Embed Size (px)
Citation preview

Md
ATB
a
ARR2AA
KSAISXS
1
tlKtdphfr1bt
DRrt
L
h0
Carbohydrate Polymers 139 (2016) 10–19
Contents lists available at ScienceDirect
Carbohydrate Polymers
j ourna l ho me pa g e: www.elsev ier .com/ locate /carbpol
olecular rearrangement of waxy and normal maize starch granulesuring in vitro digestion
nju Teng, Torsten Witt, Kai Wang, Ming Li, Jovin Hasjim ∗
he University of Queensland, Centre for Nutrition and Food Sciences, Queensland Alliance for Agriculture and Food Innovation,risbane 4072, QLD, Australia
r t i c l e i n f o
rticle history:eceived 23 July 2015eceived in revised form3 November 2015ccepted 24 November 2015vailable online 2 December 2015
a b s t r a c t
The objective of the present study is to understand the changes in starch structures during digestionand the structures contributing to slow digestion properties. The molecular, crystalline, and granularstructures of native waxy maize, normal maize, high-amylose maize, and normal potato starch granuleswere monitored using SEC, XRD, DSC, and SEM. The amylose and amylopectin molecules of all fourstarches were hydrolyzed to smaller dextrins, with some having linear molecular structure. Neither the A-nor B-type crystallinity was resistant to enzyme hydrolysis. Starch crystallites with melting temperatureabove 120 ◦C appeared in waxy and normal maize starches after digestion, suggesting that the linear
eywords:tarch granulesmylose
n vitro digestionize-exclusion chromatography-ray diffractometrycanning electron microscopy
dextrins retrograded into thermally stable crystalline structure. These crystallites were also observed forhigh-amylose maize starch before and after digestion, contributing to its low enzyme digestibility. Onthe contrary, the enzyme-resistant granular structure of native normal potato starch was responsible forits low susceptibility to enzyme hydrolysis.
© 2015 Elsevier Ltd. All rights reserved.
. Introduction
Starch is the major energy source in most staple foods. Its diges-ion rate and extent of digestion in the small intestine have aarge impact on human health and nutrition. According to Englyst,ingman, and Cummings (1992), starch can be categorized into
hree groups, which are rapidly digestible starch (RDS), slowlyigestible starch (SDS) and resistant starch (RS). It has been pro-osed that RDS is completely digested within 20 min, causingigh postprandial glycemic and insulinemic responses and there-
ore increasing the risks of metabolic disorders such as insulinesistance, diabetes, and obesity (Byrnes, Br& Miller, & Denyer,
995; Ludwig, 2002; Willett, Manson, & Liu, 2002). SDS is digestedetween 20 and 120 min after consumption and RS is the remnantshat are not digested after 120 min. Both SDS and RS can lowerAbbreviations: CLD, chain length distribution; DP, degree of polymerization;SC, differential scanning calorimetry/calorimeter; RDS, rapidly digestible starch;S, resistant starch; SDS, slowly digestible starch; SEC, size-exclusion chromatog-aphy; SEM, scanning electron microscopy; Tc , conclusion temperature; To, onsetemperature; Tp , peak temperature; XRD, X-ray diffractometry.∗ Corresponding author. Current address: Roquette Management (Shanghai) Co.,td., Shanghai 200031, China. Tel.: +86 21 2422 9723; fax: +86 21 6142 3072.
E-mail address: [email protected] (J. Hasjim).
ttp://dx.doi.org/10.1016/j.carbpol.2015.11.062144-8617/© 2015 Elsevier Ltd. All rights reserved.
postprandial glycemic and insulinemic responses with SDS pro-longing the supply of glucose to the body, especially to the brain,which is particularly beneficial for maintaining cognitive functions.Similar to other dietary fibers, RS escapes the digestion in the smallintestine and reaches the colon, where it serves as an importantsubstrate for gut fermentation producing short-chain fatty acidswith the potential to prevent the development of colon cancer cells(Ferguson, Tasman-Jones, Englyst, & Harris, 2000; Zhao et al., 2011).
The structures of native starch granules can be simplified intofive levels (Dona, Pages, Gilbert, & Kuchel, 2010; Tran et al., 2011).Linear branches of starch molecules (Level 1 structure) are madeof glucose monomers linked together by �-(1 → 4) glycosidic link-ages, and these linear branches are connected together by �-(1 → 6)glycosidic linkages forming individual fully branched molecules(Level 2 structure). These lowest two levels are molecular lev-els composed of mainly amylose and amylopectin. Amylose isprimarily linear with a few long branches, whereas amylopectinis highly branched with a vast number of short branches and amolecular size two orders of magnitude larger than amylose. Somebranches of amylopectin are arranged in a double helical conforma-
tion, packing into crystallites that form alternating crystalline andamorphous layers (semi-crystalline structure, Level 3). In contrast,amylose is typically amorphous or in a single helical complex withlipid molecules. The semi-crystalline structures are arranged into
te Pol
crs
blIG(a22(2dsmw2Sdlf(Glswtdibe
rtidussvbslstmcs(
2
2
nsLfSA(L
A. Teng et al. / Carbohydra
oncentric growth rings, which alternate with amorphous growthings around the amorphous hilum (Level 4 structure) in a singletarch granule (Level 5 structure).
The rate and extent of starch digestion are largely influencedy starch structures, including the branch-chain length of amy-
ose and amylopectin molecules (Srichuwong, Sunarti, Mishima,sono, & Hisamatsu, 2005; Syahariza, Sar, Hasjim, Tizzotti, &ilbert, 2013), molecular size (Li et al., 2015a), amylose content
Li, Jiang, Campbell, Blanco, & Jane, 2008; Syahariza et al., 2013),mylose–lipid complexes (Ai, Hasjim, & Jane, 2013; Hasjim et al.,010b), crystalline structure (Hasjim & Jane, 2009; Jane et al.,003; Sievert & Pomeranz, 1990), granule size and surface structureDhital, Shrestha, & Gidley, 2010; Hasjim, Srichuwong, Scott, & Jane,009). The changes in starch structure during in vitro and in vivoigestions have recently been investigated. The rearrangement oftarch molecules during in vitro digestion of extruded high-amyloseaize starches to form enzyme-resistant crystalline structuresas reported by several authors (Htoon et al., 2009; Li et al.,
015a; Lopez-Rubio, Flanagan, Shrestha, Gidley, & Gilbert, 2008b;hrestha et al., 2010). The dextrins produced after prolonged in vitroigestion of either extruded maize starches with various amy-
ose contents or native normal maize starch granules were bothound to be predominately linear with degree of polymerizationDP) X ∼ 50 (Hasjim, Cesbron Lavau, Gidley, & Gilbert, 2010a; Witt,idley, & Gilbert, 2010). Similarly, the digesta collected from the
ower-half small intestine of the pigs fed with native normal maizetarch granules contained substantial amounts of linear dextrinsith DP X ∼ 50 (Hasjim et al., 2010a). Combining the results from
hese studies, it can be assumed that the linear dextrins pro-uced during starch digestion are able to rearrange themselves
nto highly ordered crystalline structures that are less suscepti-le to enzyme hydrolysis. However, this relationship has not beenxplicitly proven in a single study before.
The aim of this study is to understand the molecularearrangement of starch during digestion and the structures con-ributing to slow digestion properties by investigating the changesn the molecular, crystalline, and granular structures at differentigestion times. Native starch granules (Level 5 structure) weresed as the substrates for an in vitro digestion. Although nativetarch granules are less common in human food than gelatinizedtarch, they are present in animal feed, fresh produce (fruits andegetables), and low-moisture foods (such as biscuits and somereakfast cereals), and the lack of retrograded starch in nativetarch granules allows the distinct observation of starch molecu-ar rearrangement (retrogradation) during digestion. The moleculartructures (Levels 1 and 2) of the starch digesta collected duringhe in vitro digestion were characterized using size-exclusion chro-
atography (SEC), while their crystalline structures (Level 3) wereharacterized using X-ray diffractometry (XRD) and differentialcanning calorimetry (DSC). The changes on the granular structureLevel 5) were identified using scanning electron microscopy (SEM).
. Materials and methods
.1. Materials
Four types of native starch granules were used: waxy maize,ormal maize, high-amylose maize (Gelose 50), and normal potatotarches. The maize starches were obtained from Penford Australiatd. (Lane Cove, NSW, Australia). Normal potato starch, pancreatinrom porcine pancreas, and LiBr (ReagentPlus) were obtained from
igma-Aldrich (Castle Hill, NSW, Australia). Amyloglucosidase fromspergillus niger, isoamylase from Pseudomonas sp., and d-glucoseGOPOD Format) kit were purchased from Megazyme Internationaltd. (Bray, Co. Wicklow, Ireland). Dimethyl sulfoxide (DMSO, GR forymers 139 (2016) 10–19 11
analysis ACS) was from Merck and Co., Inc. (Kilsyth, VIC, Australia).Other chemicals were reagent grade and used as received.
2.2. In vitro starch digestion
Native starch granules (1.5 g, dry weight basis) were suspendedin sodium acetate buffer (5.0 mL, 0.2 M, pH 6.0) containing 200 mMCaCl2, 0.49 mM MgCl2, and 0.02% NaN3. An enzyme solution(5.0 mL) containing 60 mg pancreatin and 300 �L amyloglucosidasein sodium acetate buffer solution was added to the starch suspen-sion after being equilibrated at 37 ◦C with stirring for 5 min. Themixture was incubated at 37 ◦C with stirring. The digestion wasstopped by adding 30 mL absolute ethanol to the mixture at 0, 2, 4,8, and 16 h for waxy maize starch; 0, 2, 4, 8, 16, and 24 h for nor-mal maize starch; and 0, 16, 24, and 48 h for high-amylose maizeand normal potato starches. The starch digesta (precipitate) wascollected by centrifugation at 4000 × g for 10 min and dried in afume hood at room temperature. The structure of starch digestawas characterized using SEC, DSC, XRD, and SEM. The amount ofglucose in the supernatant was determined using the Megazymed-glucose kit following the instructions from the manufacturer andconverted to the amount of digested starch using the factor of 0.9(the conversion factor for glucose to anhydroglucose unit in starch).The degree of starch hydrolysis was calculated as follows:
% hydrolysis
= total weight of glucose in supernatant × 0.9dry weight of starch
× 100%
The concentrations of pancreatin and amyloglucosidase (drystarch weight basis) were higher than previous studies (Hasjimet al., 2010a; Sopade & Gidley, 2009; Witt et al., 2010) in orderto produce substantial digestion of the native high-amylose maizeand normal potato starch granules, which contain large amountsof RS (Dhital et al., 2010; Jane et al., 2003; Tester, Qi, & Karkalas,2006). The in vitro digestion steps simulating the digestion in themouth and the stomach were not used in the present study as thestarch granules entering the small intestine from a previous in vivostudy were not significantly digested in the mouth or the stomach(Hasjim et al., 2010a). The digestion times were selected based onthe digestibility of the starches. Because the native high-amylosemaize and normal potato starch granules are more resistant toenzyme hydrolysis than the native waxy and normal maize starchgranules, the digestions of the native high-amylose maize and nor-mal potato starch granules were carried out for longer times.
2.3. Size-exclusion chromatography and amylose content
Starch digesta (∼2 mg) were dissolved in 1 mL DMSO solu-tion containing 0.5% w/w LiBr for 24 h at 80 ◦C and 350 rpm forthe characterization of whole molecular structure (Level 2 struc-ture) following the method of Syahariza, Li, and Hasjim (2010).The preparation of the individual branches of starch molecules(Level 1 structure) for characterization involved dissolving ∼6 mgstarch digesta in 1 mL DMSO solution containing 0.5% w/w LiBrfor 24 h at 80 ◦C and 350 rpm, precipitated using 5 mL absoluteethanol, dissolved in 0.9 mL distilled water in a boiling water bath,and then debranched using 2.5 �L isoamylase at 37 ◦C for 3 h (pHwas adjusted using 0.1 mL 0.1 M acetate buffer of pH 3.5) follow-ing the method described elsewhere (Hasjim et al., 2010a; Tranet al., 2011). The debranched starch was neutralized to pH ∼ 7 with100 �L 0.1 M NaOH, heated at 80 ◦C for 2 h, and freeze dried. The
dried debranched starch was redissolved in 1 mL DMSO solutioncontaining 0.5% w/w LiBr at 80 ◦C and 350 rpm for 24 h.The structures of whole and debranched starch molecules werecharacterized in duplicate using SEC, also known as gel permeation

1 ate Pol
c1wJG2dwcruawuh(
mpoaiuswe
V
wMscaei
b0ttm
2
tugXbadsdalL
2
aegm
2 A. Teng et al. / Carbohydr
hromatography (GPC). The SEC system consisted of an Agilent100 Series (Agilent Technologies, Waldbronn, Germany) equippedith a refractive index detector (RID-10A, Shimadzu Corp., Kyoto,
apan) following the method described elsewhere (Cave, Seabrook,idley, & Gilbert, 2009; Liu, Halley, & Gilbert, 2010; Witt et al.,010). GRAM 30 and GRAM 3000 SEC columns (Polymer Stan-ards Service GmbH, Mainz, Germany) were used to separatehole starch molecules, whereas GRAM 100 and GRAM 1000 SEC
olumns (Polymer Standards Service GmbH) were used to sepa-ate debranched starch molecules. DMSO with 0.5% w/w LiBr wassed as the eluent with flow rates of 0.3 and 0.6 mL/min for wholend debranched starch molecules, respectively. Pullulan standardsith peak molecular weights ranging from 342 to 2.35 × 106 weresed to obtain the universal calibration to convert elution volume toydrodynamic volume (Vh) or corresponding hydrodynamic radiusRh), where Vh = 4/3� Rh
3.The size distributions of whole and debranched starch
olecules are presented as the SEC weight distributions, w(log Vh),lotted against Rh, and normalized to either the yield of digestar the highest peak. The DP of debranched starch molecules waslso included in the plot as the secondary X-axis. Although theres no unique relationship between hydrodynamic size and molec-lar weight for any branched polymer, Vh or Rh of linear polymers,uch as those in debranched starch, can be converted to moleculareight or equivalent DP X using the Mark–Houwink equation (Liu
t al., 2010), as follows:
h = 2KM1+a
5NAwith M = 162 (X − 1) + 180
here NA is the Avogadro’s constant, K and ̨ are theark–Houwink parameters (0.0150 mL/g and 0.743 for linear
tarch, respectively), 162 is the molecular weight of anhydroglu-ose monomer in starch, and 180 is the molecular weight of glucoset the reducing end. The DP obtained using the Mark–Houwinkquation is, however, prone to error for small molecules and thuss only semi-quantitative.
The amylose content was analyzed from the SEC weight distri-ution of debranched starch molecules from the digesta collected at
h in vitro digestion. It was calculated as the ratio of the area underhe curve (AUC) of amylose branches to the AUC of the whole dis-ribution (both amylopectin and amylose branches) following the
ethod described elsewhere (Vilaplana, Hasjim, & Gilbert, 2012).
.4. X-ray diffractometry
XRD was performed using an XPert Pro multi-purpose diffrac-ometer (PANanalytical, Almelo, The Netherlands). The instrumentsed a copper long fine focus tube with an incident beam diver-ence slit, a diffracted beam scatter slit fixed at 0.125◦, and an’celerator high speed detector. The starch digesta were observedetween the angular range of 4◦ and 35◦ with a step size of 0.0334◦
nd a count time of 220 s per point. Approximately 2 g of each starchigesta, obtained by combining several replicates from the in vitrotarch digestion, was used to give a sample depth of 4 mm. Theata were analyzed using PeakFit v. 4.12 (Systat, San Jose, CA, USA),nd the percentages of A-, B- and V-type crystallinity were calcu-ated following the method described elsewhere (Li et al., 2015b;opez-Rubio, Flanagan, Gilbert, & Gidley, 2008a).
.5. Differential scanning calorimetry
The thermal properties of native starch and starch digesta were
nalyzed in duplicate using a DSC (DSC 1, Mettler Toledo, Schw-rzenbach, Switzerland). The thermal properties of native starchranules after being annealed at 37 ◦C for 16 h (waxy and nor-al maize starches) or 48 h (high-amylose maize and normalymers 139 (2016) 10–19
potato starches) without digestive enzymes were also determinedin duplicate to differentiate between the effects of incubation at ele-vated temperature and those of enzyme digestion on the thermalproperties of starch granules. Precisely weighed starch (approxi-mately 4–6 mg, dry weight basis) with excess water (three times theweight of starch) was placed in a high-pressure crucible pan withgold-plated copper sealing disk, and allowed to equilibrate for atleast 1 h at room temperature. The heating profile consisted of hold-ing the sample at 10 ◦C for 1 min, heating from 10 to 180 ◦C at a rateof 10 ◦C/min, rapid cooling from 180 to 10 ◦C at a rate of 40 ◦C/min,and reheating from 10 to 180 ◦C at a rate of 10 ◦C/min. The reheatingwas used to determine the endotherm associated with the meltingof amylose–lipid complexes, which reformed spontaneously dur-ing rapid cooling (Li et al., 2008). Indium was used for calibration,and an empty crucible pan with gold-plated copper sealing diskwas used as the reference. Onset temperature (To), peak tempera-ture (Tp), conclusion temperature (Tc), and enthalpy change (�H)were determined from each endotherm using the built-in soft-ware (STARe system, Mettler Toledo). Due to the overlapping ofstarch gelatinization and the melting of amylose–lipid complexesin the high-amylose maize starch granules, the �H of starch gela-tinization from the first heating was corrected by subtracting the�H of the melting of amylose–lipid complexes obtained from thereheating step. Furthermore, the high-pressure crucible pans usedin this study were made of stainless steel, which has poorer heatconductivity compared with those made of aluminum. Due to thislimitation, it was difficult to obtain precise information from broadsmall endotherms, such as the melting of amylose–lipid complexes.Hence, only the endotherm from starch gelatinization was dis-cussed quantitatively, whereas other smaller, broader peaks werediscussed qualitatively.
2.6. Scanning electron microscopy
Native starches and starch digesta were dried in a freeze dryer(BenchTop 2K, VirTis, Gardiner, NY, USA) overnight, and then placedonto specimen stubs with double-sided carbon tape. Each starchsample was coated three times with gold to a thickness of 20 nmusing a sputter coater (SPI-Module, SPI Supplies, West Chester, PA,USA). The direction of specimen was rotated to obtain an evenlayer on the surface. The granule morphology of starch samples wasexamined using an SEM (XL30, Philips, Eindoven, The Netherlands)with an accelerating voltage of 3 kV and a spot size of 6 nm.
2.7. Statistical analysis
The mean values of the gelatinization properties of starch wereanalyzed with Minitab 16 (Minitab Inc., State College, PA, USA)using the General Linear Model and Tukey’s Pairwise Comparisonswith a confidence level of 95.0% in performing an analysis of vari-ance (ANOVA) test.
3. Results
3.1. In vitro digestion of native starch granules
The digestograms of native waxy maize, normal maize, high-amylose maize, and normal potato starch granules are shownin Fig. 1A–D, respectively. The digestion of waxy and normalmaize starch granules reached completion with ∼100% of starchhydrolyzed at 8 and 24 h, respectively. Waxy and normal maizestarch granules also displayed rapid digestion from 0 to 4 h fol-
lowed by slower digestion after 4 h. On the other hand, the digestionof high-amylose maize and normal potato starch granules onlyreached 82% and 71% hydrolysis, respectively, after 48 h with nei-ther digestogram showing a plateau.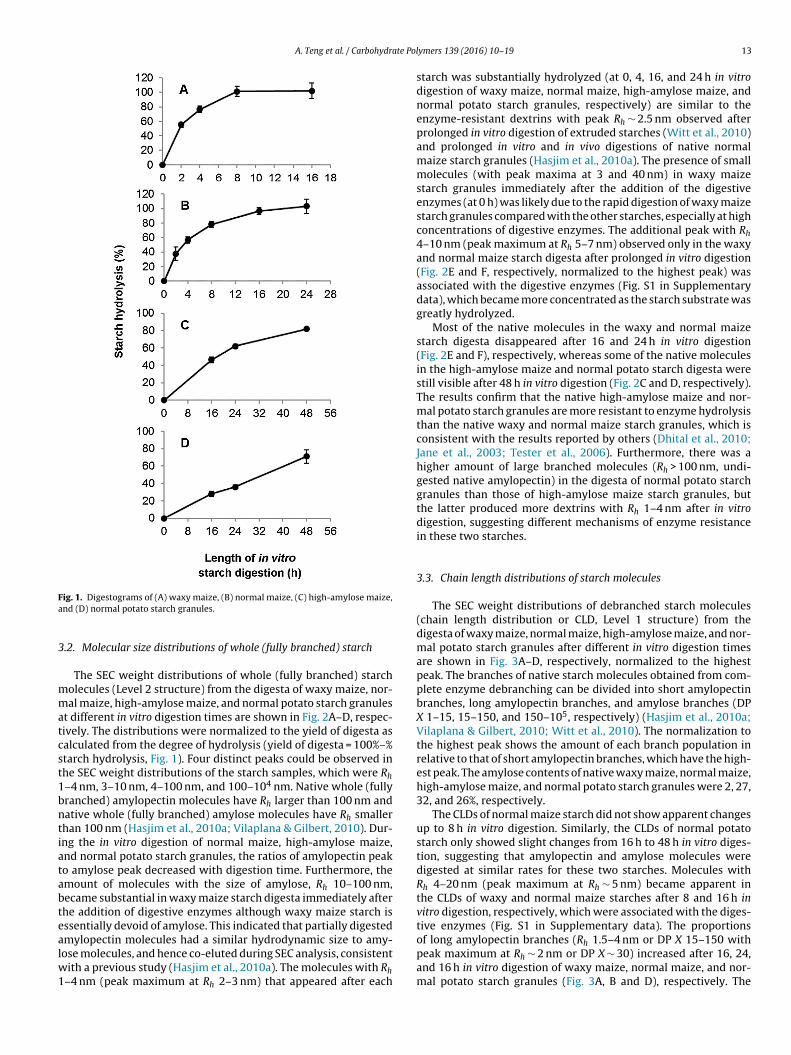
A. Teng et al. / Carbohydrate Pol
Fa
3
mmatcst1bntiatabtealw1
of long amylopectin branches (R 1.5–4 nm or DP X 15–150 with
ig. 1. Digestograms of (A) waxy maize, (B) normal maize, (C) high-amylose maize,nd (D) normal potato starch granules.
.2. Molecular size distributions of whole (fully branched) starch
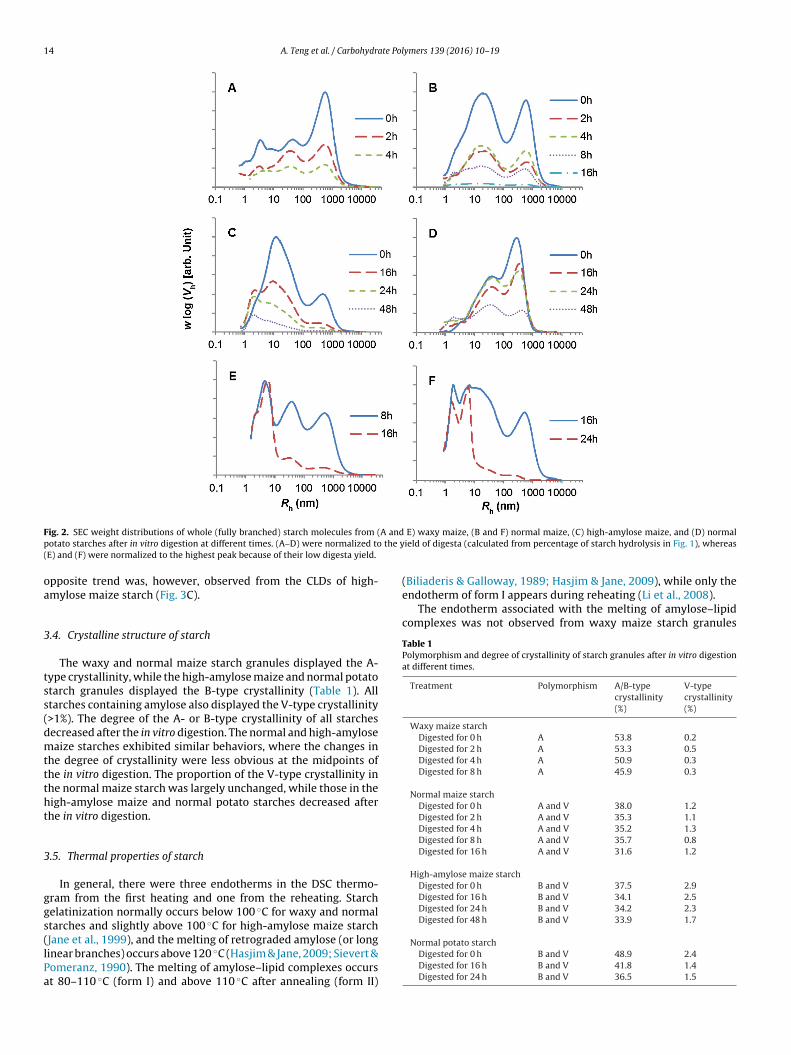
The SEC weight distributions of whole (fully branched) starcholecules (Level 2 structure) from the digesta of waxy maize, nor-al maize, high-amylose maize, and normal potato starch granules
t different in vitro digestion times are shown in Fig. 2A–D, respec-ively. The distributions were normalized to the yield of digesta asalculated from the degree of hydrolysis (yield of digesta = 100%–%tarch hydrolysis, Fig. 1). Four distinct peaks could be observed inhe SEC weight distributions of the starch samples, which were Rh–4 nm, 3–10 nm, 4–100 nm, and 100–104 nm. Native whole (fullyranched) amylopectin molecules have Rh larger than 100 nm andative whole (fully branched) amylose molecules have Rh smallerhan 100 nm (Hasjim et al., 2010a; Vilaplana & Gilbert, 2010). Dur-ng the in vitro digestion of normal maize, high-amylose maize,nd normal potato starch granules, the ratios of amylopectin peako amylose peak decreased with digestion time. Furthermore, themount of molecules with the size of amylose, Rh 10–100 nm,ecame substantial in waxy maize starch digesta immediately afterhe addition of digestive enzymes although waxy maize starch isssentially devoid of amylose. This indicated that partially digestedmylopectin molecules had a similar hydrodynamic size to amy-
ose molecules, and hence co-eluted during SEC analysis, consistentith a previous study (Hasjim et al., 2010a). The molecules with Rh–4 nm (peak maximum at Rh 2–3 nm) that appeared after each
ymers 139 (2016) 10–19 13
starch was substantially hydrolyzed (at 0, 4, 16, and 24 h in vitrodigestion of waxy maize, normal maize, high-amylose maize, andnormal potato starch granules, respectively) are similar to theenzyme-resistant dextrins with peak Rh ∼ 2.5 nm observed afterprolonged in vitro digestion of extruded starches (Witt et al., 2010)and prolonged in vitro and in vivo digestions of native normalmaize starch granules (Hasjim et al., 2010a). The presence of smallmolecules (with peak maxima at 3 and 40 nm) in waxy maizestarch granules immediately after the addition of the digestiveenzymes (at 0 h) was likely due to the rapid digestion of waxy maizestarch granules compared with the other starches, especially at highconcentrations of digestive enzymes. The additional peak with Rh4–10 nm (peak maximum at Rh 5–7 nm) observed only in the waxyand normal maize starch digesta after prolonged in vitro digestion(Fig. 2E and F, respectively, normalized to the highest peak) wasassociated with the digestive enzymes (Fig. S1 in Supplementarydata), which became more concentrated as the starch substrate wasgreatly hydrolyzed.
Most of the native molecules in the waxy and normal maizestarch digesta disappeared after 16 and 24 h in vitro digestion(Fig. 2E and F), respectively, whereas some of the native moleculesin the high-amylose maize and normal potato starch digesta werestill visible after 48 h in vitro digestion (Fig. 2C and D, respectively).The results confirm that the native high-amylose maize and nor-mal potato starch granules are more resistant to enzyme hydrolysisthan the native waxy and normal maize starch granules, which isconsistent with the results reported by others (Dhital et al., 2010;Jane et al., 2003; Tester et al., 2006). Furthermore, there was ahigher amount of large branched molecules (Rh > 100 nm, undi-gested native amylopectin) in the digesta of normal potato starchgranules than those of high-amylose maize starch granules, butthe latter produced more dextrins with Rh 1–4 nm after in vitrodigestion, suggesting different mechanisms of enzyme resistancein these two starches.
3.3. Chain length distributions of starch molecules
The SEC weight distributions of debranched starch molecules(chain length distribution or CLD, Level 1 structure) from thedigesta of waxy maize, normal maize, high-amylose maize, and nor-mal potato starch granules after different in vitro digestion timesare shown in Fig. 3A–D, respectively, normalized to the highestpeak. The branches of native starch molecules obtained from com-plete enzyme debranching can be divided into short amylopectinbranches, long amylopectin branches, and amylose branches (DPX 1–15, 15–150, and 150–105, respectively) (Hasjim et al., 2010a;Vilaplana & Gilbert, 2010; Witt et al., 2010). The normalization tothe highest peak shows the amount of each branch population inrelative to that of short amylopectin branches, which have the high-est peak. The amylose contents of native waxy maize, normal maize,high-amylose maize, and normal potato starch granules were 2, 27,32, and 26%, respectively.
The CLDs of normal maize starch did not show apparent changesup to 8 h in vitro digestion. Similarly, the CLDs of normal potatostarch only showed slight changes from 16 h to 48 h in vitro diges-tion, suggesting that amylopectin and amylose molecules weredigested at similar rates for these two starches. Molecules withRh 4–20 nm (peak maximum at Rh ∼ 5 nm) became apparent inthe CLDs of waxy and normal maize starches after 8 and 16 h invitro digestion, respectively, which were associated with the diges-tive enzymes (Fig. S1 in Supplementary data). The proportions
hpeak maximum at Rh ∼ 2 nm or DP X ∼ 30) increased after 16, 24,and 16 h in vitro digestion of waxy maize, normal maize, and nor-mal potato starch granules (Fig. 3A, B and D), respectively. The
14 A. Teng et al. / Carbohydrate Polymers 139 (2016) 10–19
F A andp the y(
oa
3
tss(dmtttht
3
ggs(lPa
(Biliaderis & Galloway, 1989; Hasjim & Jane, 2009), while only theendotherm of form I appears during reheating (Li et al., 2008).
The endotherm associated with the melting of amylose–lipidcomplexes was not observed from waxy maize starch granules
Table 1Polymorphism and degree of crystallinity of starch granules after in vitro digestionat different times.
Treatment Polymorphism A/B-typecrystallinity(%)
V-typecrystallinity(%)
Waxy maize starchDigested for 0 h A 53.8 0.2Digested for 2 h A 53.3 0.5Digested for 4 h A 50.9 0.3Digested for 8 h A 45.9 0.3
Normal maize starchDigested for 0 h A and V 38.0 1.2Digested for 2 h A and V 35.3 1.1Digested for 4 h A and V 35.2 1.3Digested for 8 h A and V 35.7 0.8Digested for 16 h A and V 31.6 1.2
High-amylose maize starchDigested for 0 h B and V 37.5 2.9Digested for 16 h B and V 34.1 2.5Digested for 24 h B and V 34.2 2.3Digested for 48 h B and V 33.9 1.7
ig. 2. SEC weight distributions of whole (fully branched) starch molecules from (otato starches after in vitro digestion at different times. (A–D) were normalized toE) and (F) were normalized to the highest peak because of their low digesta yield.
pposite trend was, however, observed from the CLDs of high-mylose maize starch (Fig. 3C).
.4. Crystalline structure of starch
The waxy and normal maize starch granules displayed the A-ype crystallinity, while the high-amylose maize and normal potatotarch granules displayed the B-type crystallinity (Table 1). Alltarches containing amylose also displayed the V-type crystallinity>1%). The degree of the A- or B-type crystallinity of all starchesecreased after the in vitro digestion. The normal and high-amyloseaize starches exhibited similar behaviors, where the changes in
he degree of crystallinity were less obvious at the midpoints ofhe in vitro digestion. The proportion of the V-type crystallinity inhe normal maize starch was largely unchanged, while those in theigh-amylose maize and normal potato starches decreased afterhe in vitro digestion.
.5. Thermal properties of starch
In general, there were three endotherms in the DSC thermo-ram from the first heating and one from the reheating. Starchelatinization normally occurs below 100 ◦C for waxy and normaltarches and slightly above 100 ◦C for high-amylose maize starch
Jane et al., 1999), and the melting of retrograded amylose (or longinear branches) occurs above 120 ◦C (Hasjim & Jane, 2009; Sievert &omeranz, 1990). The melting of amylose–lipid complexes occurst 80–110 ◦C (form I) and above 110 ◦C after annealing (form II)E) waxy maize, (B and F) normal maize, (C) high-amylose maize, and (D) normalield of digesta (calculated from percentage of starch hydrolysis in Fig. 1), whereas
Normal potato starchDigested for 0 h B and V 48.9 2.4Digested for 16 h B and V 41.8 1.4Digested for 24 h B and V 36.5 1.5

A. Teng et al. / Carbohydrate Pol
Fig. 3. Chain length distribution obtained by SEC analysis of debranched starchm(t
bnvgdtSmhtwmao
st
olecules from (A) waxy maize, (B) normal maize, (C) high-amylose maize, andD) normal potato starches after in vitro digestion at different times, normalized tohe highest peak.
ecause of the absence of amylose (Fig. 3A). In addition, it wasot observed from normal potato starch granules because onlyery small amount of lipids is present in normal potato starchranules although the V-type crystallinity was visible from XRDiffractograms (Table 1), probably due the higher sensitivity of XRDechnique or the presence of non-complexing single helices (Dhital,hrestha, Hasjim, & Gidley, 2011; McPherson & Jane, 1999). Theelting temperatures of amylose–lipid complexes of normal and
igh-amylose maize starch granules did not seem to be altered byhe in vitro digestion (Table S1 in Supplementary data), and henceere not further discussed. However, as expected, annealing of nor-al maize starch granules increased the melting temperature of
mylose–lipid complexes, which was attributed to the conversion
f form I complex to form II complex.The gelatinization temperatures of the waxy and normal maizetarch granules, in general, increased with the in vitro digestionime (Table 2). The increase in the gelatinization temperature was
ymers 139 (2016) 10–19 15
observed at a smaller extent from the high-amylose maize starchgranules, with only the native starch displaying a lower To, whereasthe normal potato starch granules did not show any significantchanges in the gelatinization temperature after the in vitro diges-tion. The gelatinization temperatures of waxy and normal maizestarch granules after 16 h in vitro digestion as well as the To ofhigh-amylose maize and normal potato starch granules after 48 hin vitro digestion were higher than their counterparts after beingannealed at the same temperature and time without the addition ofthe digestive enzymes. The �H of gelatinization of all starches didnot significantly change at the beginning of the in vitro digestionand then significantly decreased after prolonged in vitro digestion.Furthermore, the �H of gelatinization of waxy and normal maizestarch granules after 16 h in vitro digestion were significantly lowerthan their counterparts annealed at 37 ◦C without the addition ofdigestive enzymes. The results indicated that the changes in thegelatinization properties of starch granules, which were related tothe crystalline structure, after the in vitro digestion were not solelydue to the effects of annealing.
The starch crystallites with a high melting temperature(To > 120 ◦C), normally assigned as retrograded amylose (Hasjim& Jane, 2009; Sievert & Pomeranz, 1990), were detected in thedigesta of waxy maize starch granules, indicating that these crys-tallites were not solely originated from amylose chains. Althoughthe peak was small, it appeared consistently in all digesta of waxymaize starch granules at similar temperature range and was notdetected in the native and annealed waxy maize starch granules.Normal maize starch granules also displayed a high-temperatureendotherm after the in vitro digestion in similar temperaturerange to that of the digesta from waxy maize starch granules,with neither the native nor annealed counterparts displaying thisendotherm. High-amylose maize starch granules displayed similarhigh-temperature endotherm in all samples including the nativeand annealed starches, indicating that the highly ordered struc-ture formed during the in vitro digestion of waxy and normalmaize starch granules overlapped with the amylose crystallitespresent in the native high-amylose maize starch granules. The nor-mal potato starch granules did not show the high-temperatureendotherm before and after the in vitro digestion. There was no cleartrend between the �H from the melting of the high-temperatureendotherm and the in vitro starch digestion time; the inability toobserve any trends in the �H may be due to the small amount ofthe retrograded amylose/linear chains present in the ungelatinizedstarch granules, leading to the difficulties in accurately measuringthe �H.
3.6. Granule morphology of starch
The SEM images of native starch granules from waxy maize, nor-mal maize, high-amylose maize, and normal potato are shown inFig. 4A–D, respectively. The granules of waxy and normal maizestarch digesta displayed large pores on granule surface after thein vitro digestion (Fig. 4E and F, respectively). The pores on the sur-face of high-amylose maize and normal potato starch granules after24 h in vitro digestion were less apparent (Fig. 4G and H, respec-tively), but the granule surface was evidently rougher than that ofthe native starch granule counterparts, which was more obviousfor high-amylose maize starch digesta than normal potato starchdigesta. It is well documented that the surface of the B-type starchgranules (such as high-amylose maize and normal potato starches)is more resistant to enzyme hydrolysis than that of the A-typestarch granules (such as waxy and normal maize starches), preven-
ting the enzymes to hydrolyze the inner part of the granules (Dhitalet al., 2010). The rough granule surface of the B-type starch digestawas the result of enzyme hydrolysis as the enzymes were not ableto penetrate into the starch granules. Furthermore, the granules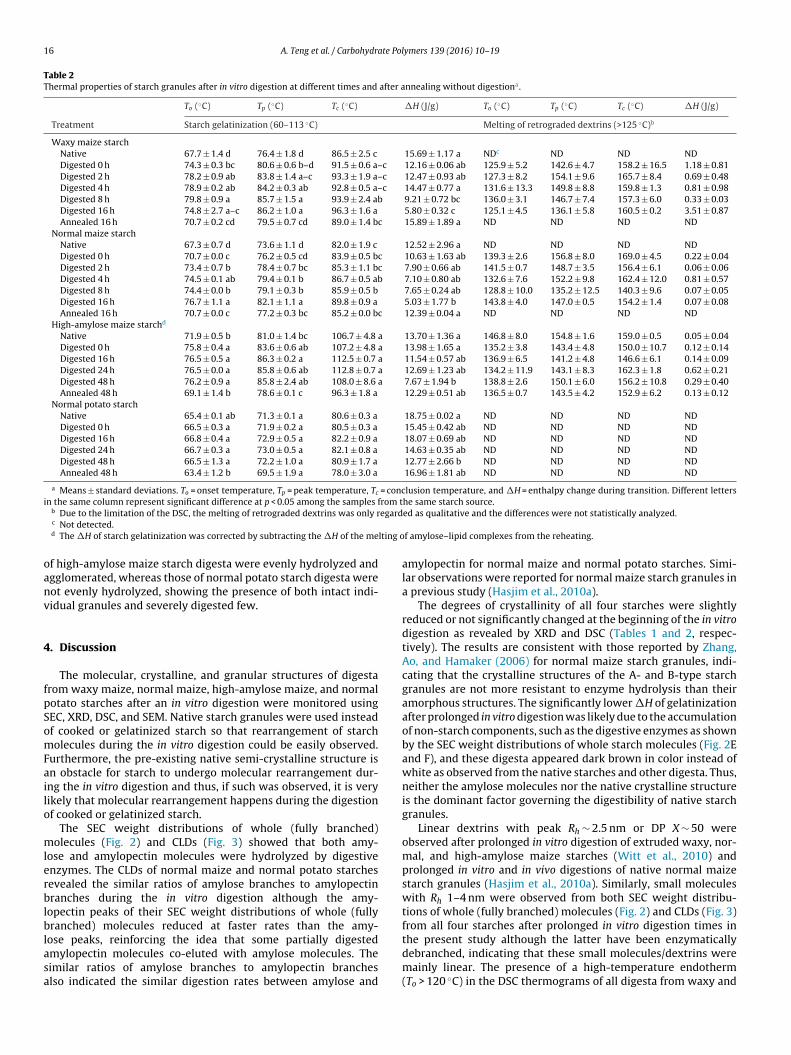
16 A. Teng et al. / Carbohydrate Polymers 139 (2016) 10–19
Table 2Thermal properties of starch granules after in vitro digestion at different times and after annealing without digestiona.
To (◦C) Tp (◦C) Tc (◦C) �H (J/g) To (◦C) Tp (◦C) Tc (◦C) �H (J/g)
Treatment Starch gelatinization (60–113 ◦C) Melting of retrograded dextrins (>125 ◦C)b
Waxy maize starchNative 67.7 ± 1.4 d 76.4 ± 1.8 d 86.5 ± 2.5 c 15.69 ± 1.17 a NDc ND ND NDDigested 0 h 74.3 ± 0.3 bc 80.6 ± 0.6 b–d 91.5 ± 0.6 a–c 12.16 ± 0.06 ab 125.9 ± 5.2 142.6 ± 4.7 158.2 ± 16.5 1.18 ± 0.81Digested 2 h 78.2 ± 0.9 ab 83.8 ± 1.4 a–c 93.3 ± 1.9 a–c 12.47 ± 0.93 ab 127.3 ± 8.2 154.1 ± 9.6 165.7 ± 8.4 0.69 ± 0.48Digested 4 h 78.9 ± 0.2 ab 84.2 ± 0.3 ab 92.8 ± 0.5 a–c 14.47 ± 0.77 a 131.6 ± 13.3 149.8 ± 8.8 159.8 ± 1.3 0.81 ± 0.98Digested 8 h 79.8 ± 0.9 a 85.7 ± 1.5 a 93.9 ± 2.4 ab 9.21 ± 0.72 bc 136.0 ± 3.1 146.7 ± 7.4 157.3 ± 6.0 0.33 ± 0.03Digested 16 h 74.8 ± 2.7 a–c 86.2 ± 1.0 a 96.3 ± 1.6 a 5.80 ± 0.32 c 125.1 ± 4.5 136.1 ± 5.8 160.5 ± 0.2 3.51 ± 0.87Annealed 16 h 70.7 ± 0.2 cd 79.5 ± 0.7 cd 89.0 ± 1.4 bc 15.89 ± 1.89 a ND ND ND ND
Normal maize starchNative 67.3 ± 0.7 d 73.6 ± 1.1 d 82.0 ± 1.9 c 12.52 ± 2.96 a ND ND ND NDDigested 0 h 70.7 ± 0.0 c 76.2 ± 0.5 cd 83.9 ± 0.5 bc 10.63 ± 1.63 ab 139.3 ± 2.6 156.8 ± 8.0 169.0 ± 4.5 0.22 ± 0.04Digested 2 h 73.4 ± 0.7 b 78.4 ± 0.7 bc 85.3 ± 1.1 bc 7.90 ± 0.66 ab 141.5 ± 0.7 148.7 ± 3.5 156.4 ± 6.1 0.06 ± 0.06Digested 4 h 74.5 ± 0.1 ab 79.4 ± 0.1 b 86.7 ± 0.5 ab 7.10 ± 0.80 ab 132.6 ± 7.6 152.2 ± 9.8 162.4 ± 12.0 0.81 ± 0.57Digested 8 h 74.4 ± 0.0 b 79.1 ± 0.3 b 85.9 ± 0.5 b 7.65 ± 0.24 ab 128.8 ± 10.0 135.2 ± 12.5 140.3 ± 9.6 0.07 ± 0.05Digested 16 h 76.7 ± 1.1 a 82.1 ± 1.1 a 89.8 ± 0.9 a 5.03 ± 1.77 b 143.8 ± 4.0 147.0 ± 0.5 154.2 ± 1.4 0.07 ± 0.08Annealed 16 h 70.7 ± 0.0 c 77.2 ± 0.3 bc 85.2 ± 0.0 bc 12.39 ± 0.04 a ND ND ND ND
High-amylose maize starchd
Native 71.9 ± 0.5 b 81.0 ± 1.4 bc 106.7 ± 4.8 a 13.70 ± 1.36 a 146.8 ± 8.0 154.8 ± 1.6 159.0 ± 0.5 0.05 ± 0.04Digested 0 h 75.8 ± 0.4 a 83.6 ± 0.6 ab 107.2 ± 4.8 a 13.98 ± 1.65 a 135.2 ± 3.8 143.4 ± 4.8 150.0 ± 10.7 0.12 ± 0.14Digested 16 h 76.5 ± 0.5 a 86.3 ± 0.2 a 112.5 ± 0.7 a 11.54 ± 0.57 ab 136.9 ± 6.5 141.2 ± 4.8 146.6 ± 6.1 0.14 ± 0.09Digested 24 h 76.5 ± 0.0 a 85.8 ± 0.6 ab 112.8 ± 0.7 a 12.69 ± 1.23 ab 134.2 ± 11.9 143.1 ± 8.3 162.3 ± 1.8 0.62 ± 0.21Digested 48 h 76.2 ± 0.9 a 85.8 ± 2.4 ab 108.0 ± 8.6 a 7.67 ± 1.94 b 138.8 ± 2.6 150.1 ± 6.0 156.2 ± 10.8 0.29 ± 0.40Annealed 48 h 69.1 ± 1.4 b 78.6 ± 0.1 c 96.3 ± 1.8 a 12.29 ± 0.51 ab 136.5 ± 0.7 143.5 ± 4.2 152.9 ± 6.2 0.13 ± 0.12
Normal potato starchNative 65.4 ± 0.1 ab 71.3 ± 0.1 a 80.6 ± 0.3 a 18.75 ± 0.02 a ND ND ND NDDigested 0 h 66.5 ± 0.3 a 71.9 ± 0.2 a 80.5 ± 0.3 a 15.45 ± 0.42 ab ND ND ND NDDigested 16 h 66.8 ± 0.4 a 72.9 ± 0.5 a 82.2 ± 0.9 a 18.07 ± 0.69 ab ND ND ND NDDigested 24 h 66.7 ± 0.3 a 73.0 ± 0.5 a 82.1 ± 0.8 a 14.63 ± 0.35 ab ND ND ND NDDigested 48 h 66.5 ± 1.3 a 72.2 ± 1.0 a 80.9 ± 1.7 a 12.77 ± 2.66 b ND ND ND NDAnnealed 48 h 63.4 ± 1.2 b 69.5 ± 1.9 a 78.0 ± 3.0 a 16.96 ± 1.81 ab ND ND ND ND
a Means ± standard deviations. To = onset temperature, Tp = peak temperature, Tc = conclusion temperature, and �H = enthalpy change during transition. Different lettersin the same column represent significant difference at p < 0.05 among the samples from the same starch source.
egard
lting o
oanv
4
fpSomFailo
mlerblblasa
b Due to the limitation of the DSC, the melting of retrograded dextrins was only rc Not detected.d The �H of starch gelatinization was corrected by subtracting the �H of the me
f high-amylose maize starch digesta were evenly hydrolyzed andgglomerated, whereas those of normal potato starch digesta wereot evenly hydrolyzed, showing the presence of both intact indi-idual granules and severely digested few.
. Discussion
The molecular, crystalline, and granular structures of digestarom waxy maize, normal maize, high-amylose maize, and normalotato starches after an in vitro digestion were monitored usingEC, XRD, DSC, and SEM. Native starch granules were used insteadf cooked or gelatinized starch so that rearrangement of starcholecules during the in vitro digestion could be easily observed.
urthermore, the pre-existing native semi-crystalline structure isn obstacle for starch to undergo molecular rearrangement dur-ng the in vitro digestion and thus, if such was observed, it is veryikely that molecular rearrangement happens during the digestionf cooked or gelatinized starch.
The SEC weight distributions of whole (fully branched)olecules (Fig. 2) and CLDs (Fig. 3) showed that both amy-
ose and amylopectin molecules were hydrolyzed by digestivenzymes. The CLDs of normal maize and normal potato starchesevealed the similar ratios of amylose branches to amylopectinranches during the in vitro digestion although the amy-
opectin peaks of their SEC weight distributions of whole (fullyranched) molecules reduced at faster rates than the amy-
ose peaks, reinforcing the idea that some partially digestedmylopectin molecules co-eluted with amylose molecules. Theimilar ratios of amylose branches to amylopectin brancheslso indicated the similar digestion rates between amylose and
ed as qualitative and the differences were not statistically analyzed.
f amylose–lipid complexes from the reheating.
amylopectin for normal maize and normal potato starches. Simi-lar observations were reported for normal maize starch granules ina previous study (Hasjim et al., 2010a).
The degrees of crystallinity of all four starches were slightlyreduced or not significantly changed at the beginning of the in vitrodigestion as revealed by XRD and DSC (Tables 1 and 2, respec-tively). The results are consistent with those reported by Zhang,Ao, and Hamaker (2006) for normal maize starch granules, indi-cating that the crystalline structures of the A- and B-type starchgranules are not more resistant to enzyme hydrolysis than theiramorphous structures. The significantly lower �H of gelatinizationafter prolonged in vitro digestion was likely due to the accumulationof non-starch components, such as the digestive enzymes as shownby the SEC weight distributions of whole starch molecules (Fig. 2Eand F), and these digesta appeared dark brown in color instead ofwhite as observed from the native starches and other digesta. Thus,neither the amylose molecules nor the native crystalline structureis the dominant factor governing the digestibility of native starchgranules.
Linear dextrins with peak Rh ∼ 2.5 nm or DP X ∼ 50 wereobserved after prolonged in vitro digestion of extruded waxy, nor-mal, and high-amylose maize starches (Witt et al., 2010) andprolonged in vitro and in vivo digestions of native normal maizestarch granules (Hasjim et al., 2010a). Similarly, small moleculeswith Rh 1–4 nm were observed from both SEC weight distribu-tions of whole (fully branched) molecules (Fig. 2) and CLDs (Fig. 3)from all four starches after prolonged in vitro digestion times in
the present study although the latter have been enzymaticallydebranched, indicating that these small molecules/dextrins weremainly linear. The presence of a high-temperature endotherm(To > 120 ◦C) in the DSC thermograms of all digesta from waxy and
A. Teng et al. / Carbohydrate Polymers 139 (2016) 10–19 17
F maizes orma
notssahaX(pg
tts
ig. 4. SEM images of native (A) waxy maize, (B) normal maize, (C) high-amylose
tarch granules after 4 hour in vitro digestion and (G) high-amylose maize and (H) n
ormal maize starch granules, which was not observed in thosef the native and annealed starch granule counterparts, suggestedhat the linear dextrins produced during the in vitro digestion oftarch granules could rearrange into thermally stable crystallinetructure. This crystalline structure is similar to that of retrogradedmylose, which is thermally stable and highly resistant to enzymeydrolysis (Gidley et al., 1995; Sievert & Pomeranz, 1990). It waslso reported that enzyme-resistant retrograded amylose had DP
∼ 50 after its amorphous fraction was removed by �-amylaseJane & Robyt, 1984), similar to the size of linear dextrins afterrolonged in vitro digestions of waxy and normal maize starchranules.
Different from the waxy and normal maize starch granules,he high-amylose maize starch granules displayed the high-emperature endotherm in the DSC thermograms at the nativetate and after the in vitro digestion. Amylose double helices were
, and (D) normal potato starch granules as well as (E) waxy and (F) normal maizel potato starch granules after 24 hour in vitro digestion.
reported to be present in the native high-amylose maize starchgranules, responsible for their high RS contents (Jiang, Srichuwong,Campbell, & Jane, 2010). Furthermore, the ratio of the dextrins withRh 1–4 nm to the native molecules (Rh > 4 nm) in the SEC weightdistributions of whole (fully branched) molecules for the high-amylose maize starch was evidently increased with the in vitrodigestion time (Fig. 2C), indicating that the native molecules mightbe hydrolyzed to form enzyme-resistant linear dextrins with Rh1–4 nm. The increase of the linear dextrins with Rh 1–4 nm was,however, not observed from the CLDs of the high-amylose maizestarch (Fig. 3C), which could be due to the rapid hydrolysis oflong amylopectin branches to similar size as the short amylopectin
branches. Therefore, the presence of enzyme-resistant crystallinestructure with high melting temperature (To > 120 ◦C) is mainlyresponsible for the low digestibility of native high-amylose maizestarch granules.
1 ate Pol
mgDTabttbuwstspHuoabtoedtpdfrms
hd(oaimgetsh
5
ntTntshymemmsese
8 A. Teng et al. / Carbohydr
Despite having higher enzyme resistance than the high-amyloseaize starch granules (Fig. 1C and D), the normal potato starch
ranules did not display the high-temperature endotherm in theirSC thermograms before and after the in vitro digestion (Table 2).he digesta of normal potato starch also contained a smallermount of dextrins with Rh 1–4 nm, but a higher amount of largeranched molecules (Rh > 100 nm, undigested native amylopectin)han those of high-amylose maize starch (Fig. 2D and C, respec-ively). Furthermore, the CLDs showed that the ratio of amyloseranches to amylopectin branches of normal potato starch gran-les did not change much during the in vitro digestion (Fig. 3D),hereas that of high-amylose maize starch granules changed with
hort amylopectin branches dominating after 48 h in vitro diges-ion (Fig. 3C). The high-amylose maize starch granules had rougherurface structure and were more agglomerated than the normalotato starch granules after 24 h in vitro digestion (Fig. 4G and, respectively), suggesting that the normal potato starch gran-les were more resistant to enzyme hydrolysis. Thus, the presencef large amylopectin in normal potato starch digesta could bettributed to the granular structure protecting the molecules fromeing hydrolyzed by enzymes, reducing the amount of linear dex-rins with Rh 1–4 nm in the digesta. Differences in the susceptibilityf granules from different starches to enzyme hydrolysis are wellstablished, with potato starch granules particularly resistant asemonstrated by the lack of surface pores and internal void struc-ure (Dhital et al., 2010; Jane et al., 2003; Tester et al., 2006). Theresence of intact granular structure in the normal potato starchigesta could also create a restriction for the linear dextrins toorm thermally stable crystalline structure. This is also the maineason for the difference between the digestion profiles of nor-al maize and normal potato starch granules although they have
imilar amylose contents (Fig. 1B and D, respectively).In general, the gelatinization temperatures of waxy, normal, and
igh-amylose maize starch granules increased during the in vitroigestion although the extents depended on the types of starchTable 2). The increase could be due to the more rapid hydrolysisf less perfectly aligned crystallites compared with more perfectlyligned crystallites, which the latter had higher thermal stabil-ty. The hydrolysis could also reduce the steric restriction for the
olecules to rearrange into more perfectly aligned crystallites. Theelatinization temperature of normal potato starch granules, how-ver, did not significantly change during the in vitro digestion ashe highly enzyme-resistant granular structure of normal potatotarch protected the less perfectly aligned crystallites from beingydrolyzed by the enzymes.
. Conclusions
All starches showed that amylose and amylopectin molecules inative starch granules were hydrolyzed to smaller dextrins duringhe in vitro digestion, and some small dextrins had linear structure.he degrees of crystallinity of all starches were slightly reduced orot significantly changed after digestion. However, the gelatiniza-ion temperatures of the waxy, normal, and high-amylose maizetarches increased after the in vitro digestion, indicating that theeat labile crystallites were more susceptible to enzyme hydrol-sis and/or the hydrolyzed starch molecules could rearrange intoore perfectly aligned crystalline structure. A high-temperature
ndotherm appeared in the DSC thermograms of waxy and normalaize starches after the in vitro digestion as well as in high-amyloseaize starch before and after the in vitro digestion, which was
imilar to that of retrograded amylose, suggesting that the lin-ar dextrins could form highly ordered crystalline structure withlow digestion properties and were mainly responsible for the highnzyme resistance of native high-amylose maize starch granules.
ymers 139 (2016) 10–19
On the other hand, normal potato starch granules did not show thehigh-temperature endotherm before and after digestion althoughthey had higher enzyme resistance than high-amylose maize starchgranules. The presence of intact individual granules lacking ofsurface pores in the normal potato starch digesta indicated thatthe surface structure of normal potato starch granules protectedthe molecules inside the granules from being hydrolyzed by theenzymes.
Acknowledgements
The authors thank the Australian Research Council (DP0985694and LP0990769) for funding.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.carbpol.2015.11.062.
References
Ai, Y., Hasjim, J., & Jane, J. (2013). Effects of lipids on enzymatic hydrolysis andphysical properties of starch. Carbohydrate Polymers, 92(1), 120–127.
Biliaderis, C. G., & Galloway, G. (1989). Crystallization behavior of amylose-Vcomplexes: Structure–property relationships. Carbohydrate Research, 189,31–48.
Byrnes, S. E., Brand Miller, J. C., & Denyer, G. S. (1995). Amylopectin starchpromotes the development of insulin resistance in rats. Journal of Nutrition,125(6), 1430–1437.
Cave, R. A., Seabrook, S. A., Gidley, M. J., & Gilbert, R. G. (2009). Characterization ofstarch by size-exclusion chromatography: The limitations imposed by shearscission. Biomacromolecules, 10(8), 2245–2253.
Dhital, S., Shrestha, A. K., & Gidley, M. J. (2010). Effect of cryo-milling on starches:Functionality and digestibility. Food Hydrocolloids, 24(2–3), 152–163.
Dhital, S., Shrestha, A. K., Hasjim, J., & Gidley, M. J. (2011). Physicochemical andstructural properties of maize and potato starches as a function of granule size.Journal of Agricultural and Food Chemistry, 59(18), 10151–10161.
Dona, A. C., Pages, G., Gilbert, R. G., & Kuchel, P. W. (2010). Digestion of starch:In vivo and in vitro kinetic models used to characterise oligosaccharide orglucose release. Carbohydrate Polymers, 80(3), 599–617.
Englyst, H. N., Kingman, S. M., & Cummings, J. H. (1992). Classification andmeasurement of nutritionally important starch fractions. European Journal ofClinical Nutrition, 46(Suppl 2), S33–S50.
Ferguson, L. R., Tasman-Jones, C., Englyst, H., & Harris, P. J. (2000). Comparativeeffects of three resistant starch preparations on transit time and short-chainfatty acid production in rats. Nutrition and Cancer, 36(2), 230–237.
Gidley, M. J., Cooke, D., Darke, A. H., Hoffmann, R. A., Russell, A. L., & Greenwell, P.(1995). Molecular order and structure in enzyme-resistant retrograded starch.Carbohydrate Polymers, 28(1), 23–31.
Hasjim, J., Cesbron Lavau, G., Gidley, M. J., & Gilbert, R. G. (2010). In vivo andin vitro starch digestion: Are current in vitro techniques adequate?Biomacromolecules, 11(12), 3600–3608.
Hasjim, J., & Jane, J. (2009). Production of resistant starch by extrusion cooking ofacid-modified normal-maize starch. Journal of Food Science, 74(7), 556–562.
Hasjim, J., Lee, S.-O., Hendrich, S., Setiawan, S., Ai, Y., & Jane, J. (2010).Characterization of a novel resistant-starch and its effects on postprandialplasma-glucose and insulin responses. Cereal Chemistry, 87(4), 257–262.
Hasjim, J., Srichuwong, S., Scott, M. P., & Jane, J. (2009). Kernel composition, starchstructure, and enzyme digestibility of opaque-2 maize and quality proteinmaize. Journal of Agricultural and Food Chemistry, 57(5), 2049–2055.
Htoon, A., Shrestha, A. K., Flanagan, B. M., Lopez-Rubio, A., Bird, A. R., Gilbert, E. P.,et al. (2009). Effects of processing high amylose maize starches undercontrolled conditions on structural organisation and amylase digestibility.Carbohydrate Polymers, 75(2), 236–245.
Jane, J., Ao, Z., Duvick, S. A., Wiklund, M., Yoo, S.-H., Wong, K.-S., et al. (2003).Structures of amylopectin and starch granules: How are they synthesized?Journal of Applied Glycoscience, 50(2), 167–172.
Jane, J., Chen, Y. Y., Lee, L. F., McPherson, A. E., Wong, K. S., Radosavljevic, M., et al.(1999). Effects of amylopectin branch chain length and amylose content on thegelatinization and pasting properties of starch. Cereal Chemistry, 76(5),629–637.
Jane, J., & Robyt, J. F. (1984). Structure studies of amylose-V complexes andretrograded amylose by action of alpha amylases, and a new method forpreparing amylodextrins. Carbohydrate Research, 132(1), 105–118.
Jiang, H., Srichuwong, S., Campbell, M., & Jane, J. (2010). Characterization of maizeamylose-extender (ae) mutant starches. Part III: Structures and properties ofthe Naegeli dextrins. Carbohydrate Polymers, 81(4), 885–891.
Li, L., Jiang, H., Campbell, M., Blanco, M., & Jane, J. (2008). Characterization of maizeamylose-extender (ae) mutant starches, Part I: Relationship between resistant

te Pol
L
L
L
L
L
L
M
S
S
S
A. Teng et al. / Carbohydra
starch contents and molecular structures. Carbohydrate Polymers, 74(3),396–404.
i, M., Witt, T., Xie, F., Warren, F. J., Halley, P. J., & Gilbert, R. G. (2015).Biodegradation of starch films: The roles of molecular and crystallinestructure. Carbohydrate Polymers, 122(0), 115–122.
i, M., Xie, F., Hasjim, J., Witt, T., Halley, P. J., & Gilbert, R. G. (2015). Establishingwhether the structural feature controlling the mechanical properties of starchfilms is molecular or crystalline. Carbohydrate Polymers, 117(0), 262–270.
iu, W.-C., Halley, P. J., & Gilbert, R. G. (2010). Mechanism of degradation ofstarch, a highly branched polymer, during extrusion. Macromolecules, 43(6),2855–2864.
opez-Rubio, A., Flanagan, B. M., Gilbert, E. P., & Gidley, M. J. (2008). A novelapproach for calculating starch crystallinity and its correlation with doublehelix content: A combined XRD and NMR study. Biopolymers, 89(9), 761–768.
opez-Rubio, A., Flanagan, B. M., Shrestha, A. K., Gidley, M. J., & Gilbert, E. P. (2008).Molecular rearrangement of starch during in vitro digestion: Toward a betterunderstanding of enzyme resistant starch formation in processed starches.Biomacromolecules, 9(7), 1951–1958.
udwig, D. S. (2002). The glycemic index: Physiological mechanisms relating toobesity, diabetes, and cardiovascular disease. Journal of the American MedicalAssociation, 287(18), 2414–2423.
cPherson, A. E., & Jane, J. (1999). Comparison of waxy potato with other root andtuber starches. Carbohydrate Polymers, 40(1), 57–70.
hrestha, A. K., Ng, C. S., Lopez-Rubio, A., Blazek, J., Gilbert, E. P., & Gidley, M. J.(2010). Enzyme resistance and structural organization in extruded highamylose maize starch. Carbohydrate Polymers, 80(3), 699–710.
ievert, D., & Pomeranz, Y. (1990). Enzyme-resistant starch, II. Differential
scanning calorimetry studies on heat-treated starches and enzyme-resistantstarch residues. Cereal Chemistry, 67(3), 217–221.opade, P. A., & Gidley, M. J. (2009). A rapid in-vitro digestibility assay based onglucometry for investigating kinetics of starch digestion. Starch, 61(5),245–255.
ymers 139 (2016) 10–19 19
Srichuwong, S., Sunarti, T. C., Mishima, T., Isono, N., & Hisamatsu, M. (2005).Starches from different botanical sources I: Contribution of amylopectin finestructure to thermal properties and enzyme digestibility. CarbohydratePolymers, 60(4), 529–538.
Syahariza, Z. A., Li, E., & Hasjim, J. (2010). Extraction and dissolution of starch fromrice and sorghum grains for accurate structural analysis. CarbohydratePolymers, 82(1), 14–20.
Syahariza, Z. A., Sar, S., Hasjim, J., Tizzotti, M. J., & Gilbert, R. G. (2013). Theimportance of amylose and amylopectin fine structures for starch digestibilityin cooked rice grains. Food Chemistry, 136(2), 742–749.
Tester, R. F., Qi, X., & Karkalas, J. (2006). Hydrolysis of native starches withamylases. Animal Feed Science and Technology, 130(1–2), 39–54.
Tran, T. T. B., Shelat, K. J., Tang, D., Li, E., Gilbert, R. G., & Hasjim, J. (2011). Milling ofrice grains. The degradation on three structural levels of starch in rice flour canbe independently controlled during grinding. Journal of Agricultural and FoodChemistry, 59(8), 3964–3973.
Vilaplana, F., & Gilbert, R. G. (2010). Two-dimensional size/branch lengthdistributions of a branched polymer. Macromolecules, 43(17), 7321–7329.
Vilaplana, F., Hasjim, J., & Gilbert, R. G. (2012). Amylose content in starches:Toward optimal definition and validating experimental methods. CarbohydratePolymers, 88(1), 103–111.
Willett, W., Manson, J., & Liu, S. (2002). Glycemic index, glycemic load, and risk oftype 2 diabetes. The American Journal of Clinical Nutrition, 76(1), 274S–280S.
Witt, T., Gidley, M. J., & Gilbert, R. G. (2010). Starch digestion mechanisticinformation from the time evolution of molecular size distributions. Journal ofAgricultural and Food Chemistry, 58(14), 8444–8452.
Zhang, G., Ao, Z., & Hamaker, B. R. (2006). Slow digestion property of native cereal
starches. Biomacromolecules, 7(11), 3252–3258.Zhao, Y., Hasjim, J., Li, L., Jane, J., Hendrich, S., & Birt, D. F. (2011). Inhibition ofazoxymethane-induced preneoplastic lesions in the rat colon by a cookedstearic acid complexed high-amylose cornstarch. Journal of Agricultural andFood Chemistry, 59(17), 9700–9708.





























