase crude extracts from Serratia spp.and reference strains. Lane 1, AmpC β-lactamase (39.2 kDa) fromE. cloacae 11-14 and TEM-1 β-lactamase (28.9 kDa) from E. coli JM83pBR322; lane 2, β-lactamase (37.0 kDa) from S. marcescens CCUG 6;lane 3, marker (175 kDa, 83 kDa, 62 kDa, 47.5 kDa, 32.5 kDa, 25 kDa);lane 4, S. ficaria ATCC 33105T; lane 5, S. odorifera ATCC 33077T;lane 6, S. rubidaea ATCC 27593T; lane 7, S. plymuthica ATCC 183T;lane 8, S. fonticola ATCC 29844T.
Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022

I. Stock et al.
882
likely to produce chromosomal AmpC β-lactamases.4 Thisfinding is in surprising agreement with the particular taxo-nomic position of S. fonticola within the genus Serratia:although it belongs clearly to the genus Serratia,19 DNA–DNA hybridization studies revealed a relatively low level ofDNA relatedness to other Serratia species.66
Apart from the β-lactams, there were significant species-related differences in natural susceptibility to several otherantibiotics. It was shown that there are differences in suscep-tibility to tetracyclines, the molecular basis of which is un-known. The natural and acquired tetracycline resistance inEnterobacteriacecae species is mainly due to the expressionof tetracycline-specific efflux (Tet) proteins.67 In addition,more or less non-specific multidrug transport systems and theouter membrane contribute to resistance to these antibiotics.68
It is likely that the natural low-level tetracycline resistanceof S. ficaria, S. odorifera and S. rubidaea is attributed to amultidrug transport system rather than to a chromosomallyencoded Tet protein expressed at low levels. In contrast toS. fonticola and S. plymuthica, these species showed de-creased susceptibilities to chloramphenicol and to the quino-lones tested, indicative of a multidrug efflux mechanism.68
Supporting this idea, the species-dependent levels of tetra-cycline resistance corresponded to the respective levels of thequinolone and chloramphenicol MICs (Table 2). Naturallyoccurring low-level resistance to tetracyclines combined witha decreased susceptibility to quinolones and chloramphenicolseems to be a typical feature of several Serratia spp. and islikely to be restricted to the genus Serratia, since this pheno-type has been described recently in S. marcescens, but not inthe S. liquefaciens group4 or in other Enterobacteriaceae.
Further interesting species-related differences in naturalsusceptibilty were seen with some aminoglycosides. Althoughthere was no uniform natural resistance to any amino-glycoside, the decreased susceptibility of S. odorifera tostreptomycin and spectinomycin and the decreased suscep-tibility of S. rubidaea to streptomycin, spectinomycin,kanamycin and ribostamycin indicates the presence of severalspecies-specific aminoglycoside-modifying enzymes (AMEs)expressed at low levels. With respect to the underlyingsusceptibility patterns, it can be hypothesized that S. odo-rifera expresses a chromosomally encoded enzyme identicalor similar to ANT (3′′-I), which is the only described AMEwith an activity against streptomycin and spectinomycin andis widely distributed in Gram-negative bacteria.69 The pheno-typic susceptibility pattern of S. rubidaea is difficult toexplain. It is possible that strains of these species express anANT (3′′-I) enzyme in addition to a further naturally occur-ring AME. However, in contrast to our data, all known AMEswith activity against kanamycin and ribostamycin are alsoactive against neomycin.69 An assumed novel AME shouldtherefore have no activity against neomycin or should beactive against all four substrates, provided that ANT (3′′-I)
is not present in S. rubidaea. Data similar to the results ofthis study were found in our recent study with respect toS. marcescens.4 The decreased susceptibility of S. marces-cens to amikacin, kanamycin, netilmicin and tobramycinwithout any obvious resistance was in agreement with the fea-tures of its AAC(6′)-Ic enzyme, a chromosomal 6′-N-acetyl-transferase,70,71 expressed naturally at low levels.72
In conclusion, the data presented in this study compriseinformation about the natural susceptibility to a wide rangeof antimicrobial agents of S. ficaria, S. fonticola, S. odorifera,S. plymuthica and S. rubidaea. These data can be applied tovalidate forthcoming susceptibility data of these serratiae andmight contribute to their reliable identification, supported bya number of species-related differences in antibiotic sus-ceptibility. Evidence of novel mechanisms of resistance, inparticular species-specific β-lactamases that have beendescribed in the present study for the first time, should alsodirect the interest in research on ‘unusual’ Serratia spp. forwhich isolation from clinical specimens can no longer beregarded as anecdotal1 and which are likely to be importanthuman pathogens.
Acknowledgements
Thank you very much to all who put their strains at our dis-posal. The technical assistance of Gerrit Loch and Jens Illigenis gratefully acknowledged. This study was supported byMerlin-Diagnostika, Bornheim, Germany.
References
1. Grimont, F. & Grimont, P. A. D. (1992). The genus Serratia. InThe Prokaryotes. A Handbook on the Biology of Bacteria: Eco-physiology, Isolation, Identification, Applications (Balows, A.,Trüper, H. G., Dworkin, M., Harder, W. & Schleifer, K. H., Eds),Vol. 1, pp. 2822–48. Springer, New York, NY, USA.
2. Anahory, T., Darbas, H., Ongaro, O., Jean-Pierre, H. & Mion, P.(1998). Serratia ficaria: a misidentified or unidentified rare cause ofhuman infections in fig tree culture zones. Journal of Clinical Micro-biology 36, 3266–72.
3. Livermore, D. M. (1995). β-Lactamases in laboratory andclinical resistance. Clinical Microbiology Reviews 8, 557–84.
4. Stock, I., Grüger, T. & Wiedemann, B. (2003). Natural antibioticsusceptibility of strains of Serratia marcescens and the S. lique-faciens complex: S. liquefaciens sensu stricto, S. proteamaculansand S. grimesii. International Journal of Antimicrobial Agents, inpress.
5. Angerer, A., Klupp, B. & Braun, V. (1992). Iron transportsystems of Serratia marcescens. Journal of Bacteriology 174,1378–87.
6. Kalbe, C., Marten, P. & Berg, G. (1996). Strains of the genusSerratia as beneficial rhizobacteria of oilseed rape with antifungalproperties. Microbiological Research 151, 433–9.
Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022

Antimicrobial susceptibility of unusual Serratia spp.
883
7. Martinez-Garcia, E., Tormo, A. & Navarro-Llorens, J. M. (2001).Further studies on RpoS in enterobacteria: identification of rpoS inEnterobacter cloacae and Kluyvera cryocrescens. Archives ofMicrobiology 175, 395–404.
8. Singh, B. R., Singh, Y. & Tiwari, A. K. (1997). Characterizationof virulence factors of Serratia strains isolated from foods. Inter-national Journal of Food Microbiology 34, 259–66.
9. Grimont, P. A. D., Grimont, F. & Starr, M. P. (1979). Serratiaficaria sp. nov., a bacterial species associated with smyrna figs andthe fig wasp Blastophaga psenes. Current Microbiology 2, 277–82.
10. Badenoch, P. R., Thom, A. L. & Coster, D. J. (2002). Serratiaficaria endophthalmitis. Journal of Clinical Microbiology 40, 1563–4.
11. Bollet, C., Freney, J., de Micco, P. & Grimont, P. A. D. (1990).Un nouveau case de contamination par Serratia ficaria. Revue de lalitérature et écologie. Médecine et Maladies Infectieuses 20,97–100.
12. Brouillard, J. A., Hansen, W. & Compere, A. (1984). Isolation ofSerratia ficaria from human clinical specimens. Journal of ClinicalMicrobiology 19, 902–4.
13. Darbas, H., Jean-Pierre, H., Boyer, G. & Riviere, M. (1993).Pyocholécyste à Serratia ficaria. Médecine et Maladies Infectieuses23, 269–70.
14. Darbas, H., Jean-Pierre, H. & Paillisson, J. (1994). Case reportand review of septicemia due to Serratia ficaria. Journal of ClinicalMicrobiology 32, 2285–8.
15. Gill, V. J., Farmer, J. J., III, Grimont, P. A. D., Asbury, M. A. &McIntosh, C. L. (1981). Serratia ficaria isolated from clinical speci-men. Journal of Clinical Microbiology 14, 234–6.
16. Pien, F. D. & Farmer, J. J., III (1983). Serratia ficaria isolatedfrom a leg ulcer. Southern Medical Journal 76, 1591–2.
17. Richard, C., de Coquet, J. & Suc, C. (1989). Serratia ficaria:mise au point á propos du premier isolement chez l’homme enFrance. Médecine et Maladies Infectieuses 19, 45–7.
18. Gavini, F., Ferragut, C., Izard, D., Trinel, P. A., Leclerc, H.,Lefebvre, B. et al. (1979). Serratia fonticola, a new species fromwater. International Journal of Systematic and Evolutionary Micro-biology 29, 92–101.
19. Müller, H. E., Fanning, G. R. & Brenner, D. J. (1995). Isolationof Serratia fonticola from mollusks. Systematic and Applied Micro-biology 18, 279–84.
20. Müller, H. E., Steigerwalt, A. G. & Brenner, D. J. (1986). Isola-tion of Serratia fonticola from birds. Zentralblatt für Bakteriologie,Mikrobiologie und Hygiene (Serie A) 261, 212–8.
21. Bollet, C., Gainnier, M., Sainty, J. M., Orbesser, P. & De Micco,P. (1991). Serratia fonticola isolated from a leg abscess. Journal ofClinical Microbiology 29, 834–5.
22. Farmer III, J. J., Davis, B. R., Hickman-Brenner, F. W.,McWhorther, A., Huntley-Carter, G. P., Asbury, M. A. et al. (1985).Biochemical identification of new species and biogroups of Entero-bacteriaceae isolated from clinical specimens. Journal of ClinicalMicrobiology 21, 46–76.
23. Grimont, P. A. D., Grimont, F., Richard, C., Davis, B. R.,Steigerwalt, A. G. & Brenner, D. J. (1978). Deoxyribonucleic acidrelatedness between Serratia plymuthica and other Serratiaspecies, with a description of Serratia odorifera sp. nov. (Type strain
ICPB 3995). International Journal of Systematic and EvolutionaryMicrobiology 28, 453–63.
24. Cook, M. A. & Lopez, J. J., Jr (1998). Serratia odoriferabiogroup I: an emerging pathogen. Journal of the American Osteo-pathology Association 98, 505–7.
25. Ceccaldi, B., Cariou, B., Soullie, B., Le Berre, J. P., Mayaudon,H., Dupuy, O. et al. (1999). Serratia odorifera biovar 1 bronchialinfection. Revue de Médecine Interne 20, 84–5.
26. Chmel, H. (1988). Serratia odorifera biogroup 1 causing aninvasive human infection. Journal of Clinical Microbiology 26, 1244–5.
27. Frean, J. A., Arntzen, L., Rosekilly, I. & Isaacson, M. (1994).Investigation of contaminated parenteral nutrition fluids associatedwith an outbreak of Serratia odorifera septicemia. Journal of Hospi-tal Infection 27, 263–73.
28. Glustein, J. Z., Rudensky, B. & Abrahamov, A. (1994).Catheter-associated sepsis caused by Serratia odorifera biovar 1 inan adolescent patient. European Journal of Clinical Microbiologyand Infectious Diseases 13, 183–4.
29. Mermel, L. A. & Spiegel, C. A. (1992). Nosocomial sepsisdue to Serratia odorifera biovar 1. Clinical Infectious Diseases 14,208–10.
30. Sader, H. S., Perl, T. M., Hollis, R. J., Divishek, D., Herwaldt, L.A. & Jones, R. N. (1994). Nosocomial transmission of Serratiaodorifera biogroup 2: case report demonstration by macrorestrictionanalysis of chromosomal DNA using pulsed-field gel electro-phoresis. Infection Control and Hospital Epidemiology 15, 390–3.
31. Berg, G. (2000). Diversity of antifungal and plant-associatedSerratia plymuthica strains. Journal of Applied Microbiology 88,952–60.
32. Carrero, P., Garrote, J. A., Pacheco, S., Garcia, A. I., Gil, R. &Carbajosa, S. G. (1995). Report of six cases of human infection bySerratia plymuthica. Journal of Clinical Microbiology 33, 275–6.
33. Clark, R. B. & Janda, J. M. (1985). Isolatation of Serratiaplymuthica from human burn site. Journal of Clinical Microbiology21, 656–7.
34. Domingo, D., Limia, A., Alarcón, T., Sanz, J. C., Del Rey, M.-C.& López-Brea, M. (1994). Nosocomial septicemia caused bySerratia plymuthica. Journal of Clinical Microbiology 32, 575–7.
35. Horowitz, H. W., Nadelman, R. B., Van Horn, K. G., Weekews,S. E., Goyburu, L. & Wormser, G. P. (1987). Serratia plymuthicasepsis associated with infections of central venous catheter. Journalof Clinical Microbiology 25, 1562–3.
36. Martinez, J. & Carrascosa, M. (1997). Shock séptico porSerratia plymuthica. Enfermedades Infecciosas y MicrobiologiaClinica 15, 114–5.
37. Nough, F. & Bhandari, S. (2000). CAPD peritonitis due toSerratia plymuthica. Peritoneal Dialysis International 20, 349.
38. Reina, J., Borell, N. & Lompart, I. (1992). Community-acquiredbacteremia caused by Serratia plymuthica. Case report and reviewof the literature. Diagnostic Microbiology and Infectious Disease 15,449–52.
39. Zbinden, R. & Blass, R. (1988). Serratia plymuthica osteomyelitisfollowing a motorcycle accident. Journal of Clinical Microbiology 26,1409–10.
Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022

I. Stock et al.
884
40. Grimont, P. A. D., Grimont, F. & Starr, M. P. (1981). Serratiaspecies isolated from plants. Current Microbiology 5, 317–22.
41. Bollet, C., Giulian, C., Mallet, M. N., Estrangin, E. & deMicco, P.(1989). Isolation of Serratia rubidaea (Stapp) from fresh and spoiledcoconuts. Tropical Agriculture 66, 342–5.
42. Okada, T., Yokota, E. & Matsumoto, I. (2002). Communityacquired sepsis by Serratia rubidaea (article in Japanese). Kansen-shogaku Zasshi 76, 109–12.
43. Ursua, P. R., Unzaga, M. J., Melero, P., Iturburu, I., Ezpeleta, C.& Cisterna, R. (1996). Serratia rubidaea as an invasive pathogen.Journal of Clinical Microbiology 34, 216–7.
44. Grimont, P. A. D., Jackson, T. A., Ageron, E. & Noonan, M. J.(1988). Serratia entomophila sp. nov. associated with amber dis-ease in the New Zealand grass grub Costelytra zealandica. Inter-national Journal of Systematic and Evolutionary Bacteriology 38,1–6.
45. Stock, I. & Wiedemann, B. (2001). Natural antibiotic sus-ceptibility of Klebsiella pneumoniae, K. oxytoca, K. planticola,K. ornithinolytica and K. terrigena strains. Journal of Medical Micro-biology 50, 396–406.
46. Stock, I. & Wiedemann, B. (1998). Natural antibiotic susceptibil-ity of Providencia stuartii, P. rettgeri, P. alcalifaciens and P. rustigia-nii strains. Journal of Medical Microbiology 47, 629–42.
47. Stock, I. & Wiedemann, B. (2000). Natural β-lactam suscep-tibility and mechanisms of β-lactam resistance in Yersinia entero-colitica. Reviews in Medical Microbiology 11, 197–210.
48. Deutsches Institut für Normung e.V. (1998). Susceptibilitytesting of pathogens to antimicrobial agents, part 4: evaluationclasses of the minimum inhibitory concentration, supplement 1:MIC breakpoints of antibacterial agents. DIN document 58940–4(Table 1). Deutsches Institut für Normung e.V., Berlin, Germany.
49. Comité de l’Antibiogramme de la Société Française de Micro-biologie Communiqué. (1998). Statement. Pathologie Biologie 46,I–XVI.
50. Olsson-Liljequist, B., Larson, P., Walder, M. & Miörner, H.(1997). Antimicrobial susceptibility testing in Sweden. Part 3:methodology for susceptibility testing. Scandinavian Journal ofInfectious Diseases 105, Suppl., 13–23.
51. National Committee for Clinical Laboratory Standards. (2000).Minimum inhibitory concentration (MIC) interpretive standards(µg/ml) for Enterobacteriaceae. NCCLS document M7-A5 (Table 2A).National Committee for Clinical Laboratory Standards, Villanova,PA, USA.
52. National Committee for Clinical Laboratory Standards. (2000).Minimum inhibitory concentration (MIC) interpretive standards(µg/ml) for Neisseria gonorrhoe. NCCLS document M7-A5(Table 2F). National Committee for Clinical Laboratory Standards,Villanova, PA, USA.
53. National Committee for Clinical Laboratory Standards. (2000).Minimum inhibitory concentration (MIC) interpretive standards(µg/ml) for Staphylococcus spp. NCCLS document M7-A5(Table 2C). National Committee for Clinical Laboratory Standards,Villanova, PA, USA.
54. Troxler, R., von Graevenitz, A., Funke, G., Wiedemann, B. &Stock, I. (2000). Natural antibiotic susceptibility of Listeria species:L. grayi, L. innocua, L. ivanovii, L. monocytogenes, L. seeligeri andL. welshimeri strains. Clinical Microbiology and Infection 6, 525–35.
55. Peter, K., Korfmann, G. & Wiedemann, B. (1988). Impact of theampD gene and its product on β-lactamase production in Entero-bacter cloacae. Reviews of Infectious Diseases 10, 800–5.
56. O’Callaghan, C. H., Morris, A., Kirby, S. M. & Shingler, A. H.(1972). Novel method for detection of β-lactamases by using achromogenic cephalosporin substrate. Antimicrobial Agents andChemotherapy 1, 283–8.
57. Lowry, O. H., Rosenbrough, N. J., Farr, A. L. & Randall, R. J.(1951). Protein measurement with the Folin phenol reagent. Journalof Biological Chemistry 193, 265–75.
58. Sherwood, K. J., Wiegand, I., Westphal, K., Wagner I. &Wiedemann, B. (2002). Molecular identification of Escherichia coliand Klebsiella with suspected extended spectrum and AmpC beta-lactamases. Clinical Microbiology and Infection 8, Suppl. 1, 69(Poster P435).
59. Nakajima, H., Itoh, K.-I., Arakawa, E., Inoue, M., Mori, T. &Watanabe, H. (1994). Degradation of a polymerase chain reaction(PCR) product by heat-stable deoxyribonuclease (DNase) pro-duced from Yersinia enterocolitica. Microbiology and Infection 38,153–6.
60. Richmond, M. H. & Sykes, R. B. (1973). The β-lactamases ofGram-negative bacteria and their possible physiological role.Advances in Microbial Physiology 9, 31–88.
61. Nomura, K. & Yoshida, T. (1990). Nucleotide sequence of theSerratia marcescens SR50 chromosomal ampC β-lactamase gene.FEMS Microbiology Letters 70, 295–300.
62. Farzaneh, S., Peduzzi, J., Demachy, M. C., Barthelemy, M. &Labia, R. (1995). Identification dans une souche clinique de Serratiafonticola d’une céphalosporinase sensible à l’acide clavulanique.Pathologie et Biologie (Paris) 43, 315–9.
63. Peduzzi, J., Farzaneh, S., Reynaud, A., Barthelemy, M. &Labia, R. (1997). Characterization and amino acid sequence ana-lysis of a new oxyimino cephalosporin-hydrolyzing class A beta-lactamase from Serratia fonticola CUV. Biochimica et BiophysicaActa 15, 58–70.
64. Bellais, S., Poirel, L., Fortineau, N., Decousser, J. W. &Nordmann, P. (2001). Biochemical–genetic characterization of thechromosomally encoded extended-spectrum class A β-lactamasefrom Rahnella aquatilis. Antimicrobial Agents and Chemotherapy45, 2965–8.
65. Tzelepi, E., Arvanitidou, M., Tzouvelekis, L. S., Mavroidi, A. &Tsakris, A. (2000). The isoelectric focusing patterns of β-lactam-ases of Yersinia intermedia isolates from aquatic environments.Journal of Antimicrobial Chemotherapy 46, 513–9.
66. Grimont, P. A. D. & Grimont, F. (1984). Genus VIII. SerratiaBizio 1823, 288A. In Bergey’s Manual of Systematic Bacteriology(Krieg, N. R. & Holz, J. G., Eds), pp. 477–8. Williams & Wilkins,Baltimore, MD, USA.
67. Chopra, I., Hawkey, P. M. & Hinton, M. (1992). Tetracyclines,molecular and clinical aspects. Journal of Antimicrobial Chemo-therapy 29, 245–77.
68. Schnappinger, D. & Hillen, W. (1996). Tetracyclines: antibioticaction, uptake and resistance mechanisms. Archives of Micro-biology 26, 1414–5.
Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022

Antimicrobial susceptibility of unusual Serratia spp.
885
69. Shaw, K. J., Rather, P. N., Hare, R. S. & Miller, G. H. (1993).Molecular genetics of aminoglycoside resistance genes and familialrelationships of the amninoglycoside-modifying enzymes. Micro-biological Reviews 57, 138–63.
70. Champion, H. M., Bennett, P. M., Lewis, D. A. & Reeves, D. S.(1988). Cloning and characterization of an AAC(6′) gene fromSerratia marcescens. Journal of Antimicrobial Chemotherapy 22,587–96.
71. Shaw, K. J., Rather, P. N., Sabetelli, F. J., Mann, P., Munayyer,H., Mierzwa, R. et al. (1992). Characterization of the chromosomalaac(6′)-IC gene from Serratia marcescens. Antimicrobial Agentsand Chemotherapy 36, 1447–55.
72. Hawkey, P. M. & Constable, H. K. (1988). Selection of netilmicinresistance, associated with increased 6′-aminoglycoside activity, inSerratia marcescens. Journal of Antimicrobial Chemotherapy 21,535–44.
Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022

Dow
nloaded from https://academ
ic.oup.com/jac/article/51/4/865/745277 by guest on 07 January 2022





























