Embed Size (px)
Citation preview

MOLECULARPHYLOGENETICSAND
Molecular Phylogenetics and Evolution 32 (2004) 375–382
EVOLUTION
www.elsevier.com/locate/ympev
Molecular phylogeny of the antitropical genusPseudolabrus (Perciformes: Labridae): evidence
for a Southern Hemisphere origin
Kohji Mabuchi,a,* Tetsuji Nakabo,b and Mutsumi Nishidaa
a Ocean Research Institute, University of Tokyo, 1-15-1 Minamidai, Nakano, Tokyo 164-8639, Japanb The Kyoto University Museum, Kyoto University, Yoshida, Sakyo, Kyoto 606-8501, Japan
Received 1 October 2003; revised 6 January 2004
Available online 1 March 2004
Abstract
The genus Pseudolabrus comprises 11 species of marine nearshore fishes which are antitropically distributed: two species occur in
East Asia, the remaining nine species being distributed in the Southern Hemisphere, mainly in the temperate Pacific. The distri-
butions of their closely allied genera, collectively called ‘‘pseudolabrines’’ are, however, restricted to the Australia-New Zealand
region. The molecular phylogeny of six of the 11 Pseudolabrus species from both Hemispheres and four of the five other pseudo-
labrine genera was reconstructed from nucleotide sequence data from mitochondrial DNA 12S rRNA, tRNAVal, and 16S rRNA
genes. Both parsimony and Bayesian analyses were performed. Results are not consistent with a previous phylogenetic hypothesis
based on osteological data, particularly in the relationship between Pseudolabrus and Notolabrus, indicating a probable need for
reviewing the status of Notolabrus (or the delimitations of both Pseudolabrus and Notolabrus). The two Northern Hemisphere
species of Pseudolabrus were monophyletic and nested deep into the clade of the Southern Hemisphere pseudolabrines, which in-
dicates that both pseudolabrines and Pseudolabrus originated in the Southern Hemisphere. A dispersal rather than vicariance ex-
planation for the antitropical distribution of Pseudolabrus is more parsimonious given the number of dispersal events, extinctions,
and evolutionary adaptations required under the phylogeny. Based on molecular clock calibrations, the transequatorial divergence
was suggested to be early to mid Pliocene at the earliest.
� 2004 Elsevier Inc. All rights reserved.
Keywords: Antitropical distribution; Labridae; Pseudolabrus; Notolabrus; Mitochondrial DNA
1. Introduction
The occurrence of the same taxa to north and south
of the tropics but not the intervening region is one of themost interesting phenomena of biogeography, and such
disjunct distributions are observed for a variety of taxa,
both terrestrial and aquatic (Briggs, 1995; Randall,
1981). There are many variants to this kind of distri-
bution, especially related to latitude: polar, boreal, cool
temperature, warm temperature, and subtropical. For
all these patterns collectively, Hubbs (1952) proposed
the encompassing term ‘‘antitropical,’’ and the term hascome into general use (Randall, 1981). As seen in the
* Corresponding author. Fax: +81-3-5351-6822.
E-mail address: [email protected] (K. Mabuchi).
1055-7903/$ - see front matter � 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.ympev.2004.01.008
fact that Darwin (1859) already mentioned it in his ‘‘On
the origin of species,’’ these disjunct distribution patterns
have long-attracted biologists as interesting problems to
resolve.Several hypotheses have been proposed for antitro-
pical distributions of marine taxa (Burridge, 2002).
Species intolerant of tropical temperatures may have
dispersed across equatorial waters at shallow depths
during periods of cooler climate, such as glaciations
(Ekman, 1953; Lindberg, 1991), or by descent into
cooler waters at tropical latitudes (Ekman, 1953; Hubbs,
1952). Dispersal may have also been facilitated overlonger time scales via island integration (Rotondo et al.,
1981; Springer, 1982). Alternatively, vicariance mecha-
nisms may have isolated Northern and Southern
Hemisphere taxa by extirpation of intervening tropical
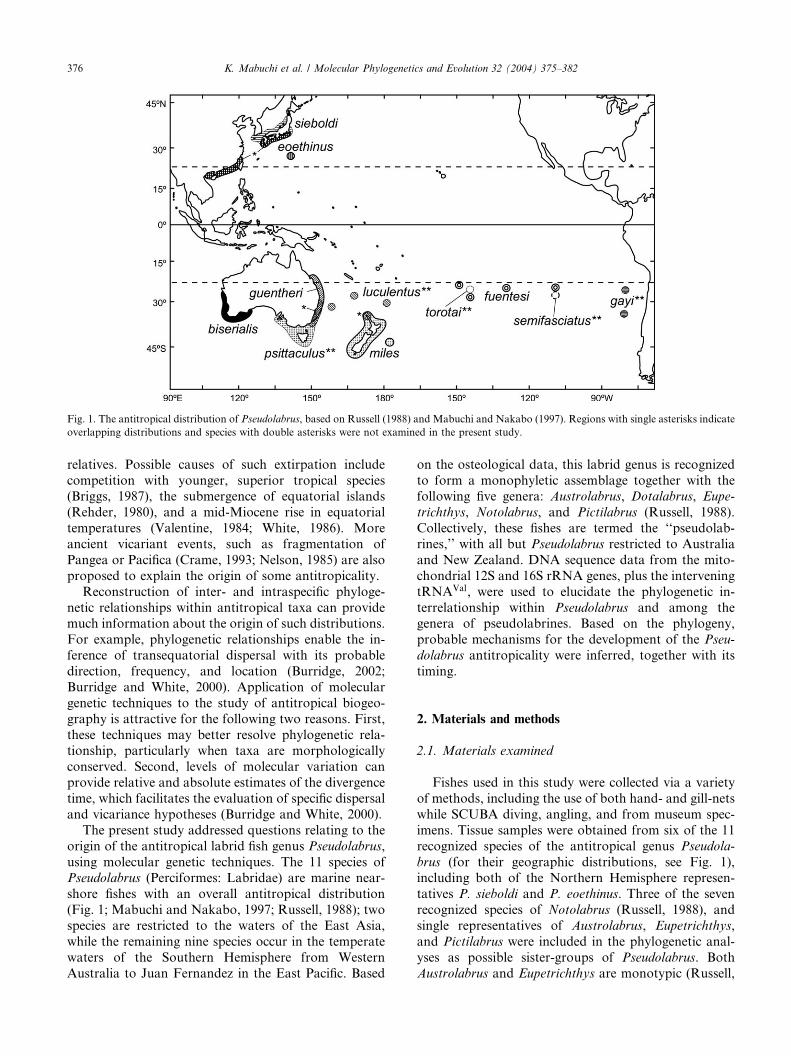
Fig. 1. The antitropical distribution of Pseudolabrus, based on Russell (1988) and Mabuchi and Nakabo (1997). Regions with single asterisks indicate
overlapping distributions and species with double asterisks were not examined in the present study.
376 K. Mabuchi et al. / Molecular Phylogenetics and Evolution 32 (2004) 375–382
relatives. Possible causes of such extirpation include
competition with younger, superior tropical species(Briggs, 1987), the submergence of equatorial islands
(Rehder, 1980), and a mid-Miocene rise in equatorial
temperatures (Valentine, 1984; White, 1986). More
ancient vicariant events, such as fragmentation of
Pangea or Pacifica (Crame, 1993; Nelson, 1985) are also
proposed to explain the origin of some antitropicality.
Reconstruction of inter- and intraspecific phyloge-
netic relationships within antitropical taxa can providemuch information about the origin of such distributions.
For example, phylogenetic relationships enable the in-
ference of transequatorial dispersal with its probable
direction, frequency, and location (Burridge, 2002;
Burridge and White, 2000). Application of molecular
genetic techniques to the study of antitropical biogeo-
graphy is attractive for the following two reasons. First,
these techniques may better resolve phylogenetic rela-tionship, particularly when taxa are morphologically
conserved. Second, levels of molecular variation can
provide relative and absolute estimates of the divergence
time, which facilitates the evaluation of specific dispersal
and vicariance hypotheses (Burridge and White, 2000).
The present study addressed questions relating to the
origin of the antitropical labrid fish genus Pseudolabrus,
using molecular genetic techniques. The 11 species ofPseudolabrus (Perciformes: Labridae) are marine near-
shore fishes with an overall antitropical distribution
(Fig. 1; Mabuchi and Nakabo, 1997; Russell, 1988); two
species are restricted to the waters of the East Asia,
while the remaining nine species occur in the temperate
waters of the Southern Hemisphere from Western
Australia to Juan Fernandez in the East Pacific. Based
on the osteological data, this labrid genus is recognized
to form a monophyletic assemblage together with thefollowing five genera: Austrolabrus, Dotalabrus, Eupe-
trichthys, Notolabrus, and Pictilabrus (Russell, 1988).
Collectively, these fishes are termed the ‘‘pseudolab-
rines,’’ with all but Pseudolabrus restricted to Australia
and New Zealand. DNA sequence data from the mito-
chondrial 12S and 16S rRNA genes, plus the intervening
tRNAVal, were used to elucidate the phylogenetic in-
terrelationship within Pseudolabrus and among thegenera of pseudolabrines. Based on the phylogeny,
probable mechanisms for the development of the Pseu-
dolabrus antitropicality were inferred, together with its
timing.
2. Materials and methods
2.1. Materials examined
Fishes used in this study were collected via a variety
of methods, including the use of both hand- and gill-nets
while SCUBA diving, angling, and from museum spec-
imens. Tissue samples were obtained from six of the 11
recognized species of the antitropical genus Pseudola-
brus (for their geographic distributions, see Fig. 1),including both of the Northern Hemisphere represen-
tatives P. sieboldi and P. eoethinus. Three of the seven
recognized species of Notolabrus (Russell, 1988), and
single representatives of Austrolabrus, Eupetrichthys,
and Pictilabrus were included in the phylogenetic anal-
yses as possible sister-groups of Pseudolabrus. Both
Austrolabrus and Eupetrichthys are monotypic (Russell,

Table 1
List of species used in the study, with DDBJ/GenBank/EMBL accession numbers
Classification Species Accession No.
Order Perciformes
Suborder Labroidei
Family Pomacentridae Abudefduf vaigiensis AB121234
Family Labridae
Tribea Hypsigenyins Choerodon azurio AB121235
Tribea Julidines Halichoeres tenuispinnis AB121236
Tribea Labrichthyines Ophthalmolepis lineolata AB121237
Suezichthys gracilis AB121238
Tribea Pseudolabrines Notolabrus gymnogenis AB121239
Notolabrus parilus AB121240
Notolabrus tetricus AB121241
Pseudolabrus biserialis AB121242
Pseudolabrus eoethinus AB121243
Pseudolabrus fuentesi AB121244
Pseudolabrus guentheri AB121245
Pseudolabrus miles AB121246
Pseudolabrus sieboldi AB121247
Pictilabrus laticlavius AB121248
Austrolabrus maculatus AB121249
Eupetrichthys angustipes AB121250
aLabrid tribes after Gomon (1997).
K. Mabuchi et al. / Molecular Phylogenetics and Evolution 32 (2004) 375–382 377
1988), while Pictilabrus consists of three species
(Hutchins and Morrison, 1996). The two species of
Dotalabrus were not examined. To root correctly the
phylogenetic network within the pseudolabrines, the
following four labrid species, Suezichthys gracilis, Oph-
thalmolepis lineolata, Halichoeres tenuispinnis, andChoerodon azurio were also included in the analysis,
because the former three species belong to the genera
which were recognized to be closely related to the
pseudolabrines based on osteological data (Russell,
1988), and the last species is classified into the Hypsig-
enyini which is believed to be most primitive among
labrid tribes (Gomon, 1997). In addition, the poma-
centrid, Abudefduf vaigiensis was used as an outgroup ofthe labrid species, on the basis of traditionally accepted
classification: Pomacentridae and Labridae both belong
to the suborder Labroidei (Nelson, 1994). Species used
in this study were listed in Table 1.
2.2. DNA amplification and sequencing
Genomic DNA was extracted from muscle tissuepreserved in 99.5% ethanol, using the Qiagen DNeasy
tissue kit. An approximately 1500 bp fragment of the
mitochondrial genome including the latter half of 16S
rRNA, the entire tRNAVal, and former half of the 12Sr
RNA gene was amplified using the following two sets of
primers: L1083-12S (ACA AAC TGG GAT TAG ATA
C)+H1903-16S (GTA GCT CGT YTA GTT TCG GG)
and L1803-16S (AGT ACC GCA AGG GAA AGCTGA AA)+H2590-16S (ACA AGT GAT TGC GCT
ACC TT) (Miya and Nishida, 2000). The two primer-
sets amplified two different fragments partially over-
lapping each other. PCR proceeded for 30 cycles on a
Model 9700 thermal cycler (Applied Biosystems), with
denaturation at 94 �C for 10 s, annealing at 45 �C for
10 s, and extension at 72 �C for 30 s, the final cycle beingfollowed by an extension at 72 �C for 5min. The PCR
products were electrophoresed on a 1% agarose gel,
purified using a Pre-Sequencing Kit (USB), and then
sequenced with dye-labeled terminators (Applied Bio-
systems). Primers used were the same as those for PCR
and each of the two partially overlapping fragments was
sequenced for both strands. All sequencing reactions
were performed according to the manufacturer�s in-structions. Labeled fragments were analyzed on a Model
3100 DNA sequencer (Applied Biosystems). All
sequences are available from DDBJ, EMBL, and Gen-
Bank under accession numbers shown in Table 1.
2.3. DNA alignment
DNA sequences were edited using the computer pro-grams EditView ver. 1.01 (Applied Biosystems), Auto-
Assembler ver. 2.1 (Applied Biosystems), and DNASIS
ver. 3.2 (Hitachi Software Engineering). The accuracy of
base determination was checked by comparison of light
and heavy complementary DNA strands. The two rRNA
and single tRNA genes, together with their secondary
structures, were identified by homology to other verte-
brates (for rRNA, European ribosomal RNA database:http://oberon.fvms.ugent.be:8080/rRNA/index.html; for
A
B
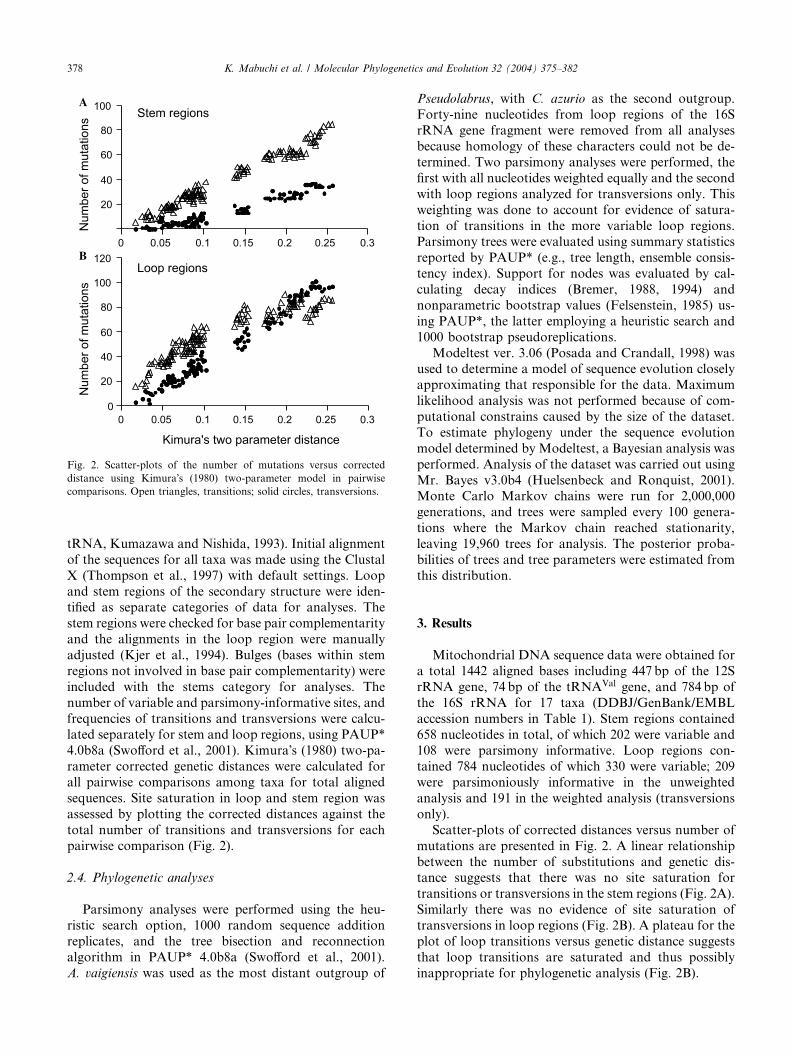
Fig. 2. Scatter-plots of the number of mutations versus corrected
distance using Kimura�s (1980) two-parameter model in pairwise
comparisons. Open triangles, transitions; solid circles, transversions.
378 K. Mabuchi et al. / Molecular Phylogenetics and Evolution 32 (2004) 375–382
tRNA, Kumazawa and Nishida, 1993). Initial alignment
of the sequences for all taxa was made using the Clustal
X (Thompson et al., 1997) with default settings. Loop
and stem regions of the secondary structure were iden-tified as separate categories of data for analyses. The
stem regions were checked for base pair complementarity
and the alignments in the loop region were manually
adjusted (Kjer et al., 1994). Bulges (bases within stem
regions not involved in base pair complementarity) were
included with the stems category for analyses. The
number of variable and parsimony-informative sites, and
frequencies of transitions and transversions were calcu-lated separately for stem and loop regions, using PAUP*
4.0b8a (Swofford et al., 2001). Kimura�s (1980) two-pa-rameter corrected genetic distances were calculated for
all pairwise comparisons among taxa for total aligned
sequences. Site saturation in loop and stem region was
assessed by plotting the corrected distances against the
total number of transitions and transversions for each
pairwise comparison (Fig. 2).
2.4. Phylogenetic analyses
Parsimony analyses were performed using the heu-
ristic search option, 1000 random sequence addition
replicates, and the tree bisection and reconnection
algorithm in PAUP* 4.0b8a (Swofford et al., 2001).
A. vaigiensis was used as the most distant outgroup of
Pseudolabrus, with C. azurio as the second outgroup.Forty-nine nucleotides from loop regions of the 16S
rRNA gene fragment were removed from all analyses
because homology of these characters could not be de-
termined. Two parsimony analyses were performed, the
first with all nucleotides weighted equally and the second
with loop regions analyzed for transversions only. This
weighting was done to account for evidence of satura-
tion of transitions in the more variable loop regions.Parsimony trees were evaluated using summary statistics
reported by PAUP* (e.g., tree length, ensemble consis-
tency index). Support for nodes was evaluated by cal-
culating decay indices (Bremer, 1988, 1994) and
nonparametric bootstrap values (Felsenstein, 1985) us-
ing PAUP*, the latter employing a heuristic search and
1000 bootstrap pseudoreplications.
Modeltest ver. 3.06 (Posada and Crandall, 1998) wasused to determine a model of sequence evolution closely
approximating that responsible for the data. Maximum
likelihood analysis was not performed because of com-
putational constrains caused by the size of the dataset.
To estimate phylogeny under the sequence evolution
model determined by Modeltest, a Bayesian analysis was
performed. Analysis of the dataset was carried out using
Mr. Bayes v3.0b4 (Huelsenbeck and Ronquist, 2001).Monte Carlo Markov chains were run for 2,000,000
generations, and trees were sampled every 100 genera-
tions where the Markov chain reached stationarity,
leaving 19,960 trees for analysis. The posterior proba-
bilities of trees and tree parameters were estimated from
this distribution.
3. Results
Mitochondrial DNA sequence data were obtained for
a total 1442 aligned bases including 447 bp of the 12S
rRNA gene, 74 bp of the tRNAVal gene, and 784 bp of
the 16S rRNA for 17 taxa (DDBJ/GenBank/EMBL
accession numbers in Table 1). Stem regions contained
658 nucleotides in total, of which 202 were variable and108 were parsimony informative. Loop regions con-
tained 784 nucleotides of which 330 were variable; 209
were parsimoniously informative in the unweighted
analysis and 191 in the weighted analysis (transversions
only).
Scatter-plots of corrected distances versus number of
mutations are presented in Fig. 2. A linear relationship
between the number of substitutions and genetic dis-tance suggests that there was no site saturation for
transitions or transversions in the stem regions (Fig. 2A).
Similarly there was no evidence of site saturation of
transversions in loop regions (Fig. 2B). A plateau for the
plot of loop transitions versus genetic distance suggests
that loop transitions are saturated and thus possibly
inappropriate for phylogenetic analysis (Fig. 2B).

Fig. 3. A single most-parsimonious tree with loop regions analyzed for transversions only. Abudefduf vaigiensis was used as the most distant out-
group. Numbers above nodes indicate bootstrap values from 1000 pseudoreplications (only values 50% are shown), numbers below nodes indicate
decay indices. TL¼ 915, CI¼ 0.53, RI¼ 0.52. Posterior probabilities from 19,960 trees produced by a Bayesian analysis under TrN+ I+G are also
indicated in the parentheses along the branches (only values 80% are shown).
K. Mabuchi et al. / Molecular Phylogenetics and Evolution 32 (2004) 375–382 379
Parsimony analysis of the nucleotide data excluding
loop transitions resulted in a single most-parsimonious
tree (Fig. 3: TL¼ 915, CI excluding uninformative
characters¼ 0.53, RI¼ 0.52). The species of pseudolab-
rines were monophyletic with a bootstrap value of 84and a decay index of 3. The pseudolabrines clade was
divided into two clades, Pseudolabrus+Notolabrus clade
and Austrolabrus+Eupetrichthys+Pictilabrus clade, the
former clade supported by a bootstrap value of 97 and a
decay index of 7, and the latter clade by a bootstrap
value of 98 and a decay index of 9. Pseudolabrus is pa-
raphyletic with respect to Notolabrus: Pseudolabrus
biserialis and P. miles form a monophyletic assemblagetogether with three Notolabrus species, with a bootstrap
value of 80 and a decay index of 2. The two Northern
Hemisphere species, Pseudolabrus eoethinus and P. sie-
boldi, were monophyletic with a bootstrap value of 94
and a decay index of 5, and the Northern Hemisphere
clade was sister to an Eastern Pacific species, P. fuentesi
with a bootstrap value of 99 and a decay index of 8.
Pseudolabrus guentheri was sister to the latter threePseudolabrus species with a bootstrap value of 67 and a
decay index of 1. The pseudolabrines clade was sister to
S. gracilis with a bootstrap value of 99 and a decay in-
dex of 13, and the pseudolabrines +S. gracilis clade was
sister to O. lineolata+H. tenuispinnis clade with a
bootstrap value of 98 and a decay index of 11.
Parsimony analysis of the unweighted nucleotide data
resulted in three equally most-parsimonious trees (TL ¼1192, CI excluding uninformative characters¼ 0.53,
RI¼ 0.54). One of the three most-parsimonious trees
had the same topology as the single most-parsimonious
tree resulting from the weighted parsimony analysis.
Among the three equally most-parsimonious trees, both
of the Pseudolabrus+Notolabrus clade and Austrola-
brus+Eupetrichthys+Pictilabrus clade were repro-
duced, and, within the former clade, the monophyly of
the two Northern Hemisphere species of Pseudolabrus,
its sister relationship to the East Pacific congenericspecies, and the monophyly of P. miles, P. biserialis,
and monophyletic three Notolabrus species were also
recovered.
Results from Modeltest (Posada and Crandall, 1998)
indicated that the model TrN+ I+G provided the best fit
to the data as determined by the likelihood ratio test. The
Bayesian analysis produced 20,000 trees of which 40 were
discarded as burn-in. This left 19,960 trees to estimatetopology and tree parameters. The percentage of times a
node occurs within these 19,960 trees was interpreted as
the posterior probability of the node (see Fig. 3). Esti-
mated model parameters were� ln l ¼ 7292:298� 4:737,A ¼ 0:360� 0:011, C ¼ 0:249� 0:010, G ¼ 0:188�0:009, T ¼ 0:202� 0:009, a ¼ 0:555� 0:126. Topologyof 50% consensus tree was identical to that of the most-
parsimonious tree from weighted analysis (see Fig. 3),except Pseudolabrus biserialis and P. miles were mono-
phyletic only in the former. The other major clades de-
scribed above were all supported by 100% posterior
probability (see Fig. 3).
4. Discussion
4.1. Taxonomic implications
The results of this study support pseudolabrines� re-lationships that do not agree with the previous hy-
pothesis based on osteological data (Russell, 1988)

Dotalabrus*
Notolabrus
Pseudolabrus
Pictilabrus
Austrolabrus
Eupetrichthys
Fig. 4. Alternative phylogenetic hypothesis for pseudolabrines based
on comparative osteology (Russell, 1988). The genus with asterisk was
not examined in the present study.
380 K. Mabuchi et al. / Molecular Phylogenetics and Evolution 32 (2004) 375–382
(Fig. 4). The most notable difference between the rela-
tionships based on molecular data and that on osteo-
logical data involves the phylogenetic relationships
between Pseudolabrus and Notolabrus. In Russell�s(1988) hypothesis, Pseudolabrus is monophyletic, andphylogenetically separated from Notolabrus: Pseudola-
brus is sister to a Austrolabrus+Pictilabrus clade, and
the Pseudolabrus+Austrolabrus+Pictilabrus clade is
sister to Eupetrichthys, Notolabrus being sister to the
clade consisting of these four genera. It was based on
these relationships that Russell (1988) erected Notola-
brus, and its constituent species were previously allo-
cated to Pseudolabrus. Our molecular analyses, however,demonstrated that Pseudolabrus was paraphyletic to
Notolabrus, and the Pseudolabrus+Notolabrus clade
was sister to the Austrolabrus+Pictilabrus+Eupetrich-
thys clade (Fig. 3). Although a final decision cannot be
reached now owing to incomplete taxon sampling, the
present molecular phylogeny seems to indicate that the
generic status of Notolabrus (or generic delimitations of
both Notolabrus and Pseudolabrus) requires review.
4.2. Probable mechanisms for the development of the
Pseudolabrus antitropicality
Phylogenetic analyses based on the molecular data
demonstrated that the two Northern Hemisphere species
of Pseudolabrus were monophyletic and nested deep
within the clade of Southern Hemisphere pseudolabrines(Fig. 3). Mapping species distributions onto the MP tree
indicated that both pseudolabrines and Pseudolabrus
originated in the Southern Hemisphere. Although some
Pseudolabrus species were not examined here, this
inference will not be upset by any addition of the yet to
be examined species of Pseudolabrus, because such spe-
cies all occur in the Southern Hemisphere. However, the
possibility of independent southern origins of theNorthern Hemisphere species cannot be discounted
(e.g., Burridge and White, 2000; Grant and Bowen,
1998).
Several mechanisms have been proposed for the de-velopment of antitropical distributions, divisible into
two classes, dispersal and vicariance (Burridge, 2002).
The dispersal mechanism assumes that a taxon was
originally represented on one side of the tropics, and
subsequently moved across the equator to colonize the
opposite hemisphere. On the other hand, the vicariance
mechanism assumes that taxa once occupied the tropics
but were later extirpated, resulting in isolated Northernand Southern Hemisphere populations. In the present
case, the vicariance mechanism with tropical origin of
Pseudolabrus seems to be unlikely, because such a
mechanism requires both multiple independent adapta-
tions to temperate waters and highly biased allotment of
species to each Hemisphere for explaining the distribu-
tion of Pseudolabrus. The vicariance mechanism with the
Southern Hemisphere origin of Pseudolabrus also seemsto be unlikely, because it requires the common ancestor
of the Northern Hemisphere species, once adapted to
tropical waters, to colonize temperate Asia without
leaving any descendants in the tropical region.
The dispersal mechanism with the Southern Hemi-
sphere origin of Pseudolabrus, seems more likely than
the above two vicariance mechanisms. In our result, the
two Northern Hemisphere species were monophyleticand derived relatively recently during the radiation of
Pseudolabrus (Fig. 3). Therefore, the dispersal mecha-
nism can explain the Pseudolabrus antitropicality by
only a single (two even at maximum) transequatorial
dispersal(s).
Within the antitropical marine fish groups studied to
date using molecular data, nearly half of the estimates of
transequatorial divergences fall within Pleistocene, per-haps implicating cooler temperatures during glaciations
for transequatorial dispersal (Burridge, 2002). However,
transequatorial divergence during recent glaciations
does not appear to be the case for Pseudolabrus. The
transequatorial divergence in partial 16S rRNA gene of
Pseudolabrus (divergences between Pseudolabrus fuentesi
and each of the two Asian Pseudolabrus species) ranges
from 3.8 to 4.1% [corrected by Kimura�s (1980) two-parameter model]. Applying a 16S rRNA molecular
clock calibration of 1.0% sequence divergence (for all
base substitutions) per million years derived from gem-
inate amphi-American Centropomus (Tringali et al.,
1999), the transequatorial divergence of Pseudolabrus is
estimated as 3.8–4.1 million years ago, corresponding to
early to mid Pliocene, which is older than Pleistocene or
late Pliocene. However, more recent transequatorialdivergences may become apparent with the analysis of
remaining five Pseudolabrus species.
Although some Southern Hemisphere species of
Pseudolabrus were not examined in this study, it is very
interesting that the monophyletic two Northern Hemi-
sphere species of Pseudolabrus were sister not to Aus-
tralian or New Zealand species, but to an East Pacific

K. Mabuchi et al. / Molecular Phylogenetics and Evolution 32 (2004) 375–382 381
species, Pseudolabrus fuentesi, from Easter Island (Figs.1 and 3). A similar phenomenon was found in the de-
mersal nearshore fish subgenus Goniistius (family
Cheilodactylidae, order Perciformes) (Burridge and
White, 2000), which is similar to Pseudolabrus in dis-
tribution and habitat. Within the antitropical subgenus,
one of the three Asian species, Cheilodactylus (Goniis-
tius) zebra showed the highest sequence similarity not
with other Japanese or Australian-New Zealand species,but with a species from Easter Island, Cheilodactylus
(G.) plessisi (Burridge and White, 2000). This fact to-
gether with our result for Pseudolabrusmay indicate that
some transequatorial dispersals undertaken by Asian
species might have occurred from the East Pacific rather
than the West Pacific, although Burridge (2002) still
considered Goniistius a West Pacific divergence. Inter-
estingly, Parrish et al. (1989) suggested that transequa-torial divergences were more predominant in the coastal
East Pacific, owing to cooler water temperature, nar-
rower tropical zone, etc. The timing of the transequa-
torial divergence between C. (G.) zebra and C. (G.)
plessisi was estimated as late Miocene-early Pliocene
(Burridge and White, 2000), which is, interestedly,
comparable to that of transequatorial divergence for
Pseudolabrus.
Acknowledgments
We thank Australian Museum, Osaka Aquarium
KAIYUKAN, H. Ikeda, and R. Robertson for pro-
viding tissue or fresh whole specimens. We also thankthe following individuals and institutions for their help
in collecting the specimens: Y. Yanagisawa, J. Paxton,
M. McGrouther, S. Reader, M. Uno, J.B. Hutchins,
S.M. Morrison, Weekly Sunday Fishing, T. Kudo, K.
Matsumoto, T. Mukai, and The University of Queens-
land. Thanks are also due to C.P. Burridge, B.C. Rus-
sell, and J.G. Inoue for providing useful information
and for discussions of the results. We are grateful toC.P. Burridge for correcting English. This study was
supported partly by Research Fellowships of the Japan
Society for the Promotion of Science for Young Scien-
tists (No. 72503), and Grants-in-Aid from the Ministry
of Education, Culture, Sports, Science and Technology,
Japan (12NP0201).
References
Bremer, K., 1988. The limits of amino acid sequence data in
angiosperm phylogenetic reconstruction. Evolution 42, 795–803.
Bremer, K., 1994. Branch support and tree stability. Cladistics 10, 295–
304.
Briggs, J.C., 1987. Antitropical distribution and evolution in the Indo-
West Pacific Ocean. Syst. Zool. 36, 237–247.
Briggs, J.C., 1995. Global Biogeography. Elsevier, New York.
Burridge, C.P., 2002. Antitropicality of Pacific fishes: molecular
insights. Environ. Biol. Fish. 65, 151–164.
Burridge, C.P., White, R.W.G., 2000. Molecular phylogeny of the
antitropical subgenus Goniistius (Perciformes: Cheilodactylidae:
Cheilodactylus): evidence for multiple transequatorial divergences
and non-monophyly. Biol. J. Linn. Soc. 70, 435–458.
Crame, J.A., 1993. Bipolar molluscs and their evolutionary implica-
tions. J. Biogeogr. 20, 145–161.
Darwin, C., 1859. On the origin of species by means of natural
selection, or the preservation of forward races in the struggle for
life. London.
Ekman, S., 1953. Zoogeography of the Sea. Sidwick and Jackson,
London.
Felsenstein, J., 1985. Confidence limits on phylogenetics: an approach
using the bootstrap. Evolution 39, 783–791.
Gomon, M.F., 1997. Relationships of fishes of the labrid tribe
Hypsigenyini. Bull. Mar. Sci. 60, 789–871.
Grant, W.S., Bowen, B.W., 1998. Shallow population histories in
deep evolutionary lineages of marine fishes: insights from
sardines and anchovies and lessons for conservation. J. Hered.
89, 415–426.
Hubbs, C.L., 1952. Antitropical distribution of fishes and other
organisms. Symposium on the Problems of Bipolarity and of
Pantemperate Faunas. In: Proceeding of the Seventh Pacific
Science Congress (Pacific Science Association), vol. 3, pp. 324–
329.
Huelsenbeck, J.P., Ronquist, F.R., 2001. MRBAYES: Bayesian
inference of phylogenetic trees. Bioinformatics 17, 754–755.
Hutchins, J.B., Morrison, S.M., 1996. Pictilabrus brauni, a new species
of labrid fish from South-western Australia. Revue Fr. Aquariol.
23, 39–42.
Kimura, M., 1980. A simple method for estimating evolutionary rate
of base substitutions through comparative studies of nucleotide
sequences. J. Mol. Evol. 16, 111–120.
Kjer, K.M., Baldridge, G.D., Fallon, A.M., 1994. Mosquito large
subunit ribosomal RNA: simultaneous alignment of primary and
secondary structure. Biochem. Biophys. Acta 1217, 147–155.
Kumazawa, Y., Nishida, M., 1993. Sequence evolution of mitochon-
drial tRNA genes and deep-branch animal phylogenetics. J. Mol.
Evol. 37, 380–398.
Lindberg, D.R., 1991. Marine biotic interchanges between Northern
and Southern Hemispheres. Paleobiology 17, 308–324.
Mabuchi, K., Nakabo, T., 1997. Revision of the genus Pseudolabrus
(Labridae) from the East Asian waters. Ichthyol. Res. 44, 321–334.
Miya, M., Nishida, M., 2000. Use of mitogenomic information in
teleostean molecular phylogenetics: a tree-based exploration under
the maximum-parsimony optimality criterion. Mol. Phylogenet.
Evol. 17, 437–455.
Nelson, G.J., 1985. A decade of change: the future of biogeography.
Earth Sci. Hist. 4, 187–196.
Nelson, J.S., 1994. Fishes of the World, third ed. Wiley, New York.
Parrish, R.H., Serra, R., Grant, W.S., 1989. The monotypic sardines,
Sardina and Sardinops: their taxonomy, distribution, stock struc-
ture, and zoogeography. Can. J. Fish. Aquat. Sci. 46, 2019–
2036.
Posada, D., Crandall, K.A., 1998. Modeltest: testing the model of
DNA substitution. Bioinformatics 14, 817–818.
Randall, J.E., 1981. Examples of antitropical and antiequatorial
distribution of Indo-West-Pacific fishes. Pac. Sci. 35, 197–209.
Rehder, H.A., 1980. The marine mollusks of Easter Island (Isla de
Pascua) and Sala y G�omez. Smithsonian Contrib. Zool. 289, 1–167.
Rotondo, G.M., Springer, V.G., Scott, G.A.J., Schlander, S.O., 1981.
Plate movement and island integration—a possible mechanism in
the formation of endemic biotas, with special reference to the
Hawaiian Islands. Syst. Zool. 30, 12–21.
Russell, B.C., 1988. Revision of the labrid fish genus Pseudolabrus and
allied genera. Rec. Aust. Mus., Suppl. 9, 1–72, pls. 1–4.

382 K. Mabuchi et al. / Molecular Phylogenetics and Evolution 32 (2004) 375–382
Springer, V.G., 1982. Pacific Plate biogeography, with special reference
to shorefishes. Smithsonian Contrib. Zool. 367, 1–182.
Swofford, D.L., Waddell, P.J., Huelsenbeck, J.P., Foster, P.G., Lewis,
P.O., Rogers, J.S., 2001. Bias in phylogenetic estimation and its
relevance to the choice between parsimony and likelihood methods.
Syst. Biol. 50, 525–539.
Thompson, J.D., Gibson, T.J., Plewniak, E., Jeanmougin, F., Higgins,
D.G., 1997. The ClustalX-Windows interface: flexible strategies for
multiple sequence alignment aided by quality analysis tools.
Nucleic Acids Res. 25, 4876–4882.
Tringali, M.D., Bert, T.M., Seyoum, S., Bermingham, E., Bartolacci,
D., 1999. Molecular phylogenetics and ecological diversification of
the transisthmian fish genus Centropomus (Perciformes: Centrop-
omidae). Mol. Phylogenet. Evol. 13, 193–207.
Valentine, J.W., 1984. Neogene marine climate trends: implications for
biogeography and evolution of shallow-sea biota. Geology 12, 647–
650.
White, B.N., 1986. The isthmian link, antitropicality and American
biogeography: distributional history of the Atherinopsinae (Pisces:
Atherinidae). Syst. Zool. 35, 176–194.





























