Embed Size (px)
Citation preview

Analele ştiinţifice ale Universităţii “Al. I. Cuza” Iaşi Tomul LV, fasc. 2, s.II a. Biologie vegetală, 2009
MORPHOLOGY AND STRUCTURE OF GLANDULAR HAIRS IN DEVELOPMENT OF THYMUS VULGARIS L.
IRINA BOZ*, LUIS NAVARRO**, RAMONA GALEŞ*, CLAUDIA PĂDURARIU*
Abstract: At Lamiaceae species the morphology, distribution and frequency of secretory hairs are used as taxonomic characters. At the most species from this family they are two types of secretory hairs: peltate and capitate, this types of hair can easily be distinguished after the number of cells of the secretory gland. For the investigation of secretory structures of Thymus vulgaris L. seedlings of different ages have been obtained. The secretory hairs from analyzed species present a basal cell, un unicellular stalk and based on numbers of cells ho form the gland, the hair can be: hairs with an unicellular gland presents on all vegetative organs, hairs with an bicellular gland presents only at the level of leafs primordium and hairs with an pluricelullar glands presents on all vegetative organs. Key words: Thymus, glandular hair, structure
Introduction
Plants produce a great number of secondary compounds utilized in the
pharmaceutical, chemical and food industries. These compounds have also many biological activities, probably as a result of the co-evolution of plants and different pathogens, herbivores, pollinators and other organisms [12]. The Lamiaceae plant family includes numerous aromatic and medicinal plants having great economical importance. Many species belonging to this family secrete volatile oils synthesized by the glandular hairs located on the surface of the aerial vegetative organs and even on the reproductive organs. Bearing in mind the economical and medicinal importance of the volatile oil of the species in the Lamiaceae family we have noticed the scarce amount of papers approaching the ultrastructural aspects of the glandular hairs [1,2, 6, 16, 17, 13, 3]. In the studies on the Lamiaceae family members carried out until the present time two types of glandular hairs are described (peltate and capitate) on the surface of the aerial vegetative organs, especially on the leaf surface. A typical glandular hair (peltate or capitate) has a basal region (formed of one or more cells), a stalk (unicellular or multicellular) and a head (uni or multicellular) [9]. Beside these structures, some authors consider the epidermis cells radialy surrounding the base of the glandular hair forming an integral part of the glandular hair. This kind of cells has been observed in Origanum [8, 9] and in species of the genus Thymus [15]. It has been noticed that these basal cells do not function as typical epidermis cells but they constitute an accessory of the glandular hair, having a role in the volatile oil secretion. The size, shape, distribution, vacuolization, the pericline walls plasmodesm density, of these cells probably contributes to collect the photosynthesis products in the mesophyll and to their transport to the basal cells of the glandular hairs. Subsequently these products will reach the head of the glandular hair through the stalk cells, where they play a role in the elaboration of the volatile oil under the action of the “enzymatic machine” of the secretory cells [9]. The capitate glandular hairs are much smaller than the peltate hairs and they possess a great morphological variability. Werker et al. [17] distinguished three major types * „Al. I. Cuza” University, Faculty of Biology, Iasi, Romania ** University of Vigo, Vigo, Spain
81

of capitate hairs in the Lamiaceae family: type I (short) having a basal cell, a uni- or bicellular stalk and a one or two secretory cell head; type II (medium) having a unicellular base, a unicellular or multicellular stalk and a unicellular gland; type III (long) having a basal cell and a stalk consisting of two to five cells. In the species taken into study the most encountered type is the first one (Thymus, Origanum) or it can be accompanied by the second type (Melissa). Compared to the peltate glandular hairs, considered to be involved exclusively in the production of volatile oil, the capitate glandular hairs in the Lamiaceae species secrete a so called “primary” volatile oil [1, 5], or mucilage [10, 14].
Matherial and methods
Structural and ultrastructural description of glandular hairs from Thymus vulgaris L. was made using electron microscopy – in the Electronic Microscopy Laboratory, Faculty of Vigo, University of Vigo, Spain): Surface electron microscopy research: a) Fixation: The material cut into pieces is fixated in a glutaraldehyde solution 2,5% in cacodilat buffer 0,1 M.; b). Dehydration: The material is dehydrated, successively dipping it in ethylic alcohol (35%, 50%, 70%, 90%, 95%); c) Substitution: Replacing the ethylic alcohol with amiloacetate; d). Desiccating: Replacing liquids with CO2, using a critical pint chamber; e). Montage of the tissue fragments: The processed samples are mounted on top of the carbonate filters on double adhesive props; f) Sample plating: The samples are coated in a thin layer of gold in vacuum atmosphere; g). Sample analysis: The processed vegetal material is analysed and photographed using a surface electron microscope. Transmission electronic microscopy analysis: a) Fixation: The cut material is fixed in a glutaraldehyde solution 2,5 % in cacodilat buffer 0,1 M.; b). Dehydration: The material is dehydrated, successively dipping it in ethylic alcohol (35%, 50%, 70%, 90%, 95%); c). Inclusion of the vegetal material in resins: the samples are successively dipped in Epon resins and acetone of different concentrations and placing them afterwards in small silicone trays at 60°C, for 48 hours.; d). Pyramid molding of the resin blocks: The blocks are molded in the pyramidothome, clearing off the resin excess that might interfere in getting them sectioned; e). Sectioning the blocks: The blocks are sectioned using a ultramicrothome to obtain semifine sections (0,70µm- sectioning using a glass knife) and ultrafine sections (70 nm- sectioning using a diamond blade knife). The semifine sections are mounted on blades, colored using methilene blue, analysed using the photonic microscope and photographed; f). Montage of the samples on grids: The ultrafine sections are mounted on grids; g). Contrasting the sections: The samples are contrasted using uranium acetate and lead citrate; h). Analysing the sections using transmission electron microscopy:
Results and discussions
The leaves of Th. vulgaris L. seedling are covered with a thick cuticle. A uni-layered epidermis is present on the abaxial and adaxial surface. In a front side view the epidermis consists in irregular-shaped cells, with waved lateral walls. The stomata of diacytic type are present in both epidermises, so the limb is of amfistomatic type. The leaves have non-glandular and glandular trichomes on both sides of the lamina. Non-glandular trichomes are
82

present on the veins and leaf margins and they are unbranched. The glandular hairs from Thymus vulgaris L. seedlings analyzed present a unicellular base, un unicellular stalk and a gland form by 1, 2, 4, 8 or 12 cells. Based on numbers of cells that form the gland, the glandular hairs observed by us can be: i. glandular hairs with unicellular gland; ii. glandular hairs with bicellular gland and iii. glandular hairs with un pluricellular gland.
The glandular hairs with a unicellular gland are present on all vegetative organs, with maxim density on the level of foliar primordia. On this level the secretory hairs presents, very rare, un stalk bi- or tricellular; the glandular cell produce volatile oil that spread through celluloses thin wall and swell out the cuticle that cover the gland.
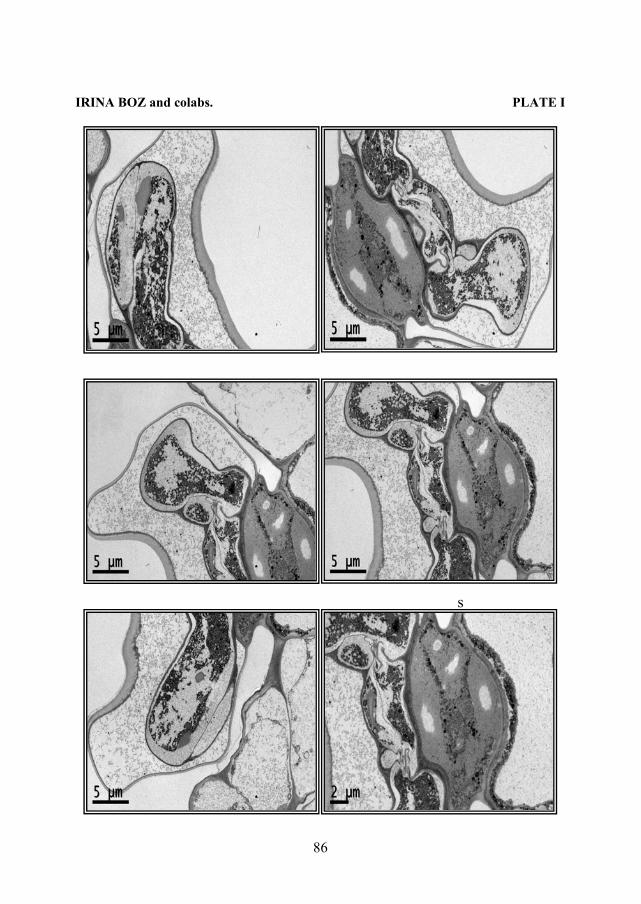
The glandular hairs with a bicellular gland can be observed only on the level of foliar primordia; then cuticle that cover the gland is detach from de cellular wall. The glandular hairs with a pluricellular gland are present on all vegetative organs. On the level of foliar primordia and on the epidermis that cover the top of the stem can be observed different stages from the development of glandular trichomes. The glandular trichomes with an octocellular gland, being in different stage of development, are distinguish by the ones with uni- or bicellular gland through the shape of base cell and of stalk, as well as the glandular cells did not have a distinguish cuticle. When the glandular hair in completely form, the cellular glands start to produce essential oil. This oil spread through external wall and swell out the cuticle that cover the gland. This glandular hairs are very abundant on young leaves, but they remain relative frequents on the mature leaves. Also, these glandular hairs are frequent on the top of the stem and the number of this hair is regressing to base. The secretory hairs, in different stage of development, can be identifying on all vegetative organs. Also, these secretory hairs are present on the level of cotyledons, epicotyls and hypocotyls (PLATE I, II).
Conclusions
Based on the number of cells that form the gland, the secretory hairs observed from
us are: secretory hairs with unicellular gland present on all vegetative organs, with maxim density on the level of foliar primordial, glandular hairs with a bicellular gland observed only on the level of foliar primordia; then cuticle that cover the gland is detach from de cellular wall and glandular hairs with a pluricellular gland present on all vegetative organs.
REFERENCES
1. AMELUNXEN F., 1964 - Elektronenmikroskopische Untersuchungen an den Drusenhaaren von Mentha piperita L, Planta Med., 12: 121–139;
2. AMELUNXEN F., 1965 - Elektronenmikroskopische Untersuchungen an den Drüsenschuppen von Mentha piperita L., Plant Med., 13: 457-473;
3. ANTUNES T., SERVINATE-PINTO I., 1991- Glandular trichomes of Teucrium scodonia L. morphology and histochemistry, Flora, 185: 65-70;
4. ASCENSÃO L., MARQUES N., PAIS M.S., 1995 - Glandular trichomes on vegetative and reproductive organs of Leonotis leonurus (Lamiaceae), Ann. Bot., 75: 619-626;
5. BISIO A., CORALLO A., GASTALDO P., ROMUSSI G., CIARALLO G., FONTANA N., DE TOMMASI N., PROFUMO P., 1999 - Glandular hairs and secreted material in Salvia blepharophyll Brandegee ex Epling grown in Italy, Ann Bot,, 83: 441–452;
6. BOSABALIDIS A.M., TSEKOS I., 1982 - Glandular scale development and essential oil secretion in Origanum dictamnus L., Planta, 156: 496–504;
83

7. BOSABALIDIS A.M., TSEKOS I., 1984 - Glandular hair formation in Origanum species, Annals of Botany, 53: 559-563;
8. BOSABALIDIS A.M., KOKKINI S., 1997 - Intraspecific variation of leaf anatomy in Origanum vulgare grown wild in Greece, Bot. J. Linn. Soc. (London), 123: 353 – 362;
9. BOSABALIDIS A. M, 2002 - Structural features of Origanum sp., In Oregano: the genra Origanum and Lippia, Taylor and Francis
10. BOURETT T., HOWARD R.J., O´KEEFE D. P., HALLAHAN D.L., 1994 - Gland development on leaf surfaces of Nepeta racemosa, Int. J. Plant. Sci., 155: 623-232;
11. CORSI, G., BOTTEGA S., 1999 - Glandular hairs of S. officinalis: New data on morphology localization and histochemistry in relation to function, Ann. Bot., 84: 657-664;
12. DUKE S.O., 1994 - Commentary: Glandular trichomes – a focal point of chemical and structural interactions, International Journal of Plant Sciences, 155:: 617-620;
13. DUDAI N., WERKER E., PUTIEVSKY E., RAVID U., PALEVITICH D., HALEVY A.H., 1988 - Glandular hairs and essential oils in the leaves and flowers of Majorana syriaca, Israel Journal of Botany, 37: 11–18;
14. MODENESI P., SERRATO-VALENTI G., BRUNI A., 1984 - Development and secretion of clubbed trichomes in Thymus vulgaris L., Flora, 175: 211–219;
15. STAHL-BISKUP E., 2002 - Essential oil chemistry of the genus Thymus - a global view In The genus Thymus, Taylor and Francis
16. WERKER E., RAVID U., PUTIEVSKY E., 1985a - Structure of glandular hairs and identification of the main components of their secreted material in some species of the Labiatae, Isr. J. Bot., 34: 31-45;
17. WERKER, E., RAVID, U., PUTIEVSKY, E., 1985b - Glandular hairs and their secretion in the vegetative and reproductive organs of Salvia sclarea and Salvia dominica, Israel Journal of Botany, 34: 239–252.
Explanation of plates:
Plate I - Thymus vulgaris L.: 1. Hypocotyl – general view; 2. Secretory hair with pluricellular gland from hipocotyl; 3. Secretory hair with bicellular gland from epicotyl; 4. Secretory hair with pluricellular gland from cotyledon; 5. Secretory hair with unicellular gland from cotyledon; 6. Leaf– general view. Plate II Thymus vulgaris L. - Ultrafine sections through a secretory hair with octocellular gland. The following structures may be observed: cuticle, subcuticular space, secretory cell, lipid inclusions, stalk cell, basal cell.
84

IRINA BOZ and colabs. PLATE I
65
43
21
85
IRINA BOZ and colabs. PLATE I
s
86





























