Embed Size (px)
Citation preview

J. Embryol. exp. Morpli. Vol. 21, I, pp. 1-21, February 1969
Printed in Great Britain
Morphogenetic analysisof the effects of juvenile hormone analogues and
other morphogenetically active substances onembryos of Schistocerca gregaria (Forskal)
By VLADIMIR J. A. NOVAK1
From the Department of Physiology, Institute of Entomology,Czechoslovak Academy of Sciences, Prague
One of the basic assumptions of the author's gradient-factor theory of insectmorphogenesis is that the effects of the hypothetical gradient-factor on tissuegrowth can be reproduced by the juvenile hormone, not only during post-embryonic development, but also in the course of embryogenesis (Novak,1951 a, b, 1956, 1966). This concept was originally based on the more or lessindirect evidence supplied by the findings of Pflugfelder (1947) in Dixippusmorosus and by those of Novak (1951 b) in Oncopeltus fasciatus.
Recently, however, direct evidence has been made available by the work ofSlama & Williams (1966) on Pyrrhocoris apterus and of Riddiford (1966) onHyalophora cecropia. Using the 'paper factor' in the first case and cecropia oilin the second, the above authors succeeded in demonstrating that these sub-stances were able to block embryogenesis when applied to the egg surface orto the female before egg-laying commenced. Most of the pathological morpho-genetic effects associated with inability to hatch were obtained with only minuteamounts of the given substances. The results indicate that these substancesought to prove very effective as ovicides in actual insect pest control. No attempthas been made to explain the mechanism of their action on embryos, however,or to compare it with their effect on larvae.
Locusts provide suitable material for the study of insect embryogenesis andof factors affecting it, both because of their large size and of their high resistanceto many types of experimental treatment. By exposing embryos of Schistocercagregaria to the action of the corpus allatum hormone and by applying farnesolmethyl ether and other JH analogues to the egg surface at different intervalsduring morphogenesis, the author succeeded in obtaining a wide variety ofembryonic morphogenetic aberrations of varying kinds and degrees (Novak,1969a).
1 Author's address: Odd. Fysiologie, Entomologicky Ustav CSAV, Na Folimance 5,Praha 2, Czechoslovakia.
I J E E M 2 I

V. J. A. NOVAK
The aim of the present paper is to classify these morphogenetic effects. It alsoattempts to explain the findings and the mechanism of the action of the sub-stances employed in terms of the author's gradient-factor theory.
MATERIAL AND METHODS
Eggs of Schistocerca gregaria laid in glass tubes 12 cm long and 4 cm indiameter, filled with moist sterilized sand, were incubated at 32 °C; at thistemperature they hatch in 12 days. Experimental eggs were washed in watercontaining 1 % nipagin to prevent the growth of moulds and small amounts ofpenicillin and streptomycin to prevent bacterial infections; they were then placedin Petri dishes on several sheets of filter paper moistened with the samesolution.
The chemicals to be tested were applied to the egg surface as drops by meansof a thin capillary tube. As most of them were lipophilic in character, the eggsurface had to be roughly dried just before applying them. Solid substanceswere either dissolved in an appropriate solvent, using the solvent only as thecontrol, or a small particle was applied to the egg surface with the tip of a needle,followed by a drop of the solvent from the capillary tube.
The following substances were tested:Farnesol trans-trans (a British product received from Dr D. B. Carlisle).Farnesol methyl ether (synthesized from the above by Dr D. B. Carlisle in
the laboratories of the Anti-Locust Research Centre, London).Dihydrochloromethyl farnesoate (synthesized by Dr J. Romariuk of the Institute
of Organic Chemistry, Czechoslovak Academy of Sciences, Prague).Queen honey-bee extract (ethanol extract of 200 fertilized queens aged 3-5
years, evaporated in vacuo and dissolved in 96% ethanol).9-oxotrans-2-decenoic acid (synthesized by Dr K. Streibl, Lipids Department,
Institute of Organic Chemistry, Czechoslovak Academy of Sciences, Prague).9-hydroxy-2-decenoic acid (same source).lO-hydroxy-2-decenoic acid (same source).Cecropia oil (extract of male abdomina of Hyalophora cecropia, received from
Professor H. A. Schneiderman, Developmental Biology Center, Western ReserveUniversity, Cleveland, Ohio, and from Dr G. B. Staal, Institute of Entomology,Agricultural University, Wageningen, Netherlands).
Paper factor (methanol extract of pages of a journal of American origin(Scientific American); this was diluted with water and extracted with petroleumether, which was afterwards evaporated off (Slama & Williams, 1965).
Control substances: pure 96% ethanol and olive oil.To study the effects of morphogenetically active substances the arrested eggs
were dechorionated by immersing them in a 2% solution of sodium hypo-chlorite, usually 1 or 2 days after the control eggs hatched. When thoroughlyrinsed in water, normal eggs continued to develop and hatched without detectable

Juvenile hormone effects 3
injury after this treatment. Only specimens used for histological sections weredissected from the egg membranes.
Eggs used for paraffin sections were fixed in Bouin's fluid, dehydrated throughthe usual series of alcohols, embedded in paraffin after Peterfi's methylbenzoatetreatment and stained after rehydration by Ehrlich's haematoxylin-eosin method.
Selected specimens, after dissection from the egg membranes and fixation,were preserved in 70% ethanol containing 3% glycerin.
RESULTS
Specimens treated with JH analogues may stop developing at any stage ofembryogenesis, from cleavage of the original egg cell (zygote) to the fullydeveloped embryo. They can easily be distinguished from normal specimens ofthe corresponding stage by the following features:
(i) They are still alive when the controls of the same age have hatched andoften for a long time afterwards.
(ii) They are usually more heavily pigmented and their compound eyesundergo partial differentiation and become pigmented at a much earlier stagethan in normal specimens.
(iii) Less of the yolk is consumed and the yolk often has a specifically granu-lated structure.
(iv) When they have reached a more advanced stage of development, theembryonic moulting process takes place at an earlier stage than in normalspecimens, so that we often find experimental embryos invested with a trans-parent exuvia which never, of course, hatch.
When estimating the morphogenetic effect of a test substance in a given case,the following features were taken into account: (a) the occurrence and timingof the embryonic moulting process; (b) the degree of pigmentation in normaland abnormal areas; (c) the amount of unconsumed yolk and the size of theembryo; (d) unsuccessful blastokinesis; (e) absence of the white cuticle; (/) trans-parency of various areas of the body; (g) the presence of white urate depositsand their position and amount; (h) the time at which changes occurred in theexperimental specimens (i.e. after application of a test substance), comparedwith the time of changes in the controls.
CLASSIFICATION OF MORPHOLOGICAL EFFECTS
The morphological changes produced by the application of active substancesto over 1500 eggs (not counting the controls) form a more or less complete seriesof transitions ranging from the arrest of development in the initial stage ofembryogenesis (during cleavage) to the complete development of apparentlynormal embryos. It was nevertheless thought that it would be useful, in orderto facilitate the description and analysis of these effects, to arrange them in ascale of representative types. Changes of a purely teratological character (e.g.

4 V. J. A. NOVAK
asymmetrical development or the obliteration of various organs) were consideredseparately. The following classification was therefore used, although, of course,it must be borne in mind that the individual grades are only arbitrary steps in awhole series of effects.
A. Symmetrical changesI. Fully developed embryos without detectable morphological aberrations,
but failing to hatch (Plate 1F).
PLATE 1
Eggs of Schistocerca gregaria treated with farnesol methyl ether and dechorionated withsodium hypochlorite (2%). A, Normal egg, outset of embryogenesis; B, type XI. (beforeblastokinesis); C, abnormal blastokinesis (note granulated yolk); D, type VI (mid-blasto-kinesis); E, type IV (miniature embryo); F, type I, fully developed embryo.

Juvenile hormone effects 5
II. Embryos with abbreviated appendages and usually with increased pig-mentation but no qualitative morphological abnormalities, with completedorsal closure and with no yolk (Plate 3H).
III. More or less fully differentiated embryos, but distinctly shorter than thelength of the egg. Communication of the digestive tube with the rest of the yolksac through a dorsal opening in the occipital part of the head. Abbreviatedappendages, increased red-brown pigmentation (Plates 2E, 3F).
PLATE 2
Embryos of Schistocerca gregaria dissected from eggs treated with farnesol methyl ether.A, Type X, cyclopoid embryo with pigmented compound eyes; B, type VII, outset of blasto-kinesis; C, type VI; D, type V; E, type V, dorsal view (note pigmentation of margins of yolk-sac opening); F, type IIT, miniature embryo with remains of yolk sac.

V. J. A. NOVAK
PLATE 3
Embryos of Schistocerca gregaria juvenilized with farnesol m. e., dissected from egg. A, TypeX, lateral view (note residue of serosal sac; B, type IX; C, type VII; D, type VI; E, type V;F, type IV (note marked sclerotization); G, advanced stage of type III (note scleroticpleuropods); H, type II.

Juvenile hormone effects 7
TV. Dwarf embryos with incomplete differentiation of segments of antennaeand maxillary palps, tarsi, etc., heavily pigmented. Yolk sac occupying morethan one-third of length of egg and communicating with body via head andpart of thorax (Plates 1 E, 2F).
V. Incompletely differentiated embryos with reduced pigmentation comparedwith IV, but greater than in the untreated controls. Incomplete blastokinesis(end of abdomen bent dorsally) (Plate 3E).
VI. Embryos with more severe morphological defects, often with a reducednumber of abdominal segments, in mid-blastokinesis. Only the compound eyespartly pigmented (Plates ID, 3D).
VII. Polypod embryos, incipient blastokinesis, head in direction of hydropyle,towards ventral side of egg. Two-thirds or more of yolk still remaining (Plates2C, 3C).
VIII. Protopod embryos with undeveloped abdominal appendages, beforeblastokinesis. Body in dorsal position, head directed towards hydropyle, onlythe compound eyes pigmented (Plate 1B).
IX. Embryos immersed in yolk, only slight differentiation of thorax, abdomenin form of thin plate. Length of embryo less than one-fifth of that of the egg(Plate 3 B).
X. Cyclopoid embryos with undifferentiated head and body and no appen-dages, not more than one-tenth of length of egg. Often connected with yolk bymeans of thin tube (formed of serosa?). Compound eyes often distinguishableand slightly pigmented (Plates 2 A, 3 A).
XI. Blastoderm stage, with more or less distinct germinal band. Roundhydropyle, blastoderm usually detached from white cuticle (Plate IB;Text-fig. 6).
XII. Pre-blastoderm stage without secondary egg membranes (yellow andwhite cuticle). Yolk granulated and usually partly consumed by hypertrophiedcleavage cells (Cf. Plate 1 B, C).
B. Asymmetrical changes
I. Reduction of segments in appendages only.II. Reduction of parts of body laterally.
III. Reduction of parts of body along anterio-posterior axis.The types described above are merely arbitrary steps in the continuous series
of morphological changes caused by the action of JH analogues. In addition,morphologically identical specimens may display a number of profound physio-logical differences, to be enumerated below. For example, types IV or V mayor may not be in an advanced stage of the moulting process (detached or notdetached from the transparent embryonic cuticle), or may be more or lesspigmented than the controls. Similarly, types VI-IX may differ as regards thestate of blastokinesis, the degree of transparency of the body or the amount ofurates deposited, types X-XII may or may not have a granulated yolk as a

8 V. J. A. NOVAK
result of survival or non-survival of cleavage cells, and may also have uratedeposits, while type X may or may not have distinctly pigmented compoundeyes. Greater or lesser asymmetry may likewise occur in most types. It willtherefore be useful to give a detailed description of several specific individualsand of the sequence of changes in various parts of the body.
DESCRIPTION
Closing of dorsal body walls and communication with yolksac. Treated specimensof types III and V may differ as regards the size of the dorsal orifice, whichcommunicates with the yolk. In some specimens it is limited to the anteriorpart of the occiput, while in others it may extend further back, to the thorax
B
Text-fig. 1. Juvenilized embryos with remains of yolk sac communication withdigestive tube through dorsal opening. A, Opening limited to anterior part of occiput(type III); B, opening in head and pronotum (type IV). Margins of openingmarkedly sclerotic. Farnesol methyl ether.
(Text-fig. 1 A, B). Although the size of the orifice varies considerably, it is easyto see that it is relatively larger, on average, in less differentiated embryos andthat the anterior margin is always on the occiput, while the position of theposterior margin changes with the size of the opening. This underlines the factthat in normal development the dorsal walls of the body close from the posteriorto the anterior end of the body.
Growth of appendages. When comparing the state of development of theantennae, palps, mandibles and legs in individuals, a number of simple rules
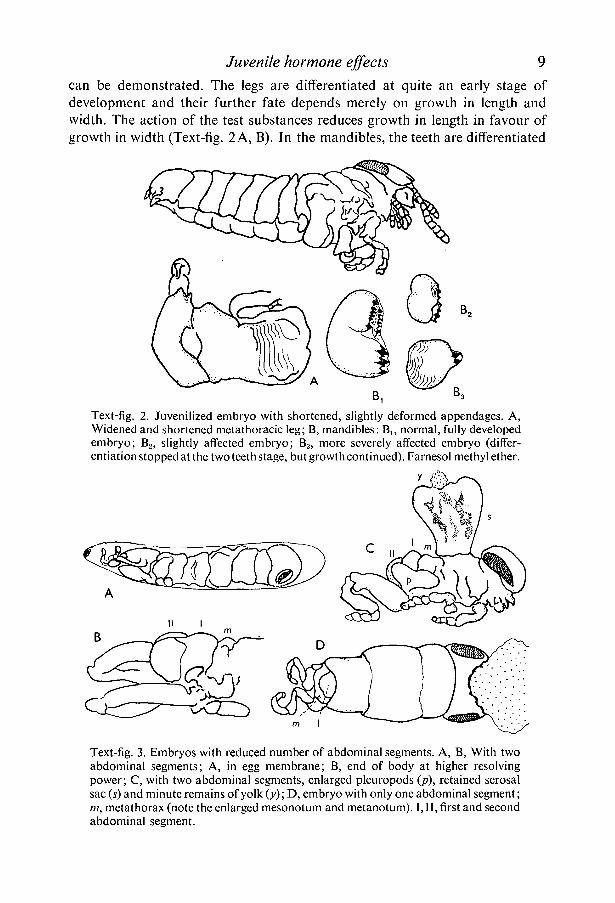
Juvenile hormone effects 9
can be demonstrated. The legs are differentiated at quite an early stage ofdevelopment and their further fate depends merely on growth in length andwidth. The action of the test substances reduces growth in length in favour ofgrowth in width (Text-fig. 2 A, B). In the mandibles, the teeth are differentiated
Text-fig. 2. Juvenilized embryo with shortened, slightly deformed appendages. A,Widened and shortened metathoracic leg; B, mandibles: B]5 normal, fully developedembryo; B2, slightly affected embryo; B3, more severely affected embryo (differ-entiation stopped at the two teeth stage, but growth continued). Farnesol methyl ether.
Text-fig. 3. Embryos with reduced number of abdominal segments. A, B, With twoabdominal segments; A, in egg membrane; B, end of body at higher resolvingpower; C, with two abdominal segments, enlarged pleuropods (p), retained serosalsac (s) and minute remains of yolk (y); D, embryo with only one abdominal segment;in, metathorax (note the enlarged mesonotum and metanotum). I, II, first and secondabdominal segment.

10 V. J. A. NOVAK
successively, one after the other, together with growth of the appendage inlength; short or long mandibles, with two or more teeth, can thus be obtained,according to the time at which the substance takes effect.
Differentiation of body segments. According to the strobilation theory (cf.Beklemishev, 1958), the segments of the body are differentiated progressively,from the anterior towards the posterior end of the body. This was also clearlydemonstrated in a number of cases in which, after application of the test sub-stance at a more advanced stage of embryogenesis (4th-5th day after egg-laying,
Text-fig. 4. Asymmetrical, severely deformed embryo with reduced thorax developingdirectly in yolk. Head developing separately from the rest of body without appen-dages. Crude queen-bee extract.
Text-fig. 5. Severely affected embryos in egg shell. A, Type VI; B, type X, but withtip of abdomen bent dorsally as at outset of blastokinesis.
room temperature), embryos with a reduced number of abdominal segmentswere obtained (Text-fig. 3 A-D), while the thoracic appendages were sometimeswell developed. A curious structure found in one of these specimens (Text-fig.3C) was undoubtedly the residue of the serosal walls of the original yolk sac.
Asymmetrical specimens were obtained mostly with large doses of stronglyactive substances acting in the later stages of development. A very peculiar caseof this type is the specimen in Text-fig. 4, in which the head and the well-differ-entiated abdomen were found to be separately immersed in the yolk, and thethorax was not to be found at all. The head had the form of a heart-shaped plate,with no appendages, but with relatively well-differentiated and pigmented
Juvenile hormone effects 11
compound eyes. The abdomen carried a pair of pleuropods on its roundedanterior end and laterally a branched segmented appendage was present on theleft side only, between the first and second abdominal segments.
Various stages of blastokinesis were preserved in specimens of types V-VII.Sometimes, however, well-differentiated type IV or even type II embryos werefound, with the head and thorax in the normal position, but with the abdomenin the same position as before blastokinesis (Plate 3D; Text-fig. 5A). In these,blastokinesis was evidently inhibited in about mid-course, while growth of the
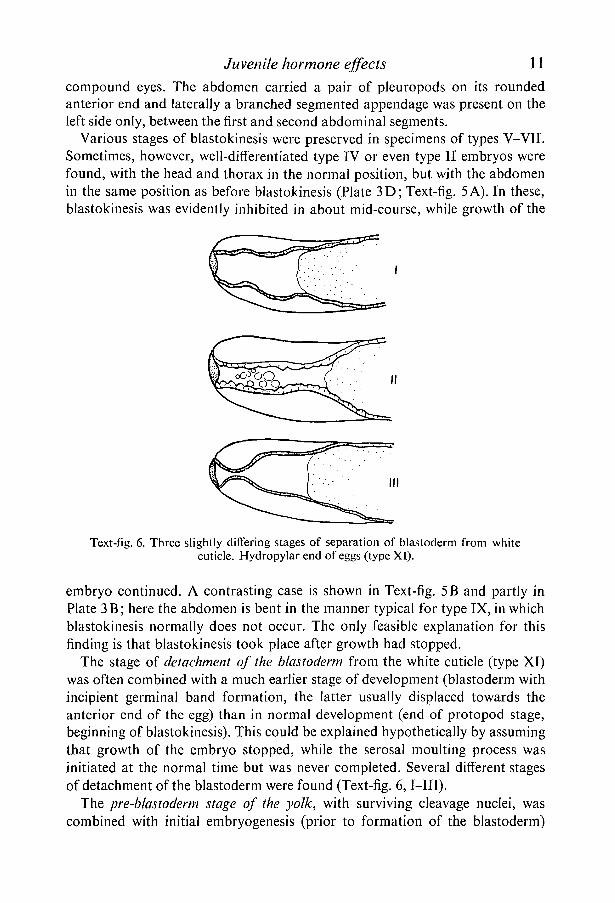
Text-fig. 6. Three slightly differing stages of separation of blastoderm from whitecuticle. Hydropylar end of eggs (type XI).
embryo continued. A contrasting case is shown in Text-fig. 5B and partly inPlate 3B; here the abdomen is bent in the manner typical for type IX, in whichblastokinesis normally does not occur. The only feasible explanation for thisfinding is that blastokinesis took place after growth had stopped.
The stage of detachment of the blastoderm from the white cuticle (type XI)was often combined with a much earlier stage of development (blastoderm withincipient germinal band formation, the latter usually displaced towards theanterior end of the egg) than in normal development (end of protopod stage,beginning of blastokinesis). This could be explained hypothetical ly by assumingthat growth of the embryo stopped, while the serosal moulting process wasinitiated at the normal time but was never completed. Several difTerent stagesof detachment of the blastoderm were found (Text-fig. 6,1—III).
The pre-blastoderm stage of the yolk, with surviving cleavage nuclei, wascombined with initial embryogenesis (prior to formation of the blastoderm)

12 V. J. A. NOVAK
(type XII). This is why the white cuticle was not formed in such eggs. In somecases, however, this state of the yolk was also found in post-blastoderm stages(types XI and X), together with a small, but more or less differentiated, embryowith a well-formed white cuticle. The presence of the white cuticle is amplyevident after dechorionation with NaClO. It forms a transparent but tough
B
Text-fig. 7. Giant cleavage cells. A, Cells of various sizes with intervening free space(consumed yolk). B, Enlarged cleavage cell at high resolving power; note the nucleus,reduced cytoplasm and three types of inclusions (type XII).
Text-fig. 8. Incomplete transverse bar with circular opening.Note the highly sclerotic margins.
membrane by which the egg keeps its original form. In its absence, the yolk,invested only with the thin vitelline membrane, collapses after the chorion hasbeen dissolved. This stage can be recognized by the coarsely granular appearanceof the yolk. Microscopic examination shows that the reason for this appearanceof the yolk is that it is composed of separate rounded cleavage cells of increasednumber and size interspersed with the rest of the yolk mass, while the remainingspace is filled with a transparent fluid. The latter is no doubt the outcome ofconsumption of part of the yolk by the cells. The largest cells are 80-100 /*• indiameter. They probably develop by cryptomitosis. Their cytoplasm contains

Juvenile hormone effects 13
three types of inclusions—large white spheres, small yellow spheres and irregulartransparent granules of gelatinous appearance and intermediate size (Text-fig. 7).
In a number of eggs, specific sclerotic, mainly heavily pigmented bars appeared,dividing the inside of the egg into two separate compartments. The bars wereoften incomplete, leaving a round central opening which sometimes had a moresclerotic margin (Text-fig. 8). In some eggs, two or more of these bars wereobserved. In several cases a small, reduced embryo was found in the largestcompartment. These bars also appeared in eggs attacked by moulds; this seemsto indicate that they developed as a result of cuticulogenic activity of the blasto-derm regenerating after annular surface injury by the mould. They appearedonly in embryos severely damaged either by a test substance or by a mould.
TESTS OF VARIOUS JUVENILE HORMONE ANALOGUES
To compare the morphogenetic activity of various substances known for theirjuvenile hormone-like effects, eggs of about the same age (1-2 days after laying)were treated with approximately the same amount of the substance (2-3 micro-drops, applied with a thin capillary). The results were evaluated 1-2 days afterhatching of the corresponding controls. The latter were eggs of the same pods,either untreated or treated with the solvent used in the experiment (acetone,liquid paraffin, ethanol). Only experiments in which the controls were com-pletely negative were taken into account. The effects of large doses of ethanoland liquid paraffin were studied separately.
The survey of the results, given in Table 1, shows that the most active testsubstances were farnesol methyl ether and other farnesol compounds, such asdihydrochloromethyl farnesoate (R 24). Crude queen-bee extract had a verypronounced and specific effect. The synthetic queen inhibitory substance(9-oxotrans-2-decenoic acid) seems to give less clear results, however, partlybecause of the small number of specimens and partly because of the necessity ofapplying the substance in ethanol, which appears to have some effect itself.This question will be studied separately (Novak, 1968). The effect of applyingcecropia oil, whatever its origin, was very slight. The paper factor gave com-pletely negative results. In further experiments farnesol methyl ether was usedalmost exclusively, as the most reliable agent (cf. Table 1, no negative results).
Influence of the amount of substance used
To obtain at least an approximate idea of the quantitative aspect of theeffects, eggs of the same age were treated after laying with varying amounts(different numbers of drops of roughly the same size) of farnesol methyl ether.The results are given in Table 2.
Even if the number of experiments in some of the series might be thought tobe not altogether adequate, the agreement between the increasing dose and the

Tab
le 1
. C
ompa
riso
n oj
the
eff
ects
oj
vari
ous
JH a
nalo
gues
on
the
embr
yoni
c de
velo
pmen
t o
/Sch
isto
cerc
a gr
egar
ia(2
-3 d
rops
oj
undi
lute
d su
bsta
nce
per
egg)
(In
the
case
of
solid
sub
stan
ces
a pa
rtic
le w
as p
lace
d on
the
egg
surf
ace
and
was
dis
solv
ed w
ith a
dro
p of
96
% a
lcoh
ol.)
Subs
tanc
e
Far
neso
l tr
ans-
tran
sF
arne
sol
met
hyl
ethe
rR
24
Cec
ropi
a oi
lP
aper
fac
tor
Que
en-b
ee e
xtra
ct9-
Oxo
tran
s-2-
dece
noic
aci
d9-
Hyd
roxy
dec-
2-en
oic
acid
10-H
ydro
xyde
c-2-
enoi
c ac
id
(Far
neso
l m
ethy
l et
her
n n 71 65 19 22 43 35 10 26
0 3—
6 4 15 5 2— —
1 — 3 5 3 31 10 — 13
II 3 3 3 4 — 4 4 4 7
III
— 4 4 — — —7
— —
IV
2 113 3—
3 2 1—
Typ
e of
effe
ct
V 4 12 6 3—
3— — —
VI 6 3
— — —2
— —
VIT — — 2 — — — — — —
Tab
le 2
. In
flue
nce
ojth
e am
ount
ojj
uven
iliz
ing
subs
tanc
e us
ea
appl
ied
undi
lute
d0-
1 da
yaf
ter
egg-
layi
ng.)
The
mea
ns w
ere
obta
ined
vi i
r ix
— 3 13
6 —
—
——
—
2—
1
—
—2
—
X 1 2 6 — —3
— — —
by m
ulti
plyi
ng t
he
XI
20 10 — —2 1 3
—•
num
ber
indi
vidu
als
of e
ach
type
by
the
corr
espo
ndin
g ty
pe n
umbe
r (1
-XII
) an
d di
vidi
ng th
e re
sult
by th
e to
tal n
umbe
r of
spe
cim
ens
eval
uate
d
Am
ount
n
1 dr
op
972
drop
s 13
3 dr
ops
384
drop
s 58
5 dr
ops
116
drop
s 53
0 I
20
10—
—
—
——
—
—
.——
—
II 9— — — — —
Hi
10 —5
— —3
IV 14 7 2 1 1 2
Typ
e of
effe
ct
V 4 2 2 8 — 2
VI 5
— — 11 — —
VII 6
— 10 15 2 8
VII
I
4 — 2 11 •— 9
IX
2 2 1 3—
2
X
XI
5
—
—6
—6
24 8
13
XII 8 2 4 1 4 6
XII 7 3 — 4 —3 2
— of
Mea
n
4 5 6 7 8 9
> OVAK

Juvenile hormone effects 15
average effect is very clear. This is confirmed by a number of other, less complete,experiments, which are not given here.
When considering these results the following points must be borne in mind.The effects of an active substance on the specimen can be modified by threefactors: (i) by its concentration, (ii) by the length of time for which an activeconcentration is maintained, (iii) by the interval between application of thesubstance to the egg surface and the attainment of an active concentration in agiven structure. With reference to the last point, differences in this respectbetween various parts of the embryo could be responsible for the production ofasymmetrical effects if the substance is applied near the crucial period fordetermination of a particular morphogenetic process.
Table 3. The influence of time of application
(Farnesol methyl ether, 2-3 drops per egg applied.)
nType of effect
01IIIIIIVVVIVIIVIIIIXXXIXII
Mean
24
51
—
—26635083
1088
36
14
—
—2
———
3———
548
Time of application (h)
48
15
—
—231
—22
—2327
62
.14
—
—13212
—22
—16
110
14
—
—22
—13
—11136
160
23
—
22332
———
5246
182
11
—
——
31
————
4126
230
14
—
—22
—2
———
4—
4(7)
Influence of the time of application
The time of application of the active substance, in relation to the stage ofmorphogenesis, is also of primary importance, as well as the amount. This wastested by subjecting eggs to the action of equal amounts of farnesol methylether (2-3 drops per egg) at different periods after laying. The eggs were keptat a constant temperature of 32 °C before and after treatment; under theseconditions the untreated controls hatched in about 12 days. The results aregiven in Table 3.
Here again, many more experiments, at shorter intervals, would be neededto obtain an exact idea of the time relations of this type of morphogenetic action.A detailed analysis of this question is given in Novak (1969 &). The present results,

16 V. J. A. NOVAK
however, like a number of others not given in the table, seem to show a clearinverse correlation between the activity of the substance and the time at whichit is administered after egg-laying.
CONCLUSIONS
The material studied allows us to conclude that embryonic morphogenesiscan be arrested at any stage by the application of substances with juvenilehormone-like activity. Development can afterwards continue, under differentconditions, when the substance ceases to act. Any structure characteristic ofembryonic development, starting with the cleavage cells, can thus be kept alivefor a period equal to the hatching time of the control embryos. In addition, awide range of new, abnormal structures can appear, either as a result ofheterochronia (in the sense of Novak, 1966) or of direct morphogenetic action.
When comparing juvenilizing effects in embryos with those in larvae, astriking contradiction can be seen. In post-embryonic development, the adminis-tration of juvenile hormone or an analogous substance at a time in developmentwhen it is normally not active (e.g. the beginning of metamorphosis) inhibitsfurther morphogenesis (metamorphosis) and results in the appearance of giantlarvae. Conversely, removal of the source of the hormone leads to precociousmetamorphosis, resulting in miniature adults. In embryogenesis, however, theaction of a juvenilizing substance quite frequently results in the formation offully differentiated dwarf embryos (types III-V).
The following considerations, in terms of the author's gradient-factor theory(Novak, 1966, 1967), nevertheless suggest that this contradictory effect is notdue principally to a difference in the mode of action of the hormone or analogoussubstances, but that it simply arises from the changed physiological and morpho-logical situation, i.e. the different intra-environment (the micro-environment ofSpratt, 1965; cf. Novak, 1967).
It has been shown that miniature embryos appear after the application ofsmall amounts of the substance to freshly laid eggs. Its effect thus seems toconsist, as in larvae, in causing the survival and further growth of that part ofthe embryonic blastoderm which would normally die and disintegrate duringgerminal band formation. There is an important difference between this situationand the situation in the larval period, however. The prolonged growth of larvaltissue necessarily results in the deflexion to larval structures of the part of thefood supply which would otherwise be available for the imaginal structures (cf.the law of correlation in consumption, Novak, 1966), thus limiting the growthof imaginal structures.
The situation in early embryos is quite different because of the presence ofan unlimited food supply in the form of the yolk mass. This alone wouldadequately explain why further differentiation of the germinal band is notstopped unless the effect of the JH analogue is strong or persistent. If the sub-

Juvenile hormone effects 17
stance takes effect after differentiation of the germinal band has commenced,but before it has spread to all the blastoderm cells involved in normal develop-ment, its juvenilizing action prevents the differentiation process from spreadingto further cells. When the action of the substance ceases, the embryo would thusdevelop from a smaller number of cells than normally; this could be the onlyreason for the limited size of juvenilized embryos under the above conditionsand for the associated incomplete consumption of the yolk.
E -b"5
jrm
al
rosa
l m
ou
ltin
z a
E ;- i
r Abn
oEo • ;
^ v S
*" 5r
J
" : ^
IV VI VIII IX XII 1 2 3 4 5 6 7 8 9 10
Text-fig. 9. Diagrammatic illustration of relation of juvenilized forms to stages ofnormal development. On left, different degrees of abnormal development; on right,stages of normal development; in centre, time (stage) span of some of processes. IVand XII, proportionate increase in body size caused by juvenilizing factor; IX,asymmetrical increase.
The same principle also undoubtedly applies at a later stage, in the case ofreduction in the number of body segments (e.g. reduction of the abdomen toonly two metameres). The metameres present at the moment the juvenilizingsubstance starts to take effect are preserved and can later differentiate appendages,but segmentation of the body does not continue. This accounts for the produc-tion of embryos with only one or two abdominal segments (Text-fig. 3). Thedifferentiated segments can continue to grow in their original shape (Text-fig. 9).Agreement with the juvenilizing effect in the post-embryonic period is clearlyevident here.
More experimental evidence will be needed before all the details observedcan be fully explained. It seems clear, however, that all aberrations producedby the action of the juvenile hormone and its analogues in embryos could becaused by the same mechanism as that which produces the changes observed
2 JEEM 21

18 V. J. A. NOVAK
in larval development and metamorphosis and that the differences betweenthem are due only to the specific intra-environmental conditions and the timingof action of the substance.
It can be assumed from the above considerations that the main types ofjuvenilizing action are as follows.1
(a) Full and persistent action of the juvenilizing factor. This results in thepreservation of structures existing at the moment the factor starts to take effect,which in normal development would disintegrate. The structures thus preservedcontinue to grow as long as the given factor is active and the necessary foodsupply (yolk) is available, with the result that they may attain a size several timeslarger than normal.
(b) Partial action of the juvenilizing factor, in which only some parts of thebody are more or less juvenilized, while differentiation of the others is notaffected. This is the origin of various forms of heterochronia (cf. Novak, 1956,1966) found both in the post-embryonic and the embryonic period. These partialeffects have two different causes: (i) the amount of the active substance is toosmall to produce a complete effect, (ii) the action of the factor is only temporary,as a result, for example, of its elimination by some of the tissues. It is impossibleto decide from the available evidence, however, which of these causes appliesin any given case.
(c) Local action of the juvenilizing factor. This may be due either to limiteddistribution of the factor, or perhaps more frequently to a low rate of spreadingof the factor inside the body, with the result that different morphogenetic pro-cesses are affected, even in symmetrical organs. It can therefore be assumed thatstriking changes originate only during the period immediately preceding a deter-mination process. Local action undoubtedly accounts for the various asym-metrical changes observed above.
SUMMARY
1. A number of juvenile hormone analogues and other morphogeneticallyactive substances (e.g. the queen-bee inhibitory substance) were shown toproduce juvenilizing effects in embryos of Schistocerca gregaria.
2. In about 1500 experimental specimens all transitions were found, from theinitial stages of embryogenesis, starting with preblastoderm cleavage cellsmaintained by the juvenilizing effect, to the normal, fully developed embryo.In addition, other deformities, some of them asymmetrical, were found.
3. To facilitate description of the results, a classification of juvenilizing effectswas suggested, comprising twelve types related to the extent of the morpho-logical changes produced.
1The term 'juvenilizing factor' covers any factor resulting in juvenilization in the senseemployed above, including the corpus allatum hormone itself and all its analogues, togetherwith external environmental factors such as temperature (cf. Wigglesworth, 1952) and thenot yet fully understood effect of parasitization by some protozoans (microsporidia) andhelminths (Mermis) (cf. Fischer & Sanborn, 1964).

Juvenile hormone effects 19
4. A number of substances were tested and compared with each other withreference to the degree of their juvenilizing effects. Farnesol methyl ether andother farnesol derivatives were found to be the most effective. Distinct specificactivity was also obtained with crude queen bee extract. More evidence isneeded to elucidate the question of the activity of 9-oxodec-2-enoic acid andsome of its derivatives. Cecropia oil and the paper factor (juvabion) of Slama& Williams had practically no effect in these experiments.
5. The juvenilizing effect was found to increase with the amount of substanceapplied and to decrease as the age of the tested eggs rose.
6. Several specific cases of juvenilized locust embryos are described in greaterdetail, including fully differentiated miniature embryos, embryos lacking someof the body segments and surviving giant cleavage cells from the preblastodermperiod.
7. The seeming contradiction between the effects of JH analogues on embryos(miniature, fully differentiated embryos) and their effects in the post-embryonicperiod (giant larvae with inhibited differentiation) can be explained on the basisof the author's gradient-factor theory as the results of exactly the same modeof action under different environmental conditions. The fact that embryonicdevelopment can be influenced by juvenilizing factors in the same way as post-embryonic morphogenesis is important evidence in support of the gradient-factortheory.
RESUME
Analyse morphogenetique des effets des analogues de Vhormone juvenile etd'autres substances morphogenetiquement actives chez les embryons de Schistocercagregaria (Forskdl)
1. Un certain nombre d'analogues de l'hormone juvenile ainsi que d'autressubstances morphogenetiquement actives (par exemple la substance inhibitricede reine d'abeille) produisent des effets juvenilisants chez les embryons deSchistocerca gregaria.
2. Chez 1500 specimens experimentaux, on trouve tous les etats intermedi-aries entre les stades initiaux de l'embryogenese — tel le maintien, du a l'effetjuvenilisant, du stade preblastoderme avec cellules de segmentation — etl'embryon normal completement developpe. En plus de ces effets, d'autresdeformations, dont certaines sont asymetriques, ont ete observees.
3. Pour faciliter la description des resultats, une classification des effetsjuvenilisants est suggeree: 12 stades sont proposes, en fonction du taux dechangements morphologiques produits. Le stade I represente les individus lesmoins atteints, le stade XII les individus les plus atteints.
4. Un certain nombre de substances ont ete eprouvees et comparees entreelles au point de vue de leur effet juvenilisant. L'ether methylique de farnesolet d'autres derives du farnesol sont les plus efflcaces. Une activite specifiquenette a egalement ete obtenue avec l'extrait brut de reine d'abeille. Des preuves

20 V. J. A. NOVAK
supplementaires sont necessaires pour mettre en evidence l'activite de l'acide9-oxodec-2-enoique et de certains de ses derives. L'huile de Cecropia et lefacteur extrait de la pate a papier par Slama & Williams (juvabion) sont quasi-ment inactifs dans ces experiences.
5. L'effet juvenilisant augmente en fonction de la quantite de substanceappliquee et diminue lorsque l'age des oeufs experimentaux augmente.
6. Plusieurs cas speciaux d'embryons d'acridien juvenilises sont decrits avecplus de detail. Parmi eux, les plus importants sont les embryons parfaitementdifferencies mais nains, les embryons depourvus de certains segments du corps,et les cellules geantes de segmentation, survivantes de la periode preblastoderme.
7. L'examen des resultats obtenus permet de formuler quelques conclusionsimportantes sur le mode d'action des facteurs juvenilisants. Les effets apparem-ment contradictoires produits sur les embryons par des analogues de l'hormonejuvenile (embryons nains completement differencies) et ceux qui sont decritscomme resultant de Faction des memes facteurs pendant la periode post-embryonnaire (larves geantes dont la differentiation est inhibee) peuvent etreexpliques en se basant sur la theorie du facteur gradient proposee par l'auteur:ces effets resulteraient d'une seule et meme action s'exercant dans des conditionsd'environnement differentes.
8. Le fait que le developpement embryonnaire peut etre influence par desfacteurs juvenilisants de la meme maniere que la morphogenese post-embryon-naire est une preuve importante en faveur de la theorie du facteur gradient.L'analyse detaillee des phenomenes observes constitue un argument de pluspour la theorie et ouvre un domaine nouveau d'investigation pour l'etude desproblemes de morphogenese.
Part of the work discussed in this paper was carried out during tenure of a grant from theOrganisation for Economic Co-operation and Development at the time of my stay in theAnti-Locust Research Centre, London, in the winter of 1965-6. I should like to express mygratitude to the Director, Dr P. T. Haskell, for providing facilities, for his generous helpduring my work at the Centre, and for editing and improving the typescript. Further thanksare due to Dr D. B. Carlisle, D.Sc, for his interest in my work and for providing technicalequipment and chemicals, and to all members of the staff; to Professor H. A. Schneiderman,Dr I. G. B. Staal, Dr K. Streibl and Dr K. Slama for providing some of the substancestested; and to Mrs Margaret Schierlova for corrections to the English of the text.
REFERENCES
BEKLEMISHEV, V. N. (1958). Grundlagen der vergleichenden Anatomie der Wirbellosen. Berlin:VEB Deutscher Verlag der Wiss. 471 pp.
FISHER, F. M. & SANBORN, R. C. (1964). Nosema, a source of juvenile hormone in parasitisedinsects. Biol. Bull. mar. biol. Lab., Woods Hole 126, 235-52.
NOVAK, V. J. A. (1951 a). New aspects of the metamorphosis in insects. Nature, Lond. 167,132.NOVAK, V. J. A. (19516). The metamorphosis hormones and morphogenesis in Oncopeltus
fasciatus Dal. Acta Soc. zool. csl. pp. 1^48.NOVAK, V. J. A. (1956). Versuch einer zusammenfassenden Darstellung der postembryonalen
Entwicklung der Insekten. Beitr. Ent. 6, 205-493.NOVAK, V. J. A. (1966). Insect Hormones, 3rd ed. London: Methuen. 478 pp.

Juvenile hormone effects 21NOVAK, V. J. A. (1967). The juvenile hormone and the problem of animal morphogenesis.
In Insects and Physiology. Oliver & Boyd, Edinburgh and London, pp. 119-132.NOVAK, V. J. A. (1969 a). The action of the juvenile hormone as a special type of derepression
activity. Proc. Int. Symp. Insect Endocrin., Brno 1966. (In the Press.)NOVAK, V. J. A. (19696). Morphogenetical analysis of the effects of the juvenile hormone
analogues on embryos of Schistocerca gregaria. II and III. (In preparation.)PFLUGFELDER, O. (1947). Die Entwicklung embryonaler Teile von Dixippus morosus in der
Kopfkapsel von Larven und Imagines. Biol. Zbl. 66, 372-87.RIDDIFORD, L. M. (1966). The action of the juvenile hormone on silkworm embryos. Proc.
Int. Symp. Insect Endocrin., Brno, 1966. (In the Press.)SLAMA, K. & WILLIAMS, C. M. (1965). Juvenile hormone activity for the bug Pyrrhocoris
apterus. Proc. natn. Acad. Sci. U.S.A. 54, 411-14.SLAMA, K. & WILLIAMS, C. M. (1966). 'Paper factor' as an inhibitor of the embryonic
development of the European bug Pyrrhocoris apterus. Nature, Lond. 210, 329-330.SPRATT, N. T. (1965). Introduction to Cell Differentiation. London: Chapman & Hall.WIGGLESWORTH, V. B. (1952). Endocrine regulation during development. I. Hormones in
relation to metamorphosis. Gen. Comp. Endocrin., suppl. 1, pp. 316-21.
{Manuscript received 20 November 1967, revised 2 August 1968)





























