Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, Aug. 1983, p. 505-5110021-9193/83/080505-07$02.00/0Copyright C 1983, American Society for Microbiology
Vol. 155, No. 2
Naphthalene Dioxygenase: Purification and Properties of aTerminal Oxygenase Component
BURT D. ENSLEYt AND DAVID T. GIBSON*Center for Applied Microbiology, and Department of Microbiology, The University of Texas at Austin,
Austin, Texas 78712
Received 11 March 1983/Accepted 16 May 1983
Naphthalene dioxygenase from Pseudomonas sp. strain NCIB 9816 is amulticomponent enzyme system that oxidized naphthalene to cis-(lR, 2S)-diohydroxy-1,2-dihydronaphthalene. The terminal oxygenase component B waspurified to homogeneity by a three-step procedure that utilized ion-exchange andhydrophobic interaction chromatography. The purified enzyme oxidized naphtha-lene only in the presence ofNADH, oxygen, and partially purified preparations ofcomponents A and C. An estimated Mr of 158,000 was obtained by gel filtration.Polyacrylamide gel electrophoresis in the presence of sodium dodecyl sulfaterevealed the presence of two subunits with molecular weights of ca. 55,000 and20,000, indicative of an a2cB2 quaternary structure. Absorption spectra of theoxidized enzyme showed maxima at 566 (shoulder), 462, and 344 nm, which werereplaced by absorption maxima at 520 and 380 nm when the enzyme was reducedanaerobically by stoichiometric quantities of NADH in the presence of the othertwo components of the naphthalene dioxygenase system. Component B boundnapthalene. Enzyme-bound napthalene was oxidized to product upon the additionof components A and C, NADH, and 02. These results, together with thedetection of the presence of 6.0 g-atoms of iron and 4.0 g-atoms of acid-labilesulfur per mol of the purified enzyme, suggest that component B of thenaphthalene dioxygenase system is an iron-sulfur protein which functions in theterminal step of napthalene oxidation.
The initial reaction in the degradation of naph-thalene by soil pseudomonads involves the en-zymatic incorporation of one molecule of oxy-gen into the aromatic nucleus to form cis-(lR,2S)-dihydroxy-1,2-dihydronaphthalene (6, 15,16). Preliminary studies have shown that inPseudomonas sp. strain NCIB 9816 this reactionis catalyzed by a multienzyme system termednaphthalene dioxygenase. This system consistsofthree protein components, A, B, and C, whichare essential for enzymatic activity (12). In thisrespect, naphthalene dioxygenase is similar tothe multicomponent enzyme systems that formcis-dihydrodiols from benzene (2, 10), toluene(9, 13, 20, 21, 26), and 5-amino-4-chloro-2-phe-nyl-2H-pyridazin-3-one (19). Each of these en-zyme systems consists of a flavoprotein reduc-tase and a plant-type ferredoxin which arerequired for the transfer of electrons fromNADH to the terminal dioxygenase or to otherelectron acceptors such as cytochrome c. Thedioxygenases are iron-sulfur proteins that re-quire exogenous iron for optimum activity.However, naphthalene dioxygenase is not stim-
t Present address: Applied Molecular Genetics, Inc., Thou-sand Oaks, CA 91320.
ulated by exogenous iron. In addition, one of itscomponents (A) is an iron-containing flavopro-tein that will transfer electrons directly fromNADH to cytochrome c (unpublished data).These properties are similar to those reportedfor the two component benzoate dioxygenase(23-25) and 4-methoxybenzoate 0-demethylase(4-5) enzyme systems. The only multicompon-ent system that requires an iron flavoprotein andtwo additional proteins for activity is the meth-ane monooxygenase from Methylococcus cap-sulatus Bath (8, 9).We now report the purification and some
properties of component B in the naphthalenedioxygenase enzyme system. The oxidized en-zyme binds naphthalene and can accept twoelectrons from NADH in the presence of compo-nents A and C. The results presented suggestthat component B functions as the terminaldioxygenase component in naphthalene dioxy-genase. Preliminary characterization studiesshow that component B is an iron-sulfur proteinthat we have designated ISPNAP.
MATERLILS AND METHODSPseudomonas sp. strain NCIB 9816 was provided
by W. C. Evans, The University College of North
505 Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
16
Febr
uary
202
2 by
218
.57.
73.2
53.

506 ENSLEY AND GIBSON
Wales, Bangor, Wales. The maintenance and growthof the organism, as well as the preparation of crudecell extracts, were as described previously (12). Toensure enzyme stability, crude extracts were preparedin 50 mM Tris-hydrochloride buffer (pH 7.8) contain-ing 10%o (vol/vol) ethanol, 10%o (vol/vol) glycerol, and0.5 mM dithiothreitol (TEG buffer).Enzyme assays. Naphthalene dioxygenase activity
was determined by measuring the accumulation ofnonvolatile metabolites from ["4C]naphthalene (12).During purification of the terminal component of thenaphthalene dioxygenase system (component B), thisprocedure was used except that partially purifiedcomponent A and partially purified component C wereadded to the reaction mixture. Reactions (0.4 ml each)contained 50 mM Tris-hydrochloride buffer (pH 7.5),100 ,ug of component C, 60 Rg of component A, 1.0nmol of flavin adenine dinucleotide, 1.0 ,mol ofNADH and appropriate amounts of component B. Thereaction was started by the addition of 100 nmol of[1(4,5,8)-14C]naphthalene (1.1 x 106 dpm) in 10 RI ofdimethylformamide. One unit of enzyme activity isdefined as that amount of protein required to convert1.0 nmol of naphthalene to nonvolatile metabolites in 1min. NADH-cytochrome c reductase activity wasmeasured as described previously (12). Protein con-tent in cell extracts and enzyme fractions was deter-mined by a microbiuret procedure (14). Bovine serumalbumin (BSA) was used as a protein standard.
Enzyme-substrate binding measurements. The bind-ing of naphthalene to purified component B (ISPNAP)was measured by diluting 0.1 ml of TEG buffer con-taining 2.9 nmol of the enzyme with 0.9 ml of 50 mMTris-hydrochloride buffer, pH 7.5. The enzyme solu-tion was incubated with 50 nmol of [14C]naphthalene(55 x 105 dpm) in 5.0 RI of dimethylformamide for 20min at room temperature. Unbound naphthalene wasseparated from the protein by passing the solutionover a column of Sephadex G-25 (1 by 9 cm) which hadbeen previously equilibrated in 50mM Tris-hydrochlo-ride buffer, pH 7.5. Fractions of 1.1 ml were collected,and ISPNAP was detected by measuring the absor-bance of each fraction at 460 nm. The radioactivity ineach fraction was determined by transferring 0.1 mlinto 5.0 ml of Aquasol scintillation cocktail. Measure-ments were made in a liquid scintillation counter.Control experiments utilized the same procedure, withISPNAP replaced by BSA. The presence of BSA incolumn fractions was detected by measuring the ab-sorbance at 280 nm.The oxidation of enzyme-bound naphthalene was
measured by incubating 1.0 ml of a column fractionprepared in the above procedure with the other twocomponents of the naphthalene dioxygenase systemand NADH. A 1.0-ml column fraction, which con-tained 2.3 nmol of [14C]naphthalene bound to 2.5 nmolof purified ISPNAP, was incubated with 48 ,g ofpartially purified component A, 60 ,ug of partiallypurified component C, and 0.5 ,mol of NADH for 10min at room temperature. The reaction mixture wasthen rechromatographed, and fractions containing ra-dioactivity were pooled and extracted twice with equalvolumes of ethyl acetate. The organic phases werecombined, dried over anhydrous sodium sulfate, andevaporated to dryness under a stream of air at roomtemperature. The residue was dissolved in 100 p.1 ofethyl acetate, and 50 ,ul was applied to the origin of a
precoated silica gel chromatography plate. Authenticsamples of cis-naphthalene dihydrodiol, 1-naphthol,and naphthalene were applied to the same spot on thechromatogram, which was developed twice in a sol-vent system of chloroform-acetone (8:2, vol/vol). Thelocation of the standards was detected by viewingunder UV light. Areas of the chromatogram corre-sponding to each standard were transferred into 7.0 mlof scintillation fluid, and the amount of radioactivity ineach of the areas was determined by standard scintilla-tion counting procedures.
Spectrophotometric measurements. All spectropho-tometric measurements were made with an AmincoDW-2a double-beam spectrophotometer. Sampleswere contained in quartz cuvettes with a 1.0-cm lightpath. Analyses of anaerobic samples were made inanaerobic cuvettes (Hellma Cells, Inc., Jamaica,N.Y.) fitted with rubber septa. The samples wererendered anaerobic by alternate evacuation of thecuvettes and sparging with argon which had beenscrubbed over a column of hot copper pellets.
Analytic methods. Polyacrylamide gel electrophore-sis was performed on 8 and 10% acrylamide gels inTris-glycine buffer, pH 8.3, as described by Davis (11).Sodium dodecyl sulfate gels used for molecular weightdeterminations were prepared according to Weber andOsborn (22). Cytochrome c, ovalbumin, catalase, andBSA were used as protein standards. The nativemolecular weight of the protein was estimated bychromatography over a calibrated Sephadex G-200column as described by Andrews (1). Inorganic ironmeasurements were performed with a Perkin-Elmermodel 400 atomic absorption spectrophotometer.Acid-labile sulfide was measured by the method ofKing and Morris (17).
Materials. The following items were from thesources indicated: cytochrome c, NADH, flavin ade-nine dinucleotide, Trizma base, DNase, BSA, andDEAE-Sephadex G-25, Sigma Chemical Co., St. Lou-is, Mo.; Octyl-Sepharose 4B, Sephadex G-75, andSephadex G-25, Pharmacia Inc., Piscataway, N.J.;DEAE-cellulose (DE-52), Whatman, Inc., Clifton,N.J.; XM-50 ultrafiltration membranes, AmiconCorp., Lexington, Mass.; [1(4,5,8)-14C]naphthalene(specific activity, S mCi/mmol), Amersham Corp.,Arlington Heights, Ill.; precoated thin-layer chroma-tography sheets, Silica Gel 60 F254, MCB Manufac-turing Chemists, Inc., Cincinnati. cis-Naphthalene di-hydrodiol was prepared as described previously (18).All other chemicals were of the highest purity com-mercially available.
RESULTSPurification of the terminal oxygenase compo-
nent. Crude cell extract prepared from 25 g (wetweight) of cells (71.0 ml, 2.20 g of protein) wasapplied to a column of DEAE-Sephadex G-25(2.5 by 11.0 cm) which had been previouslyequilibrated with TEG buffer. Components Aand B of the naphthalene dioxygenase systemdid not bind to the column and were collected asa single fraction. The column was then washedwith 200 ml ofTEG buffer, followed by 200 ml ofTEG buffer containing 0.125 M KCl. ComponentC was eluted from the column with 0.2 M KCI in
J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
16
Febr
uary
202
2 by
218
.57.
73.2
53.

VOL. 155, 1983
TEG buffer. Fractions of 10 ml were collectedand assayed for naphthalene dioxygenase activi-ty in the presence of components A and B.Those fractions containing component C werepooled and stored at -20°C.Components A and B were separated by ion-
exchange chromatography. Protein which didnot bind to the DEAE-Sephadex G-25 column(75.0 ml, 2.0 g of protein) was applied to acolumn (4.0 by 8.0 cm) of DEAE-cellulose (DE-52). The column was washed with 200 ml ofTEGbuffer. Components A and B were then elutedwith a linear gradient (800 ml) of KCI (0.0 to 0.12M) in TEG buffer. Fractions (11.0 ml each) werecollected and assayed for naphthalene dioxygen-ase and NADH-cytochrome c reductase activi-ty. Those fractions that contained both activitieswere pooled and stored at -20°C as a source ofcomponent A. Fractions 70 to 84 were pooled togive a red-brown protein solution (153 ml, 230mg of protein) that showed naphthalene dioxy-genase activity only when assayed in the pres-ence of components A and C. The protein solu-tion obtained from fractions 70 to 84 wasconcentrated to 26.0 ml over an Amicon XM-50membrane and then brought to 40% saturationwith ammonium sulfate. A slight precipitateformed which was removed by centrifugation at10,000 x g for 15 min. The supernatant solutionwas applied to a column (1.5 by 20.0 cm) ofoctyl-Sepharose 4B which had previously beenequilibrated with TEG buffer containing 40%ammonium sulfate. During this procedure, adark-red band of protein was observed to con-centrate at the top of the column. The columnwas washed with 50 ml ofTEG buffer containing40%o ammonium sulfate, and this was followedby 200 ml of the same buffer containing 30%oammonium sulfate. Fractions (10.0 ml each)were collected and assayed for naphthalenedioxygenase activity which was detected in frac-tions 16 to 25. The contents of these tubes werepooled, dialyzed overnight against 2.0 liters ofTEG buffer, and concentrated to 9.5 ml over anAmicon XM-50 membrane. A summary of thepurification procedure is shown in Table 1. A 36-fold purification of the enzyme was achieved,with a recovery of 30%o of the activity present inthe crude cell extract.
Polyacrylamide gel electrophoresis of the pu-rified enzyme on 8 and 10%o gels gave a singleband which stained for protein.
Properties of component B. The molecularweight of purified component B was determinedby gel filtration on a standardized column ofSephadex G-200. From the results obtained, themolecular weight of the native protein was cal-culated to be 158,000. Polyacrylamide gel elec-trophoresis in the presence of 0.1% sodiumdodecyl sulfate and 0.1% 2-mercaptoethanol re-
NAPTHALENE DIOXYGENASE 507
TABLE 1. Purification of naphthalene dioxygenase
Purification stepa Protein Activity Sp act Recovery(mg) (U) (U/mng) ()
1. Crude extract 2,215 84,000 37.92. DEAE-Sephadex 1,987 81,000 10.8 96
eluate3. DEAE-cellulose 230 73,400 319.0 87
chromatography4. Octyl-Sepharose 19 26,030 1,370.0 31
a Activity determined in the presence of partiallypurified components A and C and flavin adeninedinucleotide as described in the text.
vealed the presence of two subunits with molec-ular weights of 55,000 and 20,000. These obser-vations suggest that the native protein has ana2P2 composition.
Preliminary analyses revealed the presence of4.0 g-atoms of acid-labile sulfur and 5.6 g-atomsof iron per 158,000 g of protein. These resultsindicate that component B of the naphthalenedioxygenase system is an iron-sulfur protein thatwe have designated ISPNAP-
Spectral studies. The absorption spectrum ofthe purified enzyme showed maxima at 566(shoulder), 462, and 334 nm (Fig. 1). The calcu-lated extinction coefficients at these wave-lengths were 3.8, 9.0, and 21.0 mM-1 cm-1,respectively. The protein could be enzymatical-ly reduced by NADH in the presence of catalyticamounts of components A and C under anaero-bic conditions (Fig. 2), with a loss of absorptionat 566 and 462 nm and the appearance of newmaxima at 520 and 380 nm (Fig. 2, line 10).When absorbance at 462 nm was plotted as afunction of the NADH added (Fig. 3), a linear
LO
WAVELENGTH, nm
FIG. 1. Absorption spectrum of oxidized ISPNAP.The sample cuvette contained 40 nmol of purifiedenzyme in 1.0 ml of TEG buffer. The inset shows theresult of electrophoresis of purified component B in an8% polyacrylamide gel at pH 8.3.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
16
Febr
uary
202
2 by
218
.57.
73.2
53.
508 ENSLEY AND GIBSON
wz 0.12
0co) 0.08-
0.04-
340 380 420 460 500 540 580 620WAVELENGTH, nm
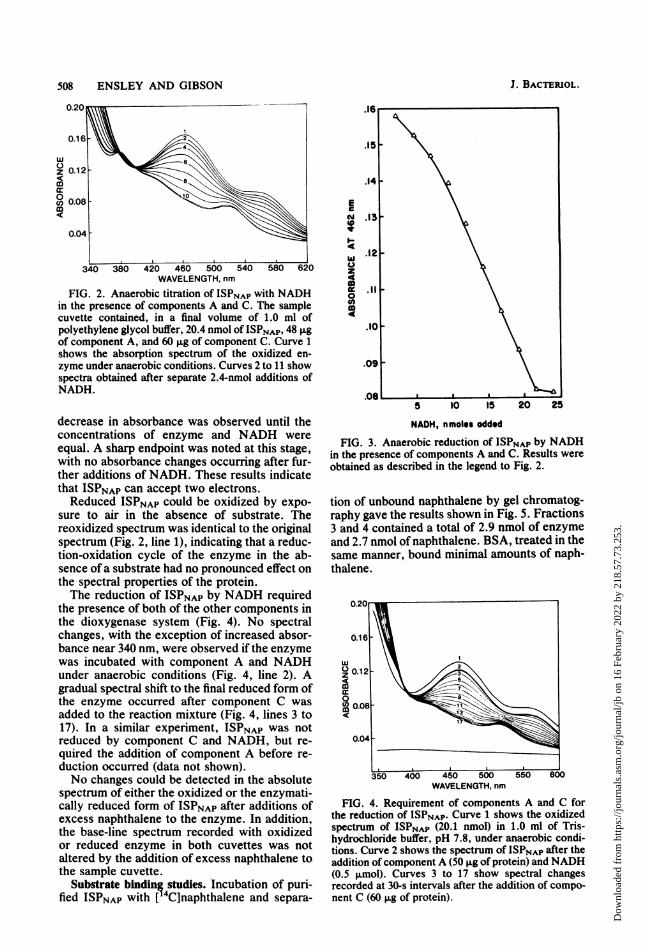
FIG. 2. Anaerobic titration of ISPNAP with NADHin the presence of components A and C. The samplecuvette contained, in a final volume of 1.0 ml ofpolyethylene glycol buffer, 20.4 nmol of ISPNAP, 48 Agof component A, and 60 ,ug of component C. Curve 1shows the absorption spectrum of the oxidized en-zyme under anaerobic conditions. Curves 2 to 11 showspectra obtained after separate 2.4-nmol additions ofNADH.
decrease in absorbance was observed until theconcentrations of enzyme and NADH wereequal. A sharp endpoint was noted at this stage,with no absorbance changes occurring after fur-ther additions of NADH. These results indicatethat ISPNAP can accept two electrons.Reduced ISPNAP could be oxidized by expo-
sure to air in the absence of substrate. Thereoxidized spectrum was identical to the originalspectrum (Fig. 2, line 1), indicating that a reduc-tion-oxidation cycle of the enzyme in the ab-sence of a substrate had no pronounced effect onthe spectral properties of the protein.The reduction of ISPNAP by NADH required
the presence of both of the other components inthe dioxygenase system (Fig. 4). No spectralchanges, with the exception of increased absor-bance near 340 nm, were observed if the enzymewas incubated with component A and NADHunder anaerobic conditions (Fig. 4, line 2). Agradual spectral shift to the final reduced form ofthe enzyme occurred after component C wasadded to the reaction mixture (Fig. 4, lines 3 to17). In a similar experiment, ISPNAP was notreduced by component C and NADH, but re-quired the addition of component A before re-duction occurred (data not shown).No changes could be detected in the absolute
spectrum of either the oxidized or the enzymati-cally reduced form of ISPNAP after additions ofexcess naphthalene to the enzyme. In addition,the base-line spectrum recorded with oxidizedor reduced enzyme in both cuvettes was notaltered by the addition of excess naphthalene tothe sample cuvette.
Substrate binding studies. Incubation of puri-fied ISPNAP with [ 4C]naphthalene and separa-
EC4
N ~ 10 IS 20 2
40
4
0
10
.09
.085 10 15 20 2
NADH, nmoles added
FIG. 3. Anaerobic reduction of ISPNAP by NADHin the presence of components A and C. Results wereobtained as described in the legend to Fig. 2.
tion of unbound naphthalene by gel chromatog-raphy gave the results shown in Fig. 5. Fractions3 and 4 contained a total of 2.9 nmol of enzymeand 2.7 nmol of naphthalene. BSA, treated in thesame manner, bound minimal amounts of naph-thalene.
WAVELENGTH, nm
FIG. 4. Requirement of components A and C forthe reduction of ISPNAP. Curve 1 shows the oxidizedspectrum of ISPNAP (20.1 nmol) in 1.0 ml of Tris-hydrochloride buffer, pH 7.8, under anaerobic condi-tions. Curve 2 shows the spectrum of ISPNAP after theaddition of component A (50 ,ug of protein) and NADH(0.5 ,umol). Curves 3 to 17 show spectral changesrecorded at 30-s intervals after the addition of compo-nent C (60 ,ug of protein).
J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
16
Febr
uary
202
2 by
218
.57.
73.2
53.

NAPTHALENE DIOXYGENASE 509
B
4
2t
49-
I
z
.4
z
FRACTION NUMBR
FIG. 5. Binding of [14C]naphthalene to oxidizedISPNAP. Experimental conditions are described in thetext. Symbols: AL, ISPNAJP, absorbance at 460 am; 0,radioactive naphthalene.
After incubation of the enzyme-bound naph-thalene with component A, component C, andNADH, followed by rechromatography, the re-sults shown in Fig. 6 were obtained. All of thenaphthalene was converted to a product whichno longer coeluted with the protein, but elutedsooner than did unreacted naphthalene. Analy-sis of this compound by thin-layer chromatogra-phy revealed that it comigrated with cis-naph-thalene dihydrodiol.
DISCUSSIONNaphthalene dioxygenase has been shown to
be a multicomponent enzyme system which cat-alyzes the NADH-dependent oxidation of naph-thalene to cis-1,2-ihydroxy-1,2-dihydronaphth-alene (12). Rapid purification of the separatecomponents in this system is essential due to thereported instability of the enzyme (7). The puri-fication scheme presented in this report resultedin the isolation of the terminal component of thenaphthalene system in purified form within 4days. Stability of the enzyme during purificationrequired the presence of 10% ethanol and 10%oglycerol (12). The same solvents have been usedduring the purification of the terminal compo-nent of toluene dioxygenase (20).
Purified ISPNAP has several properties in com-mon with the terminal components of the ben-zene (2, 10), pyrazon (19), and toluene (13, 20)dioxygenase systems. All of these proteins arecolored with absorption maxima in the visibleregion near 550 and 450 nm. The enzymaticreduction of the terminal component in ISPNAPresulted in the loss of absorption at 566 and 462nm, with the appearance of new maxima at 520and 380 nm. Similar results were observed withthe pyrazon and- benzene dioxygenases and theterminal monooxygenase component of the 4-methoxybenzoate 0-demethylase system (3-5).
/ /P .x\ . I1 5 10 15
FRACTION NUMBER
FIG. 6. Oxidation of enzyme-bound naphthalene inthe presence of NADH and components A and C.Symbols: 0, ISPNP, absorbance at 460 nm; A, radio-active metabolite. Experimental conditions are de-scribed in the text.
The enzymatic reduction of ISPNAP requiredthe presence of components A and C andNADH. Complete reduction of the protein re-sulted after the addition of an equimolar amountof NADH, as has been observed with benzeneand toluene dioxygenases (10, 13), indicatingthat two electrons are required for completereduction. The absorption maxima at 380 and520 nm, characteristic of the reduced protein,did not begin to appear until after half of the totalNADH had been added (Fig. 2, lines 6 to 10).These results suggest that a nonequivalent re-duction of iron-sulfur centers occurs in thisprotein.The native and subunit molecular weights of
ISPNAP are similar to those reported for toluenedioxygenase (13, 20), whereas the pyrazon andbenzene dioxygenases have molecular weightsof 180,000 and 215,000, respectively.
Oxidized ISPNAP will bind stoichiometricamounts of naphthalene, whereas the very simi-lar toluene dioxygenase must be reduced beforesubstrate binding can occur (20). The monooxy-genase in the 4-methoxybenzoate 0-demethyl-ase system will also bind a substrate when theenzyme is oxidized (4), but in marked contrast tothis enzyme, ISPNAP does not undergo anyspectral changes after incubation with naphtha-lene. Apparently, substrate binding by ISPNAPdoes not result in conformational changes whichaffect the iron-sulfur chromophores in this pro-tein. Complete oxidation of enzyme-boundnaphthalene to cis-naphthalene dihydrodiol afterincubation with components A and C andNADH demonstrates that the bound naphtha-lene is associated with the catalytic site onISPNAP and that all of the bound substrate canbe oxidized to product. Substrate binding by
VOL. 155, 1983
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
16
Febr
uary
202
2 by
218
.57.
73.2
53.

510 ENSLEY AND GIBSON
ISPNAP is reversible, since enzyme-bound,[14C]naphthalene can be replaced by unlabelednaphthalene in exchange experiments (unpub-lished data).ISPNAP was found to contain 5.6 g-atoms of
iron and 4.0 g-atoms of acid-labile sulfur perenzyme molecule. This is the highest iron con-tent reported for any of the dioxygenase iron-sulfur proteins. The terminal components of thebenzene (10) and toluene (13, 20) dioxygenasesystems each contain four atoms of iron andacid-labile sulfur per enzyme molecule, whereaspyrazon dioxygenase contains 1.96 to 2.33 molof iron and 1.6 mol of inorganic sulfur per mol ofenzyme (19). The iron contents reported forbenzene, toluene, and pyrazon dioxygenase maynot reflect the iron content of the catalyticallyactive protein, since all of these dioxygenasesrequire the addition of exogenous iron to reac-tion mixtures for maximum activity. In contrast,naphthalene dioxygenase activity is only mini-mally stimulated by exogenous iron (12).Putidamonoxin, the monooxygenase in the 4-
methoxybenzoate 0-demethylase system, hasbeen reported to contain equimolar amounts ofiron and acid-labile sulfur (3, 5). Bernhardt andMeisch have recently proposed that this proteincontains, in each active site, a 2Fe-2S center andan additional iron bound by a mercaptide group(4). The extra iron is essential for catalyticactivity in the proposed model. An analogouscatalytic system in ISPNAP, with two iron-sulfur(2Fe-2S) chromophores in each enzyme mole-cule, would result in a protein which contains sixatoms of iron and four atoms of acid-labile sulfurper molecule. Additional studies of the iron-sulfur centers in ISPNAP will be necessary beforethe interaction of iron and sulfur in the catalyticcenter(s) of this molecule can be resolved.The data presented in this study confirm the
presumed arrangement of the components in thenaphthalene dioxygenase system. Component Awill oxidize NADH in the presence of an elec-tron acceptor such as cytochrome c, whereascomponents B and C do not oxidize NADH (12).The requirement of both components A and Cfor the reduction of ISPNAP and the binding ofnaphthalene by ISPNAP indicate that the flow ofelectrons in the naphthalene dioxygenase sys-tem follows the pattern: NADH -* A -- CCISPNAP. Reduced ISPNAP reacts with boundnaphthalene and oxygen to form cis-naphthalenedihydrodiol.The apparent stability of catalytic iron in the
terminal component of naphthalene dioxygenasemakes this protein particularly useful for studiesof the interactions between iron, oxygen, andsubstrate which are involved in dioxygen fixa-tion. The availability of purified ISPNAP willgreatly facilitate efforts to elucidate the mecha-
nism of dioxygenation reactions in microbialenzyme systems.
ACKNOWLEDGMENTSThis investigation was supported in part by Public Health
Service grant GM 29909 from the National Institute of GeneralMedical Sciences, grant F-440 from the Robert A. WelchFoundati6n, the Exxon Education Foundation, and grantBRSG S07RR07091 from the Biomedical Research SupportGrant Program, Division of Research Resources, NationalInstitutes of Health, to The University of Texas at Austin.B.D.E. was a postdoctoral trainee supported by Public HealthService grant T32 CA09182 from the National Cancer Insti-tute.We thank V. Subramanian and B. E. Haigler for helpful
advice and assistance and Roberta DeAngelis and LindaFenton for preparing the manuscript.
LITERATURE CITED1. Andrews, P. 1964. Determination of molecular weight of
proteins. Biochem. J. 91:222-233.2. Axcell, B. C., and P. J. Geary. 1975. Purification and
some properties of a soluble benzene oxidizing systemfrom a strain of Pseudomonas. Biochem. J. 146:173-183.
3. Bernhardt, F. H., E. Heymann, and P. S. Traylor. 1978.Chemical and spectral properties of putidamonoxin, theiron-containing and acid-labile sulfur-containing mono-oxygenase of a 4-methoxybenzoate 0-demethylase fromPseudomonas putida. Eur. J. Biochem. 92:209-223.
4. Bernhardt, F. H., and H. U. Melach. 1980. Reactivationstudies on putidamonoxin: the monooxygenase of a 4-methoxybenzoate 0-demethylase from Pseudomonas pu-tida. Biochem. Biophys. Res. Commun. 93:1247-1253.
5. Bernhardt, F. H., H. Pachowsak, and H. Staudager. 1975.A 4-methoxybenzoate 0-demethylase from Pseudomonasputida. Eur. J. Biochem. 57:241-256.
6. Catterali, F. A., K. Murray, and P. A. Williams. 1971.The configuration of the 1,2-dihydroxy-1,2-dihydronaph-thalene formed in the bacterial metabolism of naphtha-lene. Biochim. Biophys. Acta 237:361-364.
7. Catterall, F. A., and P. A. Wilama. 1971. Some proper-ties of naphthalene oxygenase from Pseudomonas sp.NCIB 9816. J. Gen. Microbiol. 67:117-125.
8. Colby, J., and H. Dalton. 1978. Resolution of the methanemonooxygenase of Methylococcus capsulatus (Bath) intothree components. Biochem. J. 171:461-468.
9. Colby, J., and H. Dalton. 1979. Characterization of thesecond prosthetic group of the flavoenzyme NADH-acceptor reductase (component c) of the methane mon-ooxygenase from Methylococcus capsulatus. Biochem. J.177:903-908.
10. Crutcher, S. E., and P. J. Geary. 1979. Properties of theiron-sulfur proteins of the benzene dioxygenase systemfrom Pseudomonas putida. Biochem. J. 177:393-400.
11. Davia, B. J. 1964. Disc electrophoresis method and appli-cation to human serum proteins. Ann. N.Y. Acad. Sci.121:404 427.
12. Enaley, B. D., D. T. Gibson, and A. L. Laborde. 1982.Oxidation of naphthalene by a multicomponent enzymesystem from Pseudomonas sp. strain NCIB 9816. J.Bacteriol. 149:948-954.
13. Gibson, D. T., W.-K. Yeh, T.-N. Liu, and V. Subraman-ian. 1982. Toluene dioxygenase: a multicomponent en-zyme system from Pseudomonas putida, p. 51-62. In M.Nozaki, S. Yamamoto, Y. Ishimura, M. J. Coon, L.Ernster, and R. W. Estabrook (ed.), Oxygenases andoxygen metabolism. Academic Press, Inc., New York.
14. Itzhaki, R. F., and D. M. GDI. 1964. A microbiuret meth-od for estimating protein. Anal. Biochem. 9:401-410.
15. Jeffrey, A. M., H. J. C. Yeh, D. M. Jerlna, T. R. Patel,J. F. Davey, and D. T. Gibson. 1975. Initial reactions inthe oxidation of naphthalene by Pseudomonas putida.Biochemistry 14:575-584.
J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
16
Febr
uary
202
2 by
218
.57.
73.2
53.

VOL. 155, 1983
16. Jerina, D. M., J. W. Daly, A. M. Jeffrey, and D. T. Gib-son. 1971. cis-1,2-Dihydroxy-1,2-dihydronaphthalene: abacterial metabolite from naphthalene. Arch. Biochem.Biophys. 142:394-3%.
17. King, T. E., and R. 0. MorrI. 1967. Determination ofacid-labile sulfide and sulfhydryl groups. Methods Enzy-mol. 10:634-646.
18. Patel, T. R., and D. T. Gibson. 1974. Purification andproperties of (+)-cis-naphthalene dihydrodiol dehydro-genase ofPseudomonas putida. J. Bacteriol. 119:879-888.
19. Sauber, K., C. Frabner, G. Roseberg, J. Eb ,and F. Lingens. 1977. Purification and properties of pyra-zon dioxygenase from pyrazon-degrading bacteria. J. Bio-chem. 74:89-97.
20. Subraoanan, V., T.-N. Liu, W.-K. Yeb, and D. T. Gib-son. 1979. Toluene dioxygenase: purification of an iron-sulfur protein by affinity chromatography. Biochem.Biophys. Res. Commun. 91:1131-1139.
21. Subrama, V., T.-N. Li, W.-K. Yeb, M. Narro, andD. T. GPison. 1981. Purification and properties ofNADH-fefredoxinTOL reductase: a component of toluene dioxy-
NAPTHALENE DIOXYGENASE 511
genase from Pseudomonas putida. J. Biol. Chem.256:2723-2730.
22. Weber, K., and M. Osborn. 1969. The reliability ofmolecular weight determinations by dodecyl sulfate-poly-acrylamide gel electrophoresis. J. Biol. Chem. 244:4406-4412.
23. Yam dl, M., and H. Fucisiwa. 1978. Characterizationof NADH-cytochrome c reductase, a component of ben-zoate 1,2-dioxygenase system from Pseudomonas arvillaC-1. J. Biol. Chem. 253:8848-8853.
24. Yanu_achi, M., and H. Fujiawa. 1980. Purification andcharacterization of an oxygenase component in benzoate1,2-dioxygenase system from Pseudomonas arvilla C-1. J.Biol. Chem. 255:5058-5063.
25. Yamaguchi, M., and H. Fujisawa. 1981. Reconstitution ofiron-sulfur cluster of NADH-cytochrome c reductase, acomponent of benzoate 1,2-dioxygenase system fromPseudomonas arvilla C-1. J. Biol. Chem. 256:6783-6787.
26. Yeb, W.-K., D. T. Gibson, and T.-N. Ln. 1977. Toluenedioxygenase: a multicomponent enzyme system. Bio-chem. Biophys. Res. Commun. 78:401-410.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
16
Febr
uary
202
2 by
218
.57.
73.2
53.





























