Embed Size (px)
Citation preview

1329RESEARCH ARTICLE
INTRODUCTIONAxons ensheathed with myelin propagate action potentials in asaltatory manner resulting in significantly accelerated conductionvelocity and reduced energy requirements. In the peripheralnervous system (PNS), myelin is elaborated by Schwann cells.Their precursors emerge from the neural crest early in developmentand distribute amongst elongating axons, where they proliferateand migrate with advancing axons (Jessen et al., 1994; Wanner etal., 2006). As development proceeds they form discrete Schwanncell families that encircle bundles of axons, where they continue toproliferate as well as extend and retract radially oriented processesbetween axons, ultimately ensheathing all large caliber axons in aprocess referred to as radial sorting (Webster et al., 1973; Grove etal., 2007; Woodhoo et al., 2009) (for a review, see Woodhoo andSommer, 2008).
To initiate axon ensheathment, a Schwann cell develops alongitudinal groove that deepens as it extends processescircumferentially to fully enwrap the axon (Webster, 1971; Websteret al., 1973; Webster, 1984). Through mechanisms that remainpoorly understood (Bunge et al., 1989), the Schwann cellmembrane spirally wraps around the axon, following which,interlamellar Schwann cell cytoplasm is extruded and the apposingmembrane layers are stabilized by myelin-associated proteins(Wood et al., 1990). In addition to their seemingly lamellipodia-like mesaxons, Schwann cells generate microvilli at theelectrophysiologically specialized nodes of Ranvier that are locatedbetween successive myelin sheaths (Gatto et al., 2003).
The migratory, membrane-extending and microvilli-generatingcapacities of Schwann cells suggest an integral role for cytoskeletalremodeling in Schwann cell maturation. This possibility issupported by the growth cone-like properties displayed by theterminal processes of migrating Schwann cell precursors in vivo(Wanner et al., 2006) and by both the structure and composition ofthe mesaxons that are elaborated by myelinating Schwann cells invitro (Bacon et al., 2007). Also, in Schwann cell-neuron co-cultures, myelin formation is attenuated when actin polymerizationis disrupted by cytochalasin D (Fernandez-Valle et al., 1997).Similarly, in mice, myelin formation is attenuated when the Cdc42or Rac1 GTPase cytoskeletal regulators are deleted (Nodari et al.,2007; Benninger et al., 2007). Additionally, Rho GTPase and itskinase, ROCK, have been implicated in Schwann cell- and/oroligodendrocyte-mediated myelination (Melendez-Vasquez et al.,2004) (for a review, see Feltri et al., 2008).
Although a link between cytoskeletal dynamics and myelinationis widely appreciated, the specific effectors that modulatecytoskeletal reorganization in developing and myelinating Schwanncells are not well defined. However, one likely effector is theneuronal Wiskott-Aldrich syndrome protein [N-WASp; also known
Development 138, 1329-1337 (2011) doi:10.1242/dev.058677© 2011. Published by The Company of Biologists Ltd
1Department of Haematology, Xijing Hospital, Fourth Military Medical University,Xi'an, China. 2Departments of Medicine, Immunology and Molecular Genetics,University of Toronto, Samuel Lunenfeld and Toronto General Research Institutes,600 University Avenue, Toronto, ON M5G 1X5, Canada. 3Samuel LunenfeldResearch Institute, Mount Sinai Hospital, and Department of Molecular Genetics,University of Toronto, Toronto, ON M5G 1X5, Canada. 4Institute for Cell Biology,University of Bonn, Ulrich-Haberland-Str. 61A, 53121 Bonn, Germany. 5Division ofGenetics and Cell Biology, San Raffaele Scientific Institute, 20132 Milan, Italy.6Laboratory Neurobiologie et Transgenese, UPRES-EA3143, INSERM, CentreHospitalier Universitaire, 49033 Angers, France. 7Laboratory of DevelopmentalBiology, Royal Victoria Hospital, MUHC, McGill University, Montreal, QC H3A 1A1,Canada.
*These authors contributed equally to this work†Present address: Howard Hughes Medical Institute, Janelia Farm Research Campus,19700 Helix Drive, Ashburn, VA 20147, USA‡These authors co-directed this work§Authors for correspondence ([email protected]; [email protected])
Accepted 4 January 2011
SUMMARYSchwann cells elaborate myelin sheaths around axons by spirally wrapping and compacting their plasma membranes. Althoughactin remodeling plays a crucial role in this process, the effectors that modulate the Schwann cell cytoskeleton are poorly defined.Here, we show that the actin cytoskeletal regulator, neural Wiskott-Aldrich syndrome protein (N-WASp), is upregulated inmyelinating Schwann cells coincident with myelin elaboration. When N-WASp is conditionally deleted in Schwann cells at theonset of myelination, the cells continue to ensheath axons but fail to extend processes circumferentially to elaborate myelin.Myelin-related gene expression is also severely reduced in the N-WASp-deficient cells and in vitro process and lamellipodiaformation are disrupted. Although affected mice demonstrate obvious motor deficits these do not appear to progress, themutant animals achieving normal body weights and living to advanced age. Our observations demonstrate that N-WASp plays anessential role in Schwann cell maturation and myelin formation.
KEY WORDS: Myelination, Schwann cells, N-WASp (Wasl), Mouse
N-WASp is required for Schwann cell cytoskeletal dynamics,normal myelin gene expression and peripheral nervemyelinationFuzi Jin1,2,*, Baoxia Dong1,2,*, John Georgiou3, Qiuhong Jiang2, Jinyi Zhang2, Arjun Bharioke2,†, Frank Qiu2,Silvia Lommel4, M. Laura Feltri5, Lawrence Wrabetz5, John C. Roder3, Joel Eyer6, Xiequn Chen1,Alan C. Peterson7,‡,§ and Katherine A. Siminovitch2,‡,§
DEVELO
PMENT

1330
as Wiskott-Aldrich syndrome-like (Wasl) – Mouse GenomeInformatics], a member of the WASp family of cytoskeletalregulators. Like other family members, N-WASp links extracellularstimuli and actin polymerization (Wegner et al., 2008) via thebinding of its verprolin homology/connecting/acid region domainto the Arp2/3 complex. In cultured Schwann cells, N-WASplocalizes to the leading edge of extending processes where itsactivity depends upon interaction with Cdc42, a protein withessential roles in Schwann cell development (Benninger et al.,2007; Bacon et al., 2007). Further, the myelinating capacity ofcultured rat Schwann cells is impaired by an inhibitor of N-WASpactivity (Bacon et al., 2007), as is oligodendrocyte-mediatedmyelination in the central nervous system (CNS) when anotherWASp family member, Wave1, is deleted in vivo (Kim et al., 2006).Additionally, process remodeling in mature oligodendrocytesinvolves a complex including FAK, Fyn, the intracellular domainof Dcc and N-WASp (Rajasekharan et al., 2009).
Here, to investigate the in vivo role of N-WASp duringmyelination, we used conditional gene targeting to generate micein which N-WASp (Wasl) is deleted in Schwann cells as they initiatemyelin elaboration. N-WASp-deficient Schwann cells achieve anormal 1:1 relationship with large caliber axons, albeit on a delayedschedule. However, their capacity to elaborate myelin sheathsappears to be arrested completely and their expression of myelin-related genes is severely reduced. Additionally, the production oflamellipodia-like processes in vitro is markedly impaired. Althoughthese mutant mice manifest motor deficits characterized by tremorand mild ataxia, they are viable into advanced age. Theseobservations demonstrate that N-WASp is required for normalSchwann cell maturation and that it plays an essential role inmyelin formation, potentially by regulating the cytoskeletalremodeling that is required for the spiral extension of Schwann cellprocesses around axons.
MATERIALS AND METHODSConditional knockout and transgenic miceN-WASp floxed (N-WASpflx/flx), mPoTOTA-Cre (P0-Cre) and Z/EG reportermice have been reported by Lommel et al. (Lommel et al., 2001), Feltri etal. (Feltri et al., 1999a; Feltri et al., 1999b) and Novak et al. (Novak et al.,2000), respectively. C57BL/6 mice heterozygous for the N-WASp floxedallele were crossed with C57BL/6/P0-Cre mice to derive N-WASpflx/+/P0-Cre. These were crossed with N-WASpflx/flx mice to generate N-WASpflx/flx/P0-Cre (N-WASp–/– mutant) mice. Z/EG homozygous mice weremated to P0-Cre mice to derive Z-EG/P0-Cre mice for assessing P0-Creactivity. Genotypes were determined by PCR analysis of tail and/or sciaticnerve genomic DNA using primers that yield amplicons of 200 bp (wild-type), 306 bp (floxed) or 408 bp (recombined) from the N-WASp allele: 5�-TAACTCACATCCATAGTATC-3�, 5�-ACTTGGTCTGCTATGTATCGG-3� and 5�-TTGCACAGAGAGAATGAATG-3�.
To test gait abnormalities, paint was applied to paws of mutants andcontrol littermates and the footprint patterns generated by walking acrossa 12�2 inch Plexiglas track evaluated. Tremor activity was assessed usingan SDI Tremor Monitor (SD Instruments), which detects movementamplitude via a force transducer. For rotarod testing, mice were placed ona rod rotating at 5-7 rotations per minute (Columbus Instruments) andtested three times per day over 3 days for the length of time that theyremained on the rod.
ReagentsReagents included rabbit monoclonal (Cell Signaling Technology) andpolyclonal (Lommel et al., 2004) anti-N-WASp antibodies; rat monoclonalMbp antibody (Millipore); mouse monoclonal P0 and neurofilamentantibodies (Developmental Studies Hybridoma Bank); mouse monoclonalS100 and -actin antibodies (Sigma); rat monoclonal BrdU and mousemonoclonal Mag antibodies (Abcam); goat polyclonal antibodies to
Krox20 (Santa Cruz Biotechnology), Oct6 (Abcam) and Sox10 (R&DSystems); FITC- or Cy3/5-conjugated secondary antibodies (JacksonImmunoResearch); FITC-phalloidin (Molecular Probes); laminin (Sigma);and DAPI (Roche).
Immunofluorescence analyses and primary culturesSciatic nerve sections were fixed in 4% paraformaldehyde (PFA), blockedfor 1 hour with 10% goat serum/0.1% Triton X-100 in PBS and incubatedovernight at 4°C with primary antibodies. After washing in PBS, tissuesections were incubated for 1 hour with secondary antibody. Alternatively,Schwann cells were isolated from P4 sciatic nerves following nervedigestion with 1 mg/ml collagenase/dispase and 2.5% trypsin and Thy1.2antibody/complement-mediated fibroblast lysis (Honkanen et al., 2007) andmaintained in culture in DMEM supplemented with 10% fetal calf serum(FCS)/antibiotics. Dorsal root ganglia (DRG) were dissected as previouslydescribed from E13.5 wild-type mice (Päiväläinen et al., 2008) andcultured for 2 days in neural basal medium containing 10% FCS followedby supplementation with 10–5 M uridine/5�-fluoro-2�-deoxyuridine(Sigma). Schwann cells and DRG were then co-cultured in myelination-promoting media containing 50 g/ml ascorbic acid (Eldrige et al., 1987).For staining, Schwann cells or Schwann cell/DRG co-cultures were fixedin 4% PFA for 10 minutes at room temperature, the cells thenpermeabilized with 0.2% Triton X-100 for 5 minutes at room temperatureand incubated with primary antibody in blocking buffer (1% BSA in PBS)overnight at 4°C, followed by incubation with secondary antibodies inblocking buffer for 1 hour at room temperature. Staining was visualizedusing an Olympus FluoView 1000 inverted confocal microscope equippedwith fluorescence optics and Olympus FluoView FV100 software.
Immunoblotting analysisSciatic nerves were homogenized with a chilled mortar and pestle in RIPAbuffer containing protease inhibitors (Roche). The lysate was suspended inloading buffer, electrophoresed through a 10% polyacrylamide gel andtransferred to nitrocellulose. After overnight blocking, as previouslydescribed (Badour et al., 2004), membranes were incubated for 1 hour withprimary antibody followed by incubation for 1 hour with a 1/3000 dilutionof horseradish peroxidase-conjugated secondary antibody (Bio-Rad), thenECL substrate (Amersham Biosciences) for 1 minute, followed by a 1- to10-minute exposure to X-ray film (Eastman Kodak).
Morphological analysisSciatic nerves or spinal roots were immersion-fixed in either 4% PFA orby trans-cardiac perfusion with 2.5% glutaraldehyde/0.5% PFA in 0.1 Mphosphate buffered saline (pH 7.4) and subjected to alcohol dehydrationand Epon embedding. Alternatively, specimens were fixed for 2 hours in2% glutaraldehyde in 0.1 M sodium cacodylate buffer, dehydrated andembedded in Spurrs resin (Electron Microscopy Sciences). Semi-thinsections were stained with Toluidine Blue or Methylene Blue and examinedby light microscopy. Ultrathin sections were stained with uranyl acetate andlead citrate and examined by electron microscopy (JEOL JEM2010electron microscope).
Proliferation and TUNEL assaysMice were injected intraperitoneally with BrdU (300 mg/kg body weight)and sacrificed 24 hours later. The labeled sciatic nerves were then fixedwith 4% PFA, embedded in paraffin and 5 m sections stained with BrdUantibody. Apoptotic cell death was measured in transverse sciatic nervesections by TUNEL labeling using calf thymus TdT, biotin-labeled UTPand horseradish peroxidase-streptavidin (Boehringer). Counterstaining wasperformed with Haematoxylin. Sections were dehydrated and coverslippedand the percentage of TUNEL-positive cells in a sample of 500 wasdetermined.
Gene expression assaysTotal RNA (200 ng) prepared from sciatic nerve samples using the QiagenRNeasy Kit was reverse transcribed with Superscript II (Invitrogen)according to the manufacturer’s protocol. SYBR Green-based qRT-PCRwas performed using the ABI 7900HT system (Applied Biosystems).Primers were: Mag forward 5�-GTGGAGCTGAGTGTCATGTATG-3� and
RESEARCH ARTICLE Development 138 (7)
DEVELO
PMENT

reverse 5�-CATAGATGACCGTGGCTAGGA-3�; Mbp forward 5�-GT -GCCACATGTACAAGGACTC-3� and reverse 5�-TTGAAGA AATG -GACTACTGGGTT-3�; and P0 forward 5�-GATTCGGTAC TGCT -GGCTG-3� and reverse 5�-CATACAGCACTGGCGTCTG-3�. Levels ofmRNA were calculated by the comparative cycle threshold method (Pfaffl,2001) and expressed as fold increase relative to Gapdh levels.
RESULTSN-WASp is expressed in myelinating Schwann cellsN-WASp is expressed in many regions of the developing and maturebrain and also in cultured neurons, oligodendrocytes and Schwanncells (Tsuchiya et al., 2006; Bacon et al., 2007). To determine whenN-WASp might participate in Schwann cell development in vivo, wefirst evaluated the developmental programming of N-WASpaccumulation in the PNS of postnatal mice. N-WASp was readilydetectable by immunocytochemistry in sciatic nerves at postnatal day(P) 28, a time when myelin sheaths are approaching a mature state.
As demonstrated by its co-localization with S100, the N-WASpsignal was localized largely to a cytoplasmic domain adjacent to theexternal Schwann cell membrane. Intensely labeled profilesfrequently contained Schwann cell nuclei with the N-WASp signalextending from the perinuclear cytoplasm to define much, but not all,of the fiber circumference. Similarly intense N-WASp labeling wasnot observed in either the Schwann cell cytoplasm adjacent to theaxon nor in nuclei (Fig. 1A).
To determine when N-WASp expression initiates, weinvestigated sciatic nerve samples obtained from mice at earlierstages of postnatal development (Bray et al., 1977). As early as P1,coinciding with the initiation of PNS myelin elaboration, N-WASpwas readily detectable by immunofluorescence imaging (Fig. 1B).A broad ring of N-WASp staining partially overlapping that ofmyelin basic protein (Mbp) and extending to the cell periphery wasthe most characteristic labeling pattern observed at this age (seeFig. S1 in the supplementary material). Staining intensity appeared
1331RESEARCH ARTICLEN-WASp is required for PNS myelination
Fig. 1. N-WASp expression isdevelopmentally regulated in Schwanncells and its conditional deletion isassociated with motor impairment.(A)Confocal images of sciatic nerve transversesections from wild-type mouse reveal N-WASpto be localized around individual axons in apattern distinct from those of S100 andDAPI. (B)N-WASp immunofluorescenceincreases between P1 and P5, diminishes byP13 and remains low at P28. (C)qRT-PCRanalysis shows changes over time in N-WASp(left) and Mbp (right) mRNA levels in wild-typesciatic nerves (mean of three independentexperiments expressed relative to Gapdh +s.d.). (D)Two-month-old (left) and 7-month-old (right) mutant mice have a wider andoutward-pointing hindpaw stance. Footprintpatterns recorded from forepaws (red) andhindpaws (blue) reveal a widened ataxic gaitcompared with the narrow, even stride ofcontrols (see Movies 1 and 2 in thesupplementary material). (E)Rotarod testsperformed on three consecutive days (threetrials per day) showing significantly reducedmean (± s.e.m.) hold times in 2-month-oldmutant mice. (F)Tremor activity in 2-month-old mutant mice shown as a representativerecording of movement amplitude and agraph indicating numbers of tremor events ateach frequency (mean ± s.e.m.) (datarepresentative of three independentexperiments).
DEVELO
PMENT

1332
to increase to peak levels near P5 and moderated by P13, at whichtime a distribution pattern characterized by thinner and incompleteperipheral rings, typical of more mature fibers, was common (Fig.1B). Quantitative real-time PCR (qRT-PCR) assays revealed thatN-WASp and Mbp mRNA accumulated on similar programs andindicated that N-WASp expression was coincident with, orpreceded, the initiation of myelination (Fig. 1C).
Mice with N-WASp-deficient Schwann cellsTo explore the role of N-WASp in Schwann cell-mediatedmyelination, we derived mice in which N-WASp was specificallyinactivated in premyelinating Schwann cells. Mice homozygous foran N-WASp floxed allele (N-WASpflx/flx) were bred with N-WASpflx/+
mice carrying a P0-Cre transgene (Feltri et al., 1999a; Lommel etal., 2001). As the P0 (Mpz – Mouse Genome Informatics) sequencedrives expression in Schwann cells during the perinatal period(Feltri et al., 1999a; Feltri et al., 1999b), we expected accumulatedCre to delete floxed N-WASp at the beginning of PNS myelination.In sciatic nerve samples, both a Cre-sensitive GFP reporter and Creexpression followed the expected developmental program, bothbeing detectable at P1, increasing through P5 and stabilizingthereafter (see Fig. S2 in the supplementary material). Evaluationof Cre expression revealed ~65% of all DAPI-labeled nuclei to beCre-positive at P1, increasing to 99% by P28, thus suggesting thatmost Schwann cells express effective levels of Cre.
Crosses involving floxed N-WASp and P0-Cre alleles resultedin the predicted Mendelian ratios of wild-type, floxed,heterozygous and homozygous mutant (N-WASpflx/flx/P0-Cre,referred to as N-WASp–/–) offspring. The deleted N-WASp allelewas detected only in Schwann cells from mice bearing the P0-Cre transgene and the floxed N-WASp allele (see Fig. S3A in thesupplementary material). Consistent with widespreadinactivation of the N-WASp gene in Schwann cells, N-WASpprotein levels were profoundly reduced in mutant sciatic nerves(see Fig. S3B in the supplementary material). Moreover, inimmunolabeled sciatic nerve samples, the perineurium labeledintensely, whereas N-WASp was undetectable in all but rareSchwann cells (see Fig. S3C in the supplementary material).
Motor impairment in N-WASp–/– miceN-WASp–/– mice appeared normal at birth but, by P14, showedlocomotor anomalies including a wide-based ataxic gait withextraneous steps and impaired rotarod endurance (Fig. 1D,E andsee Movies 1 and 2 in the supplementary material). Mutants alsomanifested an abnormally wide hindlimb stance while sitting andlow frequency (10-20 Hz) tremors (Fig. 1D,F). Interestingly, thesemice achieved normal body weights and their behavioralphenotypes progressed minimally over an 8-month follow-upperiod.
N-WASp-null Schwann cells fail to elaboratemyelinAs shown in Fig. 2, all large caliber (>2 m) axons coursingthrough mutant sciatic nerves (Fig. 2A,B) and ventral roots (Fig.2C) were ensheathed. However, most fibers were amyelinated(>99% in sciatic nerve; see Table S1 in the supplementary material)and, as seen in many demyelinating or amyelinating conditions,axons without normal myelin sheaths failed to undergo normalradial growth (see Table S1 in the supplementary material).Exceptional myelinated fibers were encountered in the mutants(<1%; see Table S1 in the supplementary material) and, consistentwith the enhanced radial growth of myelinated axons, such fibers
appeared as prominent components in nerve transverse sections(Fig. 2C). Such myelin sheaths appeared qualitatively normal andrevealed normal interlamellar spacing (13.17±0.48, versus13.41±0.46 nm in controls) suggesting that these rare Schwanncells might have escaped N-WASp inactivation. Nonetheless,relative to the calibers of their associated axons, these sheaths wereof modestly reduced thickness, achieving g-ratios of 0.79±0.06,versus 0.67±0.06 in controls (mean of 31 myelinated fibers).Finally, regional heterogeneity may exist in the frequency at whichmyelin sheaths arise in this model as they were encountered inmost spinal roots but were virtually absent in sciatic nerves (Fig.2A,B). Interestingly, enlarged blood vessels were also prominentin mutant spinal roots (Fig. 2C).
At birth, many axons in normal sciatic nerves remain closelypacked into bundles surrounded by Schwann cell families. Througha complex program involving process extension/retraction and celldivision, Schwann cells segregate such bundles to arrive at a 1:1relationship with large caliber axons. This radial sorting wassuccessfully achieved in sciatic nerves of mature N-WASp–/–
animals. However, in mutant nerves, axon bundles were stillprominent in the sciatic nerves of P5 mice, whereas in control miceat this age radial sorting was largely complete (Fig. 2A,B; see Fig.S4A and Table S1 in the supplementary material). Thus, the mutantSchwann cells remained capable of completing the radial sortingprocess, albeit over an extended period.
Mutant fibers that failed to elaborate myelin sheathsdemonstrated numerous ultrastructural features shared withpromyelinating fibers in normal perinatal mice (Webster, 1971).Processes enwrapping axons often comprised inner- and outer-facing Schwann cell membrane with little cytoplasm between.Where their processes confronted each other, no evidence forcontinued circumferential movements was apparent. Instead, suchmesaxon contacts remained in apposition (Fig. 2E,F) or developedmultiple short interdigitations (Fig. 2G). Also common were oneor more Schwann cell processes extending away from the axon intointerfiber space (Fig. 2E,F). All outward-oriented Schwann cellmembrane was associated with a continuous and non-redundantbasal lamina and collagen fibrils were deposited densely betweenthe amyelinated fibers. In Remak fibers, where Schwann cellsensheath multiple small diameter C-fibers, but elaborate no myelin,qualitative anomalies were not obvious (see Fig. S4B and Table S1in the supplementary material) and neither the number nor caliberof the ensheathed axons was remarkable (see Table S1 in thesupplementary material).
With the exception of Schwann cells, N-WASp was expressedwidely in mutant mice bearing both the P0-Cre transgene andfloxed N-WASp alleles (data not shown). Therefore, theamyelination observed in the PNS of N-WASp mutants was likelyto be the result of a Schwann-cell-autonomous defect. To test thisconclusively, we used mixed cultures in which mutant or wild-typeSchwann cells were assessed for their capacity to myelinate axonsextending from wild-type dorsal root ganglia neurons. Consistentwith a Schwann-cell-autonomous defect, myelination, as assessedby Mbp immunoreactivity, was detected only in cultures containingwild-type Schwann cells (Fig. 2H).
N-WASp deficiency alters Schwann cell numbersand proliferationWe next compared Schwann cell numbers (Brown and Asbury,1981), proliferation and apoptosis in mutant versus control-typenerves. Cell numbers were equivalent at P1 (Fig. 3A,B), but by P7and at all later ages examined the mutant nerves contained
RESEARCH ARTICLE Development 138 (7)
DEVELO
PMENT

significantly more cells and these demonstrated greaterproliferation as revealed by Ki67 labeling and bromodeoxyuridine(BrdU) incorporation (Fig. 3B,C). By contrast, TUNEL assaysrevealed no differences in the percentages of TUNEL-positive cellsin mutant compared with control (6.58±0.54% versus 6.64±0.83%,P0.81) nerves (data not shown). As the extent of caspase 3staining was also similar in mutant and control samples (data notshown), the enhanced Schwann cell proliferation observed inmutant nerves does not appear to be in response to increasedSchwann cell death. Rather, their increased proliferation isconsistent with the limited capacity of non-myelin-forming, ascompared with myelin-forming, Schwann cells to extend for longdistances along their associated axons. Thus, relatively more non-myelin-forming Schwann cells are required to ensheath acomparable length of axon.
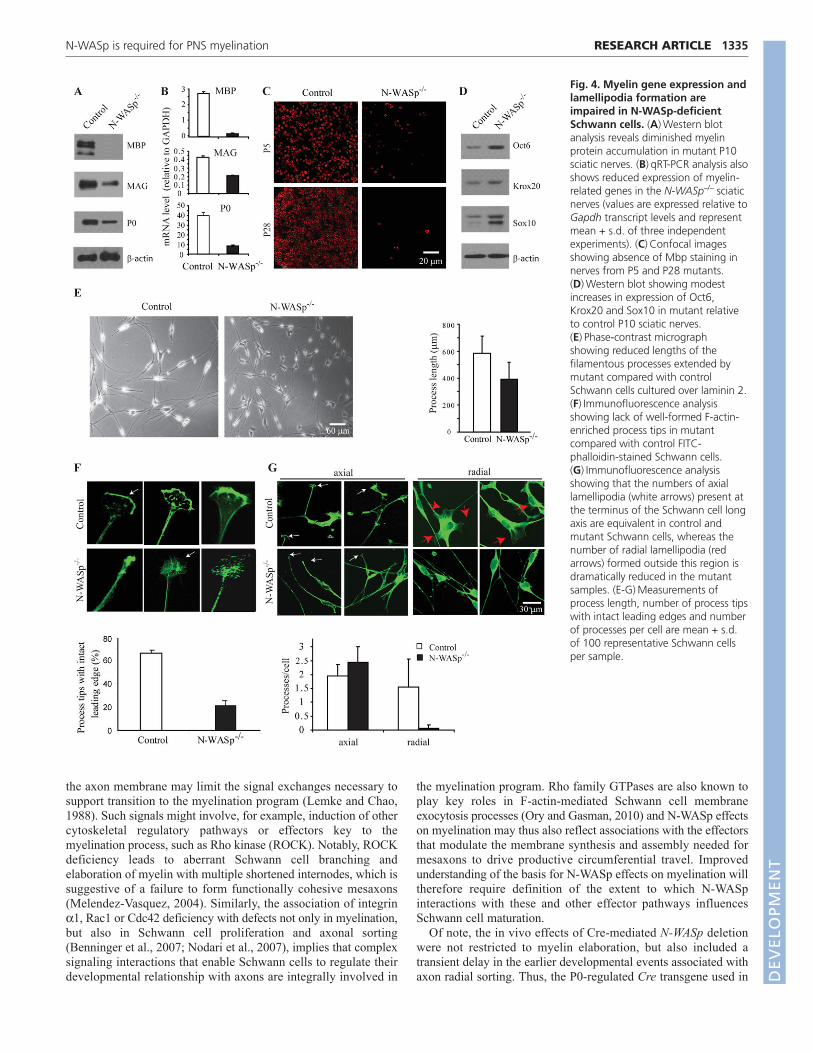
N-WASp modulates myelin-related geneexpressionGenes encoding a group of myelin proteins are coordinatelyupregulated as Schwann cells initiate myelination. To determinewhether the lack of N-WASp modulates this expression program,levels of three such myelin proteins were assessed in P5 sciaticnerve samples. In the hypercellular mutant samples, bothimmunoblotting and qRT-PCR analyses revealed reduced
accumulation of Mag and P0 (Fig. 4A,B), whereas only traceamounts of transcript and negligible protein were apparent for thelater-accumulating Mbp (Fig. 4A-C). Notably, these findingscannot be explained by a generalized transcription defect, asexpression of Oct6 (Pou3f1 – Mouse Genome Informatics),Krox20 (Egr2 – Mouse Genome Informatics) and Sox10, which areall promyelinating transcription factors with key roles in Schwanncell differentiation and myelin gene activation (Svaren and Meijer,2008; Ghislain and Charnay, 2006), was increased in the mutantcompared with control samples (Fig. 4D).
N-WASp modulates lamellipodia formationCultured N-WASp-deficient Schwann cells were assessed for theircapacity to form the actin-based processes that are thoughtnecessary for spiral membrane wrapping and myelin formation(Bunge et al., 1989; Feltri et al., 2002). After 16 hours in culture,similar numbers of processes extended from the main cell axis ofboth mutant and wild-type cells, but those extending from themutant cells were only ~60% the normal length (Fig. 4E).Although phalloidin staining revealed that an equivalent number oflamellipodia formed at cell termini, only wild-type cells elaboratedwell-defined, F-actin-rich leading edges (Fig. 4F,G). Additionally,the number of processes extending from the periphery of the cellbody was reduced in N-WASp–/– cells.
1333RESEARCH ARTICLEN-WASp is required for PNS myelination
Fig. 2. N-WASp is required for Schwann cellmyelination. (A-C)Light microscopy of semi-thin transverse Toluidine Blue-stained sectionsfrom control and mutant P5 (A) and P60 (B)sciatic nerves and P60 L3 ventral roots (C). AtP5, axons in control nerves are sorted and manyhave myelin sheaths, whereas in mutantsamples bundles of unsorted axons remainwithin Schwann cell families. Asterisks indicateunsorted axon bundles. At P60, all large caliberaxons in control nerves are myelinated, whereasin the mutant only rare myelinated fibers areobserved amongst ensheathed amyelinatedaxons (see Table S1 in the supplementarymaterial). (D-G)Electron micrographs of control(D) and mutant (E-G) fibers in P60 samples.Although axon membranes are coveredcompletely in both control and mutant samples,mutant Schwann cells do not show spiralmembrane wrapping. Where mesaxons abut,they show little evidence of furthercircumferential extension (F), often elaboratingmultiple short interdigitating processes (G). Inboth mutant and control nerves, all fibers aresurrounded by basal lamina and the interfiberspace is richly invested with collagen fibrils.(H)Immunofluorescence analysis of control andmutant Schwann cells co-cultured with controldorsal root ganglia neurons co-stained withDAPI and antibodies to neurofilament and Mbp.The numbers of DAPI-stained nuclei andneurofilament-stained axons are equivalent inboth culture types, but mutant cells do notproduce Mbp-stained internodes (datarepresentative of five independentexperiments).
DEVELO
PMENT

1334
DISCUSSIONOur study of Schwann cell maturation in the context of N-WASpdeficiency identifies crucial roles for N-WASp in the elaborationof PNS myelin. The amyelination phenotype manifested by themutant Schwann cells studied here is likely to be attributable, atleast in part, to loss of N-WASp actin polymerization activity. Thecytoskeleton supporting the growth cones elaborated by migratingand elongating Schwann cells is normally enriched in actin, andperturbation of the actin cytoskeleton is associated with defectiveSchwann cell morphology and myelin production (Fernandez-Valleet al., 1997; Melendez-Vasquez et al., 2004; Nodari et al., 2007;Benninger et al., 2007) (for a review, see Feltri et al., 2008). Thus,N-WASp deficiency might result in an impaired cytoskeletalcapacity to evoke the inner Schwann cell mesaxon movement thatdrives the elaboration of spirally wrapped membrane needed formyelin sheath production. This possibility is supported by theabsence of mesaxon-mediated plasma membrane circumnavigationfollowing axon ensheathment in the N-WASp-deficient cells invivo and also by the multiple cytoskeletal anomalies manifested bythe N-WASp-deficient Schwann cells cultured from mutant nerves.Thus, in contrast to the actin-rich growth cones observed in bothaxial and radial lamellipodia of cultured wild-type Schwann cells,the axial processes elaborated by cultured N-WASp-deficient cells
were reduced in length and lacked defined F-actin-rich leadingedges. Numbers of radial lamellipodia also were decreaseddramatically in the mutant cells. These findings are consistent withthe aberrant process extension observed in cultured Schwann cellsand oligodendrocytes treated with an N-WASp inhibitor (Bacon etal., 2007) and suggest that the crucial role for N-WASp in Schwanncell-mediated myelination reflects, at least partially, the regulationof actin rearrangements that are integral to the formation andfunction of Schwann cell lamellipodia.
In addition to actin regulatory roles for N-WASp at the Schwanncell membrane, the localization of N-WASp to multiple cellularcompartments, including the perinuclear cytoplasm external to themyelin sheath, suggests that the influence of N-WASp onmyelination might be realized through multiple effects on Schwanncell biology. When co-cultured with neurons, for example,Schwann cells normally undergo long extensions along the axon ofcontact prior to initiating the circumferential mesaxonal movementthat drives deposition of spirally wrapped plasma membrane(Bunge et al., 1989). By contrast, as is consistent with the extensiveSchwann cell hypercellularity observed in mutant nerves,longitudinal extension of N-WASp-deficient Schwann cells ismarkedly foreshortened. The consequent failure of pre-myelinatingSchwann cells to achieve significantly extensive apposition with
RESEARCH ARTICLE Development 138 (7)
Fig. 3. Increased number and proliferation of N-WASp-deficient Schwann cells. (A)Haematoxylin- and Eosin-stained sciatic nerve transverse sections show increasednumbers of Schwann cell nuclei in mutant nerves at alltimepoints studied (bar chart). (B,C)The number ofproliferating cells is increased in mutant compared withcontrol nerve sections as assessed by DAPI and Ki67 stainingat P5 and P28 (B) and by BrdU staining of cells 24 hoursafter BrdU intraperitoneal injection at P5 and P24 (C). Dataare expressed as mean + s.e.m. of at least four independentexperiments.
DEVELO
PMENT
the axon membrane may limit the signal exchanges necessary tosupport transition to the myelination program (Lemke and Chao,1988). Such signals might involve, for example, induction of othercytoskeletal regulatory pathways or effectors key to themyelination process, such as Rho kinase (ROCK). Notably, ROCKdeficiency leads to aberrant Schwann cell branching andelaboration of myelin with multiple shortened internodes, which issuggestive of a failure to form functionally cohesive mesaxons(Melendez-Vasquez, 2004). Similarly, the association of integrin1, Rac1 or Cdc42 deficiency with defects not only in myelination,but also in Schwann cell proliferation and axonal sorting(Benninger et al., 2007; Nodari et al., 2007), implies that complexsignaling interactions that enable Schwann cells to regulate theirdevelopmental relationship with axons are integrally involved in
the myelination program. Rho family GTPases are also known toplay key roles in F-actin-mediated Schwann cell membraneexocytosis processes (Ory and Gasman, 2010) and N-WASp effectson myelination may thus also reflect associations with the effectorsthat modulate the membrane synthesis and assembly needed formesaxons to drive productive circumferential travel. Improvedunderstanding of the basis for N-WASp effects on myelination willtherefore require definition of the extent to which N-WASpinteractions with these and other effector pathways influencesSchwann cell maturation.
Of note, the in vivo effects of Cre-mediated N-WASp deletionwere not restricted to myelin elaboration, but also included atransient delay in the earlier developmental events associated withaxon radial sorting. Thus, the P0-regulated Cre transgene used in
1335RESEARCH ARTICLEN-WASp is required for PNS myelination
Fig. 4. Myelin gene expression andlamellipodia formation areimpaired in N-WASp-deficientSchwann cells. (A)Western blotanalysis reveals diminished myelinprotein accumulation in mutant P10sciatic nerves. (B)qRT-PCR analysis alsoshows reduced expression of myelin-related genes in the N-WASp–/– sciaticnerves (values are expressed relative toGapdh transcript levels and representmean + s.d. of three independentexperiments). (C)Confocal imagesshowing absence of Mbp staining innerves from P5 and P28 mutants.(D)Western blot showing modestincreases in expression of Oct6,Krox20 and Sox10 in mutant relativeto control P10 sciatic nerves.(E)Phase-contrast micrographshowing reduced lengths of thefilamentous processes extended bymutant compared with controlSchwann cells cultured over laminin 2.(F)Immunofluorescence analysisshowing lack of well-formed F-actin-enriched process tips in mutantcompared with control FITC-phalloidin-stained Schwann cells.(G)Immunofluorescence analysisshowing that the numbers of axiallamellipodia (white arrows) present atthe terminus of the Schwann cell longaxis are equivalent in control andmutant Schwann cells, whereas thenumber of radial lamellipodia (redarrows) formed outside this region isdramatically reduced in the mutantsamples. (E-G)Measurements ofprocess length, number of process tipswith intact leading edges and numberof processes per cell are mean + s.d.of 100 representative Schwann cellsper sample.
DEVELO
PMENT

1336
deriving the mutant Schwann cells investigated here must beexpressed at levels sufficient to cause deletion of floxed N-WASpalleles prior to the initiation of myelination, when genes such as P0are upregulated. Although the sorting anomaly implies a role forN-WASp in early Schwann cell maturation events, the eventualrealization of the mature relationship between each Schwann celland a single axon demonstrates that Schwann cells deficient in N-WASp do successfully complete radial sorting. Schwann cellmitosis is also not adversely affected by N-WASp deficiency, asintense Schwann cell proliferation accompanies the normal sortingprocess. In further support of this conclusion, the size of theSchwann cell population in the amyelinated nerves of maturemutant mice was markedly increased.
N-WASp deficiency in Schwann cells was also associated withdiminished expression of multiple myelin-associated genes,including Mag, P0 and Mbp. This finding was, however, notindicative of a global transcription deficit as expression of thepromyelinating transcription factors Oct6, Knox20 and Sox10appeared normal or increased. Thus, failure of the N-WASp-deficient Schwann cells to realize a normal myelin gene expressionprogram could reflect their apparent developmental immaturity(Fernandez-Valle et al., 1997). However, evidence is growing fordirect roles of nuclear actin and its modulating partners intranscriptional regulation (for a review, see Gieni and Hendzel,2009); both N-WASp and related WASp molecules are nowdirectly implicated in transcriptional activities relevant to theircellular functions. WASp, for example, has recently been shown todirectly regulate epigenetic changes crucial to the transcriptionalinduction of the TBX21 gene required for the differentiation of Thelper cells (Taylor et al., 2010). Similarly, N-WASp has beenidentified within a transcription complex that is recruited inresponse to retinoic acid to the HoxB2 gene enhancer, and the actinpolymerization evoked by N-WASp is required for the induction ofthese genes (Ferrai et al., 2009). The extent to which transcriptionalregulatory roles for N-WASp might contribute to the impairedexpression of myelin genes in N-WASp-deficient Schwann cellsrequires further investigation.
Interestingly, the multiple Schwann cell deficits associatedwith N-WASp deficiency appear to be tolerated relatively wellby the mutant mice, in which neither the amyelinated PNSphenotype nor the motor abnormalities significantly evolvedwith age. Such stability is remarkable as it contrasts with thesevere axon pathology and progressive neuropathies associatedwith many myelin gene mutants (for a review, see Nave, 2010).The stability of this phenotype provides a unique opportunity toinvestigate the regulation of peripheral nerve fiber maturation inthe amyelination context and also renders these mutant mice ofparticular value in delineating the molecular pathways thatconnect N-WASp and/or N-WASp-modulated cytoskeletalremodeling to myelination and the control of myelin geneexpression.
AcknowledgementsThis work was supported by grants from the Canadian Institute for HealthResearch (MOP12136) to K.A.S. and the Multiple Sclerosis Society of Canadato K.A.S. and A.C.P. K.A.S. holds a Canada Research Chair and the ShermanFamily Chair in Genomic Medicine.
Competing interests statementThe authors declare no competing financial interests.
Supplementary materialSupplementary material for this article is available athttp://dev.biologists.org/lookup/suppl/doi:10.1242/dev.058677/-/DC1
ReferencesBacon, C., Lakics, V., Machesky, L. and Rumsby, M. (2007). N-WASP regulates
extension of filopodia and processes by oligodendrocyte progenitors,oligodendrocytes, and Schwann cells-implications for axon ensheathment atmyelination. Glia 55, 844-858.
Badour, K., Zhang, J., Shi, F, Leng, Y., Collins, M. and Siminovitch, K. A.(2004). Fyn and PTP-PEST-mediated regulation of Wiskott-Aldrich syndromeprotein (WASp) tyrosine phosphorylation is required for coupling T cell antigenreceptor engagement to WASp effector function and T cell activation. J. Exp.Med. 199, 99-112.
Benninger, Y., Thurnherr, T., Pereira, J. A., Krause, S., Wu, X., Chrostek-Grashoff, A., Herzog, D., Nave, K. A., Franklin, R. J., Meijer, D. et al.(2007). Essential and distinct roles for cdc42 and rac1 in the regulation ofSchwann cell biology during peripheral nervous system development. J. Cell Biol.177, 1051-1061.
Bray, G., Perkins, S., Peterson, A. and Aguayo, A. (1977). Schwann cellmultiplication deficit in nerve roots of newborn dystrophic mice. J. Neurol. Sci.32, 203-212.
Brown, M. J. and Asbury, A. K. (1981). Schwann cell proliferation in thepostnatal mouse: timing and topography. Exp. Neurol. 74, 170-186.
Bunge, R. P., Bunge, M. B. and Bates, M. (1989). Movements of the Schwanncell nucleus implicate progression of inner (axons related) Schwann cell processduring myelination. J. Cell Biol. 109, 273-284.
Eldrige, C. F., Bunge, M. B., Bunge, R. P. and Wood, P. M. (1987).Differentiation of axon-related Schwann cells in vitro. I. Ascorbic acid regulatesbasal lamina assembly and myelin formation. J. Cell Biol. 105, 1023-1034.
Feltri, M. L., D’Antonio, M., Previtali, S., Fasolini, M., Messing, A. andWrabetz, L. (1999a). P0-Cre transgenic mice for inactivation of adhesionmolecules in Schwann cells. Ann. N. Y. Acad. Sci. 883, 116-123.
Feltri, M. L., D’Antonio, M., Quattrini, A., Numerato, R., Arona, M., Previtali,S., Chiu, S. Y., Messing, A. and Wrabetz, L. (1999b). A novel P0 glycoproteintransgene activates expression of lacZ in myelin-forming Schwann cells. Eur. J.Neurosci. 11, 1577-1586.
Feltri, M. L., Porta, D. G., Previtali, S. C., Nodari, A., Migliavacca, B., Cassetti,A., Littlewood-Evans, A., Reichardt, L. F., Messing, A., Quattrini, A. et al.(2002). Conditional disruption of beta 1 integrin in Schwann cells impedesinteractions with axons. J. Cell Biol. 156, 199-209.
Feltri, M. L., Suter, U. and Relvas, J. B. (2008). The function of RhoGTPases inaxon ensheathment and myelination. Glia 56, 1508-1517.
Fernandez-Valle, C., Gorman, D., Gomez, A. M. and Bunge, M. B. (1997).Actin plays a role in both changes in cell shape and gene expression associatedwith Schwann cell myelination. J. Neurosci. 17, 241-250.
Ferrai, C., Naum-Onganía, G., Longobardi, E., Palazzolo, M., Disanza, A.,Diaz, V. M., Crippa, M. P., Scita, G. and Blasi, F. (2009). Induction of HoxBtranscription by retinoic acid requires actin polymerization. Mol. Biol. Cell 20,3543-3551.
Gatto, C. L., Walker, B. J. and Lambert, S. (2003). Local ERM activation anddynamic growth cones at Schwann cell tips implicated in efficient formation ofnodes of Ranvier. J. Cell Biol. 162, 489-498.
Ghislain, J. and Charnay, P. (2006). Control of myelination in Schwann cells: aKrox20 cis-regulatory element integrates Oct6, Brn2 and Sox10 activities. EMBORep. 7, 52-58.
Gieni, R. S. and Hendzel, M. J. (2009). Actin dynamics and functions in theinterphase nucleus: moving toward an understanding of nuclear polymeric actin.Biochem. Cell Biol. 87, 283-306.
Grove, M., Komiyama, N. H., Nave, K. A., Grant, S. G., Sherman, D. L. andBrophy, P. J. (2007). FAK is required for axonal sorting by Schwann cells. J. CellBiol. 176, 277-282.
Honkanen, H., Lahti, O., Nissinen, M., Myllylä, R. M., Kangas, S.,Päiväläinen, S., Alanne, M. H., Peltonen, S., Peltonen, J. and Heape, A. M.(2007). Isolation, purification and expansion of myelination-competent, neonatalmouse Schwann cells. Eur. J. Neurosci. 26, 953-964.
Jessen, K. R., Brennan, A., Morgan, L., Mirsky, R., Kent, A., Hashimoto, Y.and Gavrilovic, J. (1994). The Schwann cell precursor and its fate: a study ofcell death and differentiation during gliogenesis in rat embryonic nerves. Neuron12, 509-527.
Kim, H. J., DiBernardo, A. B., Sloane, J. A., Rasband, M. N., Solomon, D.,Kosaras, B., Kwak, S. P. and Vartanian, T. K. (2006). WAVE1 is required foroligodendrocyte morphogenesis and normal CNS myelination. J. Neurosci. 26,5849-5859.
Lemke, G. and Chao, M. (1988). Axons regulate Schwann cell expression of themajor myelin and NGF receptor genes. Development 102, 499-504.
Lommel, S., Benesch, S., Rottner, K., Franz, T., Wehland, J. and Kuhn, R.(2001). Actin pedestal formation by enteropathogenic Escherichia coli andintracellular motility of Shigella flexneri are abolished in N-WASp-defective cells.EMBO Rep. 2, 850-857.
Lommel, S., Benesch, S., Rohde, M., Wehland, J. and Rottner, K. (2004).Enterohaemorrhagic and enteropathogenic Escherichia coli use differentmechanisms for actin pedestal formation that converge on N-WASP. Cell.Microbiol. 6, 243-254.
RESEARCH ARTICLE Development 138 (7)
DEVELO
PMENT

Melendez-Vasquez, C. V., Einheber, S. and Salzer, J. L. (2004). Rho kinaseregulates Schwann cell myelination and formation of associated axonal domains.J. Neurosci. 24, 3953-3963.
Nave, K. A. (2010). Myelination and the trophic support of long axons. Nat. Rev.Neurosci. 11, 275-283.
Nodari, A., Zambroni, D., Quattrini, A., Court, F. A., D’Urso, A., Recchia, A.,Tybulewicz, V. L., Wrabetz, L. and Feltri, M. L. (2007). Beta1 integrinactivates Rac1 in Schwann cells to generate radial lamellae during axonal sortingand myelination. J. Cell Biol. 177, 1063-1075.
Novak, A., Guo, C., Yang, W., Nagy, A. and Lobe, C. G. (2000). Z/EG, a doublereporter mouse line that expresses enhanced green fluorescent protein uponCre-mediated excision. Genesis 28, 147-155.
Ory, S. and Gasman, S. (2010). Rho GTPases and exocytosis: what are themolecular links? Semin. Cell Dev. Biol. 22, 27-32.
Päiväläinen, S., Nissinen, M., Honkanen, H., Lahti, O., Kangas, S. M.,Peltonen, J., Peltonen, S. and Heape, A. M. (2008). Myelination in mousedorsal root ganglion/Schwann cell cocultures. Mol. Cell. Neurosci. 37, 568-578.
Pfaffl, M. W. (2001). A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 29, 9.
Rajasekharan, S., Baker, K. A., Horn, K. E., Jarjour, A. A., Antel, J. P. andKennedy, T. E. (2009). Netrin 1 and Dcc regulate oligodendrocyte processbranching and membrane extension via Fyn and RhoA. Development 136, 415-426.
Svaren, J. and Meijer, D. (2008). The molecular machinery of myelin genetranscription in Schwann cells. Glia 56, 1541-1551.
Taylor, M. D., Sadhukhan, S., Kottangada, P., Ramgopal, A., Sarkar, K.,D’Silva, S., Selvakumar, A., Candotti, F. and Vyas, Y. M. (2010). Nuclear roleof WASp in pathogenesis of dysregulated TH1 immunity in human Wiskott-Aldrich syndrome. Sci. Transl. Med. 2, 37ra44.
Tsuchiya, D., Kitamura, Y., Takata, K., Sugisaki, T., Taniguchi, T., Uemura, K.,Miki, H., Takenawa, T. and Shimohama, S. (2006). Developmental expressionof neural Wiskott-Aldrich syndrome protein (N-WASP) and WASP familyverprolin-homologous protein (WAVE)-related proteins in postnatal rat cerebralcortex and hippocampus. Neurosci. Res. 56, 459-469.
Wanner, I. B., Mahoney, J., Jessen, K. R., Wood, P. M., Bates, M. and Bunge,M. B. (2006). Invariant mantling of growth cones by Schwann cell precursorscharacterize growing peripheral nerve fronts. Glia 54, 424-438.
Webster, H. (1984). Development of peripheral nerve fibers. In PeripheralNeuropathy (ed. P. T. Dyck, E. H. Lambert and R. Bunges), pp. 329-359.Philadelphia, PA: W. B. Saunders.
Webster, H. D. (1971). The geometry of peripheral myelin sheaths during theirformation and growth in rat-sciatic nerves. J. Cell Biol. 48, 348-367.
Webster, H. D., Martin, R. and O’Connell, M. F. (1973). The relationshipsbetween interphase Schwann cells and axons before myelination: a quantitativeelectron microscopic study. Dev. Biol. 32, 401-416.
Wegner, A. M., Nebhan, C. A., Hu, L., Majumdar, D., Meier, K. M., Weaver,A. M. and Webb, D. J. (2008). N-WASp and the Arp2/3 complex are criticalregulators of actin in the development of dendritic spines and synapses. J. Biol.Chem. 283, 15912-15920.
Wood, P., Moya, F., Eldridge, C., Owens, G., Ranscht, B., Schachner, M.,Bunge, M. and Bunge, R. (1990). Studies of the initiation of myelination bySchwann cells. Ann. N. Y. Acad. Sci. 605, 1-14.
Woodhoo, A. and Sommer, L. (2008). Development of the Schwann cell lineage:from the neural crest to the myelinated nerve. Glia 56, 1481-1490.
Woodhoo, A., Alonso, M. B., Droggiti, A., Turmaine, M., D’Antonio, M.,Parkinson, D. B., Wilton, D. K., Al-Shawi, R., Simons, P., Shen, J. et al.(2009). Notch controls embryonic Schwann cell differentiation, postnatalmyelination and adult plasticity. Nat. Neurosci. 12, 839-847.
1337RESEARCH ARTICLEN-WASp is required for PNS myelination
DEVELO
PMENT





























