Embed Size (px)
Citation preview

Research Article
Molecular Profiling of Premalignant Lesions in LungSquamous Cell Carcinomas Identifies MechanismsInvolved in Stepwise Carcinogenesis
Aik T. Ooi1, Adam C. Gower9, Kelvin X. Zhang4, Jessica L. Vick9, Longsheng Hong3, Brian Nagao3,W. Dean Wallace3, David A. Elashoff5,7, Tonya C. Walser2,7, Steven M. Dubinett2,3,7, Matteo Pellegrini6,8,Marc E. Lenburg9, Avrum Spira9, and Brigitte N. Gomperts1,2,7,8
AbstractLung squamous cell carcinoma (SCC) is thought to arise from premalignant lesions in the airway
epithelium; therefore, studying these lesions is critical for understanding lung carcinogenesis. Previous
microarray and sequencing studies designed to discover early biomarkers and therapeutic targets for lung
SCC had limited success identifying key driver events in lung carcinogenesis, mostly due to the cellular
heterogeneity of patient samples examined and the interindividual variability associated with difficult to
obtain airway premalignant lesions and appropriate normal control samples within the same patient. We
performedRNA sequencing on laser-microdissected representative cell populations along the SCCpathologic
continuum of patient-matched normal basal cells, premalignant lesions, and tumor cells. We discovered
transcriptomic changes and identified genomic pathways altered with initiation and progression of SCC
within individual patients. We used immunofluorescent staining to confirm gene expression changes in
premalignant lesionsand tumor cells, including increasedexpressionofSLC2A1,CEACAM5,andPTBP3at the
protein level and increased activation of MYC via nuclear translocation. Cytoband enrichment analysis
revealed coordinated loss and gain of expression in chromosome 3p and 3q regions, respectively, during
carcinogenesis. This is the first gene expression profiling study of airway premalignant lesions with patient-
matched SCC tumor samples. Our results provide much needed information about the biology of prema-
lignant lesions and themolecular changes thatoccur during stepwise carcinogenesis of SCC, and it highlights a
novel approach for identifying some of the earliest molecular changes associated with initiation and
progression of lung carcinogenesis within individual patients. Cancer Prev Res; 7(5); 487–95. �2014 AACR.
IntroductionLung cancer is the most deadly cancer worldwide,
accounting for more deaths than prostate cancer, breastcancer, pancreatic cancer, and colon cancer combined(1). Squamous cell carcinoma (SCC) is a common type
of non–small cell lung cancer that accounts for 30% of alllung cancers and is frequently associated with smoking(2). In general, despite current therapeutic strategies ofchemotherapy, radiotherapy, and trials with targetedtherapies, the overall survival of patients with lung can-cer, including SCC, is still very poor with a 5-year survivalrate of 15.9% (3).
SCC often arises centrally from a large airway, usually abronchus. Ongoing injury of airway epithelia leads to repairand regeneration that can give rise to a phenotype ofsquamous metaplasia and subsequently to dysplasia, bothof which are histologic features seen in the airways ofsmokers (4, 5). It is believed that SCC develops througha series of genetic and epigenetic changes that alter theepithelium from squamous metaplasia, then to dysplasia,carcinoma in situ and finally to invasive carcinoma (6).
Although there have been studies devoted to discoveringthe genetic and molecular changes observed in lung cancer,few studies have directly investigated changes associatedwith squamous metaplasia or dysplasia (7–9). In fact, it isnot known with certainty whether premalignant lesions ofthe airway are the direct progenitors of invasive SCC. This ismainly due to the challenge inherent in following airway
Authors' Affiliations: 1Department of Pediatrics, Mattel Children's Hos-pital; 2Division of Pulmonary and Critical Care Medicine, David GeffenSchool of Medicine; Departments of 3Pathology and Laboratory Medicineand 4Biological Chemistry, HowardHughesMedical Institute; Departmentsof 5Biostatistics, 6Molecular, Cell, and Developmental Biology; 7The LungCancer Research Program of the Jonsson Comprehensive Cancer Center;8Broad Stem Cell Research Center, University of California, Los Angeles,California; and 9Section of Computational Biomedicine, Department ofMedicine, Boston University School of Medicine, Boston, Massachusetts
Note:Supplementary data for this article are available atCancer PreventionResearch Online (http://cancerprevres.aacrjournals.org/).
A. Spira and B.N. Gomperts share senior authorship for this article.
Corresponding Author: Brigitte N. Gomperts, Mattel Children's Hospital,University of California, LosAngeles, 10833 LeConteAvenueA2-410MDCC,Los Angeles, CA 90095. Phone: 310-206-0772; Fax: 310-206-8089; E-mail:[email protected]
doi: 10.1158/1940-6207.CAPR-13-0372
�2014 American Association for Cancer Research.
CancerPreventionResearch
www.aacrjournals.org 487
Research. on May 27, 2018. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2014; DOI: 10.1158/1940-6207.CAPR-13-0372

premalignant lesionsseriallyover time in the largeairways todeterminewhether a particular lesion is destined to developintoSCC.Tobetter understand theprocess of carcinogenesisleading to SCC, especially those steps involved in the earlyand precancerous stages, a comprehensive study of themolecular alterations that characterize premalignant lesionsis needed alongwith adirect comparisonwith themolecularchanges found in SCC from that same individual.
Basal cells of the airway are known to be stem/progenitorcells required for airway epithelial repair (10), and wehypothesize that premalignant lesions arise from aberrantrepair in these cells (11). Therefore, we profiled the tran-scriptome of airway basal cell, premalignant lesions, andtumors from the same patients to improve our understand-ing of the stepwise carcinogenesis in SCC and to aid in theidentificationofnewdiagnostic and therapeutic approachesfor SCC and novel chemopreventive strategies.
Materials and MethodsCase selection and histology review
Resected tissue blocks fromSCC caseswere reviewedwithtwo pathologists to identify regions of normal airway epi-thelium, squamousmetaplasia or dysplasia, or carcinomas.Patients with fresh-frozen or formalin-fixed paraffin-embedded (FFPE) tissue blocks containing all three regionswere selected for the study. Immunofluorescent staining ofKRT5 was performed to validate the identification of select-ed lesions. Fresh-frozen tissues were used for RNA sequenc-ing, whereas FFPE tissues were used for validation of inde-pendent cases with quantitative real-time PCR (qRT-PCR)and immunofluorescent staining.
Laser capture microdissectionTissues were sectioned at a 7-mm thickness and mounted
on regular uncharged glass slides for patients 1, 2, and3, andon polyethylene napthalate membrane slides (Leica) forpatient 4, followedbyhematoxylin and eosin staining. Lasercapture microdissection was performed using the ArcturuseIIx for patient 1 and 2, Zeiss PALM for patient 3, and LeicaLMD7000 for patient 4. A tissue area of 800,000 to1,200,000mm2wasdissectedandcollected fromeach lesion.
RNA extraction and sequencing library preparationRNA was extracted from laser-microdissected cells using
the RNeasy Micro Kit (Qiagen). The cDNA was generatedusing the Ovation RNA-Seq System (NuGEN) for patients 1and 2 and the Ovation RNA-Seq V2 System (NuGEN) forpatients3and4.Forpatients1and2,cDNAofapproximately200 bpwas selected by gel purification. For patients 3 and 4,the cDNA was sheared to 140 to 180 bp using the Covarisfocused-ultrasonicatorwiththefollowingsettings:dutycycle10%; intensity 5; cycles per burst 200; total time 6 minutes.The size range of the sheared cDNA was confirmed byBioanalyzer analysis before library construction using theEncore Library System (NuGEN). The average size of eachlibrary was estimated by Bioanalyzer analysis, and the con-centration of each was measured on the Qubit fluorometer(Invitrogen).
Sequence analysisSequencing libraries from patients 1 and 2 were each
sequenced on a single flow cell lane of an IlluminaGenomeAnalyzer IIx, generating 36-base single-end reads,and libraries frompatients 3 and 4were each sequenced ona single flow cell lane of an Illumina HiSeq 2000, gener-ating 50-base single-end reads. All reads were trimmed to35 bases before alignment. In the case of patient 1, the firstbase of each read was also trimmed-off due to a problemwith the first sequencing cycle. Reads that failed the chas-tity filter of Illumina [brightest intensity/(brightest inten-sityþ secondbrightest intensity)<0.6 for at least twoof thefirst 25 cycles] were automatically removed during pre-processing. The remaining reads were aligned to thehuman genome (build hg19) using Bowtie v0.12.7 (12),allowing only unique alignments and up to two mis-matches per read. Reads aligning to the mitochondrialgenome were removed from further analysis. Gene expres-sion estimates were then computed by measuring thecoverage of each of 55,841 Ensembl Gene loci (Ensemblbuild 69) using the BEDTools software suite (13). Thecoverage for each Ensembl Gene locus in each sample wasthen normalized to the size of the locus and the totalnumber of readsmapping uniquely to the nuclear genometo obtain an RPKM (14) value for each gene in eachsample. RPKM values were seventh root-transformedbefore analysis to produce an approximately normal dis-tribution of (nonzero) gene expression values.
Statistical analysisAll models were created using the R environment for
statistical computing (version 2.12.0). Linear mixed-effectsmodels were created using the nlme R package (version 3.1-97) and negative binomial models were created using theMASS R package (version 7.3-7). The Student two-sample ttest with equal variance (or, in the case of GDS1312,Student paired t test) was used to assess the significance ofdifferential expression of candidate genes in Gene Expres-sion Omnibus (GEO) datasets. Analysis of GEO datasetswas performed using the preprocessed expression levelsgenerated using default Affymetrix probeset mappings(averaging across multiple probesets to obtain a singleexpression value for each gene).
Gene set enrichment analysisPositionally defined (cytoband) Ensembl Gene sets were
created using the biomaRtRpackage to extract chromosomalband annotation for Ensembl Gene identifiers usingEnsembl version 69. These gene sets were then used toperform preranked GSEA (gene set enrichment analysis;ref. 15) using lists of all Ensembl Genes ranked by the tstatistics from the linear mixed-effects models, to identifycytobands that were overrepresented among genes coordi-nately up- or downregulated in premalignant or tumor cellscompared with normal basal cell. Analysis was performedusing GSEA v2.0.8 (build 14) with 1,000 permutations,removal of gene sets with >500 genes, and a random seed of1,234.
Ooi et al.
Cancer Prev Res; 7(5) May 2014 Cancer Prevention Research488
Research. on May 27, 2018. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2014; DOI: 10.1158/1940-6207.CAPR-13-0372

Quantitative real-time PCRAmplified cDNAgenerated during the library preparation
for patients 3 and 4 was used for qRT-PCR analysis. Inaddition, normal basal cell andpremalignant lesions from4independent patients were laser microdissected from FFPEtissues, and cDNAwas generated using theOvationRNASeqFFPE System. TaqMan Gene Expression Assays (Life Tech-nologies) were used to examine the expression levels ofselected candidate genes (CEACAM5, SLC2A1) in normalbasal cell and premalignant lesions. b2-microglobulin(B2M) was used as an endogenous control. Statistical anal-ysis was performed using the sign test.
Immunofluorescent stainingFFPE tissues were sectioned at a 5-mm thickness and
stained as previously described (11). Antibodies used are:rabbit anti-KRT5,mouse anti-KRT5,mouse anti-PTBP3, andrabbit anti-c-myc (Abcam); mouse anti-CEACAM5 (Pro-Mab Biotechnologies Inc.); rabbit anti-SLC2A1 (AlphaDiagnostic International); anti-rabbit Cy3, anti-mouseAlexa Fluor 647, anti-mouse Cy3, and anti-rabbit AlexaFluor 647 (Jackson ImmunoResearch). Immunostainedtissues were visualized on an Axiocam system (Zeiss), andimages were taken using the Axiovision software.
ResultsStudy population, sample acquisition, and sequencealignmentFresh-frozen tissue blocks were obtained from 4 indivi-
duals with lung SCC (patients 1–4) at the time of tumorresection, and regions of normal basal cell, premalignant(squamous metaplastic and dysplastic) cells, and tumorcells were successfully captured from sectioned tissues bylaser microdissection (Supplementary Fig. S1). The demo-graphic information and clinical characteristics of thesepatients, as well as a description of the histology of eachmicrodissected premalignant region, are presented in Sup-plementary Table S1. Sequencing libraries of the expectedconcentration and cDNA size ranges were generated fromRNA isolated from the microdissected cells. All sequencedsamples produced reads with mean Phred quality scoresabove 25, indicating that it was possible to generatesequencing libraries of good quality with our method ofisolating RNA from laser-microdissected materials.A table of the number of reads that aligned uniquely
within each sample is shown in Supplementary Table S2.The fraction of reads aligning to the mitochondrial genomevaried considerably among samples. Inpatients 1 and2, thisfraction varied from 7% to 28%, but in patients 3 and 4,mitochondrial reads comprised from 22% to 65% ofuniquely aligned reads, with the highest fraction found inthe tumor samples from patient 4 (57%–65%). Because ofthis large amount of variability, reads aligning to the mito-chondrial genome were discarded from analysis after align-ment, and RPKM (reads/kb/millions of reads) values werecomputed relative to the total number of reads aligninguniquely to the nuclear genome.
Identification of genes associated with carcinogenesisTo identify SCC-associated geneswhose expression is also
associatedwithprogression fromnormal airwaybasal cell topremalignant(metaplasticordysplastic) lesions,amultistepprocedure was used as outlined in Fig. 1A. First, EnsemblGenes with zero aligned reads in all samples from at least 1patient were removed from analysis (to ensure that allpatients contributed evidence to each result), leaving20,817 genes for analysis. This list was then filtered toconsider only those genes with substantial evidence ofexpression(medianofgreaterthan50uniquelyalignedreadsacross all samples), leaving 7,025 genes for analysis. Usinglinear mixed-effects models and negative binomial general-ized linearmodels (see SupplementaryMethods for details),we then identified 626 early-stage genes (significantlydifferentially expressed in a similar manner in both prema-lignant lesionsand tumorcomparedwithnormalbasal cell),730 late-stage genes (significantly differentially expressedin a similar manner in tumor compared with both prema-lignant lesions and normal basal cell), and 68 "stepwise"genes (significantly differentially expressed in both of thedescribed stages of carcinogenesis; Fig. 1B; SupplementaryTable S3).
Experimental and computational validation ofcandidate genes
Three genes were selected for further validation: CEA-CAM5, SLC2A1, and PTBP3. These genes, whose expressionwas upregulated in premalignant lesions and tumor cellscompared with normal basal cell, were chosen because oftheir potential roles in the biology of lung carcinogenesis.The expression of CEACAM5 and SLC2A1was measured byperforming qRT-PCR on remaining material from thesequencing libraries of patients 3 and 4, as well as onlaser-microdissected RNA from four additional indepen-dent cases (patients 5–8). In each case, the mRNA level ofeach gene was significantly higher (sign test P < 0.05) in thepremalignant lesion than in normal basal cell (Fig. 2A).
BecausemRNA and protein levels may not always be wellcorrelated (16–18), immunofluorescent staining was per-formed in sections of normal epithelium, premalignantlesion, and carcinoma from 2 independent cases (patients9 and 10). CEACAM5 and SLC2A1 were not detectable inthe normal epithelia, but they were highly expressed in cellswithin both metaplastic lesions and the SCC tumors (Fig.2B and C). SLC2A1 was expressed throughout the KRT5þ
component of the tumor, whereasCEACAM5was expressedin some, but not all, KRT5þ tumor cells. PTBP3was stronglyexpressed in premalignant lesions and tumor cells, andalthough it was strongly expressed in columnar KRT5� cellsof normal airway epithelium, its expression was undetect-able in normal basal cell (Supplementary Fig. S2).
Tobetterunderstandthebiologicrole that thesegenesmayplay in the development of lung SCC, the significance of thedifferential expression of each gene was assessed in severalGEO datasets with respect to experimental parameters rele-vant to lung SCC carcinogenesis. First, SLC2A1 and PTBP3were confirmed to be significantly upregulated (SLC2A1,
Identifying Pathways in Lung Carcinogenesis
www.aacrjournals.org Cancer Prev Res; 7(5) May 2014 489
Research. on May 27, 2018. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2014; DOI: 10.1158/1940-6207.CAPR-13-0372

P¼ 0.004; PTBP3, P¼ 0.017) in an independent set of SCCtumors (n ¼ 5) with respect to paired samples of adjacentnormal tissue (GEOdatasetGDS1312; ref. 19);however, theexpression ofCEACAM5was unchanged (P¼ 0.64). Next, acollection of SCC (n¼ 18) and adenocarcinoma (ADC; n¼40) lung tumors (GDS3627; refs. 20, 21)was interrogated todeterminethespecificityof theexpressionof thesegeneswithrespect to theSCCtumortype.TheexpressionofSLC2A1andPTBP3 was again strongly increased in SCC tumors com-pared with ADC tumors (SLC2A1, P ¼ 1.1 � 10�7; PTBP3,P ¼ 0.0004); however, CEACAM5 was moderately down-regulated inSCCrelative toADC(P¼0.08). Finally, becausepremalignant lesions in large central airways are believed toarise frominjury causedbycigarette smoking, theexpressionlevels of these genes were examined in a study of broncho-scopic brushings of healthy current (n ¼ 34), former(n ¼ 18), and never (n ¼ 23) smokers (GDS534; ref. 21).In this study, CEACAM5 and SLC2A1 were significantlyupregulated in brushings from current smokers comparedwith those from never smokers (CEACAM5, P ¼ 0.0001;SLC2A1, P ¼ 0.016), although PTBP3 was not (P ¼ 0.66).
Prediction of chromosomal gains and losses duringcarcinogenesis
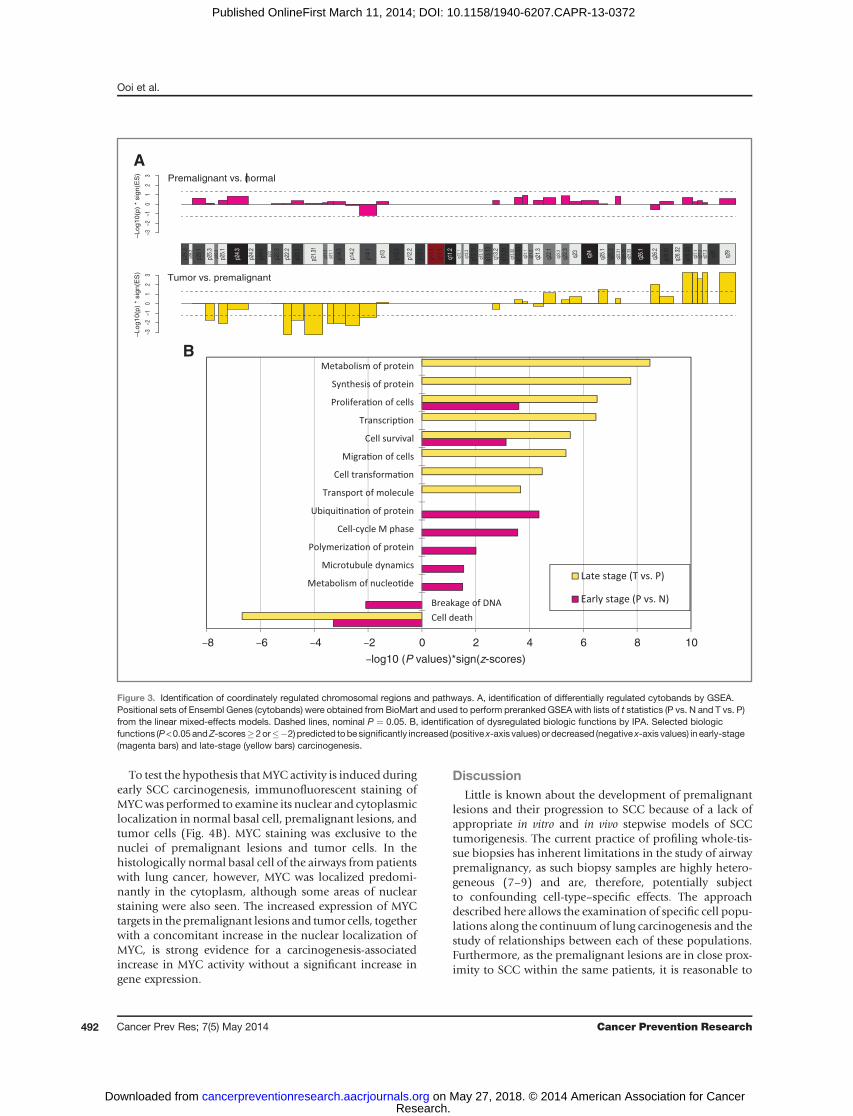
GSEA performed using positionally defined gene sets(cytobands) revealed that late-stage (but not early-stage)carcinogenesis is associated with a coordinate loss ofexpression in the p arm of chromosome 3 and an atten-dant gain of expression in 3q26.33-3q29 (Fig. 3A), whichcorresponds to previously reported observations of fre-quent 3p deletion and 3q amplification in squamoustumors (22, 23). In particular, chromosomal band3q26.33 has been reported to be consistently amplifiedin lung SCC (24).
Identification of biologic changes in early- and late-stage carcinogenesis
Ingenuity pathway analysis (IPA; Ingenuity Systems) wasused to further characterize the changes in biologic func-tions resulting from the differential expression of genesassociated with early-stage events, which contribute to theinitiation and formation of premalignant lesions, or withlate-stage events, which are involved in the progression
55,841 Ensembl genes
A B
≥ 1 Read in each subject
Median ≥ 50 reads across
all samples
Linear mixed-effects model
(7√RPKM) ~ Group ⎢Patient
Negative binomial model
Reads ~ ∑reads
+ patient + group
7,025
7,024
1,000Sign(P/N)
= sign(T/N)
Sign(P/N)
= sign(T/N)Sign(T/P) =
sign(T/N)
Sign(T/P) =
sign(T/N)
956999
626Overlap
730Overlap
662
Late
68
Stepwise
558
Early
1,143 1,836
1,178 1,859956
3,400 3,370
T vs. NP < 0.05
T vs. PP < 0.05
P vs. NP < 0.05
P vs. NP < 0.05
T vs. NP < 0.05
T vs. PP < 0.05
6,292
Numericallystable
Numericallystable
Ea
rly
Ste
pw
ise
La
te
Figure 1. Identification of genes associated with early- or late-stage SCC carcinogenesis. A, analysis flowchart. Uniquely aligned reads were assignedto 55,841 Ensembl Gene loci (Ensembl build 69). Two statistical models were then applied to identify genes with significant (P < 0.05) differential expressionbetween tumor and normal cells, as well as between premalignant and normal cells ("early" genes), between tumor and premalignant cells ("late"genes), or both ("stepwise" genes). B, expression heatmap. Root-transformed RPKM values were scaled to a mean of 0 and SD of one within each patient;red and blue indicate genes with expression that is higher or lower than the mean within each patient, respectively. Genes are hierarchically clustered withineach group (early, stepwise, and late).
Ooi et al.
Cancer Prev Res; 7(5) May 2014 Cancer Prevention Research490
Research. on May 27, 2018. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2014; DOI: 10.1158/1940-6207.CAPR-13-0372

from premalignant lesions to tumor. This analysis revealedthat the early-stage carcinogenesis was characterizeduniquely by increased protein ubiquitination and cell-cycleprogression, whereas the late-stage events were markedprimarily by increased transcriptional and translationalactivity and cellular migration and transformation (Fig.3B; Supplementary Table S4). In addition, an increase incell survival and proliferation and a corresponding down-regulation of cell deathmechanisms was observed through-out both stages of carcinogenesis. IPA was also used todetermine whether the genes identified to be differentiallyexpressed either early or late in carcinogenesis are enrichedin known targets of various transcription factors. Thisapproach revealed that the set of genes that is differentially
expressed early in carcinogenesis and remains dysregulatedin tumor cells is enriched in previously reported targets ofMYC and TP53 (Fig. 4A; Supplementary Tables S5 and S6).As MYC and TP53 are predicted to activate or repress theexpression of these targets, respectively, this suggests thatMYC activity is significantly induced (P ¼ 2.41 � 10�5,Z-score ¼ 3.789) and TP53 activity is potentially repressed(P ¼ 9.30 � 10�8, Z-score ¼ �1.034) during early carci-nogenesis, and that their activity remains altered through-out tumorigenesis. Importantly, the gene expression levelsof TP53 and MYC did not change significantly with respectto the pathologic continuum from normal to tumor, sug-gesting that the predicted changes in their activity are due toposttranscriptional regulation.
Lo
g2 (
fold
ch
an
ge
)
CEACAM5Basal
cells
CEACAM5Premalignant
lesions
SLC2A1Premalignant
lesions
SLC2A1Basal
cells
Sign test P = 0.03 Sign test P = 0.03
05
10
1
52
0
Normal airway
Premalignantlesion
Squamous cellcarcinoma
KRT5DAPI
CEACAM5DAPI
MergedBA
Normal airway
lesionPremalignant
Squamous cellcarcinoma
MergedKRT5DAPI
SLC2A1DAPI
C
Figure 2. Experimental validation of CEACAM5 and SLC2A1 expression. A, qRT-PCR. Box plots represent RNA levels of CEACAM5 and SLC2A1 in normalbasal cell and premalignant lesions from 6 patients, showing increased expression of both genes in premalignant lesions compared with normal basal cell.B2M was used as the endogenous control. B and C, immunofluorescent staining of CEACAM5 and SLC2A1. Protein staining shows increased expressionof CEACAM5 and SLC2A1 in premalignant lesions and SCC compared with basal cell in the normal epithelium. Top rows, normal airway epithelium;middle rows, premalignant lesions; bottom rows, SCC. Left columns, KRT5, stained in red as marker for basal cell, premalignant lesions, and tumorcells; middle columns, SLC2A1 or CEACAM5, stained in green; right columns, merged images of left and middle columns. DAPI (40,6-diamidino-2-phenylindole), stained in blue, as nuclear marker. White scale bars, 50 mm. Insets show close-up views of the boxed regions.
Identifying Pathways in Lung Carcinogenesis
www.aacrjournals.org Cancer Prev Res; 7(5) May 2014 491
Research. on May 27, 2018. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2014; DOI: 10.1158/1940-6207.CAPR-13-0372
To test the hypothesis thatMYC activity is induced duringearly SCC carcinogenesis, immunofluorescent staining ofMYCwas performed to examine its nuclear and cytoplasmiclocalization in normal basal cell, premalignant lesions, andtumor cells (Fig. 4B). MYC staining was exclusive to thenuclei of premalignant lesions and tumor cells. In thehistologically normal basal cell of the airways from patientswith lung cancer, however, MYC was localized predomi-nantly in the cytoplasm, although some areas of nuclearstaining were also seen. The increased expression of MYCtargets in the premalignant lesions and tumor cells, togetherwith a concomitant increase in the nuclear localization ofMYC, is strong evidence for a carcinogenesis-associatedincrease in MYC activity without a significant increase ingene expression.
DiscussionLittle is known about the development of premalignant
lesions and their progression to SCC because of a lack ofappropriate in vitro and in vivo stepwise models of SCCtumorigenesis. The current practice of profiling whole-tis-sue biopsies has inherent limitations in the study of airwaypremalignancy, as such biopsy samples are highly hetero-geneous (7–9) and are, therefore, potentially subjectto confounding cell-type–specific effects. The approachdescribed here allows the examination of specific cell popu-lations along the continuum of lung carcinogenesis and thestudy of relationships between each of these populations.Furthermore, as the premalignant lesions are in close prox-imity to SCC within the same patients, it is reasonable to
–8 –6 –4 –2 1086420
Metabolism of nucleo�de
Microtubule dynamics
Polymeriza�on of protein
Cell-cycle M phase
Ubiqui�na�on of protein
Transport of molecule
Cell transforma�on
Migra�on of cells
Cell survival
Transcrip�on
Prolifera�on of cells
Synthesis of protein
Metabolism of protein
–log10 (P values)*sign(z-scores)
Late stage (T vs. P)
Early stage (P vs. N)Breakage of DNACell death
A
B
vs. n
vs. p
–L
–L
Figure 3. Identification of coordinately regulated chromosomal regions and pathways. A, identification of differentially regulated cytobands by GSEA.Positional sets of Ensembl Genes (cytobands) were obtained from BioMart and used to perform preranked GSEA with lists of t statistics (P vs. N and T vs. P)from the linear mixed-effects models. Dashed lines, nominal P ¼ 0.05. B, identification of dysregulated biologic functions by IPA. Selected biologicfunctions (P<0.05andZ-scores�2or��2) predicted tobesignificantly increased (positive x-axis values) or decreased (negative x-axis values) in early-stage(magenta bars) and late-stage (yellow bars) carcinogenesis.
Ooi et al.
Cancer Prev Res; 7(5) May 2014 Cancer Prevention Research492
Research. on May 27, 2018. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2014; DOI: 10.1158/1940-6207.CAPR-13-0372

expect that alterations in gene expression shared betweenpremalignant and tumor cells reflectmolecular changes thatoccur during carcinogenesis.We focused specifically on the expression patterns of
three genes, CEACAM5, SLC2A1, and PTBP3, that areupregulated in the premalignant lesions (and, in the caseof SLC2A1, further upregulated in tumor cells). CEA-CAM5, a cell surface glycoprotein that plays a role in celladhesion and intracellular signaling, has been shown tobe important in other epithelial cell cancers, such ascolon cancer (25). SLC2A1 (also known as glucose trans-porter 1, or GLUT1) is a facilitative glucose transporterassociated with hepatocellular cancer and head and neckSCC (26, 27). PTBP3 (also known as regulator of differ-entiation 1, or ROD1) is an RNA-binding protein thatregulates pre-mRNA alternative splicing and plays a rolein the regulation of cell proliferation and differentiation(28, 29). The protein level expression of each gene wassubstantially increased in premalignant lesions andtumor cells, although the expression of CEACAM5 withinthe tumor cells was more heterogeneous than that of theother genes. In addition, although PTBP3 was stronglyexpressed in normal airway epithelium, its expression wasrestricted to columnar KRT5� cells.We also examined the expression of these genes in pub-
licly available microarray datasets related to SCC carcino-
genesis. In one such experiment, the genes SLC2A1 andPTBP3 were significantly upregulated in lung SCC tumorsrelative to matched adjacent normal tissue, but unexpect-edly, CEACAM5 was not. However, that study profiledtumor biopsies, which often contain significant stromalcontamination; moreover, we observed substantial hetero-geneity of CEACAM5 immunostaining in SCC tumor tissuein this study. The identification of CEACAM5 as an early-stage marker of squamous lung carcinogenesis in this studymay, therefore, be attributable to the careful laser micro-dissection of SCC tumor cells from the surrounding stroma.
Because lung SCC is strongly associated with a history oftobacco smoking, we examined the relationship betweensmoking history and the expression of these genes in aprevious study of bronchoscopic brushings. In that study,CEACAM5 and SLC2A1 were significantly upregulated inbrushings from current smokers compared with those fromnever smokers. In a subsequent study from the same authors(30), the expression of CEACAM5 was reported to beirreversibly altered in former smokers for up to severaldecades after smoking cessation, suggesting that a smok-ing-associated increase in CEACAM5 expression in histo-logically normal airway epithelium may be an early eventassociated with carcinogenesis in these individuals.
WeusedGSEA to identify chromosomal regions thatwereenriched in differentially expressed genes, which suggested
Downregulated Upregulated
Predicted inhibited
Predicted activated
Activates expression
Inhibits expression
Early carcinogenesis
A
Normal airway
Premalignant lesion
KRT5
DAPI
MYCDAPI
MERGED
Squamous cellcarcinoma
B
Figure 4. Identification of MYC as a dysregulated transcription factor in early carcinogenesis. A, identification of dysregulated transcription factors byIPA. Expression patterns of known downstream targets of TP53 and MYC suggest the inhibition of TP53 and the activation of MYC during earlycarcinogenesis. B, experimental validation of MYC activation in premalignant lesions and SCC. Immunofluorescent staining shows detection of MYCexpression in the nuclei of premalignant lesions and SCC compared with cytoplasmic localization in basal cell. Top rows, normal airway epithelium; middlerows, premalignant lesion; bottom rows, SCC. Left columns, KRT5, stained in green as marker for basal cell, premalignant lesion, and tumor cells;middle columns, MYC, stained in red; right columns, merged images of left and middle columns. DAPI, stained in blue, as nuclear marker in all images. Whitescale bars, 50 mm. Insets show close-up views of the boxed regions.
Identifying Pathways in Lung Carcinogenesis
www.aacrjournals.org Cancer Prev Res; 7(5) May 2014 493
Research. on May 27, 2018. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2014; DOI: 10.1158/1940-6207.CAPR-13-0372

that the frequent 3p loss and 3q amplification that arecharacteristic of SCC (and rare in ADC; refs. 24, 31, 32) arelate-stage events in SCC carcinogenesis. A relevant work byvan Boerdonk and colleagues presented a longitudinalstudy of 6 patients with squamous metaplastic lesions thatshowed carcinoma in situ or carcinoma at follow-up bron-choscopy (33). These lesions showed 3p loss and 3q gainwhen compared with 23 lesions from subjects with no signof cancer in their follow-up bronchoscopy. The potentialdiscordant result about the timing of this genomic ampli-fication of chromosome 3 (early event vs. late event in lungcarcinogenesis)maybe a result of the different experimentaldesigns of both studies. The van Boerdonk study foundchromosomal changes in squamous metaplastic lesions ofsubjects that had follow-up carcinomas when comparedwith lesions from subjects with no follow-up carcinomas,but they did not compare the premalignant lesions tonormal airway epithelium or SCC from these same indivi-duals to establish the potential stepwise chronology of thismolecular event. In our study, we compared premalignantlesions with matched normal basal cells (early-stage) andmatched carcinomas (late-stage), all within individualpatients with SCC. Our data show chromosome 3 abnor-malities during late-stage carcinogenesis, suggesting thepossibility that the observed 3p loss and 3q gain couldhave happened at any time point during the progressionfrom squamousmetaplasia to carcinomas.We alsoused IPAto identify biologic functions and regulators that wereoverrepresented among the genes associated with early- orlate-stage carcinogenesis. This analysis revealed that early-stage carcinogenesis is marked primarily by increased fluxthrough the cell cycle, but that cellular proliferation con-tinues throughout late-stage carcinogenesis.
Finally, we used IPA to make predictions about theupstream regulators that might be responsible for thesechanges, and identified the transcription factors TP53 andMYC as likely candidates based on the coordinate differ-ential expression of their target genes. Nuclear expression ofMYC in premalignant lesions and tumor cells, suggestingthat activation ofMYC by nuclear translocation could be animportant event contributing to dysregulated cell-cycleprogression during SCC carcinogenesis. Previous reportshave also identified the potential importance of MYC inpremalignant lesions of lung carcinoma (31) and breastcancer (34). Further analysis of other datasets is needed tovalidate the results.
Although this study represents a novel approach foridentifying driver molecular events associated with squa-mous cell lung carcinogenesis, there are a number ofimportant limitations to the work. Ourmodel assumes thatthere is a molecular relationship between the premalignantand tumor cells foundwithin the airwayof the samepatient,although the lesions may develop from disparate clonalpopulations, thereby limiting the interpretation of thosechanges as reflecting a stepwise change between lesions.Longitudinal studies of premalignant lesions resampledover time are needed to identify molecular alterationsassociated with progression or regression within a clonal
populationof cells. Furthermore, our group andothers havepreviously reported molecular alterations throughout thehistologically normal airway of smokers with lung cancer(35). Those molecular events in the histologically normal"field of injury" may reflect some of the earliest events incarcinogenesis and will not be captured directly by ourapproach. Finally, our study did not evaluate the potentialrole of stromal cells in initiation and progression of SCC.
In summary, we present a novel approach to identify themolecular alterations that characterize premalignantlesions and carcinogenesis in lung SCC. By isolating andtranscriptome profiling a progenitor cell population withinnormal airway epithelium, premalignant lesions and SCCfrom the same individual, we were able to provide uniqueinsight into stepwise molecular alterations that occur dur-ing lung carcinogenesis. Our analysis identified coordinatechanges in the activity of upstream regulators and theexpression of downstream genes within the same patientduring early- and late-stage carcinogenesis. Further studiesthat profile molecular alterations within an individualpremalignant lesion followed serially over time (as it pro-gresses or regresses) will provide further resolution to themolecular events associatedwith lung carcinogenesis. Addi-tional work will also be necessary to determine whether anyof the genes identified in our study can be used to distin-guish premalignant lesions that will progress to cancer fromthose that will regress. Genes identified and validated in thismanner might serve as early biomarkers for SCC detectionand targets for SCC chemoprevention.
Disclosure of Potential Conflicts of InterestM.E. Lenburg has received a commercial research grant from Johnson and
Johnson, has ownership interest (including patents) and is a consultant/advisory board member of Allegro Diagnostics. A.E. Spira has ownershipinterest (including patents) and is a consultant/advisory board member ofAllegro Diagnostics. No potential conflicts of interest were disclosed by theother authors.
Authors' ContributionsConception and design: A.T. Ooi, D.A. Elashoff, T.C. Walser, S.M. Dubi-nett, A. Spira, B.N. GompertsDevelopment of methodology: A.T. Ooi, A.C. Gower, B.N. GompertsAcquisition of data (acquired and managed patients, providedfacilities, etc.): A.T. Ooi, J.L. Vick, B. Nagao, T.C. Walser, A. SpiraAnalysis and interpretation of data (e.g., statistical analysis, bio-statistics, computational analysis): A.T. Ooi, A.C. Gower, K.X. Zhang,W.D. Wallace, D.A. Elashoff, M. Pellegrini, M.E. Lenburg, B.N. GompertsWriting, review, and/or revision of the manuscript: A.T. Ooi,A.C. Gower, W.D. Wallace, D.A. Elashoff, M. Pellegrini, M.E. Lenburg,B.N. GompertsAdministrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases): A.T. Ooi, A.C. Gower, L. HongStudy supervision: A.T. Ooi, M.E. Lenburg, A. Spira, B.N. Gomperts
AcknowledgmentsThe authors thank Gang Liu and Lingqi Luo for performing the RNA-seq
of patients 1, 2 and 4, UCLA Broad Stem Cell Research Center High-Throughput Sequencing Core Resource for performing the RNA-seq forpatient 3, as well as Frank Schembri for technical assistance in optimizingRNA extraction and Josh Campbell and Nacho Caballero for assistance withnegative binomial analysis. Histologic services were performed by the UCLATranslational Pathology Core Laboratory (TPCL) and laser microdissectionwas performed through the TPCL and the Advanced Light Microscopy Core.The statistical analysis for qRT-PCR was performed by Tristan Grogan andsupported by the NIH/National Center for Advancing Translational Science(NCATS) UCLA CTSI grant number UL1TR000124.
Ooi et al.
Cancer Prev Res; 7(5) May 2014 Cancer Prevention Research494
Research. on May 27, 2018. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2014; DOI: 10.1158/1940-6207.CAPR-13-0372

Grant SupportThis work was supported by the Department of Defense (DOD) CTRA
LC090615 (to S.M. Dubinett, A. Spira, B.N. Gomperts, and T.C. Walser),Tobacco-Related Disease Research Program (TRDRP; 19FT- 0046; to A.T.Ooi), NationalHeart, Lung, andBlood Institute (NHLBI) R01HL094561-01,CIRM RN2-009-04 (to B.N. Gomperts), the National Cancer Institute (NCI;#U01-CA152751; to S.M. Dubinett, A. Spira, B.N. Gomperts, and T.C.Walser), the Department of Defense (DOD; #W81XWH-10-1-1006; toS.M. Dubinett, A. Spira, and T.C. Walser), Department of Veteran Affairs
(VA; #5I01BX000359; to S.M. Dubinett and T.C. Walser), and TRDRP(#20KT-0055; to T.C. Walser).
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received October 23, 2013; revised February 24, 2014; accepted February25, 2014; published OnlineFirst March 11, 2014.
References1. Siegel R, Naishadham D, Jemal A. Cancer statistics, 2012. CA Cancer
J Clin 2012;62:10–29.2. Stat bite: mortality from lung and bronchus cancer by race/ethnicity,
1998–2002. J Natl Cancer Inst 2006;98:158.3. Ettinger DS, Akerley W, Borghaei H, Chang AC, Cheney RT, Chirieac
LR, et al. Non-small cell lung cancer. J Natl Compr Canc Netw2012;10:1236–71.
4. Colby TV, Wistuba II, Gazdar A. Precursors to pulmonary neoplasia.Adv Anat Pathol 1998;5:205–15.
5. Kerr KM. Pulmonary preinvasive neoplasia. J Clin Pathol 2001;54:257–71.
6. Peebles KA, Lee JM, Mao JT, Hazra S, Reckamp KL, Krysan K, et al.Inflammation and lung carcinogenesis: applying findings in preventionand treatment. Expert Rev Anticancer Ther 2007;7:1405–21.
7. Kettunen E, Anttila S, Seppanen JK, Karjalainen A, Edgren H, Lind-strom I, et al. Differentially expressed genes in nonsmall cell lungcancer: expression profiling of cancer-related genes in squamous celllung cancer. Cancer Genet Cytogenet 2004;149:98–106.
8. Seo JS, Ju YS, Lee WC, Shin JY, Lee JK, Bleazard T, et al. Thetranscriptional landscape and mutational profile of lung adenocarci-noma. Genome Res 2012;22:2109–19.
9. Xi L, Feber A, Gupta V, Wu M, Bergemann AD, Landreneau RJ, et al.Whole genome exon arrays identify differential expression of alterna-tively spliced, cancer-related genes in lung cancer. Nucleic Acids Res2008;36:6535–47.
10. Hong KU, Reynolds SD,Watkins S, Fuchs E, Stripp BR. Basal cells areamultipotent progenitor capable of renewing the bronchial epithelium.Am J Pathol 2004;164:577–88.
11. Ooi AT, Mah V, Nickerson DW, Gilbert JL, Ha VL, Hegab AE, et al.Presence of a putative tumor-initiating progenitor cell populationpredicts poor prognosis in smokers with non–small cell lung cancer.Cancer Res 2010;70:6639–48.
12. Langmead B, Trapnell C, Pop M, Salzberg SL. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome.Genome Biol 2009;10:R25.
13. Quinlan AR, Hall IM. BEDTools: a flexible suite of utilities for comparinggenomic features. Bioinformatics 2010;26:841–2.
14. Mortazavi A,WilliamsBA,McCueK, Schaeffer L,WoldB.Mapping andquantifying mammalian transcriptomes by RNA-Seq. Nat Methods2008;5:621–8.
15. SubramanianA, TamayoP,Mootha VK,Mukherjee S, Ebert BL,GilletteMA, et al. Gene set enrichment analysis: a knowledge-based approachfor interpreting genome-wide expression profiles. Proc Natl Acad SciU S A 2005;102:15545–50.
16. Greenbaum D, Colangelo C, Williams K, Gerstein M. Comparingprotein abundance and mRNA expression levels on a genomic scale.Genome Biol 2003;4:117.
17. Taniguchi Y, Choi PJ, Li GW, Chen H, Babu M, Hearn J, et al.Quantifying E. coli proteome and transcriptome with single-moleculesensitivity in single cells. Science 2010;329:533–8.
18. Gygi SP, Rochon Y, Franza BR, Aebersold R. Correlation betweenprotein andmRNAabundance in yeast.MolCell Biol 1999;19:1720–30.
19. Wachi S, YonedaK,WuR. Interactome–transcriptomeanalysis revealsthe high centrality of genes differentially expressed in lung cancertissues. Bioinformatics 2005;21:4205–8.
20. Kuner R, Muley T, Meister M, Ruschhaupt M, Buness A, Xu EC, et al.Global gene expression analysis reveals specific patterns of celljunctions in non-small cell lung cancer subtypes. Lung Cancer2009;63:32–8.
21. Spira A, Beane J, Shah V, Liu G, Schembri F, Yang X, et al. Effects ofcigarette smoke on the human airway epithelial cell transcriptome.Proc Natl Acad Sci U S A 2004;101:10143–8.
22. Brunelli M, Bria E, Nottegar A, Cingarlini S, Simionato F, Calio A, et al.True 3q chromosomal amplification in squamous cell lung carcinomaby FISH and aCGH molecular analysis: impact on targeted drugs.PLoS ONE 2012;7:e49689.
23. Partridge M, Kiguwa S, Langdon JD. Frequent deletion of chromo-some 3p in oral squamous cell carcinoma. Eur J Cancer B Oral Oncol1994;30B:248–51.
24. Bass AJ, Watanabe H, Mermel CH, Yu S, Perner S, Verhaak RG, et al.SOX2 is an amplified lineage-survival oncogene in lung and esoph-ageal squamous cell carcinomas. Nat Genet 2009;41:1238–42.
25. Pignatelli M, Durbin H, Bodmer WF. Carcinoembryonic antigen func-tions as an accessory adhesion molecule mediating colon epithelialcell-collagen interactions. Proc Natl Acad Sci U S A 1990;87:1541–5.
26. Amann T, Maegdefrau U, Hartmann A, Agaimy A, Marienhagen J,Weiss TS, et al. GLUT1 expression is increased in hepatocellularcarcinoma and promotes tumorigenesis. Am J Pathol 2009;174:1544–52.
27. Heikkinen PT, Nummela M, Jokilehto T, Grenman R, Kahari VM,Jaakkola PM.Hypoxic conversion of SMAD7 function from an inhibitorinto a promoter of cell invasion. Cancer Res 2010;70:5984–93.
28. Sadvakassova G, DobocanMC, DifalcoMR, Congote LF. Regulator ofdifferentiation 1 (ROD1) binds to the amphipathic C-terminal peptide ofthrombospondin-4 and is involved in its mitogenic activity. J CellPhysiol 2009;220:672–9.
29. Yamamoto H, Tsukahara K, Kanaoka Y, Jinno S, OkayamaH. Isolationof a mammalian homologue of a fission yeast differentiation regulator.Mol Cell Biol 1999;19:3829–41.
30. Beane J, Sebastiani P, Liu G, Brody JS, Lenburg ME, Spira A. Revers-ible and permanent effects of tobacco smoke exposure on airwayepithelial gene expression. Genome Biol 2007;8:R201.
31. Massion PP, Zou Y, Uner H, Kiatsimkul P, Wolf HJ, Baron AE, et al.Recurrent genomic gains in preinvasive lesions as a biomarker of riskfor lung cancer. PLoS ONE 2009;4:e5611.
32. Massion PP, KuoWL, Stokoe D, Olshen AB, Treseler PA, Chin K, et al.Genomic copy number analysis of non–small cell lung cancer usingarray comparative genomic hybridization: implications of the phos-phatidylinositol 3-kinase pathway. Cancer Res 2002;62:3636–40.
33. van Boerdonk RA, Sutedja TG, Snijders PJ, Reinen E, Wilting SM, vande Wiel MA, et al. DNA copy number alterations in endobronchialsquamous metaplastic lesions predict lung cancer. Am J Respir CritCare Med 2011;184:948–56.
34. Ling H, Sylvestre JR, Jolicoeur P. Notch1-induced mammary tumordevelopment is cyclin D1-dependent and correlates with expansion ofpremalignant multipotent duct-limited progenitors. Oncogene 2010;29:4543–54.
35. Spira A, Beane JE, Shah V, Steiling K, Liu G, Schembri F, et al. Airwayepithelial gene expression in the diagnostic evaluation of smokerswithsuspect lung cancer. Nat Med 2007;13:361–6.
www.aacrjournals.org Cancer Prev Res; 7(5) May 2014 495
Identifying Pathways in Lung Carcinogenesis
Research. on May 27, 2018. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2014; DOI: 10.1158/1940-6207.CAPR-13-0372

2014;7:487-495. Published OnlineFirst March 11, 2014.Cancer Prev Res Aik T. Ooi, Adam C. Gower, Kelvin X. Zhang, et al. CarcinogenesisCell Carcinomas Identifies Mechanisms Involved in Stepwise Molecular Profiling of Premalignant Lesions in Lung Squamous
Updated version
10.1158/1940-6207.CAPR-13-0372doi:
Access the most recent version of this article at:
Material
Supplementary
1
http://cancerpreventionresearch.aacrjournals.org/content/suppl/2014/03/12/1940-6207.CAPR-13-0372.DCAccess the most recent supplemental material at:
Cited articles
http://cancerpreventionresearch.aacrjournals.org/content/7/5/487.full#ref-list-1
This article cites 35 articles, 12 of which you can access for free at:
Citing articles
http://cancerpreventionresearch.aacrjournals.org/content/7/5/487.full#related-urls
This article has been cited by 9 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerpreventionresearch.aacrjournals.org/content/7/5/487To request permission to re-use all or part of this article, use this link
Research. on May 27, 2018. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst March 11, 2014; DOI: 10.1158/1940-6207.CAPR-13-0372





























