Embed Size (px)
Citation preview

The UOEH Association of Health Sciences
NII-Electronic Library Service
The UOEHAssociation ofHealth Sciences
J UOEH M(4): 253-260 (1992) 253
Reduction and Inactivation of the Sarcoplasmic
by 2-Mercaptoethanol-Contrast
to the (Na', K') ATPase-
[Original)
Ca2'-ATPase
Yoshinobu MUTOHi'2, Atsuko SUGINOt, Ken HIGASHI3, Masayuki TAKASUGI2,
Akio KUROIWA? and Masaru KAWAMURAiiDivision
ofBiotqgy,"Second
Dopartment of internal Medic'ine3Dapartment
ofBiochemistty, St,hoot of Medicine, Chtiversity of Occiipational and Envirenmental Hbatth,
.kipan. Ylahatanishi-ku, Kitalpashu 807, .J4pan
Abstrace :
Key words:
Tbe sarcoplusmic CaP''-ATPase was reduccd with 300 rnM 2-mercaptoethanol at elcvated
terriperatures (40 - 45℃ ) w{th a concornitant loss of iXTPase activity. Thc reductioii and
inaetivation of the Ca!+-ATPase proceeded rapidiy in the absence of Ca2'. '1'he
Ca'+-ATPase
was also inactivated with 2-mcrcaptocthanol in the presence of diluted SDS (O.4・ mg/ml) even at
20℃ . In contrast to thc (Na', KF') A'i'Pase, the inactivated CaUT-A'1'Pase in the prcsence of
diluted Sl)S was sedimented by the centrifugation at 100,OOO × g tbr 30 min.
Ca2F-ATPase, disutfide bond, 2-mercaptoethanol, (Nai, K') ATPase.
(Received 17 August 1992, accepted 1 october 1992)
Introduction
There is a close structural similarity between the Ca'"'-ATPase of sarcoplasmic
reticulum and the a-subunit of the (Nai, K") ATPase of plasma membrane, including
transmembrane topology as well as highly conserved domains fbr ATP binding and
phosphory]ation [1], On the other hand, while the CaL'-ATPase is composed solely of
110 kDa poiypeptide, the (Na', K') ATPasc requircs the glycosylated f?-subunit in addition
to thc cr-subunit. The B-subunit is indispensable in the biogenesis of thc (Na', Ki)
ATPase, but its role in the catalytic cycle is still unknown l2-51.
Kawamura and Nagano [6] and recently Kirlcy [7I have shown that the reduction ot' thc
disulfide bonds in the B-subunit resulted in the inactivation of the (Na', K') ATPasc.
The disulfide bonds in the a-subunit were lcft intact, It ufould be, interesting to discover
whether reducing agcnts, such as 2-mercaptoethanol, inactjvate Ca"-A'l'Pase without the B-subunit Here, we report that the Ca"-ATPase was reduced by 2-mercaptoethanol with a
concomitant loss of ATPasc activity.

The UOEH Association of Health Sciences
NII-Electronic Library Service
The UOEHAssociation ofHealth Sciences
・254 Y MuToH et at
Materials and Methods
Proparation ofsarcoplasmic reticulum
Sarcoplasmic'reticulum (SR) was preparcd from back muscles of New Zealand white
rabbits as dcscribed by Kanazawa et al [8], SR was further trcated with either
octacthylenedodecylethcr (C]!Es) or deoxycholate (DOC), Estimation of the disulfide
bond was carried out by using I)OC-solubilized enzyme that was composcd of a single band
of 1 10 kDa polypeptide on sodium dodecylsulfate polyacr>JIamide gcl.
7Yeatment u.:ith re(incing crgents at elevatecX temperatec'res
Ci2E"-solubilized Ca2'-ATPase (O.2-O.3 mg/ml) was incubated in 10 mM TES butrer
(pH 7.5) containing 100 mM KCI, 1 mM CaC17, and O.1 mg/ml Tween 80 in the presenceand absence of reducing agents (300 mNI 2-mercaptoethanol or 4 mM tributhylphosphine)
at elevated tempcratures, At the end of incubation, 100 gl aliquots of incubation mixturc
was withdrawn and added to 900 pt1 Ca?'-ATPase assay medium containing 50 mM
Tris/maleate (pH 7.0), 100 mM KCI, 5 mM MgCl?, lmM EGTA and O,47 mM CaC12.
ATPase activity was determined as the rate of rclcase of inorganic phosphate at 37℃ [81.For the controls, the rate of re]ease of inorganic phosphattr in thc absence of O.47 mM CaClp
was determined and subtracted tfom the above,
Labeling qf disuUide bonds
Labeling of disuliide bonds was carried out as described by Kirley [9]. DOC-
solubilized CaL'-ATI'ase (3 mg/ml) was alkylated with 4 mM lodoacetamide in e.5 ml of
200 mM TrisfHCI (pH 8.3), 5 mM EDTA, 5 M urea and 4.0% sodium dodecylsul{'ate
(SDS) in the dark at 37℃ I'or 1 h, 'I]he
alkylated Cti2'-ATPase was applied on a co}umn
of Bio-Gel P-6 DG and concentra{ed using (1]entricon 30 microconccntrators. 'Lihe
alk}ilation with iodoacetamide was repeated again. To 3.5 ml of Lhe alkylated protein, O.5
ml of a freshly mixed solution containing IM NaBOi (pH 8.0), 8 mlVI EDTA, 2 rnrvl tribu-
tylphosphine (TBP) and 2 mM 4-<aminosulfbnyl)-7-fluoro-2, 1, 3-benzoxadiazole (ABD-F)was added and heated at 60℃ for 1 h.
'FBP-reduced
and ABD-F-alkylated Ca'i-ATI'asc,
whose frec sulthydryl groups had been alkylated with iodeac;etamide, was then applied en a
column of TSK-GS"J-3000 ancl the peak fractions of (:a"'-ATPase wcrc pooled and assayed
fbr protein and ABD-F (absorbance at 385 nm). In another set of experiments, I)OC-
solubilized Ca"-ATPase (3 mg/ml) was incubated at 43℃ in 10 m)vr TrislHCI (pH 7.5),IOO mM KCI, 1 mM CaCIL, with or without 4 mM TBP in the presence of 2 mM ABD-
F. Aftcr 60 min, the paired samples were cooled to room tempcraturc and applied on a
column of Bio-Gel P-6 DG. Thc samp]es were solubilized with 4% SDS, separated by'IiSK-GSW-300e
chromatograph>, and assayed fbr protein and ABD-F as above,

The UOEH Association of Health Sciences
NII-Electronic Library Service
The UOEHAssociation ofHealthSciences
InacLivation of Ca2+-ATPase 2.55
Other methocts'
Protein was determined by the method of Lowry et al [10I with bovinc serum albumin
as a standard. SDS-polyacrylamidc ge] electrophoresis was carried out by the method of
Laemmli [l ll using a separating gcl of 1O%.
Results
inactivation ofthe Cke'i -A7T'ase lij, redatcing a.aents at etevated temperalures
The thermal inactivation of the Ca2'-ATPasc in the presence oJ' 300 mpt1 2-mercaptoe-
thanol was determined by measuring the activity at 37℃ after exposure to inactivating
temperatures. As it was rel)orted by Lepock et al I12] that Ca2i protects the Ca2'-ATPase
fi/om thermal inactivation, the sarnples were trcated with and without l mM Call'. Figure
1 illustrates the results obtaincd in the presencc of l mrv{ Ca2'. Figure l (A) shows the
time course of inactivation of the Ca"-ATPase activity at 43℃ . The inactivation by 2-
mercaptoethanol obeys pseudo first-order kinetics and was almost completed after 15 min
exposure to 43℃ . The other reducing agent, TBP (4 mM), induced similar inactivation
of thc Ca'F-AFIil]ase, though to a lesser extent. Without 2-mercaptoethanel, the rate of
inactivation at 43℃ was slow as shown in Fig, 1 (A). An adclition of ethanol (350 mM)
did not havc any signifieant efibet on thermal {nactivation. These results suggested Lhat
the inactivation due to the reducing agents resulted from the reduction of disulfide bond(s)
of the Ca?'-ATPase. Figure 1 (B & C) shows the efl'ec'ts oftemperature and concentration
of 2-mercaptoethanol, rcsl]cctively, on t,he itiactivation of ATPase activity,
" t' IE・ 6o 6o
"U
di
of' e
.
0 o 10 lto 30 35 40 4s O O.1 O.2 O.3
Time(min) TeinperatureC(;) 2-mercaptoethanol(Dvl)
Fig. 1. Inactivation of' Ca7+-ATPase with reducing agents at elevatcd temperaLures. CmEH-
solubilized enzymc (O.2-O,3 mg/ml) was incubated in the medium containing IU rntL'1 TES
huffer (pH 7.5}, 100 mM KCI, ] mM CIaCIL, and O,1 mg/m] Twcen 80 in thc presenctr and
absence of a reducing agent. For (iN), incuhation pcriods were varied with a uonstant
{toi]centrution of eiihcr 300 rnN{ 2-mercaptoethanot (O) er 4 mptt 'l'BP
(A) and without
reducing agents (()) at 43℃ . Fur {B), incubation temperaturcs wcre ehangeti in the
preseiice (O) and absence {C)) of 300 m)vl 2-mcrcaptoethanol. Incubatiun periods werc
15 min. For (C), 2-mercaptoethanol conccntrations "'ere var{ed and incubations Ns,erc
performed at 43 ℃ ibr 15 min,
The UOEH Association of Health Sciences
NII-Electronic Library Service
The UOEH Assoolatlon of Health Solenoes
YMUIOHetal256
In the abscncc of Cd2 +
(1n the prcsencc of l mM EGTA ), mactlvaUon of the CaP+ −
ATPase in the prescncc ot 300 mM 2−mercaptoethanol was rapid and completed withm a 5
mln expo 〜ure to 43℃ (data not shown )
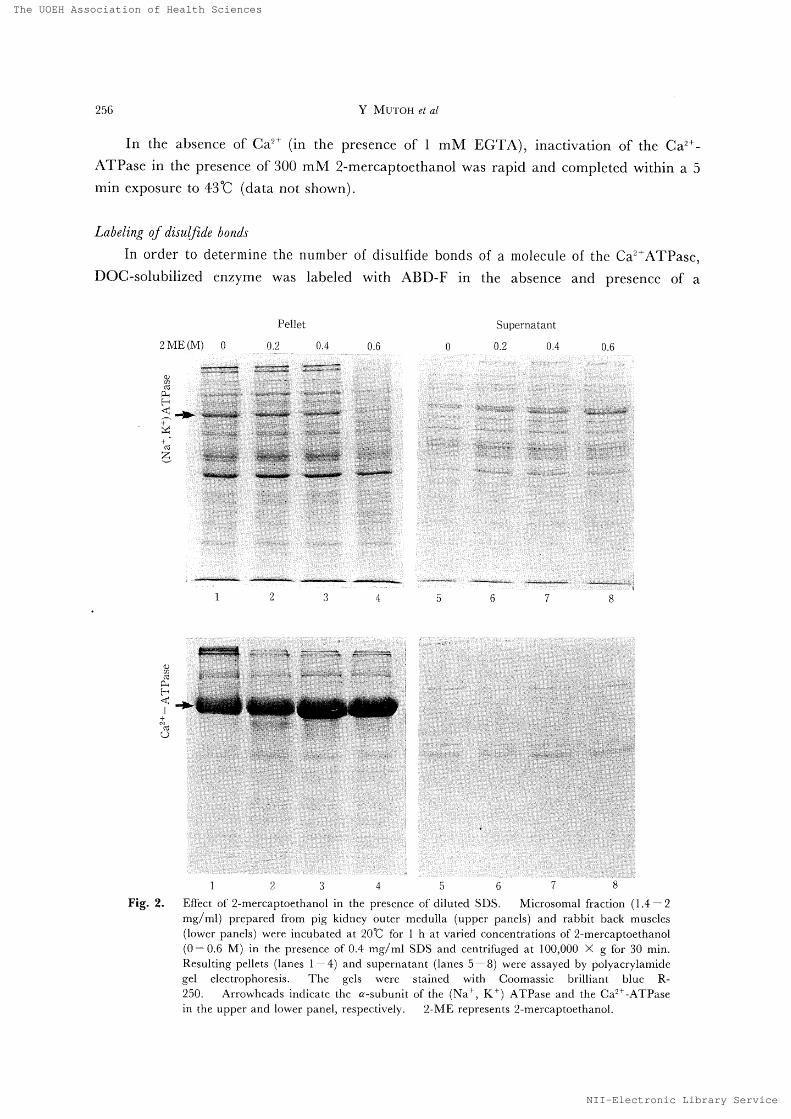
Labelzng cf dtsulfide bond・
In order to determlne thc number Qf disulfide bondg of a molecule of the Ca2+ATPase ,
DOC −solublllzed cn7yme was labeled wlth ABD −F ln thc absence and pregcncc of a
纛巽
。
携
∵驚
惣
贈謹
機
髏
繋
躡
↑
ヒ
}窶 レ
品
,鑼
婆糞
ゆ,
}
さ
図
驚耀
鑾蠶ノ
ニ3
.
鵡驪
覊壷
繋鸚
∵爨
÷
も
ゑ
ヨ
嚢鱗
灘
.
、
、
鞍
篷
ー
…
、
…
、贓
撫遷瀞
鍵、
劉
∴蜘
蠡
罵醤
獄撫
称
∴∴
∵熊
域∴撫∵》∵び
、・
嘆鴨
懸
触
霧
ヒ賢・
銃
竃
篝
∴鱗
}
ギ
邪韓
∵∵黴
、
Φω
山
←<
1一 」
評
鴎輩
♂ 階
…
卓轟
声
驫
纛韈
謙鱗
Fi92
1 2 3 4 5 b 7
H 麁dof 2 mercaptoethanol ln the presence of dlluted SDg Mlcroromal fractlon([4− 2
mg /m り p 正 epared from plg kldney {〕utcr mcdulla 〔upper [)aIlels )and rabbit back musclcs
(lo“ er panelr)“ cre mcub 飢 cd at 20℃ f()r l h at vancd (oncentrdttonb of 2 mercaptoethanol
(0 − 06M )m 且hq )resenLe of O 4 mg /ml SD 〜 and Lentnfi ユged at 100,000 × gfor 30 mm
Resulting peHe〔s (lanes 1− t)and supernatarlt (lanes 5
− 8)were absayed b》 po且v 肌 rylamide
gel el 〔 cLrophoIeslr Thc gtl〜 w 唖 re btd ]ned 、〜 Lth Coomass艮e bmlliant blue R250 Arrowheddb lndl 〔ate the α subumt ot the (丶 a
+ K +
)ATPare and the Ca 片
へTPasem 山 euppe1 and lower ptuiel, iespecu 、 ely 2ME repr じ bents 2 mertaptoethanol
N 工工一Eleotronlo Llbrary

The UOEH Association of Health Sciences
NII-Electronic Library Service
The UOEHAssociation ofHealth Sciences
Inactivation of Ca?'-ATPase 2:')7
reducing agent. Prior to labeling, the SH groups of the Ca2"-ATPase were alkylated with
iodoacetamide in the presence ofSDS, TBP was used as the reducing agent since it does
not react with ABD-F. Reduction with TBP and alkylation with .ABD-F should be
performed simultaneously so that reoxidation to disulfide during labeling can be avoided [9).The number of disulfide bonds, thus estimated, was 3,22 a molecule of the Ca?'-ATPase
(average of 3.13 ancl 3.31 in two separate determinations). X'Vhcn the native DOC-
solubilized enzyme was labeled with ABD-F in the presence ancl absencc of TBP at 48℃ for
60 min, the diflerencc in the number of ABD-F incorporated was O.97 (avcrage of O.94 and
1.0 in two separate determinations) between the two samples. (Since the inactivation by'IiBI'
was slow as shown in Fig. 1 (A), the enzyme was incubated for 60 min at 43℃ under
the condition where the inactivation was almost complete.) These results suggested that
at least one disulfide bond out of three in thc Ca?"-ATPase was clcaved upon treatment
with a reducing agent at elevated temperatures,
Effect of2-mercoptoethanol on the dn2'-ATPase in the presence of'diluted SDS
We have already observed that the (Na', K') ATPase lost its activity when treated
with 2-mercaptoethanol even at room temperature ifdiluted SDS was present [13, 14], and
that the inactivated enzyme was no longer sedimented by centrifugation at 100,OOO × g fbr
30 min [13], The C`solubilization"
of the (Na', K') AFIiPase was confirrned by using crude
microsomes from pig kidney outer meclulla (Fig. 2, upper panels). When microsomes
from rabbit skeletal muscles were treated with 2-mercaptoethanol in the presencc of O,4
mg/ml SDS at 20℃ for lh, the ATPase activity was nearly completely lost (data not
shown) and the `Csolubilization"
of the Ca?'-ATPase was not observed at all (Fig, 2, lower
panels).
Discussion
The SR Ca2+-ATPase is inactivated by reducing agents at elevated temperatures (Fig.1). The inactivation preceeds more rapidly in the absence of Ca'L than in its presence,
ln the ubsence of Ca'!' an irreversible confbrmatienal change is induced even at moderatc
temperatures in the region of the calcium binding sites ol' the Ca"-ATPase []5]. It is
interesting to assume that the contbrmational challge would expose disulfide bond(s) of the
CaU'-ATPase to 2-mercaptoethanol, which would result in the rapid reduction of disulfide
bond(s) leading to the rapid inactivation of the ATPase activity.
The Ca'LATPase is composed of a single polypeptide and lacks thc polypeptide
equivalent to the B-subunit of the (Na', K') ATPase. Under the conclition where the
Ca?'-AFI]Pase is inactivated with reducing agents, at least one disulfide bond of the Ca2'-
ATPase is reduced. It $cems likely that the reduction of the disulfide bond within a
molecule of the Ca"" -A[[iPase is rcspensible for the inactivation.
The (Na', K') ATPase whose a-subunit has a similar transmembrane topology to the

The UOEH Association of Health Sciences
NII-Electronic Library Service
The UOEHAssociation ofHealth Sciences
258 Y)v[uToH et al
Ca'"'-A[I]Pase, is also inactivated upon thermal treatment in the presence of reducing agents.
Contrast to the Ca2'-ATPase, no change in the disulfide bond ofthe cr-subunit ofthe (Na',Ki) ATPase occurs, but instead the B-subunit is reduced l6, 7]. That is, for the (NaT,Ki) ATPase, the reduction of the disulfide bonds of t,he B-subunit is responsible fbr the
thermal inactivation with rcduc;ing agents.
One possible exp]anation is that the two protcins arc difl'erent in structure with respect
to the locations of the disulfide bridge, even if both proteins contain the samc numbcr ol'
disulfide bonds, 'I'he
nurnbers of the disu}fidc bond of the a-subunit of thc (Na', K")
iNTPase so far reported arc at least one [16], two [17] and three l71. Thc disulfidc bonds of
thc cr-subunit of thc (Nai, K') ATPase may bc occludcd and not bc susceptiblc to 2-
rnercaptoethanol while, for the CaU'-ATI'ase, at least one of the disulfide bonds may be
cxposed and susceptiblc to the reducing agent, If this is the case, the CaU'-ATPase has
uniquc structural featurcs around thc disulflde bonds that are not shared by the (Na', K')
AFIiPase. Alternatcly, it may bc possib]c that the B-subunit pret,ects the a-subunit firom
reduction by forming the crP complex in the (Na', K") A'i'Pase, even if disulfidc bonds of
the a-subunit are present at identical locations te those in the Ca?'-ATPase. Thereforc,
the Ca2T' -ATPase without the protector might bc easily reduced, leading to inactivation. ・
Thc a-subunit of the (Na", K') ATPasc was "solubilized"
by diluted SDS in the
prescnce of 2-mercaptoethanol, whe,rcas the, Ca2"-ATPase rcmained in the membranc.
Thc mechanism by which thc reducin.cr. agents {'acilitate the `Csolubilization"
of the (Na",K") A'l'Pase with diluted SDS is interesting but still an open question. Howcvcr, these
rcsults may be taken as evidence, although circumstantial, that the Ca2'-ATI'ase and the
(Na', K') Arl'Pase a-subunit are somewhat diflerent in sttuctural fcatures. Recent
immunological and biochemical studies on the CaL''-ATPase draw similar conclusions [17,l8],
Acknowledgments
This work was supported by Grant-in-Aid lbr Scicntific Rcsearch to M. K. from the
Ministry of Eclucation, Science and Culture of Japan. 1,Vc thank Dr. S. Noguchi of
Kyushu Institute ofTechnology for his usefu1 discussions and suggestions.
References
1. Jergenscn P I. & Andcrseii j I' (l988): Structural basis for El-E2 conformational transttion in Na, K-
pump and Ca-pump preteins, J rvtembrane Biol 1011: 95-1202. Noguchi S, pt'Iishina rvt, Kawamura M et al C1987): Exprcssion of functional CNa', K')-ATPase fi'om
cloi'ied eDNAs. FEBS Lctt 225/ 27-32
3, lvlcl)c)nough A A, C;eering K & Farley R A (199U)/
'1'hc sodium pump lleeds iLs i3-suburiit. FASEBJ
4i 1598-1605

The UOEH Association of Health Sciences
NII-Electronic Library Service
TheUOEHAssociation ofHealth Sciences
Tnactivation ofCR2'-ATPasc 259
4. Geering K (]990): Subunit assernbly and funcLional rrmturation of Na, K-ATPasc, J Membrar}e Biol
115: 109 -
E21 '
5. Kawamura M & Nog'uchi S (199I): Possible role of thc B-subunit in the expression of the sodium
purnp. In: The Sodium Purnp/ Stru{/turc, Mcchanism ancl Regulation (Kaplan J H, DeWecr P, ed).
The Rockcfcllcr University Press, New York pp 46-61
6. Kawamura pt'I & Nag'ano K (1984・): Evidence for essential disulfide bonds in the P-subunit of <Na', KT)-ATPase. Biochim Biophys Acta 774: 188-192
7. Kirley 'E' I. (1990): Inactivation of (Nai, K")-ATPase by "-mercaptocthanol. ,I Biol Chem
265i rl-227-,1・232
8. Kanazawa T, Ynmuda S, Yamamoto T et al (1971): Rcaction mechanism ot' the Ca? i-dependent
ATPasc
ofsarcoplasmic reticulurn frorTi skeletal musclc. j B{ochem 70: 95T12S
9. Kirley T L 0989): Determination of three disulfide bonds and one fi'ee sulfhydryL in thc P-subunit of'
(Na, K)-ATPasc. J Biol Chem 264・: 7185'7192
10. Lowry O H, Rosebrough .J J, Farr A L et at (1951): Protein measurement with the Folin phenol reagenL
J Biol Chem 193: 265-275
11. Laemirili U K (1970): Cleavage ofstructural protcins during the assembly of the hcad ofbacLeriophage
T4, Naturc (Lond) 227: 680r685
12, I,cpock J R, Rodahl A M, Zhang (: et al (I990): Thermal dcnaturation of the Ca!'i-fYl'1'ase of
sarcoplasinic Feticulurri reveals tN・vo thermoclynarriically irLdependent domains. Biochcn]istry
29: 68] -689
13. Kawamura }v(, Ohta 'I'
& paagano K (l980): Eil'ect of reclucing agcnts on the solubi]ization of rcnal
sodium and potassiurn ATPasc with cletcrgcnt. J Biecliem 87: 1327-1333
14・. Kawarnura pt{, Olimizo K, "torohashi ts4 et al (1985): Protcctive elll]ct of Na' and Ki against
illactivation of Na-, K+-ATPasc by high conccntrations oi' 2-mercaptoethanol at high ternpcratures.
Bioc,him Biophys Acta 821/ ll5-I20
15. (:heng K EI & Lepock J R (1992)/ Inactivation of calcium uptake b>, EGTA is due to an irreversibie
Lh(/,rmoLropic confbrmationa} chan.v.e in the calcium binding clomain of the Ca?'i-ATPasc.
Biochemistry 31:4074-4080
16. Gcvondyan N )v'I, Govonclyan V S, GavrilyeiJa E E et at (1989): Analysis of free sulfhydryl groups and
disulfide bonds in Nai, K'-ATI'ase. FEBS Lett 255: 265"'26817. Miller R P & J"arley R A (1990); B-subunit of CNa', KM)-ATPase contains thrcc disult"ide
bonds. Biochernistry 29: l52・1- -1532
18. rvtolnar E, Varga S & pt'tart,onosi A (1991): I)iflbrenccs in susceptibiliLy of various cation transport
A'I'1'ases te vanadntc-catalyzcd photoclcaN,agc, Biochim Biophys Acta 1068: 17-26I9. "・Iolnar E, Varg'a S,.)ona 1 et at (1992)/ lmmunologica] relatcdncss of thc sarcoplasmic ret{(/ulum Ca'i-
A'FPase and thc Na" , K
i -ATPasc. Biochim Biophys Acta 1 103: 2Hl-295

The UOEH Association of Health Sciences
NII-Electronic Library Service
The UOEH Assooiation of Health Soienoes
260 Y .MUTOH et al
筋小胞体 Ca2+ −ATP ア
ーゼ の 2一メ ル カ プ トエ タ ノ ー ル に よ る還元 と失活
(Na〒, K
+
)ATP アーゼ と の 対比
武藤 可信1・2
, 杉野 厚子1
, 東 監3
, 高杉 昌幸z
,黒岩 昭夫2
川村 越1
L産業医科大 学生 物学教室
2産業医科大学第二 内科学教室
3産業医科大学生化学教室
要 旨 : ウ サ ギ 骨格筋筋小胞体 Cai’+.ATP ア
ーゼ を 40 − 45℃ で 300 mM 2.メ ル カ プ トエ タ ノ ール
で 還 元 す る と,(Na+, K
+
)ATP ア ーゼ の 場合 と 同様 に,ATP アーゼ 活性が 消失 し た. Ca2+ −
ATP アー
ゼ の 還 元 ・失 活 は Ca2+不 在 下 で 迅 速 に 進行 し た. Ca2+ ・ATP ア ーゼ は 20℃ で も
希薄な SDS (0,4 mg /lnl )が存在する と 失活 した.〔Na+ K
−)ATP ア
ーゼ も同条件下で 失活す
る が .失活 し た (Na+
, K+
)ATP アー
ゼ が 10000 ×g,30min の 遠心 で 上 清 に 回 収 さ れ る の
に 対 し,Ca2+.ATP ア ーゼ は 沈澱 に 回収 さ れ た、
JUOEH (産業医大誌),14〈4 ): 253− 26D (1992>
N 工工一Eleotronio Library







![Prevention of doxorubicin-induce renal function abnormalities ......ATPase, Mg2+-ATPase and Na+, K+-ATPase activities [15, 16]. Turmeric is a golden spice derived from the rhizome](https://img.dokumen.tips/doc/110x75/61385b7c0ad5d20676493447/prevention-of-doxorubicin-induce-renal-function-abnormalities-atpase-mg2-atpase.jpg)

![OleicAcidInducesLungInjuryinMicethrough ...2 Mediators of Inflammation [16–18]. Oleic acid (OA) is an inhibitor of the Na/K-ATPase activity in bovine serum [19] and is also a Na/K-ATPase](https://img.dokumen.tips/doc/110x75/60c8bb5d6300d261b231ffb8/oleicacidinduceslunginjuryinmicethrough-2-mediators-of-iniammation-16a18.jpg)



















