Embed Size (px)
Citation preview

C. V. WardDepartments of Anthropologyand Pathology andAnatomical Sciences,107 Swallow Hall,University of Missouri,Columbia, Missouri 65211,U.S.A. E-mail:[email protected]
M. G. LeakeyDivision of Palaeontology,National Museums of Kenya,P.O. Box 40658, Nairobi,Kenya. E-mail:[email protected]
A. WalkerDepartments ofAnthropology and Biology,Pennsylvania StateUniversity, University Park,Pennsylvania 16802, U.S.A.E-mail: [email protected]
Received 6 March 2001Revision received25 June 2001 andaccepted 27 June 2001
Keywords: Australopithecusanamensis, Kanapoi,Allia Bay, description,morphology.
Morphology of Australopithecus anamensisfrom Kanapoi and Allia Bay, Kenya
The hominid species Australopithecus anamensis was originallydescribed in 1995, with new specimens and more secure dates givenin 1998. This paper lists all fossils attributed to A. anamensis, andprovides anatomical descriptions of those not yet described in detailwith photographs of all but undiagnostic fragments. We also providecomparative analysis of these specimens. The A. anamensis holotypemandible was found at Kanapoi, as were most of the paratypes. TheAllia Bay sample is less well represented, and does not preserve manyanatomical elements diagnostic of this species. Still, the Allia Baysample most closely resembles that from Kanapoi, and we suggestthat for the time being it be retained as A. anamensis. A. anamensismost closely resembles A. afarensis, but can be distinguished from itin many features. Most of these features are inferred to be primitivefor the genus. Based on the limited postcranial evidence available,A. anamensis appears to have been habitually bipedal, although itretained some primitive features of its upper limbs. A. anamensisdiffers from A. afarensis in having narrower, more parallel jaws with avery slightly more ape-like canine/premolar complex than is found inA. afarensis, although not as ape-like as in Ardipithecus ramidus. It hadslightly larger lower lateral incisors, a unique upper canine mor-phology, and a different structure of the lateral nasal aperture than A.afarensis. A. anamensis had at least as great a range of body size, andperhaps slightly greater canine dimorphism, although this is difficultto determine. At present, there appears to be no autapomorphiesprecluding A. anamensis from ancestry of A. afarensis.
� 2001 Academic Press
Journal of Human Evolution (2001) 41, 255–368doi:10.1006/jhev.2001.0507Available online at http://www.idealibrary.com on
Introduction
The holotype and paratype series ofAustralopithecus anamensis were found atKanapoi, Kenya. Other fossils attributed toA. anamensis have been found at Allia Bay,Kenya. This species was first described inLeakey et al. (1995). Additional specimensand more secure dates were given threeyears later (Leakey et al., 1998).
All fossils attributed to A. anamensis dateto between 3·9 and 4·2 Ma. The older of thetwo sites is the type site of Kanapoi. Withthe exception of one specimen, a mandible(KNM-KP 29287), all Kanapoi hominins
0047–2484/01/100255+114$35.00/0
come from strata dated to between4·17�0·03 and 4·07�0·03 Ma (Leakeyet al., 1998). The mandible KNM-KP29287 derives from the paleosol developedon the upper tuff, and therefore cannot bemuch younger.
The Allia Bay fossils are slightly youngerthan those from Kanapoi. At Allia Bay, mostof the hominids were found at the 261-1site. This site lies just below the MoitiTuff, dated elsewhere in the region to3·94�0·03 Ma (Leakey et al., 1995). Whiteet al. (1993) report an age of 3·89�0·02 fortuff VT-1 in the Maka area of Ethiopia, atuff that on chemical grounds is regarded as
� 2001 Academic Press

256 . . ET AL.
Specimens attributed to Australopithecus anamensis
Specimen number Figure Element
KanapoiKNM-KP 271 13 L distal humerusKNM-KP 29281 1, 2 Holotype mandible & L temporal fragmentKNM-KP 29282 — LM1 or M2
KNM-KP 29283 3 MaxillaKNM-KP 29284 4 RC & RP3 germsKNM-KP 29285 14, 15 R proximal & distal tibiaKNM-KP 29286 5 Mandible fragments & associated dentition
(RI1, L & RI2-M3)KNM-KP 29287 6 Mandible with teethKNM-KP 30498 7 L & R maxillary fragments & associated dentitionKNM-KP 30500 8 Mandibular fragments & associated dentitionKNM-KP 30502 4 Associated mandibular tooth fragmentsKNM-KP 30503 16 Proximal manual phalanxKNM-KP 30505 4 Partial M germKNM-KP 30942 — 5 tooth fragmentsKNM-KP 31712 9 Associated juvenile mandibular & dental fragmentsKNM-KP 31713 10 R mandible with tooth fragmentsKNM-KP 31714 4 Ldm2
KNM-KP 31715 — LM1 or M2 fragment & two other tooth fragmentsKNM-KP 31716 — P3 or P4 fragment & C/ fragmentsKNM-KP 31717 4 LM3, RM3 & LM2 fragmentsKNM-KP 31718 — R Mandibular fragment (M2–3)KNM-KP 31719 — I1
KNM-KP 31720 — Maxillary M fragmentKNM-KP 31721 — RM2 & M3 partial crownsKNM-KP 31723 4 RM3
KNM-KP 31724 17 L capitateKNM-KP 31726 4 RP4
KNM-KP 31727 — RC
KNM-KP 31728 4 LM1
KNM-KP 31729 4 Rdm2
KNM-KP 31730 4 LM2 & RP3
KNM-KP 31732 — Tooth fragmentsKNM-KP 34725 11 Associated juvenile dentition and skull fragmentsKNM-KP 35838 — LM3
KNM KP 35839 12 Associated LI1, RC & LP3
KNM-KP 35840 — LM3 & maxillary M fragmentsKNM-KP 35841 — M crownKNM-KP 35842 4 R maxillary MKNM-KP 35844 — M fragmentKNM-KP 35845 — M fragmentKNM-KP 35847 4 LM2
KNM-KP 35850 — Maxillary M fragmentKNM-KP 35851 — LM2 or M3 fragmentKNM-KP 35852 4 LC
KNM-KP 37522 — L mandibular molarKNM-KP 37523 — M fragmentKNM-KP 37524 — Tooth fragments
Allia BayKNM-ER 7727 — LM2
KNM-ER 20419 22 L radiusKNM-ER 20420 — LM2
KNM-ER 20421 — RM3
Table 1

257AUSTRALOPITHECUS ANAMENSIS
Continued
Specimen number Figure Element
Allia Bay (Continued)KNM-ER 20422 — LM1
KNM-ER 20423 — LM2
KNM-ER 20427 — LM1
KNM-ER 20428 — LM3
KNM-ER 20432 — L mandibular fragment (P3–4)KNM-ER 22683 — LP4
KNM-ER 24148 — Ldm2
KNM-ER 30200 18 L maxillary fragment (M1–2, partial M3)KNM-ER 30201 19 LM1
KNM-ER 30202 19 RIl
KNM-ER 30731 19 RC
KNM-ER 30744 — RC
KNM-ER 30745 20 L maxillary fragment (partial C, P3–M1, partial M2, M3)KNM-ER 30747 — LP4
KNM-ER 30748 — L maxillary M fragmentKNM-ER 30749 19 LM1
KNM-ER 30750 — RC
KNM-ER 35228 21 RP4
KNM-ER 35229 — L mandibular M fragmentKNM-ER 35230 — M fragmentKNM-ER 35231 19 RM1 or M2
KNM-ER 35232 19 LM1
KNM-ER 35233 19 LM2
KNM-ER 25234 — LP3
KNM-ER 35235 19 LM2
KNM-ER 35236 19 LM3
KNM-ER 35238 19 RM1
Table 1
a correlative of the Moiti Tuff. The ageof the bone bed bearing the majority ofhominin fossils can be extrapolated tohave been 3·95�0·05 Ma (C. S. Feibel,personal communication). KNM-ER30200, KNM-ER 30201, KNM-ER 30744were found about 1 km to the east of the261-1 site in a small drainage channel thatcut through the Moiti Tuff. The strata atSibilot, near site 261-1, yielded a homininradius attributed on the basis of age toA. anamensis, and is estimated to be3·9�0·1 Ma (Heinrich et al., 1993).
The paleoenvironmental reconstructionsfor Kanapoi and Allia Bay have been men-tioned in Coffing et al. (1994), Leakey et al.(1995), Ward et al. (1999b), and Wynn(2000). As far as can be determined, thehabitats sampled at these sites were similarto those described for A. afarensis at Hadar,
Ethiopia, and Laetoli, Tanzania; fairlywooded regions that included woodland andbushland, sometimes with edaphic grasslandand often well watered (Reed, 1997). Theydiffer from the more closed woodland habi-tats reconstructed for the earlier Ardipithecusramidus site of Aramis, Ethiopia (Wolde-Gabriel et al., 1994). In one respect, Wynn’s(2000) conclusion, based on paleosols andtheir carbon isotopes, is slightly at variancewith the australopithecine habitats inter-preted by Reed (1997). The hominin fossilsat Kanapoi are associated with soils charac-teristic of semi-arid vegetational mosaics.Wynn (2000) suggests that this indicatesthat the hominins thrived in mosaic settings,whereas Reed (1997) concludes that ‘‘theenvironment in which these Australopithecusspp. existed was fairly static’’ and ‘‘con-strained by minimum and maximum

258 . . ET AL.
Description of Fossils
Each fossil is described first without specificcomparative references to provide an inven-tory of the preservation and morphology ofeach. Descriptions are provided by site,separated into craniodental and postcranialsections. Specimens are described innumeric order within these sections. Allspecimens that have been attributed to A.anamensis are listed by anatomical elementin Table 1.
Measurements are given in millimetersunless otherwise indicated. Standard metricdata are listed in Tables 2, 3 & 4, and otherdata are included in the text. Dentalmeasurements we previously reported(Leakey et al., 1995; Leakey et al., 1998)were taken following Wood (1991). Dentaldata published for A. afarensis, however,were taken using different techniques(White, 1977) that yield slightly differ-ent results. To permit metric comparisonwith other East African Australopithecusspecimens, tooth measurements reported(Table 2) in this paper were taken followingWhite (1977). These data should be con-sidered the definitive metric sample forcomparative purposes.
Anatomical terminology follows Johansonet al. (1982). Abbreviations used in thetext and tables are as follows (most followJohanson et al., 1982):
AP= anteroposterior/lyML= mediolateral/ly
SI= superoinferior/lyMD= mesiodistal/lyBL= buccolingual/ly
LaL= labiolingual/lyPD= proximodistal/lyDP= dorsopalmar/lyPrd= protoconid
Med= metaconidHyd= hypoconidEnd= entoconidHld= hypoconulid
Pa= paraconePr= protocone
Me= metaconeHy= hypoconeFa= anterior foveaFc= central foveaFp= posterior foveaC6= tuberculum sextum
Mmr= mesial marginal ridgeDmr= distal marginal ridgeMlg= median longitudinal grooveCo= crista obliqua
IPF= interproximal wear facetL= leftR= right
Kanapoi
At Kanapoi, specimens that were found inclose proximity and that appeared to repre-sent a single individual were given oneaccession number. Many of these have morethan one field number. Other specimensthat share a field number may representmore than one individual and therefore havedifferent accesssion numbers. If fragmentsthat make up a specimen cannot be certainlyassociated, different accession numbers havebeen assigned. In cases where more than one
amounts of rainfall and tree cover.’’ Wecannot be absolutely sure of the specifichabitats favored by the Kanapoi hominins,however, because most of the hominin fos-sils show signs of carnivore damage, andthere might be a depositional bias caused bythe behavioral proclivities of the predatorsand/or scavengers, rather than of thehominins.
Preliminary reports of the A. anamensisfossils are found in the original publications,and Leakey & Walker (1997) and Wardet al. (1999b). This paper describes themorphology of these fossils in detail, putsthe collection in comparative context, andmakes some functional and evolutionaryinterpretations.

259AUSTRALOPITHECUS ANAMENSIS
Tab
le2
Den
tal
met
rics
for
Au
stra
lop
ith
ecu
san
amen
sis
Spe
cim
enS
ide
I1C
P3
P4
M1
M2
M3
MD
LaL
MD
LaL
MD
BL
MD
BL
MD
BL
ant
BL
post
MD
BL
ant
BL
post
MD
BL
Max
illar
ype
rman
ent
dent
itio
nK
NM
-ER
7727
L12
·314
·714
·3K
NM
-ER
2042
0R
16·3
KN
M-E
R20
421
R11
·213
·7K
NM
-ER
2042
2L
11·9
10·9
KN
M-E
R20
428
L15
·713
·7K
NM
-ER
3020
0R
10·3
11·7
11·3
11·4
13·2
11·6
KN
M-E
R30
202
R10
·5a
9·3
KN
M-E
R30
744
L(7
·8)
(9·0
)K
NM
-ER
3074
5L
(11·
0)9·
913
·08·
813
·912
·214
·113
·9(1
3·2)
12·7
(14·
5)K
NM
-ER
3523
1R
11·7
12·5
11·9
KN
M-E
R35
232
L12
·9(1
2·2)
KN
M-E
R35
235
R(1
2·9)
KN
M-E
R35
236
L11
·513
·0K
NM
-ER
3523
8R
(10·
0)12
·812
·3K
NM
-KP
2928
3L
(9·3
)12
·17·
5[7
·8]
(12·
8)12
·914
·713
·313
·014
·7K
NM
-KP
2928
3R
7·9
[8·1
]11
·3[1
1·9]
13·2
12·2
13·2
14·8
13·3
KN
M-K
P30
498
L10
·98·
2K
NM
-KP
3049
8R
(10·
6)10
·28·
713
·211
·613
·912
·611
·6K
NM
-KP
3171
7L
11·1
13·8
KN
M-K
P31
723
R(1
2·7)
(15·
7)K
NM
-KP
3172
6L
7·2
KN
M-K
P34
725
R14
·214
·9K
NM
-KP
3583
9L
12·4
8·9
11·8
KN
M-K
P35
839
R11
·711
·2K
NM
-KP
3584
2R
11·6
KN
M-K
P35
852
L(1
1·0)
(10·
1)

260 . . ET AL.
Tab
le2
Con
tin
ued
Spe
cim
enS
ide
I 1I 2
CP
3P
4M
1M
2M
3
MD
LaL
MD
LaL
MD
LaL
MA
XM
INM
DB
LM
DB
LM
DB
LM
DB
L
Man
dibu
lar
perm
anen
tde
ntit
ion
KN
M-E
R20
423
L12
·3K
NM
-ER
2043
2L
12·0
8·6
9·7
11·9
KN
M-E
R22
683
L8·
7[9
·2]
10·4
KN
M-E
R30
201
L11
·610
·2K
NM
-ER
3073
1R
8·3
10·6
KN
M-E
R30
750
R6·
69·
2K
NM
-ER
3522
8R
7·4
9·6
KN
M-E
R35
233
L14
·813
·313
·0K
NM
-KP
2928
1L
5·9
[6·6
]7·
56·
5[7
·5]
8·6
8·8
9·6
11·7
8·5
7·9
[8·2
]9·
811
·9[1
2·7]
11·9
13·4
[13·
7]12
·614
·412
·2K
NM
-KP
2928
1R
6·1
[6·9
]7·
36·
9[7
·8]
8·6
8·7
9·4
11·5
8·7
7·8
[8·4
]10
·312
·0[1
2·6]
12·0
13·5
[13·
9]12
·614
·311
·9K
NM
-KP
2928
4R
9·8
11·4
(11·
3)(8
·6)
KN
M-K
P29
286
L7·
3[8
·7]
7·8
10·4
12·4
9·4
9·5
11·3
12·3
11·9
13·8
14·3
13·2
KN
M-K
P29
286
R8·
010
·311
·212
·39·
19·
511
·712
·311
·814
·514
·014
·113
·4K
NM
-KP
2928
7L
6·8
8·6
8·5
9·8
11·4
13·6
12·8
13·6
KN
M-K
P29
287
R8·
210
·3(1
4·2)
13·1
KN
M-K
P30
500
L13
·48·
913
·313
·515
·815
·1K
NM
-KP
3050
0R
13·8
13·1
15·9
14·7
17·0
13·4
KN
M-K
P31
712
L6·
612
·110
·5K
NM
-KP
3171
2R
11·5
10·5
KN
M-K
P31
717
R13
·712
·4K
NM
-KP
3173
0L
(10·
0)K
NM
-KP
3472
5L
7·3
8·5
12·1
13·7
12·2
13·0
14·0
KN
M-K
P35
847
L13
·6

261AUSTRALOPITHECUS ANAMENSIS
Tab
le2
Con
tin
ued
Spe
cim
enS
ide
di2
MD
LaL
Dec
iduo
usm
axill
ary
dent
itio
nK
NM
-KP
3472
5R
(4·5
)(4
·0)
Spe
cim
enS
ide
dcdm
1dm
2
MD
LaL
MD
BL
MD
BL
Dec
iduo
usm
andi
bula
rde
ntit
ion
KN
M-K
P31
712
10·0
7·9
KN
M-K
P31
729
(9·7
)7·
9K
NM
-KP
3472
5R
6·8
(5·5
)K
NM
-KP
3472
5L
(6·7
)5·
6(9
·3)
(6·7
)
Mea
sure
men
tsm
ainl
yfo
llow
Whi
te(1
977)
,exc
ept
teet
har
em
easu
red
aspr
eser
ved
wit
hco
rrec
tion
sfo
rin
ters
titi
alw
ear
give
nin
[],
and
BL
dim
ensi
ons
for
M1
and
M2
give
nac
ross
mes
ial
and
dist
alcu
sps
inde
pend
entl
y.A
llm
easu
rem
ents
give
nin
mm
.(
)=va
lue
esti
mat
eddu
eto
dam
age.
a=to
oth
mea
sure
das
pres
erve
dde
spit
ein
ters
titi
alw
ear;
esti
mat
ing
orig
inal
dim
ensi
onre
liabl
yis
not
poss
ible
.

262 . . ET AL.
individual has been recovered from a site, anote is given at the beginning of the speci-men description with possible associationsindicated. A map showing the relationshipof several of the hominin discoveries foundclose to the type mandible is given in Wynn(2000, Figure 2).
Table 3 Mandibular metrics for Australopithecus anamensis*
KNM-KP 29281 KNM-KP 29287 KNM-ER 31713
Minimum breadth between crownsP3 25·9 — —P4 28·1 — —M1 26·4 — —M2 26·8 — —
Breadth between crown centersP3 35·9 — —P4 37·2 — —M1 37·6 — —M2 38·6 — —
Symphysis dimensionsSI height 27·0 27·0 27·0AP length 32·2 38·0 29·0Maximum length 41·0 42·0 33·8Minimum breadth 21·0 19·5 17·7Angle to alveolar margin 141 135 142
Left Right Left Right Right
Corpus dimensions at P4/M1 junctionSI height 34·0 34·0 — 42·3 29·0ML breadth 19·6 18·5 — 22·5 19·7Minimum breadth 17·9 17·7 — 19·7 17·8
Height of mental foramen centerFrom alveolar margin 18 19 20 23 —From base of mandible — 16 — 22 17
Mental foramen SI height 2·2 2·8 2·6 — —Mental foramen AP length 4·1 3·3 4·6 — —
*Mandibular metrics follow Wood (1991).
Craniodental FossilsKNM-KP 29281—Holotype mandibleand temporal fragment (Figures 1 and 2).The two pieces of this specimen were foundonly a few centimetres apart and are there-fore included under one accession numberas a single individual.
A: Holotype mandible (L and RI1–M3)
Preservation. This adult mandible consistsof most of the body with all the permanentteeth, which are complete except for someminor damage. It is missing its rami, angles,and the base posterior to M1. The left andright sides of the body are separated by alarge crack, and the two halves do not matchperfectly. There are several puncture markson the lateral and medial surfaces of thebody, probably carnivore tooth marks. Sev-eral cracks are probably weathering damagecaused as the mandible dried on a landsurface. Minor distortion is caused by cracksthat have opened throughout the body. Theincisors and canines are hinged anteriorly intheir alveoli, particularly on the left side, andmatrix has filled in the gaps posterior to

263AUSTRALOPITHECUS ANAMENSIS
Table 4 Metrics for new Australopithecus anamensis postcranial elements
Tibia KNM-KP 29285 Phalanx KNM-KP 30503
Proximal Maximum fragment length 32·9Maximum fragment length 103 Midshaft DP depth 7·0Plateau maximum ML (70) Midshaft ML breadth 9·7Plateau maximum AP (46) Distance from flexor ridges from
proximal joint surface center24
Medial condyle AP (45) Proximal end DP depth (9)Medial condyle ML (27) Proximal end ML breadth 12·7Lateral condyle AP ((37))Lateral condyle ML ((26))Minumum intercondylar distance 16Plateau to distalmost point on tuberosity 31Maximum shaft breadth at break 33Minimum shaft breadth at break 21Semimembranosus m. insertion site SI 15Semimembranosus m. insertion site ML 18
DistalCapitate KNM-KP 31724
Maximum fragment length 96 PD length 21·8AP shaft breadth at break 22 DP depth (14·8)ML shaft breadth at break 22 ML width of head (12·5)Maximum AP depth metaphysis 33Maximum ML breadth metaphysis 43AP depth of talar facet 24ML width of talar facet at center 26ML width of talar facet at lateral edge 13ML width of talar facet at medial edge 21SI length of malleolus 14Maximum AP depth of malleolus 13SI height of fibular facet 7AP depth of fibular facet (16)Width of flexor m. groove 8
( ) indicates estimate, (( )) more tentative estimate.
them. The left I1 and I2 have shiftedanteriorly by about 4·3, the right I1 by a littleless, and the right I2 by only a little. The leftincisors have also rotated slightly laterally.The right /C has shifted forward for 2·0, theleft /C by 3·9. The true positions can befound by realignment of casts of the teethusing the IPFs as a guide. The labial sides ofthe incisor and canine alveoli are broken off,as is a piece on the inferior side of thesymphysis. In posterior view, it is apparentthat the left M3 is in a piece of the body thathas been rotated slightly clockwise a fewdegrees.
Lateral aspect. The tooth rows show a mildheliocoidal occlusal wear plane and a weak
Curve of Spee. The alveolar margin isstraight and parallels the occlusal marginwhere it is preserved. Although the rami aremissing, in places a narrow extramolarsulcus is visible. A mild jugum for P4 and aslightly larger one for the mesiobuccal P3
root interrupt the body surface. The caninejuga are missing, but would have formedrounded anterolateral corners to the man-dibular body.
The lateral superior torus is merely theextension of the oblique line for the attach-ment of the buccinator muscle, and existsonly as a rounded eminence that disappearsbelow M1. A narrow marginal torus runsanteriorly, passing just inferior to the mental

264 . . ET AL.
Figure 1. Holotype mandible, KNM-KP 29281A. Top row, occlusal and basal views; middle row, rightand left lateral views; bottom row, right and left medial views.
foramen and fades out below P3. It is separ-ated from the base by a shallow groove.Anterior to the lateral torus and above the
marginal torus is a broad, shallow intertoralsulcus centered superior and anterior to themental foramen. There is a single mental

265AUSTRALOPITHECUS ANAMENSIS
foramen on each side. Each is centeredbetween P4 and M1 directly above the baseof the symphysis, 17·5 down from thealveolar margin on the left and 19·2 on theright. They open anteriorly. The left one hasabraded margins, but the right one measures3·5 AP by 2·6 SI. The mental surfaceis strongly but smoothly convex in bothdirections.
Posterior aspect. The postincisive planum isabout 24 long, reaching posteriorly as far asthe level of the P4s and is concave in bothdirections. It ends in a strong superior trans-verse torus, which swings laterally from themidline to merge into the medial body wallat the level of the junction between M1 andM2. There is an even stronger inferiortransverse torus centered 16 inferiorly andposteriorly from the superior one, and separ-ated from it by a marked genial fossa. Theinferior torus reaches as far posteriorly as themesial edge of the M1s at midline, con-tinuing almost directly laterally to mergewith the body also level with the M1s. Thereare faint digastric impressions on the under-side of the inferior torus on each side, whichmeet in the midline to form a crest in thecenter of the torus.
Medial aspect. The symphyseal cross-section is strongly inclined, slopingposteriorly with its long axis running 141�to the alveolar margin. The entire externalcontour is smoothly convex through itsrounded inferior margin, with no flatteningevident basally. The alveolar margins arestraight. The alveolar torus is mild through-out its length except below M3, where itbecomes more prominent. The alveolartorus is only about 15 SI here, but deepensanteriorly to merge with the superior trans-verse torus. Inferior to the alveolar torus, thesubalveolar fossa is deepest posteriorlybecoming shallow anteriorly, where its in-ferior edge swings up and over the inferiortransverse torus allowing it to blend into thegenioglossal fossa. No mylohyoid line isevident.
Basal aspect. Although much of the base ismissing, the preserved contours suggestthat it was everted posterolaterally. Thepreserved basal contour is rounded.
Occlusal aspect. The postcanine tooth rowsare almost parallel and set close together.The original minimum distance between thecanine crowns before distortion can be esti-mated at about 20. Minimum distancesbetween the lingual sides of the oppositeteeth are: P3–P3 25·3, P4–P4 27·2, M1–M1
25·2, M2–M2 25·4, M3–M3 about 26·5.This last measurement is an estimatebecause of the slight distortion here.
Teeth. Morphological descriptions of theteeth are given for the best preservedside, with any differences between sidesnoted.
The LI1 is complete, with two transversecracks through its root that cause no signifi-cant distortion. The RI1 has alveolar boneadhered to its medial surface. It is missingthe inferior labial enamel, and some othersmall chips. It is split longitudinally, andtransverse cracks across its root areexpanded causing distortion. The left side isdescribed.
This tooth is worn, exposing an area ofdentine across its occlusal plane that is 5·4MD by 2·5 LaL, with only a thin strip ofenamel around its margin. Only 6·9 of labialenamel and 6·5 of lingual enamel remains.The occlusal plane is mildly concave inlabial or lingual view and angled slightlydistally and lingually. The crown flares outmesially and distally from 4·3 wide mesio-lingually at its base to 5·9 at its occlusaledge, and would have been wider before itwas worn. Lingual and labial surfaces aretransversely convex. The lingual surface isconcave SI above its prominent basal bulge.Traces of mesial and distal lingual groovesare left just below the occlusal margin. Atiny area of the IPF remains at the occlusaledge of the mesial surface. Faint traces oftransverse hypoplastic lines are visible onthe labial face. The root is strongly convex

266 . . ET AL.
labially and concave lingually, measuringabout 17·5 in length from the labial side ofthe cervix. It swings mesially just at its apexand there is a distinct mesial groove. At thecervical margin, it measures 4·1 MD by 7·3LaL.
The right I2 is broken in a similar way tothe right I1. It is missing the mesiolingualcorner of its crown, so the completely pre-served left I2 is described. This tooth has alongitudinal crack, but has suffered negli-gible distortion. It has dentine exposedacross the entire flattened occlusal surface(5·9 MD by 3·4 LaL) except for a narrowrim of enamel. Only 6·5 of enamel is left SIlabially, and 7·4 lingually. Both sides of thecrown flare from the cervix increasing MDfrom 4·8 to 6·6. The occlusal plane appearsto slope slightly mesially and lingually, butthis impression is mainly due to the higherdistolabial corner. Its labial face is uniformlyconvex in all directions. The lingual facepreserves shallow mesial and distal groovesand a mild median ridge above the basalswelling. Interproximal wear has marked themesial margin of the occlusal face, but nofacet is measurable. The root is 4·5 MDnear the cervical margin, but mandibularbone and matrix obscures its LaL dimen-sions. It is 17·4 long and swings distally at itsapex.
The left /C is missing its mesiolabialcorner, and other small areas of enamel. Theright one is complete, so is described. Thiscrown is a rounded rhombus in occlusaloutline. Its occlusal wear plane runs throughthe high Mmr, and across the crownthrough the low Dmr located near thecervical margin. The wear plane is angledslightly to slope lingually but is horizontal ina mesiodistal direction, except where it turnsto slope sharply inferiorly along the distalmargin of the crown. The exposed dentineis hollowed out inside the narrow enamelrim. Although the mesial and distal basalmarginal ridges are worn at their highestpoints, they are intact and confluent as
they swing around the base of the crownlingually. Labially, they are faint, as are thelabial grooves. The lingual ridge is asym-metrical, and has a sharp distal margin alonga narrow distal lingual fossa, but a flat mesialedge that merges gently with the distal edgeof the broad, shallow mesiolingual fossa.The labial surface is tightly convex trans-versely, and somewhat so SI. Hypoplasticlines are visible on labial and lingual faces,and perikymata are apparent on the labialface. Root dimensions cannot be measured,but on the right side the canine root isexposed for most of its length, and can beseen swinging gently distally.
Both P3s are well preserved, but the rightone shows slightly less wear so the descrip-tion is based on this. The occlusal outlinewould be rhomboidal but for the inflateddistolingual corner. This tooth has one maincusp near the center. Dentine is exposed in a1 wide pit on the Prd, and along the distalocclusal ridge. The mesial and distal occlu-sal ridges and transverse crest are sharpand prominent. The transverse crest has acuspule representing the Med where itmeets the Mmr and Dmr. Lingually theMmr is narrow and low, arching towards thebase of the crown, and bounding a narrowFa that opens lingually close to the Med.The Dmr is stronger and higher, andbounds a larger Fp. Buccally, there is apit-like mesiobuccal groove alongside theMmr. The Mmr swings inferiorly and dis-tally to merge with the basal enamelswelling. There is only a shallow trace ofa distobuccal groove. The buccal faceslopes strongly toward the cusp apex, and isbiconvex. An IPF measuring 4·7 BL hasflattened the distal margin of the tooth.Three roots are present. The two distinctbuccal roots diverge apically.
The P4s are also well preserved, butslightly more occlusal enamel remains onthe left one on which the description isbased. The occlusal outline is asymmetrical.The left P4 is rhomboidal in outline,

267AUSTRALOPITHECUS ANAMENSIS
although the right one has a less extensivemesiolingual corner and presumably asmaller Med originally. There are two cusps.The Prd is large and the Med much smaller.The Prd is almost completely worn off,exposing dentine that extends to the mesialand distal margins of the tooth, widening to2·1 BL at the cusp center and making itlower than the Med. A pinpoint of dentine isvisible on the Med. A pit-like remnant of theFa remains, as does a lower and largerFp surrounded by a prominent Dmr. Thelingual face is straight vertically, and mildlybilobate in occlusal profile, with two faintvertical enamel wrinkles. The labial face isbiconvex and slopes towards the buccalcusp. A faint distobuccal groove remains,indenting the occlusal profile of the crown.Interproximal wear has flattened mesial anddistal margins of the tooth. The mesial IPFis 4·7 LaL, and the distal one is 6·0 LaL.
Both M1s are well preserved, but the leftone is missing a small piece of enamel fromits mesiobuccal corner. Description is basedon the right, even though it is missing apiece of enamel along its distal margin. Theocclusal outline is a rounded rectangle. Thebuccal side is worn more heavily than thelingual side, so that the buccal cusps, Hldand all foveas have been replaced by anelongate, hollowed area of dentine. Thelingual cusps have pinpoints of dentineexposed, and on the Med, this area ofdentine extends halfway down towards theFc. Wear has obscured details of relativecusp sizes but the Med is the highest, and islarger in area than the End. The Mmr isalmost worn through by interproximal wear.The Dmr is intact on the unbroken left side.The mesiobuccal groove is set slightly mesialto the lingual one. It continues across thevertically convex buccal face where itbecomes doubled, and forms a stronglybilobate occlusal profile as it continues tothe cervix. The lingual groove is more pro-nounced occlusally, and is sharp as it cutsthe occlusal margin and passes along the
vertical, bilobate lingual face to the cervicalline. Interproximal wear has left a broadconcavity in the mesial occlusal profile,and has flattened the distal profile of theunbroken left side. On the right, the mesialIPF measures 6·1.
Although both M2s are completely pre-served, the right one is described because itis slightly less worn. This tooth is a roundedrectangle with bilobate sides with a barelyconcave mesial margin caused by inter-proximal wear. The buccal cusps are muchlarger than the lingual ones, and togetherwith the Hld comprise more than half thecrown. The lingual ones are higher as pre-served, with the Med the tallest. The entiresurface is worn, but dentine is exposed in a1·7 MD slit on the right Prd. The slit-like Fais located to the lingual side of the MD axis,as are the Fc and Fp. Only a trace of the Fpremains. The lingual and mesiobuccalgrooves are continuous across the crown.The lingual groove sharply incises theocclusal margin, and continues to the cervix.Faint vertical enamel wrinkles mark thevertical lingual face. The mesiobuccalgroove splits just before it passes the occlusalmargin, and has a mesial branch that termi-nates occlusally at a large protostylid, and adistal branch that continues to the cervixalong the distal margin of the protostylid.Immediately posterior to this branch arethree small, vertical enamel grooves extend-ing along the superior half of the slopingbuccal crown face. The distobuccal grooveis largely obliterated by occlusal wear, and isfaint along the side of the crown. The mesialIPF indents the mesial occlusal profile andmeasures about 5·0 BL. A distal IPF isobscure, although one is visible on the leftM2.
The M3s are both missing pieces ofenamel from their distobuccal corners, butthe right one is missing more. The left onehas a crack through the mesial cusps and thetip of its Med damaged. Description isbased on the left. This tooth is an elongate

268 . . ET AL.
Figure 2. Holotype temporal, KNM-KP 29281B in lateral and inferior views.
rectangle in occlusal outline. The occlusalsurface is worn with polishing on all cuspsand flattening of the buccal cusps and Hld,but no dentine is exposed. The Med ishigher than the others, but the Prd is largestin area. There is a large Hyd and a smallerEnd. Each is divided into mesial and distalparts by grooves. A C6 is present and largerthan the Hld. A deep, transversely wide Faruns from the MD axis of the crown to thelingual margin, bounded by the Mmrmesially and a transverse ridge connectingthe mesial cusps distally. There is no Fp.The lingual and mesiobuccal grooves aredeep and continuous, meandering across thecrown, incising the occlusal marginsstrongly and continuing to the cervicalmargins. Vertical enamel grooves are visibleon the buccal, lingual and distal faces, someof which incise the occlusal margin. Noprotostylid is visible on the left M3, althougha distinct one is seen on the right one. TheM3 roots incline strongly buccally andmildly distally, as seen on the damaged rightside.
B: Left temporal fragment—This pieceof left temporal bone consists of a maximum
of about 52 AP. About 25 SI of thesquamous portion remains, as does much ofthe temporomandibular joint and thetympanic with the external acoustic porus.Laterally, the zygomatic process is brokenoff at its base from the prearticular areaposteriorly, cutting through the mandibularfossa and postglenoid process. Several largeair cells are exposed here. The lateral marginof the external acoustic porus is preserved atits inferior edge, but the rest of the margin isbroken away. The inferior surface of thefragment extends posteriorly from the sphe-nosquamosal suture. It preserves much ofthe articular surface, part of the prearticulararea and inferior surface of the tympanic.The sphenotemporal suture is preservedabout 25 anteriorly from the tympanic. Themedial surface of the fragment shows amixture of broken bone pieces and matrix.
The small remaining piece of the squamehas fine ridges running superoposteriorly,especially on its posterior part, that followthe course of the temporalis musclefibers. There is a small tubercle about 4·5anterior to the most anterior portion of thezygomatic root, in the position to have been

269AUSTRALOPITHECUS ANAMENSIS
associated with a septum that divided thetemporalis muscle. It lies at the anterior endof a faint linear ridge that, like the striationsfurther posteriorly, runs superoposteriorlyacross the squame. The external auditoryporus is small and elliptical in outline. Itslong axis is oriented 90� to the superiorsurface of the zygomatic origin. The porusmeasures a maximum of 8·9 by 5·7 aspreserved.
The articular tubercle is flat, with only amild convexity demarcating it from the in-ferior temporal surface. The tympanic platemakes a large angle with the articular surfaceof the temporomandibular joint, so that thewhole of this medial part of the temporo-mandibular joint is just one shallow, cup-like cavity. It is not possible to estimate howmuch of the articular surface extendedlaterally past the squame, but about 20 ispreserved medially. The squamotympanicfissure is represented by a 6·4 long slit in themiddle of the cavity. The postglenoid pro-cess is fused to the tympanic, and wouldhave continued laterally to it. As preserved,the tympanic plate measures 11·6 AP fromthe squamotympanic fissure to the brokenedge of the stylomastoid foramen and 26·3perpendicular to this to the edge of theacoustic porus. The sheath of the styloidprocess is partly preserved on the posterioraspect of the tympanic.
On the medial surface, some remainingpieces of bone appear to be endocranialbone and one has a trace of a meningealvessel groove on it, but on the whole thisfractured surface is uninformative.
KNM-KP 29282: LM1 or M2 (not figured).This is a broken molar, either M1 or M2,preserving a worn crown and partial roots. Itwas discovered during screening, just wash-ing into the sandy stream channel at the baseof the slope from which the type specimen,KNM-KP 29281, and the worn premolarand molar, KNM-KP 31730 were found(Wynn, 2000, Figure 2). It is possible that
this specimen is the same individual asKNM-KP 31730, but separate accessionnumbers have been allocated because of thelarge distance, from the base to the top ofthe slope (approximtely 10 m), separatingthe two specimens.
The molar is the most heavily wornmandibular molar in the sample, withdentine exposed over the entire occlusalsurface. The roots are nearly completebuccally, but are largely missing lingually.Mandibular bone is preserved between theroots, but only the buccal cortex remains.The occlusal outline is rectangular. Thebuccal side is broken, and enamel is missingfrom the mesiobuccal corner, and distal sideof the crown. The smaller area of lingualdentine is set at higher plane than the muchlarger buccal portion, which is stronglyhollowed out transversely. On the buccalface, the mild mesiobuccal groove extendsto the cervical margin. Wear and damageobscures any potential evidence of a proto-stylid. A tiny portion of the mesial IPF isvisible and contacts the occlusal margin.Distally no IPF is preserved. The remainingmesiobuccal root is at least 14·5 long andless distally inclined than is the remaining11·5 long distobuccal one.
KNM-KP 29283: Maxilla (Figure 3). Thismaxilla was a surface discovery, but the I1
and both lateral incisors were recoveredseparately through excavation.
Preservation. This is a weathered adultmaxilla with all teeth except the left I1 andright M3. The maxilla is preserved in twohalves. Both preserve the intermaxillarysuture anterior to the nasal aperture.Posterior to this, the left side preserves thesuture inferiorly along the palatine process.The right side, however, is broken 3 laterallyfrom the midline on the palatine process.Therefore, the two halves can be articulatedonly anterior to the incisive foramen andnasal aperture. The superior portion of theclivus is missing on both sides of the midline

270 . . ET AL.
Figure 3. KNM-KP 29283 maxilla with associated I2s. LI2 in lingual, distal and mesial views; RI2, inlingual, distal and mesial views; maxilla in facial view at top, second row, superior and palatal views; thirdrow, left and right lateral views; bottom row, left and right medial views.

271AUSTRALOPITHECUS ANAMENSIS
within the nasal aperture. The incisor alveoliare partially preserved, with their labialportions broken away. The labial and buccalmargins of the canine alveoli are brokenaway over the entire length of the canineroot on the right, and for much of it on theleft. On the right side, the P3 buccal rootsare exposed and much of the P4 roots, but itis impossible to tell how much of the rootexposure is due to damage and not resorp-tion. The right I1, both I2s and the RC/ werefossilized separately. On the left side, themaxillary tuberosity is eroded, exposingcancellous bone. Only about 10 of bone ispreserved above the alveolar margins. Thereis lateral exposure of the distal M1 root,probably due to abrasion.
Facial aspect. The anterior margin of themaxilla is slightly convex transversely wherepreserved. There is a mild, vertical depres-sion laterally where the bone flares aroundthe canine jugum on each side. The I2
alveoli are set lateral to the nasal cavity. Theinflated canine juga and the anterior P3 jugaform large rounded anterolateral corners tothe profile of the bone. The nasal aperture issmoothly continuous with the external sur-face of the bone. No ridges or crests delimitthe preserved portion laterally or inferiorly.The lateral walls of the aperture extend 12·5superior to the floor on the right and abouthalf that on the left. The nasal aperture is22·3 wide. A depression about 8 wide justinside the lower lateral margin of the nasalaperture is delimited posteriorly by arounded ridge that runs posteriorly andsuperiorly from the posterior edge of theclivus.
Lateral aspect. The lateral contour of thealveolar process is gently convex. Thecanine fossae are asymmetrical and quiteirregular. On the right side, the canine fossais deep and measures just over 11�11.There may be some pathology, also sug-gested by an irregular and convoluted floorof the maxillary sinus on this side comparedwith that of the left. On the right side, the
alveolar bone along P3 to M1 is resorbed,exposing the bases of the roots. Just postero-inferior to the canine fossa, right above thetips of the distobuccal P3 and single buccalP4 roots, there is a circular depressionmeasuring 5 in diameter of obscure origin.The root of the zygomatic is preserved as araised area of abraded bone. Inferior to it isa shallow depression, bounded posteriorlyby the rounded jugum for the distobuccalM1 root. The jugum for the distobuccal rootof M2 is pronounced. These juga give thelateral margin of the alveolar process amildly undulating profile. Similar contoursare seen on the more abraded left side. Onthe left, however, the canine fossa appears tohave been smaller, although it is brokensuperiorly. There is also less root exposure.
Inferior aspect. The postcanine tooth rowsare almost parallel to each other, and areslightly concave lingually. Minimumbreadths of the palate between teeth are: C/34, P3 38, P4 38·5, M1 38, and M2 35·5.The palatine processes rise graduallyposteriorly, so that the palatal depthincreases steadily. As measured from toothcervices, palatal depth is 6 at P3, 8 at P4,10·5 at M1, and 12·5 at M2. The length ofthe tooth row from the mesial side of C/ todistal M3 is 63·3. The incisive foramen isabout 5 in transverse diameter and is cen-tered 17 from the alveolar margin, betweenthe P3s. The greater palatine foramen is notpresent, but the vascular groove is, lying atthe inflection between the alveolar and pala-tine processes. On the left side, a severealveolar resorption pit measuring 15 AP hasexposed up to 9·0 of the M1 roots and 4·0 ofthe P4 root. On the right side, resorption isless extreme, but is present in roughly thesame location.
Superior aspect. The nasal cavity is narrow-est at a point just posterior to the internalridge, where it is about 21·5 wide, broaden-ing posteriorly to about 27 above the level ofthe M1. On the left, the maxillary sinus as itis exposed is 46 AP, running from a point

272 . . ET AL.
level with P4 to the posterior break. It is upto 15 ML, with its widest point above M1. Itis divided into a large anterior cavity, a smallmiddle cavity superior to M3, and an evensmaller one posterior to that. There is a faintdivision of the larger cavity separating theroots of M1 and M2. The right maxillarysinus reveals a more complex set of irregularloculi than the left, with a deep anterolateralpocket reaching down towards the caninefossa, and two smaller pockets preservedsuperior to the canine root. The sinus ispreserved for 37 AP, running from the levelof P3 only to M2. It is up to 17 ML at the M1
level.Medial aspect. Subnasal anatomy is
exposed on the midline break. The incisivecanal is 3·2 wide in the sagittal plane nearthe foramen and expands posteriorly toabout 6 near the nasal cavity. The long axisof the canal runs superiorly and posteriorlyat an angle of about 25� to the inferiorsurface of the palatine process. The naso-alveolar clivus is a maximum of 11·4 thick.From the anterior margin of the nasalaperture, the floor of the nasal aperturepasses backwards about 13 before droppinginferiorly about 12 to the palatine processes.The left palatine process, which preservesthe midline suture, expands in thicknessposteriorly from a thin apex at the incisivecanal to 6 at the level of M2 at midline. Theopening of the incisive canal is just over 12across, and it opens posteriorly and laterallyto the widest portion of the nasal cavity.
Teeth. The teeth are heavily worn,especially anteriorly. The right ones are lessheavily worn, so these are described whereboth sides are preserved. Little detailedocclusal morphology remains. There is aclear heliocoidal wear pattern along eachtooth row.
The right I2 consists of the root and thelabial portion of the crown. The labialenamel is only preserved for an arc 3·2mesially and 2·2 distally. The lingual side isworn well below the original cervical
margin, so that the occlusal plane wasoriented superolingually and slightly dis-tally. The root is broken superiorly, but canbe replaced into the preserved portion of itsalveolus. The root is 22 long. It is triangularin outline near the cervix and becomesrounder apically. The root is convex in out-line on its labial side and concave on thealveolar side. A mesial groove is present.
The I2s are morphologically similar, sothe more complete left one is described.Only 2·4 of enamel remains labially on theleft, although 3·8 remains on the right. Theroot is worn down below the original cervi-cal margin lingually, so that the occlusalwear surface is strongly angled labially. It isalso angled to slope lingually and distally. Anarrow band of enamel remains mesially,but almost none distally. The root is 22 longfrom the labial enamel line. It is stronglycompressed MD, measuring 4·6 MD by8·2 LaL near the cervix, and is hookedlabially at its tip. The preserved labialand lingual margins of the root are mildlyconvex, so that it widens to a maximum of9·0 LaL. The root has mild mesial and distalgrooves.
The canine has only 3·5 of the labialenamel and a thin ribbon of distal enamelremaining. Its occlusal surface is planar andslopes lingually. Almost the entire root isexposed. It is fairly straight but convexlabially, 28·6 long. It is oval in section,measuring 9·0 MD by 11·5 LaL near thecervix. The root has a mesial groove. It isnearly vertical, with its long axis orientedat over 85� to the cervical margins of thepostcanine teeth in lateral view.
All that remains on the P3 crown is a verythin rim of enamel. It had about 6 of buccalenamel remaining, but most of the buccalenamel has been broken away. The dentineexposure is hollowed out. Mesial and buccallabial grooves extend about 2 superiorlyfrom the occlusal plane, and there is apronounced basal enamel bulge. It has threedistinct roots. On the right, the roots are

273AUSTRALOPITHECUS ANAMENSIS
exposed for 13·0 mesially and 12·7 distally,which is most of their length. The mesio-buccal and distobuccal roots diverge atabout 30�.
The right P4 is displaced superiorly. It hasenamel chips missing from its buccal andlingual surfaces. The left P4 has lost enamelfrom the mesiolingual corner. Some occlusalenamel remains between the cusps and inthe Fp. A 3·2 wide area of dentine isexposed lingually, but the buccal exposurecannot be measured owing to damage.Extensive contact facets are evident, alteringthe mesial and distal faces of the crown.
The left M1 is missing enamel on most ofthe mesial half of the tooth, and the rightone is missing a small piece from the disto-lingual corner. In occlusal outline the M1
tapers distally. The lingual half of the crownis a heavily excavated dentine basin sur-rounded by a 2 high thin rim of enamel.Wear has flattened the buccal cusps.Dentine exposure is difficult to assess due toweathering, but there is a clear 1·3 widedentine pit on the Pa. The two buccal cuspsare mostly preserved, with the Me the larger.A pit-like Fp remains bounded by a Dmr.The buccal groove is pronounced, and con-tinues to the cervical margins making thebuccal face strongly bilobate. Interproximalwear has removed much of the mesialenamel.
The occlusal outline of the M2 also tapersdistally. The preserved buccal cusps areroughly the same size and flattened bywear. Dentine is exposed as a continuousexcavated strip along the two lingual cusps.There is also a 1 wide dentine pit on the Pa.The shallow remains of a Fp are present.Both lingual and buccal faces are bilobate,with lingual and buccal grooves extending tothe cervical margin. A weathered distal IPFis visible but cannot be measured.
The M3 has a strongly tapered occlusalprofile, with a rounded distal margin. Themesial cusps are substantially larger thanthe distal ones, occupying most of the
crown. The Pa is higher than the distalcusps. The occlusal surface is flattened bywear and there are two mesial dentineexposures, 1·9 BL on the Pa and nearly 5and deeply excavated on the Pr. The lingualand buccal grooves are mild, and extend allthe way up the crown faces.
KNM-KP 29284: R/C and RP3 germs(Figure 4). This specimen was found in situin an excavation to recover further partsof the adult maxilla, KNM-KP 29283.Because these two teeth are germs they areclearly from a different individual.
A: R/C. This is the nearly completecrown or a /C germ that is slightly weatheredand cracked around its cervical half. Thereis no trace of any root formation. It isstrongly asymmetrical. In occlusal outline, itis almost a quadrilateral with corners mesio-labially, mesiolingually, distolabially andthe most prominent one distolingually. Itslingual face is marked by pronounced mesialand distal lingual fossae and a sharp lingualridge, all of which are concave distally. Themesial edge of the lingual ridge is rounded,but its distal edge is sharp. The high Mmris faint as it passes the mesial edge. Theincisive and cervical halves of the sharpmesial margin are set at about 30� to oneanother in lingual view. The cervical portionalmost parallels the distal margin of thecrown and the incisive portion angles dis-tally towards the apex. The sharp distalmargin of the crown is concave, swingingout inferiorly to meet a strong distal basaltubercle 10·0 from the crown tip and 3·3from the preserved base. This tubercle existsas the pinnacle of the Dmr, which inclinesinferiorly both lingually and labially, butmore steeply labially. Lingually, it meets thebase of the lingual ridge. Nested just abovethe distal tubercle there is a smaller tubercle.Labially, there is a broad distolabial groovewith a sharp distal edge and a gentler mesialedge that blends into the labial face. Thelabial face is tightly convex transversely, but

274 . . ET AL.
Figure 4. Dental specimens from Kanapoi not associated with jaw fragments. KNM-KP 29284A inlingual, distal and occlusal views, KNM-KP 29284B in lingual and occlusal views, KNM-KP 30505 inocclusal view, KNM-KP 30502D, KNM-KP 30502E, KNM-KP 31714, KNM-KP 31723 in occlusalview, KNM-KP 31717A in mesial, distal and occlusal views, KNM-KP 31717B and KNM-KP 31726 inocclusal view, KNM-KP 31729 in buccal, distal and occlusal views, KNM-KP 31728, KNM-KP 35842,KNM-KP 31730A and KNM-KP 31730B in occlusal view, KNM-KP 35847 in buccal and occlusal view,and KNM-KP 35852 in mesial and occlusal views.

275AUSTRALOPITHECUS ANAMENSIS
is almost straight SI. There is almost nomesiolabial groove, only a sharp, straightedge inferior to the Mmr. Hypoplastic linesare evident on the lower half of the labialface, with a particularly strong one 2·4above the inferior margin. Perikymata arealso visible.
B: RP3. The crown of this germ is almostcomplete with no trace of root development.It is split in two by a vertical crack passingmesiodistally through its middle, but it is notdistorted. Lingual enamel is broken awayfrom the sides adjacent to the basins,although more extensively so around the Fp.Enamel is also missing from the inferioredge of the distal margin of the crown. Thecrown has an ovoid occlusal margin, withthe long axis running mesiobuccally todistolingually. The large Prd is located in thecenter of the tooth. The mesial and buccalocclusal ridges and transverse crest aresharp. The transverse crest meets a tinyMed. The mesial ridge is continuous withthe sharp Mmr. The distal ridge meets theDmr at a distal tubercle. The Fa is BLnarrow, and the Fp is broader with its sidesmarked by radial wrinkles. The buccal sur-face of the tooth is convex transversely. It ismildly convex SI, inclining strongly towardthe Prd. Mesial and distal labial grooves arebounded by sharp marginal ridges. Faintvertical enamel ridges are visible on thebuccal face.
KNM-KP 29286: Mandible fragmentswith associated mandibular dentition(I1, L & R I2–M3) (Figure 5). Apart fromthe fragments that make up KNM-KP29286A, this specimen was recoveredthrough screening and comprises a completeset of lightly worn associated teeth. Severalhave their roots embedded in expanded andcracked mandibular bone and matrix. Sev-eral tooth fragments that do not appear tobelong to this specimen were also recoveredbut given a separate accession number,KNM-KP 31732.
A: R mandibular fragments (I2–P3).This piece of mandible is badly broken. Theonly remaining cortex is about 1 along itsinferior edge below I2 and much of the areabelow P3, although here it is weathered. Thealveolar margin between I2 and /C is mildlyabraded but close to preserving its originalcontour. Most of the rest of the bone isbadly splintered, distorted and intermingledwith matrix. The mesial wall of the P4
alveolus is exposed at the posterior break.The tip of the I2 crown is broken off, with
enamel preserved only 3·9 labially and 5·7lingually. The break exposes the pulp cavity.The root is complete although the tip isseparated from the rest of the tooth by a 3wide transverse crack. The superior halfis displaced slightly labially relative to theapical end, which appears to be in itsoriginal position. Most of the tip is obscuredby matrix. The I2 lingual face preserves amild basal tubercle, above which it is gentlyconcave SI and flat transversely. The gentlyconvex labial face has faint vertical enamelwrinkles and perikymata visible. The rootmeasures 5·1 MD by 8·0 LaL near thecervical margin. The exposed side of theroot has a mesial groove.
The canine crown is complete except for asmall area of enamel at its distolabial cervi-cal margin. The distal half of its root iscomplete, but the mesial half is missingabout 11 from the mesial cervical margin,exposing the pulp cavity throughout itslength. The canine crown measures 14·3high labially as preserved, and was probablyabout 15 before wear. It is highly asym-metrical. The tip of the cusp is worn trans-versely, preserving a tiny spot of dentine,and polishing extends most of the way alongthe distal crown margin. The mesial marginis sharp, making a 30� bend at the Mmr inlingual view at midcrown. The Mmr swingsdistally and cervically, terminating inferiorto the wide and deep mesiolingual fossa. Inlingual view, the distal margin of the crownbecomes concave towards the distal basal

276 . . ET AL.
Figure 5. KNM-KP 29286 associated mandible fragments and mandibular dentition. On left, (A), (B),(C), (D), (G), (H), and (I) in occlusal view; (J) in lingual view; (E) in two pieces, mandible fragment withI2 in labial view, and LC in mesial view; bottom right, (A) in lateral view.
tubercle. The Dmr is faint, extending fromthis tubercle mesially. The lingual ridge issharp at its top, but widens inferiorly. Thedistal lingual fossa is a narrow slit. The labialface is transversely convex and mildly so SI.There is almost no mesial labial groove, buta wide, shallow distal labial groove. There islittle basal swelling of the crown on any side.Hypoplastic lines mark the labial surface.The root measures 8·0 MD by 11·7 BL nearthe cervical line. It has a pronounced mesialgroove. The canine alveolus measures about30 deep. The pulp cavity is about 5·5 wideLaL.
The P3 crown is complete, with somehairline cracks through it that cause nodistortion. Most of the middle portion of itsmesiobuccal root is missing. This lightlyworn P3 has a large Prd occupying thecenter of the tooth. The mesial and distalocclusal ridges and transverse crest aresharp, as are the marginal ridges. All showsome polishing from occlusal wear. There isan incipient Med. The Fa opens lingually,because the Mmr dips almost to the cervicalline. The Fp is larger and opens superiorly,as the Dmr is more pronounced and higherthan the Mmr. Buccally, the Mmr and Dmr

277AUSTRALOPITHECUS ANAMENSIS
merge with a horizontal basal enamel swell-ing. Above it, the strongly sloping buccalface is convex transversely and straight SI.The labial grooves are moderate.
B: RP4. This is the crown with up to 4·3of the root. Cracks run through the crownbut cause no distortion. The occlusal outlineis a broad oval that is slightly broaderlingually. It is lightly worn. This tooth isbicuspid. A large Prd occupies about halfthe crown, its apex about one third of theway from the buccal margin in occlusalview. The Med is smaller but almost as high.Mesial and distal occlusal ridges arerounded. The transverse crest is notched inmesial or distal view. The shallow Fa issmaller and higher than the Fp, but both aremarked with enamel wrinkles. The Mmr isnarrow and meets the base of the Med. Thestronger Dmr meets the Med nearer its apexand close to a pit that is separate from theFp. The buccal face is convex transversely.It is nearly straight vertically, slopingstrongly towards the apex, with a gentlebasal swelling that connects the mesial anddistal labial grooves. The distal labial grooveis more pronounced than the mesial one,and delineates a small cuspule at theocclusal margin. An obliquely set mesialIPF measures 2·8 BL by 1·4 SI. Anotheroblique, lingually placed distal IPFmeasures 4·1 by 2·1. The preserved top ofthe root has grooves mesially, mesiolinguallyand distally.
C: RM1. This crown is complete andpreserves up to 2 of the root. The crown isalmost square but slightly elongate MD.From largest to smallest the cusp areas are:Prd, Med, Hyd, End, Hld. All show wear;the lingual cusps are facetted and the buccalflattened with small areas of dentineexposed. A protostylid is present, delineatedfrom the Prd by a narrow slit along itsmesiobuccal corner. The Fa exists as a slitrunning up the Med nearly to its apex fromthe Mlg. Distal to it is a less extensivegroove, bounded posteriorly by a ridge
connecting the two mesial cusps. The Fp is acleft connecting the End and Hld. It istraversed by the distal lingual groove, whichdisappears about halfway down the disto-lingual crown face. The sharp lingualgroove deeply incises the occlusal marginbut disappears halfway down the verticallingual face. The mesiobuccal groove is deepnear the occlusal margin, but is only a broadand shallow depression towards the cervix.The buccal face is strongly bilobate, and thelingual face is straighter. The distobuccalgroove terminates near the occlusal margin.A mesial IPF measures 4·7 BL by 2·3 SI anddoes not meet the occlusal plane. A faintdistal IPF measures about 3 BL by 2 SI.
D: R mandibular fragment (M2–3).This is a piece of badly splintered mandibu-lar bone that preserves no morphology. Thecomplete M2 and M3 crowns are present,with roots partially encased in extensivelycracked bone.
The M2 tapers distally with a roundeddistal margin. From largest to smallest, thecusp areas are: Prd, Med, End, Hyd andHld. The tooth is lightly worn; the lingualcusps are facetted and the buccal flattenedbut without dentine exposure. Its foveasresemble those of the M1. The mesiobuccalgroove passes the protostylid to fade outtoward the cervical margin. The lingualgroove incises the occlusal margin stronglybut is faint on the lingual face. The disto-buccal groove is shallow on the side of thecrown and disappears just past the occlusalmargin. The protostylid is found only on thebuccal face of the Prd and is most pro-nounced at the mesiobuccal groove. Thebuccal and lingual faces are like those of M1.Vertical enamel wrinkles mark the buccalface. The mesial IPF is weathered andcannot be reliably measured. The distal IPFis a sliver 3·2 BL along the occlusal margin.
The M3 is lightly worn on all cusps. It ismesiodistally elongate with a receding,rounded distobuccal corner. Relative cuspsizes and shapes are the same as for M2,

278 . . ET AL.
except that the Hld is smaller and there is asmall C6 adjacent to its pit-like distal fovea.The occlusal surface enamel is crenulated.The Fa is a broad single slit markedby vertical enamel wrinkles. The deepmesiobuccal groove continues to a pointnear the cervix, just distal to a distinctprotostylid that flanks the entire buccal faceof the Prd. The lingual and distobuccalgrooves both disappear midcrown. Themesial and buccal sides resemble those ofthe other molars. Faint vertical enamelwrinkles mark all sides of the crown.
E: L mandibular fragment (I2–/C).This fragment of mandible is badly splin-tered. Part of the distal wall of the I2 alveolusand mesial wall of the /C alveolus remain.The I2 has its crown broken diagonally sothat the exposed face is angled distally. Thepulp cavity is exposed. Up to 7·4 of enamelis preserved lingually and 6·5 labially alongthe mesial edge. A chip of enamel is missingat the mesiolabial corner. A 1 wide trans-verse crack divides the root just below thecrown lingually and 4·4 below it labially.The apical end of the root is missing pastabout 7 from the crown. The /C is missingthe distobuccal corner and superior half ofits crown. The root is preserved for up to13·3 of its length. The morphology of theseteeth is like those of the right side.
F: L mandibular fragment (P3, rootsP4). Small amounts of cortex remain on thisfragment, but no morphology is preserved.The P3 is complete. Two broken roots of P4
are visible. The P3 is like the right one, but ismore compressed MD, with a slightly lowercrown and lower Dmr.
G: LP4. This is the crown with up to 4·5of its root. It cannot be joined to fragment Fdescribed above. The crown is split by a thinmesiodistal crack, but has suffered negligibledistortion. An obliquely set mesial IPFmeasures about 2 BL by 1 SI. A moredistinct, oblique, lingually placed distal IPFmeasures 2·9 by 1·8. In all other aspects itresembles the RP4.
H: L mandibular fragment (M1). Thisis the crown and partial roots of the toothencased in badly splintered mandibularbone that preserves no morphology. A smallpiece of the lateral surface of the body about9 long is visible, but not in its originalposition. The M1 crown is nearly the mirrorimage of the right one, except that a distincttransverse pit is located along the disto-buccal groove incising the edges of the Hydand Hld. The protostylid is more extensiveextending almost to the mesiobuccal groove.Vertical enamel wrinkles are visible on thebuccal and lingual faces.
I: L mandibular fragment (M2–3).These complete M2 and M3 crowns havepart of their roots preserved, encased in abadly fragmented piece of mandibular bone.The only original bone that remains inposition is the alveolar margin around M2.Details of the occlusal groove pattern differbetween sides, and the left ones are lessworn, but these teeth are essentially mirrorimages of the right ones. The M3 has a largerHld and a double C6, contributing to itsmore rectangular occlusal outline comparedto the right.
J: I1. This is most of the crown weatheredin patches. The long IPF on one sidesuggests that it may be a right, as listedin Leakey et al. (1995), but because theother side of the crown is too weathered toassess, this tooth cannot be attributed to aside with certainty. The top of the root isvisible mesially and distally, but the crowndoes not reach the cervical line labiallyor lingually. The tooth is worn, exposing astrip of dentine. The lingual side has gentle,broad mesial and distal lingual fossae. Thelabial side is straight and flat. The IPFmeasures 5·2 SI by 1·6 LaL, and occupiesmost of the right side of the crown in lingualview.
KNM-KP 29287: Mandible with teeth(Figure 6). This specimen was largelyrecovered through screening. The initial

279AUSTRALOPITHECUS ANAMENSIS
Figure 6. KNM-KP 29287. Top row left, occlusal view of mandible KNM-KP 29287 A and B, and right,incisors KNM-KP 29287 C, H, D from right to left in lingual view, and below KNM-KP 29287 C inmedial view. Middle row, mandible KNM-KP 29287 A and B in left lateral views. Bottom row, mandiblein right and left medial views.
discovery, two fragments of left and rightedentulous mandible, were found close tothe to the origin of a small stream channel.Subsequent screening for about 50 m along
this channel led to the recovery of numerousbone and tooth fragments that were recon-structed to make up this almost completemandibular body.

280 . . ET AL.
A: L mandible (alveoli I2–/C, partialcrowns P4–M2, roots P3, partial rootsM3). Preservation. This is a mandible of anadult. The left body is more poorlypreserved than the right, so mandibularmorphology is described for the right side(B). The left portion of the mandible ispreserved anteriorly from a point just lateralto the symphysis internally and about 14anterior to the mental foramen externally,and extends to the third molar distal rootsposteriorly. The left body is damagedanteriorly, with a break passing transverselythrough the alveolus of the I2 and /C. Thebase is missing, with cortex preserved for amaximum of 28 inferiorly from the alveolarmargin on the lingual face. The alveolarmargin is intact only along the lingual side ofM2, and it is abraded everywhere else. Thepreserved cortex is cracked throughout andweathered in places, but distortion appearsminimal. A carnivore tooth mark is visibleanterosuperior to the mental foramen.
Morphology. The mandibular canal isexposed along the broken inferior surfacefrom the posterior break to a point inferiorto the M1–M2 junction, and is about 5·6 indiameter. At the posterior break, thereappears to be a crypt situated just lingual tothe mandibular canal for a supernumerarymolar or a Stafne’s or other type of develop-mental cyst. The canal is at least 7·6 indiameter and immediately lateral to the dis-tal root of M3, which itself is seen inlongitudinal section at the posterior break.
The single mental foramen is oval insection. Its posteroinferior margin isabraded, but it appears to have openedslightly superiorly.
The I2 alveolus is vertically oriented, witha MD width of about 5 and an estimatedminimum depth for the I2 root of 14·6. Theextremely large left canine alveolusmeasures about 10·5 in maximum diametertransversely, in an axis running diagonallyfrom the middle of the I2 alveolus to justbuccal to the P3 root. Although a small
triangle of bone is missing from its lateralmargin, the alveolus is apparently not dis-torted, because a CT image of the preservedright canine root and alveolus reveals com-parable dimensions (F. R. Spoor, personalcommunication). The LP3 has no crown. Asingle mesial root is visible running behindthe canine alveolus. A separate distal, plate-like root appears in the broken cross-sectionof the alveolar margin, but this root bifur-cates deeper in the mandible, as suggestedon the right side and visible on a CT image(F. R. Spoor, personal communication).
The LP4 is almost complete, missingenamel only along its distolingual corner. Itsocclusal outline is a rounded rhombus. ThePrd is the larger of the two cusps and is setdirectly buccal to the Med. The only appar-ent occlusal wear is a flattened light facet onthe Prd that has exposed a small, circulararea of dentine about 0·4 across. The facet-ting runs from the tip of the Prd slightly ontothe transverse crest, and more so mesiallyonto the Mmr although it does not contactthe mesial IPF. The Med and all ridges andcrests are polished. The Fa is smaller andsituated higher than the Fp. The transversecrest is split, so that there is a small Fcbetween them. The Dmr is stronger andwider than is the mesial Mmr, and it mergesbuccally with the buccal marginal ridge. Theoval mesial IPF is 3·3 BL by 2 high. Itcontributes to the flat occlusal outline of themesial side of the crown. There is a singleslight lingual groove, distal to the lingualcusp. On the buccal face, there is a distinctdistobuccal groove but only a trace of amesiobuccal groove.
The LM1 is missing enamel along itsentire mesial margin to the center of the twomesial cusps, so the Mmr and Fa cannot beseen. The crown is a rounded rectangle inocclusal outline with mildly bilobate sides.The tooth has two relatively high facettedlingual cusps, two worn and flattened rela-tively low buccal ones, a flattened Hld and asmall polished C6. The Med is the dominant

281AUSTRALOPITHECUS ANAMENSIS
cusp but the relative cusp areas cannotbe determined due to the missing mesialportion of the tooth. The buccal cusps areset mesial to the lingual ones. Wear facetsare seen on all cusps, with wear striae run-ning predominantly BL. Small, circularareas of dentine are exposed on the twobuccal cusps, with the Prd exposure biggerthan that on the Hyd, measuring about0·5 in diameter. The strong lingual groovetraverses the tooth surface to continue as themesiobuccal groove, but it becomes faintacross an isthmus joining the two buccalcusps. There are cingular remnants extend-ing from the mesiobuccal groove, andanother extending mesially from the disto-buccal groove. The buccal and lingualenamel lines are fairly straight wherepreserved.
The LM2 crown is broken through themesial cusp tips and is missing its mesialportion. Its morphology is similar to thatdescribed for the right M2. A small, singledistal IPF is visible.
B: R mandible (alveoli I2, brokenroots I1, /C–P3, partial crowns P4–M2).Preservation. The right body is nearly com-plete from the symphysis to the mesial partof the M3 alveolus. It has many weatheringcracks and fragments of bone missing. Thealveolar margin is complete only on thelingual side of M2 and between M1 and M2.A large SI crack runs through the caninealveolus across the entire bone, separatinganterior and posterior portions that havebeen joined together. Cracking and abrasionalong the margins of the two pieces disruptthe contours of the bone, especiallymedially, but this region does not otherwiseappear distorted.
Lateral aspect. What is preserved of theextramolar sulcus opens anteroinferiorlyadjacent to M2. A strong lateral torus marksthe root of the ramus, and is located abouthalfway down the body from the alveolarmargin. There is a faint concavity just belowthe torus. The oblique line is obscured by
damage to the cortex, but on the left side canbe seen to be short, extending anteriorly towhere it terminates just below the mesialedge of M1. There appears to be a slighthollowing of the bone posterosuperior to themental foramen. The single mental foramenis large with abraded margins, but probablywould have been about 4 in diameter. Thedirection it opened cannot be determined. Itis situated beneath the distal root of P3 justbelow the SI midpoint of the corpus. Antero-superior to this, the contours of the bone flareout slightly toward what would have been afairly prominent canine jugum, although thebone overlying the canine root is largely miss-ing. Only a weak marginal torus is apparentwhere the base is broken posteriorly. Thecorpus does not appear to deepen posteriorly,although much of the inferior margin is lost.At the symphyseal midline the height is esti-mated at 43, posterior to I2 it is 42, /C 44, P3
43, and P4 42·3.Posterior aspect. The postincisive planum is
hollowed ML and almost straight AP, untilit curves into the superior torus at about thelevel of the P3 roots. The superior torus isstrong and high, located almost halfwaytoward the base from the alveolar rim. It isseparated from the strong inferior torus by ashallow and rounded genioglossal pit. Theinferior torus is as posteriorly extensive asthe superior one. The area of the sublingualfossa is obscured by a large crack.
Medial aspect. The symphysis is long andlow, sloping posteriorly with its long axisrunning 135� to the alveolar margin. Itsexternal contour is uniformly convex, arch-ing smoothly around its inferior margin. Themylohyoid line is partly obscured by cracksand spalling, but appears as a rounded ridgerunning anteroinferiorly from the posteriorbreak. The alveolar prominence is fairly flatinferior to P4, gently curving into a shallow,anterior subalveolar fossa that appears torun parallel to the base.
Basal aspect. At the short section of thebasal contour below the mandibular tori is a

282 . . ET AL.
pronounced concavity leading to a strongdigastric tubercle situated about 5 from themidline. Because bone is missing here, anycrest along the mandibular base is obscured.The mandibular body is noticeably evertedbetween M2 and M3. The small preservedportion of the mandibular base is smooth,forming a rounded ridge along its inferior-most extent. There is a small, longitudinalridge that begins near the anterior-mostpreserved portion of the base.
Occlusal aspect. Corpus thickness can beestimated at P3 (22), at P4 (20), at M1 (21),and at M2 (23), but these estimates arerough due to spalling and weathering.
Teeth. On the right no canine crownremains, but pieces of the right canine rootare present for most of its length, especiallylingually. The actual length of the R/C rootis 24·4. Apart from the Dmr and distal faceof the P4 that are almost completely pre-served, the P4 and M1 look like theirantimeres.
The M2 has 4 main cusps, of which theMed is the dominant, a small Hld, and a C6
made up of 2 cuspules. From largest tosmallest the cusp areas are: Med, Prd, Hyd,End, Hld, C6. There are wear facets andpolish on all cusps, and the buccal cusps areslightly flattened but with no dentine expo-sure. The Mlg runs from the shallow andpoorly delimited Fa to the crenulated Fc,where it disappears. Two secondary fissuresrun mesially from the tip of the Med towardsthe Fa and the Mlg, and others run pos-teriorly from close to the tip of the Medto the Fc and lingual groove. Secondaryfissures also run similarly down the distalcusps into the Fc, the lingual groove and theFp. The lingual and mesiobuccal groovesare continuous across the crown. Thelingual groove extends onto the lingual faceand terminates near the cervix. Themesiobuccal groove deeply incises thebuccal face, and terminates at a protostylidthat continues mesially around the Prd anddistally as a cingulum around the Hyd as far
as a break on the MD corner. A small part ofthe mesial IPF is visible, but most is missingbecause of the broken chip of tooth. On thedistal margin of the tooth an irregularlyshaped distal IPF has two distinct parts.Both M3s were evidently erupted and incontact with the M2s.
C: LI2. This I2 is complete except forchips of enamel lost along the edges of abreak running across the crown, and somebone along a crack across the root. Thetooth is lightly worn, exposing a ribbon ofdentine about 5 long and 0·5 wide on themesial incisive edge. The lateral two-fifths ofthe edge slopes downwards, away from theincisive wear plane at about 45�. There ispolishing in the proximity of the occlusaledge on both the lingual and labial faces.The mesial margin of the tooth is fairlystraight, and the tooth expands slightly inwidth mesially from the cervix. The distalmargin flares out more substantially to makethis tooth 8·5 MD. The labial surface of theI2 is concave, with a hint of a distolabialgroove. The mesial surface has a reciprocalfacet for the central incisor. The lingualsurface is concave in both major directions.The basal tubercle is present but notaccentuated. The distal IPF is visible but notclearly demarcated. It is about 2·9 by 2. Theroot is strongly compressed MD with longi-tudinal grooves on both mesial and distalfaces. It is 16 long measured on the mesialsurface from the incisal angle to the root tip.
D: RI2. This specimen preserves thelingual half of the crown; most of the labialsurface is broken away. In its preservedparts, this tooth is the mirror of the left one.
E: RP3. This partial crown is missing awedge from the buccal face extending fromthe cervical margin most of the way to theapex, and another piece from the distalmargin. Enamel is preserved to the cervicalmargin only on the distolingual corner.There is no dentine exposure, but the ridgesand crests show polishing. The surface ofthe tooth shows perikymata which are worn

283AUSTRALOPITHECUS ANAMENSIS
away apically, most likely as the result ofantemortem wear. The tooth is oval in oc-clusal outline with a more tightly convexdistolingual portion. It has one central cusp.The mesial occlusal ridge joins a short Mmrthat encloses a small Fa. The transverse crestruns lingually for about 5 before it bifurcatesand sends a mesial branch to the cervix and adistal branch to join a Dmr. The distal oc-clusal ridge runs distally to join a short Dmrthat encloses a larger Fp. A portion of thedistal IPF can be seen on the Dmr adjacentto the break of the missing fragment. Notrace of the canine contact facet is apparent,although that area of tooth is preserved.
F: RM3. This crown has lost the enamelaround the entire cervical margin; slightly solingually, about halfway up mesially andbuccally, and all the way to the occlusalsurface distally. No root is preserved. Theocclusal outline is a rounded rectangle, withwhat would have been a convex posteriormargin. It has four main cusps, of which thePrd is the largest and the others are subequalin area. All cusps are traversed by secondaryfissures, and accessory cuspules are visible inthe Fc. As on the M2, the mesiobuccalgroove is continuous across the central basinwith the lingual groove. The Mmr extendspartially around the Med. There are threedevelopmental pits around the Med andwear facets on both mesial cusps and on theHyd. The facet on the Hyd exposes dentine,and is associated with a large chip of enamelthat was broken from the buccal side of thiscusp during life. This happened in life,rather than post mortem, because this is theonly cusp with dentine exposure in a facet,its antimeric cusp has almost the full thick-ness of enamel left and the edges of thebroken enamel defect are smoothed over. Ifthis had occurred by post mortem rolling itwould also have abraded the edges of thewear facets, but these are still sharp. Asfor the M2, the Mlg terminates at the Fcafter meeting the continuous lingual andmesiobuccal grooves. The lingual groove
extends to the break close to the cervix. TheV-shaped mesiobuccal groove deeply incisesthe buccal face.
G: LM3. This is the buccal and distal por-tion of the left M3 crown. It is similar to itsantimere. The buccal surface of the toothretains almost the entire extent of its enamel,and has a less pronounced mesiobuccalgroove than its antimere. It shows a proto-stylid that extends mesially from the mesio-buccal groove. Distally, a cingulum bordersthe Hyd. Many cuspule-like vertical striationsindent the buccal and distal faces. Both pre-served cusps show wear, extending from theHyd into the central basin.
H: LI1 crown. This crown is missing thecervical half of the lingual surface. A tinyportion of root is preserved on the mesio-labial corner. The crown is worn, exposinga ribbon of dentine about 0·5 wide. The facetfor the other I1 is an elongated oval, measur-ing about 4·0 long SI by 1·4 LaL, withweakly defined edges. The I2 facet is clearerand roughly circular, measuring about 2 indiameter. The labial surface is smoothly con-vex, with well-marked perikymata on the cer-vical half, but these have been worn off theincisive half. The lingual surface is almostflat to slightly concave, with light undulatingridges running from the wear plane towardsthe marginal tubercle. There are roughlysymmetrical mesial and distal groovesseparating similarly sized mesial and distalridges. The mesial and distal margins aresubparallel, so the tooth has a roughlyrectangular outline in lingual view, and doesnot splay out markedly from the cervix.
I: 7 crown fragments (not figured).These fragments include parts of twoincisors, a canine, and three molars orpremolars. One is indeterminate.
KNM-KP 30498—L and R maxillaryfragments and associated maxillarydentition (Figure 7: G–L not figured).
This specimen was discovered washingdown the southern slope of a small hillock

284 . . ET AL.
Figure 7. KNM-KP 30498 associated maxillary dentition. (A) in labial, lingual, mesial and occlusal views,(B) in labial, lingual, mesial and occlusal views, (C) in distal and occlusal views, (D) in occlusal view,(E) in mesial and occlusal views, and (F) in occlusal view.
65 m from the site of discovery of the typemandible, KNM-KP 29281 (Wynn, 2000,Figure 2). Apart from the initial isolatedteeth, the majority of these teeth wererecovered through screening. SpecimensKNM-KP 30505, which includes a molargerm, and KNM-KP 30942, tooth frag-ments, were also recovered in the screening.
A: L premaxillary fragment (I1, rootI2). This is a complete I1 crown and root,along with maxillary bone lateral to it hous-ing just over half of the I2 root. The I1 crown
is cracked and weathered, but the mor-phology is preserved. On the labial andmesial faces, cracks runs through the crownvertically from the root widening to about0·2 at the occlusal surface. The crown isheavily worn almost to the basal tubercle.Dentine exposure is 2·5 at its maximum LaLby 9·3 MD. Both the mesial and distalmargins of the crown flare out strongly fromthe cervix almost symmetrically, but thedistal margin slightly more so. The basaltubercle is swollen, and the Mmr and Dmr

285AUSTRALOPITHECUS ANAMENSIS
fade into it. There are traces of three lingualgrooves running into the tubercle from thewear facet. Both IPFs are indistinct due toweathering, but the distal one can be seenset distolingually on the Dmr. Mesial anddistal enamel lines are strongly concavetowards the occlusal surface. The lingualenamel line is mildly convex, but the labialone is damaged. The root measures 17 longfrom the cervical margin labially. The root is7·7 LaL and 6·8 MD at the cervix.
Only 12·5 of the I2 root is preserved fromits apex, and its original length is estimatedto have been 18. It is oval at its brokensection, with only a faint distal groovetraversing its distal side. It measures about5·3 MD by 7·7 LaL at its break. Its apex isblunt, and the root appears to convergeupon the I1 root apically.
B: RC/. This tooth is missing only theapex of its root, and the enamel at its distaledge obscuring any Dmr. The crown isweathered, and has a 0·2 wide crack runningLaL through its center. It flares out mesiallyand distally from the root, below its archedcervical margins. The crown is worn acrossthe tip, exposing a tear-dropped area ofdentine measuring 5·2 MD by 2·8 LaL thatslopes distally and lingually. The crown isasymmetrical, with the Mmr separated fromthe main cusp by a shallow marginal groove.This pronounced marginal ridge and mar-ginal groove continue across the occlusalsurface and faintly onto the labial surface,forming a prominence and concavity,respectively, in the occlusal outline in labialand lingual views. The labial surface is uni-formly convex, with perikymata exposednear its cervical margin. The lingual surfacehas a swollen cervical tubercle that is con-tinuous with the shallow mesial and distalmarginal grooves, and with the pronounced,rounded median lingual ridge that is locatedtowards the mesial edge of the toothimmediately adjacent to the mesial marginalgroove. Distal to the median lingual ridge,and inferior to the cervical swelling the
lingual face of the tooth is flat. There is nodiscernible facet for the I2. The root is 9·1MD by 7·5 LaL at the cervix. The premolarfacet cannot be seen on the canine becauseof the missing enamel in this area.
C: R maxillary fragment (P3). This is acompletely preserved crown, with partialroots surrounded by some maxillary bone.Part of the canine alveolus is preservedmesially, indicating the position of thispremolar in the maxilla. Much of the speci-men is weathered, obscuring the occlusalmorphology. The crown has two tiny expan-sion cracks running across its partially wornocclusal surface that would affect measure-ments minimally. It is a bicuspid tooth witha tall, conical Pa that occupies more thanhalf of the occlusal area, and a low roundedPr. There are indistinct mesial and distalgrooves adjacent to the rounded Mmr andDmr respectively. Wear has flattened bothcusps and there is a tiny area of dentineexposure on the tip of the Pa, but none onthe Pr. Ridges and crests are polished bywear. Both buccal and lingual faces slopeinwards towards their tips, but the buccalone slopes more strongly. The lingual sur-face is smoothly convex all around, but thebuccal face has slight mesial and distalgrooves and ridges. There is a strong mesio-cervical enamel extension of the buccal face.The crown flares out mesially and distallyfrom its root about equally, and bulgesjust below the cervical margin labially andlingually. The mesial IPF is set midway onthe mesial border; it is small but indistinctdue to weathering. The P3 has two roots,with distinct buccal and lingual roots. Thelingual one is missing only its apex, andmeasures 14·2 long as preserved, which isalmost all of its original length. It angleslingually from the crown and somewhatdistally. Only about 2·2 of the buccal root isleft. Part of the alveolus for P4 is visible, andabout 5 of the P4 root near its apex isadhered to the bone surrounding the P3
roots.

286 . . ET AL.
D: L maxillary fragment (M3). This isa fragment of maxilla with no preservedcortex, and the lingual two-thirds of a M3
crown with its roots embedded in themaxillary bone. The surface of the tooth isquite weathered. Enough of the crown ispreserved to show that it is an almostsymmetrical mirror copy of the other one(E), although the details of the cuspules arenot identical.
E: R maxillary fragment (M3, rootM2). This is a complete M3, except forfragments of the enamel broken from thecervical margin lingually and distally. It hasa single root that is encased in maxillarybone on all but its distal sides. A sliver of thedistolingual root of M2 is preserved mesially.The occlusal outline of the M3 is a roundedtriangle, with the blunt apices mesio-lingually, mesiobuccally and distally, so it isformed by the two mesial cusps and anirregular set of distal cuspules. The occlusalsurface is lightly worn with flattening andpolishing of mesial cusps and ridges. The Pais the larger of the two mesial cusps. There isa small Fa 7·2 wide cut into by the only wearfacet that is across the mesial part of thetooth. No dentine is exposed. Cuspules arearranged in a series of bead-like structurescentered on a Fc. The lingual face of thecrown is corrugated by four distinct verticalridges and adjacent grooves. The M3 root isin the shape of an irregular cone that isabout 11 long. The mesial IPF is oval, andmeasures 3·3 BL by 1·9 high.
F: L maxillary fragment (M1, rootsP4). This specimen consists of a completeworn and cracked M1 crown, its brokenroots, most of the P4 roots and interveningmaxillary bone. The M1 crown is complete.Its occlusal outline is a rounded rhombus,with the buccal cusps set mesial to thelingual ones. From largest to smallest thecusp areas are Pr, Pa, Me, Hy. Wear hasflattened all cusps. Dentine is exposed on allcusps except the Me. The Pa dentine pit iscircular and measures 0·7, the Pr one is
tear-dropped measuring 1·9 by 2·3, and theHy one is a tiny circular one of 0·2. Thecrown has a strongly sloping lingual side andmore vertical buccal side. The buccal grooveis obscured on the buccal surface by a crackrunning through the crown. A lingual groovesets off the Hy. Not much remains of thegroove system except for the trace of thebuccal groove extending along the occlusalsurface almost to the center of the crown.There is no trace of a Fa, and the Fp ispresent as a small V-shaped depression setabout 4 mesially from the distal margin. Themesial IPF is about 6 wide but obscured byadhering matrix that retains the impressionof the distal P4 enamel.
G: RI2. The root of this tooth is almostcomplete, but the badly cracked andweathered crown preserves only a sliver ofenamel on the distobuccal corner and thelingual portion near the cervical margin.The latter has a swollen basal portion, andthere is a trace of the Dmr before the break.The root is weathered and cracked withsmall pieces missing. It measures 7·7 LaL by4·8 MD at the cervix, and it is 15 long. TheLaL widest point on the root is near itsmidpoint, where it measures about 9·3 by4·5.
H: M fragment. This is a fragment of amolar, possibly the mesial side and adjacentocclusal surface. It is worn nearly flat, with agroove separating two partial cusps. Thebeginnings of a root flare strongly away fromthe crown. At the edge of the preservedocclusal margin, the enamel measures amaximum of 1 thick.
I: RP4. This is the buccal half of a crackedcrown with a short fragment of root,measuring a maximum of 5 from the cervix.Weathering has obscured details of thecrown, but it appears that the buccal facehad distinct mesial and distal grooves. Thebuccal face is also weathered and cracked,but appears to have had mesial and distalgrooves running towards its occlusal margin.What appears to be the buccal half of the

287AUSTRALOPITHECUS ANAMENSIS
Figure 8. KNM-KP 30500 partial mandibular denti-tion with associated mandibular bone, in occlusal view.
mesial IPF is visible. The distal IPF is large,and although only half is preserved, is abroad band 2·1 high across the distal surfaceof the tooth. No dentine is exposed.
J: LP3. This is most of the distal halfof the LP3 crown. Morphology is like theright P3.
K: LC/. This is an 18 long, badly crackedand weathered root with neither the cervicalmargin nor apex preserved. Its dimensionsare obscured by damage.
L: Molar root. This is a badly cracked,weathered and spalled molar root withneither end preserved, measuring 14·9 longas preserved.
KNM-KP 30500: Mandibular fragmentsand associated mandibular dentition(Figure 8; H–L not figured). This specimenwas recovered as a surface find andadditional pieces were found throughscreening. There are no associatedspecimens.
A: L mandibular fragment (M1 andM2). This specimen consists of part of amandibular body extending to a point justinferior to the tips of the molar roots, alongwith M1 and M2 crowns. The cortex of thebody is badly weathered. Laterally it swellsbelow M2 to the lateral torus at the pointwhere it extends posterosuperiorly to meetthe ramus. The internal margin of the bodyis mildly concave AP.
The LM1 is complete except for smallpieces of enamel missing from the edges ofcracks running through the surface. Itssurface is weathered, but most morphologyis preserved. In occlusal outline it is arounded square with an expanded disto-lingual corner. The Med is the highest andlargest, and the central basin is large. Fromlargest to smallest the cusp areas are: Med,End, Prd, Hyd, Hld. The tooth is wornon all cusps, with flattening of the buccalcusps and the Hld which all have smalldentine pits measuring less than 1 in diam-eter. The Fa is represented only by small,
irregular pits, and the Fp by a 2·4 long cleftdelineating the Dmr. The lingual surface ofthe tooth is quite vertical, but the buccal oneslopes considerably. A distinct protostylidborders the Prd. The lingual groove runsacross the central basin, and is largely con-tinuous with the mesiobuccal groove, whichends in a distinct pit at the protostylid. Ashort distobuccal groove indents the buccalface distally. A distinct distal IPF is about4·8 wide BL.
The LM2 is also slightly weathered, andhas cracks crossing it diagonally in bothdirections. Its occlusal outline is a roundedrectangle. Polishing indicates light wear butno dentine is exposed. From largest tosmallest the cusp areas are: Med, End, Prd,Hyd, Hld. Like the M1, the central basin is

288 . . ET AL.
large and mildly crenulated, and is dividedby the continuous lingual and mesiobuccalgrooves. These grooves are more evidentthan on the M1 because this tooth is lessworn. A distinct shelf-like protostylid,bordered distally by the mesiobuccal groove,delimits the Prd. The distobucccal grooveincises the buccal face distally, terminatingat a V-shaped notch. The cingular remnantsalong the buccal face are stronger than onthe M1. The Fa is again a series of 4 pits, setbehind the Mmr, and the Fp is a short cleftdelineating the Dmr. The mesial IPF isabout 4·8 wide BL. Weathering obscuresany traces of a distal IPF.
B: LM3. This tooth is complete except forenamel missing from the distal and lingualsurfaces, and it has cracks running throughit. It is morphologically similar to the RM3
(E), which is more complete.C: R mandibular fragment (M1). This
specimen preserves a badly weathered, smallsection of the lateral surface of the mandibu-lar body around the M1 roots. The fragmentmeasures up to about 17 AP and 12 SI. TheM1 crown is missing its mesiolingual corner,and is cracked. The root apices are brokenaway. Wear has flattened the two buccalcusps and exposed small islands of dentineon the Hld. The roots are broad and plate-like narrowing apically and distally inclined.A marked furrow indents the mesial face ofthe mesial root. The rest of the morphologyresembles the LM1 (A).
D: RM2. This tooth is cracked andweathered but the crown is complete. Theroots are preserved, and surround somemandibular bone. They are broad andplate-like, and bifurcate at the tips. Theroots incline distally. Crown morphologyresembles the LM2 (A).
E: RM3. This tooth is complete except forenamel missing from its distalmost surface.Cracks run through its weathered crown.The occlusal outline is more triangular thanthat of the other molars, resembling anelongate rectangle mesially, that tapers to a
distal apex. There are 5 main cusps, thelargest and most salient of which is the Med.From largest to smallest the cusp areas areMed, Prd, Hyd, End, Hld. Although thereis no dentine exposure there appears to bepolishing on all cusps and some flatteningof the buccal cusps. A small protostylidborders the Prd, terminating distally at thedistinct mesiobuccal groove. The Facomprises a series of distinct pits, as for theM1 and M2, but in this case the pits arejoined by a meandering furrow. The Fc isextensively crenulated, and comprised of aseries of fissures and inflated ridges thatradiate from the cusp tips to the basin. TheHld is cracked and weathered, but it appearsto be partly separated from the other cusps.The lingual surface of the crown is steep,and the buccal surface sloping. The mesialroots are broad and plate-like. The singledistal root is roughly triangular in section,following the contour of the posteriormargin of the crown, with longitudinalfurrows on either side, particularly thebuccal one. The apices of the roots arebroken.
F: LP3. This crown is almost complete,but is missing its cervical enamel along itsdistal margin up to the Dmr. Parts of theroots are present, extending about 3–4 fromthe cervix inferiorly on most sides, but toonly about 1·5 lingually. Large crackstraverse the crown, with the largest missing achip of enamel at the tip of the main cusp.The occlusal outline is a triangle with apicesmesially, distolingually and distobuccally.The tooth is weathered, but does not appearto have been worn although slight polishingis apparent on the Prd. There is a singlecusp. The transverse crest bifurcates into amesial branch that runs mesially to join theMmr and encloses the Fa. The distal branchjoins the Dmr to enclose a larger, deeper Fp.The Fp has several small furrows emanatingfrom the center, but the Fa is relativelysmooth. There was a single mesial root anda plate-like distal root that already shows

289AUSTRALOPITHECUS ANAMENSIS
two separate pulp cavities indicating that itprobably divided lower down the root. Theenamel is weathered, obscuring any possiblecanine facet. The missing distal enamelobscures any potential distal IPF. Aweathered hypoplastic groove traverses thebase of the crown along its buccal side.
G: LP4. This crown is complete exceptfor part of the lingual face that is broken andhas enamel missing along the distal surface.A 1 wide piece of root is preserved mesially.This is a bicuspid tooth, with the Prd largerthan the Med. The occlusal surface isweathered, and only slight wear is apparentin some flattening of the Med and lightpolishing of ridges. The smaller, smooth Fais set higher than the larger Fp, which hassmall fissures and ridges running into itscenter from the two cusps. The mesial IPF isoval, and about 3·5 BL by 2·1 high. Thebuccal face shows two indistinct mesial anddistal ridges.
H: I root. This is a piece of incisor rootthat is compressed MD. It is 17·5 long aspreserved. No crown is preserved, and theroot tips are missing. Near its center it is 8·7LaL by 4·1 MD.
I: R/C. This is a fragment of a rightcanine, preserving the distolingual corner ofthe crown with some root attached. The tipof the crown is broken off. The tooth isweathered. Little can be said about its mor-phology, although on the lingual face, adistinct distal marginal groove is borderedby a Dmr.
J: C root. This is a fragment of canineroot split longitudinally. It is cracked andweathered, and preserves little morphologybut shows the pulp cavity.
K: L/C. This specimen is a broken canine,preserving the distobuccal corner of thecrown and about 7·5 of the adjacent root.The entire specimen is traversed by a large,longitudinal crack and is weathered. Theroot appears to have a slight transversebulge, but this is greatly enhanced dueto postdepositional taphonomic processes.
Although broken away, the shape of theremaining crown indicates that there was adistal cuspule or strong ridge. The occlusaloutline takes a sharp turn between buccaland distal faces. A distal IPF is apparent.
L: 7 mandible and root fragments.These are small fragments of mandibularbone and tooth root that cannot be reliablyassigned to particular teeth.
KNM-KP 30502: Associated mandibulartooth fragments (Figure 4; A–C, E, J andK not figured). Apart from the initial frag-ment, these tooth fragments were recoveredthrough screening.
A: LP4. This is the lingual part of a P4
crown with partial basins preserved. There isa small Med separated from the broken Prdby a notch in the transverse crest. The Fa ishigher and smaller than the Fp. The Fp hassmall ridges extending into it from eachcusp. The lingual surface of the crown isevenly convex SI.
B: RM1. This is the mesiobuccal cornerof a RM1 crown, preserved not quite downto its cervical margin. The entire Prd ispreserved, along with the mesial and buccalparts of the Med and part of the centralbasin. The Med was the higher of the twopreserved cusps. There is dentine exposureon the Prd, which is circular and 0·8 indiameter. The Med has a groove runningfrom its tip into the Fa, which is bounded bya mild Mmr. There is a trace of a smallprotostylid on the mesiobuccal corner andposteriorly as a distinct but short shelforiginating at the base of the buccal notch.The mesiobuccal groove extends from thebuccal notch into the Fc. The buccal side ofthe tooth slopes strongly toward the occlusalsurface. There is a tear-drop shaped mesialIPF 5 wide by 1·6 high placed centrally onthe mesial surface of the crown.
C: RM2. This is a wedge-shaped piece ofan M2 crown preserving the buccal side intothe central basin, but not all the way to thecervical margin. Half of the Prd, all of the

290 . . ET AL.
Hyd, and most of the Hld remain. Faintwear facets are visible on the Hyd. It has astrongly sloping buccal side, which iscracked and slightly weathered. The deepmesiobuccal groove extends from the buccalnotch where there is a cingular remnant toFc. Secondary fissures that pass into the Fcform radial ridges emanating from the cusps.
D: RM3. This crown of a germ preservesalmost the entire unworn occlusal surface,except for the mesiolingual slope of theMed, and it is missing its cervical enamel. Alarge transverse crack divides the mesialportion of the crown. The tooth is a roundedrectangle in occlusal view, with five maincusps. Numerous secondary fissures forminflated ridges, and smaller cuspules that fillthe basins. The mesiobuccal groove is con-tinuous across to the crown with the lingualgroove, terminating buccally at a proto-stylid. The deep Fa is 3·8 wide and demar-cated by a Mmr. A Mlg runs from it to thedeep, sinuous Fp that is about 3 wide. Thebuccal side of the tooth slopes strongly.
E: LM3. This is most of the unworncrown of a germ, missing the mesial edgeand mesiolingual corner, as well as someenamel along the cervix. Although details ofthe central basin cuspules also differ slightly,this tooth is morphologically similar to theright one. Unlike the right M3, the Fp isbroad and shallow, but has two small cus-pules bounding it mesially.
KNM-KP 30505: Partial molar germ(Figure 4). This specimen was discovered inthe screening for the adult isolated teeth,KNM-KP 30498. The tooth is clearlyfrom another juvenile individual. Severaladditional tooth fragments, KNM-KP30942, that do not appear to belong toeither of these specimens, were alsorecovered.
This is a partial mandibular molar germwith the remains of 2 half cusps. It is prob-ably the mesial half of the crown because thefovea is so clearly delineated. It is probably a
left, because the slope of the buccal portionis stronger than the small slope of the lingualface. There are numerous secondary fissuresand accessory cuspules on the occlusal sur-face. This germ is associated with a smallpiece of deciduous molar root.
KNM-KP 30942: Five molar fragments(not figured). These fragments were dis-covered in the screening for KNM-KP30498. As they cannot be associated withany certainty to this specimen or the toothgerm, KNM-KP 30505, also recovered inthe screening, they are given a separateaccession number.
These are five molar fragments. Each haspart of the crown. The largest appears to bemost of the mesial or distal face of a molar.Little informative morphology is preserved.The dentoenamel junction is clear on sev-eral, but because none preserve unworncusp tips standard enamel thicknessmeasurements are not possible.
KNM-KP 31712: Associated juvenilemandibular and dental fragments(Figure 9; B, C, J and K not figured). Apartfrom the initial mandibular fragment, thisspecimen was recovered through extensivescreening of a steep slope and gulley. Noadditional specimens were recovered in thescreening. KNM-KP 31714, 31728 and35842 were recovered from the vicinity ofthis site, but are not considered to representthis individual.
A: R mandibular fragments (dm1,roots dm2, P3 & P4 in crypts). This speci-men includes the medial cortex of a rightmandible measuring 16·2 SI by 23 AP, apartial crown of the Rdm1, roots of dm2,and the crowns of P3 and P4 in their crypts.The mesial margin of the M1 alveolus isvisible on the posterior side of this specimenand part of the canine alveolus just belowthe P3 germ. The lingual alveolar margin ismissing, leaving a straight contour that is aminimum of 2·1 lingually from the tooth

291AUSTRALOPITHECUS ANAMENSIS
Figure 9. KNM-KP 31712 associated juvenile dentition. (G), (E), (F) and (H) in lingual views, (I) inlingual, labial, distal and occlusal views, (D) in occlusal, lingual and distal views, (A) in occlusal andmedial views.
roots. In occlusal view, the anterior end ofthe mandibular bone curves medially towardthe symphysis along its inferior edge, but thesuperior edge is almost straight. The pre-served part of the alveolar eminence issmooth, with a slight bulge adjacent to thedeveloping P4. Laterally, the buccal sides ofthe deciduous molar roots are exposed. Theinferior margins of the developing P3 and P4
crowns are visible tucked within the roots ofthe dm1 and dm2 respectively. Just inferiorto the P3 crown, the medial wall of what isprobably the /C crypt is preserved for 5 APby 4·3 SI.
The dm1 crown is broken. Most of theenamel is broken or spalled off, but someremains on the mesial, distal, lingual andocclusal faces. A matrix-filled crack, 0·5
wide, runs transversely BL through themesial cusps. The occlusal outline is anarrow, elongate ovoid oriented MD witha fairly straight lingual side and roundedmesial margin. Some of a high, pointed Medis preserved just mesial to the MD midpointof the lingual occlusal margin. Also pre-served is the lingual side of the Hyd, whichwas worn to expose a pinpoint of dentine.The floor of the Fa is lingual to the MD axisand terminates distally in a Dmr that issmooth from wear. A basal bulge of thecrown is evident on the mesial and lingualsides. Both roots of this tooth run almoststraight inferiorly, but the distal one anglesslightly distally toward its apex.
The dm2 crown is completely brokenaway, but it would have been larger in all

292 . . ET AL.
dimensions and relatively broader BLthan the dm1 crown. Its mesial root runsinferiorly, hooking distally at its apex andthe distal root angles distally.
B: R mandibular fragment. This is thelateral surface of mandibular bone, whichalmost fits on A, but cannot be joineddirectly. It comprises two pieces joined at alarge, uneven crack running vertically pastthe posterior edge of the well preservedmental foramen, and is up to 1 wide. Thefragment measures 18·8 SI and 37·6 AP.Posteriorly, the beginning of the extramolarsulcus appears medial to a gently roundedlateral torus that becomes sharper as itbegins to form the base of the ramus at itsposterior extent. Little or no oblique line isevident. The alveolar margin is broken, butadjacent to the dm1 and dm2 it appears to beclose to its original contours, as the internalcontours of this fragment match the con-tours of the deciduous molar roots of A. Anapparent crypt is present mesially, possiblyfor the permanent canine. The anteriorbreak of this fragment slopes anteriorly andinferiorly and the posterior break posteriorlyand inferiorly. The anterior portion of thefragment is flat SI but begins to curve medi-ally at its superior edge and laterally along itsinferior edge. The posterior part is verticallyconvex. There is a shallow depressionimmediately superior to the mental foramen.The mental foramen measures 3·1 by 2·0,its long axis running posteroinferiorly toanterosuperiorly.
C: RI1 or I2. This is a crown and root,with mandibular bone adhering to the distalportion of the root. The distal part of thecrown is broken and largely missing, and thebroken face is vertically concave. Enamel iscompletely missing from the lingual face,apart from a thin strip along the mesial edge.The root is largely complete but hiddenapically by mandibular bone adhering to it.A flattened surface at the inferior and mesialcorner of this adhering bone may be the wallof a permanent incisor crypt. The crown is
slightly worn across the occlusal surface.The labial face is vertically convex, andmarked by faint vertical grooves. It exhibitsno basal swelling. A small mesial IPF isvisible mesially, running 1 down from theocclusal edge. The root measures 4·4 LaLby 3·3 MD near the cervical margin.
D: Ldm2. This is the crown with up toabout 4 of the root all around. Enamel ismissing from the mesiobuccal and mesio-lingual corners up to the cusp tips andacross the mesial face except for a narrow,vertical isthmus, but the floor of the Fa ispreserved. A crack runs up the lingual sideof the End to its tip. The occlusal outline isa rounded rectangle. The size of the brokenPrd cannot be assessed but from largest tosmallest the areas of the remaining cuspsare: Med, Hyd, End, Hld. The two lingualcusps are taller and slightly more distallyplaced than the buccal ones. All show somewear, with the buccal cusps slightly flattenedand a pinpoint of dentine is exposed on theHyd. A large Fa is bounded by a large butbroken Mmr, and separated from the mildlycrenulated Fc by a sharp crest connectingthe two mesial cusps. This crest is notchedby the broad Mlg. A small Fp sits betweenEnd and Hld, inside a Dmr. The lingualgroove is distal to the mesiobuccal one andnotches the occlusal margin as it continuesto the cervical margin. The mesiobuccalgroove is pronounced and terminates in adeep notch close to the superior margin ofthe buccal face, but inferior to this the twolobes of this face continue to be divided bya shallow furrow. The distobuccal grooveends in a small pit. The lingual and buccalfaces are both bilobate, but the buccal one ismore sloping towards the occlusal surfacethan are the lingual and distal sides. Amesial IPF is just visible on the isthmus ofenamel preserved. None is present distally.
E: I1. This is a partial unerupted crownmissing its left edge in lingual view. It is tooincomplete to attribute to the right or leftside. Three mammelons mark the occlusal

293AUSTRALOPITHECUS ANAMENSIS
margin, which are the continuations of faintlingual ridges. The lingual side is verticallyconcave and only mildly so transversely.The labial surface is smoothly convex, andmarked by faint vertical furrows and ridges.Perikymata are apparent on this surface.
F: I1. This fragment is also too incom-plete to attribute further. It was recovered intwo pieces that have subsequently beenjoined along a vertical crack that divides thecentral portion from the left side in lingualview. Its morphology resembles E.
G: Mandibular fragment (LI2 incrypt). This fragment consists of theexternal cortex just lateral to the symphysis,about 13·3 SI and 14·9 ML. The I2 is in itscrypt, and the anterior wall of the /C crypt isvisible. The preserved anterior cortex issmooth and flat, flaring slightly laterally justat its inferior edge.
The I2 crown is incomplete. Verticalcracks that run up the right and left sidesfrom the crown base diverge towards themesial and distal margins at the occlusalsurface. Most of the mesial half of theocclusal margin is broken away, except atthe mesial corner. On the preserved distalportion, two small mammelons are present.The distal corner of the occlusal surface isrounded, while the mesial one is sharp.Vertically elongate mesial and distal lingualfossae are visible. The lingual face is concavevertically, except for a ridge along theocclusal surface, but horizontally is onlyslightly so. The labial face is convex horizon-tally and vertically and perikymata arevisible. The superior half of the labial crownsurface is marked by small vertical groovesleading towards the mammelons on thepreserved part. An area 6·8 SI by 6·0 MLof the crypt for the L/C is visible at themedial edge of the posterior side of thisfragment.
H: RI2. This triangular shaped fragmentof the unerupted RI2 preserves the unwornocclusal margin, but both mesial and distalsides are missing. The mesial and distal
breaks, as on the other incisors, are slightlycurved, widening from the base towards theocclusal margin. The occlusal margin curvesgently down towards its distal margin. Fourmammelons make up the occlusal margin,with faint vertical ridges running inferiorlyfrom them on the labial side. The labialsurface is convex SI and MD, and thelingual surface is concave in the samedimensions except for a ridge of enamel atits occlusal edge. As on the other incisors,perikymata are visible on the labial face.
I: R/C. This is most of an uneruptedcrown. A 0·4 wide crack at the apex splitsthe crown into mesial and distal halves. Theocclusal outline is strongly convex labially,and flat lingually. The crown is asymmetri-cal, with the apex situated slightly distal tothe LaL axis. The labial surface is smoothand tightly convex transversely, except for avertical flattening toward the apex. Themesial and distal labial grooves are weaklydeveloped. The distal margin of the labialsurface is longer than the mesial one. Themesial margin terminates in a sharp Mmr,5·4 from the crown apex that continues ontothe lingual face, angling inferiorly and dis-tally to the break, standing well out from thetooth surface. No distal basal tubercle ispreserved, as this crown is incompletelyformed. In lingual aspect the prominent,sharp, lingual ridge is eccentrically placedclose to and parallel with the distal margin,so that most of the lingual face is flat,measuring a maximum of 6·0 MD. Thedistal lingual groove is deep, narrow andV-shaped, and extends from the base of thecrown almost to its apex.
J: RM1. This is the unerupted crown withonly a 1 long piece of root preserved on themesial side. It has a crack running obliquelyfrom the mesiolingual corner of the tooththrough the tip of the Prd. Its outline is anelongated, rounded trapezoid with long axisrunning mesiolingually to distobuccally,with an inflated mesiobuccal corner andbilobate lingual and buccal sides. The Med

294 . . ET AL.
is slightly taller than the other cusps. Fromlargest to smallest the cusp areas are: Med,Prd, Hyd, End, Hld. The tooth was not inwear. The occlusal surface is extensivelycrenulated by secondary fissures that formenamel ridges and cuspules. The ovoidbuccal portion of the Fa is bounded by theMmr, which is incised by two shallowfurrows but barely separated from the Fc. Itcontinues lingually as a slit anterior to theMed. The Fp is a narrow pit, which incisesthe edges of the End and Hld. It is boundedby the Dmr, which has a pit-like slit along itsdistal side. The Mlg extends the length ofthe crown. The mesiobuccal groove isdeeper and more mesially placed than thelingual one. It terminates in a pit boundedby a cingular remnant less than halfwaydown the crown, which is confluentanteriorly with a ridge-like protostylid. Thisprotostylid, in turn, traverses the Prd toeventually meet the Mmr. The distobuccalgroove ends in a more restricted cingularremnant in the form of a small cuspule lessthan halfway down the crown. The lingualgroove extends to the cervical margin. Allsides of the crown are convex, but the buccalone is the most sloping.
K: LM1. This is the unerupted crown,with 1–3 of root preserved mesially. There isa crack running through the Fa from mesialto buccal that has a sliver of enamel up to1·3 wide missing from it along its mesialface. A second crack runs transverselythrough the distal cusps and is widest lin-gually, a maximum of 0·3 across. This toothis a mirror copy of J, except for some detailsof the groove system that differ.
L: Mandibular fragments. These areseveral additional fragments of mandiblethat were recovered with this specimen butare too fragmentary to be diagnostic.
KNM-KP 31713: R mandible with toothfragments (Figure 10; B and C notfigured). This was a surface find and subse-quent screening led to the recovery of the
two partial teeth. No additional specimenswere recovered.
A: Mandible—Preservation. This man-dible is preserved from the left I2 alveolusthrough an area close to the distal M2
medially and adjacent to the M2/M3 junc-tion laterally. Its surface is weathered andtraversed by many cracks, the most signifi-cant of which runs horizontally along theinferolateral side of the body and is about 1wide. Bone is missing from the anterior andlateral surface of the alveolar region fromthe level of the mental foramen anteriorly,exposing all four incisor alveoli and thebroken canine root. A small amount ofthe alveolar margin remains mesial to thecanine, but along the mesial sides ofthe incisor alveoli the bone is abraded, andthe alveolar margins are broken off alongboth sides of the premolars and molars. Thetooth crowns are almost all broken away,and the remaining parts of the teeth arecracked and split. Only the lingual half ofthe canine root remains for most of itspreserved length, exposing the pulp cavity,but the root is complete near its apex. Thebuccal P3 root is broken away. The buccalsurface of the P4 root is exposed where thebone is missing. The posterior break exposesthe mesial wall of the alveolus for the distalM2 root.
Anterior aspect. There is a mild basalincisura. The I1 alveoli are about 11·3 deep.Their inferior margins diverge at theirdeepest point.
Lateral aspect. The extramolar sulcusbegins next to M1. Anterior to it, therounded ramal contour continues as theoblique line, which arcs inferiorly and fadesbelow M1. The lateral torus is broad andblends smoothly with the base. The surfacebecomes slightly concave along its anteriorbreak, just posterior to the mental foramen.The mental foramen is located at thejunction between P3 and P4, a minimum of17 from the base. Only its posteroinferiormargin is visible, but this is quite sharp

295AUSTRALOPITHECUS ANAMENSIS
Figure 10. KNM-KP 31713 edentulous mandible. Top row, occlusal, basal and anterior views; bottomrow, lateral and medial views.
so it must have opened anteriorly and/orsuperiorly.
Posterior aspect. The weak superior trans-verse torus is difficult to discern because thebone is largely broken here, but it appears toreach posteriorly only to P3. The post-incisive planum is short and mildly hol-lowed. A broad and shallow genioglossalfossa has a small supraspinous foramen in it.The inferior transverse torus is stronger thanthe superior one, extending to the level ofthe junction between P4 and M1, and issharply convex superior to the mandibularbase. The edge of a genial tubercle ispreserved next to the break.
Medial aspect. Although this mandible isbroken to the left of the midline, symphysealcontours can be appreciated. The symphysis
is inclined sharply inferiorly and posteriorly,its long axis can be estimated to have beenset 142� to the postcanine alveolar margins.It is vertically convex except for a milddepression inferior to the incisor alveoli.The mylohyoid line is barely visible as aroughened line running posterosuperiorlyfrom the symphysis to the posterior extent ofthe bone. The alveolar prominence is broadand evenly rounded, and is continuouswith the weak superior transverse torus.A distinct, continuous subalveolar fossais present, tapering anteriorly below themylohyoid line.
Basal aspect. The basal contour is noteverted. It is rounded in the symphysealregion, becoming slightly sharper inferiorto the canine jugum. A digastric fossa is

296 . . ET AL.
evident, bounded anterolaterally by a small,sharp ridge, but its posterior margin isunclear.
Occlusal aspect. The canines and incisorsform a smooth arc anteriorly. Although thebone is missing here, preserved tooth rootsindicate that the canine jugum would haveextended as far or further laterally than didthe P3 jugum, so that the lateral alveolarcontour would have swung medially anteriorto the /C jugum.
B: R/C. This is a partial canine crownthat cannot be joined to its root in themandible. Most of the mesial side of thecrown is broken away. Enamel remains onlyon the labial and distal surfaces, but doesnot extend to the occlusal margin at anypoint, and is missing along its distolingualcorner of the crown to within about 3 of thecervical line. Many vertical cracks traversethe tooth.
The amount of wear cannot be deter-mined although dentine exposed by wear isvisible superior to the broken distal tubercle.The labial surface is smooth. About 4 of theroot remains on the labial side, and can beestimated to be about 10·3 by 7·5 near thecervical margin.
C: RM fragment. This is a badlyweathered fragment, probably the disto-lingual corner of a mandibular right molar.The End can be seen, with parts of the Medand distal marginal ridge present, along withthe depth of the Fc. The crown is notpreserved to its cervical margin. Wear isdifficult to assess due to weathering, butlittle had taken place because the disto-lingual cusp is still fairly pointed. There isno dentine exposure on the occlusal surface.
KNM-KP 31714: Ldm2 (Figure 4). Thisspecimen was recovered in the vicinityof, but some distance from, the juvenilespecimen, KNM-KP 31712. No additionalfragments were found in the screening, butKNM-KP 31728 and 35842 were recoveredfrom higher on the slope. This specimen
could conceivably be associated with one orother of these specimens.
This is a weathered partially worn crownthat has almost the entire occlusal surfacepreserved and up to 5·6 of the distal root.The mesial face is missing, as is the enamelalong the cervical margin of much of theother sides. An oblique crack runs throughthe apex of the Prd and just mesial to theHyd tip. The crack is widest (0·4) at thePrd. The distal cusp has its apex chipped off
and the margins weathered. The occlusaloutline is a rounded rectangle with a convexdistal margin. Small dentine pits areexposed on all five cusps. From largest tosmallest the cusp areas are: Med, Prd, Hyd,End, Hld. The lingual cusps are higher andmore pointed, and set distal to the buccalones. The four main cusps all have smalldentine pits exposed. The broken Fa is abroad ovoid with crenulated sides and floor.The Fc is large, and positioned distal to theMD midpoint and lingual to the BL mid-point of the crown. The Fp is a pit-likedepression between the End and Hld,bounded distally by a high Dmr. The lingualface of the tooth is also less sloping than thebuccal one. On the buccal face, themesiobuccal groove disappears at a smallbut deep pit midcrown, but on the lingualface, the lingual one continues to the brokenedge of the fragment.
KNM-KP 31715: LM1 or M2 fragmentand two other tooth fragments (notfigured). These two fragments were found1·5 m distant from each other. Extensivescreening did not produce any additionalpieces.
A: LM1 or M2. This is a LM1 or M2,preserving the buccal portion of the crownwith parts of the Prd, Hyd and End, and thebuccal sides of 4·5 of the mesial root, andmost of the distal root. It is worn to justexpose islands of dentine on both buccalcusps. The mesiobuccal groove is mild anddisappears halfway down the buccal face.

297AUSTRALOPITHECUS ANAMENSIS
The buccal face is sloping, gently bilobate inocclusal view and convex in distal view. Thedemarcation between dentine and enamel isapparent on the broken surface, as is theroot canal. The edge of an IPF is just visibleon the lingual side of the broken distalsurface.
B: M fragment. This is a partial lingualfragment of a mandibular molar, preservingthe lingual groove occlusally and the centralportion of the lingual face to the cervix.
C: Tooth fragment. This tooth fragmentpreserves a portion of the mesial or distalface of a molar with a partial IPF.
KNM-KP 31716: P3 or P4 fragment andC/ fragments (not figured). This specimenwas found on the surface of the base of theslope below the excavation for the juvenile,KNM-KP 34725 (Wynn, 2000, Figure 2),the worn M2, KNM-KP 35847, and theadult associated teeth, KNM-KP 35839.It does not appear to belong with any ofthese individuals, and was at least 10 metresdistant from the majority of these fragments.
A: P3 or P4. This is the distal portion ofthe crown and about 8 of the root. Bothpartially preserved cusps are worn to thesame height, almost down to the level of thebase of the Fp and for the Pr to the dentine.A distal IPF measuring 3·7 BL by 2·1 SI sitsjust to the lingual side of center, and iswidest in its lingual half. The root flaresslightly distally and buccally towards itsapex. Enamel weathering has exposedperikymata on the distal face.
B: C/. This is a small fragment of C/preserving only a limited area of enamel onthe lingual face, with part of the basal tuber-cle. Inferior to this, the lingual face is nearlyflat. A faint distal labial groove is visible, asare perikymata.
KNM-KP 31717: Associated molars(Figure 4). This specimen was foundwashing down a slope about 10 m to thewest of the type mandible KNM-KP 29281.
After screening, a trench was excavated(Wynn, 2000, Figure 2) but no addtionalfragments were recovered in situ.
A: LM3. This is a crown with partialroots. The Pa and adjacent root are separ-ated from the rest of the tooth by thincracks. The occlusal outline is a roundedtriangle with a convex distobuccal side. Themesial edge shows a slight concavity at theIPF, which measures 4·8 BL and 2·8 SI, butmay have been larger as it is slightlyobscured by weathering. There is slight wearof the occlusal surface that has flattenedand polished the cusps and ridges and hasmade cusp sizes difficult to determine andobscured details of basin morphology. Nodentine is exposed. There appears to be aslit-like Fa, but the Fp does not appear tohave been distinct from the Fc. The Fcshows some crenulations. The doublelingual groove disappears half-way up thelingual face. There is a small cuspulebounded by vertical incisions near theocclusal margin of the Pr. There is anothersmall pit on the mesial aspect of the cuspadjacent to the occlusal plane. Faint verticalenamel ridges ring the sides of the crown,especially occlusally. The roots appear tohave angled distally. They measure 13·3 BLby a maximum of 9·6 MD near the cervix.
B: RM3. This is the crown, missing itsmesiolingual corner, and preserving about 3of the root at the distobuccal corner. Wearhas flattened the buccal cusps but dentineis exposed only barely on the Prd apex.Secondary fissures run into the centralbasin. The small Fa is slit-like, and the Fp isobscured by wear. Faint traces of fissuressuggest that a large, possibly double C6 waspresent. The distal margin of the toothtapers to a rounded point in occlusal view.The mesiobuccal groove is deep, forming apit-like depression close to the buccalocclusal margin and continuing about half-way down the buccal face. This depressionis bounded mesially by a protostylid made oftwo cuspules, a smaller distal and larger

298 . . ET AL.
mesial one. There is another small pit mesialto the protostylid. There is also a distinctdistobuccal groove that terminates a shortway down the crown. The lingual groove isdistinct near the lingual margin of theocclusal surface, but fades half-way downthe lingual face. Faint vertical enamel ridgesring all sides of the crown. An IPF is presentalong the mesial edge, producing a visiblemild concavity in occlusal view, but cannotbe measured.
C: RM2. This is the mesiolingual quarterof a crown, missing a triangular wedge ofenamel from the mesiolingual corner withabout 4 of the root preserved mesially. Thecrown is worn, largely obliterating occlusalmorphology, although there is no dentineexposure. A rounded lingual groove dividesthe lingual surface, making it bilobate.Division between enamel and dentine isapparent on the broken surfaces, andenamel is preserved up to 1·6 thick along themesial occlusal margin and 1·5 along thelabial margin.
KNM-KP 31718: R mandibular frag-ment (M2–3) (not figured). This was asurface recovery with no associated pieces.
This is a heavily cracked piece of man-dible with M2–3. The cracks are up to 1·5wide and run throughout the crown, rootand bone, splitting them into small frag-ments and obscuring nearly all the mor-phology. The incomplete M2 is also badlycracked and split. Only the distal portion ofthe crown is preserved, with one piece con-sisting of the End and sides of the toothseparated by a 1·6 split from a distal portionpreserving the Hld and a sliver of the Hyd.Some facetting of the Hld and Hyd indicatesslight occlusal wear, but there is no obviousdentine exposure. A faint distal IPFmeasures 2·2 SI by 2·4 BL.
The M3 crown is complete but severelydamaged by cracking obscuring details ofsurface morphology and occlusal wear. Themesial portion of the crown was wider than
the distal part, and the buccal surface moreSI convex and sloping than the lingual one.The lingual cusps are higher than the buccalones. There is a small C6.
KNM-KP 31719: I1 (not figured). This wasa surface discovery in the vicinity ofKNM-KP 29286. The tooth is badlyweathered and broken. Only a thin strip ofenamel remains at the lingual margin andalong one side at the enamel line. A faintbasal enamel bulge is evident. About 12of the weathered root is preserved. It istriangular in cross-section.
KNM-KP 31720: Maxillary M fragment(not figured). This was a surface discoveryin the vicinity of KNM-KP 34725 and closeto KNM-KP 35850, 35851 and 35852. It isa badly weathered molar fragment that pre-serves a complete lingual root and adjacentportion of a heavily worn crown. Overallweathering and cracking through the crownobscure its morphology. The root is about16·5 long.
KNM-KP 31721: RM2 & M3 partialcrowns (not figured). This was a surfacediscovery about 20 m to the west ofKNM-KP 30498 and 40 m to the east of thetype mandible, KNM-KP 29281 (Wynn,2000, Figure 2). Screening of this arearevealed additional specimens includingKNM-KP 31723 and 31724. None of thesecan be certainly associated.
A: RM2. This large mesial portion of thecrown is broken diagonally through the Paand Hy. There is matrix and some damageto the enamel on the mesial half of the Pr.Wear has flattened the cusp apices, particu-larly the Pr but no dentine is exposed.Striations are visible passing BL across thePr. A small Fa is preserved bounded bythe Mmr. A tiny enamel pit is visible on themesial side of the Pr. The lingual groovedisappears about half-way up the side of thecrown. The lingual face of the tooth is

299AUSTRALOPITHECUS ANAMENSIS
strongly sloping, convex in distal view, andmildly bilobate in occlusal view. Faintvertical enamel ridges are visible on thelingual side, and faint perikymata areapparent near the cervical margin. Themesial IPF is 5·9 BL by 2·9 SI, and reachesto the occlusal surface.
B: RM3. This is a strip of lightly wornmolar crown, about 5·6 wide. It is probablythe distobuccal portion of an RM3. It pre-serves the distal part of the Pa and threeother smaller cusps. The two occlusalgrooves defining the Me disappear just overthe occlusal margin. The central basin issmooth and lacks crenulations. Faint verti-cal enamel wrinkles mark the distobuccalface of the tooth.
KNM-KP 31723: RM3 (Figure 4). Thiswas a surface discovery about 20 m to thewest of KNM-KP 30498 and 45 m to theeast of the type mandible, KNM-KP 29281(Wynn, 2000, Figure 2). Screening of thisarea led to the recovery of additionalspecimens including KNM-KP 31721 and31724. None of these can be certainlyassociated.
This complete worn crown preservesabout 3 of the root along the lingual anddistal sides. The surface is weathered, andthere is a crack through the Pa runningdiagonally towards the Hy. Enamel is miss-ing along the cervical margin of the mesialhalf of the tooth. The occlusal outline istrapezoidal, tapering distally. The tooth isworn nearly flat, obscuring most details ofocclusal morphology although there is nodentine exposed. The buccal cusps arehigher and less worn than the lingual ones.The two buccal grooves define a cuspulebetween the large Pa and small Me, andboth grooves disappear about halfway up thebuccal face. Faint vertical enamel wrinklesare visible on the sides of the crown, and theremains of a mesial IPF are visible, thoughnot measurable, despite the weatheredenamel.
KNM-KP 31726: RP4 (Figure 4). Thisspecimen was found in the screening forKNM-KP 31727 but is considered torepresent a second individual.
This is a worn P4 with most of the crownand 7·5 of the lingual root preserved. Theenamel is missing from the buccal and mostof the distal sides. As preserved, the occlusaloutline is a rounded rectangle with its longaxis running BL. The Pr is worn to expose ahollowed-out area of dentine 3·3 BL by 4·4MD and preserves only a narrow band ofenamel around its margins. The higher Pa isalso worn, exposing an area of dentine 2·6MD that continues to the broken edge ofthe tooth. A concave mesial IPF measures5 BL by 1·7 SI and extends to the occlusalmargin. The lingual side of the crown bulgesout from the base, and is tightly convex indistal view. There were two distinct rootsthat diverged just above the crown. Thepreserved lingual root diverges stronglylingually from the crown. It measures 5·9BL by 4·9 MD near the cervix.
KNM-KP 31727: R/C (not figured). Thisspecimen was a surface find. A second speci-men, KNM-KP 31726, which is consideredto represent a second individual, was foundin the subsequent screening.
This /C lacks its entire mesial portion,and much of the crown. The distobuccalportion of the remaining crown is lost, sothat whereas 7 of the crown is preservedlingually, only 5 remains buccally. Enamel isworn away from the enamel line, but itsprobable original extent is discernable. Adistinct distal basal tubercle is preservedwith a distinct and deep distal lingual grooveadjacent to it, and a sharp median lingualridge bounding the groove mesially. Mesialto this, a small, flat area of the lingual faceremains. The root extends to a point 22·8from the cervix just below the distinct basaltubercle. It measures a maximum of 9·7near the cervix, roughly BL, broadening to amaximum of 10·5 about one third of the way

300 . . ET AL.
down its length, after this it tapers towardsits apex.
KNM-KP 31728: LM1 (Figure 4). Thisspecimen was found in the vicinity of thejuvenile mandible KNM-KP 31712, andhigher on the slope than the worn dm2,KNM-KP 31714 and the unworn molarKNM-KP 35842. It is possible that thisspecimen could be associated with one orother of the latter.
This is an unworn crown, possibly a germ.None of the root is present. Enamel isalmost completely missing from the mesialand distal sides, and towards the base of thebuccal and lingual sides. None of thecervical margin remains. The Hld is partiallypreserved. The mesial half of the Mmr isbroken away. The occlusal outline is arounded rhomboid. The four main cuspsare subequal in area, but the lingual onesare slightly taller. The occlusal surface istraversed by secondary fissures. The cuspsare high and pointed, and demarcated bydistinct grooves. A small and slit-like grooveis separated from the Fc by a ridge connect-ing the mesial cusps. This groove is incisedby the Mlg. Two tiny remnants of the Fa arevisible anterior to this. The lingual andmesiobuccal grooves are nearly continuous.The fissure between the End and Hld termi-nates distally in a tiny V-shaped pit repre-senting the Fp. The mesiobuccal groovedeeply incises the buccal face, making itstrongly bilobate. The lingual groove is lesspronounced on the lingual face, disappear-ing about 2 from the occlusal margin. Thebuccal side is more sloping than the lingual.
KNM-KP 31729: Rdm2 (Figure 4). Thiswas a surface recovery from the westernbank of the small drainage channel close tothe type mandible, KNM-KP 29281. Nofurther specimens were found in thescreening.
This tooth consists of the crown and up to8·0 of the distal and 6·5 of the lingual root.
Enamel is missing from the mesial face. Theocclusal outline is a rounded, elongatetrapezoid with convex mesial and distal sideswidening distally. Wear has exposed dentineislands, each about 1 in diameter, on thebuccal cusps, but the lingual cusps have onlypinpoints of dentine exposed, with polishingextending from the Med apex towards thecentral longitudinal groove. From largest tosmallest the cusp areas are: Med, Hyd, Prd,End and Hld. The taller lingual cusps are setdistal to the buccal ones. The Fa is deep butsmall, and bounded by a thin Mmr that iscrenulated at its occlusal edge. The Fa isbordered distally by a sharp crest runningbuccally from the Med. The Fp exists onlyas a distal extension of the Fc. The occlusalgrooves are deep and incise the occlusalmargins, but the lingual groove is deeperthan the mesiobuccal one. The lingual,buccal and distobuccal grooves form adistinct Y-pattern. The mesiobuccal grooveterminates in a small cleft bounded by a 2long ridge-like protostylid, and is mesial tothe lingual one. The buccal side of thecrown is more sloping and vertically convexthan the lingual one. The distal IPF is 2·1 SIby 3·1 BL, and located lingually, not quitereaching the occlusal rim. The roots divergestrongly, with the mesial one projectingalmost directly inferiorly, but the distal oneinclining distally. They measure a minimumof 8·3 MD near the cervix, where they areboth 6·8 BL. Both have longitudinalgrooves, but the distal one is faint.
KNM-KP 31730: LM2 & RP3 (Figure 4).These two teeth were found on the sameslope above the type specimen, KNM-KP29281, and above the isolated worn molarKNM-KP 29282. It is possible thatKNM-KP 29282 represents the same indi-vidual but the distance separating thesespecimens was too large to make this likely.KNM-KP 31730A was a surface discoverywhereas KNM-KP 31730B was discoveredin situ in an excavation extending from this

301AUSTRALOPITHECUS ANAMENSIS
spot to the southern side of the slope,where the juvenile, KNM-KP 34725 wasrecovered.
A: LM2. This is a very worn, weatheredand broken tooth. The mesial root has itsbuccal half broken away and much of therest of the roots are encased in matrix,except along the distolingual and mesiobuc-cal faces. Most of the enamel is broken awayfrom the sides of the crown. Only an 8·7MD by up to 5·5 BL portion of enamelremains on the occlusal surface between thelingual cusps. This portion is flattened andpolished from wear, obscuring any meaning-ful morphology. A worn area of dentineremains buccal to this enamel island, and inthe position of the Pr this is a deeply incisedpit. The roots are inclined distally andbuccally from the crown, and are roughlyparallel.
B: RP3. This weathered tooth is worn,and preserves 8·5 of its single root buccallyby only 2·5 lingually. Enamel is missingfrom the distal and lingual sides. Dentine isexposed over most of the flattened occlusalsurface. Enamel remains at the depth ofeach basin. The buccal face was stronglysloping towards the occlusal surface, and atrace of the mesiobuccal groove and Mmrare apparent. The enamel bulged from theroot on all sides.
KNM-KP 31732: Tooth fragments (notfigured). These tooth fragments were recov-ered in the screening for the isolated lowerdentition, KNM-KP 29286. Because noneof them obviously appears to belong tothis individual they were given a separateaccession number. However, these eightfragments may not be associated and couldwell represent more than one individual.
A: L/C. This is approximately the distalone-third of a crown, which has been brokentransversely through its tip and verticallythrough the buccal and lingual faces. Itpreserves most of the distal margin which ispolished. The Dmr forms a small tubercle at
the cusp’s distal margin, visible in lingualand labial views. This ridge, which has asmall IPF for P3 just lingual to the tubercle,sweeps along the cervical margin of thecrown, fading toward the crown center. Agentle distal lingual fossa is visible, butotherwise the lingual face is transversely flatand SI concave. The labial face is convextransversely and flatter vertically. Hypoplas-tic lines are visible across the labial face andperikymata can be seen along the polisheddistal ridge.
B: Tooth fragments. These are sevensmall fragments of molars and premolars.
KNM-KP 34725: Associated juveniledentition and skull fragments (Figure 11;F, J, K, N, O, Q and V not figured). Thisspecimen was largely recovered in situ.Through screening, a number of skullfragments were found washing down a slopeafter eroding from a bed close to the top ofa small hillock (Wynn, 2000, Figure 2).Excavation led to the recovery of the isolatedteeth associated with this specimen as wellas dental parts of additional specimens.These include KNM-KP 35839, 35840,35847, 37522 and 37523.
A: Rdi2. This is the lightly worn crownand 10·3 of its root, with a piece of maxillarybone adhering to the mesial surface of theroot. The tooth is weathered over its entiresurface and enamel is lost from the mesialand distal faces. The occlusal surface slopesfrom the superior mesial corner inferiorlyand distally, from 2·9 to 0·75 below themesial enamel line. A narrow strip ofdentine is exposed over the entire occlusalsurface. The Mmr and Dmr are faint,running from the basal tubercle to theocclusal margin along the vertically concavelingual face. The labial side is convex, andmarked by fine vertical furrows. The crownflares out from its root mildly, and only onthe mesial side. The enamel line archeshigher mesially than distally. The root isstraight, and measures 3·3 MD by 4·1 LaL.

302 . . ET AL.
Figure 11. KNM-KP 34725, associated juvenile partial dentition and cranial fragments, (A) in lateral,labial and lingual views, (B) in lingual, distal and labial views, (C) in lingual, labial and mesial views, (D)in occlusal, buccal, distal and lingual views, (E) and (G) in occlusal view, (H) in mesial, lingual and labialviews, (I) in distal and lingual views, (L) in occlusal and labial views, (P), (S) and (T) in occlusal view.(R) upper row in lingual and occlusal views, lower row in buccal and distal views, (U) in internal view.
The slit-like pulp cavity is visible on thesuperior break and measures just over 1LaL. The root projects directly superiorlyfrom the crown.
B: Rdc. This is the crown and completeroot. It is the mirror version of the left d/c
(C), but less worn. The crown is worn towithin 4·4 of the base distally, almost to thedistal basal tubercle, and 5·6 mesially. A 3·2wide vertical strip of enamel has spalled off
the labial surface, and another up to 1·4wide off the distolingual corner. Even if

303AUSTRALOPITHECUS ANAMENSIS
there had been no missing enamel, the toothis narrower LaL than MD. A sharply distinctdistal basal tubercle and a less distinctmesial basal tubercle are present, the latterleading to a short lingual marginal ridge.The mesiolingual fossa is broad and shallow,about 3·7 wide at the worn occlusal margin.The distal lingual groove is deep and nar-row, only 1·3 wide at the occlusal margin,because the median lingual ridge closelyapproximates the distal margin of the crown.The root measures 18·2 long, and 6·0 LaLby 5·25 MD at the cervix. Its apex curvesdistally.
C: Ldc. This specimen preserves thecrown and root. It is worn to within 3 of thebase of the crown lingually and 5 labially,with a hollowed 4·0 MD by 3·4 LaL area ofdentine exposed on the distally angledocclusal surface. A basal tubercle is presentat the distal corner of the crown. The lingualsurface is vertically straight below the basalbulge, and this surface slopes labially towardthe crown. The rounded labial surface ismore vertical and smooth, with distinctdistal and faint mesial grooves. It is the moreworn antimere of the right one (B).
D: Ldm1. This tooth consists of a crownand two roots, the mesial one measuring 7·9and the distal one 11·2 long. The roots aremissing their apices. Enamel is missing fromthe mesiobuccal corner and much of thelingual face. The crown is ovoid in occlusalview, almost twice as long MD as it is BL,and wider mesially than distally. It is heavilyworn so that dentine is exposed on most ofthe occlusal surface. Wear is oblique, slop-ing inferiorly and buccally, and the lingualcusps are higher than the buccal. The mesialcusps occupy about two-thirds of the crownsurface, with their apices near the MD mid-point of the tooth, although the lingual oneis slightly distal to the buccal one. Thereappears to have been an extensive Mmr withthe remains of an incipient mesial accessorycuspule. Both buccal and lingual faces of thecrown are roughly vertical in mesial view,
initially bulging out slightly from the root,and then sloping slightly towards each other.The lingual groove notches the lingualocclusal margin and barely continues ontothe lingual face. Just below the cervix, theroots measure a total of 8·9 MD, and bothare 5 wide BL. The mesial root anglesslightly mesially toward its apex, while thedistal one diverges distally. Both inclineslightly buccally away from the crown.
E: Rdm2. This is the mesiolingual cornerof the worn crown, preserving the Med.The Fa and part of the Fc are preserved.Dentine is exposed on the Prd continuingdistally to the break. The Med has a smalldentine pit at its tip and an elongatedexposure running down its buccal sidereaching almost to the dentine exposure onthe Prd. There are two tiny tubercles on theside of the Med. The top of the root is alsopreserved adjacent to this cusp. The lingualface was bilobate, divided by a distinctlingual groove.
F: RI2. This is the distal third of thecrown of the I2 germ. The distal margin isconcave in labial or lingual view, with itsmost widely flaring point about midcrown.The lingual face has a sharp edge, whichextends superiorly from the Dmr. The Dmrdemarcates a distinct distal lingual fossa thatextends about 3 from the basal break, abouthalf way towards the occlusal surface.Mesially the crown is broken through thedistal shoulder of a well-developed medianlingual ridge. Superior to this point, thelingual surface is flat in all dimensions. Thelabial surface is smooth and horizontallyconvex, except for a gentle but wide distallabial groove. It is straight in mesial viewexcept at the basal bulge and adjacent to theapex where it becomes convex.
G: RM2. This germ consists of a completecrown with its root just beginning to form. Itis a rounded rhomboid in occlusal view, withbilobate labial and lingual margins, and con-vex mesial and distal ones. It is heavilycrenulated, obscuring the divisions between

304 . . ET AL.
cusps and foveas. A small Fa can be seen,but the broad Fp is merged with the Fc. Thelingual side of the Hy is damaged, obscuringenamel relief. The Mmr and Dmr aremarked by vertical wrinkles. A distinctcingulum with vertical wrinkles traverses thelingual side of the tooth from the mesialside of the Pr, and continues as a cingularremnant to a point parallel with the Hy. Thelingual groove is complex, and terminates atthis feature. The buccal groove is not visibleon occlusal surface, but exists as a gentlefurrow on the buccal side forming thebilobate contour. The buccal, lingual anddistal sides of the crown are vertically con-vex. The mesial side is straighter and anglesmesially from the enamel line. The buccalside is more sloping than the lingual one.Vertical enamel wrinkles ring the crown.Perikymata are visible on all sides near theenamel line, and a possible hypoplastic lineis apparent 4·6 below the cervix on the distalside of the crown.
H: LI1. This is a complete unwornunerupted crown with 3·4 of its root, asmeasured on the lingual side. The crownmeasures 13·3 SI lingually. The occlusalsurface inclines slightly distally and in-feriorly. Five distinct mammelons disruptthe occlusal edge, with correspondingvertical grooves on labial and lingual sur-faces of the tooth extending almost halfwaytoward the root. The lingual face is distallydished, flaring sharply towards the cervix.The labial face is only slightly convex.Curved hairline cracks diverging superiorlyare visible on both labial and lingual facesseparating the central portion from themesial and distal. The mesial and distalmargins of the tooth flare out from the basegently, so that the MD diameter of the toothincreases from 5·6 at the base to 7·3 at theocclusal edge. Horizontal perikymata arevisible on the inferior half of the labial andlingual surfaces, and fine vertical grooves onthe inferior half of the labial side. Theenamel line arches superiorly about 5 at the
incisal angle of either side of the tooth. Theroot measures 8·9 LaL by 4·6 MD.
I: RI2. This is the distal two thirds of agerm made up of two pieces joined togetherat a hairline crack with no distortion. Thecrown is formed down to the labial cervicalmargin. Several mammelons are present,one on either side of the crack dividing thetwo fragments, and some more minor oneslaterally along the occlusal edge. The distalmargin of the occlusal surface curves inferi-orly to the vertical distal side of the tooth.The lingual fossa extends over most of thelingual surface. The labial surface is convexexcept for the shallow distal labial fossa thatruns along the upper half of the crown.Perikymata are clearly visible on the distaland labial surfaces. The distal enamel line isstrongly arched.
J: LI2. This is a small fragment of theocclusal half of the distal portion of I2,measuring only 8 SI by 4 MD. It is similar tothe I2 of the right side (I).
K: Mandibular I fragments. These arefive incisor crown fragments that are toosmall to attribute to a specific tooth. Twohave basal tubercles and the left side of thecrown in lingual view. One is the centralportion of the occlusal surface, and anarrower section of the middle of the crown.Another is a broken distolabial portion ofthe crown, with enamel missing from itsdistal surface. The last fragment is the leftside of a crown when viewed lingually.None of these pieces can be joined with eachother or other incisor fragments from thisspecimen. There is no reason to presumethat any belong to another individual,however. Mammelons are present on thetwo preserving some occlusal margin, andperikymata are visible on all.
L: R/C. This is an unerupted R/C crownmissing most of its lingual face. It is splitdown the buccal face with a thin cracksuperiorly but half-way down, the enamel ismissing in a wedge shaped piece about 2·6wide at the base of the preserved crown. Part

305AUSTRALOPITHECUS ANAMENSIS
of the lingual face remains mesially anddistally. The crown is 13·4 high as pre-served, pointed and unworn at the tip. It issimilar to that of KNM-KP 29286 in sizeand morphology, but is unworn. The labialface is smooth and tightly convex in occlusalview, with strong perikymata runninghorizontally across it. There is a distinctdistal labial groove that deepens cervically,whereas the mesial labial groove is indistinctand terminates at a Mmr. On the lingualface, the deep distal lingual groove is clearlydemarcated by sharp parallel vertical ridges,comprising the distal margin and centrallingual ridge, which run either side of it. Amesial lingual groove is visible just mesial tothe broken section. The preserved distalportion of the lingual face is flat, with asharp distal border. At the distal break, thebase of a sharp distal cingular ridge is visible.The mesial margin of the crown is convex inlabial or lingual view, with the Mmr andanother, cuspule-like feature further apicallyalong it. The distal border of the crown isalso sharp, and is concave in labial andlingual view.
N: L/C. This is a 10 SI by 7 MD piece ofthe superior labial surface of the left caninepreserving much of the distal lingual grooveand bordering the central lingual ridge anddistal margin. It is a good mirror of the rightone (L).
O: LP3. This is all but the mesiolingualcorner and distal margin of the crown of theP3 germ. The Prd is centrally placed, andthe Med exists only as a tubercle on themedian lingual ridge. The mesial and distalridges and transverse crest are sharp. Thereis a mildly crenulated, deep Fp. The buccalsurface of the crown is convex in occlusaland mesial views. It is bounded mesially anddistally by sharp, vertical marginal ridges.Perikymata traverse the buccal face, as dofaint vertical enamel wrinkles.
P: RP4. This is the broken unworn crownof a P4, germ comprising two pieces with aninsubstantial join. The mesiolingual corner
is missing, as is the distal margin and disto-buccal corner. The crown would have hadan oval occlusal outline. On the occlusalface, the deep Fa is delimited mesially by adistinct Mmr, and distally by a sharp trans-verse crest. The Fp is deep, set lower thanthe Fa, and crenulated around its distalmargin. Two distinct inflated ridges descendfrom both cusps to its base. There is adistinct but small End on the lingual end ofthe sharp Dmr. Mesial and distal buccalgrooves are visible on the buccal face andthere are horizontal perikymata near thebase.
Q: LP4. This is the distobuccal fragmentof an LP4 preserving the distal part of thePrd, Dmr, and part of the Fp. The Fp iswrinkled and deep. A distal labial groovefades out just at the inferior edge of thefragment. Faint vertical enamel ridges andperikymata are visible on its face.
R: Mandibular fragment (RM1). ThisM1 is complete. It sits in a piece of mandiblethat is 23 long, and extends 16·8 inferior tothe alveolar border. The lateral surface ofthe mandible is missing so that both rootsare exposed buccally. Part of the distal sideof the P4 alveolus and proximal wall of theM2 crypt are preserved. Medially, there isa deep subalveolar fossa inferior to a pro-nounced alveolar swelling. The anterioredge of the subalveolar fossa curves mediallyadjacent to the anterior break. The molarcrown is a rounded rectangle in occlusalview with a convex distal margin. The fourmajor cusps are subequal in area and theHld is slightly smaller, although it is almostas large as the Hyd. The mesial cusps areplaced next to each other, but the End sitsslightly distal to the Hyd. All cusps and theFc show occlusal wear, and small islands ofdentine are exposed on the buccal cusps.The Med is only lightly worn and remainshigh and pointed, and both lingual cuspsremain higher than the buccal ones. The Fais an elongate slit just distal to a thin Mmr, itis continuous with a larger pit between the

306 . . ET AL.
two mesial cusps. A faint ridge connectingthese cusps barely divides the Fa from theFc. The Fp is small, and lingual in position.The crenulated enamel of this tooth makesdivisions among foveas difficult to discern.The mesiobuccal groove terminates in a pitbordered by the distinct cingulum thatextends mesially along the buccal surface ofthe tooth to become a protostylid ridge. Thisridge is in turn continuous with the Mmr,so that the tooth is ringed mesially andlingually with a nearly continuous cingulum-like structure that extends distally nearly tothe Dmr. The lingual groove is mild anddisappears by mid-crown on the lingualsurface. Faint vertical enamel ridges ring thesides of the crown. A distinct mesial IPFmeasures 5·0 BL and 1·7 high. It is confluentwith the occlusal margin, and cuts into thecrown margin in occlusal view. There is nodistal IPF. The enamel line is straight on allsides. The roots adjacent to the enamel linemeasure 10·8 MD by 10·8 BL. The mesialroot is 11·2 long and the distal one 12·0 long.Both curve distally near their apices. Themesial one extends almost directly inferiorly,while the distal one angles distally. Above thepoint where the root is hidden by the invest-ing mandibular bone, the mesial root can beseen to have a longitudinal groove separatingits mesial and buccal portions.
S: LM1. This is a partial crown, preserv-ing the distolingual corner and most of thebuccal side. The mesial cusps are missing,and enamel is lost from most of the distalface and the distobuccal corner. It is amirror of the R side.
T: LM2. This germ consists of a crownwith its roots just beginning to form. Itsocclusal outline is a rounded rectangle withconvex distal margin. The occlusal surface ishighly crenulated by numerous secondaryfissures and inflated enamel ridges andcuspules. The mesial cusps are larger thanthe distal ones, of which the Med is largest.A wrinkled, BL elongate Fa is evident. TheMed is higher than the others. There is a
small C6 present between the Hld and End,in place of a Dmr. A Mmr is present, as is avertically wrinkled cingulum along the entirebuccal surface. This cingular ridge is strong-est where the mesiobuccal and distobuccalgrooves terminate in it. Its most promi-nent section occupies the region usuallyattributed to a protostylid. The lingual sur-face is divided halfway down by a lingualgroove. Perikymata ring the crown towardsthe enamel line, and a possible hypoplasticline is apparent 4·6 from the crown,measured at the center of the distal edge.
U: Occipital fragment. This is a pieceof left lateral occipital squame, preservedfrom almost the midline to asterion, andmissing its superior portion. It measures upto 33·1 ML by 29·2 SI. The lambdoidsuture is preserved for only about 15 medialto asterion, and the occipitomastoid suturefor about 11 inferiorly. The sutural surfacesare weathered. The external surface is alsoweathered and preserves little morphology,but a faint, rounded superior nuchal line isdiscernable. Above it, the bone is convex SI,and below it almost flat. Endocranially, thelateral margin of the groove for the superiorsagittal sinus can be seen. A distinct groovefor the transverse venous sinus extendsacross the middle of the fragment. Superiorto it, the internal surface of the bone isconcave in all dimensions. Inferior to thesinus there is also a concavity that extends tothe break. The cranial vault bone is thickersuperior than inferior to the transverse sinusgroove, perhaps suggesting a relativelyposteriorly-positioned cerebellar fossa. Nopart of any other sinus groove remains.
V: Cranial vault fragments. These aremore than 15 fragments of cranial vaultassociated with this specimen, some ofwhich can be joined. None can be attributedto a specific bone.
KNM-KP 35838: LM3 (not figured). Thisspecimen was a surface recovery. No otherspecimens are associated with it.

307AUSTRALOPITHECUS ANAMENSIS
Figure 12. KNM-KP 35839 associated maxillary teeth.(A) in labial, distal, lingual and mesial views, (B) inlabial, lingual and distal views, and (C) in mesial andocclusal views.
The specimen is a broken and worn crownand complete roots. The crown lacks themesial and lingual margins. The roots arecracked and weathered. The Hyd and Hldare preserved, along with parts of the End.Wear has flattenened the occlusal surfaceand obscured much of the morphology. Alarge area of dentine 2·6 in diameter isexposed on the Prd at the break, and a tiny,pinpoint of dentine is exposed on the Hyd.A faint Mlg is visible, with traces of the wornmesial and distal lingual grooves visiblelingual to it. Part of a C6 is visible at thebroken distal edge of the tooth. Themesiobuccal groove incises the occlusal rim.It extends onto the buccal face where itbecomes a shallow furrow halfway downthe side of the crown contributing to thebilobate contour of this side of the tooth.Fainter grooves delineating the Hld arevisible, and continue a short way down thesides of the tooth. The mesial roots extendalmost straight inferiorly, and are too poorlypreserved to measure accurately, buttheir lengths can be estimated at about 13lingually and 11·5 buccally. The distal rootbends distally towards its apex and its broadside is angled slightly buccally. It is 13·2long, and about 9·0 BL near the cervix.
KNM KP 35839: associated LI1, RC� &LP3 (Figure 12). A and B of this specimenwere found on the surface close to theexcavation of KNM-KP 34725 (Wynn,2000, Figure 2). KNM-KP 35839C wasfound in situ about 20 cm below the level ofthe lowest fragment of KNM-KP 34725.Additional specimens found either inscreening or excavation were KNM-KP35840, 35847, 37522, and 37523.
A: LI1. This is a complete crown and upto 11·8 of root. Light wear has just flattenedthe occlusal margin but has not exposeddentine. There is a developmental defect,such that the crown is bent distally near itscervical margin. A large groove running 8·7from the occlusal edge circles the base of the
crown, and marks the inflection point of thebend in the crown. The enamel is pitted onthe distal side inside the defect concavity. A3·9 SI by 1·8 LaL IPF is present on thelower half of the mesial surface. If this facetis placed vertically as if at the LaL axis, theroot angles strongly laterally in labial view.The vertically crenulated lingual face is con-cave SI and MD, with a distinct gingivaleminence. The mesial and distal lingualfossae are bounded by marginal ridges. Themedian lingual ridge is better developed inthe SI center of the crown, because betweenthis point and the cervix the defect hasdistorted the morphology. The labial surface

308 . . ET AL.
is convex MD, and almost straight SI excepttowards the cervix, where the defect also hasdistorted the surface. A vertically elongatedpit measuring 2·7 SI by 0·8 MD is visible onthe mesial portion of the crown near thecervix, which has almost no bulge fromthe root. Perikymata are also visible near thecervical margin. Incremental lines are visibleon the root running perpendicular to thelong axis.
B: RC/. This is an unworn completecrown, about 14·6 high, with up to about 12of a weathered root. A vertical cracktraverses the lingual surface but obscures nomorphology. The surface of the root isweathered and cracked. The crown is ovoidin occlusal view with the long axis runningmesiolabially to distolingually. It is diamondshaped in lingual view and unworn,although some polishing is visible at thecusp apex, distal margin and distal tubercle.Distinct basal tubercles are present. Sharplydelineated marginal ridges extend linguallyto the basal bulge of the crown, boundingthe superior edges of the mesial and distallingual fossae, and merging at a swollenbasal tubercle. Labially, marginal ridgesextend for 5·6 distally from the mesial basaltubercle and 3·1 mesially from the distaltubercle. The median lingual ridge isrounded, and set just 4·2 from the mesialmargin, so that the distal lingual fossa ismuch larger than the mesial one. The distalmargin of the crown is predominantlystraight but convex near the apex and con-cave towards the basal tubercle. The straightmesial margin is more vertical than the distalmargin, giving the crown an asymmetricallingual profile. The labial surface is convexMD marked by faint vertical enamel ridges,and traversed by uneven perikymata andpossible hypoplastic lines. The rootmeasures a maximum of 10·9 by a minimumof 9·0 adjacent to the crown. It is inclineddistally away from the crown.
C: LP3. This is the crown and up to 4·6 ofits root in places. It is missing enamel on its
distal surface. It also exhibits enamel defectsin the form of a pit on the distal half of thebuccal face, and a deep indentation in themesial face along the cervical line, just abovea malformed Mmr. This defect has affectedthe occlusal outline mesially. The Pa isabout twice as tall as the Pr, which is littlemore than a tubercle at the lingual end of asharp transverse crest. Little or no wear isevident. The Fa exists only as a small slitdue to the pathological shape of the crown.The Fp is deeper, and is extensive andindented by transverse fissures. The Dmr isbroken. Aside from the defect, the buccalface of the tooth is strongly sloping andstraight, beginning at the basal bulge. Thelingual side is less sloping, but also bulgesnear the root. The buccal face tapers root-ward and the enamel line is strongly convexwith a mesiocervical rootward enamelextension. Perikymata are visible on thebuccal face. The root is MD compressed,and appears to have been affected by thepathology of the mesial crown, so that itsmesial side appears pushed inward. Itmeasures 10·2 BL by 6·2 MD as preservednear the cervix.
KNM-KP 35840: LM3 & M fragments(not figured). This specimen was found onthe surface close to the excavation ofKNM-KP 34725 and 35839 (Wynn, 2000,Figure 2). Additional specimens foundeither in screening or excavation of this siteinclude KNM-KP 35847, 37522, and37523. It is possible that this is the sameindividual as KNM-KP 35839 although itshows no signs of the pathology so evi-dent on the incisor and premolar of thisspecimen.
A: LM3. This is a LM3 preserving 5·8 ofthe buccal root and 9 of the lingual one. Theocclusal surface is lightly worn with onlyslight polishing. The crown is missing atriangular chip on the mesiolingual cornerand a larger piece is lost distobuccally. Itsocclusal outline is a triangle with straight

309AUSTRALOPITHECUS ANAMENSIS
mesial side, slightly rounded lingual side andconvex distobuccal side. It is broader BLthan MD. The Pa is largest and tallest, andboth mesial cusps are larger in area than thedistal ones. Slight wear is apparent on thecusps. The occlusal surface is distinctlycrenulated, and the Fc is indented bysecondary fissures and inflated enamelridges. The Fa is a shallow, narrow slit nearthe mesial edge of the crown, and separatedfrom the Fc by a pronounced, broad, incisedridge connecting the mesial cusps. No Fp ispreserved. The lingual groove is distinct,incising the occlusal margin to terminatehalf-way up the crown in a wrinkled, shelf-like cingular remnant. The buccal groove isfainter, but still incises the occlusal marginand disappears half-way up the buccalface. The lingual side of the tooth is moresloping than the buccal, and has more of abasal swelling. A mesial IPF is present, andalthough weathering obscures its dimen-sions was at least 2·5 MD by 1·5 SI. It didnot meet the occlusal surface. The rootswere widely divergent, and adjacent to thecrown can be estimated to be 14·1 BL by9·4 MD.
B: L maxillary M fragment. This is asmall maxillary left molar fragment compris-ing part of the root and adjacent buccal faceof the crown as well as a triangular wedge ofworn occlusal surface, including the Fc andpartial Pr. The latter has a small area ofdentine exposed.
C and D: M fragments. These are twosmall, indeterminate molar fragments.
KNM-KP 35841: M crown (not figured).This was a surface recovery in the vicinity ofKNM-KP 34725 but at the base of the slopeto the east.
The specimen is a fragment of molarcrown, consisting of the side of a tooth and asliver of worn occlusal surface, measuring7·1 transversely by 5·8 high, and 3·1 ofadjacent root. It is too fragmentary toattribute to a specific tooth.
KNM-KP 35842: R maxillary M (Figure4). This specimen was found in vicinity ofKNM-KP 31712, 31714 and 31728. Someof these specimens could represent oneindividual, but they were widely dispersedand there are no obvious associations.
This is a right M1 or M2 crown lackingroots. It is unworn and slightly weathered.Most of the enamel is missing from aroundits cervical margin, except along most themesial and labial faces and the cusp apicesand several cuspules are chipped. It isrectangular in occlusal outline, with aninflated mesiolingual corner and bilobatebuccal edge. The buccal cusps are set mesialto the slightly taller lingual ones. Fromlargest to smallest the cusps are; Pr, Pa, Meand Hy. Secondary fissures form smallcuspules and inflated ridges of enamel overthe occlusal surface. The Fa comprises twosmall but deep pits, bounded by the Mmr,and a sharp ridge connecting the mesialcusps bounding it distally. These ridgesappear to be cuspidate, but the cuspules ortubercles along them are weathered. The Cois clearly defined and transected by a Mlgconnecting the deep Fc and equally deepFp. The Fp is slit-like, bounded distally by astrong Dmr. The lingual groove is truncatedon the occlusal surface but on the lingualface terminates in a distinct V-shaped notchhalf-way up the crown. The more mesiallyplaced buccal groove is distinct on theocclusal surface but only faintly visible onthe buccal face, continuing as a shallowfurrow to the break. There is an enamel piton the mesial side of the Pr.
KNM-KP 35844: M fragment (notfigured). This is a small fragment of wornmolar crown lacking roots.
KNM-KP 35845: M fragment (notfigured). This specimen was found in thescreening of a site approximately 20 mdistant from KNM-KP 31712. It is a smallfragment of worn molar or premolar crown.

310 . . ET AL.
KNM-KP 35847: LM2 (Figure 4). Thisspecimen was found in situ in the excavationfor KNM-KP 34725 and 35839 (Wynn,2000, Figure 2). Because it is worn it cannotrepresent the same individual as either ofthese specimens. Other specimens recoveredeither in the screening or excavation at thissite are KNM-KP 35840, 37522, 37523.
This is a worn crown and complete roots.The roots surround some mandibular bone.The lingual side of the crown is brokenaway. However, it can be seen that theocclusal outline of the crown was a rounded,MD elongate rectangle with a mildlybilobate buccal face. The Med appears tohave been the most extensive. Dentineexposures on the Prd, Hyd and Hld are 1·5,1·4 and 1·1 in diameter, respectively. Thereare small exposures of dentine on theremaining portions of the two lingual cusps,but most of these cusps are missing. Wearhas flattened the buccal cusps and obscuredthe foveas and grooves, although a Y-patternis apparent. Traces of a large shelf-like pro-tostylid are visible along most of the buccalside of the Prd. The faint mesiobuccal anddistobuccal grooves disappear just over theocclusal edge on the buccal face. The buccalface has a basal bulge, and then slopesocclusally. A crack divides the mesial faceand the large, obliquely oriented IPF. Thisfacet measures 5·2 BL and 2·3 SI and runsobliquely toward the crown, meeting theocclusal surface only in its buccal half. Thedistal IPF is 4·2 BL by 2·2 SI, and extendsto the occlusal margin. The deeply groovedmesial root is 18·3 long and angles distallytowards its bifid apex. Near the cervix, itmeasures 12·6 MD, and is 12·4 wide bulg-ing to 13·3 midway down its length. The tipsswing buccally and slightly distally. Thedistal root is 18·9 long. It is more angleddistally and buccally, with only a faintlongitudinal groove merging to a single tipthat is mainly formed by its buccal portion.Near its cervix it is 11·3 wide bulging to 12·4near its midpoint.
KNM-KP 35850: maxillary M fragment(not figured). This was a surface recov-ery in the vicinity of KNM-KP 34725,and close to KNM-KP 31720, 35851 and35852. It is an extremely rolled andweathered fragment of worn maxillarymolar crown with no details of occlusalmorphology remaining.
KNM-KP 35851: LM2 or M3 fragment(not figured). This was a surface recoveryin the vicinity of KNM-KP 34725, and closeto KNM-KP 31720, 35850 and 35852. Itis an unworn distobuccal portion of amaxillary molar crown. It is too incompleteto assign with certainty to a particular tooth,but is probably M2 or M3. It preserves theMe and Dmr. The bases of the Hy, Pr andPa are preserved with much of the Fc. TheDmr delineates a narrow Fp, and is largelyoccupied by a distal conule traversed by twolight grooves. This distal conule is separatedfrom the Hy by a continuation of the deepMlg that traverses the fragment and termi-nates half-way up the distolingual crownface. The buccal groove continues from theFc, notching the occlusal margin to rundown the length of the preserved buccalface. Faint vertical fissures mark the pre-served sides of this crown and all of thecusps.
KNM-KP 35852: LC/ (Figure 4). This wasa surface recovery in the vicinity ofKNM-KP 34725 and close to KNM-KP31720, 35850 and 35851. It is theweathered, worn crown and up to about 9·5of the root. A 1·8-wide crack divides thelabial surface of the whole tooth, where mostof the surface enamel is missing. The root isbroken at an angle, exposing an oval-shapedoblique section of root canal. The crown isworn to the level of the mesial and distalbasal tubercles, but slightly more so distally,with a mildly sinusoidal wear plane. Dentineexposure measures 9·8 MD by 4·1 LaL.The pulp cavity is exposed in a small circle

311AUSTRALOPITHECUS ANAMENSIS
Postcranial FossilsKNM-KP 271: L distal humerus (Figure13). This fossil was found by the HarvardExpedition of 1965 (Patterson & Howells,1967) and has been discussed by severalresearchers (e.g., Day, 1978; Senut, 1980;Senut & Tardieu, 1985; Lague & Jungers,1996) and has been described and discussedby Hill & Ward (1988). Because of this weare not redescribing it here. Hill & Ward
at the center of the occlusal surface. Theundamaged distal portion of the lingualsurface is worn smooth, but there is a hintof a distal basal bulge present connectingMmr and Dmr. A vertically concave mesiallingual groove notches the occlusal surface.A distal IPF meets the occlusal margin, andmeasures 3·7 LaL and a maximum of 1·8high. Near the cervix, the root measures 8·2LaL by 10·0 MD. Its surface is weathered.At the distal break the pulp cavity is difficultto measure accurately because it is brokenobliquely, but is estimated at about 2 indiameter.
KNM-KP 37522: L mandibular molar(not figured). This specimen was recoveredin situ in the excavation for KNM-KP 34725and 35839. The occlusal wear indicatesthat it is unlikely to belong to either to one ofthese and it cannot be associated with anycertainty to any of the other specimensrecovered through either excavation orscreening of this site. These includeKNM-KP 35840, 35847 and 37523.
This partial crown is made up of fourjoined fragments that comprise the buccaland most of the distal sides of a mandibularleft molar crown, probably M2. The Prd isbroken through near its tip but the Hyd andHld are complete, as are the bases of thelingual cusps. The lingual cusps would havebeen taller than the buccal ones. The Prd islarger than the Hyd. The buccal cusps arelightly worn and polished, and a tiny spot ofdentine is exposed on the Prd. A pit-like Fpis present. There is also a pit in which thedistobuccal groove terminates. The pro-nounced mesiobuccal groove intersects theMlg in the Fc. The mesiobuccal groovecontinues down the entire preserved buccalface, giving it its bilobate contour. A smallprotostylid is located about half-way downthe Prd from its tip. Small patches of surfaceenamel on the buccal face are pitted.Perikymata are visible over most of thebuccal face.
KNM-KP 37523: R mandibular M frag-ment (not figured). This specimen wasrecovered in the screening for KNM-KP34725 and 35839. The occlusal wear indi-cates that it is unlikely to belong to eitherone of these specimens and it cannot beassociated with any certainty to any of theother specimens recovered through eitherexcavation or screening of this site. Theseinclude KNM-KP 35840, 35847 and37522.
The specimen appears to be the lightlyworn lingual half of a R mandibularmolar, probably M2 or M3. There is a com-plete End, a small part of the Med, andwhat appears to be a C6. Some polishing isvisible.
KNM-KP 37524: tooth fragments (notfigured). These are four tooth fragments.They were found with KNM-KP 34725,KNM-KP 37522 and KNM-KP 37523.The first is a partial labial and mesial orlateral face of a mandibular incisor or caninewith about 5 of the root. The second is apartial buccal face of a mandibular premolarcrown. The third is the inferior half of themesiolingual corner of a molar crown with6·3 of adjacent root. The crown bulges outfrom its root and is vertically convex, andshows faint vertical enamel ridges andperikymata. The last is a fragment of wornmolar crown with an IPF partly preservedon one side. Although it is slightly worn, theenamel measures 2 thick near the preservedcusp tip.

312 . . ET AL.
Figure 13. KNM-KP 271 humerus. The distal portion of the lateral trochlear keel is missing. In proximalview, the extremely thick cortical bone is apparent.
(1988) note that ‘‘The specimen is cleanlyfractured along an oblique plane that beginsproximally about 5 cm above the medialepicondyle and terminates about 3 cm abovethe same point.’’ This, apparently, was oncetrue, but in its present condition, and asillustrated by Hill & Ward (1988), the cleanfracture was clearly made by a rock saw. Wehave no information about this cut, whichwas made while the specimen was on loan toProfessor Bryan Patterson. It was presum-ably to take a thin section of the shaft. Castsmoulded before this cut was made show ajagged break exposing a roughly cylindricalmedullary cavity about 7 in diameter on thediagonal break that is elliptical in cross-section. Several millimeters of bone were cutfrom the shaft, the greatest loss being on themedial side.
KNM-KP 29285: R proximal anddistal tibia (Figures 14 and 15). This was asurface find. The proximal fragment was
discovered on the bank of a small channel,whereas the distal fragment had washed intothe channel. In spite of extensive screeningthe middle portion of this tibia was notrecovered.
A: proximal tibia. This is the proximalportion of a right tibia, probably about aquarter of the total length, measuring 104. Itis complete except for some occasional mildabrasion along the margins of the condyles.Little of the medial condyle is missing, andcontours are preserved, even in the postero-lateral corner where it is most heavilyabraded. Laterally, all of the cortex alongthe anterior, lateral and posterior margins ofthe subchondral surface are abraded and/orbroken away so that the cancellous bone isuniformly exposed, obscuring the originalmargins of this joint surface. The top of theintercondylar eminence, the anteromedialmargin of the lateral condyle, and theposterolateral margin of the medial condyleare also abraded.

313AUSTRALOPITHECUS ANAMENSIS
Figure 14. KNM-KP 29285A, right proximal tibia.
There is some irregular bone depositedalong the anterior margin of the plateauextending nearly to the fibular facet, whichappears pathological in origin. Its extentis primarily along the area for knee jointcapsular attachment. This pathology was
apparently not severe, as it has affected noother morphology. The lateral side of theplateau is roughened, but intact. Just distalto it, the bone is abraded in places, andseparated from the shaft inferiorly by a largecrack up to 3 in diameter, which has
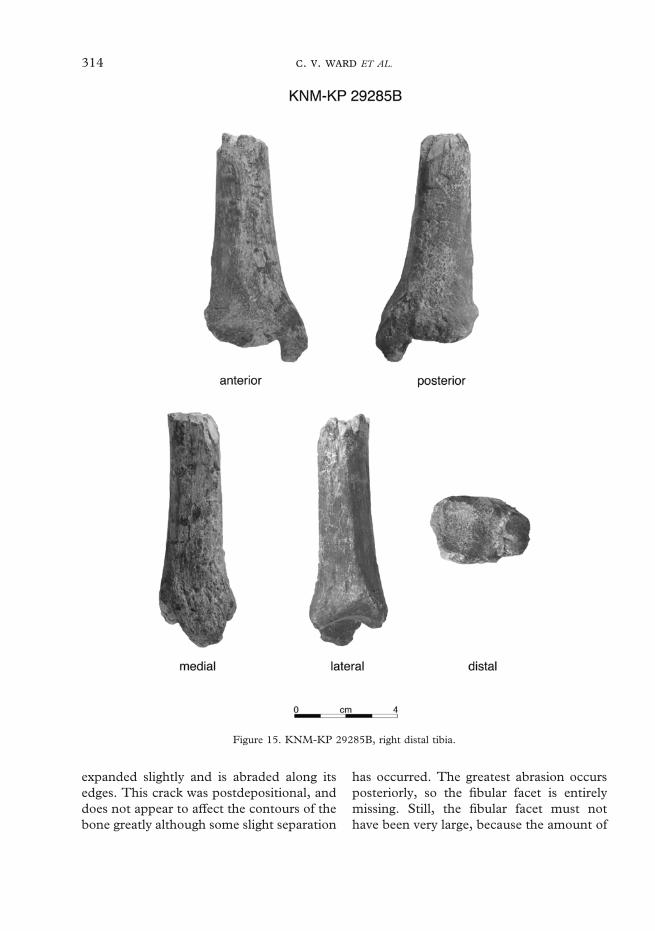
314 . . ET AL.
Figure 15. KNM-KP 29285B, right distal tibia.
expanded slightly and is abraded along itsedges. This crack was postdepositional, anddoes not appear to affect the contours of thebone greatly although some slight separation
has occurred. The greatest abrasion occursposteriorly, so the fibular facet is entirelymissing. Still, the fibular facet must nothave been very large, because the amount of

315AUSTRALOPITHECUS ANAMENSIS
bone missing is not extensive. The area ofinsertion of the iliotibial tract is pronounced,and sits upon a large, raised area withabrasion of only the cortex around itsmargin.
The cortex of the shaft is weathered butintact. The distal break is nearly transverse,but is slightly lower on the medial side,and about half of the thickness of corticalbone anteriorly is broken off for about 10,tapering toward the break along the medialside. In distal view, the endosteal surfacesare marked by some cancellous bone, butare generally visible so that a reasonablecross-section can be obtained. The bone isradiographically opaque.
Both condyles are concave in mostdimensions, but the lateral one becomesAP convex just along its most posteriorpreserved margin. The relative size of thecondylar surfaces cannot be accuratelydetermined due to damage to the lateralone. In proximal view, a distinct line,originating on the anterior-most point of themedial condyle runs laterally and posteriorlyto the most anterior points on the lateralcondyle and separates the rugose areaanterior to it from the gently concave inter-condylar area. The anterior attachment forthe medial meniscus is a deep groove justposterior to the medial extent of the linedemarcating the rugosity. The intercondylararea is peppered with nutrient foramina, andseveral faint puncture marks. Clear facetsdemarcating ligamentous and meniscalattachments cannot be detected, but thearea for attachment of the anterior cruciateligament is slightly depressed. The posteriorintercondylar area is similarly preserved, butin this case the groove on the posteriormargin for attachment of the posterior cru-ciate ligament is pronounced and clearlyvisible in proximal and posterior views.There is no apparent notch in the postero-medial margin of the lateral condyle for aposterior attachment of the lateral meniscus.The posterior attachment area for the
medial meniscus is also unclear. The plateauis set at about a 78� angle to the preservedanterior border of the shaft in medial view.
The tibial tuberosity is broad and pro-nounced but fairly smooth. The rugositysuprajacent to the tibial tuberosity along theanterior margin of the plateau obscures theline demarcating the tendinous and bursalareas of the tuberosity. The rugose areaprojects anteriorly from the surface ofthe bone further than the tuberosity. Thetuberosity is continuous distally as a sharpand laterally deflected anterior border,with a faint longitudinal ovoid depressionflanking it medially for the pes anserinusmuscles. The anterolateral surface of theshaft is convex transversely and the medialborder is mildly concave. The interosseusborder is rounded. The semimembranosusattachment forms a large, biconcave, almosttriangular depression with a raised rim thatis more distinct anteriorly than posteriorly.This facet sits distal to an overhang of thecondylar surface that projects about 3·5medial to the muscle attachment surface.
In posterior (or anterior) view, themetaphyseal region slopes gently fromthe plateau to the shaft with only a mildconcavity. The spiral line is rugose. It runsobliquely across the posterior side of theshaft. It extends distally from a point about10 mm below the disjunction along themissing portion of subcondylar bone wherethe fibular facet would have been, runninginferiorly to about the center of the shaft atthe distal break. Medial to the soleal linenear its proximal extent, the bone is gentlyconcave until it meets a mild ridge thatextends distal to the posterior cruciateligament attachment. This ridge meets thespiral line at the distal break. No nutrientforamen is preserved on the shaft.
B: Distal tibia. This is the distal portionof the same tibia, probably also about aquarter of the total length. It measures 83from the center of the talar surface along thekeel to the most proximal point preserved,

316 . . ET AL.
and 100 in maximum length. It is completeexcept for some minor spalling along theshaft, abrasion of cortex around the distalend, and some more extensive damage alongthe inferior and anterior margins of themalleolus, which obscure the original con-tours. Cortex is missing from the medial sideof the malleolus, but the general contours ofthe bone are discernible. Part of the distalmargin of the malleolar articular surface isabraded. Cortical bone is largely intactaround the proximal break, except for alongthe anteromedial corner. Still, a good cross-section can be seen and cortical thicknessesmeasured accurately.
The shaft is angled 93� medially and 90�posteriorly from the talar articular surface.The metaphysis flares out significantly pastthe articular margins on all sides. Therounded triangular cross-section measures20·6 AP by 22·5 ML at the break. Theanterolateral and posterolateral sides of theshaft are convex transversely, and the medialside is almost flat. The medial side of thebone is bounded anteriorly and posteriorlyby smoothly rounded borders. The interos-seus border forms a rounded ridge at theproximal end of the specimen, sharpening toa roughened crest about 36 above the distalend. It is mildly concave anteriorly as itpasses from the AP center of the lateral shaftproximally to the top of the synovial fibularfacet distally. The widest and most proximalpoint on the fibular facet is located abouttwo-thirds of the way from the anteriormargin of the bone along the interosseusmargin. Its dimensions can be estimated tobe about 9·4 SI by 17·8 AP. The fibularfacet is set at an angle of about 145� to thetalar facet.
A flexor hallucis longus groove is pre-served along the posterior margin of themalleolus, but its medial margin is abradedfor most of its length. The preserved portionof the inferior malleolar margin is concave,and angled from anteroinferior to posterosu-perior, and is not articular. As preserved, the
malleolus extends 13·4 from the adjacenttalar surface, but this is an underestimate ofthe original length since the end of themalleolus is missing. As preserved, themalleolus is 21·7 AP by 12·4 ML; probablyonly a slight underestimate of the originalvalues. The articular surface of the malleolusmakes a rounded but distinctly acute anglewith the talar surface.
The talar joint surface is almost square,measuring 23·0 AP at its midpoint, with thelateral part widening to 24·1 AP. ML, it is24·4 at its midpoint, 22·9 posteriorly and26·7 anteriorly. In distal view, the posteriormargin of the facet is straight, and theanterior is mildly bilobate. The medial sideis straight and oriented AP, and the lateralside flares anteriorly. There is a distinctkeel dividing the joint surface into roughlyequal halves. The two halves are gentlyconcave ML. They are more so AP, with theconcavity measuring a maximum of 5 deep.
KNM-KP 30503: Proximal manualphalanx (Figure 16). This was a surfacefind and no additional pieces were found inthe screening.
The specimen is most of the proximalthree-quarters of a manual proximal phalanxthat is 32·9 in maximum length. It is pre-served distally to an oblique line runningjust distal to the terminal end of the flexorridge on the left and about 5 distal to theflexor ridge on the right when viewedplamarly. The cortex is cracked and in sev-eral areas small spalls are missing, but it islargely intact except at the distal end whereclose to the break the dorsal cortex is lost. Alarge longitudinal crack runs along the pal-mar surface of the shaft, but does notobscure the dimensions of the bone. Morethan half of the proximal joint surface ismissing, and all its margins are abraded off.Cancellous bone is exposed along the entireproximal dorsal surface and half-way alongthe right and left sides, as well as in a smallarea palmarly adjacent to the longitudinal

317AUSTRALOPITHECUS ANAMENSIS
Figure 16. KNM-KP 30503, proximal manual phalanx in dorsal, palmar, side and proximal views.
crack. Most of the palmar tubercle on theright side of the base is preserved, but not onthe left.
Based on a comparison with the morpho-logically similar A. afarensis phalanges AL333-19 and AL 333-63, this phalanx wouldhave been roughly 40–42 long originally. Itis not possible to assign it to a particular ray.The palmar aspect of the shaft is ML convexbetween the flexor ridges proximally, andalmost flat near the distal break. Narrowgrooves flank the distinct flexor ridges on thepalmar side. The flexor ridges are hard tomeasure precisely since their proximal endsgrade into the sides of the shaft, but areabout 14 long on the left, and 17 on theright, the right one extending furtherproximally. In side view, the shaft ismarkedly convex dorsally, and slightly con-cave palmarly especially adjacent to theproximal end. In dorsal view, the shaft isslightly ML constricted before the meta-physeal flare. A cross-sectional outline isdifficult to estimate due to missing boneexternally and an ill-defined endostealmargin. Shaft dimensions at the distalmostpoint at which cortex is preserved, about 7from the distalmost point along the break,are 6·2 DP by 9·7 ML. At the center of theflexor ridges, these dimensions are 6·5
by 9·8, and proximally are 7·8 by 9·3.What little that remains of the proximal jointsurface suggests that this surface was mildlyconcave in all dimensions. Maximum MLwidth of the base can be estimated at about13 ML, but the dorsopalmar dimension istoo damaged to estimate accurately.
KNM-KP 31724: L capitate (Figure 17).This was a surface discovery about 20 m tothe west of KNM-KP 30498 and 40 m tothe east of the type mandible, KNM-KP29281 (Wynn, 2000, Figure 2). Screeningof this area led to the recovery of additionalspecimens including KNM-KP 31721 and31723. None of these can be certainlyassociated.
This specimen is nearly complete butmost of its surface is abraded away, andmuch of the preserved portion is weathered.The palmar surface of the bone is com-pletely missing, except for a transverseisthmus one-third of the way from theproximal end. Most of the dorsal surface ismissing as well. Laterally, the scaphoid facetis preserved except along its dorsal marginwhich is broken away, and an area along itsdistopalmar margin. The MC 2 facet is onlypreserved along its palmar portion to aboutthe dorsopalmar midpoint of the bone, and

318 . . ET AL.
Figure 17. KNM-KP 31724, left capitate.
not quite to the distal edge. Medially, thehamate facet is nearly complete except forabrasion along its dorsal edge that does notaffect its original contours, and weatheringalong its palmar margin that does obscurethe margin of the subchondral bone here.Proximally, the head is missing its palmarmedial edge, obscuring the palmar marginsof the scaphoid and lunate facets. Distally,only the center of the third metacarpal facetis preserved along the dorsal margin to apoint about two-thirds palmarwards.
The bone measures 21·6 in proximodistallength. Because of breakage and abrasion,almost no other measurements can be takenreliably (Table 4). The hamate facet is com-prised of a single articular surface in twoparts. It is about 7 wide.
In medial view, the facet runs straightdistally from the margin of the head to apoint just more than halfway along thebone, where it curves and runs distally anddorsally to the corner of the bone. Theproximal portion is SI flat. There is a slightelevation near the inflection point, where abroad, low, rounded ridge occurs at thedorsal edge of the facet. Distal to this,the facet inclines dorsally and curvesmedially, most tightly near its distal extent.The palmar surface of the bone is almostcompletely abraded, and the little cortex
remaining along the distal edge is heavilyweathered.
The scaphoid facet occupies about two-thirds of the lateral surface of the bone, andall of the head in palmar view. The scaphoidfacet faces mainly dorsally and laterally, andhas no proximally directed area on the head.The facet swings laterally along its dorso-distal edge to form a distinct saddle. Palmarto this, a 3-long arm of subchondral bonecontinues distally and palmarly to the deepintercarpal ligament pit that marks thecenter of the distal half of this surface. Justdistal to the intercarpal ligament pit, most ofthe palmar half of the second metacarpalfacet is preserved, except for the distopalmarcorner. This facet is oriented about 90� tothe third metacarpal facet, facing directlylaterally. In distal view, the preserved part ofthe second metacarpal facet is only mildlyconcave. This facet was continuous frompalmar to dorsal, with no intervening pitfor the intercarpal ligament, which in thisspecimen is located proximal to the MC 2facet. The MC 3 facet is concave ML andalmost flat dorsopalmarly. Damage obscuresthe original contours of this surface.
The lunate facet terminates not quite aquarter of the way from the proximal end.The scaphoid facet is also visible in palmarview, owing to its oblique orientation.

319AUSTRALOPITHECUS ANAMENSIS
The lunate facet occupies most of thehead. It is tightly curved dorsopalmarly. Itextends about 3 distally on the dorsal side,where there is a tiny remnant of a ridgealong its presumed palmar margin, but thisis difficult to ascertain with certainty due todamage to the specimen. It extends about6 distally on the medial side. On thehead, the junction between scaphoid andlunate facets is mildly crested, and runs fromdorsomedial to palmolateral. The lunatefacet is narrowest dorsally, measuring anestimated 6·5 ML here, widening palmarlyso that it appears roughly triangular inproximal view.
Allia Bay
Figure 18. KNM-ER 30200, maxillary fragment with M1–3.
Craniodental FossilsKNM-ER 7727: LM2. This was the firstfossil to be found in Area 261 at Allia Bayand was described by Leakey & Walker(1985) and Coffing et al. (1994).
KNM-ER 20420 LM2, KNM-ER 20421RM3, KNM-ER 20422 LM1, KNM-ER20423 LM2, KNM-ER 20427 LM1,KNM-ER 20428 LM3, KNM-ER 20432 L
mandibular fragment (P3–4), KNM-ER22683 LP4, & KNM-ER 24148 Ldm2.These specimens were found over severalyears by surface collecting at the main AlliaBay site in Area 261, East Lake Turkana.They were described and discussed byCoffing et al. (1994) and are not redescribedhere.
KNM-ER 30200: L maxillary fragment(M1–2, partial M3) (Figure 18).
A: L maxillary fragment (M1–2). Thisadult specimen preserves part of the alveolarprocess of the maxilla with M1 and M2. Thelateral side of the bone is missing. Thepalatine process is preserved for about 15ML and up to 23·8 AP, but it is missing itscortex except for a diamond-shaped area 12AP and ML that extends from the mesialroot of M2 to the midline. Superiorly, thefloor and part of the lateral wall of themaxillary sinus remain, and a ML convexsection of the nasal floor is preserved for13·1 anteroposteriorly by 9·0 ML. A tinyportion of the midline suture remains nearthe posterior edge of the fragment, at thelevel of M2. The alveolar margins are brokenaway medially, so that 4·8 of the lingual M1

320 . . ET AL.
root and the entire lingual M2 root areexposed. The distobuccal M2 root is brokenoff. The lingual M2 root is exposed by theloss of a flake of bone of the sinus wall. TheM1 is complete except for the very tips of thebuccal roots. The M2 is complete except formost of the distobuccal root, the tip of themesiobuccal root, and some enamel missingadjacent to the distolingual cervix.
The palate arcs up to a depth of about 10above the cervical margins of the teeth. Thepalatal process is 8·0 thick at the midline.This was a narrow palate relative to its toothsize. An estimate of the horizontal distancefrom the lingual cervix of M2 to the midlineis 13. The thin plate of bone that separatedthe nasal aperture from the maxillary sinus isbroken off. Part of the sinus wall remains,narrowing to an isthmus about 7 long AP onits medial wall, and lengthening to about 20AP along its floor. The exposed sinus floor isrelatively smooth and does not invaginatedeeply between the molar roots. Anteriorly,parts of the P4 alveoli are preserved.
The occlusal outline of M1 is a roundedtransverse rectangle. From smallest tolargest the cusp areas are: Me, Pr, Hy, Pa.The buccal cusps are higher than the lingualones and set mesial to them. A broad Co ispresent. A tiny pit is visible on the mesio-lingual corner of the tooth. The tooth islightly worn across its occlusal surface, withflattening of both lingual cusps, the Pa andthe mesial marginal ridge and polishing ofadditional ridges and crests. Pinpoints ofdentine are exposed on the mesial cusps.There is a trace of a slit-like 3 wide Faremaining. The Fp is in the form of adistinct pit, bounded posteriorly by a thickDmr marked by a tiny cleft that runstowards the distal IPF. The buccal groove ismesial to the lingual one. It deeply incisesthe occlusal margin but becomes fainttowards the cervix. The lingual groove ismilder occlusally and interrupted by anenamel ridge at the occlusal margin, but it isstrong on the lingual face. Both lingual and
buccal faces are bilobate. The lingual facehas a series of small enamel wrinkles about2·5 inferiorly from the cervix. The mesialIPF measures 4·7 by 2·4 and incises theocclusal surface. The buccal enamel linearcs up over each root, but the lingual lineis fairly straight. The single lingual rootdisplays a longitudinal groove. The verticallyimplanted buccal roots diverge at an angle of25�. The roots were not much longer thantheir preserved lengths of just more than10·6 for the mesiobuccal one and 11 for thedistobuccal one.
The M2 crown is similar to that of the M1
except in a few features, and because it isless worn the features are easier to discern. Itis more rhomboidal in outline. There isslight flattening of the lingual cusps and allcusps are polished but no dentine isexposed. The slit-like Fa measures 3·9 wideand clearly demarcates the Mmr. The Fp isdeeper and BL broader. The lingual andbuccal grooves are stronger on the occlusalsurface. A small distal IPF measures 2·3 BLby 1·4 SI, and is set lingual to the MD axisof the tooth and does not meet the occlusalplane. The lingual root measures 8·0 MDnear the cervix. It is about 15 long, andis marked by a lingual groove. It angleslingually towards its apex. The mesiobuccalroot extends directly superiorly.
B: M3. This tooth is missing its mesio-buccal corner to its center. About 10 of itslingual root remains, but only 4 of its rootbuccally. Judging by the curvature of theremaining part of the Pr it appears that thetooth had two larger mesial cusps and twosmaller distal ones. The distal margin of thetooth is rounded. There is light polishingand some facetting of the cusps and ridges.There is a small pit at the mesiolingualbreak. The lingual groove terminates in aseries of vertical enamel wrinkles midway upthe crown. There are fainter wrinkles on thedistal face that incise the distal occlusalmargin, one of which ends as a small cleft inthe margin. The small Me has grooves on

321AUSTRALOPITHECUS ANAMENSIS
Figure 19. Isolated dental specimens from Allia Bay. KNM-ER 30201 in occusal view, KNM-ER 30202in labial and lingual views, KNM-ER 30731 in distal and lingual views, KNM-ER 30749 in buccal view,KNM-ER 35231 and KNM-ER 35232 in occlusal view, KNM-ER 35233 in buccal and occlusal views,KNM-ER 35235 in occlusal view, KNM-ER 35236 in mesial and occlusal views, and KNM-ER 35238 inmesial and occlusal views.
either side and an apex at the edge of thecrown. The remaining mesial face of thecrown does not extend far enough to seeits IPF.
KNM-ER 30201: LM1 (Figure 19). Thisis a lightly worn and weathered crownpreserved almost to the cervical line. It ismildly elongate with 5 cusps. There is light

322 . . ET AL.
polishing of the buccal cusps and Hld. TheMed is largest, followed by the subequalbuccal cusps, and the End and Hld. Thefoveas are crenulated. The Fa is situatedtoward the lingual side of the tooth, extend-ing almost to the Med apex. A secondaryfissure traverses the mesiobuccal Med face.Another groove swings up the distal side ofthe Med almost to its apex, and is nearlycontinuous with the Fa, leaving an almostunbroken rim of enamel around the Med.The Fp is a deep pit. The faint lingualgroove is set just distal to the buccal one.Both disappear midcrown on the sides of thetooth, as does the distobuccal groove. Thebuccal side of the crown is trilobate, deeplyincised by the buccal and distobuccalgrooves. The lingual side is straighter. A pitis present on the mesiobuccal corner of thetooth.
KNM-ER 30202: RIl (Figure 19). Thiscrown is complete, with only some slightabrasion of the cervical enamel linelingually, and some hairline cracks whichcause no distortion. About 9 of its root ispreserved, and shows polish from rolling.The crown is worn occlusally, exposing astrip of dentine 9·0 MD by 1·3 LaL insidea thick enamel rim. The crown widensfrom 6·8 MD at its base to 10·5 at theocclusal margin, and would probably havebeen wider before it was worn. The occlusalplane slopes slightly distally and lingually.The lingual face is slightly concave in bothdirections, with little basal enamel swelling,an indistinct lingual ridge, and faint mesialand distal lingual fossae and ridges. Thelabial face is smoothly convex in both direc-tions with faint vertical enamel wrinkles. Amesial IPF measures 3·2 LaL by 2·5 SI,extending to the occlusal plane. A distal onedoes not meet the occlusal plane, andmeasures 2·1 LaL by 3·4 SI. The root haspronounced mesial and distal grooves, andthe pulp cavity is exposed on the brokenroot.
KNM-ER 30731: R/C (Figure 19). This isa complete crown and most of its root,missing only its apex. The crown is rolledand polished, and the root is spalled in manyplaces. Its occlusal outline is a MD com-pressed oval. The crown is worn at its apex,with the wear surface exposing a tear-dropped area of dentine that slopes distallyand cervically from its mesiolabial corner. Itflares out from the root more stronglymesially than distally. The Mmr and Dmrare both strong on the lingual face, andmerge with the moderately well definedbasal tubercle. These ridges, along with astrong lingual ridge, help to demarcateclearly defined mesial and distal lingualfossae. The Mmr extends much furthersuperiorly than does the Dmr, giving thetooth an asymmetrical appearance in lingualand labial views. The labial surface of thetooth is smoothly convex SI, and muchmore tightly curved transversely. The dis-tolabial groove is deeper than the mesio-labial one, but both extend to the occlusalsurface as preserved. They are continuouswith one another across the base of thecrown, forming a cingulum-like structure.The inferior portion of the labial surfaceshows transverse grooves that may representhypoplastic features, and weathering inplaces has exposed perikymata over partsof the surface. Rolling has obscured anytrace of IPFs. The root is compressedMD, measuring 6·6 MD by 10·2 LaL. Itscontours are obscured by damage to thespecimen. It is broken towards its apex, anda small area of the pulp cavity is exposed.
KNM-ER 30744: RC/ (not figured). Thisworn and rolled tooth preserves about 12of the root, but only 3 of enamel is leftbuccally, and 2 mesially and lingually. Noenamel is left distally where the cervix archestoward the crown. The pulp cavity isexposed and secondary dentine is fillingit. The root is hemicylindrical, roundedmesially and grooved distally.

323AUSTRALOPITHECUS ANAMENSIS
Figure 20. KNM-ER 30745, maxilla, in palatal and medial views.
KNM-ER 30745: L maxillary fragment(C/, P3–M1, partial M2, M3) (Figure 20).Preservation. This is a left maxilla, preservingthe palatine process to the midline, andno maxillary bone on the lateral surface.The buccal tooth roots have been lost toweathering and exposure, but the lingualroots are mostly preserved. The I1 alveolusis partially preserved, with part of its lingualmargin remaining. The interalveolar processbetween I1 and I2 appears complete. The I2
alveolar margin is preserved mesially andlingually, but distally its dimensions aredifficult to estimate due to damage here andaround the canine root. The rest of thealveolar margin is abraded throughoutmost of its extent, and only completelypreserved between P3 and M1, and along thedistal portion of M2. Only the distobuccalcorner of the canine and a short segment ofadjacent root remain. The premolars arecomplete. The M1 is missing a small sliver ofbone from the mesiobuccal corner, the M2 ismissing the entire buccal half, and the M3
is missing the mesiobuccal corner.Description. The bone of the palate is in a
friable and delicate state. It is cracked andslightly weathered, but the contours arepreserved. The midline suture can be
followed from near alveolare to a positionlevel with M2. The incisive canal opens intoa smooth incisive fossa so that its distalmargin is at the level of the P3–4 junction. Itsaperture was about 5 long AP, and, as far ascan be judged, was probably not this wide.The anterior margin of the incisive canalruns up and backwards at an angle ofabout 42� from the plane of the palate. Thethickness of the palatine process cannot bemeasured.
Palate widths from midline to the teethare about: at P3 14, P4 14·5, and M1 16.Distances at M2 and M3 cannot bemeasured accurately because of missingbone in this region. The palate is shallowanteriorly, sloping steadily up and back-wards until at the level of M3 it is about12 deep at the greater palatine groove.Just lateral to the M3 lingual root, asmall, biconcave section of the floor of themaxillary sinus remains, measuring 8·3 by4·3.
An estimate of the I1 alveolus diameternear the margin is 7. The distal half of thecanine is preserved, but the tip and lingualsides have been broken off. Its LaL diameteris about 11 at the cervix. Enamel on thelingual side is eroded, but enough remains

324 . . ET AL.
to show that there was a shallow distalgroove ending superiorly in a distal depres-sion. The root adjacent to the distobuccalcorner is preserved for about 5.
The P3 crown is complete, and is slightlyexpanded by a MD crack running throughthe groove separating the two cusps. No rootremains buccally. Its crown is ovoid inocclusal outline with a flattened, slightlyoblique mesial edge. The Pa is the larger ofthe two cusps, and is more projecting. Wearwas slight but cannot be fully assessed dueto weathering of the occlusal surface. TheFa is small, and restricted to a small pitimmediately mesial to the Pa and borderedmesially by a thin rather slight Mmr. Theslit-like Fp is 3·5 wide and is connected tothe Fa by an irregular furrow. It is bordereddistally by a wide Dmr. One or two smallgrooves run from each cusp towards theMlg. The buccal surface slopes markedlyupwards and has two shallow groovesdemarcating moderate mesial and distalbuccal ridges. The buccal enamel line isconvex rootward with a strong mesiocervicalenamel extension. The lingual surface isonly slightly sloping.
The P4 crown is complete, but its buccalroot is also missing. Its occlusal outline isalso a MD compressed oval, with an almostpointed apex buccally and a mesial surfaceflattened at the IPF. Wear was light withpolishing across the occlusal surface andsome flattening of the lingual cusp. The P4
crown is less asymmetrical than the P3
crown, with the two cusps being subequal insize. There is a small pit-like Fa, but the Fpis indistinctly defined. Both the Mmr andDmr are well developed, the latter the moreso. The lingual and buccal slopes areapproximately the same. The mesial anddistal buccal ridges are weakly developed.Buccal enamel extends rootwards, butnot to the extent seen in the P3. A singledistal lingual groove marks the lingual face.Occlusally, the groove separating the cuspsruns from a slit-like curved 2·7-long Fa to
a more irregular but longer Fp. Severalaccessory cuspules fill the Fp.
The M1 is a rectangular tooth with astrongly sloping lingual surface. Its occlusaloutline is a rectangle slightly compressedMD with a rounded distal margin. All cuspsare lightly worn with extensive polishing andflattening of the lingual cusps. The Pr is thelargest cusp, with the other three subequalin size. The buccal cusps are taller than thelingual ones. All cusps are worn, but onlythe Pr has dentine exposed in a small areaabout 0·4 wide. The tiny, slit-like Fa is onlyabout 1·5 long. The Fp forms a Y-shapedbifurcation of the Mlg that separates the twodistal cusps and sets apart a hypoconule.The buccal groove starts in a shallow pit inthe Fc, and terminates about half-way upthe buccal surface. The lingual groove isfaint at the occlusal margin, but is deep anddouble-furrowed before it terminates half-way up the lingual face. Other, fainterlingual grooves mark the lingual face of thetooth.
Only the Pr and part of the Hy of the M2
remain. Both are flattened by wear butneither has dentine exposed. Part of a smallFa is apparent at the front of the tooth. Acingular remnant can be seen at the mesio-lingual corner and slightly at the terminationof the lingual groove about midway up thecrown. The lingual groove has two parts onthe lingual face. One extends on the occlusalsurface to the break, and the other is a shortbranch swinging mesially, but not reachingthe occlusal surface.
The M3 is missing its mesiobuccal corner.It is a broad oval in occlusal outline, with itslong axis running mesiobuccally to disto-lingually. Its Pr is largest, and the Pa is onlyslightly smaller. All cusps are lightly worn,the Pr is extensively flattened, the Hy less so,and the lingual cusps are facetted. The distalmargin of the tooth is rounded, due to theweak development of the distal cusps thatexist primarily as the Dmr. The Fp is largeand well developed, and is connected to the

325AUSTRALOPITHECUS ANAMENSIS
Fc by a groove separating the Me and Hy.The buccal groove extends from the Fc toterminate halfway up the buccal face. Thelingual groove terminates in a series of soft,irregular grooves on the lingual face. Thedistolingual margin of the tooth has irregularvertical ridges running down the enamel.The lingual root of M3 is completelyexposed. It is missing only its tip, andmeasures 12·7 as preserved. It angles gentlydistally from the crown.
KNM-ER 30747: LP4 (not figured). This isthe distolingual half of a small crown and upto about 4 of the adjacent root only. It iscracked through the Med, and preserves theFp. The tooth was lightly worn. Radialridges lead from the Med and Dmr into thebasin. Two lingual grooves mark the lingualsurface extending to the occlusal rim. Adistal IPF measures about 3·4 BL by 2·1high. The crown does not flare out stronglyfrom the root.
KNM-ER 30748: L maxillary M frag-ment (not figured; see Figure 31). This is arolled and lightly worn maxillary molar frag-ment consisting of the mesiolingual cornerof a left molar, and the lingual half of themesial root. The wear facet cuts into theslit-like Fa, and there is a cingular remnanton the lingual corner. Part of the IPF isvisible lingual to the break. The cervicalmargin is straight in the parts preserved, andthe roots diverge strongly toward their api-ces. This specimen has been sectioned foranalysis of enamel structure by M. C. Dean.The section is shown in Figure 31 anddiscussed in the comparative morphologysection below.
KNM-ER 30749: LM1 (Figure 19; see alsoFigure 31). This is the distolingual half of asmall unworn LM1, broken diagonally frommesiolingual to distobuccal, preserving thelingual cusps, the End and the Hld, half ofthe Med and a sliver of the Hyd. It has up to
6·5 of its root preserved at its distolingualcorner, extending into the groove betweenthe roots lingually, and just past a longi-tudinal depression between the buccal andlingual portions of the distal root. The cuspsshow light polishing which could be due towear. The main cusps are demarcated bydistinct fissures, with accessory fissures andridges running from the cusps to the basins.The lingual groove extends from the Fc toterminate about half-way down the side ofthe crown, as do the two grooves delineatingthe Hyd. Perikymata are visible on thelingual and distal sides. No IPF is visible.This specimen has been sectioned foranalysis of enamel structure by M. C. Dean.The section is shown in Figure 31.
KNM-ER 30750: R/C (not figured). This isa R/C crown and about 11 of its root. Itsocclusal outline is a MD compressed oval.The crown was worn, with a tip facet setobliquely, running from the mesial side atthe apex inferiorly and distally. The enamelon the labial side is about 1 thick. There isa chip of enamel missing from the mesialside of the apex. The buccal surface iscylindrical with only a distal labial grooveadjacent to a sharply defined Dmr that has asmall tubercle at its base. On the lingualsurface, there is a strong distal groove andonly a mild, shallow mesial groove. Loss ofenamel obscures any Mmr. The crownbulges out only slightly from its root on allsides. The enamel line is fairly straighton three sides, but arches towards thecrown mesially. The root is compressedMD, measuring 5·6 MD by 8·3 LaL at thecervix, and curves gently distally towards itsapex.
KNM-ER 35228: RP4 (Figure 21). This isthe almost complete slightly rolled crown ofa germ, missing a small amount of enamelfrom the cervical margin on the mesial side.The top of the root was just beginning toform.

326 . . ET AL.
Figure 21. KNM-ER 35228, RP4, in occlusal and distalviews. Drawings by Susan Alta Martin.
Although this tooth has an ovoid crownoutline, it is unlikely to be a P3 for severalreasons. The Med is well developed, andalthough the Prd is large, its apex is placedmesial to the mesiodistal midpoint of thecrown. The Mmr is high, and there is adeep Fa that opens superiorly, ratherthan mesiolingually. The transverse crest isdistinctly notched. The distolabial groove ispronounced, delineating a distal cuspule.There is no mesiolabial groove. Its mor-phology is described here without referenceto it being a P3 or P4.
This is a very small tooth. In occlusalview, the ovoid crown outline has its longaxis running mesiobuccally to distolingually,with the distal surface slightly more convexthan the mesial one. The Prd is located nearthe center of the tooth on the BL axis and atabout the mesial third on the MD axis. Itclosely approximates the much smaller Med,which is set almost directly lingual to it. Twodistal cuspules representing the Hyd andEnd, are set farther apart than the maincusps, and sit along the pronounced Dmrseparated from the main cusps by smallfurrows. The End cuspule is slightly lower,and set just mesial to the Hyd one, so that itsits almost on the distal slope of the Med.The Fa is much smaller and set higher thanthe Fp. The Fp is wider BL than MD. Both
the Fa and Fp are located on the lingual halfof the tooth and are separated by a strong,notched transverse crest. The Fa is round inocclusal outline and almost featureless. Inmesial or distal view, the buccal surface ofthe tooth slopes strongly towards the Prdcusp tip. It bulges just above the cervix andagain at the cusp apex, and is mildly concavein between. The mesial, lingual and buccalsurfaces are all similarly concave SI. Thebuccal surface is marked by a broad disto-buccal groove that meets the occlusalsurface but fades out before the cervix.
KNM-ER 35229: L mandibular M frag-ment (not figured). This is the distolingualcorner of an M1 or M2 germ. It is brokenthrough the End and preserves most ofthe Hld. There is a tiny cupsule adjacentto the End on the Dmr. The Fp is apit. Crown formation is incomplete. Thedentine–enamel junction is not discernable.
KNM-ER 35230: M fragment (notfigured). This hexagonal fragment consistsof the central portion of an unworn molarcrown, probably a lower molar. It is about 6in diameter, and preserves the Fc, and basesof five cusps. It is not crenulated.
KNM-ER 35231: RM1 or M2 (Figure 19).This is a slightly worn complete crown withup to 1–2 of root preserved. Areas of enamelare missing around the cervix, so that thecrown appears to have more of a basalbulge than it would have had when intact.Its occlusal outline is a rounded trapezoid,with the mesiobuccal corner inflated relativeto the other three, so that the crown isbroader mesially than distally. From largestto smallest the cusp areas are: Pr, Pa, Me,Hy. The two lingual cusps are morerounded, and set somewhat distally to thebuccal ones, but all are roughly equal inheight. A tiny spot of dentine is exposed onthe Pa tip, but the tip of the Me is brokenoff, rather than being worn. Facetting is

327AUSTRALOPITHECUS ANAMENSIS
visible on the occlusal surfaces of bothmesial cusps, and just onto the mesial andlingual marginal ridges. The Fa is boundedby a smooth, continuous Mmr. The Fp,which is bounded distally by the strongDmr, is continuous with the deep Mlgconnecting it with the Fa. The Mlgseparates the Pr from Me, completelybisecting the weakly developed Co. Thefoveas are slightly crenulated. The lingualgroove is deep, and terminates midwayalong the lingual face. The buccal groove issimilarly developed on the occlusal surface,but less marked on the buccal face. Thelingual side of the crown slopes stronglytowards the occlusal surface and is convex inmesial and distal view, whereas the buccalsurface is flatter and less strongly sloping.The distal side of the crown is rounded inlateral view, while the mesial one flaresmesially towards the occlusal surface. Anovoid mesial IPF measuring 1·3 SI by 2·6BL sits just buccal of the MD axis of thetooth, but none is apparent distally.
KNM-ER 35232: LM1 (Figure 19). This isa slightly worn crown with 2–4 mm of root.Enamel is missing around the entire cervicalmargin. In occlusal view, the crown is arounded trapezoid in outline, broader dis-tally than mesially. The buccal cusps andHld show wear at the tips, but dentine isexposed only on the Prd as a tiny point. TheMed is higher and more pointed than theother four cusps, which are roughly equal inheight and is also larger in occlusal view.From largest to smallest the cusp areas areMed, End, Hyd, Prd, Hld. The deep Fa isbounded mesially by a strong Mmr anddistally by an incised crest connecting theMed and Prd. The Fc is large and traversedby numerous secondary fissures and inflatedridges and cuspules. There is a small, trans-verse Fp and Dmr marked by two fainttransverse grooves. The shallow lingualgroove terminates halfway down the lingualsurface, whereas the deep mesiobuccal
groove divides the entire buccal side of thecrown. A deep distobuccal groove separatesthe Hyd and Hld on the buccal face justabove the break. Each of these groovesoriginates in the Fc and incises the occlusalmargin. The buccal face slopes morestrongly than the nearly vertical lingual face.A mesial IPF measuring 1·4 SI by 1·6 BL isvisible.
KNM-ER 35233: LM2 (Figure 19). Thistooth is nearly complete, but rolled,weathered and polished so that it lacks muchsurface detail. Its roots are partially investedin bone. Enamel is broken along the cervicalmargin, and some of the surface of the rootabraded off beneath the enamel junction.The occlusal outline is a rounded rectanglewith convex distal margin. The five primarycusps are bounded by distinct groovesdelineating them in a Y-pattern. C6 and C7
are small, but present. A trace of a proto-stylid is visible. The lingual cusps are higherand more pointed than the buccal ones,and were probably only slightly worn. Thebuccal ones are worn but no dentine is ex-posed. From largest to smallest, in occlusalview, the cusps are Med, Prd, Hyd, End andHld. A pronounced Mmr bounds the Fa,which is a transverse slit. The Fc is extensive,but the pit-like Fp is small. It is boundedposteriorly by a Dmr and the C6. The lingualgroove continues half-way down the lingualface. Another, less extensive vertical furrowruns mesial to C7. The mesiobuccal grooveextends to the cervix, contributing to thebilobate shape of this face in occlusal view.A distobuccal groove extends a short wayonto the buccal face, terminating in a smallcingular remnant. The buccal face is morestrongly sloping than the lingual one, and istightly convex in distal view compared withthe nearly straight contour of the lingualface. Mesial and distal IPFs are visible butdo not reach the occlusal surface. The mesialone measures 3·1 SI by 4·0 BL, and thedistal one 2·5 SI and 2·5 BL.

328 . . ET AL.
There are distinct mesial and distal rootsof approximately the same length. Themesial root is BL broader than the distalone. Each has a mild but distinct longi-tudinal groove, and each terminates in a pairof apical tips. Their length is difficult tomeasure accurately because of the missingcervical enamel and weathering, but themesial one was probably about 18 and thedistal one about 16 long. They are mildlyinclined buccally, and more so distally.
KNM-ER 25234: LP3 (not figured). This isa badly rolled lower crown missing most ofits enamel, and its nearly complete root. Itsocclusal outline is a rounded triangle. Noobservable crown morphology is preserved.This tooth had a single root that is curvedslightly distally near its apex.
KNM-ER 35235: LM2 (Figure 19). This isan extremely rolled and polished crown withup to 3 of root preserved distally and 7lingually. Most of the crown morphology islost and the occlusal surface has littleremaining topography. The occlusal outlineis a rounded trapezoid that is broadermesially than distally. Cusps are barelydelineated, and from largest to smallestappear to have been: Pa, Pr, Hy and Me.The Mmr and Fa are barely distinguishable.The median longitudinal groove incises thestrong Co. There is a strong Dmr boundinga deep Fp. The distobuccal and lingualroots diverge strongly. The lingual rootappears as two fused columns, separated bya longitudinal groove.
KNM-ER 35236: LM3 (Figure 19). Thisslightly weathered tooth is missing only thedistobuccal root and a small piece of enamelat the mesiobuccal cervical margin. Thecrown is a rounded trapezoid in occlusalview tapering distally, and wider BL thanMD. Wear has polished the occlusal surfacebut there is no dentine exposure. Fromlargest to smallest the cusp areas are: Pr, Pa,
Me, Hy. A small, slit-like Fa is positionedbuccally. The deeper, elongated pit-like Fpis delineated by a strong Dmr that has asmall cuspule situated at its center. The Fc isinterrupted by a broad Co, which is incisedby a Mlg connecting lingual and buccalgrooves. The buccal and lingual groovesoriginate at mid-crown, but the lingual oneis more deeply incised on the side of thetooth where it terminates in a crenulated de-pression. Faint vertical enamel wrinkles ringthe crown along the occlusal surface. Periky-mata are occasionally visible on the sides,particularly near the cervical margin, and areespecially evident on the distobuccal corner.Slightly deeper and more irregular transversefurrows near the base may be mild hypo-plastic features. The buccal face slopes morestrongly than the lingual one. A 3·2 SI by 5·0BL IPF is visible on the straight mesial face.The distal face is convex in lingual view. Theroots are angled distally from the cervix, anddiverge BL towards their tips. The lingualone is double, incised by a median fissure,and notched at its tip. It is 12·3 long. Themesiobuccal root is 12·6 long. Only 4·1 ofthe distobuccal root is preserved.
KNM-ER 35238: RM1 (Figure 19). Thistooth has the crown and roots preserved, butis weathered and missing most of its detailedmorphology. In occlusal outline, it is arounded, transversely wide rectangle withmesial and distal sides made flatter bya pronounced concave mesial IPF and asmaller distal one. The crown is slightlybroader mesially than distally. The Pr is thelargest cusp, and the other three appear sub-equal in area. The tooth is worn, exposingcircles of dentine at each cusp tip, about 2wide on the lingual cusps and 1 wide on thebuccal cusps. Evidence of the Fa is obscuredby wear, and the Fp and Dmr are barelydistinguishable. The crown bulges outslightly from the root on all faces. In distalview, the lingual face is the most convex.The buccal groove terminates half-way up

329AUSTRALOPITHECUS ANAMENSIS
the crown in a cingular remnant. The lingualgroove is faint, and also terminates near thecrown midpoint. A 3·5 SI by 6·7 BL IPF hasworn away almost the entire mesial marginof the tooth, and can be seen in occlusal viewas a large, gentle concavity. A smaller IPF,2·9 SI by 5·8 BL, flattens the distal margin ofthe crown. The lingual root is single andgrooved along its buccal face. It is angledslightly distally, and diverges markedly lin-gually while the buccal roots ascend straightup from the crown in distal view. Themesiobuccal root is much shorter than thedistobuccal one. The lingual root is 12·4long, the distobuccal one 11·4 and themesiobuccal one 9·8 long.
Postcranial FossilsKNM-ER 20419: L radius (Figure 22).This fossil was recovered from sediments onthe northern flanks of Sibilot Hill, about20 km northeast of the Area 261-1 excava-tion. It has been described by Heinrich et al.(1993), so is not redescribed here.
Subsequent to that publication, however,another small shaft fragment was found thatarticulates with the proximal portion justdistal to the radial tuberosity. This fragmentdoes not join the proximal and middleportions, but suggests that the originallength of the bone was closer to the upperend of the 265–275 mm estimate given byHeinrich et al. (1993).
Figure 22. KNM-ER 20419, Sibilot radius showingadditional fragment at the distal end of the proximalfragment.
Comparative morphology
The morphology of the Kanapoi and AlliaBay hominin fossils is described here inreference to other relevant early hominins,as well as to other relevant fossil andextant hominoids. In this section, we treatA. anamensis as a distinct species that mayhave been ancestral to A. afarensis. Thetaxonomic assignation is covered further inthe discussion section. Although this mayeventually be proven incorrect, at present itis a plausible hypothesis that is not falsified
by data, and so provides a useful compara-tive framework from which to compare themorphology of these taxa.

330 . . ET AL.
OccipitalAlthough little morphology is preservedon the left occipital squame fragment(KNM-KP 34725U), a large groove for thetransverse sinus is apparent. There is noconfluens area remaining, so it is notpossible to determine the origin of the bloodthat drained into the transverse sinus. Thearea where an occipital marginal sinusgroove would be is not preserved. Theenlarged transverse sinus does not necessar-ily imply the lack of an enlarged occipitalmarginal system in this individual, however.Both sinus systems are enlarged in a numberof australopithecine specimens (Falk &Conroy, 1983; Falk, 1986, 1988), and atleast one A. boisei specimen (KNM-ER23000) has a transverse/sigmoid pattern onone side and an occipital/marginal patternon the other (Brown et al., 1993). A domi-nant transverse/sigmoid sinus system is thetypical pattern for African apes and humans,and is likely to be the primitive hominincondition (Kimbel, 1984), but the observedvariability in Australopithecus should under-score the need for caution when using thischaracter for taxonomic purposes.
TemporalThe small temporal fragment associatedwith the holotype mandible gives a few cluesabout the condition of the temporo-mandibular joint and the external acousticporus. It shows that the temporal squamewas well pneumatized in this adult indi-vidual. Placing the specimen in its correctorientation is difficult in the absence ofwhole crania of the species, but using otherAustralopithecus crania as a guide, we canmake some statements with comparativecertainty.
The articular eminence of A. anamensis isless well developed than in other Australo-pithecus and Homo species. In A. robustus andA. boisei the eminence develops into a strongcylinder anterior to the mandibular fossa.The KNM-KP 29281B eminence is even
less well developed than in the only cranialspecimen of A. aethiopicus, KNM-WT17000 (Leakey & Walker, 1988). Eminencedevelopment is somewhat variable, however,with the A. afarensis specimen A.L. 333-45being flatter than other A. afarensis and laterhominins and most closely approaching thecondition seen in A. anamensis. The pre-served portion of the temporomandibularjoint represents perhaps half of the originaljoint surface, although this is extremely dif-ficult to estimate. The preserved surface issituated directly inferior to the temporalsquame, rather than further laterally as isseen in most robust australopithecines. It isdifficult to measure the angle at which thetympanic plate meets the fossa because ofproblems with orientation and the fact thatneither is perfectly flat. Despite this, theKanapoi temporal gives the strong impres-sion that the two meet at a very obtuseangle, closer to 180� than in any otherhominin.
KNM-KP 29281B has a smaller externalacoustic porus than any other hominin, andin this respect is more like that of greatapes (Leakey et al., 1995) (Table 5). Thus,A. anamensis retains the primitive condition.This preserved diameter of the porus is thetrue maximum, because the outer edge oftwo-thirds of the bony porus is preserved.The margin shows the characteristic rugoseappearance of the edge that is seen in allcomplete hominoid temporal bones. Theposterior wall of the porus is broken, but athin sliver of bone remains stuck to thematrix filling the canal, which confirms thatthe canal itself had a small diameter.
MaxillaA difference between A. anamensis and A.afarensis previously cited (Leakey & Walker,1997) was that KNM-KP 29283 had amore vertical maxillary canine root thandoes A. afarensis. The robust canine rootis indeed more vertical than the majority ofA. afarensis specimens (Leakey et al., 1995),

331AUSTRALOPITHECUS ANAMENSIS
Table 5 External auditory porous dimensions
n
SI height AP breadth Area (SI�AP)
Mean S.D. Min Max Mean S.D. Min Max Mean S.D. Min Max
A. anamensis 1 8·9 — — — 5·7 — — — 50·7 — — —A. afarensis 4 10·9 1·23 9·5 12·3 10·1 1·73 6·4 11·8 101·6 34·3 60·8 139·2Pan troglodytes 9 8·1 1·33 5·6 9·8 6·7 1·24 5·2 9 55·1 14·2 29·1 72·0Gorilla gorilla 9 9·0 0·91 8·1 10·7 8·3 1·54 6·5 10·3 75·3 19·7 53·3 110·2Pongo pygmaeus 9 7·0 1·03 4·9 8·1 6·2 1·14 4·6 7·5 43·5 10·6 27·4 56·9Homo sapiens 9 10·6 1·36 7·6 12·5 7·3 0·89 5·6 8·5 77·1 13·7 57·0 89·8
Only adult A. afarensis specimens were used. A. afarensis data kindly provided by W. H. Kimbel.n=sample size, S.D.=standard deviation, Min=minimum, Max=maximum.
but is close to the orientation seen in AL333-2, and is now known to match oneunpublished specimen (W. H. Kimbel, per-sonal communication). Still, A. anamensisdiffers from the majority of preservedA. afarensis maxillae in this feature.
Commensurate with the robust verticalcanine roots are the large canine juga ofKNM-KP 29283. These juga contribute tothe rounded lateral margins of the nasalaperture, which are smooth with no lateralnasal crest (Leakey et al., 1995) (Figure 23).This condition contrasts with all A. afarensisspecimens (McCollum et al., 1993), exceptfor the single Laetoli specimen from Garusi(Kohl-Larson, 1943; Puech, 1986; Puechet al., 1986; Leakey & Walker, 1995; Senut,1999). Except for the Garusi specimen,A. afarensis has distinct lateral nasal creststhat are structurally distinct and situatedmedial to the canine juga. The Garusimaxilla and A. anamensis share a roundednasal aperture confluent with the caninejuga with most African apes. A. africanusspecimens, with the exception of STS 52,have rounded lateral nasal margins relatedto the presence of anterior pillars, but thesestructures are distinct from the canine juga(McCollum et al., 1993). This implies thatA. afarensis may be derived in its lateral nasalaperture contour relative to the other earlyaustralopithecine taxa.
The rest of the subnasal region ofKNM-KP 29283 resembles that described
for A. afarensis (McCollum et al., 1993),and is also similar to that found in Africanapes. It has a stepped nasal cavity floor witha broad incisive fossa, but damage alongthe midline obscures any evidence of anincisive crest on the vomer. The root of thezygomatic is partly broken away, but appearsto be adjacent to M1, as in A. afarensis.
The A. anamensis palates (KNM-KP29283, KNM-ER 30200 and KNM-ER30745) all preserve at least parts of themidline. The postcanine tooth rows ofKNM-KP 29283 were originally reported toconverge slightly posteriorly (Ward et al.,1999b). This specimen is weathered, how-ever, and closer inspection reveals thatthe intermaxillary suture is less completeposteriorly than originally reported. Thischanges the measurements slightly, so thatthis specimen can now be reconstructed tohave nearly parallel, but very slightly diver-gent maxillary postcanine tooth rows, fallingat the narrow end of the A. afarensis distri-bution (Table 6). Palate length/breadthindex (Table 6) shows that the A. anamensispalate is similar in length/breadth relationsto that of A. afarensis and chimpanzees, andvery similar to that of gorillas.
KNM-ER 30200 appears particularlynarrow (Figure 18). In this respect, it issimilar to the Garusi specimen (Puech,1986; Puech et al., 1986) which is also verynarrow (Figure 23). Unfortunately, it is notpossible to compare them directly because

332 . . ET AL.
Figure 23. The KNM-KP 29283 A. anamensis maxilla compared with the AL 200-1 maxilla from Hadarand the Garusi maxillary fragment from Laetoli, both of which are attributed to A. afarensis. BothKNM-KP 29283 and Garusi have rounded lateral nasal apertures lacking crests, unlike the AL 200-1.They also have large canine juga contributing to this margin, whereas the juga are anatomically distinctfrom it in AL 200-1. The Garusi specimen is the only A. afarensis to share these features with A. anamensis.Garusi also appears to have very narrowly set postcanine tooth rows, like some A. anamensis and few otherA. afarensis maxillas. Drawn by Susan Alta Martin.

333AUSTRALOPITHECUS ANAMENSIS
Tab
le6
Pal
atal
wed
gin
gan
dm
easu
rem
ents
use
din
its
calc
ula
tion
n
X:
Bi-
cani
nebr
eadt
h*Y
:B
i-M
2br
eadt
h†A
Pdi
stan
cefr
omX
toY
lines
Pal
ate
leng
th/b
read
thX
/YP
alat
alw
edgi
ng‡
Mea
nS
.D.
Min
Max
Mea
nS
.D.
Min
Max
Mea
nS
.D.
Min
Max
Mea
nS
.D.
Min
Max
Mea
nS
.D.
Min
Max
Mea
nS
.D.
Min
Max
A.
anam
ensi
sK
NM
-KP
2928
334
·5—
——
33·7
——
—33
·2—
——
1·02
——
—1·
02—
——
�1·
4—
——
KN
M-E
R30
745
33·8
——
—34
·0—
——
36·2
——
—0·
94—
——
0·99
——
—0·
3—
——
A.
afar
ensi
s7
27·7
4·03
23·0
35·0
32·0
5·26
24·0
41·0
35·1
3·13
31·0
40·0
0·91
0·11
0·73
1·03
0·87
0·08
0·78
1·04
7·0
4·4
�1·
712
·9P
.tr
oglo
dyte
s24
38·1
2·20
34·6
41·8
36·1
2·10
32·8
41·1
36·9
2·60
32·8
42·2
0·98
0·08
0·80
1·12
1·06
0·06
0·94
1·16
�3·
03·
0�
8·7
3·8
G.
gori
lla24
41·1
3·57
35·9
50·8
36·0
3·34
30·1
42·9
49·9
4·52
42·6
58·0
0·72
0·09
0·59
0·98
1·15
0·07
1·02
1·31
�5·
92·
7�
11·2
�0·
7H
.sa
pien
s15
23·4
1·38
20·6
25·1
40·2
2·38
35·3
43·6
29·1
1·79
25·1
31·4
1·39
0·11
1·18
1·59
0·58
0·03
0·54
0·63
32·2
3·2
27·5
38·2
*Min
imum
dist
ance
betw
een
cani
necr
owns
.†M
inim
umdi
stan
cebe
twee
nM
2cr
owns
.‡P
alat
alw
edgi
ngca
lcul
ated
usin
gfo
rmul
afr
omD
iGio
vann
iet
al.
(198
9).
Dat
afig
ured
inW
ard
etal
.(1
999b
).A
.af
aren
sis
data
kind
lypr
ovid
edby
W.
H.
Kim
bel.

334 . . ET AL.
they do not preserve relevant similar parts.Although it is missing anterior to M2,KNM-ER 30200 does not appear to havehad divergent tooth rows. KNM-ER 30745is difficult to assess in this respect alsobecause it is missing palatal bone adjacent toM2 and M3. Its reconstruction shows somedistortion in tooth orientation. Still, theoverall impression it gives is of parallel post-canine tooth rows. A. anamensis can still becharacterized as having a primitive palateshape with little postcanine tooth row diver-gence. The closely set, parallel mandibulartooth rows support this interpretation (seebelow).
Figure 24. Symphyseal profiles of A. anamensis mandibles in top row, compared with those of A. afarensisin bottom row. Adapted from White et al. (2000). Note the sloping orientation of the A. anamensissymphyses, especially the external contour basally, and the elongate anteroposterior extent of thesymphyses relative to their height.
MandibleThe mandible of A. anamensis is distinctfrom that of other hominin taxa. The man-dible attributed to A. bahrelghazali (Brunetet al., 1996) is morphologically equivalent to
those of A. afarensis, and at present weconsider them synonymous.
Most notably, all three sufficiently pre-served Kanapoi mandibles (KNM-KP29281, 29287 and 31713) have stronglysloping symphyses with strongly recedingsymphyseal contours, overlapping with onlyone other Australopithecus specimen, AL198-1 (Leakey et al., 1995; Ward et al.,1999b) (Figure 24; Table 7). The slopingsymphysis is reflected in occlusal view as along postincisive planum extending furtherposteriorly than is typical for any otherspecies of Australopithecus (Figure 25). Theexternal symphyseal contour is distinctive,being convex throughout its extent. Africanapes and other hominids tend to becomeflatter in section basally, with a more abruptbasal margin. A. anamensis is unique amongextant hominoids and fossil hominins in itssmooth convexity across this contour.

335AUSTRALOPITHECUS ANAMENSIS
Table 7 Angle of mandibular symphysis relativeto tooth row
Australopithecus Specimen Angle
A. anamensis KNM-KP 29281 141KNM-KP 29287 135KMN-KP 31713* (142)
Mean 139
A. afarensis LH 4 107AL 198-1 138AL 266-1 122AL 277-1 121AL 288-1 100AL 333-12 130
Mean 120
Extant taxa Mean S.D. Min Max nPan troglodytes 134 6 123 149 39Gorilla gorilla 137 7 123 154 37Pongo pygmaeus 135 7 125 154 51Homo sapiens 88 14 57 118 46
Midline symphyseal outlines were obtained by mold-ing Coltene President putty around the symphysis. Awooden rod was placed at the alveolar margin of thesymphysis and directed lingually to rest upon a secondrod spanning the posterior margin of the tooth row.Data from Sherwood et al. (in prep).
*KNM-KP 31713 is edentulous, so the valueobtained represents an estimate.
This shape appears to be associatedwith the narrowly set, parallel postcaninetooth rows (Figure 25; see below). Thismorphology is typical of ape mandibles,but again is unmatched among otherAustralopithecus or Homo species, all ofwhich have tooth rows that divergeposteriorly. This morphology reflects thatsuggested by the maxillary dental arcadesdiscussed above.
The holotype mandible (KNM-KP29281) is slightly distorted, so that itsincisors and, to a lesser extent, its caninesare inclined further anteriorly than theywould have been originally. This may give amistaken impression of more procumbentincisors than is accurate. Figure 26 shows aphoto of the KNM-KP 29281 mandiblewith the incisors replaced closer to theiroriginal positions. Incisor orientation in A.anamensis does not appear to have differedfrom that of other early hominins.
The A. anamensis mandibles also show thenarrow, parallel postcanine tooth rows evi-denced in the maxillary dental arcade, whichis a major difference between the mastica-tory apparatus of A. anamensis and A.afarensis (Figure 25). The larger I2 andmandibular canine teeth (see below) of A.anamensis must contribute to this shape inA. anamensis by making the anterior toothrow mesiodistally broad relative to theposition of the molar teeth, which are notexpanded relative to those of A. afarensis.
This morphology is reflected in theexternal alveolar contour as well, which isdistinctive in A. anamensis relative to otherhominins. In occlusal view, the most laterallyprojecting portion of the alveolar process is atthe canine juga. The bony contour andtoothrow itself turns medially anterior to thecanines, and posterior to them continues in aroughly straight line posteriorly and slightlylaterally (Figure 25). The canine juga aremore laterally projecting than are the P3 juga.In contrast, East African A. afarensis man-dibles and the specimen from Chad (Brunetet al., 1996) have toothrows that turn medi-ally at P3, with the corresponding alveolarcontour curving medially anterior to the P3
jugum. So, in contrast to A. anamensis, the P3
juga are situated lateral to the canine juga inthese other taxa. This disparity is not duesimply to expanded canine roots, becauseeven presumed female A. anamensis(KNM-KP 29284 and KNM-KP 31713)specimens with relatively small canines dis-play this morphology while large, presumedmale, A. afarensis mandibles do not.
The narrow jaws of A. anamensis mayhave influenced chewing mechanics andtemporomandibular joint form and func-tion. When tooth rows are narrow andparallel there are two main consequences.First, the amount of wishboning due to thelateral pull of the external masticatorymuscles (Hylander et al., 2000) is greaterthan if the tooth rows diverge. This isbecause the masticatory muscle lever arms

Figure 25. Occlusal views of the A. anamensis holotype mandible, KNM-KP 29281A, compared with theA. afarensis holotype, LH 4 from Laetoli, and several A. afarensis specimens from Hadar. The other twoA. anamensis mandibles are not figured here because of uncertaintly in reconstructing the angle of theirpostcanine toothrows accurately. Note the narrowly set, parallel postcanine toothrows of KNM-KP 29281compared with the others. Also apparent in this comparison is that the broadest point across the externalalveolar profile anteriorly is at the canine juga in A. anamensis, but always posterior to this near P3 in A.afarensis. AL 128-23 appears close, but still does not match A. anamensis in this contour. This contour canalso be seen in KNM-KP 29287 in Figure 6, and in KNM-KP 31713 in Figure 10. A. anamensis and LH4drawn by Susan Alta Martin, Hadar specimens adapted by her from drawings by Luba Gudz featured inWhite & Johanson (1982).

337AUSTRALOPITHECUS ANAMENSIS
Figure 26. KNM-KP 29281 with anterior teeth recon-structed in their original position. These teeth are notpreserved in their original positions, but have beenrotated anteriorly and inferiorly several millimeters.
Permanent tooth sizeExternal crown dimensions of A. anamensisteeth are generally similar to those of A.afarensis (Table 9). A. anamensis teeth aregenerally larger than those presently docu-mented for Ardipithecus ramidus, with a fewexceptions. The only dental dimensions inwhich Ar. ramidus exceeds A. anamensis isthe upper canine MD length and BLbreadth, and the M2 BL breadth. The twotaxa are equivalent in /C BL breadth, P4 andM2 dimensions. The greatest differencebetween A. anamensis and A. afarensis is thatA. anamensis has larger lower lateral incisors(Figure 28; Table 9), mainly due toexpanded MD dimensions. This almostcertainly contributes to its distinctivemandibular shape. In addition, while A.anamensis P3s overlap the size range of A.afarensis values, they are larger on average
decrease in length by the value x=1(1�Cos �), where 1=mandibular lengthfrom center of rotation of symphysis tocenter of combined wishboning muscleattachment and � is the angle of divergencefrom the midline. The value x reaches abouta quarter of the original length if the diver-gence is about 40�. Parallel tooth rows arefound in great apes and in A. anamensis,and in those species this is accompanied bythe addition of bone between the bodies inthe form of strong posteriorly developed tori,which can best resist the greater wishboning.
Second, dorsoventral shear forces at thesymphysis are due to the different forces ofrotation caused by muscles from the balanc-ing or working sides of the mandible(Walker, 1978; Hylander et al., 2000).These forces are more concentrated in thesagittal plane in parallel-sided and close-together tooth rows, as is the case with
scissors. As tooth rows diverge, the shearforces at the symphysis spiral more stronglyin opposite directions. The amount ofbone needed to resist these forces is alsogreater in forms with parallel tooth rows.In all cases, however, resisting bone canbe in any position away from the center ofshear and not necessarily concentratedposteriorly.
The A. anamensis mandible is less robustin proportion than that seen in any laterhominin on average (Teaford & Ungar,2000), overlapping the A. afarensis and A.africanus distribution. This can be quantifiedby the breadth/height index at the level ofM1, and is close to the values typical ofliving and fossil apes (Figure 27; Tables 3& 8). This may indicate that on average,A. anamensis mandibles tended not to becapable of resisting such high levels of trans-verse bending and torsion as those of laterhominins (Daegling & Grine, 1991). Thisslight difference may be related to a trend indiet to harder or tougher foods in these laterhominins than was typical for A. anamensis(Teaford & Ungar, 2000).

338 . . ET AL.
Figure 27. Mandibular corpus cross-sections at the first molar mid-crown perpendicular to the lingualalveolar margin. Adapted from White et al. (2000). The A. anamensis mandibular corpus is generally lessrobust on average in breadth relative to height than in A. afarensis, although there is overlap. Data listedin Table 8.

339AUSTRALOPITHECUS ANAMENSIS
Tab
le8
Com
par
ativ
em
and
ibu
lar
corp
us
size
and
shap
eat
M1
Tra
nsve
rse
brea
dth
Sup
eroi
nfer
ior
heig
htS
hape
[(br
eadt
h/he
ight
)�10
0]
nM
ean
S.D
.M
ax/M
in�
100
nM
ean
S.D
.M
ax/M
in�
100
nM
ean
S.D
.M
ax/M
in�
100
A.
anam
ensi
s3
18·6
1·0
110·
83
35·3
6·5
144·
13
53·6
6·8
129·
6A
.af
aren
sis
1219
·02·
214
9·4
1032
·93·
413
5·2
1057
·86·
314
2·4
A.
afri
canu
s6
21·5
2·2
125·
04
36·1
1·7
110·
64
59·8
5·9
121·
8P
.tr
oglo
dyte
s20
14·2
1·3
132·
220
28·8
2·7
143·
820
49·5
5·5
158·
1G
.go
rilla
2019
·32·
215
5·6
2038
·24·
615
3·7
2050
·94·
714
6·2
Com
para
tive
data
from
Dae
glin
g&
Gri
ne(1
991)
and
Loc
kwoo
det
al.
(200
0).
n=
sam
ple
size
,S
.D.=
stan
dard
devi
atio
n,M
in=
min
imum
,M
ax=
max
imum
.

340 . . ET AL.
Tab
le9
Com
par
ativ
ep
erm
anen
tto
oth
met
rics
Mes
iodi
stal
Lab
io/B
ucco
lingu
alC
row
nA
rea
(MD
�B
L)
Cro
wn
Sha
pe(M
D/B
L)
nM
ean
S.D
.M
inM
axn
Mea
nS
.D.
Min
Max
nM
ean
S.D
.M
inM
axn
Mea
nS
.D.
Min
Max
Max
illar
yde
ntit
ion
I1
Kan
apoi
211
.7—
10.9
12.4
28.
6—
8.2
8.9
299
.9—
89.4
110.
42
1.36
—1.
331.
39A
llia
Bay
110
.5—
——
19.
3—
——
197
.7—
——
11.
13—
——
Ar.
ram
idus
110
.0—
——
27.
8—
7.5
8.2
175
.0—
——
11.
25—
——
A.
afar
ensi
s6
10.6
0.91
9.0
11.8
88.
40.
717.
19.
76
89.7
13.5
63.9
101.
96
1.26
1.10
10.8
01.
40
C/ K
anap
oi2
11.2
—10
.611
.72
10.7
—10
.211
.22
119.
6—
108.
113
1.0
21.
04—
1.04
1.04
Alli
aB
ay0
——
——
111
.0—
——
0—
——
—0
——
——
Ar.
ram
idus
211
.4—
11.2
11.5
211
.4—
11.1
11.7
212
9.4
—12
4.3
134.
52
1.00
—1.
010.
98A
.af
aren
sis
1310
.00.
688.
911
.613
10.9
1.02
9.3
12.5
1310
8.9
16.7
82.8
145.
013
0.92
0.55
0.81
1.01
P3
Kan
apoi
39.
91.
648.
711
.82
12.7
—12
.113
.22
113.
7—
112.
511
4.8
20.
71—
0.66
0.77
Alli
aB
ay1
9.9
——
—1
13.0
——
—1
128.
7—
——
10.
76—
——
Ar.
ram
idus
17.
7—
——
112
.5—
——
196
.3—
——
10.
62—
——
A.
afar
ensi
s9
8.7
0.53
7.5
9.3
912
.40.
6311
.313
.49
108.
111
.284
.812
0.9
90.
710.
025
0.66
0.74
P4
Kan
apoi
37.
70.
467.
28.
10
——
——
0—
——
—0
——
——
Alli
aB
ay1
8.8
——
—1
13.9
——
—1
122.
3—
——
10.
63—
——
Ar.
ram
idus
18.
4—
——
111
.3—
——
194
.9—
——
10.
74—
——
A.
afar
ensi
s17
9.1
0.70
7.6
10.8
1112
.40.
8411
.114
.511
112.
217
.984
.415
6.6
110.
730.
040.
660.
81
M1 K
anap
oi3
11.7
0.17
11.6
11.9
313
.30.
5612
.813
.92
159.
2—
157.
116
1.2
20.
870.
047
0.83
0.90
Alli
aB
ay5
11.4
1.24
10.0
12.9
512
.70.
9011
.714
.14
149.
021
.812
0.5
172.
04
0.93
0.08
70.
871.
06A
r.ra
mid
us0
——
——
0—
——
—0
——
——
0—
——
—A
.af
aren
sis
1512
.11.
0310
.513
.812
13.4
0.92
12.0
15.0
1116
7.8
22.4
129.
620
2.5
110.
920.
580.
851.
04

341AUSTRALOPITHECUS ANAMENSIS
Tab
le9
Con
tin
ued
Mes
iodi
stal
Lab
io/B
ucco
lingu
alC
row
nA
rea
(MD
�B
L)
Cro
wn
Sha
pe(M
D/B
L)
nM
ean
S.D
.M
inM
axn
Mea
nS
.D.
Min
Max
nM
ean
S.D
.M
inM
axn
Mea
nS
.D.
Min
Max
Max
illar
yde
ntit
ion
cont
inue
dM
2 Kan
apoi
114
.2—
——
214
.9—
14.8
14.9
319
8.9
11.4
189.
621
1.6
30.
100.
040
0.88
0.95
Alli
aB
ay4
12.9
1.45
11.4
14.8
413
.50.
8012
.914
.73
176.
023
.515
0.5
196.
83
0.94
0.15
0.84
1.11
Ar.
ram
idus
211
.8—
11.8
11.8
214
.6—
14.1
15.0
217
1.7
—16
6.4
177.
02
0.81
—0.
840.
79A
.af
aren
sis
1013
.00.
6212
.114
.111
14.7
0.64
13.4
15.8
1019
1.0
17.0
162.
122
2.8
100.
880.
025
0.84
0.91
M3 K
anap
oi5
12.0
0.82
11.1
13.0
414
.60.
8113
.815
.73
185.
018
.316
4.5
199.
43
0.83
0.04
50.
800.
88A
llia
Bay
311
.80.
7911
.212
.73
13.7
0.75
13.0
14.5
316
2.4
19.0
149.
518
4.2
30.
860.
036
0.82
0.88
Ar.
ram
idus
110
.2—
——
112
.3—
——
112
5.5
——
—1
0.83
——
—A
.af
aren
sis
1112
.51.
3610
.914
.811
14.3
1.13
13.0
16.3
1117
9.9
33.7
141.
724
1.2
110.
880.
038
0.81
0.95
Man
dibu
lar
dent
itio
nI 1
Kan
apoi
46.
90.
296.
67.
33
7.8
0.64
7.3
8.5
354
.07.
049
.562
.10
0.90
0.04
50.
860.
95A
llia
Bay
0—
——
—0
——
——
0—
——
—0
——
——
Ar.
ram
idus
16.
0—
——
0—
——
—0
——
——
0—
——
—A
.af
aren
sis
66.
50.
935.
68.
06
7.4
0.29
6.9
7.7
548
.18.
938
.661
.65
0.88
0.13
0.75
1.04
I 2K
anap
oi6
7.9
0.78
6.6
8.7
58.
30.
377.
88.
64
68.1
3.6
64.5
73.1
40.
980.
110.
871.
12A
llia
Bay
0—
——
—0
——
——
0—
——
—0
——
——
Ar.
ram
idus
0—
——
—0
——
——
0—
——
—0
——
——
A.
afar
ensi
s7
6.3
0.82
5.0
7.2
78.
00.
726.
78.
86
49.7
7.8
41.5
596
0.82
0.14
0.57
0.93
C/ K
anap
oi6
9.8
0.92
8.7
11.0
510
.30.
929.
411
.45
100.
916
.381
.811
5.4
50.
940.
086
0.83
1.09
Alli
aB
ay2
7.5
—6.
68.
32
9.9
—9.
210
.62
74.4
—60
.788
.02
0.75
0.04
60.
720.
78A
r.ra
mid
us0
——
——
110
.0—
——
0—
——
—0
——
——
A.
afar
ensi
s12
9.0
1.14
7.5
11.7
1410
.51.
158.
812
.49
95.5
19.5
66.8
121.
79
0.89
0.16
0.77
1.13

342 . . ET AL.
Tab
le9
Con
tin
ued
Mes
iodi
stal
Lab
io/B
ucco
lingu
alC
row
nA
rea
(MD
�B
L)
Cro
wn
Sha
pe(M
D/B
L)
nM
ean
S.D
.M
inM
axn
Mea
nS
.D.
Min
Max
nM
ean
S.D
.M
inM
axn
Mea
nS
.D.
Min
Max
Man
dibu
lar
dent
itio
nco
ntin
ued
P3
Kan
apoi
712
.10.
7011
.313
.46
8.9
0.34
8.5
9.4
610
7.4
9.7
97.2
119.
36
1.36
0.07
31.
311.
51A
llia
Bay
18.
6—
——
112
.0—
——
110
3.2
——
—1
1.40
——
—A
r.ra
mid
us2
7.9
—7.
58.
22
10.7
—9.
911
.52
84.3
—74
.294
.32
0.74
—0.
760.
71A
.af
aren
sis
259.
51.
047.
912
.625
10.5
0.87
8.9
12.6
2510
0.3
14.6
70.3
133.
625
0.91
0.10
70.
711.
19
P4
Kan
apoi
59.
10.
738.
29.
87
10.7
0.76
9.8
11.7
599
.414
.980
.411
1.7
50.
830.
020
0.81
0.86
Alli
aB
ay4
8.5
1.10
7.4
9.7
410
.21.
269.
011
.94
88.1
21.7
70.2
115.
44
0.83
0.05
20.
770.
88A
r.ra
mid
us2
8.2
—7.
58.
92
10.7
—9.
911
.52
78.8
—71
.286
.32
0.85
—0.
790.
92A
.af
aren
sis
239.
81.
037.
711
.420
11.0
0.77
9.8
12.8
2010
7.6
16.7
77.0
134.
520
0.89
0.07
60.
741.
01
M1 K
anap
oi10
12.8
0.78
11.5
13.8
1012
.00.
9810
.513
.510
152.
724
.110
5.0
180.
810
1.06
0.04
60.
991.
15A
llia
Bay
211
.8—
11.6
11.9
210
.6—
10.2
10.9
112
9.7
——
—1
1.09
——
—A
r.ra
mid
us2
11.1
—11
.011
.12
10.3
—10
.210
.32
113.
3—
113.
211
3.3
21.
08—
1.08
1.08
A.
afar
ensi
s27
13.0
0.96
10.1
14.8
2112
.30.
7711
.014
.821
165.
718
.512
4.3
197.
121
1.05
0.04
20.
971.
13
M2 K
anap
oi8
14.3
10.4
013
.015
.99
13.7
0.86
12.6
15.1
620
0.8
29.4
12.6
238.
66
1.05
0.06
30.
931.
10A
llia
Bay
111
.6—
——
211
.3—
10.2
12.3
111
8.3
——
—1
1.14
——
—A
r.ra
mid
us1
13.0
——
—1
11.9
——
—1
154.
7—
——
11.
09—
——
A.
afar
ensi
s30
14.3
1.23
12.1
16.5
2613
.41.
0011
.115
.226
190.
027
.113
7.6
234.
126
1.06
0.06
20.
931.
18
M3 K
anap
oi6
14.6
1.19
13.7
17.0
612
.80.
6611
.913
.47
182.
126
.615
3.2
227.
87
1.1
0.15
0.80
1.27
Alli
aB
ay1
15.7
——
—1
13.7
——
—1
215.
1—
——
11.
15—
——
Ar.
ram
idus
112
.7—
——
111
.0—
——
113
9.7
——
—1
1.15
——
—A
.af
aren
sis
2215
.11.
1813
.417
.419
13.4
0.99
11.3
15.3
1819
9.0
27.0
151.
426
6.2
181.
130.
062
0.99
1.21

343AUSTRALOPITHECUS ANAMENSIS
Figure 28. Category scatterplots of lower lateral incisor dimensions in A. anamensis and A. afarensis. Pointsrepresent individual specimens. Data listed in Table 9. A. anamensis LI2s are broader mesiodistally,although they overlap in labiolingual thickness. This makes them more elongate in occlusal profile thanthose of A. afarensis. It also makes them larger on average in occlusal area, although the distributionsoverlap in this dimension.
than those of A. afarensis. This is mainlybecause they are more MD elongate.Finally, A. anamensis has a more mesio-distally elongate C/ crown than doesA. afarensis.
The preserved A. anamensis caninecrowns and roots are similar in absolute sizeto those in the A. afarensis sample, withsimilar size ranges represented (Table 9;Ward et al., 1999b). These tooth crown androot data seem to suggest that there was asimilar amount of canine size dimorphism inA. anamensis and A. afarensis. The largecanine alveolus of KNM-KP 29286, how-ever, exceeds the size that would fit even thelargest preserved A. anamensis canine root(Ward et al., 1999b). Because in extantAfrican apes and humans canine rootdimensions are tightly correlated withcrown dimensions (Pearson’s r2=0·9931,P<0·001, n=60), it is reasonable to assumethat lower canine crown size dimorphism
exceeded anything we are able to measuredirectly in A. anamensis.
Although they are broadly similar inabsolute size of preserved canine crowns androots, when canine size is considered relativeto the size of the postcanine dentition withinindividuals, it is apparent that A. anamensishas relatively larger canines than does A.afarensis (Ward et al., 1999b; Table 10).This seems to be supported by KNM-KP31713, the edentulous mandible. Eventhough it appears to have been from afemale, this specimen also has a large partialcanine root preserved (Figure 10). It doesnot appear larger in dimensions than that ofKNM-KP 29281, however, so apparentlywould not affect estimates of overall caninesize range in A. anamensis. The presence ofrelatively larger canine teeth in A. anamensis,and perhaps a greater degree of canine sizedimorphism, is more primitive than found inlater hominins.

344 . . ET AL.
Deciduous dentitionThe deciduous teeth of A. anamensis arealso similar in size to those of A. afarensis(Table 11). The most notable feature ofthe A. anamensis deciduous dentition isthe narrow dm1 (KNM-KP 34725). Thespecimen is worn, but displays a poorlydifferentiated Fp (Leakey et al., 1998). Thistooth is more similar in shape to that of Ar.ramidus and the African apes, lacking the BLexpansion typical of all other hominins. It isalmost as long MD as the longest A. afaren-sis tooth, but narrower BL than the smallest.The KNM-KP 34725 dm1 is more similarto those of A. afarensis in overall size,however (Table 12).
Crown heightsCrown heights of the incisors and caninesappear equivalent in A. anamensis and A.afarensis, although this is difficult to quantifydue to wear on most specimens. In contrast,A. anamensis molars are lower-crownedthan those of A. afarensis (Leakey et al.,1995; Hlusko, 1998). Their mandibularmolars have more sloping lingual facesand the maxillary molars more slopingbuccal faces (Leakey et al., 1995). This
Mandibular canine crown area relative to crown areas of associatedpostcanine teeth
Taxon Specimen no. C/P3 C/P4 C/M1 C/M2 C/M3
A. anamensis KNM-KP 29281 0·82 1·00 0·55 0·48 0·48KNM-KP 29284 1·08 — — — —KNM-KP 29286 0·99 1·04 0·79 0·57 0·61
Mean 0·96 1·02 0·67 0·53 0·55
A. afarensis AL 128-23 0·85 0·87 0·54 0·44 —AL 198-1 0·88 0·90 — 0·51 0·44AL 333w-58 1·20 — — — —AL 400-1a 0·70 0·68 0·46 0·36 0·37LH 3 0·71 0·74 0·67 — —LH 14 0·85 0·75 — — —
Mean 0·87 0·79 0·56 0·44 0·41
Ratios are of crown areas calculated as MD�BL.Measurements represent average of right and left sides when both were available.A. afarensis data kindly provided by W. H. Kimbel.Data figured in Ward et al. (1999b).
Table 10
appears to be due to a superobuccal expan-sion of the protoconid on the A. anamensismandibular molars (Hlusko, 1998), andpresumably a complementary apical expan-sion of the maxillary molars. The resultingdifferences in crown height may have func-tional implications, supporting interpreta-tions of selection for altered tooth use, andpresumably diet, in A. afarensis as comparedwith A. anamensis.
The discrepancy between incisor andmolar tooth crown heights, combined withwhat appear to be slightly larger canine teethon average in A. anamensis, suggest that thediscrepancy between the level of the incisaland postcanine occlusal wear planes mayhave been slightly greater in A. anamensisthan A. afarensis.
Canine/premolar complexThe maxillary canine of A. anamensis isunique in form, with strong basal tuberclesfound both mesially and distally. KNM-KP35839, the only unworn upper canine ofA. anamensis, has its mesial and distal basaltubercles at nearly the same distance fromcrown tip, sitting immediately adjacent tothe cervical margin. Both A. afarensis and

345AUSTRALOPITHECUS ANAMENSIS
Tab
le11
Com
par
ativ
ed
ecid
uou
sto
oth
met
rics
Mes
iodi
stal
Lab
io/b
ucco
lingu
alC
row
nsi
ze(M
D�
BL
)C
row
nsh
ape
(MD
/BL
)
nM
ean
Min
Max
nM
ean
Min
Max
nM
ean
Min
Max
nM
ean
Min
Max
di2 A
.an
amen
sis
14·
5—
—1
4·0
——
118
·0—
—1
1·1
——
A.
afar
ensi
s—
——
——
——
——
——
——
——
—
d cA
.an
amen
sis
26·
86·
76·
82
5·6
5·5
5·6
237
·536
·938
·12
1·22
1·22
1·21
A.
afar
ensi
s2
6·6
6·5
6·6
25·
85·
85·
82
38·0
37·7
38·3
21·
131·
121·
14
dm1 A.
anam
ensi
s1
9·3
——
16·
7—
—1
62·3
——
11·
39—
—A
.af
aren
sis
39·
18·
59·
53
7·9
7·7
8·0
371
·868
·075
·13
1·16
1·06
1·22
dm2 A.
anam
ensi
s2
9·9
9·7
10·0
27·
97·
97·
92
77·8
76·6
79·0
21·
251·
231·
27A
.af
aren
sis
211
·711
·611
·72
9·6
9·4
9·7
211
1·3
109·
011
3·5
21·
221·
231·
21
n=
sam
ple
size
,M
in=
min
imum
,M
ax=
max
imum
.

346 . . ET AL.
Tab
le12
Low
erfi
rst
dec
idu
ous
mol
arm
etri
cs
n
Mes
iodi
stal
leng
thB
ucco
lingu
albr
eadt
hC
row
nar
ea(M
D�
BL
)C
row
nsh
ape
(MD
/BL
)
Mea
nS
.D.
Min
Max
Mea
nS
.D.
Min
Max
Mea
nS
.D.
Min
Max
Mea
nS
.D.
Min
Max
A.
anam
ensi
s1
9·3
——
—6·
7—
——
62·3
——
—1·
39—
——
KN
M-K
P34
725
A.
ram
idus
17·
3—
——
4·9
——
—35
·8—
——
1·49
——
—A
RA
-VP
-1/1
29A
.af
aren
sis
49·
20·
508·
510
7·9
0·40
7·6
8·4
72·5
5·70
68·0
80·6
1·16
0·07
1·06
1·22
A.
afri
canu
s5–
78·
80·
208·
49
7·6
0·40
7·1
8·1
66·6
5·50
59·6
73·7
1·16
0·02
1·12
1·18
P.
pani
scus
217·
40·
566·
39
5·1
0·31
4·4
5·5
37·6
4·74
27·7
48·4
1·46
0·08
1·35
1·60
P.
trog
lody
tes
298·
10·
577·
09
5·2
0·35
4·6
5·8
42·2
5·19
32·9
54·5
1·58
0·10
1·42
1·83
G.
gori
lla20
11·0
0·69
9·8
127·
50·
556·
78·
982
·310
·71
71·4
108·
61·
480·
081·
321·
63P
.py
gmae
us6
9·2
0·67
8·4
107·
10·
616·
48·
166
·210
·31
53·8
82·6
1·30
0·04
1·24
1·36
H.
sapi
ens
218·
40·
477·
49
7·2
0·41
6·4
8·1
60·4
6·09
47·4
69·9
1·18
0·06
1·06
1·30
Com
para
tive
data
kind
lypr
ovid
edby
T.
D.
Whi
te.

347AUSTRALOPITHECUS ANAMENSIS
Figure 29. Upper canines of A. anamensis (KNM-KP 35839), A. ramidus (ARA-VP-6/1) and A. afarensis(AL 333x-3, LH 6 and LH 3) compared in lingual view. A. anamensis is unique in having two low butstrong lingual and distal basal tubercles, and a symmetrical profile. Adapted from White et al. (1994) bySusan Alta Martin.
Figure 30. Lower P3 and P4 of A. anamensis (KNM-KP 29286 and KNM-ER 20432), compared withthose of A. ramidus (ARA-VP-6/1) and A. afarensis (LH 3). A. anamensis resembles A. ramidus in havingunicuspid P3s and P4s with less expanded talonids than A. afarensis. Adapted from White et al. (1994) bySusan Alta Martin.
Ar. ramidus (White et al., 1994) tend tohave crown shoulders that are placed furtherapically, and that are not in the form ofdistinct tubercles (Figure 29). KNM-KP35839 also has less distinct apical crests thandoes Ar. ramidus, especially mesially. It ismore symmetrical in lingual or labial viewthan the Hadar canines, and matched mostclosely by the mostly unworn canines LH 3and 6 in symmetry. At Hadar, the mesialtubercle and marginal ridge are furtherinferiorly along the crown towards the apexthan are the distal ones. LH 3 and 6 aresimilarly symmetrical, however no publishedA. afarensis specimen displays the strongbasal tubercles seen in KNM-KP 35839
(White et al., 1994), nor as distinct a basaltubercle or ridge lingually. Metrically,the maxillary canine tooth of A. anamensisis MD elongated relative to those ofA. afarensis (Table 9), perhaps reflectingthe pronounced basal tubercles (Figure29).
Mandibular canines of A. anamensis aresimilar to those of other early hominins inform, but are MD narrower and more blade-like in shape. Their distal slope is concavein lingual or labial view, terminating in astrong basal tubercle (Ward et al., 1999b).The most similar A. afarensis specimens areLH 2 and 3, but even these have distalslopes that are convex towards the cusp apex

348 . . ET AL.
Tab
le13
Non
met
ric
crow
nfe
atu
res
(fol
low
ing
Woo
d&
Ab
bot
,19
83,
and
Woo
d,
1991
)
Max
illar
yde
ntit
ion
Man
dibu
lar
dent
itio
n
Spe
cim
enC
arib
elli
cris
taob
liqua
Spe
cim
enF
issu
repa
tter
nC
6C
7P
roto
styl
id
M1
M1
KN
M-E
R20
427
12
KN
M-E
R20
422
+0
02,
3,4
KN
M-E
R30
200
12
KN
M-E
R35
232
Y0
04
KN
M-E
R30
745
1,2
2K
NM
-KP
2928
1W
orn
Wor
nW
orn
KN
M-E
R35
238
2,3
(Wor
n)—
KN
M-K
P29
286L
Wor
n0
02
KN
M-K
P30
498R
22
KN
M-K
P29
286R
Y0
02
M2
KN
M-K
P29
287L
Wor
n1
02
KN
M-E
R77
27?0
1(W
orn)
KN
M-K
P29
287R
—1
—1
KN
M-E
R20
420
Wor
nW
orn
KN
M-K
P30
500L
Wor
n—
03
KN
M-E
R30
200
12
KN
M-K
P30
500R
Wor
n—
03
KN
M-E
R30
745
1,2
—K
NM
-KP
3171
2LT
oow
rink
led
00
3K
NM
-ER
3523
5W
orn
Wor
nK
NM
-KP
3171
2RT
oow
rink
led
00
3K
NM
-KP
3472
52,
3T
oow
rink
led
KN
M-K
P31
728
Y0
0—
M1
orM
2K
NM
-KP
3472
5Y
00
3K
NM
-ER
3523
11
3K
NM
-KP
3752
22
Bro
ken
KN
M-K
P35
842
1,2
2M
2
M3
KN
M-E
R20
423
Y(1
)0
3K
NM
-ER
2042
10
0K
NM
-ER
3523
3Y
11
4K
NM
-ER
3020
02
2K
NM
-KP
2928
1L—
10
0K
NM
-ER
3074
51,
22
KN
M-K
P29
281R
—1
03
KN
M-E
R35
236
2,3
2K
NM
-KP
2928
6LY
10
3K
NM
-KP
3049
8R2
Too
wri
nkle
dK
NM
-KP
2928
6RX
(1)
02
KN
M-K
P31
717
2T
oow
rink
led
KN
M-K
P29
287L
Y1
02
KN
M-K
P35
840
22
KN
M-K
P29
287R
Y1
03

349AUSTRALOPITHECUS ANAMENSIS
Tab
le13
Con
tin
ued
Max
illar
yde
ntit
ion
Man
dibu
lar
dent
itio
n
Spe
cim
enC
arib
elli
cris
taob
liqua
Spe
cim
enF
issu
repa
tter
nC
6C
7P
roto
styl
id
KN
M-K
P30
500L
Too
wri
nkle
d0
03
KN
M-K
P30
500R
Too
wri
nkle
d0
03
KN
M-K
P34
725
Too
wri
nkle
d1
03,
4?M
KN
M-K
P35
847
Y—
——
KN
M-E
R20
748
1—
M1
orM
2
KN
M-K
P29
282
Wor
nW
orn
Wor
nM
3 KN
M-E
R20
428
X1
(wor
n)0
1,4
KN
M-K
P29
281L
X1
04
KN
M-K
P29
281R
X1
03
KN
M-K
P29
286L
Y1
03
KN
M-K
P29
286R
Y1
03
KN
M-K
P29
287L
—T
oow
rink
led
—3
KN
M-K
P29
287R
Too
wri
nkle
dD
amag
ed0
2K
NM
-KP
3050
0LT
oow
rink
led
00
3K
NM
-KP
3050
0RT
oow
rink
led
00
3K
NM
-KP
3050
2LY
11
3K
NM
-KP
3050
2RY
01
3K
NM
-KP
3171
7T
oow
rink
led
Wor
nB
roke
n2
KN
M-K
P35
838
Bro
ken
0B
roke
nB
roke
n
All
occl
usal
nonm
etri
csc
orin
gsy
stem
desc
ribe
dby
Woo
d&
Abb
ot(1
983)
and
Woo
d(1
991)
.C
arib
elli
scor
ing
syst
em:0
=ab
sent
;1=
pito
rgr
oove
atm
esio
lingu
alco
rner
;2=
groo
ves
orri
dges
runn
ing
from
lingu
algr
oove
tom
esio
lingu
alco
rner
;3=
defin
ite
shel
fof
enam
elre
late
dto
prot
ocon
e.C
rist
aob
liqua
scor
ing
syst
em:
1=
pres
ent
and
cont
inuo
us;
2=
pres
ent
but
inte
rupt
ed;
3=
abse
nt.
C6
&C
7sc
orin
gsy
stem
:0
=ab
sent
;1
=pr
esen
t.P
roto
styl
idsc
orin
gsy
stem
:0
=ab
sent
;1
=sh
ape
and
bran
chin
gpa
tter
nof
the
mes
iobu
ccal
groo
vesu
gges
tive
ofpr
otos
tylid
form
atio
n;2
=sm
alls
helf
ofen
amel
belo
wa
dist
inct
bran
choff
the
mes
iobu
ccal
groo
ve;
3=
long
and
wel
l-de
velo
ped
enam
elsh
elf;
4=
vert
ical
groo
ves
and
ridg
eson
the
mes
iobu
ccal
surf
ace
ofth
epr
otoc
onid
.

350 . . ET AL.
Occlusal molar morphologyAustralopithecus anamensis specimens have asimilar pattern of molar cusp size as seen inA. afarensis specimens. The protocone tendsto be the largest cusp on the upper molars,and either the protoconid or metaconid onthe lower ones. There is variation in fissurepatterns (Table 13) within the A. anamensissample for two reasons. First, most unwornmolars show extensive secondary fissures,obscuring interpretation of cusp margins
and primary groove patterns. Second,cusp contact is generally fairly close, soalthough that X, Y and + patterns (Wood,1991) are seen, many teeth coded as X or Yare, in fact, very close to the + pattern.Variability in fissure pattern reinforcesthe need to be cautious when using suchfeatures for taxonomic purposes, at least forthis taxon.
Protostylid/Carabelli’s featuresAll but one of the 25 sufficiently well-preserved A. anamensis individuals displaysome form of cingular remnant features ontheir molars, visible as Carabelli’s featureson the maxillary teeth and protostylid com-plexes on the mandibular ones (Table 13)(see also Hlusko & Mahaney, 2000). Thesefeatures are usually in the form of ridges,although they generally do not reach thelevel of the occlusal surfaces. Almost allmandibular molars and the Kanapoi maxil-lary molars have a cuspule or ridge. There isno clear distinction between the Kanapoiand Allia Bay samples in cusp detail, but ingeneral the Allia Bay teeth show less pro-nounced features, with KNM-ER 30200,30745 and 35231 showing only pits on thesides of the teeth. The only individual in theA. anamensis sample that lacks any evidenceof a cingular feature is KNM-ER 20421, anisolated M3. The significance of the latterfor appreciating the near ubiquity of at leastsome types of these features is lessened bythe fact that third molars are variable, andin two associated dentitions from Kanapoiwhen features are present on M1 or M2, theyare missing on M3 (KNM-KP 29283 and30498).
The presence of Carabelli’s and proto-stylid features is common in A. afarensis, butis found only on about half of the molars,and then rarely expressed as more thanpits or mild ridges (White et al., 1981). A.africanus teeth from Sterkfontein andparticularly Makapansgat have particularlylarge cingular features (White et al., 1981),
with broader crowns superior to the basaltubercles.
The premolars reflect this difference incanine/premolar complex development(Ward et al., 1999b) (Figure 30). The A.anamensis third premolars all are more uni-cuspid than that of any A. afarensis, with themetaconid existing only as a tiny tubercle onthe transverse crest. The protoconid sits inthe middle of the occlusal surface and ishigh and sharp. In all A. afarensis P3s, themetaconid is relatively more developed inheight where preserved and in occlusal area.The A. anamensis P3 anterior fovea openslingually, as the thin mesial marginal ridgetends to have a notch along the mesiolingualedge of the tooth. This morphology, seen atboth at Kanapoi and Allia Bay, is mostclosely matched in ways by AL 128-23 andLH 3, but even these specimens are not asunicuspid as the A. anamensis P3s.
The P4s of A. anamensis tend to have lessexpanded posterior foveas than do those ofA. afarensis, although there is more overlapin these teeth than in the P3s. The Chadspecimen (Brunet et al., 1995, 1996) hasmuch more expanded premolar posteriorfoveae than does A. anamensis and evenmany A. afarensis specimens.
The maxillary premolars of A. anamensisare similar in morphology to those of A.afarensis. The P3s exhibit a similarly distinc-tive mesiocervical enamel extension on thebuccal face of the crown, which appears tobe plesiomorphic for hominins.

351AUSTRALOPITHECUS ANAMENSIS
often larger than those typical for A.anamensis. White et al. (1981) suggested thatthe cingular remnants in A. africanus maynot be primitive, but instead may be aby-product of crown expansion. The nearubiquity of cingular features in A. anamensisteeth suggests that if these features have anyphylogenetic signal, A. afarensis represents amore derived condition. Extra enamel onthe lingual sides of mandibular, and buccalsides of maxillary, molars may slightly slowwear on these low-crowned teeth whenmuch of the enamel is gone.
Tooth wearAustralopithecus anamensis teeth exhibit adistinctive pattern of wear. Evident in olderindividuals, the anterior teeth are worn veryheavily, much more so than the molars andpremolars. The KNM-KP 29281 anteriorteeth are heavily worn, with nearly half ofthe incisor and canine crowns obliteratedbut only about 1 mm-wide area of dentineexposed on M2 and none on M3. A similarpattern is also seen on the KNM-KP 29283maxilla, which has the crowns almost com-pletely obliterated on the incisors andcanine, but more than half of the occlusalenamel remains on M2 and M3. The othertwo mandibles and the associated dentitionsdo not show much tooth wear at all, andso were presumably from much youngerindividuals. Where significant amounts ofdentine are exposed on the postcanine teeth,they tend to be deeply hollowed, suggestingfairly intense molar tooth use.
All worn canines display apical wear com-bined with wear along their distal slopes.This is similar to the condition found inA. afarensis (White et al., 1981), althoughthe canine is slightly larger on average inA. anamensis. Although no mandible pre-serves these relationships well, the mor-phology of KNM-KP 28291 suggests thatthe canine tip was approximately levelwith or slightly above the incisor occlusalmargins, and so was a continuation of this
functional wear surface as in A. afarensis(White et al., 2000).
The A. anamensis molars also show astrong buccal/lingual wear discrepancy, alsofound in A. afarensis and other hominins(White et al., 1981). On the mandibularmolars, the buccal cusps both exposedentine before the lingual ones, and thereverse on maxillary molars. The unevenwear does not seem to be due to unequalmolar cusp height, because on unwornspecimens the lingual and buccal cusps aresubequal in height. This wear discrepancy isalso seen on A. anamensis premolars,especially on KNM-KP 29283 which hasthe P3 worn almost to its root lingually, butpreserves almost 6 mm of enamel from thecervical margin buccally. The P4 is alsoworn unevenly.
The only worn upper lateral incisors areassociated with the maxilla KNM-KP29283. These crowns are heavily worn withworn faces oriented labially and distally,presumably from contact with the lowercanine. Although these teeth have unusuallymesiodistally narrow roots, their wearpattern demonstrates that they are, indeed,maxillary incisors. This pattern is accentu-ated by the very worn state of the entiredentition. This pattern is not evident on anyA. afarensis incisors.
Enamel thicknessM. C. Dean is currently investigatingenamel thickness and structure in A.anamensis. The following discussion andmicrographs were kindly provided by him(Figure 31).
Enamel thickness is notoriously difficultto quantify. Enamel thickness increasesdistally through the tooth row and alsovaries among the cusps of molars. Beynon &Wood (1986) and Grine & Martin (1988)have previously published both linearmeasurements of enamel thickness, as wellas relative or average values of enamel thick-ness, where there has been an attempt to

352 . . ET AL.
Figure 31. KNM-ER 30748 above and KNM-ER 30749 below, sectioned though the protocone andhypoconid respectively. Both display relatively thick enamel. Metric data are given in text. Photographskindly provided by M. C. Dean, who is undertaking a more complete study of enamel structure inA. anamensis.

353AUSTRALOPITHECUS ANAMENSIS
scale for tooth size, in certain fossil homininand modern hominid molar teeth. Morerecently, Schwartz (2000) has also pub-lished a comprehensive data set of linearmeasurements of enamel thickness madefrom sections through the mesial cusps ofmodern human and great ape molar teeth.These measurements were also scaledusing the linear distance between the BLcementoenamel junctions (CEJ). Unfortu-nately, it is difficult to scale enamel thick-ness measurements on tooth fragmentswhere neither the complete dentine cap northe BL CEJ diameter is preserved. Quanti-fying enamel thickness in the A. anamensissample is difficult, as many specimens withbreaks exposing enamel sections are in non-standard orientations and not through cusptips, or are worn, obscuring accuratemeasurements of enamel thickness.
Enamel structure of two A. anamensismolar tooth fragments naturally fracturedclose to the axial plane of one cusp wereexamined by M. C. Dean (Figure 31).KNM-ER 30748 is a maxillary molar frac-tured through the axial Pr, and KNM-ER30749 is a mandibular molar, fracturedthrough the Hyd. As part of a more exten-sive study on the microstructure of fossilhominin enamel, each fractured surface waspolished until the dentine horn lay in thesurface plane and a 100 �m-thick section cutfrom the face. Linear measurements thatcorrespond to those defined by Beynon &Wood (1986) and Schwartz (2000) weremade to the nearest 0·1 from photo-montages of the thin sections with spreadingcalipers.
For the Pr, linear measurements ‘‘lingualocclusal basin’’ (LOB)=1·9, ‘‘linear cusptip’’ (LCT)=1·9, and ‘‘lingual cervical wall’’(LCW)=2·1. For the Hyd, linear measure-ments equivalent to ‘‘buccal occlusal basin’’(BOB)=1·6, ‘‘buccal cusp tip’’ (BCT)=1·0and ‘‘buccal cervical wall’’ (BCW)=1·8.(The measurement LCT of the Pr includes0·1 to compensate for estimated wear of this
cusp.) These linear measurements of enamelthickness are, on average, less than thosereported for robust Australopithecus by Grine& Martin (1988, Table 1.15, p. 30). Theirmeasurements ranged broadly between 2·0and 3·5. The closest match of measurementsmade on the A. anamensis fragments is withthose made by Grine & Martin on Stw 402,attributed to A. africanus. It seems clearthat molar enamel was thick in A. anamensis,but not absolutely as thick as in robustAustralopithecus.
Much has been made of the proportionsof cuspal and lateral molar enamel inAustralopithecus. In the A. anamensis Pr therewere approximately 400 daily enamel crossstriations between the dentine horn and thesurface. The lateral enamel contained 61striae with a periodicity of 7 days each.Thus, the time to form lateral and cuspalenamel was almost equal in this cusp. Like-wise in the Hyd, there were approximately400 days of cuspal enamel formation and 84striae with a periodicity of 7 days (588 days)in the lateral enamel. Neither of these molarcusps is likely to have included both the startand end of enamel formation but totalenamel formation times were 2·28 years inthe Pr and 2·72 years in the Hyd. Thegreater time for cusp formation in theKNM-KP 30749 Hyd than the KNM-KP30748 Pr suggests that this specimenbelongs more distal in the tooth row thandoes KNM-KP 30748. The near equaldivision of lateral and cuspal enamel for-mation times contrasts that described forrobust Australopithecus, where there is adominance of cuspal enamel and fewerstriae (that are more obliquely aligned to theenamel dentine junction) in the lateralenamel (Beynon & Wood 1987; Grine &Martin 1988).
HumerusThe A. anamensis humerus, KNM-KP 271(Figure 13), has been included in numerousstudies of distal humeral morphology. Some

354 . . ET AL.
have found this specimen more similar toHomo than Australopithecus or African apes(Senut & Tardieu, 1985; Baker et al., 1998;Senut, 1999). More thorough studies,however, have determined that KNM-KP271 cannot be distinguished from thoseattributed to A. afarensis, and is indeedmost similar to that taxon than any other(Feldesman, 1982; Hill & Ward, 1988;Lague & Jungers, 1996). The anteriormargin of the lateral trochlear keel is slightlyabraded almost to its distalmost extent,making this ridge appear less pronouncedthan it would have been (Figure 13). Thismay have contributed to the impressionthat this bone appears more like Homo.Discrepancies among these studies are alsodue to the fact that human and ape humeriare broadly similar while at the same timebeing quite variable, so that in most detailsthe ranges of variation overlap, obscuringapparent morphological distinctions.
One way in which hominin humericonsistently differ from those of Africanapes is in the lack of trochlear joint surfaceextension along the lateral margin of theolecranon fossa, a feature typical of Africanapes. This may be due to resisting loadsincurred during hyperextension of the elbowduring quadrupedalism. It is occasionallyfound to a certain extent in some Pongoindividuals, however. Not surprisingly, this
Percent cortical area at break of KNM-KP 271 and at 20% humeral location inextant hominoids
Taxon n % CA mean S.D. Min Max
KNM-KP 271* 1 86·5 — — —Pan troglodytes 20 70·0 9·8 51·5 87·7Gorilla gorilla 20 69·4 7·9 51·0 86·7Pongo pygmaeus 20 82·8 7·0 68·0 96·1Homo sapiens 40 65·2 7·6 47·5 82·9
Comparative data kindly provided by C. B. Ruff.H. sapiens sample from Pecos Pueblo, all individuals <40 years old.*Note that KNM-KP 271 is broken distal to what probably would have been 20%,
and so probably represents a minimum comparative value.n=sample size, S.D.=standard deviation, Min=minimum, Max=maximum.
Table 14
extension is absent in KNM-KP 271.KNM-KP 271 also lacks an elongate,superoposteriorly extended lateral humeralepicondyle which is noted for Ar. ramidus(White et al., 1994), resembling A. afarensisin this feature.
One feature of KNM-KP 271 not pre-viously noted is the extraordinarily thickcortical bone visible at the proximal break.Although it is broken slightly distal to whatwould have been 20% of its original length,it can be compared at least roughly toAfrican apes and humans. KNM-KP 271has a percent cortical area of about 86%,almost the maximum of the observed rangeof African apes and humans (Table 14), buttypical for Pongo. Because it is brokenslightly distal to where the cortical area wasmeasured for the extant specimens, its rela-tive value might be slightly inflated. Still, itwas unlikely that the cortical bone would bethicker here than at the true 80% location,if that was further proximal to the break.KNM-KP 271 is clearly a robust bone. Thefunctional or behavioral significance of thisextraordinary cortical thickness is difficultto interpret in such a small fragment. Whilea robust humeral shaft might appear tosuggest forelimb-dominated arboreality, itshould be noted that the humeri of Africanapes are not as robust but these apescertainly climb trees. Without bone length,

355AUSTRALOPITHECUS ANAMENSIS
RadiusKNM-ER 20419 is a long radius. If, forinstance, the individual to which it belongedhad body proportions like those of modernHomo, the estimated stature from regressiondata is as follows; 182·96�4·32 cm forwhite males, 175·61�0·43 cm for blackmales (regression equations from Trotter &Gleser, 1952). These estimates are high andwell above the stature means (132–151 cm)generated by McHenry (1992) from data forpresumed males of four Australopithecusspecies including A. afarensis. Rather thansuggest that these hominins were tall withmodern human proportions, we believeinstead that they were shorter in stature withrelatively long forearms, based on inferencefrom all other hominins prior to 2 Ma. Thisappears to be similar to the condition inA. afarensis, which also appears to havehad relatively long foreams (Kimbel et al.1994).
The radius is similar in morphology toother Australopithecus radii and is distin-guished from great apes by having a thickneck relative to the shaft and a distal shaftthat is semilunar in cross-section. It differsfrom African apes and humans in particularin the relative proportions of the articularsurface areas for the lunate and scaphoid(Heinrich et al., 1993). Only Australopithecushas a lunate contact facet that is larger thanthat of the scaphoid. Richmond & Strait(2000) recently used a cast of KNM-ER20419 in a multivariate analysis of distalradial morphology. They claim that thisbone retains morphological evidence ofknuckle-walking. They did not, unfortu-nately, correct for the missing styloid pro-cess, which could affect their angular and
breadth measurements and, thus, the out-come of their analysis. Even if they had donethis, the significance of the dorsal ridgelimiting extension in knuckle-walkers has yetto be demonstrated, so their results shouldremain tentative. The uniquely broad lunatefacet of Australopithecus must also be takeninto account in any functional analysis of theradiocarpal joint.
CapitateAbrasion of most of the cortex of thecapitate (KNM-KP 31724) obscures muchof its morphology. This bone was large,though, being similar in overall length tothat of a chimpanzee or female gorilla. Itappears to have had a large head, although itcannot be measured due to damage. What ispreserved of the head shows that almost allof its proximally-facing surface was com-prised by the lunate facet, as in otherAustralopithecus specimens (Ward et al.,1999a). In contrast, extant great apes andhumans have more extensive scaphoid facetshere. This morphology may be related to arelatively large area of contact between thelunate and radius, found in Australopithecus,but not in extant African apes and humans.
Enough of the specimen is intact toindicate that the MC 3 surface probably hadless topographic relief than that typical ofAfrican apes. Enough of the second meta-carpal surface is preserved to show that thisfacet was not complete dorsopalmarly as itis in later hominins, but instead there wasa ligamentous attachment area in thedorsopalmar center of the distal end of thebone separating two MC 2 facets (Leakeyet al., 1998).
One feature that is clear is that the MC 2facet was set at about a 90� angle to the MC3 facet (Leakey et al., 1998). This conditionis typical for African apes but not humans,who have their MC 2 facet set at a 112–155� angle to the MC 3 facet (Table 15;Figure 32). This obliquity, combined witha dorsopalmarly continuous MC 2 facet,
diaphyseal morphology is difficult to inter-pret with respect to bending rigidity and anybehavioral conclusion is difficult to makewithout detailed further research into thesignificance of cortical thickness in such asmall fragment of humerus.

356 . . ET AL.
Tab
le15
Com
para
tive
met
rics
ofth
eca
pita
te
n
MC
2/M
C3
face
tan
gle,
�D
orso
palm
ardi
amet
erof
head
,m
mP
roxi
mod
ista
lle
ngth
ofbo
ne,
mm
Mea
nS
.D.
Min
Max
Mea
nS
.D.
Min
Max
Mea
nS
.D.
Min
Max
A.
anam
ensi
s1
90—
——
14·8
——
—21
·8—
——
A.
afar
ensi
s2
120
—12
012
012
·2—
10·8
13·5
18·1
—16
·220
·0cf
.A
.af
aren
sis
111
5—
——
15·0
——
—20
·8—
——
Pan
trog
lody
tes
2086
7·2
7110
033
·32·
329
·236
·822
·91·
619
·325
·9G
orill
ago
rilla
2085
6·7
7397
45·5
6·1
36·0
56·4
26·4
3·2
22·1
32·2
Hom
osa
pien
s20
134
10·9
112
155
44·5
3·4
39·5
50·0
21·8
1·9
17·7
25·1
MC
2/M
C3
face
tan
gle
mea
sure
dfr
omta
ngen
tsof
palm
aras
pect
sof
thes
ejo
int
surf
aces
.cf
.A
.af
aren
sis
data
isfr
omth
eT
urkw
elho
min
id,
KN
M-W
T22
944
(War
det
al.,
1999
b).

357AUSTRALOPITHECUS ANAMENSIS
allows rotation at this joint (Lewis, 1977;Dubosset, 1981; Marzke, 1983, 1997;Marzke & Shackley, 1986). Such move-ment would not have been possible forthe KMN-KP 31724 individual (Leakeyet al., 1998; Ward et al., 1999a). Otheraustralopithecine specimens are intermedi-ate in orientation, with human-like continu-ous facets (Broom & Schepers, 1946; LeGros Clark, 1947; Lewis, 1973; McHenry,1983; Marzke, 1983, 1997; Ward et al.,1999a). In this respect, KNM-KP 31724is more great ape-like than any knownAustralopithecus or Homo specimens.
PhalanxThe manual phalanx, KNM-KP 30503, isvery similar to those of A. afarensis from
Figure 32. Orientation of second metacarpal facet on capitate. Measurement taken between MC 2 facetand palmar portion of MC 3 facet. Points represent individual specimens. Data listed in Table 15. A.anamensis is the only hominin specimen to have a laterally-facing MC 2 facet. All others overlap modernhumans in having a distolaterally inclined facet. In these later hominin specimens, the MC 2 facet is mildlyconcave and uninterrupted by an intercarpal ligament. In humans, this contributes to up to 20� of rotationat the second carpometacarpal joint, contributing to cupping of the palm. Although the facet is damagedobscuring potential evidence of an intercarpal ligament, the orientation and flat preserved contour of theMC 2 subchondral surface suggests that this motion was not possible in A. anamensis.
Hadar in size and morphology (Johansonet al., 1982; Bush et al., 1982). The speci-men exhibits a gentle palmar concavityand clear flexor ridges along the centralportion of its shaft. It shows no appreciabledifferences in form compared to those ofA. afarensis. This morphological similaritysuggests functional and developmentalhistory similarity, but whether this impliesarboreality or not remains a matter of debate(summarized in Stern, 2000).
TibiaThe KNM-KP 29285 tibia is similar tothose described for A. afarensis fromHadar (Lovejoy et al., 1982). The tibia givesclear signs of being adapted for bipedallocomotion, particularly in the vertical

358 . . ET AL.
Discussion
Taxonomy and evolutionary implicationsThe Allia Bay hominids have been con-sidered part of the A. anamensis hypodigmand the sample includes paratypes for this
species (Leakey et al., 1995, 1998; Wardet al., 1999b). The holotype of A. anamensisis from Kanapoi, where all of the featuresdescribed as diagnostic are found. The AlliaBay sample is less complete than that fromKanapoi, with fewer fossils. It is not that theAllia Bay fossils lack diagnostic features, it israther that the body parts and charactersnecessary to make a conclusive species attri-bution are not preserved at this site. Sizes ofthe preserved teeth from both sites overlap,as they do in all observable aspects of mor-phology. Given the present samples, theAllia Bay fossils preserve no morphologyindicating that they should not belong toA. anamensis.
Several Allia Bay specimens have possibletaxonomic implications. At first glance, itmight appear that the mandible KNM-ER20432 (Coffing et al., 1994) does not have asloping symphysis. This specimen is frag-mentary, however, and preserved only as faranteriorly as P3, so is missing enough boneto make it unclear what the symphysealcontour would have looked like. When castsof the more complete Kanapoi specimensare broken at the same points as KNM-ER30745, they too lose all evidence of theircharacteristic symphyseal morphology.Thus, symphyseal morphology cannotbe reconstructed accurately based onKNM-ER 20432.
This mandible does appear to be mor-phologically more similar to A. anamensismandibles than A. afarensis ones in pre-served morphology, however. The alveolarcontour of this specimen is similar to theKanapoi mandibles. Although the caninejugum is partly missing, it is clear that itwould have flared laterally at least as far asdoes the P3 jugum, a feature found onlyamong A. anamensis mandibles withinhominins.
The P3 of KNM-ER 20432 is quitesimilar to all Kanapoi P3s (Coffing et al.,1994, Figure 30). These teeth have onlya dominant, centrally placed protoconid,
orientation of the shaft relative to the talarfacet (Latimer et al., 1987; Ward et al.,1999b). Apes all have an obliquely orientedtibial diaphysis. KNM-KP 29285 is slightlylarger than the largest tibia recovered so farfrom Hadar, AL 333-42 (Lovejoy et al.,1982). Using McHenry’s (1992) regressionequation based on humans, KNM-KP29285 came from a 47–55 kg individual.
KNM-KP 29285 resembles A. afarensistibias in several other features including arectangular proximal epiphysis, concavecondyles that are roughly equal in area, amarked depression anteriorly for the pesanserinus muscles, a straight shaft, and asemilunar articular facet for the distal fibula.It also has swollen metaphyses that areexpanded in all dimensions past thesubchondral bone. This latter feature can befunctionally linked to bipedality as a mech-anism for expanding cancellous bone as ashock-absorbing mechanism to dissipatehigh peak loads incurred during heel strikein bipedal gait (Kunos & Latimer, 2000).
The superior fibular facet is abraded, butcannot have been very big. Together withthe small semilunar facet for the inferiorfibular articulation, this is evidence that thefibula may have been relatively small. Greatapes have a large fibula for attachment of thelarge flexor hallucis longus muscle associ-ated with a divergent hallux. A markedanterior depression for the pes anserinusinsertion (mistakenly described in Johansonet al. (1982) for the tibialis anterior inser-tion) is a common feature of great apes, andis found in all Australopithecus individuals inwhich it can be seen. The functional signifi-cance of this feature is under investigationby one of us (A.W.).

359AUSTRALOPITHECUS ANAMENSIS
with the metaconid present as only a smalltubercle along the marginal ridge, andstrongly demarcated cusp ridges. All ofthese teeth are very similar. This mor-phology is not matched by any A. afarensisspecimen, despite the great variability in P3
morphology in that species. This specimen,then, suggests that the Allia Bay hominidsshould remain with A. anamensis.
KNM-ER 20432 also preserves thebroken root of a very large canine, anotherA. anamensis feature. Because of the diffi-culty in quantifying broken canine rootdimensions, it is difficult to comparemetrically. Australopithecus anamensis does,however, appear to have had slightly largercanine teeth than did A. afarensis, so thischaracter also would appear to link Allia Baywith Kanapoi.
Another premolar with potential taxo-nomic implications is KNM-ER 35228, apremolar germ that we argue is P4, but hassome unusual morphology (Figure 21). It issimilar in some ways to A. afarensis P3s,which could lead some to doubt its assigna-tion as a P4. The somewhat unusual natureof KNM-ER 35228 is worth noting becauseif this tooth were a P3, it would suggest thatthe Allia Bay sample was aligned morpho-logically more closely with A. afarensis.
Compared with other A. anamensis andA. afarensis P4s, KNM-ER 35228 has amore ovoid occlusal outline with reducedmesiolingual corner, a more sloping buccalface and relatively large protoconid withsmaller lingual cusps than found in mostA. anamensis or A. afarensis P4s. Amongthe A. afarensis P4s, it is most similar tothe specimen AL 288-23, but still differsfrom it.
Although KNM-ER 35228 somewhatresembles some Hadar P3s in overall form,it has a notched lingual cusp ridge withthe anterior fovea opening lingually, a largedistal cuspule with a distinct distobuccalgroove and no mesiobuccal groove, and arelatively short mesial cusp ridge. These
features are found together only on A.afarensis or A. anamensis P4s. The greaterweight of the anatomical evidence suggeststhat this tooth is, indeed, a P4. We recognizethat KNM-ER 35228 differs from other A.anamensis P4s, however, in occlusal outlineand cusp structure.
Until more fossils are found with which totest the hypothesis of conspecificity of theAllia Bay and Kanapoi hominins, it seemsmost reasonable to retain the Allia Bayhominins in A. anamensis.
As noted by Leakey et al. (1995, 1998)the fossils attributed to A. anamensis displaya suite of characters which differentiatethem from all previously described species,including A. afarensis (contra Senut, 1996,1999; Wolpoff, 1999). This justified thenaming of the new species.
It is worthwhile to note that with thepossible exception of the strong basaltubercles on the male upper canine(Figure 29), and perhaps the basal convexityof the external mandibular symphysealcontour, there are no apomorphies of A.anamensis that are not present in A. afarensis.On cladistic grounds, then, some might betempted to argue that the A. anamensissamples should be referred to A. afarensis(e.g. Wolpoff, 1999) or Praeanthropusafricanus (e.g. Senut, 1996, 1999). We arguethat there is biological merit in distinguish-ing A. anamensis as a separate species. Com-bining these samples in one taxon does notpreclude the distinct possibility that they donot have an ancestor-descendent relation-ship, much less an exclusive one whichwould make it appropriate to combine them.Given the many similarities between thesetwo taxa, and the fact that most of the waysthat A. anamensis differs from A. afarensisappear to be in the retention of primitivetraits, a reasonable working hypothesis isthat A. anamensis was ancestral to A.afarensis. Until future discoveries of fossilsare made which disprove this hypothesis, itprovides a useful scientific framework with

360 . . ET AL.
which to explore ideas about the earlyevolution of Australopithecus.
This hypothesis that A. anamensis may beancestral to A. afarensis is consistent with thephylogeny recently published by Strait &Grine (1999) that placed A. anamensis asa sister taxon to A. afarensis and otherhominins. Traits from their list that can beassessed in A. anamensis are, by number(Strait et al., 1997): 4, anterior pillars—absent; 5, nasoalveolar clivus contour incoronal plane—convex; 6, protrusion ofincisor alveoli behind bicanine line (basalview)—yes; 7, nasal cavity entrance—stepped; 8, palate thickness—thin; 12,anterior palatal depth—shallow; 32, relativedepth of mandibular fossa—shallow; 34,configuration of tympanic—tubular; 44,mandibular cross section at M1—small; 45,orientation of mandibular symphysis—veryreceding; 46, direction of mental foramenopening—anterior/superior; 47, hollowingabove and behind mental foramen—present;49, mandibular deciduous canine shape—apex central, mesial convexity, low; 50,incisal reduction—moderate; 51, caninesreduced—somewhat/slightly less than inA. afarensis; 53/54, premolar/molar crownarea—like A. afarensis; 56, dmr of M2—low;57, separation of molar and premolar cuspapices—wide; 58, frequency of well devel-oped P3 metaconid—absent; 59, relativeenamel thickness—thick. This list doesnot summarize all characters visible in A.anamensis, however, not mentioning thesmall external auditory porus, roundedlateral nasal margins, rounded externalinferior mandibular symphysis contour,mandibular alveolar contour not curvingmedially until /C, slightly larger canine teethwith robust roots, narrow dm1, large C/basal tubercles, low-crowned molars withsloping sides, thick humeral cortex, laterally-oriented MC 2 facet on the capitate, forexample. All of these but the mandibularsymphyseal shape and C/ form are primitivefor hominins excluding Ar. ramidus, so no
further characters presented here wouldlikely alter the hypothesis supported by theirresults. A trait list that would enable one tomore thoroughly assess the phylogeneticposition of A. anamensis relative to Orrorinand Ardipithecus would have to includeother characters, and must wait until theselatter fossils have been further analyzed andpublished.
Senut has argued that the A. anamensissample be combined with the Garusi (A.afarensis) specimen in what would now bereferred to as Praeanthropus afarensis (Senut,1999) based on similarities between theGarusi maxilla and KNM-KP 29283. It istrue that these fossils share robust caninejuga contributing to a rounded lateralnasal margin, features not seen in otherA. afarensis specimens (Figure 23). A.anamensis overlaps morphologically withother A. afarensis specimens in otherfeatures, however, including in the maxilla.This could be attributable to a closephylogenetic or even ancestor-descendentrelationship. The process of populationdifferentiation and speciation involves shift-ing frequencies of traits that vary withinpopulations, so finding some traits whichoverlap should neither preclude taxonomicseparation or require grouping of sampleswhich are otherwise distinct. At the presenttime, we recognize these similarities betweenA. anamensis and the Garusi maxilla, butprefer that no taxonomic reallocation begiven until more is known about patternsof variation in these morphologies withinand among extant and fossil hominoidtaxa.
Some additional Ethiopian fossil samplesare broadly contemporareous with A.anamensis. One is the collection of heavilyworn teeth from Fejej (Fleagle et al., 1991)that were initially attributed to A. afarensis.The taxonomic affinity of this highly frag-mentary material is indeterminate, however,and this specimen may as well belong to A.anamensis, which was unknown at the time

361AUSTRALOPITHECUS ANAMENSIS
of their publication (Ward et al., 1999b).Other contemporaneous fossils are theBelohdelie frontal, dated to 3·9 Ma (Asfaw,1987), and the Maka femur, dated to 3·5–4·0 Ma (Clark et al., 1984), but becauseno frontals or femora are known for A.anamensis, direct comparisons cannot bemade to assess possible taxonomic affinitiesamong these samples.
DietDietary implications of the Kanapoi andAllia Bay fossils are summarized by Teaford& Ungar (2000). The thick enamel of A.anamensis teeth compared with the thinenamel of Ar. ramidus and African apessuggests a shift in emphasis towards eatingharder or more abrasive foods, while retain-ing an ability to process softer fruits. This isalso supported by the apparently reducedlevels of anterior tooth wear, slight reductionin anterior teeth, flat molars and slightlythicker mandibular corpora than found inapes. Later australopithecines appear tohave continued trends toward harder objectfeeding. Thus, selection for the ability toprocess harder food objects seems to bea trend that began in the ancestors of A.anamensis and continued in later Australo-pithecus. A. anamensis then, would perhapshave been able to process foods with a morevariable set of material properties than canAfrican apes, perhaps widening the dietaryniches and microhabitats that they werecapable of exploiting.
The dietary shift suggested by the A.anamensis dentognathic morphology may berelated to environmental shifts occurringat the time. Evidence for dietary drivenadaptive radiations at this time is not limitedto A. anamensis or even to fossil hominins.Faunal accumulations from East Africadocument a major turnover in the lateMiocene. Fauna characteristic of theMiocene was replaced by new taxa of mod-ern aspect, incorporating the last occurrenceof archaic Miocene taxa with ancestors of
the extant biota including hippos, giant pigs,grazing antelopes, true giraffes andelephants (Leakey & Harris, 2001). Thisfaunal change appears to be directly relatedto changing habitats resulting from theexpansion of C4 grasses (Cerling et al.,1997a). Evidence for changing paleo-ecology has been indicated through analysesof stable carbon isotopes in pedogeniccarbonates and in tooth enamel of herbivor-ous mammals. Both approaches indicate aglobal event, marking the expansion of C4
plants (Quade et al., 1992; Cerling et al.,1997a, 1998). This is believed to be theresult of decreasing CO2 levels reaching athreshold that favors C4 photosynthesis(Ehleringer et al., 1997; Cerling et al.,1998). Stable isotope studies of paleo-ecosystems have shown that few C4 plantswere present in significant abundance any-where prior to about 8 mya (Morgan et al.,1994; Cerling et al. 1997b). After this time,however, mixed C3/C4 habitats with a sig-nificant C4 component became widespread(Quade et al., 1992; Cerling et al., 1997a).
Although the fossil record provides littleevidence of increased diversity in non-mammalian species, evidence for faunalchange and dietary-driven adaptive radia-tions among mammals in the late Mioceneand Pliocene has been well demonstrated insites as widely separated geographically asthe Siwaliks, North America and Africa(Quade et al., 1989, 1992; Morgan et al.,1994; Cerling et al., 1997a). The earliestAfrican mammalian assemblages to exploitC4 plants are found in the Lothagamsediments and Samburu Hills succession(Cerling et al., 2001). Lothagam sedimentsrange in age from greater than 7·4 Ma at thebase of the lower Nawata Formation to4·2 Ma at the top of the Apak Member ofthe Nachukui Formation (McDougall &Feibel, 1999). Samburu Hills overlaps inage with the earliest Lothagam sediments.Several lineages of herbivores includingproboscideans, equids, rhinos and suids

362 . . ET AL.
show a change from C3 to C4 dominateddiets (Cerling et al., 2001). In contrast,analyses of pedogenic carbonates andmammalian tooth enamel from earlier sedi-ments at Nakali (Cerling et al., 2001),Ngorora (Morgan et al., 1994), and FortTernan (Cerling et al., 1997b) that predateLothagam, show little evidence of C4 photo-synthesizing plants (Cerling et al., 2001).The emergence of hominins in the lateMiocene or early Pliocene may be part ofthis global phenomenon in which changingecosystems prompted a radiation of speciesusing the newly available habitats and foodresources.
LocomotionThe Kanapoi tibia clearly demonstratesthat A. anamensis was adapted for habitualbipedality (Leakey et al., 1995; Leakey et al.,1998; Ward et al., 1999b). This tibia is theearliest conclusive postcranial evidence ofhominin bipedality documented so far,showing that habitual terrestrial bipedallocomotion was selectively advantageousfor hominins prior to 4·2 Ma. Thisadaptation may be older, as Ar. ramidus isalleged to have been bipedal at 4·4 Mabased on the position of its foramenmagnum (White et al., 1994), and analysisof its postcranial anatomy currently under-way will reveal the extent of its bipedaladaptation.
The apparently long radius and mildlycurved manual phalanx represent primitivecharacters retained in the skeleton. The sig-nificance of these features for interpretingbehavior and adaptive history is compli-cated, however. There are two major ques-tions commonly asked about locomotion inAustralopithecus. First, what type of locomo-tion had been selected for in a particularspecies, and second, what were those ani-mals capable of doing and/or spent timedoing. These are fundamentally differentquestions requiring different types of evi-dence to address. The adaptive significance
of primitive retentions is not readily testable,as it is difficult to refute either the hypothesisthat plesiomorphic characters were retainedby stabilizing selection or that they merelyhad not been selected against. This isparticularly difficult in Australopithecus inlight of the clear and strong directionalsignal towards bipedality and away fromarboreality. Because the ape-like charactersof the A. anamensis skeleton are primitive, itis not possible to say conclusively whether ornot they indicate that arboreality was anadaptively significant component of the A.anamensis behavioral repertoire at this point.It will always be difficult to say, even if A.anamensis climbed trees, whether this behav-ior enhanced their survival and reproductivesuccess, or whether they were selectivelyneutral because of their primitive nature,especially given the overwhelming shifttowards terrestrial bipedality evident in theskeleton.
The second question is slightly easier, butstill requires more comparative morphologi-cal data. Finding characters with a strongepigenetic component, and thus influencedby an individual’s actual activity patternsover its lifetime, could possibly demon-strate that an animal may have engaged ina particular behavior. Recently, possibleontogenetic influences of climbing behavioron phalangeal curvature have been sug-gested (Paciulli 1995; Richmond, 1997,1999) but have yet to be supported convinc-ingly by data in the literature. If thesedata are supported, phalangeal curvaturecould indicate occasional climbing in A.anamensis, although still we could not besure that behavior influenced the indi-vidual’s fitness. Some have argued, in fact,that not only were they not selected to do so,Australopithecus (A. afarensis and the othermore recent species, at least) could nothave climbed trees regularly (summarized byLatimer, 1991).
Clearly, bipedality had been selectedfor and positively affected individual

363AUSTRALOPITHECUS ANAMENSIS
survival and reproduction in the immediateancestors of A. anamensis. There is noconclusive evidence to suggest thatarboreality was practiced and/or remainedselectively advantageous for A. anamensis,although the possibility cannot be ruled outat present.
Body mass and sexual dimorphismThe apparently large body size range in theA. anamensis sample is analogous to thatfound in A. afarensis, which has roughly2:1 body mass sexual dimorphism (e.g.McHenry, 1992) comparable to the levelstypical of Gorilla gorilla and Pongo pygmaeus(Plavcan & Van Schaik, 1997; Smith &Jungers, 1997). This far exceeds body massdimorphism levels found in Homo andPan, suggesting that reduced body massdimorphism represents homoplasy inthese lineages (Lockwood et al., 1996).These data contradict hypotheses positingthat humans and Pan troglodytes (e.g.,Wrangham, 1999) or Pan paniscus (e.g.,Zihlman et al., 1978) share extensive aspectsof their sociality as an evolutionary heritage.
Canine dimorphism in A. anamensis wascertainly reduced compared with extant apesand even Ar. ramidus, revealing a history ofcontinued selection against large canine sizein males early in the hominin lineage. Ascanines are primarily important for intra-specific displays and combat among males inextant primates, it is reasonable to assumethat canine teeth were less valuable asinstruments in male–male combat. Thistrend continued in A. afarensis and laterhominins.
The fact that body mass dimorphismremained high until the appearance ofHomo, however, suggests that large malebody size (and/or small female size) had notbeen selected against for all known Australo-pithecus species, and may still have conferreda reproductive advantage, although, similarto interpretations of locomotor morphology,this plesiomorphic character is complicated
to assess. The presence of high levels ofbody mass dimorphism is associated withpolygynous social systems in all anthropoids(see Plavcan, 2000), and would seem tocontradict the hypothesis that these earlyhominins had a pair-bonded social system(Lovejoy, 1981), even in light of reducedcanine dimorphism (Plavcan, 2000). Largebody size appears to have retained anadvantage for males while canine size didnot, suggesting an altered emphasis onstrategies of male-male combat rather thanan elimination of such competition.
It is interesting to note that all five of theknown postcranial elements of A. anamensisare from large, presumably male, individ-uals. They all compare in size with thelargest known specimens of A. afarensis fromHadar. Any comparisons made between A.anamensis postcrania and those of other taxamust take body mass into account whenmaking functional interpretations based onthis apparently biased small sample.
Summary and conclusions
Australopitheus anamensis predates all othertaxa that have been assigned to Australo-pithecus, and in most ways it appears to bemore primitive than any other. Although it isreadily distinguishable from A. afarensis, thisis because A. anamensis is generally moreprimitive in all except its uniquely largemaxillary canine basal tubercles androunded inferior contour of the externalsurface of its mandibular symphysis. Thepossibility that A. anamensis represents anancestor of A. afarensis is not precluded bythe evidence available to date.
Because the Allia Bay sample is morepoorly represented than the Kanapoisample, it preserves few diagnostic charac-ters for A. anamensis. However, because itis most similar to the Kanapoi hominidsmorphologically, and is close to it geo-graphically, we recommend it be retained, atleast for the moment, in A. anamensis.

364 . . ET AL.
Australopitheus anamensis was a habitualbiped that retained some primitive charac-ters in its upper limb. It has thick enamel,with anterior teeth considerably reducedfrom the putative primitive condition. Thelower lateral incisors are relatively largerthan those of Australopitheus afarensis, andthe canine/premolar complex is more primi-tive with apparently slightly larger caninesrelative to the postcanine teeth and moreunicuspid lower third premolars. The man-dibular canines are narrower and moreasymmetrical in form, and the maxillarycanine displays a unique morphology withsubequal mesial and distal basal cuspules.
Australopithecus anamensis jaws are nar-rower than those of other Australopithecus,with more parallel postcanine tooth rows.Mandibular robusticity is slightly less thanin A. afarensis. The A. anamensis mandiblesalso differ from those of later hominins inthat the lateral alveolar contour swingsmesially only anterior to the canine juga, notat the P3 juga. The lateral nasal aperture isrounded and is smoothly continuous withthe robust maxillary canine juga, while thatof A. afarensis is separate and there is aprominent lateral nasal crest. The onlyexception to this in A. afarensis is the Garusispecimen from Laetoli, which more closelyresembles A. anamensis in nasal contourform.
While its thick enamel and reducedanterior dentition suggest a dietary shifttowards tougher foods from a primitive,ape-like condition, A. anamensis retains afairly ape-like structure to its jaws withnarrowly set parallel postcanine toothrows, a flat temporomandibular joint andheavy anterior tooth wear. Whatever dietarychange corresponded with the appearanceof Australopithecus, dietary adaptation con-tinued to change with A. afarensis, causingthe development of a more parabolic dentalarcade, less flat glenoid fossa and associatedmorphology. This dietary change mayreflect a long term environmental trend
occurring at the time from closed to moreopen environments. The reduced caninesexual dimorphism combined with appar-ently high levels of body mass dimorphismsuggests a unique social system character-ized by high levels of male–male competitionthat did not involve use of the canine teeth.Thus, data from A. anamensis supporthypotheses suggesting that the evolution ofhominid bipedality may have been corre-lated both with changes in diet and/or socialbehavior.
Acknowledgements
The authors would like to thank the follow-ing people and institutions for their adviceand/or support. The Government of Kenyagave us permission to carry out this researchand the National Museums of Kenya gavelogistical support. We thank the directorand staff of the National Museums ofKenya, the staff of the National Museumof Ethiopia and Ethiopian Ministry ofInformation and Culture, and the curatorsand staff of the Cleveland Museum ofNatural History. We also thank FrankBrown, Deborah Cunningham, Chris Dean,Holly Dunsworth, Victoria Emsch, CraigFeibel, Leslea Hlusko, Bill Kimbel, FrancisKirera, Charlie Kunos, Bruce Latimer,Charlie Lockwood, Ian MacDougall,Lyman Jellema, Donald Johanso, RichardLeakey, Emma Mbua, Anne Mwai, ThierraNalley, Erica Phillips, Richard Sherwood,Fred Spoor, Gen Suwa, Peter Ungar, andTim White for help. Members of the fieldexpeditions to Kanapoi (1994–1997) andAllia Bay (1995–1997) included the follow-ing: Kiptalam Cheboi, Sila Dominic, JustusEdung, Gabriel Ekalalei, John Ekamais,Christopher Epat, Nicolas Ewalan, CraigFeibel, Patrick Gathogo, Sabine Hagemann,Leslea Hlusko, John Kaatho, Peter Kapoko,Christopher Kiarie, Musa Kieva, KamoyaKimeu, Katey Coffing, Simon Komolo,Benson Kyongo, Louise Leakey, William

365AUSTRALOPITHECUS ANAMENSIS
Lokipi, Daniel Lokitar, Robert Lokure,Robert Lorinyok, Eluid Losike, NinianLowis, Simon Makathimo, Mathew Eragae,Boniface Malika, Wambua Mangao, IanMcDougall, Simon Milledge, GertrudeMugaisi, Stephen Muge, Joseph Mutaba,Mwongela Muoka, Daniel Mutinda, NzubeMutwiwa, Wycliffe Mutwiwa, NjerogeNjigi, Samuel Ngui, Blasto Onyango, FredSpoor, Kathlyn Stewart and JonathanWynn. Input by Charlie Lockwood, BillKimbel, an anonymous reviewer and TerryHarrison greatly improved this manuscript.The National Geographic Society andNational Science Foundation (NSF SBR9601025) funded the fieldwork. Caltex(Kenya) provided fuel and Richard andJonathan Leakey each allowed us the useof their aeroplanes. This research was alsosupported by NSF BNS 94-04813. SusanAlta Martin did the illustrations.
References
Asfaw, B. (1987). The Belohdelie frontal: new evidenceof early hominid cranial morphology from the Afar ofEthiopia. J. hum. Evol. 16, 611–624.
Baker, E. W., Malyango, A. A. & Harrison, T. (1998).Phylogenetic relationships and functional mor-phology of the distal humerus from Kanapoi, Kenya.Am. J. phys. Anthrop. 26 (Suppl.), 66.
Beynon, A. D. & Wood, B. A. (1986). Variation inenamel thickness and structure in East Africanhominids. Am. J. phys. Anthrop. 70, 177–193.
Beynon, A. D. & Wood, B. A. (1987). Patterns andrates of enamel growth in the molar teeth of earlyhominids. Nature 326, 493–496.
Broom, R. & Schepers, G. W. H. (1946). The SouthAfrican Ape-Men: the Australopithecinae. Pretoria:Transvaal Museum Memoirs No. 2.
Brown, B., Walker, A., Ward, C. V. & Leakey, R. E.(1993). New Australopithecus boisei calvaria from EastLake Turkana, Kenya. Am. J. phys. Anthrop. 91,137–159.
Brunet, M., Beauvilain, A., Coppens, Y., Heintz, E.,Moutaye, A. H. E. & Pilbeam, D. (1995). The firstaustralopithecine 2,500 kilometres west of the RiftValley (Chad). Nature 378, 273–275.
Brunet, M., Beauvilain, A., Coppens, Y., Heintz, E.,Moutaye, A. H. E. & Pilbeam, D. (1996). Australo-pithecus bahrelghazali, une nouvelle especed’Hominide ancien de la region de Koro Toro(Tchad). C. r. Acad. Sci., Paris. 322, 907–913.
Bush, M. E., Lovejoy, C. O., Johanson, D. C. &Coppens, Y. (1982). Hominid carpal, metacarpaland phalangeal bones recovered from the HadarFormation: 1974–1977 collections. Am. J. phys.Anthrop. 57, 651–678.
Cerling, T. E., Harris, J. M., MacFadden, B. J.,Leakey, M. G., Quade, J., Eisenmann, V. &Ehleringer, J. R. (1997a). Pattern and significance ofglobal ecologic change in the late Neogene. Nature389, 153–158.
Cerling, T. E., Harris, J. M., Ambrose, S. H., Leakey,M. G. & Solounias, N. (1997b). Dietary andenvironmental reconstruction with stable isotopeanalysis of herbivore tooth enamel from the Miocenelocality of Fort Ternan, Kenya. J. hum. Evol. 33,635–650.
Cerling, T. E., Ehleringer, J. R. & Harris, J. M. (1998).Carbon dioxide starvation, the development of C4
ecosystems, and mammalian evolution. Phil. Trans.R. Soc. Lond. B. 353, 159–171.
Cerling, T. E., Harris, J. M. & Leakey, M. G. (2001).Isotope paleoecology of the Nawata and NachukuiFormations at Lothagam, Turkana Basin, Kenya. In(M. G. Leakey & J. M. Harris, Eds) Lothagam theDawn of Humanity. New York: Columbia UniversityPress.
Clark, J. D., Asfaw, B., Assefa, G., Harris, J. W. K.,Kurashina, H., Walter, R. C., White, T. D. &Williams, M. A. J. (1984). Palaeoanthropologicaldiscoveries in the Middle Awash Valley, Ethiopia.Nature 307, 423–428.
Coffing, K., Feibel, C., Leakey, M. & Walker, A.(1994). Four-million year old hominids from EastLake Turkana, Kenya. Am. J. phys Anthrop. 93,55–65.
Daegling, D. J. & Grine, F. E. (1991). Compact bonedistribution and biomechanics of early hominidmandibles. Am. J. phys Anthrop. 86, 321–340.
Day, M. H. (1978). Functional interpretations of themorphology of postcranial remains of early Africanhominids. In (C. Jolly, Ed.) Early Hominids of Africa,pp. 311–345. New York: St. Martin’s.
DiGiovanni, B. F., Scoles, P. V. & Latimer, B. M.(1989). Anterior extension of the thoracic vertebralbodies in Scheuermann’s kyphosis: An anatomicstudy. Spine 14, 712–716.
Dubosset, J. F. (1981). Finger rotation during prehen-sion. In (R. Tubiana, Ed.) The Hand, pp. 202–206.Philadelphia: W.B. Saunders.
Ehleringer, J. R., Cerling, T. E. & Helliker, B. R.(1997). C4 photosynthesis, atmospheric CO2 andclimate. Oecologia 112, 285–299.
Falk, D. (1986). Evolution of cranial blood drainagein hominids: Enlarged occipital/marginal sinusesand emissary foramina. Am. J. phys. Anthrop. 70,311–324.
Falk, D. (1988). Enlarged occipital/marginal sinusesand emissary foramina: Their significance in hominidevolution. In (F. E. Grine, Ed.) Evolutionary Historyof the ‘‘Robust’’ Australopithecines, pp. 85–96. NewYork: Aldine de Gruyter.

366 . . ET AL.
Falk, D. & Conroy, G. C. (1983). The cranial venoussinus system in Australopithecus afarensis. Nature 306,779–781.
Feldesman, M. R. (1982). Morphometric analysis ofthe distal humerus of some Cenozoic catarrhines: theLate Divergence hypothesis revisited. Am. J. phys.Anthrop. 59, 73–95.
Fleagle, J. G., Rasmussen, D. T., Yirga, S., Bown,T. M. & Grine, F. E. (1991). New hominid fossilsfrom Fejej, Southern Ethiopia. J. hum. Evol. 21,145–152.
Grine, F. E. & Martin, L. B. (1988). Enamel thick-ness and development in Australopithecus and Paran-thropus. In (F. E. Grine, Ed.) Evolutionary History ofthe ‘‘Robust’’ Australopithecines, pp. 3–42. New York:Aldine de Gruyter.
Harris, J. M. (1987). Summary. In (M. D. Leakey &J. M. Harris, Eds) Laetoli: a Pliocene Site in NorthernTanzania, pp. 524–531. Oxford: Oxford UniversityPress.
Heinrich, R. E., Rose, M. D., Leakey, R. E. & Walker,A. (1993). Hominid radius from the middle Plioceneof Lake Turkana, Kenya. Am. J. phys. Anthrop. 92,139–148.
Hill, A. & Ward, S. (1988). Origin of the Hominidae:the record of African large hominoid evolutionbetween 14 My and 4 My. Yearb. phys. Anthrop. 31,49–83.
Hlusko, L. J. (1998). Euclidean distance matrixanalysis of Australopithecus molars. M.A. Thesis,Pennsylvania State University, University Park.
Hlusko, L. J. & Mahaney, M. C. (2000). The geneticsof expression of two dental traits in baboons. Am. J.phys. Anthrop. 30 (Suppl.), 180.
Hylander, W. L., Ravosa, M. J., Ross, C. F., Wall,C. E. & Johnson, K. R. (2000). Symphyseal fusionand jaw-adductor muscle force: an EMG study. Am.J. phys. Anthrop. 112, 469–492.
Johanson, D. C., Lovejoy, C. O., Kimbel, W. H.,White, T. D. & Ward, S. C. (1982). Morphology ofthe Pliocene partial hominid skeleton (A. L. 288-1)from the Hadar formation, Ethiopia. Am. J. phys.Anthrop. 57, 403–452.
Kimbel, W. H. (1984). Variation in the pattern ofcranial venous sinuses and hominid phylogeny.Am. J. phys. Anthrop. 63, 243–263.
Kimbel, W. H., Johanson, D. C. & Rak, Y. (1994). Thefirst skull and other new discoveries of Australo-pithecus afarensis at Hadar, Ethiopia. Nature 368,449–451.
Kohl-Larsen, L. (1943). Auf den Spuren desVormenschen; Forschungen Fahrten und Erlebnisse inDeutsch-Ostafrika. Stuttgart: Strecker & Schroder.
Kunos, C. A. & Latimer, B. (2000). Cross-sectionalgeometric properties of the distal tibial metaphysis inhumans and apes. Am. J. phys. Anthrop. 30 (Suppl.),202–203.
Lague, M. R. & Jungers, W. L. (1996). Morphometricvariation in Plio-Pleistocene hominid distal humeri.Am. J. phys. Anthrop. 101, 401–427.
Latimer, B. (1991). Locomotor adaptations in Australo-pithecus afarensis: the issue of arboreality. In (Y.
Coppens & B. Senut, Eds) Origine(s) de la Bipediechez les Hominides, pp. 169–176. Paris: CentreNational de la Recherche Scientifique.
Latimer, B., Ohman, J. C. & Lovejoy, C. O. (1987).Talocrural joint in African hominoids: implicationsfor Australopithecus afarensis. Am. J. phys. Anthrop.74, 155–175.
Le Gros Clark, W. E. (1947). Observations on theanatomy of the fossil Australopithecinae. J. Anat. 81,300–333.
Leakey, M. G. & Harris, J. M. (2001). Lothagam: TheDawn of Humanity. New York: Columbia UniversityPress.
Leakey, M. G. & Walker, A. (1997). Early hominidfossils from Africa. Sci. Am. 276, 74–79.
Leakey, M. G., Feibel, C. S., McDougall, I. & Walker,A. (1995). New four-million-year-old hominidspecies from Kanapoi and Allia Bay, Kenya. Nature376, 565–571.
Leakey, M. G., Feibel, C. S., McDougall, I., Ward,C. V. & Walker, A. (1998). New specimens andconfirmation of an early age for Australopithecusanamensis. Nature 363, 62–66.
Leakey, R. E. F. & Walker, A. (1985). Furtherhominids from the Plio-Pleistocene of Koobi Fora,Kenya. Am. J. phys. Anthrop. 67, 135–63.
Leakey, R. E. F. & Walker, A. (1988). New Australo-pithecus boisei specimens from East and West LakeTurkana, Kenya. Am. J. phys. Anthrop. 76, 1–24.
Lewis, O. (1973). The hominoid os capitatum, withspecial reference to the fossil bones from Sterkfonteinand Olduvai Gorge. J. hum. Evol. 2, 1–12.
Lewis, O. J. (1977). Joint remodelling and the evol-ution of the human hand. J. Anat. 123, 157–201.
Lockwood, C. A., Richmond, B. G., Jungers, W. L. &Kimbel, W. H. (1996). Randomization proceduresand sexual dimorphism in Australopithecus afarensis.J. hum. Evol. 31, 537–548.
Lockwood, C. A., Kimbel, W. H. & Johanson, D. C.(2000). Temporal trends and metric variation in themandibles and dentition of Australopithecus afarensis.J. hum. Evol. 39, 23–55.
Lovejoy, C. O. (1981). The origin of man. Science 211,341–350.
Lovejoy, C. O., Johanson, D. C. & Coppens, Y.(1982). Hominid lower limb bones recovered fromthe Hadar Formation: 1974–1977 collections. Am. J.phys. Anthrop. 57, 679–700.
MacFadden, B. J., Solounias, N. & Cerling, T. E.(1999). Ancient diets ecology and extinction of 5million year old horses from Florida. Science 283,824–827.
Marzke, M. W. (1983). Joint functions and grips of theAustralopithecus afarensis hand, with special referenceto the region of the capitate. J. hum. Evol. 12,197–211.
Marzke, M. W. (1997). Precision grips, hand mor-phology, and tools. Am. J. phys. Anthrop. 102, 91–110.
Marzke, M. W. & Shackley, M. S. (1986). Hominidhand use in the Pliocene and Pleistocene: evidencefrom experimental archeology and comparativemorphology. J. hum. Evol. 15, 439–460.

367AUSTRALOPITHECUS ANAMENSIS
McCollum, M. A., Grine, F. E., Ward, S. C. & Kimbel,W. H. (1993). Subnasal morphological variation inextant hominoids and fossil hominids. J. hum. Evol.24, 87–111.
McDougall, I. & Feibel, C. S. (1999). Numerical agecontrol for the Miocene-Pliocene succession atLothagam, a hominoid-bearing sequence in thenorthern Kenya Rift. J. Geol. Soc. 156, 731–745.
McHenry, H. M. (1983). The capitate of Australo-pithecus afarensis and A. africanus. Am. J. phys.Anthrop. 62, 187–198.
McHenry, H. M. (1992). Body size and proportions inearly hominids. Am. J. phys. Anthrop. 87, 407–431.
Morgan, M. E., Kingston, J. D. & Marino, B. D.(1994). Carbon isotopic evidence for the emergenceof C4 plants in the Neogene from Pakistan andKenya. Nature 367, 162–165.
Paciulli, L. M. (1995). Ontogeny of phalangeal curva-ture and positional behavior in chimpanzees. Am. J.phys. Anthropol. Suppl. 20, 165.
Patterson, B. & Howells, W. W. (1967). Hominidhumeral fragment from early Pleistocene of North-western Kenya. Science 156, 64–66.
Plavcan, J. M. (2000). Inferring social behavior fromsexual dimorphism in the fossil record. J. hum. Evol.39, 327–344.
Plavcan, J. M. & Van Schaik, D. P. (1997). Interpretinghominid behavior on the basis of sexual dimorphism.J. hum. Evol. 32, 345–374.
Puech, P.-F. (1986). Australopithecus afarensis Garusi 1,diversite et specialisation des premiers Hominidesd’apres les caracteres maxillo-dentaires. C. r. Acad.Sci., Paris 303, 1819–23.
Puech, P.-F., Cianfarani, F. & Roth, H. (1986).Reconstruction of the maxillary dental arcade ofGarusi Hominid 1. J. hum. Evol. 15, 325–32.
Quade, J., Cerling, T. E. & Bowman, J. R. (1989).Development of Asian monsoon revealed by markedecological shift during the latest Miocene in northernPakistan. Nature 342, 163–166.
Quade, J., Cerling, T. E., Morgan, M. M., Pilbeam,D. R., Barry, J., Chivas, A. R., Lee-Thorp, J. A. &van der Merwe, N. J. (1992). A 16 million yearrecord of paleodiet using carbon and oxygen isotopesin fossil teeth from Pakistan. Chem. Geol. (IsotopeGeoscience Section) 94, 183–192.
Reed, K. (1997). Early hominid evolution and ecologi-cal change through the African Plio-Pleistocene.J. hum. Evol. 32, 289–322.
Richmond, B. G. (1997). Ontogeny of phalangealcurvature and locomotor behavior in lar gibbons.Am. J. phys. Anthropol. Suppl. 24, 197.
Richmond, B. G. (1999). Reconstructing locomotorbehavior in early hominids: evidence from primatedevelopment. J. hum. Evol. 36, A20.
Richmond, B. G. & Strait, D. S. (2000). Evidence thathumans evolved from a knuckle-walking ancestor.Nature 404, 382–5.
Schwartz, G. T. (2000). Taxonomic and functionalaspects of the patterning of enamel thickness distri-bution in extant large-bodied hominoids. Am. J.phys. Anthrop. 111, 221–244.
Senut, B. (1980). New data on the humerus and itsjoints in Plio-Pleistocene hominids. Coll. Anthrop. 1,87–93.
Senut, B. (1996). Pliocene hominid systematics andphylogeny. S. Afr. J. Sci. 92, 165–166.
Senut, B. (1999). Les humains les plus anciens. LesOrigines de l’Humanite, Dossier Hors-serie Janvier,66–69.
Senut, B. & Tardieu, C. (1985). Functional aspects ofPlio-Pleistocene hominid limb bones: implicationsfor taxonomy and phylogeny. In (E. Delson, Ed.)Ancestors: The Hard Evidence, pp. 193–201. NewYork: Alan R. Liss.
Smith, R. J. & Jungers, W. L. (1997). Body mass incomparative primatology. J. hum. Evol. 32, 523–559.
Stern, J. T. (2000). Climbing to the top: A personalmemoir of Australopithecus afarensis. Evol. Anthrop. 9,113–133.
Strait, D. S. & Grine, F. E. (1999). Cladistics and earlyhominid phylogeny. Science 285, 1210–1211.
Strait, D. S., Grine, F. E. & Moniz, M. A. (1997). Areappraisal of early hominid phylogeny. J. hum. Evol.32, 17–82.
Teaford, M. F. & Ungar, P. S. (2000). Diet and theevolution of the earliest human ancestors. Proc. natl.Acad. Sci. 97, 13506–13511.
Trotter, M. & Gleser, G. C. (1952). Estimates ofstature from long bones of American Whites andNegroes. Am. J. phys. Anthrop. 10, 463–514.
Walker, A. (1978). Functional anatomy of oral tissues:mastication and deglutition. In (J. H. Shaw, E. A.Sweeney, C. C. Cappucino & S. M. Meller, Eds)Textbook of Oral Biology, pp. 227–296. Philadelphia:W. B. Saunders.
Ward, C. V., Leakey, M. G., Brown, B., Brown, F.,Harris, J. & Walker, A. (1999a). South Turkwel: Anew Pliocene hominid site in Kenya. J. hum. Evol.36, 69–95.
Ward, C. V., Walker, A. & Leakey, M. G. (1999b). Thenew hominid species Australopithecus anamensis. Evol.Anthrop. 7, 197–205.
White, T. D. (1977). New fossil hominids fromLaetolil, Tanzania. Am. J. phys. Anthrop. 46,197–230.
White, T. D. & Johanson, D. C. (1982). Pliocenehominid mandibles from the Hadar Formation,Ethiopia: 1974–1977. Am. J. phys. Anthropol. 57,501–544.
White, T. D., Johanson, D. C. & Kimbel, W. H.(1981). Australopithecus africanus: Phyletic positionreconsidered. S. Afr. J. Sci. 77, 445–470.
White, T. D., Suwa, G., Hart, W. K., Walter, R. C.& WoldeGabriel, G. (1993). New discoveries ofAustralopithecus at Maka in Ethiopia. Nature 366,261–265.
White, T. D., Suwa, G. & Asfaw, B. (1994). Australo-pithecus ramidus, a new species of early hominid fromAramis, Ethiopia. Nature 371, 306–312.
White, T. D., Suwa, G., Simpson, S. & Asfaw, B.(2000). Jaws and teeth of Australopithecus afarensisfrom Maka, Middle Awash, Ethiopia. Am. J. phys.Anthrop. 111, 45–68.

368 . . ET AL.
WoldeGabriel, G., White, T. D., Suwa, G., Renne, P.& de Heinzelin, J. (1994). Ecological and temporalplacement of early Pliocene hominids at Aramis,Ethiopia. Nature 371, 330–333.
Wolpoff, M. H. (1999). Paleoanthropology, 2nd Edition.Boston: McGraw-Hill.
Wood, B. (1991). Koobi Fora Research Project Volume 4:Hominid Cranial Remains. Oxford: ClarendonPress.
Wood, B. & Abbott, S. A. (1983). Analysis of thedental morphology of Plio-Pleistocene hominids. I.
Mandibular molars—crown area measurements andmorphological traits. J. Anat. 161, 1–35.
Wrangham, R. W. (1999). Evolution of coalitionarykilling. Yearb. phys. Anthrop. 42, 1–30.
Wynn, J. G. (2000). Paleosols, stable carbon isotopes,and paleoenvironmental interpretation of Kanapoi,northern Kenya. J. hum. Evol. 39, 411–432.
Zihlman, A., Cronin, J., Cramer, D. & Sarich, V. M.(1978). Pygmy chimpanzee as a possible prototypefor the common ancestor of humans, chimpanzeesand gorillas. Nature 275, 744–746.





























