Embed Size (px)
Citation preview

Anita. Behav., 1976, 24, 626-636
M O U N T A I N G A Z E L L E A G O N I S T I C B E H A V I O U R
BY GERALD A. GRAU* & FRITZ R. WALTHER Department of Wildlife and Fisheries Sciences, Texas, A & M University, College Station, Texas 77843
Abstract. The behaviour of mountain gazelle (Gazella gazella) in Israel was investigated from 1970 to 1972. Most agonistic encounters occur between members of the same age, sex and social class. Females and immature males did not exhibit the full repertory of display patterns. The intensity of fighting gradually increases as the males mature. However, fights between territorial males are ritualized and not as violent as fights between adult bachelor males. Some encounters, especially among the immature males, take place without recognizable function and situational motivation, while others are linked with the co-ordination of group activity, enforcement of individual distance and territory establishment and maintenance. Horn contact occurs more frequently in the encounters of immature bachelor males than in those of adults or territorial males.
The function of intraspeeific aggression has been the subject of many behavioural studies. Several authors, for example Eibl-Eibesfeldt (1961), have stated that intraspecific fights rarely end in death or serious injury, but only in the removal of one individual from the immediate surroundings. Lorenz (1967) proposed dispersion as well as sexual selection and defence of young as additional functions. Walther (1974) included co-ordination of group activity plus establish- ment and maintenance of group organization. In some cases, elaborate social organization may not be possible without aggression (Lorenz 1967).
Within agonistic behaviour, one may distin- guish forms of aggression which involve physical contact, and threat or dominance displays in which the combatants do not touch one another. Sometimes the displays constitute the entire encounter. In 'reproductive fighting' (Tinbergen 1953) the breeding adults are of equal maturity and usually territorial rivals. Therefore, threats are assumed to arise from a conflict between fight and escape tendencies (Tinbergen 1952), and it is generally agreed that threat displays may often originate from intention movements for fighting (Tinbergen 1954; Eibl-Eibesfeldt 1970).
Although much has been written on intra- specific aggression, relatively little has been published on the distribution of aggression among sex, age and social classes. This paper discusses the ontogeny of agonistic behaviour patterns and their motivation and function in the social organization of the mountain gazelle (Gazella gazella). Mountain gazelle social organi-
*Present address: Ohio Cooperative Wildlife Research Unit, The Ohio State University, Columbus, Ohio 43210, U.S.A.
zation, consisting of female and young herds, bachelor male herds, and solitary territorial males (Grau 1974; Mendelssohn 1974), is very similar to the social organization of the East African gazelles (Walther 1964, 1965, 1968; Estes 1967).
Methods Field observations from August 1970 to June 1972 were conducted in two different biotypes chosen for their convenience for watching gazelles. The northern Negev area is located in the gently rolling loess soil of the Philistian plain (Zohary 1962) along the Gaza Strip about forty kilometres west of Beersheba. Gazelle density in this area was relatively low (two and one half per square kilometre) but observation was quite easy due to the relatively flat terrain. A second study area was located in Nahal Yissakhar, a deep, wide wadi crossing the southern part of Ramat Yissakhar, in the lower Galilee. Cattle graze in the wadi all year long and the surrounding area is cultivated. Gazelle density reached levels of thirty-one per square kilometre (Mendelssohn 1972), but observations were difficult due to the rough terrain. Observa- tion in the research zoo at Tel-Aviv University supplemented the field study.
The mountain gazelle in Israel have a larger flight distance from humans than from vehicles or semipermanent structures. Therefore the field observations were made from a Land Rover or camera blind. The quantity and sequence of the behaviour patterns were recorded by tape recorder, and documented by still and motion pictures and sketches.
Based on the presence and size of the horns (Plate IX, Fig. 1), males were separated into age classes (adult, sub-adult and adolescent) similar to
626

GRAU & WALTHER: GAZELLE AGONISTIC BEHAVIOUR 627
those described by Walther (1973). T h e adult males were also classified according to their social status (territorial or bachelor). The data from observations on adolescent and sub-adult males were pooled to form a larger immature class. Females were easily separated from the males by horn size and shape (Plate IX, Fig. 1), but due to the large amount of variation in horn sizes, no attempt was made to age the females in tile field.
Fourteen territorial males and six of the twenty-five bachelor males on the northern Negev areas were known by the size and shape of their horns plus the shape of their facial mark- ings. Photos of the face and horns from two angles were mounted in a field book to assist in this identification. In principle, it is possible to identify individual females by irregularities of their horn shape, but due to the large observa- tion distances this was not a feasible field technique. This study includes observations on approximately forty females and twenty calves in the northern Negev and fifty males (of various ages and social status), sixty females, and thirty calves in Nahal Yissakhav, none of which were recognized individually.
Since unequal amounts of time were spent observing the agonistic behaviour of each sex, age and social class, the totals for the classes cannot be directly compared. Instead, the relative frequency of occurrence within each class will be compared. The following analyses are based on a total of 512 agonistic encounters during which 1259 displays, of five forms, were recorded.
Results Threat and Dominance Displays
The threat and dominance (Imponierverhalten, Walther 1974) display patterns most frequent in agonistic encounters are direct-approach, head- turned-away, high-presentation of horns, medium-presentation of horns and chasing at gallop. Other patterns such as broadside display without simultaneous head-turned-away, sub- mission, horn-sweeping, weaving, linked unna- tion-defecation, mounting, foreleg kick, grazing and grooming occur irregularly.
Direct approach is simply the addressor moving toward the addressee. The approach may be fast or slow, frontal, lateral or toward the rear of the addressee.
In the head-turned-away posture, the gazelle holds the head above shoulder level, and pres- ents the horns while facing away from the
addressee (Plate X, Fig. 2(a), (b)). In intense dis- plays the addressor may tilt his horns or turn his neck directly toward the addressee while simul- tane ously facing away. The display is usually given in broadside position to an opponent several metres away and can be maintained for several minutes. In the broadside orientation a territorial male circles the bachelor opponents but is in parallel or reverse parallel orientation to coun- ter-displaying territorial neighbours. Occasion- ally individuals may also show the head-turned- away display while approaching or following the addressee.
In the high-presentation of horns (Walther 1958, and Plate X, Fig. 2(a)), the addressor holds his head above shoulder level with the chin tucked and the horns tilted forward to a vertical position. As with the head-turned-away display, high-presentation can be given at a relatively large distance, but it can also be given at close range (2m or less). Occasionally this posture may be combined with the broadside position or during 'pursuit marches' it may infrequently occur while the gazelle is pacing away from a displaying opponent. However, it is most com- mon and effective when the addressor is facing or directly approaching the addressee, as illustrated by an example from our records (12 May 1971) involving an adolescent male in submissive posture approaching a territorial male. The territorial male presented the horns high but did not directly face the adolescent male. The adolescent male continued to ap- proach and the territorial male turned toward him and faced him with the high-presenta- tion of horns. The adolescent male immediately fled a short distance.
In medium-presentation of horns (Plate X, Fig. 2c, d), the head is below shoulder level with the horns tilted forward of the vertical position, but not yet horizontal as in the low-presentation (Walther 1968). The display usually lasts for only a brief time, and ordinarily is given frontally when the opponents are at close range. As in high-presentation of horns, this pattern is most effective when the addressor is facing the addressee as illustrated by another observation on 12 May 1971. A female (performing medium- presentation of horns) stood in front of a resting young gazelle for a short time, but the latter did not move. The female then turned her head slightly so that she looked directly at this young gazelle. The young gazelle immediately got up and fled a short distance before lying down. The female then bedded down where the

628 A N I M A L B E H A V I O U R , 24, 3
Table I. Summary of Agonistic Pattern Characteristics
Addressor
Behaviour pattern Body orientation Head and horn position Locomotion
I. Direct approach Frontal Above shoulder level and Yes horns not vertical
2. Head-turned-away Above shoulder level and Yes, during approach, lateral facing away from addressee; and following displays horns may approach a vertical position
Above shoulder level ; horns vertical
3. High presentation
4. Medium-presentation
Frontal or lateral
Frontal or while withdrawing
During approach, pursuit, and infrequently also during withdrawal
Frontal and at short Below shoulder level; horns Usually not distance from vertical or tips forward addressee of vertical
young gazelle had been lying. The above four agonistic behaviour patterns are summarized in Table I.
Individuals frequently galloped during the encounters, but to make recordings easier, chasing at gallop was only recorded when the addressor tried to make contact with or change the direction of the addressee.
Submissive behaviour with the head lowered and horns not presented is similar to the corres- ponding behaviour in Grant 's gazelle, Gazella granti (Walther 1965), and dorcas gazelle, Gazella dorcas (Walther 1966). Occasionally an immature male assumes the submissive posture while approaching or fleeing from the territorial male. After flight, the submissive animal may lie down with neck and chin stretched forward on the ground. In this case the territorial male continues with the head- turned-away or high-presentation postures while circling the immature male until it flees once more. Females may also show submissive lying in response to sexual advances of territorial males. The males usually continue to approach in courtship display and attempt to sniff or nuzzle the female's genital region.
Horn-sweeping is found in situations where two adult (territorial or bachelor) males face each other in high-presentation. From this position, one lowers his head, sweeps the horns sideways while parallel to the ground, and then returns to the high,presentation in one con- tinuous motion. His opponent may respond with the same behaviour or remain in the original posture. In the research zoo, the horn-sweeping was observed in a male only 2 months old.
Aggression against an inanimate object, including weaving (Walther 1965), and linked urination-defecation precede or follow agonistic encounters involving bachelors or territorial males during periods of low excitement. Females were observed weaving on six occasions; three of these took place when a female was herded during a sexual encounter with a territorial male, one just after a rest period and the remain- ing two during grazing. In agonistic encounters of low intensity, gazelles of each class may also graze or groom, and bachelor males may sometimes mount or kick with the foreleg.
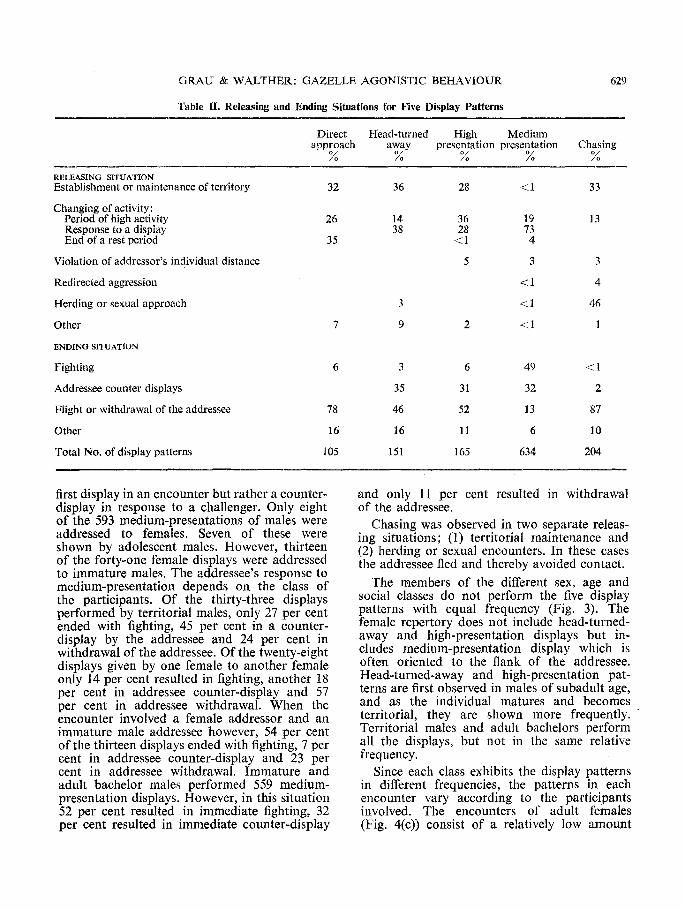
The possible motivation of direct-approach, head-turned-away, high-presentation, medium- presentation and chasing, may be surmised by examining the releasing situation and results for each individual pattern (Table II). Direct- approach, head-turned-away display and high- presentation of horns are found in two different situations: during changing of group activity and during territory establishment and main- tenance. Head-turned-away and high-presenta- tion displays, given only by males, are also frequently the response to a display by another male. All three displays often result in the with- drawal of the addressee, but about one-third of the head-turned-away and high-presentation displays lead to the addressee's counter-dis- playing. They are rarely followed by immediate fighting. Head-turned-away and high-presenta- tion are rarely addressed to females.
Medium-presentation of horns is rarely per- formed by a male toward a female. When addressed to another male it is usually not the
GRAU & WALTHER: GAZELLE AGONISTIC BEHAVIOUR
Table II. Releasing and Ending Situations for Five Display Patterns
629
Direct Head-turned High Medium approach away presentation presentation Chasing
% % % % %
RELEASING SITUATION Establishment or maintenance of territory
Changing of activity: Period of high activity Response to a display End of a rest period
Violation of addrcssor's individual distance
Redirected aggression
Herding or sexual approach
Other
ENDING SITUATION
Fighting
Addressee counter displays
Flight or withdrawal of the addressee
Other
Total No. of display patterns
32 36 28 <1 33
26 14 36 19 13 38 28 73
35 <1 4
5 3 3
<1 4
3 <1 46
7 9 2 <1 1
6 3 6 49 <1
35 31 32 2
78 46 52 13 87
16 16 11 6 10
105 151 165 634 204
first display in an encounter but rather a counter- display in response to a challenger. Only eight of the 593 medium-presentations of males were addressed to females. Seven of these were shown by adolescent males. However, thirteen of the forty-one female displays were addressed to immature males. The addressee's response to medium-presentation depends on the class of the participants. Of the thirty-three displays performed by territorial males, only 27 per cent ended with fighting, 45 per cent in a counter- display by the addressee and 24 per cent in withdrawal of the addressee. Of the twenty-eight displays given by one female to another female only 14 per cent resulted in fighting, another 18 per cent in addressee counter-display and 57 per cent in addressee withdrawal. When the encounter involved a female addressor and an immature male addressee however, 54 per cent of the thirteen displays ended with fighting, 7 per cent in addressee counter-display and 23 per cent in addressee withdrawal. Immature and adult bachelor males performed 559 medium- presentation displays. However, in this situation 52 per cent resulted in immediate fighting, 32 per cent resulted in immediate counter-display
and only 11 per cent resulted in withdrawal of the addressee.
Chasing was observed in two separate releas- ing situations; (1) territorial maintenance and (2) herding or sexual encounters. In these cases the addressee fled and thereby avoided contact.
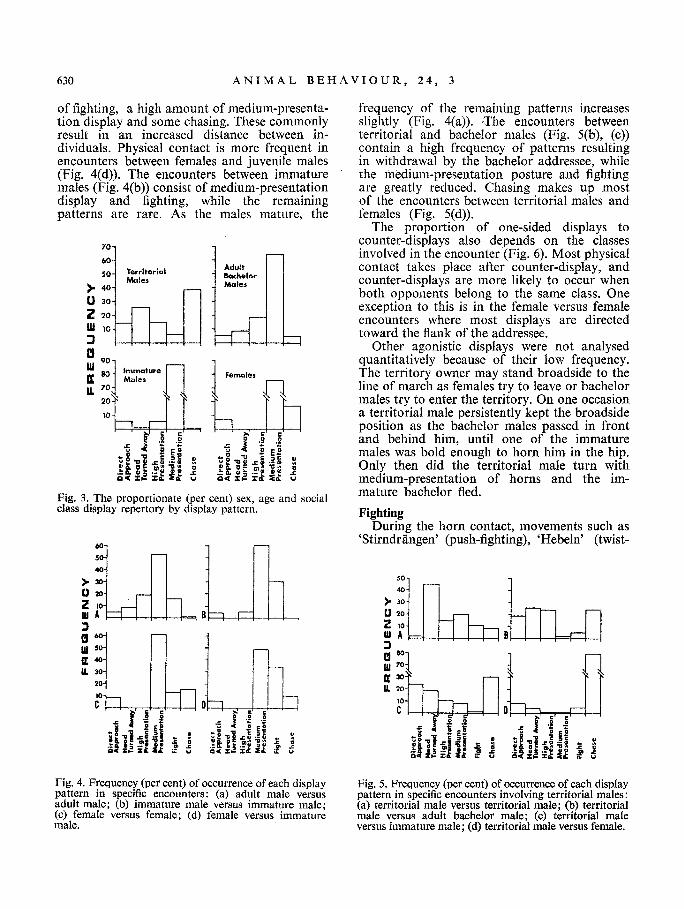
The members of the different sex, age and social classes do not perform the five display patterns with equal frequency (Fig. 3). The female repertory does not include head-turned- away and high-presentation displays but in- eludes medium-presentation display which is often oriented to the flank of the addressee. Head-turned-away and high-presentation pat- terns are first observed in males of subadult age, and as the individual matures and becomes territorial, they are shown more f r equen t ly . Territorial males and adult bachelors perform all the displays, but not in the same relative frequency.
Since each class exhibits the display patterns in different frequencies, the patterns in each encounter vary according to the participants involved. The encounters of adult females (Fig. 4(c)) consist of a relatively low amount
630 A N I M A L B E I - I A V I O U R , 24, 3
of fighting, a high amount of medium-presenta- tion display and some chasing. These commonly result in an increased distance between in- dividuals. Physical contact is more frequent in encounters between females and juvenile males (Fig. 4(d)). The encounters between immature males (Fig. 4(b)) consist of medium-presentation display and fighting, while the remaining patterns are rare. As the males mature, the
7 0 -
6 0 - $ 0 - Territorial
Nk~les ~, 40~
U ~ o ~ ~ 2 ~o~ W ~o~
Adult Bachelor I I
a~z~-ze~ v
Fig. 3. The proportionate (per cent) sex, age and social class display repertory by display pattern.
U Z I I �88
m , t z ~ - Z 't" lea" 1,7. v
frequency of the remaining patterns increases slightly (Fig. 4(a)). The encounters between territorial and bachelor males (Fig. 5(b), (c)) contain a high frequency of patterns resulting in withdrawal by the bachelor addressee, while the medium-presentation posture and fighting are greatly reduced. Chasing makes up most of the encounters between territorial males and females (Fig. 5(d)).
The proportion of one-sided displays to counter-displays also depends on the classes involved in the encounter (Fig. 6). Most physical contact takes place after counter-display, and counter-displays are more likely to occur when both opponents belong to the same class. One exception to this is in the female versus female encounters where most displays are directed toward the flank of the addressee.
Other agonistic displays were not analysed quantitatively because of their low frequency. The territory owner may stand broadside to the line of march as females try to leave or bachelor males try to enter the territory. On one occasion a territorial male persistently kept the broadside position as the bachelor males passed in front and behind him, until one of the immature males was bold enough to horn him in the hip. Only then did the territorial male turn with medium-presentation of horns and the im- mature bachelor fled.
Fighting During the horn contact, movements such as
'Stirndr/ingen' (push-fighting), 'Hebeln' (twist-
4 0
>. 3o U 20 Z i i i
0 a~ t W ro
I L 2 0
, . o
.-~g: e3 .-'~ _o~ m r Q• z~, z is ;tr .',- u
Fig. 4. Frequency (per cent) of occurrence of each display pattern in specific encounters: (a) adult male versus adult male; (b) immature male versus immature male; (e) female versus female; (d) female versus immature mate.
Fig. 5. Frequency (per cent) of occurrence of each display pattern in specific encounters involving territorial males: (a) territorial male versus territorial male; (b) territorial male versus adult bachelor male; (c) territorial male versus immature male; (d) territorial male versus female.
GRAU & WALTHER.: GAZELLE AGONISTIC BEI-[AVIOUR 63t
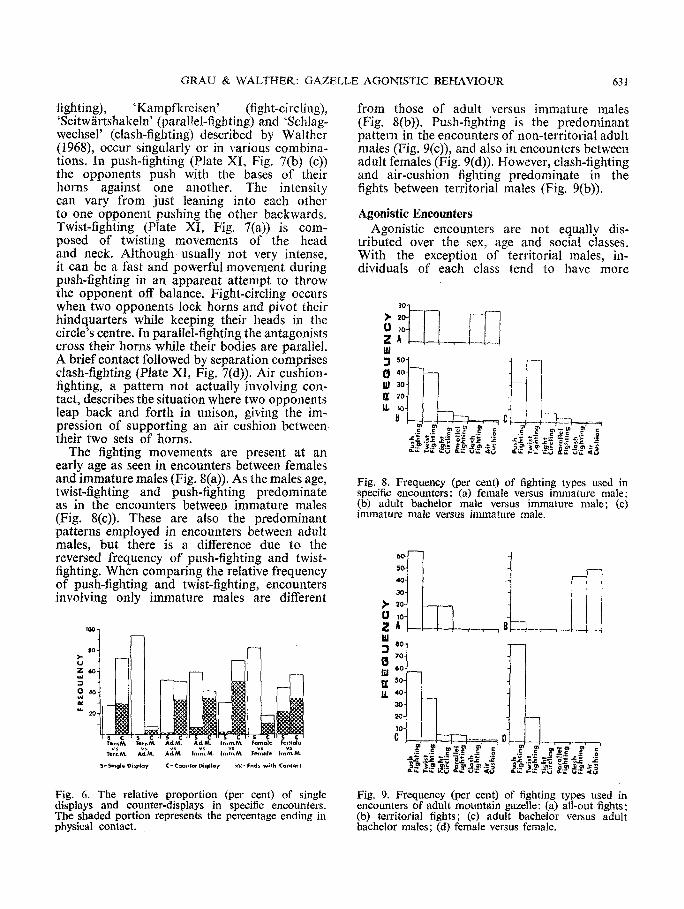
fighting), 'Kampfkreisen' (fight-circling), 'Seitw/irtshakeln' (parallel-fighting) and 'Schlag- wechsel' (clash-fighting) described by Walther (1968), occur singularly or in various combina- tions. In push-fighting (Plate XI, Fig. 7(b) (c)) the opponents push with the bases of their horns against one another. The intensity can vary from just leaning into each other to one opponent pushing the other backwards. Twist-fighting (Plate XI, Fig. 7(a)) is com- posed of twisting movements of the head and neck. Although usually not very intense, it can be a fast and powerful movement during push-fighting in an apparent attempt to throw the opponent off balance. Fight-circling occurs when two opponents lock horns and pivot their hindquarters while keeping their heads in the circle's centre. In parallel-fighting the antagonists cross their hon~s while their bodies are parallel. A brief contact followed by separation comprises clash-fighting (Plate XI, Fig. 7(d)). Air cushion- fighting, a pattern not actually involving con- tact, describes the situation where two opponents leap back and forth in unison, giving the im- pression of supporting an air cushion between their two sets of horns.
The fighting movements are present at an early age as seen in encounters between females and immature males (Fig. 8(a)). As the males age, twist-fighting and push-fighting predominate as m the encounters between immature males (Fig. 8(c)). These are also the predominant patterns employed in encom~ters between adult males, but there is a difference due to the reversed frequency of push-fighting and twist- fighting. When comparing the relative frequency of push-fighting and twist-fighting, encounters involving only immature males are different
Iit
Terr.M. Terr, M. A~M. Ad.M. I m a M , Female Ferule v s v s v s
$-Si~jle Display C-Cau~tar D~sl0lay ~'Ends wlth Co~tc~t
from those of adult versus immature males (Fig, 8(b)). Push-fighting is the predominant pattern in the encounters of non-territorial adult males (Fig. 9(c)), and also in encounters between adult females (Fig. 9(d)). However, clash-fighting and air-cushion fighting predominate in the fights between territorial males (Fig. 9(b)).
A g o n i s t i c E n c o u n t e r s
Agonistic encounters are not equally dis- tributed over the sex, age and social classes. With the exception of territorial males, in- dividuals of each class tend to have more
)- 2o U to Z A U/
0 4o W 3o
~o
Fig; 8, Frequency (per cent) of fighting types used in specific encounters: (a) female versus immature male; (b) adult bachelor male versus immature male; (c) immature male versus immature male.
u
Z
0
IT u.
2O
80- 70- 60- 50- ~ . 40 ~ ~ 2
20-1
c _ %~-y.
Fig. 6, The relative proportion (per cent) of single displays and counter-displays in specific encounters. The shaded portion represents the percentage ending in physical contact.
Fig. 9. Frequency (per cent) of fighting types used in encounters of adult mountain gazelle: (a) all-out fights; (b) territorial fights; (c) adult bachelor versus adult bachelor males; (d) female versus female.

632 ANIMAL B E H A V I O U R , 24~, 3
encounters within their own class than with members of other groups (Table III). More time was spent watching free ranging females than any other sex, age or social class but few encounters were recorded. Encounters among females in the research zoo were observed more frequently and these observations supplemented field data.
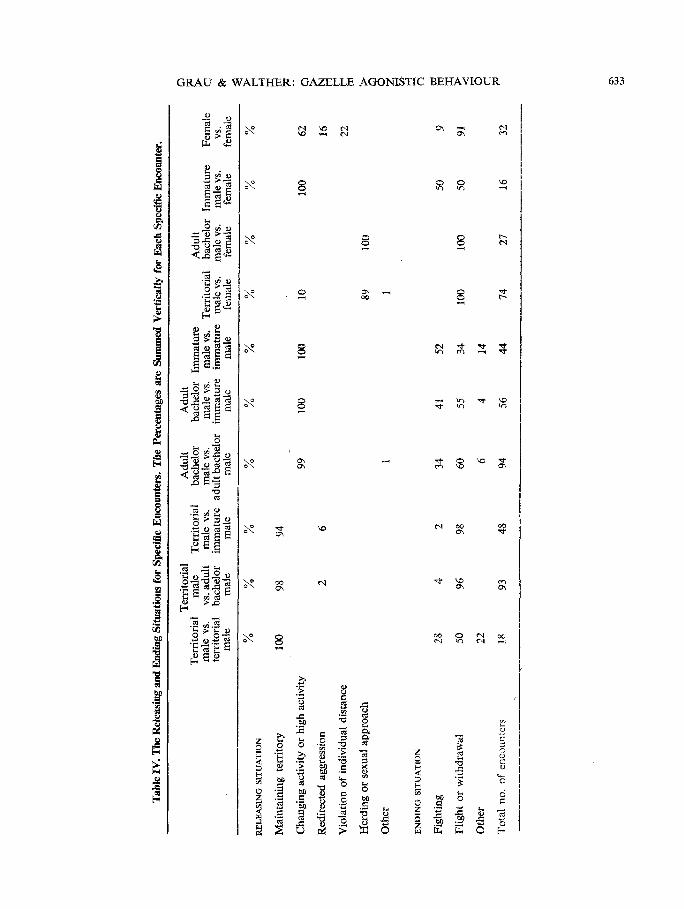
In most encounters initiated by females or bachelors, the releasing situation seems to be a change of activity (Table IV). However, in cases of female versus female, 38 per cent were due to violation of individual distance or redirected aggression ( 'Radfahrer-Reaktion' , Grzimek 1949) when one of the females was driven by a territorial male during a reproductive encounter. Sexual approaches or herding attempts by males preceded all of the encounters between females and territorial or non-territorial adult males. The establishment and maintenance of territories were the situations for encounters of territorial males versus neighbours or bachelor males.
The result of each encounter was classified as (1) fighting, (2) withdrawal by one or both individuals or (3) other endings (Table IV). In general, most encounters end with one or both individuals withdrawing before the en- counter reaches the fighting stage. However, encounters involving immature bachelor males more frequently end in fighting as compared to encounters between older males.
T y p e s o f Encounters
Encounters between territorial and bachelor males rarely end in fighting and are almost entirely composed of the territorial individual using the head-turned-away pattern while in broadside orientation. The bachelors slowly withdraw from this display, and the territorial males continue the head-turned-away while following.
On a few occasions we witnessed 'pursuit marches' involving adult males. During the march the pursuer in head-turned-away or high- presentation postures follows the addressee. When the latter stops, the addressor circles with head-turned-a~way display. Occasionally the males face each other in high-presentation display which is followed by horn-sweeping and possibly medium-presentation of the horns. In most instances the addressee withdraws before the encounter reaches the contact stages and the addressor continues following. One rather unusual encounter, with a bachelor male as addressor and a territorial male as addressee,
lasted from 06.53 hours until 15.42 hours. During this period ,,there were several periods of resting and grazing, but the actual pursuit took 5 hr and 25 min. This encounter ended in an all-out fight and the bachelor was defeated. The other 'pursuit-marches' were also of long duration, but unfortunately I did not know the individuals or observe the endings.
Encounters involving fighting can be classified into three distinct types: common fights, territorial fights and all-out fights. The common fights include encounters of bachelor males, female versus female, and female versus im- mature male. They can be serious, playful or combinations of the two. Common fights may be preceded by displays but consist mainly of physical contact. Such an encounter can last for several minutes, but the periods of contact are usually short with frequent pauses of varying lengths, during which the individuals graze, groom or stand. During these common fights the combatants usually remain within a small area.
In territorial fights there is a lengthy sequence of displays, starting with head-turned-away and gradually progressing into high-presentation of horns along with horn-sweeping, and finally into medium-presentation of horns and horn contact. The reverse of this sequence may take p lace as the opponents separate after a fight. However, the complete sequence before and after contact is not observed on occasions when
Table III. Frequency of Encounters Between Members of each Class. The Number of Encounters for Each Class is Given in the Addressor Column and the Percentages are
Summed to 100 Horizontally
Addressee
Adult Territorial bachelor Immature
Addressor male male male Female
% % % % Territorial male 8 40 21 31 N = 233
Adult bachelor 3 56 25 16 male N = 169
Immature male 9 17 64 10 N = 69
Female __ m 22 78 N = 41
Total no. of encounters = 512.
G R A U & WALTHER: GAZELLE AGONISTIC BEI-IAVIOUR 633
r ~
g
i
e~
~ o
: ~
~ ~.~
, .~ . ~
z ~ "r~ "~
~ ' ~ ~
I."5
�9 z e

634 A N I M A L B E H A V I O U R , 24, 3
the excitement rises or falls rapidly. Actual horn contact lasts for only 1 or 2 s and usually occurs only once per encounter. During the display, opponents may move over a considerable distance along the territorial boundary. Grazing, weaving and linked urination-defecation are exhibited frequently during the display.
All-out fights were observed only three times. One involved a territorial male and an adult male after a long pursuit march. The opponents in the other two encounters were unidentified adult males, and the activities preceding horn contact were not observed. In two of the all-out fights the periods of violent pushing and twisting lasted for 13 and 15 rain respectively and the third was at least 10 rain long. The fighting movements used are identical to those employed by other adult gazelle (Fig. 9(a)) and thus the all-out fights may be just a highly intense form of the common fight. However, they differ from common fights in that the horn contact is broken by several pauses of only a few seconds, and the encounter covers an area of at least 50 • 50 m. All three of these encounters ended with a chase at gallop of at least 500 m. Therefore, the all-out fight is the only type ending with one opponent completed defeated.
Discussion Mountain gazelle agonistic behaviour is closely associated with social organization. Most in- dividuals have encounters with other individuals of the same sex, age or social class, and members of the same class use similar displays and fight- ing movements. In cases where the herd is composed of one age class, there is no chance of encounter with individuals of other classes. However, bachelor herds of entirely one class are infrequent, and in herds containing males of several ages there is a marked tendency for the encounters to be among individuals of equal age. Encounters among individual.s of similar development are also known among other ungulates, for example Axis axis (Schaller 1967), Odocoileus virginianus (Michael 1968) and mountain sheep (Geist 1968a). As discussed for other species (Walther 1974), aggression contributes to the co-ordination of mountain gazelle group activities as the individuals display agonistic patterns causing other groups to rise or move. It also enforces the enlarged distances between individuals, especially among females, and functions in the establishment and main- tenance of territories. Bachelor males, especially the immature, may actually search for or invite
encounters from other males during the high activity periods. This searching for encounters supports the thesis o f spontaneous aggression (Lorenz 1967; Hinde 1970).
Mountain gazelle medium-presentation and high-presentation displays are similar to the comparative postures of other gazelle species (Walther 1968). The head-turned-away display of the mountain gazelle is similar to the early posture in the head-flagging display of Grant's gazelle (Walther 1965, Fig. 7(b); Walther 1968, Figs 25(b) and 26). Displays using postures similar to the head-turned-away may also occur in other species such as the Uganda-Kob, Kobus kob thomasi (Walther 1965, Fig. 11).
In the agonistic displays, the medium- presentation of horns is closely related to fighting behaviour since the individual can immediately perform fighting actions from this posture and 49 per cent are directly followed by fighting. High-presentation and head-turned- away do not indicate an intention for immediate attack, since very few lead directly to fighting and the posture must change to promote effective fighting position, which is in contrast to Gazella thomsoni where high-presentation frequently leads to fighting (Walther 1968). Only medium-presen- tation of horns, which usually results in immediate counter display or fighting, is used by immature males and females. Individuals in these classes fre- quently orient their displays toward the flank of the addressee. Adult territorial males show medium-presentation as well as head-turned- away display and high-presentation of horns. The latter two generally result in withdrawal of the addressee instead of fighting. All of these displays are oriented to the anterior of the opponent by the adult males. Thomson's gazelle lack the head-turned-away display, but the Grant's gazelle perform a somewhat similar display in the early stages of head-flagging. Walther (1965) found that 47 per cent of these head-flagging encounters end in withdrawal of one individual and only 11 per cent end in fighting. Older mountain gazelle males exhibit more displays, resulting in the withdrawal of the addressee, and fight less than the younger males. Males of other ungulates, for example Cervus duvauceli and Axis axis (Schaller 1967) and mountain sheep (Geist 1968b), also become more polished in social behaviour by increasing display patterns and reducing aggressive patterns as they mature.
All the gazelle fighting postures with the exception of 'H6rnerpressen' (horn pressing),

GRAU & WALTHER: GAZELLE AGONISTIC BEHAVIOUR 635
described by Walther (1968), were observed in the mountain gazelle. In contrast to display patterns all fighting movements are performed by females and immature males. As individuals mature they tend to perform some patterns more frequently than others. For example, push-fighting, the most intense form of horn contact, is used by all age groups, but pred- ominates the fighting when at least one adult individual is involved. Therefore the intensity of horn contact in the common fights increases directly with the age of the opponents. The greatest intensity occurs in all-out fights between adult males. Although fighting is most intense between adult males, relatively few encounters end in physical contact as compared to en- counters between immature males.
Mountain gazelle territorial encounters are infrequent and usually are settled by display. Fights between territorial males are rare and highly stereotyped. These fights of one or rarely two blows during clash-fighting are much shorter in duration than those of territorial Thomson's and Grant's gazelle males. As in the territorial fights of other gazelle species, there is no winner as such, but a mutual withdrawal from the area of physical contact. The mountan gazelle display after fighting is much shorter than the grazing ritual of territorial Thomson's gazelle males (Walther 1968) and apparently the separation of the two individuals is less difficult.
Certain authors have expressed the view that threat displays have evolved from conflicting tendencies to attack and to escape (Tinbergen 1952; Ewer 1968; Hinde 1970). Likewise, broad- side orientation is considered a result of the conflicting urges of fight and flight (Fraser 1957; Ewer 1968). Observations of threat behaviour in mountain gazelle do not support these theories of conflicting motivations. During encounters between territorial males and im- mature males there is no indication of escape tendencies recognizable in the territorial males. Still, the territorial males perform head-turned- away display while in broadside orientation instead of attacking. Furthermore, in the mountain gazelle, as well as many other bovid species (for example Antonius 1939; Geist 1964; Walther 1965, 1968; von Richter 1972), it is the dominant animal that uses the broadside orien- tation in encounters between unequal opponents. Subordinates rarely show it, making motivation by feat" unlikely.
In situations where aggression rises to an unusually high level, the lack of certain display
patterns observed at lower levels of excitement can be expected. However, this does not neces- sarily mean that occurrence of these displays depends on a conflict of motivations. Threat and fight undoubtedly belong to aggression as a major motivation or drive, and threat seems to correspond to a lower level of aggressive excite- ment than does intense fighting. Therefore, the occurrence of threat display instead of fighting or before fighting can be explained by the activation of a lower level of aggression rather than by a conflict in motivation. In other words, the (aggressive) excitement is simply not strong enough to release fighting immediately.
Apparently, other authors have come to similar conclusions. For example, by electrical stimulation of the cat brain, Brown & Hunsperger (1963, 444 to 445) found definite areas where threat behaviour could be eon- starttly elicited. 'This fact appears to contradict the theory that threat behaviour results from simultaneous activation and conflict of neural mechanisms for attack and escape, since only one area needs to be directly stimulated, not two.' They were unsuccessful in getting mutual inhibition between threat-attack and escape zones. However, they did find that different types of threat and their temporal relationships with attack and escape can be correlated with spatial patterns of central nervous activity.
Acknowledgments This is Texas Agricultural Experiment Station Technical Article No. 10972. The study was supported by Smithsonian Institution grants SFG-5181 and SFG-1-7066, and by the Caesar Kleberg Research Program in Wildlife Ecology.
We are grateful to Professor H. Mendelssohn (Tel-Aviv), the personnel of Tel-Aviv University and the members of Kibutz Nir Oz whose help and co-operation were essential for the study. We would like to thank Dr David Schmidly, Mr Wayne Faatz, Ms Rebecca Moore (College Station) and Mrs Brenda Grau (Colombus) for reading and correcting the manuscript.
REFERENCES Antonius, O. (1939). t)ber Symbolhandlungen und
Verwandtes bei Sfiugetieren. Z. Tierpsychol., 3, 263-278.
Brown, J. L. & Hunsperger, R. W. (1963). Nuerothology (sic) and the motivation of agonistic behaviour. Anita. Behav., 11, 439-448.
Eibl-Eibesfeldt, I. (1961). The fighting behaviour of animals. Scient. Am., 205, 112-122.
Eibl-Eibesfeldt, I. (1970). Ethology: The Biology of Behavior. New York: Holt, Rinehart & Winston.

636 A N I M A L B E H A V I O U R , 2 4 , 3
Estes, R. D. (1967). The comparative behavior of Grant's �9 and Thomson's gazelles. J. Mammal., 48, 189-209.
Ewer, R. F. (1968). Ethology of Mammals. New York: Plenum Press.
Fraser, A. F. (1957). The state of fight or flight in the bull. Brit. J. Anita. Behav., 5, 48--49.
Geist, V. (1964). On the rutting behavior of the mountain goat. Y. Mammal., 45, 551-568.
Geist, V. (1968a). On the interrelation of external appearance, social behaviour and social structure of mountain sheep. Z. Tierpsychol., 25, 199-215.
Geist, V. (1968b). On delayed social and physical matura- tion in mountain sheep. Can. J. ZooL, 46 (5), 889-904.
Grau, G. A. (1974). Behavior of mountain gazelle in Israel. Ph.D. Dissertation. College Station: Texas A & M University.
Grzimek, B. (1949). Die 'Radfahrer-Reaktion'. Z. TierpsychoL, 6, 41-44.
Hinde, R. A. (1970). Animal Behavionr; A Synthesis of Ethology attd Comparative Psychology. New York: McGraw-Hill, 2nd Edn.
Lorenz, K. (1967). On Aggression. New York: Bantam Books, Inc.
Mendelssohn, H. (1972). On the biology and ecology of gazelles in Israel. Final report on work carried out under Smithsonian grants SFG-0-5181 and SFG-I-7066. 53 pp. Xerox.
Mendelssohn, H. (1974). The development of the popula- tions of gazelles in Israel and their behavioural adaptations. In: The Behaviour of Ungulates and Its Relation to Management (Ed. by V. Geist & F. Walther), pp. 722-743. Morges, Switzerland: IUCN Publications new series, No. 24.
Michael, E. D. (1968). Aggressive behavior of white- tailed deer. Southwest. Nat., 13(4), 411-420.
Schaller, G. B. (1967). The Deer and the Tiger; A Study of Wildife in India. Chicago: University of Chicago Press.
Tinbergen, N. (1952). 'Derived' activities; their causation, biological significance, origin, and emancipation during evolution. Q. Rev. Biol., 27, 1-32.
Tinbergen, N. (1953). Social Behaviour in Animals, With Special Reference to Vertebrates. London: Methuen.
Tinbergen, N. (1954). The origin and evolution of courtship and threat display. In: Evolution As a Process (Ed. by J. Huxley, A. C. Hardy & E. B. Ford), pp. 233-250. London: Allen & Unwin.
von Richter, W. (1972). Territorial behaviour of the black wildebeest Connochaetes gnou. Zoologica Afrieana, 7, 207-231.
Walther, F. R. (1958). Zum Kampf- und Paarungsver- halten einiger Antilopen. Z. Tie~psychol., 15, 340-380.
Walther, F. R. (1964). Einige Verhaltensbeobachtungen an Thomsongazellen (Gazella thomsoni Guntehr, 1884) im Ngorongoro--Krater. Z. Tierpsyehol., 21, 871-890.
Walther, F. R. (1965). Verhaltensstudien an der Grant- gazelle (Gazella granti Brooke, 1872) im Ngoron- goro--Krater. Z. TierpsychoL, 22, 167-208.
Waltber, F. R. (1966). Von Waffen und K~impfen bei Horntieren. Kosmos, 62, 208-214.
Walther, F. R. (1968). Verhalten der Gazellen. Wittenberg- Lutherstadt: A. Ziemsen Yerlag.
Walther, F. R. (1973). On age class recognition and individual identification of Thomson's gazelle in the field. J. South Aft. Wildl. Mgmt. Ass., 2, 9-15.
Walther, F. R. (1974). Some reflections on expressive behaviour in combats and courtship of certain horned ungulates. In: The Behaviour of Ungulates and Its Relation to Management (Ed. by V. Geist & F. Walther), pp. 56-106. Morges, Switzerland: IUCN Publications new series, No. 24.
Zohary, M. (1962). Plant Life of Palestine. New York: Ronald Press.
(Received 22 May 1974; revised 10 January 1975; MS. number: A1568)





























