Embed Size (px)
Citation preview

Prq. N-Psychop hmmamL d BtoL Psychic-d_ 1996. Vol. 20. pp. 827-636
Copyrtght Q 1996 ELwvier Science Inc.
Pllntedtntheu6A. Aurighti3reserved
0276.5646/96 $32.00 + .OO
ELSEVIER PII SO278-!%46(96)00062-0
EFFECT OF ISOLATION ON MORPHINE- INDUCED RUNNING AND CHANGES IN BODY TEMPERATURE
JEc\N-PIE~ CXKIDEREAU, MARCEL DEBRAY t, CLAIRE MONIER, JEAN-MARIE BOHRRE AND HEWETTE FRANCES
JNSERM U26 - Hopital Fernand Widal - Paris, France. I Laboratoire de Biostatistiques, Fact&e de Pharmacie - Paris, France.
(Final form, March 1996)
Abstract
Coudereau, Jean-Pierre, Marcel Debray, Claire Monier, Jean-Marie Bourre and Henriette Frances: Effect of isolation on morphine-induced running and changes in body temperature. Prog. Neuro- Psychophamacol. & Biol. Psychiat. 1996,~. pp. 827-838
1, The influence of isolation of three durations 8, 15 and 30 days has been examined in mice on the effects of morphine on rectal temperature and on locomotor activity Isolated mice were compared to non isolated mice with the same age
2, Morphine (20 mg/kg ip) induced in mice an early hypothermia followed by a late hyperthermia The hypothermic effect was significantly reduced following isolation, but the duration of isolation (8, 15, 30 days) had no influence. Isolation did not modify the hyperthermic effect of morphine.
3 Morphine (40 mg/kg ip) induced in mice an increase in locomotor activity called “running”. The running activity was significantly increased following isolation. The duration of isolation (8. 15, 30 days) did not seem to influence this effect
4 These results show that isolation does not modify in the same way every effects of morphine, they suggest that isolation alters the mechanism involved in the running activity and in the hypothermic effect The nature of these mechanisms is discussed
Keywords : hyperkinesis, hyperthermia, hypothermia, isolation, morphine
Abbreviations : dopamine (DA), dihydroxyphenylacetic acid (DOPAC), 5-hydroxyindolacetic acid (SHIAA), homovanillic acid (HVA), 3-metoxytyramine (3-MT)
Introduction
Social isolation increases in rats the morphine-induced locomotor response ( Deroche et al. 1994 ), the propensy to self-administration of heroin ( Bozarth et al 1989 ) or to drink morphine ( Hadaway et al 1979, Alexander et al 1981 ) but reduces the sensitivity to the rewarding effect of heroin in the conditioned place preference model ( Schenk et al 1983 )
The effect of isolation on morphine-induced analgesia are not homogenous in isolated mice, the analgesic effect of morphine has been reported to be inexistent or greater than in non isolated mice , in
827

828 J.-P. Coudereau et aL
rats isolation has been reported to reduce the analgesic effect of morphine or to induce no change (Adler et al 1975 ; Kostovski et al., 1977 ; Panksepp et al., 1980 , Puglisi-Allegra and Ohverio 1983) The discrepancy between these results may arise from differences in the species, the duration of isolation and the experimental models. Isolation has also been reported to decrease the number of opioid receptors in rat brain (Schenk et al. 1982)
In the present study, the effect of isolation was studied on morphine-induced increase in locomotor activity and on morphine-induced changes in rectal temperature These effects were chosen because they are respectively representative of the stimulant (hyperkinesis, hyperthermia) and inhibitory (hypothermia) effects of morphine The stimulant effects of morphine do not give rise to tolerance after chronic administration and, even, may produce sensitization; on the contrary, the inhibitory effects give rise to tolerance after repeated injections. Using the same strain of mice with similar durations of isolation, the authors tried to determine whether isolation affects in the same way these two effects of morphine.
Materials and Methods
Animals
Male NMRI mice (4-5 weeks old at the beginning of the experiment), from CERI, Genest St Isle 53 940 (France) were either housed in groups of six in home cages of 30x20~10 cm or isolated in home cages of 24x10~8 cm Isolated mice were compared to non-isolated mice which were the same age and were maintened in the same conditions for the same duration. The maintenance and experimental rooms were maintened at 21+1”C with 12 hours light/dark schedule (light on from 8 a.m to 8 p.m) Food and water were freely available. There were three periods of isolation : 8, 15 and 30 days.
Morphine sulfate (Francopia, France) was dissolved in demineralized water. The volume of injection was 0 2 ml / 20 g body weight. Administrations were performed by the intraperitoneal route (i.p ).
Experimental Procedures
Assessement of Locomotor Activitv
The locomotor activity of each animal was measured individually in a photocell actimeter (APELEX). Briefly, the apparatus consisted of individual transparent covered plastic boxes (20.5 cm wide x 27 cm deep x 11 cm high) and locomotor activity was counted by two infrared photo-couplers mounted at a height of 0 8 cm and located, one in the middle of the width, the other in the middle of the length
Each animal was placed in the actimeter for 30 minutes to acclimatize the environment. Immediately after drug injection (morphine base = 40 mgikg), the animal was returned to the cage for measuring the running activity. The scores were noted every 15 minutes for 60 minutes

Morphine effects (temperature, runn~gl in isolated mice
Assessement of Rectal Temperature
The rectal temperature was measured with a thermoelectrical probe maintened at a constant depth and linked to a galvanometer. Each animal was placed in individual transparent cages (20 cm wide x 20 cm deep x 10 cm high) for 30minutes to acclimatize the environment Then, rectal temperature was measured at 30 min before and at different times ( 30, 60, 90. 120, 150, 180, 210. 240 minutes ) after injection of morphine (morphine base = 20mg/kg).
Mice were allotted to two groups in such a way that the mean rectal temperature before morphine administration was the same in each group one group received morphine, the other the vehicle
All mice were used only once Mice were killed using CO2 in a special container
Data Analvsis
Results were analyzed with a univariate (Fig. 1) and a multivariate (Fig 2) repeated measures analysis using the Huynh-Feldt probability adjustment (Morrisson, 1976). These analysis were followed by post- hoc tests of specific contrasts. The results of Fig. 3 were analyzed using an analysis of variance for repeated measures followed by the Bonferronni’s tests The area under the time-response curve (AUC) for each animal was calculated by curve fitting and integration procedures utilizing an IBM personal computer. Since the mice were handled 10 times until the end of the experiment, this may influence the rectal temperature Therefore, to take into account the effect of the repetitive handling, the following procedure was used to express the results for each non isolated control mouse, the basal temperature (time - 30 min) was subtracted from the temperature at times (t) Then, the mean (2s E.M.) temperature time curve for all non isolated control mice receiving vehicle was calculated. For each non isolated morphine treated mouse, the basal temperature (time - 30 min) was subtracted from the temperature at times (t). Then, the mean temperature-time curve of corresponding control mice was substracted from the temperature-time curve obtained for each mouse. Thus, the authors obtained an area under the curve (AUC) which was at first negative and thereafter positive The same procedure was used for isolated mice Results of Fig. 4 were analyzed using a two-way analysis of variance followed by the Bonferronni’s test.
Results were expressed with mean 2 standard error to the mean (S E M). There were 10 - 15 animals for each group.
Results
Locomotor Activity
The administration of morphine (40mg/kg) induced an increase of locomotor activity in mice (Fig 1) However, the locomotor activity of morphine-treated mice is qualitatively so different from that of control mice that some authors call it “running” (Kuchinsky and Hornikiewicz, 1974 , Brase et al , 1977). The present authors agree. Therefore, the activity of morphine treated mice was compared in isolated and non-isolated animals without subtracting the activity of control mice.
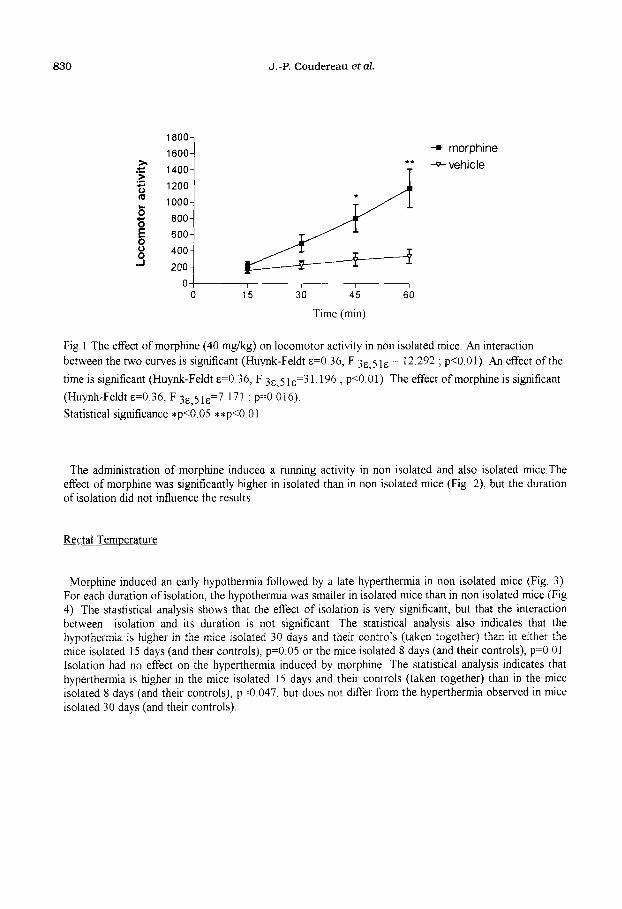
830 J.-P. Coudereau et aL
1800-
1600-
1400-
1200-
1 ooo-
800-
600-
400-
200-
- morphine
+ vehicle
01 / 0 15
I I I
30 45 60
Time (mm)
Fig. 1 The effect of morphine (40 mgIkg) on locomotor activity in non isolated mice. An interaction
between the two curves is significant (Huynk-Feldt E=O 36, F 3a,5 lE = 12.292 ; p<O.Ol). An effect ofthe
time is significant (Huynk-Feldt &=0.36, F 3s 5 ls=3 1.196 ; p<O 01). The effect of morphine is significant
(Huynh-Feldt E=O 36, F 3s,5 le37.171 ; p=O b16)
Statistical significance *p<O.O5 **p<O 01
The administration of morphine induced a running activity in non isolated and also isolated mice.The effect of morphine was significantly higher in isolated than in non isolated mice (Fig. 2) but the duration of isolation did not influence the results
Rectal Temperature
Morphine induced an early hypothermia followed by a late hyperthermia in non isolated mice (Fig 3) For each duration of isolation, the hypothermia was smaller in isolated mice than in non isolated mice (Fig 4) The stastistical analysis shows that the effect of isolation is very significant, but that the interaction between isolation and its duration is not significant The statistical analysis also indicates that the hypothermia is higher in the mice isolated 30 days and their controls (taken together) than in either the mice isolated 15 days (and their controls), p=O.O5 or the mice isolated 8 days (and their controls), p=O.Ol
Isolation had no effect on the hyperthermia induced by morphine The statistical analysis indicates that hyperthermia is higher in the mice isolated 15 days and their controls (taken together) than in the mice isolated 8 days (and their controls), p=O 047, but does not differ from the hyperthermia observed in mice isolated 30 days (and their controls).

Morphine effects (temperature, running) in isolated mice 831
1800- 1600- -+ isolated
2 1400- A +- non isolated :E *** 1200-
z m lOOO-
F BOO-
‘c
: 600-
w 400-
200-
0
&
0 30 45 60 Time (min)
1800- 1600
h 1400 B
.T 1200 l *
jlL$ ‘I;_
+- isolated
-+ non isolated
l
/ I I I 0 15 30 45 60
Time (min)
1800- 1600-
h 1400- 2 1200-
tj m lOOO-
F
‘E
3
600 Boo:
400
200
0 &
0 15 30 45 60 Time (min)
C
tt
- isolated
+-non isolated l **
Fig. 2 The effect of isolation (A 8 days, B. 15 days and C 30 days) on running activity induced by morphine (40 mgikg). The interaction (time of measurement of morphine effect 15-60 min) x (duration
of isolation 8, 15, 30 days) x (isolation, non isolation) is not significant (Huynh-Feldt &=0.466, F 6c, 162E
= 1 185 ; p=O 32) The effect of time of measurement of effect (15-60 min) is significant (Huynh-Feldt E
=0 466, F 3a,162c=208.27 ; p<O.Ol). The interaction (time of measurement of morphine effect: 15-60) x
(isolation, non isolation) is significant (Huynk-Feldt E=O 466, F 3c,t62c=32.64 ; pcO.01 The interaction
(time of measurement of morphine effect. 1 S-60) x (duration of isolation S, 15, 30 days) is not significant (Huynh-Feldt c=O 466, F 6c,t62c’O 636; p=O 58)
Statistical significance. *p<O.O5, **p<O.Ol, +**p<O 001
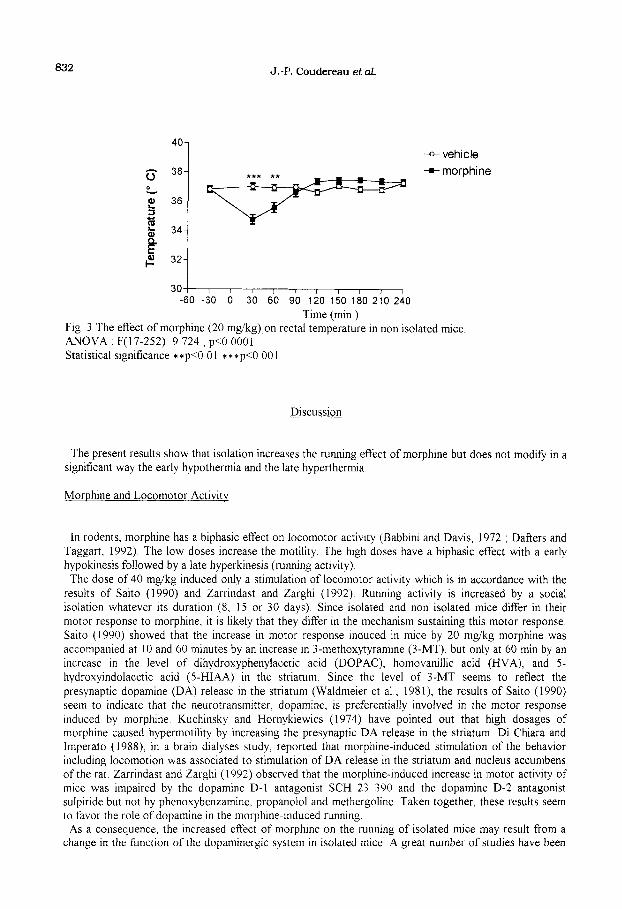
832 J.-P. Coudereau et&.
40
1 - vehicle
3a-
32-
-morphine
301 -60 -30 0 30 60 90 120 150 180 210 240
Time (min.) Fig 3 The effect of morphine (20 mg/kg) on rectal temperature in non isolated mice ANOVA. F(17-252~9 724, p<O 0001 Statistical significance **p<O 01 ***p<O.OOl
Discussion
The present results show that isolation increases the running effect of morphine but does not modify in a significant way the early hypothermia and the late hyperthermia
Morphine and Locomotor Activity
In rodents, morphine has a biphasic effect on locomotor activity (Babbini and Davis, 1972 ; Dafters and Taggart, 1992) The low doses increase the motility The high doses have a biphasic effect with a early hypokinesis followed by a late hyperkinesis (running activity)
The dose of 40 mg/kg induced only a stimulation of locomotor activity which is in accordance with the results of Saito (1990) and Zarrindast and Zarghi (1992) Running activity is increased by a social isolation whatever its duration (8, 15 or 30 days). Since isolated and non isolated mice differ in their motor response to morphine, it is likely that they differ in the mechanism sustaining this motor response. Saito (1990) showed that the increase in motor response induced in mice by 20 mg/kg morphine was accompanied at 10 and 60 minutes by an increase in 3-methoxytyramine (3-MT), but only at 60 min by an increase in the level of dihydroxyphenylacetic acid (DOPAC), homovanillic acid (HVA), and 5- hydroxyindolacetic acid (5-HIAA) in the striatum Since the level of 3-MT seems to reflect the presynaptic dopamine (DA) release in the striatum (Waldmeier et al, 198 1). the results of Saito (1990) seem to indicate that the neurotransmitter, dopamine, is preferentially involved in the motor response induced by morphine Kuchinsky and Hornykiewics (1974) have pointed out that high dosages of morphine caused hypermotility by increasing the presynaptic DA release in the striatum Di Chiara and Imperato (1988) in a brain dialyses study, reported that morphine-induced stimulation of the behavior including locomotion was associated to stimulation of DA release in the striatum and nucleus accumbens of the rat. Zarrindast and Zarghi (1992) observed that the morphine-induced increase in motor activity of mice was impaired by the dopamine D-l antagonist SCH 2 3 390 and the dopamine D-2 antagonist sulpiride but not by phenoxybenzamine, propanolol and methergoline Taken together, these results seem to favor the role of dopamine in the morphine-induced running
As a consequence, the increased effect of morphine on the running of isolated mice may result from a change in the function ofthe dopaminergic system in isolated mice A great number of studies have been

Morphine effects (temperature, running) in isolated mice
0 non isolated
EZ2J rsolated
Bypothermia Hyperthermia
300
250
200 i
0 non isolated
B ED isolated
T
Hypothermia Hyperthermia
300
C 0 non isolated 250 T EZ2 Isolated
833
Hypothermia Hyperthermia
Fig 4 The effect of isolation (A: 8 days, B: 15 days and C: 30 days) on changes in temperature induced by morphine (20 mg/kg). For hypothermia, a two way analysis of variance shows the followings. The interaction (isolation, non isolation)x(duration of isolation 8,15,30 days) is not significant F(2-84)=0 2 1 , p=O 811. The effect of isolation (isolation)x(non isolation) is very significant F( l-84)=8.46, p=O 0047 The duration of isolation (8,15,30 days) has a significant effect F(2-84)=5.35;p=O.O065 For hyperthermia, a two way analysis of variance shows the followings The interaction (isolation, non isolation)x(duration of isolation 8,15,30 days) is not significant F(2-84)=0 28 ; p=O 760. The effect of isolation (isolation)x(non isolation) is not significant F( l-84)=0.46 ; p=O 499. The duration of isolation (8,15,30 days) has a significant effect F(2-84)=3.22 ; p=O.O45

834 J.-P. Coudereau et al.
performed to tentatively associate social deprivation and DA The results are heterogeneous with, in isolated animals, similar cerebral levels of dopamine (Anisman and Sklar 198 1 , Nishikawa et al 1974) or increased levels of dopamine (Lasley and Thurmond 1985 , Welch and Welch 1968) The release of dopamine has been reported to be reduced (Andreas et al 1985 , Wutsman et al. 1987) or increased in the olfactive tubercules (Thoa et al 1977) of isolated animals The turn-over of dopamine after isolation has been described as increased (Lasley and Thurmond 1985 ; Modigh 1973 ; Valzelli 1978) or reduced (Thoa et al 1977 , Weinstock et al 1978) or unchanged (Anisman and Sklar 1981) These discordant results may be linked to global evaluations since the study of structures appears more homogeneous A 8 weeks isolation produces, in rats, a reduction in the activity of the mesocortico-frontal dopaminergic neurons as measured by the ratio DOPACIDA. whereas a rather enhanced activity of the dopaminergic neurons projecting to the nucleus accumbens or the striatum was observed (Blanc et al , 1980)
Stamford et al. (1991) showed that chronic exposure to mild unpredictable stress causes an increase in the release of DA in the nucleus accumbens of rats and since isolation is viewed as a stress, a similar release of DA may have occured in isolated mice
These data led to the suggested mechanism of action the running effect of morphine involves a dopaminergic mechanism and this effect is increased in isolated mice because isolation per se increases the DA function. This explanation, although likely, does not exclude other possibilities.
It has been shown that an opioid agonist of the delta receptors, administered in the nucleus accumbens induces a behavioral activation which is not blocked by the DA receptor antagonist thioproperazine (Dauge et al. 1989) ; this means that morphine may also stimulate behavior independently of DA.
This direct (not involving DA) action of morphine may have been altered in isolated mice since several authors have described an effect of isolation on the opiodergic system Iglesias et al (1992) have described an activation of the central proenkephalin opioid system in the hypothalamus following 14-20 days isolation Angulo et al (199 I) observed a reduction of 30% of the proenkephalin mRNA in caudate putamen and nucleus accumbens of rats isolated for 14 days
Alternatively, Deroche et al. (1994) have demonstrated that the increased locomotor response to morphine in isolated rats relative to control rats does not occur if corticosterone secretion is blocked by adrenalectomy. The social deprivation may, in fact, induce a series of events which may be independent or bound together, the blockade of one of them impairing the effect of isolation
Morphine and Rectal Temperature
The biphasic effect of morphine on rectal temperature (early hypothermia and late hypetthermia) was described by Gunne (1960) In the present experiments, the biphasic effect of morphine on rectal temperature was observed in isolated as well as in non isolated mice. In isolated mice, the hypothermic effect was signiticantlly smaller than in non isolated mice, but this effect was unrelated to the duration of isolation. It may be suggested that, if the effect of isolation on morphine hypothermia results from a stress, then the size of the stress is not different after these three durations of isolation The mechanism underlying this effect is not Kelly known. Morphine-induced hypothermia is antagonized
by naltrexone (Numan and Lal, 1981) and by naloxone (Geller et al., 1983). It was partially blocked by haloperidol (Glick, 1975). The hypothermia observed in mice isolated 30 days and their controls taken together was greater than for the other groups of mice (isolated 8 days and their controls or isolated 15 days and their controls). This may be linked to the age of the mice ; indeed, all mice were beginning the isolation period at the same age. So that after 30 days of isolation, the isolated mice were older than after 8 or 15 days of isolation. The same is true for control mice which were brought in the animal house concomitantly with the corresponding isolated mice
The hyperthermic effect was unchanged in isolated mice whatever the duration of isolation. The mechanism underlying this effect is not elucidated. The hyperthermia observed in mice isolated 15 days and their controls taken together was greater than in the group of mice isolated 8 days and their controls but did not differ from the hyperthermia observed in mice isolated 30 days and their contols. As for hypothermia, it may be suggested that this effect is related to the age of the mice. The hyperthermia is antagonized by naloxone (Geller et al. 1983 ; Vezina and Stewart, 1985) and by naltrexone (Numan and

Morphine effects (temperature. runningl in isolated mice 835
Lal, 1981) and attenuated by methergoline (Vezina and Stewart, 1985) and by physostigmine but not by neostigmine, two cholinesterase inhibitors the former (but not the latter) crossing the blood-brain barrier (Sharkawi, 1972). Morphine-induced hyperthermia is antagonized by neither of the following drugs pimozide, scopolamine, phenoxybenzamine (Vezina and Stewart, 1985). Thus, morphine-induced hyperthermia appears to depend on a central mechanism of action involving opioid receptors and modulated by the cholinergic and serotonergic systems. The lack of alteration of morphine-induced hyperthermia by isolation suggests that this form of stress does not modify the function of the receptors or of the neurochemical pathways induced in this effect.
Alternatively, an effect of isolation on the hyperthermia may have been hidden by the additive stress that represents the repetitive assessment of temperature (Zelman et al , 1985 ; Dallers and Taggart, 1992) in non isolated as well as in isolated mice since hyperthermia occurs after 90-120 minutes (or 5-6 measures)
Conclusion
The present data show that isolation does not modify in the same way three effects of morphine hypothermia, hyperthermia and running activity. Hyperthermia and hyperkinesis are stimulant effects of morphine which may be induced by administration in the same structure : the ventral tegmental area (Vezina and Stewart, 1985). Both hyperthermia and hyperkinesis give rise to sensitization after chronic treatment.
In spite of these similarities, the present results together with the data of Vezina and Stewart (1985) (antagonism by pimozide of the hyperkinesis but not of the hyperthermia) show that these two effects are not sustained by the same neuronal mechanisms and are not sensitive to the same environmental events.
Acknowledgements
This work was supported by the INSERM (Institut National de la Sante et de la Recherche Medicale) We thank FRANCOPIA (France) for its generous gift of morphine sulfate.
ADLER, M. W., MAURON, C., SAMANIN, R and VALZELLI, L (1975) Morphine analgesia in grouped and isolated rats. Psychopharmacologia 41 1 l- 14
ALEXANDER, B K., BEYERSTEIN, B. L., HADAWAY, P F and COAMBS, R. B. (1981) Effect of early and later colony housing on oral ingestion of morphine in rats Pharmacol Biochem Behav. fi 571-576
ANDREAS, K., DIENEL, A., FISHER, H D., OEHLER, J , WUSTMANN, C. and SCHMIDT, J (1985) The influence of social isolation on ethanol preference behavior and dopamine release in telencephalon slices in mice Pol J Pharmacol Pharm. 37 85 l-854
ANGULO, J.A., PRINTZ, D , LEDOUX, M. and McEWEN, B.S. (1991) Isolation stress increases tyrosine hydroxylase mRNA in the locus coeruleus and midbrain and decreases proenkephahn mRNA in the striatum and nucleus accumbens. Mol. Brain Res. n, 301-308.

836 J.-P. Coudereau et al.
ANISMAN, H and SKLAR, L. S. (1981) Social housing conditions influence escape deficits produced by uncontrollable stress : assessment of the contribution of norepinephrine. Behav. Neural. Biol 32 406-427
BABBINI, M. and DAVIS, W M (1972) Time-dose relationships for locomotor activity effects of morphine after acute or repeated treatment. Br J. Pharmacol. 46 213-224.
BLANC, G , HERVE, D , SIMON, H., LISOPRAWSKI, A, GLOWINSKI, J and TASSIN, J P (1980) Response to stress of mesocortico-frontal dopaminergic neurones in rats after long-term isolation Nature 284: 265-267.
BOZARTH, M A., MURRAY, A and WISE, R A. (1989) Influence of housing conditions on the acquisition of intravenous heroin and cocaine self-administration in rats Pharmacol Biochem. Behav ;1 903-907
BRASE, D. A., LOH, H. H and WAY, E. L. (1977) Comparison of the effects of morphine on locomotor activity, analgesia and primary and protracted physical dependence in six mouse strains J Pharmacol. exp. Therap. 201 368-374.
DAFTERS, R. and TAGGART, P (1992) Biotelemetric investigation of morphine’s thermic and kinetic effects in rats. Psychopharmacol 106 195-201
DAUGE, V., ROSSIGNOL, P. and ROQUES, B P. (1989) Blockade of dopamine receptors reverses the behavioral effects of endogenous enkephalins in the nucleus caudatus but not in the nucleus accumbens
Involvement of delta and mu opioid receptors Psychopharmacology 99 16% 175
DEROCHE, V , PIAZZA, P V , LE MOAL, M and SIMON, H (1994) Social isolation-induced enhancement of the psychomotor effects of morphine depends on corticosterone secretion Brain Res 040 136-139
DI CHIARA, G and IMPERATO, A (1988) Opposite effects of mu and kappa opiate agonists on dopamine release in the nucleus accumbens and in the dorsal caudate of freely moving rats J Pharmacol exp Ther. 244 1067-1080
GELLER, E B , HAWK, C , KEINATH, S.H., TALLARIDA, R J. and ADLER, M W (1983) Subclasses of opioids based on body temperature change in rats acute subcutaneous administration J Pharmacol exp Therap. 225 391-398
GLICK, S D (1975) Hyperthermic and hypothermic effects of morphine in mice Interactions with apomorphine and pilocarpine and changes in sensitivity after caudate nucleus lesions Arch int Pharmacodyn 213 264-271
GUNNE, L M (1960) The temperature response in rats during acute and chronic morphine administration a study of morphine tolerance Arch int pharmacodyn 12c, 4 16-428
HADAWAY, P. F , ALEXANDER, B. K , COAMBS, R.B and BEYERSTEIN, B (1979) The effect of housing and gender on preference for morphine-sucrose solutions in rats Psychopharmacol (Berlin) 66 87-91
IGLESIAS, T, MONTERO, S., OTERO, M J, PARRA, L and FUENTES, J A (1992) Preproenkephalin RNA increases in the hypothalamus of rats stressed by social deprivation Cell Molec Neurobiol 12 547-55s.

Morphine effects (temperature, runninp) in isolated mice 837
KOSTOWSKI, W., CZLONKOWSKI, A, REWERSKI, W. and PIECHOCKI, T (1977) Morphine action in grouped and isolated rats and mice Psychopharmacol s 19 I- 193
KUSCHINSKY, K. and HORNYKIEWICZ, 0 (1974) Effects of morphine on striatal dopamine metabolism : Possible mechanism of its opposite effect on locomotor activity in rats and mice Eur .I Pharmacol 26 41-50
LASLEY, S M and THLJRMOND, J. B (1985) Interaction ofdietary tryptophan and social isolation on territorial aggression, motor activity, and neurochemistry in mice Psychopharmacol Q 3 13-32 I
MODLGH, K. (1973) Effects of isolation and fighting in mice on the rate of synthesis of nor-adrenaline. dopamine and 5-hydroxytryptamine in the brain Psychopharmacologia (Berl ) ;3 1-l 7
MORRISSON D F (1976) Multivariate statistical methods, Chap 5. 236-247, ed MC Graw-Hill , New- York
NISHIKAWA, T , KAJIWARA, Y , KONO, Y . SANO, T and NAGASAKI, N (1974) Isolation- induced behavioural changes and brain monoamine levels in the rat Kurume Med 21 1 I7- 12 1
NUMAN, R and LAL, H (1981) Effect of morphine on rectal temperature after acute and chronic treatment in the rat Prog. Neuro-Psychopharmacol and Biol Psychiat 5 363-371
PANKSEPP, J, HERMAN, B H , VILBERG, T, BISHOP, P and DEESKINAZI. F G (1980) Endogenous opioids and social behavior Neurosci Biobehav Rev 4 473-487
PUGLISI-ALLEGRA, S and OLIVERIO, A (1983) Social isolation effects of pain threshold and stress-induced analgesia. Pharmacol Biochem Behav u 679-68 I
SAITO, H (1990) Inhibitory and stimulator-y effects of morphine on locomotor activity in mice biochemical and behavioral studies. Pharmacol. Biochem. Behav 35 23 l-235
SCHENK, S , BRITT, M D. and ATALAY, J. (1982) Isolation rearing decreases opiate receptor binding in rat brain Pharmacol. Biochem. Behav 16 841-842.
SCHENK, S , HUNT, T , COLLE, L. and AMIT. Z (1983) Isolation versus grouped housing in rats differential effects of low doses of heroin in the place preference paradigm. Life Sci 32 I 129-l 134
SHARKAWI, M (1972) Morphine hyperthermia in the rat its attenuation by physostigmine Br J Pharmac 44 544-548
STAMFORD, J A, MUSCAT, R., O’CONNOR, J J., PATEL, J., TROUT, S. J , WIECZOREK, W J KRUK, Z. L, and WILLNER, P (1991) Voltametric evidence that subsensitivity to reward following chronic mild stress is associated with increased release of mesolimbic dopamine Psychopharmacol IO5 275-282
THOA, N B , TIZABI, Y. and JACOBOWITZ, D. M. (1977) The effect of isolation on cathecholamine concentration and turnover in discrete areas of the rat brain. Brain Res 131 259-269.
VALZELLI, L (1978) Effect of socio-environmental isolation on brain biochemistry, behaviour and psycho-active drug activity Ann 1st Super Sanita. 14 173- I82
VEZINA, P and STEWART, J. (1985) Hyperthermia induced by morphine administration to the VT.4 of the rat brain an effect dissociable from morphine-induced reward and hyperactivity. Life Sci -33. 1095-l 105

838 J.-P. Coudereau et al.
WALDMEIER, P C , LAUBER, W , BLUM, W. and RICHTER, W J (1981) 3-Methoxytyramine Its suitability as an indicator of synaptic dopamine release Naunyn Schmiedebergs Arch Pharmacol 315 219-225
WEINSTOCK, M., SPEISER, Z and ASHKENAZI, R (1978) Changes in brain catecholamine turnover and receptor sensitivity induced by social deprivation in rats Psychopharmacology 56 205-209
WELCH, B. L. and WELCH, A S (1968) Greater lowering of brain and adrenal cathecholamines in group-housed than in individually-housed mice administered DL-alpha-methyltyrosine J Pharm Pharmaco 20 : 244-246
WUSTMANN, C, OEHLER, J, DIENEL, A, JAHKEL, M, FISCHER, H. D. and SCHMIDT, J (1987) The influence of social isolation, hypoxia and immobilization on potassium-stimulated dopamine release from telencephalon slices of mice Biomed. Biochem Acta 46 245-250
ZARRINDAST, M. R. and ZARGHI, A. (1992) Morphine stimulates locomotor activity by an indirect dopaminergic mechanism possible D-l and D-2 receptor involvement. Gen. Pharmac. 23 (N”6) 1221-1225
ZELMAN, D. C., TIFFANY, S. T. and BAKER, T. B (1985) Influence of stress on morphine-induced hyperthermia : relevance to drug conditioning and tolerance development Behav Neurosci 1 l22-
144
Inquiries and reprint requests should be addressed to
Dr J. P. Coudereau INSERM U26 Hopital Fernand Widal 75010 Paris FRANCE




![Regis Debray - Transmitir [Libro]](https://img.dokumen.tips/doc/110x75/557211ee497959fc0b8fbca3/regis-debray-transmitir-libro.jpg)
























