Embed Size (px)
Citation preview

337
1040-9238/01/$.50© 2001 by CRC Press LLC
Critical Reviews in Biochemistry and Molecular Biology, 36(4):337–397 (2001)
Molecular Mechanisms of DNA Damageand Repair: Progress in Plants*
Narendra Tuteja1,** Mohan B. Singh,2 Mithilesh K. Misra,3
Prem L. Bhalla,2 and Renu Tuteja1
Referee: Dr. Nawin C. Mishra, Professor of Genetics, University of South Carolina,Department of Biological Sciences, Columbia, SC 29208
1International Centre for Genetic Engineering and Biotechnology, Aruna Asaf AliMarg, New Delhi — 110 067, India; 2Institute of Land and Food Resources, TheUniversity of Melbourne, Australia; 3Department of Biochemistry, University ofLucknow, Lucknow, India
* Dedicated to Prof. Jan H. J. Hoeijmakers.
** To whom correspondence should be addressed. Phone: 91-11-6181242; Fax; 91-11-6162316; e-mail:[email protected]
Table of Contents
I. INTRODUCTION ............................................................................................. 339
II. DNA DAMAGES .............................................................................................. 340
A. Types of DNA Damage and Damage Products ...................................... 341
B. Free Radicals Production and Antioxidant
Defense Mechanisms ............................................................................... 343
C. Lipid Peroxidation and DNA Damage .................................................... 348
D. Ozone and DNA Damage in Plants ........................................................ 350
E. DNA Damage Response .......................................................................... 351
III. VARIOUS DNA REPAIR MECHANISMS ................................................... 351
A. Direct Reversal ........................................................................................ 351
B. Base Excision Repair ............................................................................... 353
C. Nucleotide Excision Repair ..................................................................... 356
D. Photoreactivation and Photolyases .......................................................... 362
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

338
E. Bypass Damage Repair ............................................................................ 364
F. Double-Strand Break Repair ................................................................... 365
G. Mismatch Repair ...................................................................................... 369
IV. ROLE OF DNA HELICASES IN DAMAGE
AND REPAIR .................................................................................................... 369
V. DESICCATION RELATED DAMAGES, TOLER-
ANCE AND REPAIR ....................................................................................... 371
A. In Bacterial Spores .................................................................................. 373
B. In Seeds .................................................................................................... 374
C. DNA Integrity in Pollens......................................................................... 375
VI. DNA REPAIR GENES IDENTIFIED BY
SEQUENCE HOMOLOGY ............................................................................. 375
VII. CONCLUSION AND FUTURE PROSPECTS ............................................. 377
ABBREVIATIONS: 6-4 pps, 6-4 photo products; AOS, activted oxygen species; AP, apurinic/apyrimidinic; APXs, ascorbate peroxidases; BER, base excision repair; CPD, cyclobutane pyrimi-dine dimer; CS, Cockayne syndrome; DNA-PK, DNA-dependent protein kinase; DR, direct rever-sal; DSB, double strand break; ERCC, excision repair cross complementation; FEN-1, flapendonucelase; GO, 7,8-dihydro-8-oxoguanine; GR, glutathione reductase; GSH, reduced glutathione;GSSH, oxidized glutathione; HDH, human DNA helicases; L., lipid radicals; LO., alkoxyl radicals;LOO., lipid peroxide; LOOH, lipid hydroperoxide; MDA, malondialdehyde; MGMT, methylguanine-methyltransferase; MMR, mismatch repair; MMS, methylmethanesulfonate; NER, nuleotide exci-sion repair; NHEJ, nonhomologous end-joining; O2¯
•, superoxide anion radical; •OH, hydroxylradical; PARP-1, poly(ADP-ribose) polymerase; PCNA, proliferative cell nuclear antigen ; PI3K,phosphatidylinositol 3-kinase; PUFA, polyunsaturated fatty acids; RAR, repair and recombination;RFC, replication factor C; ROS, reactive oxygen species; RPA, replication protein A; RSH, thiylradicals (‘sulfur’ containing functional group); SASP, small acid-soluble protein; SDSA, synthesis-dependent strand annealing; SOD, superoxide dismutase; SSA, single strand annealing; SSBR,single strand break repair; UV, ultraviolet; XP, Xeroderma pigmentosum.
ABSTRACT: Despite stable genomes of all living organisms, they are subject todamage by chemical and physical agents in the environment (e.g., UV and ionizingradiations, chemical mutagens, fungal and bacterial toxins, etc.) and by free radi-cals or alkylating agents endogenously generated in metabolism. DNA is alsodamaged because of errors during its replication. The DNA lesions produced bythese damaging agents could be altered base, missing base, mismatch base, deletionor insertion, linked pyrimidines, strand breaks, intra- and inter-strand cross-links.
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

339
These DNA lesions could be genotoxic or cytotoxic to the cell. Plants are mostaffected by the UV-B radiation of sunlight, which penetrates and damages theirgenome by inducing oxidative damage (pyrimidine hydrates) and cross-links (bothDNA protein and DNA-DNA) that are responsible for retarding the growth anddevelopment. The DNA lesions can be removed by repair, replaced by recombina-tion, or retained, leading to genome instability or mutations or carcinogenesis orcell death. Mostly organisms respond to genome damage by activating a DNAdamage response pathway that regulates cell-cycle arrest, apoptosis, and DNArepair pathways. To prevent the harmful effect of DNA damage and maintain thegenome integrity, all organisms have developed various strategies to either reverse,excise, or tolerate the persistence of DNA damage products by generating anetwork of DNA repair mechanisms. A variety of different DNA repair pathwayshave been reported that include direct reversal, base excision repair, nucleotideexcision repair, photoreactivation, bypass, double-strand break repair pathway, andmismatch repair pathway. The direct reversal and photoreactivation require singleprotein, all the rest of the repair mechanisms utilize multiple proteins to remove orrepair the lesions. The base excision repair pathway eliminates single damagedbase, while nucleotide excision repair excises a patch of 25- to 32-nucleotide-longoligomer, including the damage. The double-strand break repair utilizes eitherhomologous recombination or nonhomologous endjoining. In plant the latter path-way is more error prone than in other eukaryotes, which could be an importantdriving force in plant genome evolution. The Arabidopsis genome data indicatedthat the DNA repair is highly conserved between plants and mammals than withinthe animal kingdom, perhaps reflecting common factors such as DNA methylation.This review describes all the possible mechanisms of DNA damage and repair ingeneral and an up to date progress in plants. In addition, various types of DNAdamage products, free radical production, lipid peroxidation, role of ozone,dessication damage of plant seed, DNA integrity in pollen, and the role of DNAhelicases in damage and repair and the repair genes in Arabidopsis genome are alsocovered in this review.
I. INTRODUCTION
In general, the genomes of all theliving organisms, including plants, arestable. Normally the DNA of genome inthe cell replicates during cell divisionand passes all the genetic information totheir progeny. It is very important for allliving organisms to ensure proper func-tioning and propagation of their geneticinformation. However, due to constantexposure of the genome to various en-dogenous and environmental agents, theDNA gets damaged and can produce alarge variety of DNA lesions. These le-sions can affect the fidelity of DNA rep-lication (Painter, 1985), and transcrip-
tion (Protic-Sabljic and Kraemer, 1985),which can create mutations in importantprotein coding sequences. As a result,the produced mutated protein can affectvarious biological processes leading tothe genome instability. Proliferating cellsare often presumed to be more mutablethan quiescent cells because they haveless time to repair DNA damage beforeDNA replication. If these damages oc-cur in the germ cells, it can be heritableand will be harmful to the next genera-tion in passing a heritable disease. More-over, the damage in somatic cells playsan important role in the development ofcancer and aging.
The DNA damage can have genotoxicand cytotoxic effects on the cell. The real
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

340
biological consequences of these dam-aged products usually depend on thechemical nature of the lesion. In plants ifthis damage are not repaired properly theycan induce proliferation as well as playan important role in the aging ofseeds stocks and perennial crops. Thisunrepaired damage can also lead to thegeneral deterioration of cell function andcell death. This damage situation requiresconstant and accurate excision and thereplacement of damaged nucleotide byvarious DNA repair pathways. In orderto maintain the integrity of the genome,the prokaryotic and eukaryotic organismsare well equipped with several DNA re-pair mechanism pathways (Pieper et al.,1998; Britt, 1999). In general, the lesionsin the actively transcribed strand are re-paired more rapidly than the lesions inthe nontranscribed strand (Selby andSancar, 1993).
DNA repair is not only a fundamen-tal cellular process for protecting cellsagainst the damage, but it is also essen-tial to ensure the faithful transmission ofgenetic information from one genera-tion to the next. The biological conse-quence of defective DNA repair in hu-man has been reported to be the cause ofmany diseases, such as Xerodermapigmentosum (XP), Cockayne syndrome(CS), and Trichothiodystrophy (TTD)(Hoeijmakers, 1993a, b; Lehmann, 1998;Winkler et al., 1998; Lindahl and Word,1999; Tuteja and Tuteja, 2001).However, few inherited diseases are as-sociated with altered processing ofdouble-strand breaks such as Ataxia te-langiectasia and Nijmegen breakage syn-drome (Shiloh, 1997; Lindahl and Wood,1999). All the known mechanisms ofDNA repair processes have been shownto be complex and diverse and best stud-ied in microorganisms and mammals. Inhumans it is of great interest because of
the role of DNA repair in mutagenesis,in carcinogenesis, and possibly in aging(Ribeiro et al., 1998). In humans, themost important self-inflicted mutagen istobacco smoke, which is responsible formore cancer deaths. However, theprogress in plant genetic engineering andthe use of mutagenesis in the creation ofgenetic diversity for the improvementof crops have now demanded more re-search in DNA damage and repair. How-ever, DNA repair in plant has not beenwell studied, but now it is slowly pro-gressing. In this article we are describ-ing the molecular basis of DNA damageand repair and the recent advances ofDNA repair in plants. Further, we havealso focused on endogenous factors af-fecting DNA damage during naturaldesiccation process and the dependenceof desiccation tolerance mechanisms onexcision repair pathways.
II. DNA DAMAGES
The Earth’s atmosphere is gettingpolluted due to the man-made pollutantssuch as chlorofluorocarbons. As a re-sult, the stratospheric ozone layer is get-ting depleted, which causes the increasedexposure to solar ultraviolet-B (UV-B,wavelength 280 to 320 nm) on theEarth’s surface. A possible increase inUV radiation by ozone depletion is nowattracting attention of researchers. Thisincreased exposure of UV-B is harmfulto all living organisms, especially to theplants. Plants are constantly being chal-lenged by UV radiation through the sun-light because of their obligatory require-ment of sunlight for photosynthesis. Thisradiation penetrates plant tissues anddamages their genome and other cellu-lar targets such as photosystem II and
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

341
plasma membrane ATPase (Stapleton,1992). These ambient and increased so-lar radiations are responsible for retard-ing growth and development, morpho-logical and biochemical alteration, andbiomass accumulation in the plants(Bornman, 1989; Tevini and Teramura,1989; Bornman and Teramura, 1993;Teramura and Sullivan, 1994; Jansen etal., 1998; Vonarx et al., 1998).
UV-B light is potentially very dam-aging to DNA and proteins and alsoincreases the free radical production.Plants have their own way of tacklingthis problem. They have developed twoprotective mechanisms to cope withharmful effect of UV. First, the shield-ing through the production of UV-ab-sorbing compound such as flavonoidsand anthocyanins and the reflection ofUV radiation by epicuticular waxes andcuticular structures (Bornman et al.,1997; Stapleton et al., 1997). Second,removal and direct reversion by photo-reactivation of UV-induced DNA lesionsthat involve photolyase (Pang and Hays,1991; Sutherland et al., 1996; Stapletonet al., 1997; Yasui and Eker, 1998; Britt,1999). Recently, Cathie Martin’s groupat John Innes Centre, UK, have shownthat the production of UV-protectingsunscreens in Arabidopsis is controlledthrough the transcriptional repression byAtMyB4. The mutant AtMyB4 line ismore tolerant of UV-B irradiation thanwild type (Jin et al., 2000). However,the light-dependent pathway is the ma-jor mechanism of repair of DNA dam-age in plant parts exposed to sunlightinduced by UV radiation. Other path-ways such as base excision repair (BER),nucleotide excision repair (NER),postreplication, and recombinational re-pair may be more important in internaltissues where light penetration is lim-ited (Hoeijmakers and Bootsma, 1990;
Hoeijmakers, 1993 a, b; Wood, 1997;Balajee and Bohn, 2000).
Besides UV radiation, the plant ge-nomic DNA is vulnerable to DNA dam-age from a variety of endogenous reac-tive metabolites, particularly oxygen-freeradicals. The DNA in reproductive struc-tures, pollen, and seed is particularlysusceptible to endogenous DNA-dam-aging agents. In the plants there are anumber of developmental stages that areparticularly susceptible to the DNA dam-age. As a part of developmental path-way, seeds and pollens undergo the pro-cess of dehydration. This loss of thewater triggers several metabolic changes,leading to free radical formations, whichare capable of damaging DNA eitherdirectly or through lipid peroxidation.The accumulation of DNA lesions hasbeen shown to accompany the seed-ag-ing process (Boubriak et al., 1997). Thenature and the role of endogenous me-tabolites involved in plant seed DNAdamage are not clear. However, it isrelevant to note that a naturally occur-ring product of lipid peroxidation,malondialdehyde (MDA), has beenshown recently to be a major endog-enous DNA damaging agent in mam-malian systems (Fink et al., 1997;Marnett, 1999 a, b; Lindahl and Wood,1999).
A. Types of DNA Damage andDamage Products
The variety of the DNA damage pro-duced by various endogenous and exog-enous agents is corrected by different re-pair pathways. Major DNA damageinduced by different DNA damagingagents are shown in Figure 1. The endog-enously generated damage to DNA is
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

342
FIGURE 1. Major DNA damage induced by different DNA-damaging agents.
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

343
known as “spontaneous DNA damage”(Britt, 1999), which is produced by thereactive metabolites (e.g., oxygen-freeradicals, hydroxyl radicals, superperoxide,and nitric oxide) (Demple and Harrison,1994) and the defect in normal processesof DNA replication or recombination. Theenvironmental DNA-damaging agentscould be UV-light, ionizing radiation,chemical mutagens, and cross-linkingagents (e.g., mitomycin C and cisplatin),alkylating agents, aromatic compounds,and fungal and bacterial toxins (Plooy etal., 1984; Friedberg, 1985; Doetsch,1995). The various DNA lesions producedby these agents are mismatches, base al-terations, deletion of bases, strand breaks,cyclobutane pyrimidine dimers, 6-4photo-products, intra- and inter-strandcross-links, alkylated bases, and bulkyadducts (Figure 1) (Friedberg et al., 1995).
The most common natural environ-mental genotoxic agent is the UV com-ponent of sunlight. The damage inducedby UV radiation to the epidermal cellsof a plant is as inevitable as hydrolyticdamage induced by the water present inthe nucleus (Britte, 1996). Besides UVand hydrolytic damage, other damagecould be alkylation damage, oxidativedamage and the damage, produced byionizing radiation (Britte, 1996). Theoxidative damage is a major stress to thechilling-sensitive plants (Hodges et al.,1997; Pinehro et al., 1997). Low tem-peratures together with high light inten-sities induce the production of activatedoxygen species (AOS), such as superox-ide (O2¯
•), hydroxyl radicals (.OH), andhydrogen peroxide (H2O2) (Wise andNaylor, 1987). These radicals can reactwith DNA, proteins, and lipid, causingdamage. However, normally the super-oxide dismutase (SODs), ascorbate per-oxidases (APXs), and catalases play im-portant role as a defense against these
radicals (Inze and Van-Montagu, 1995).The environmental stresses to plants,such as drought, heat shock, salt stress,temperature, etc., can generate the pro-duction of AOS and are also known toinduce plant SODs (Van Camp et al.,1994).
The chemotherapeutic agent Cisplatinforms the intrastrand adduct between theadjacent purines (AG or GG) (Plooy etal., 1984). The alkylating agents such asmethylmethanesulfonate (MMS) includeN-7-alkylguanine, O6-alkyl-guanine,N-3-alkyladenine, and O4-alkylthymidineinduce the major damage (Beranek, 1990;Friedberg et al., 1995). The N-3-alkyladenine, a damage product, is highlytoxic because it acts as a block to DNAreplication by not recognizing the DNApolymerase (Britt, 1999). Alkylatingagents can also form the phosphotriestersby alkylating the phosphodiesters(Vaughan et al., 1993). The ligandBleomycin produces single- and double-strand breaks by intercalating into DNA(Urdea et al., 1988). Overall, we can saythat a single DNA-damaging agent canproduce lesions that are inconsequential,mutagenic (because of mispairing), orcytotoxic (because it blocks the DNAreplication or transcription). The mainlesions produced in DNA by hydrolysis,reactive oxygen species, and small reac-tive intracellular molecules such asS-adenosylmethionine have been re-viewed by Lindahl (1993).
B. Free Radicals Productionand Anti-Oxidant DefenseMechanisms
Free radicals are chemical species(molecules or atoms) possessing an un-paired electron in their outermost orbital.
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

344
Due to the presence of one or more un-paired electrons, these species are para-magnetic, which makes them highly reac-tive. These are conventionally representedby a superscript dot R. (Dormandy, 1980).Free radicals can be formed in a moleculeby gaining an additional electron, for ex-ample, the reduction of molecular oxygen(O2) to the superoxide anion radical (O2¯
•).
O2 + e– Æ O2¯ •
Transition metals contain unpairedelectrons and so act as free radicals, withthe sole exception of zinc. The mostimportant feature of transition metals,from the free radical point of view, istheir variable valency, which allowsthem to undergo changes in their oxida-tion state involving one electron; for ex-ample, Fe (iron) has two common va-lencies. If a solution of ferrous salt (IronII) is left in contact with the air, it slowlyoxidizes to the ferric (Iron III) state.This is a one-electron oxidation andoxygen dissolved in solution is reducedto the superoxide radical (Halliwell andGutteridge, 1986).
Fe2+ + O2 ¤ Fe2+ O2 ¤ Fe3+ O2¯
¤ Fe3+ O2¯ •
(Intermediate Complexes)
Copper (Cu) has two common va-lencies Cu+ and Cu2+. Similarly, underoxidation conditions copper salts canreceive and donate electron to superox-ide radicals O2¯
•
Cu2+ + O2¯ • Æ Cu+ + O2
H2O2 + Cu+ Æ Cu2+ + •OH
The variable valency of the transi-tion metals helps them to be effectivecatalysts in many oxidation reductionreactions, and they are present for this
purpose at the active sites of many en-zymes catalyzing such reactions. Themost important free radicals in biologi-cal systems are a derivative of oxygen.The complete reduction of O2 by univa-lent pathway results in the formation ofsuperoxide anion, hydrogen peroxide(H2O2), and other products as shownbelow (Naqvi et al., 1986; Pryor, 1986):
O2 + e– æ Ææ O2¯ •
(superoxide)
2 O2¯ • + 2H+
SOD catalysedæ Æææææ
H2O2 + 3O2
(peroxide) (triplet O2)
2 O2¯ • + 2H+ spontaneousæ Æææææ
H2O2 + 1O2(peroxide) (singlet O2)
O2¯ • + H2O2 + H+ metal catalystæ Æææææ
O2 + •OH + H2O(hydroxyl radical)
O2¯ • + •OH + H+ æ Ææ H2O + 1O2
Hydrogen peroxide is an oxidizingagent but not especially reactive, and itsmain significance lies in it being a sourceof hydroxyl radicals. This radical is anextremely reactive oxidizing radical thatreacts with most biomolecules at diffu-sion-controlled rates. It will not diffusea significant distance within a cell be-fore reacting and has an extremely shorthalf-life but is capable of causing a greatdamage within a small radius of its siteof production. Under normal circum-stances, the major source of free radi-cals in cells is electron leakage fromelectron transport chain, such as thosein mitochondria. These react with
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

345
molecular oxygen to generate superox-ide ion. Other enzymes, such as flavinoxidases located in the peroxisome, canalso produce superoxide or hydrogenperoxide. Autooxidation of certain com-pounds, including ascorbic acid (vita-min C), thiols (e.g., Glutathione, cys-teine), adrenaline, and flavin co-enzymes are yet other sources ofsuperoxide anions in the cells. Theseautooxidation reactions can be greatlyenhanced by the transition metal ioninvolvement.
Various potential sources that areresponsible for the production of super-oxide anions are as follows:
1. Enzymes: Various enzymes areinvolved in catalyzing the oxida-tion reactions, as a result there isunivalent reduction of O2 to O2¯
•,for example, xanthine oxidase, al-dehyde oxidase, dihydrocerotic de-hydrogenase, flavin dehydrogenase(Fridovich, 1976), and peroxidases(Halliwell, 1990).
2. Autooxidation: a large group ofcompounds, including epinephrine,flavins, and ferrodoxin (Misra andFridovich, 1972), reduced forms ofriboflavin and thiol compoundssuch as cysteine and glutathione,generate superoxide radicals byautooxidation.
3. Subcellular organelles: the mito-chondrial electron transport chain,endoplasmic reticulum substratehydroxylation, chloroplasts, andneutrophils are all capable of pro-ducing superoxide radicals.
4. The oxidation of hemoglobin inerythrocytes is a very rich sourceof oxygen free radicals (Misra andFridovich, 1972).
5. Irradiation: irradiation of living tis-sues also produce superoxide an-ion and other free radicals.
Superoxide anion and hydrogenperoxide react together to generatehighly reactive hydroxyl (•OH) radical(Beuchamp and Fridovich, 1973). Per-oxides also react with metal cations toproduce hydroxyl radicals in the liv-ing system. Some important mecha-nisms responsible for generation ofhydroxyl radicals are briefly under-lined.
1. Harber-Weiss reaction: in the pres-ence of copper and/or iron (metalcatalyst), H2O2 reacts with O2¯
• andforms highly reactive •OH (Harberand Weiss, 1934; Kehrer, 2000).
O2¯ • + H2O2 (metal catalyst)æ Ææææææ •OH +
Fe/Cu
–OH + O2¯ •
2. Fenton reaction: damage to DNAby oxygen radicals (oxyradicals) ismediated by metal ions, especiallyiron, as in the classic Fenton reac-tion, which is the major source ofhighly reactive •OH radicals (Barbet al., 1957). This reaction was firstobserved by its inventor H. J. H.Fenton in 1894.
O2¯ • + Fe3+ Æ O2 + Fe2+
Fe2+ + H2O2 Æ Fe3+ + OH– + •OHFe2+ + •OH Æ Fe3+ + OH–
Fe3+ + H2O2 Æ Fe2+ + HO2. + H+
H2O2 + •OH Æ HO2. + H2O
HO2. ¤ O2–
• + H+
Fe3+ + O2¯ • Æ Fe2+ + O2
Halliwell and Gutteridge (1984)discuss the significance of Fentonreaction in free radical productionin a biological system. An analogueof this reaction, where hydroper-oxides replace H2O2, is also impor-tant in biological systems, and is
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

346
considered to be a Fenton type re-action.
Fe2+ + ROOH Æ Fe3+ + RO. + OH–
However, both the Harber-Weissand Fenton reactions produce •OH,but the difference is that in theHarber-Weiss reaction, metal ionsare used as catalyst, while in Fentonreactions the metal ions are in-volved in the reaction.
3. Hydroxyl radicals are also formedduring the exposure of high-energyradiations such as X-rays or g-raysto the living tissues. Most of theenergy is absorbed by the cell saphaving very high water content. Itmay result in splitting of one of thecovalent bonds of water.
H-O-H radiationæ Ææææææ Intermediate stepæ Ææææ H. + .OH
(hydrogen radical) (hydroxyl radical)
Generally, free radicals in biologi-cal systems are extremely reactive andunstable. Most of these radicals existonly at a low concentration and they donot move far from their site of forma-tion. Many free radicals are formed bythe reaction of a free radical with anonradical compound, so a chain reac-tion is started that may be several thou-sand events long, for example, lipidperoxidation involving polyunsaturatedfatty acids. The primary free radical pro-duces only local effects at the site of itsproduction but the secondary radicalsderived from it and degradation prod-ucts of primary reaction cause moredamaging effects at sites distinct anddistant from the site where the primaryfree radical was formed. Two free radi-cals after reacting with each other forma stable molecule, which explains the
eventual termination of free radical-in-duced chain reaction. Reactive oxygenspecies (ROS), such as superoxide, H2O2,and the .OH radical, are now known toplay an important role in programmedcell death in both plants and animals(Jobs, 1999; Bethke and Jones, 2001).
Free radicals are capable of reactingwith almost every known molecule ofbiological system in their vicinity. Freeradicals damage proteins, cause break-down of DNA strands, and initiate theperoxidation of various compounds(Rosen et al., 1993). Almost all the vitalcomponents of cells are susceptible todamage by free radicals. The damagecaused by free radicals to different vitalmacromolecules and the mechanism in-volved is summarized in the Table 1.
7,8-dihydro-8-oxoguanine (GO) is acommon oxidative DNA lesion gener-ated by direct modification via reactiveoxygen species. GO lesions are mu-tagenic and can mispair with adenineduring DNA replication (Yang et al.,2001). If the resulting A/GO is not re-paired before the next round of DNAreplication, a C/GÆA/T transversionoccurs and the opportunity for repair, islost. The A/GO is repaired via base ex-cision repair which is initiated by theDNA repair enzyme adenine-DNAglycosylase (Yang et al., 2001).
As a defense against the deleteriousactions of free radicals, cells containseveral enzymes for removing free radi-cals and their products. These are knownas the antioxidant defense system or asfree radical scavengers (Sies, 1991). An-tioxidant means “against oxidation”. An-tioxidants work to protect lipids fromperoxidation by radicals. Antioxidantsare effective because they give up theirown electrons to free radicals and therebybreak the oxidation chain reaction. Someof the important ones are superoxide
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

347
TABLE 1Biological Roles of Oxy Free Radicals
Radical Damaging roles Protectorsspecies
O2¯ • RSH damage
Enzyme activation and inactivation SODFenton reaction
O2¯ • Lipid peroxidation Vit. C and E
Fenton-type reaction b-carotene
.OH Breaking of DNA strand RSH antioxidantsLipid peroxidationSecondary radical peroxidation DNA repair process
RO– Secondary radical peroxidation RSH antioxidants
ROOH Peroxidation GlutathioneFenton reaction peroxidase
H2O2 Peroxidase action CatalaseFenton reaction
dismutase (SOD), catalase, and glu-tathione peroxidase.Superoxide dismutase (SOD): Thisenzyme directly interacts with oxy freeradical to convert it to hydrogen perox-ide.
O2¯ • + 2H+
SODæ Æææ H2O2 + O2
Cytosolic enzyme contains copperand zinc (Cu/Zn-SOD) (Fee, 1982),whereas mitochondrial enzyme containsMn (Mn-SOD) (Weisigner and Fridovich,1973). In plant cells the active oxygenspecies (AOSs) are also known to beproduced under stress conditions. Re-cently, in rice seedling the responses ofSOD genes to environmental stresses,such as drought, salinity, and chilling,were analyzed by Kaminaka et al. (1999).They observed that the expression ofabscisic acid (ABA)-inducible genes, Mn-SOD gene (sodA1), and one of the cyto-solic Cu/Zn-SOD gene (sodCc2) werestrongly induced under the drought andsalinity stresses. While Fe-SOD gene
(sodB) and the other cytosolic Cu/Zn-SOD gene (sodCc1) were also reportedto be induced by ABA. However, themRNA level of sodB was decreased un-der drought stress, and sdCc1 gene didnot induce under drought and stress con-ditions. The plastidic Cu/Zn-SOD gene(sodcp) altered under salinity stresses inthe light but not in the dark. The aboveresults clearly showed that phytohormoneand AOSs are associated with the regula-tion of SOD genes under environmentalstresses (Kaminaka et al., 1999).Catalase: The enzyme catalyses thetransformation of less toxic hydrogenperoxide to water
2 H2O2 Æ 2 H2O + O2
It is a heme-containing enzymepresent in the peroxisomes and the othercell organelles.Glutathione peroxidase: This enzymecatalyzes the oxidation of reduced glu-tathione (GSH) to oxidized form (GSSG)at the expense of H2O2 thereby remov-
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

348
ing toxic H2O2. Furthermore, glutathioneperoxidase also metabolizes lipid hy-droperoxides to hydroxyl fatty acids
H2O2 + 2 GHS glutathione peroxidaseæ ÆææææææGSSG + 2H2
The supply of reduced glutathionefor continuous removal of H2O2 is main-tained by the glutathione reductase
GSSG + 2NADPH glutathione reductaseæ Æææææææ2GSH + 2NADP+
In plants free radicals are known tobe present in vivo in the young rootsenriched with oxygen (Goodman et al.,1986), in the germinating seeds subjectedto dehydration (Senaratna and McKersie,1986), in ungerminated seed and pollen,and also accumulate during artificialaging (Priestley et al., 1985). As a resultof the drought-induced impairment ofthe electron transport system, the super-oxide radical is generated, which dam-ages the membrane in droughted wheat(Halliwell, 1987; Hendry et al., 1989;Price et al., 1989). It was known thatthrough Fenton reaction, the superoxideis reduced to the highly reactive hy-droxyl radical, which causes severeperoxidative damage to lipid membranes(Price and Hendry, 1989; Price et al.,1989). The extracellular sources of acti-vated oxygen could be air pollutants suchas ozone (Mehlhorn et al., 1990;Kanofsky and Sima, 1991) and the radi-cals produced by neighbouring cellsduring the hypersensitive response(Levine et al., 1994).
Recently, Luxford et al. (1999) dem-onstrated conclusively that Histone H1and other protein and amino acid hydro-peroxides can give rise to free radicals,which might result in oxidative damageto associated DNA. The hydroperoxides
can decompose to oxygen- and carbon-centred radicals (detected by electronparamagnetic resonance spectroscopy)on exposure to Cu+ and other transitionmetal ions. These hydroperoxide-derivedradicals react readily with pyrimidineDNA bases and nucleosides to give ad-duct species (i.e., protein-DNA basecross-links). Product analysis has dem-onstrated that radicals from histone H1-hydroperoxides and other protein andamino acid hydroperoxides can alsooxidize both free 2´-deoxyguanosine andintact calf thymus DNA to give themutagenic oxidized base 8-oxo-2´-deoxyguanosine (8-oxo dG). 8-oxo dGis considered to be a precise and sensi-tive biomarker of oxidative DNA dam-age (Liu and Wells, 1995). Bialkowskiand Olinski (1999) studied 8-oxo dGlevels in DNA as a measure of oxidativeDNA damage in Cardamine pratensisplant as affected by different environ-mental parameters such as light, wateraccessibility, and low temperature. Nosignificant difference in 8-oxo dG levelbetween DNA of etiolated and light-exposed plants and between DNA ofdrought and control plants was found.However, cold temperature (10C for 28 h)-treated plants showed an increase of 8-oxo dG in their DNA, indicating oxida-tive damage to plant DNA underlow-temperature conditions.
C. Lipid Peroxidation andDNA Damage
Lipid peroxidation is a highly de-structive process that causes DNA dam-age and alters RNA transport fromnucleus to cytoplasm. It also alters thestructure and function of cellular mem-brane (Agrawal and Kale, 2001). All
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

349
the major classes of biomolecules maybe attacked by the free radicals, butlipids are probably the most suscep-tible. The cell membranes are richsources of polyunsaturated fatty acids(PUFA), which are readily attacked byoxidizing radicals. The oxidative de-struction of PUFAs, known as lipidperoxidation (Tappel, 1973), causes aloss of fluidity and a breakdown of themembrane’s secretory functions andtransmembrane ionic gradient chainreaction (Barber and Berheim, 1967).They can further oxidize PUFA mol-ecules and initiate new chain-produc-ing lipid hydroperoxides (LOOH) thatcan break down to yet more radicalsand to a wide range of compounds, no-tably aldehydes (Esterbauer et al.,1990a). Therefore, lipid peroxidationcan be used as a measure of oxidativedamage.
LH + R. Æ L. + RHL. + O2¯
• Æ LOO.
LOO. + LH Æ LOOH + L.
LOOH Æ LO., LOO., aldehydes
The end products of lipid peroxidationprocesses are aldehydes, hydrocarbongases, and various chemical residues (delMaestero et al., 1981a). These degrada-tion products can diffuse away from thesite of chain reaction and give rise to celledema, influence vascular permeability,and cause inflammation and chemotaxisin animals. These products may also alterthe activity of phospholipids and inducethe release of arachidonic acid with thesubsequent formation of prostaglandins(Hemler and Lands, 1980) and variousendoperoxides (del Maestro et al., 1981a,b).
The O2¯ • can indirectly influence
lipid peroxidation via Fenton/Harber-Weiss reaction.
O2¯ • + Fe3+ Æ O2 + Fe2+
2O2¯ • + 2H+ Æ H2O2 + O2
Fe2+ + H2O2 Æ Fe3+ + OH– + •OHFe2+ + LOOH Æ Fe3+ + OH– + LO•
The generation of O2¯ • by any source,
in the presence of metal ions, particu-larly iron ions, can lead to the formationof •OH radical and in turn may initiatelipid proxidation. However, lipidperoxidation has no intrinsic metal ionrequirement. Nonetheless, iron com-pounds can increase the rate of propaga-tion of lipid peroxidation dramaticallyby decomposing lipid hydroperoxides(LOOH) to reactive alkoxyl (LO.) orperoxyl (LOO.) radicals, which can ini-tiate new reactions (Agrawal and Kale,2001).
Lipid peroxidation causes the en-dogenous lesions by inducing the for-mation of exocylic DNA base adducts(Lindahl and Wood, 1999). A majorrecent advancement has been the de-tection of number of lesions in DNAthat can be attributed to metabolicsources (Fink et al., 1997). Lipidperoxidation, for example, leads to theformation of a complex variety of nu-cleophiles capable of reacting with pro-teins and DNA (reviewed by Marnett,1999a,b). The reaction of nucleophileswith DNA leads to formation of mu-tagenic adducts. The lipid peroxidationproduct malondialdehyde (MDA) re-acts with G residue in DNA to form apyridimidopurinone called M1G, anexocyclic adduct that can block theWatson-Crick base pairing of DNAbases and thus have the potential toblock DNA replication (Marnett,1999b). Site-directed mutagenesis stud-ies have shown that M1G is an efficientpremutagenic lesion (Fink et al., 1997).The M1G adduct is not very stable andcan decompose to a secondary ring-
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

350
opened derivative. In addition to MDA,lipid peroxidation can also produce ac-rolein and crotonaldehyde, which arereadily metabolized to epoxides. Thisresulted in the generation of exocylicetheno modifications of DNA bases(Lindahl and Wood, 1999). Two suchbases, etheno-A and etheno-C, are re-moved by DNA glycosylases (Hang etal., 1997; Saparbaev and Laval, 1998).These strongly suggest that the genera-tion of such adducts occurs at high rate(Lindahl and Wood, 1999).
Several leading laboratories aroundthe world are currently addressing theidentification of endogenous DNA le-sions in mammalian DNA and their rolein genetic disease. As lipid peroxidationand DNA damage individually have beenconsidered major determinants of seedviability loss; however, no linkage be-tween lipid peroxidation product andDNA damage in seed deterioration hasbeen contemplated so far.
D. Ozone and DNA Damage
Ozone (O3) itself is not a free radi-cal, but the damage caused by it is oftenmediated by free radical production andthereby can stimulate lipid peroxidation.In last 40 years, the trophospheric O3
concentration has increased 2- to 5-fold(Kley et al., 1999). Exposure of animalsto O3 is known to damage lungs (Mustafaand Tiernery, 1978). However, O3 posesa 2-fold challenge to plants: photosyn-thesis and growth can be impaired and itcan also produce foliar lesions in sensi-tive species and cultivars (Kley et al.,1999). O3 has also been regarded as a“wound stress” that causes necrosis byoxidizing and damaging plasma mem-branes (reviewed in Heath and Taylor,
1997). O3 responses are known to re-semble components of the hypersensi-tive response (HR) found in incompat-ible plant-pathogen interactions (Sharmaand Davis, 1997; Sandermann et al.,1998). This resemblance is most likelyrelated to the occurrence of relativeoxygen species (ROS), such as superox-ide anion radicals (O2¯
• ) and H2O2 in theapoplast, which trigger an oxidative burstin the affected cells (Pellinen et al., 1999;Rao and Davis, 1999). H2O2 has beenreported to be accumulated in responseto O3 in tobacco (Schraudner et al., 1998)and birch (Pellinen et al., 1999). O3 stressin Arabidopsis is reported to be respon-sible for cell death (Rao and Davis,1999).
O3 also activates ethylene and sali-cylate (SA) signal transduction pathwaysleading to downstream responses, suchas antioxidant and antimicrobial defenses(Sharma and Davis, 1997; Sandermannet al., 1998). The toxicity of O3 is throughchemical reaction with ethylene, yield-ing toxic products that initiate aself-propagating lipid peroxidation(Mehlhorn and Wellburn, 1987). Theemission of ethylene is a kind of wound-ing symptom in O3-stressed plants (Heathand Taylor, 1997). In pea carpel senes-cence, maize endosperm development,as well as in root aerenchyma forma-tion, ethylene is reported to regulate theprogrammed cell death (Orzaez andGranell, 1997; Young et al., 1997; Drewet al., 2000). Recently, Overmyer et al.(2000) have isolated an Arabidopsismutant, rcd1 (for radical-induced celldeath 1), which displayed O3– and O2¯
•–inducible lesion formation. These au-thors proposed a model for the relativecontribution of the different signalingpathways to ROS-driven lession propa-gation and the process of lesion contain-ment.
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

351
E. DNA Damage Response
Most organisms respond to chromo-somal insults by activating a complexdamage response pathway that regulatessome known responses such as cell-cyclearrest, apoptosis, and direct activation ofrepair networks (Zhou and Elledge, 2000).DNA damage checkpoints control theability of cells to arrest the cell cycle inresponse to DNA damage. The DNAdamage response pathway is a kind ofsignal transduction pathway consistingof sensor, transducers, and effectors. Theidentities of sensors are not yet known,but the DNA-break binding protein suchas poly (ADP-ribose) polymerase (PARP)and DNA-dependent protein kinase(DNA-PK) are the kind of DNA damagesensors. Much is known about signaltransducers, which are composed of foursets of conserved proteins with some rec-ognizable motifs. These four classes are
1. Phospho-inositide kinase (PIK)-re-lated proteins, which include ATMand ATM-Rad3-related (ATR) inmammals and their homologs inyeast (Mec1, Rad3, Tel1). Down-stream of these proteins are twocheck point kinases (chk1 andchk2).
2. BRCT-repeat containing proteins,which include budding yeast Rad9and fission yeast Crb2.
3. RFC-like proteins, for example,Rad17, RFC2.5, in mammals andRad24, Rad17, RFC2-5 and RFC3in yeasts.
4. PCNA-like proteins, for example,Rad1, Rad9, Hus1 in mammals andRad17, Rad1, Ddc1, Rad9, Mec3,and Hus1 in yeasts.
The effectors of damage responsecould be either activation of DNA repair
pathways or cell cycle arrest or apoptosisdepending on the severity and type ofthe damage (Zhou and Elledge, 2000).
III. VARIOUS DNA REPAIRMECHANISMS
For more than 2.5 billion years sincethe first appearance of living cells on Earth,they were continuously exposed to solarUV radiation and many DNA damagingagents. To protect the DNA from variousdamage, an intricate network of DNA re-pair mechanisms has evolved early in evo-lution (de Laat et al., 1999). Several DNArepair pathways known to date in differentliving cells are shown in Figure 2. Out ofthese, the nucleotide excision repair (NER)has been studied extensively. Althoughthe DNA repair mechanisms have beendescribed thoroughly in E. coli, Saccharo-myces cerevisiae, rodents, and human, littleis known still about these processes inplants. Any alteration in the DNA doublehelix, even as small is single base damage,are easily recognized. Such recognition isusually by a change in the physical struc-ture of the DNA double helix. The repairmechanisms protect the genome by re-pairing modified bases, DNA adducts,crosslinks, and strand breaks. Furthermore,these repair mechanisms are also coordi-nated with other cellular functions, in par-ticular gene transcription and cell cycle.The various basic repair mechanisms bywhich damages are eliminated from DNAare described below.
A. Direct Reversal (DR)
DR is a simple mechanism that in-volves a single-enzyme reaction for the
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

352
FIGURE 2. Several DNA repair pathways used by different livingcells. MGMT = methylguanin methyltransferase; DR = direct rever-sal; BER =base excision repair; NER =nucleotide excision repair;SSBR =single strand break repair; DSBR = double-strand breakrepair; MMR = mismatch repair; PR = photoreactivation.
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

353
removal of certain types of DNA dam-age. Alkyltransferases simply extract thealkyl group from alkylated bases that istransferred to an internal cysteine resi-due, and thus inactivate themselves (Teoet al., 1984). The best example for DR isthe correction of the miscoding alkyla-tion lesion O6-methylguanine, which isgenerated endogenously in small amountsby reactive cellular catabolites (Vaughanet al., 1993). DR is carried out by a spe-cific enzyme, called methylguanine-methyltransferase (MGMT), which re-moves the methyl group from the guanineresidue of DNA, and transfers it to one ofits own cysteine residues (Figure 2) in arapid and error-free repair process (Mooreet al., 1994). O6-methyl-guanine can pairwith both C and T and thereby causetransition mutations, which are sometimecorrected by mismatch repair mechanism(Lindahl and Wood, 1999).
Photolyases, on the other hand, re-vert UV-induced dimers in a light-de-pendent reaction called photo-reactiva-tion (PR) (Sancar, 1990; Yasui and Eeker1998; Todo, 1999), which is discussedlater.
B. Base Excision Repair(BER)
The BER mechanism is known forthe elimination of single damaged baseresidues in DNA and is considered to bean essential pathway for DNA mainte-nance. BER mainly removes the DNAdamages that are arising spontaneouslyin a cell from hydrolytic events such asdeamination or base loss, fragmentedbases resulting from ionizing radiation,and oxidative damages or methylationof ring nitrogens by endogenous agents.BER pathways involve several steps,
which include recognition and removalof damaged base, incision, gap filling,and sealing (Britt, 1996; Lehman, 1998;Lindahl and Wood, 1999). A model ofvarious steps in BER is shown in Figure3. The first step in BER involves theremoval of a single damaged basethrough the action of a specific DNAN-glycosylase, which hydrolyticallycleaves the base-deoxyribose glycosylbond of a damaged nucleotide residue,and leaves the sugar-phosphate back-bone intact. The resulting abasic sitesare then recognized by an apurinic/apyrimidinic (AP) endonuclease thatcreates an incision on the backbone ofthe DNA at the 5¢ end of AP site (Sakumiand Sekiguchi, 1990). The completionof base excision requires removal of the5 terminal deoxyribose-phosphate resi-due, which is catalyzed by the phos-phodiesterase (AP lyase) activity ofDNA polymerase b (Pol b) (Matsumotoand Kim, 1995). The resulting one-nucle-otide gap is filled by DNA Pol b andsealed by either DNA ligase I or DNAligase III with its accessory proteinXRCC1 (Figure 3) in mammalian sys-tem (Lehman, 1998).
The BER pathway has been shownpreviously to depend strongly on thepresence of nicotinamide adenine di-nucleotide (NAD+). This dependencywas proposed to be mediated throughthe catalytic activation of the nuclearenzyme poly(ADP-ribose) polymerase-1 (PARP-1) following its transient bind-ing to BER-induced DNA strand break(Satoh and Lindahl, 1992; Satoh et al.,1993). PARP-1 is an abundant nuclearenzyme found in many eukaryotes, withthe exception of yeast. This enzyme hashigh affinity for single- and double-strand DNA breaks. After binding toDNA strand breaks, PARP-1 catalyzesextensive synthesis of poly(ADP-ribose)
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

354
from NAD+ and covalently modifiesmany nuclear proteins, including itself.The massive automodification of PARP-1effects its dissociation from DNA strandbreaks and inhibition of its catalyticactivity. A modulatory role for poly-(ADP-ribose) formation and PARP-1automodification in BER has been pro-posed by Satoh and Lindahl (1992).According to this model, the unmodi-fied enzyme binds tightly to DNA strand
interruptions formed either by ionizingradiation or through incision of AP sitesduring BER and interferes with the BERprocess because the bound PARP-1 mol-ecule hinders access of the repair ma-chinery to the lesion. Automodificationand release of PARP-1 from the breaksallow the repair process to proceed. Inthe absence of NAD+ or after the inhibi-tion of poly(ADP-ribose) synthesis,PARP-1 persists on DNA breaks and
FIGURE 3. A model for base excision repair pathway. AP endonuclease = apurinic/apyrimidinic endonuclease; POL.b = polymerase b; PCNA = proliferating cell nuclearantigen; FEN-1 = flap endonuclease-1
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

355
DNA repair is abrogated. Recently, ithas been shown that the cells lackingPARP-1 have normal capacity to repairsingle-strand breaks inflicted by x-irra-diation or breaks formed during the re-pair of modified bases (Vodenicharov etal., 2000).
Different DNA glycosylases (at leasteight) are known to be present in humancell nuclei (Lindahl and Wood, 1999).These glycosylases remove different kindsof damage, and the specificity of repairpathway is determined by the type ofglycosylase involved (Seeberg et al., 1995).The mode of action of these glycosylaseand their three-dimensional structures havebeen described earlier (Cunnigham, 1997;Krokan et al., 1997; Parikh et al., 1998).These enzymes have a catalytic domain of~250 amino acid residues and for addi-tional interactions they also use theiramino- and carboxy-terminal region(Lindahl and Wood, 1999). In general,these DNA glycosylases move along theminor groove of DNA until a specific typeof damaged nucleotide is recognized. Thisenzyme then binds to the backbone of adamaged strand and slides out the dam-aged nucleoside residue to accommodatethe damaged base in a specific recognitionpocket and mediates cleavage (Parikh etal., 1998). Although these enzymes areessential for BER, in mice the knockoutsof various DNA glycosylases have beenshown to be viable (Wilson and Thomp-son, 1997), because the abasic sites arealso known to be generated by nonenzy-matic depurination.
Structurally, the AP endonucleasebelongs to the superfamily of nucleasesthat also contains pancreatic DNaseI(Gorman et al., 1997). This enzyme flipsout the base free deoxyribose residuefrom the DNA before chain cleavageand also recruits the Pol b to the site ofrepair (Bennett et al., 1997). The Pol b
contains two distinct domains, the largerdomain is the polymerase domain, whichhelps in gap filling and the small basicamino-terminal domain that contain APlyase (Phosphodiesterase) activity thatexcises the abasic sugar-phosphate resi-due at the strand break (Matsumoto andKim, 1995; Sobol et al., 1996). A knock-out mutation of Pol b in mice has beenshown to cause embyronic lethality (Guet al., 1994). This finding suggests thateither the single-patch mode of BER isessential for maintaining normal viabil-ity or that Pol b has an additional role inthe cells such as in chromosomal DNAreplication. The XRCC1 plays impor-tant role in bringing the Pol b and ligaseIII together, and it is also known to di-rectly bind to the DNA single-strandbreak (Kubota et al., 1996; Marintchevet al., 1999).
Sometimes longer repair patches of2 to 10 residues have also been observed(Matsumoto et al., 1994; Frosina et al.,1996). This might happen when the ter-minal sugar-phosphate residue has amore complex structure, which is resis-tant to cleavage by AP lyase, then theDNA strand displacement may occur.These longer repair tracts are thought toresult from a nick translation reactionaccompanied by strand displacement inthe 5¢ to 3¢ direction, thereby generatinga flap type of structure. The produceddisplacement flap structure is removedby flap endonuclease FEN-1 with thehelp of PCNA (Figure 3) (Harringtonand Lieber 1994; Wu et al., 1996;Klungland and Lindahl, 1997). In thiscase the gap of few nucleotides long isfilled by either Pol b or Pol d (Fortini etal., 1998; Dianov et al., 1999) and sealedby DNA ligase I (Figure 3).
The BER in plants still has not beenwell studied when compared with mam-malian systems, but this mechanism does
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

356
exist in plant (Talpaert-Borle and Liuzzi,1982). During the early germination ofZea mays, the formation of AP site hasbeen reported (Dandoy et al., 1987),which also suggested that plants do haveBER pathway. A specific glycosylaseenzyme (uracil-DNA glycosylase) wasfound in cultured cells of carrot thatplay a role in BER (Talpaert-Borle andLiuzzi, 1982). A cDNA clone encoding3-methyl adenine glycosylase was re-ported from Arabidopsis and the encodedprotein shown to contain glycosylase ac-tivity (Santerre and Britt, 1994). Thisgene was shown to be expressed in grow-ing tissue in Arabidopsis (Shi et al.,1997). The gene encoding DNA ligasehas also been cloned from Arabidopsisand sequence data are available inGenebank (Taylor et al., 1996b).
C. Nucleotide Excision Repair(NER)
NER is one of the most versatileDNA repair pathways operating in bothprokaryotes and eukaryotes. Unlike otherDNA repair pathways that are repair spe-cific, NER is capable of removing vari-ous classes of DNA damage, includingthose induced by UV radiation (pyrimi-dine dimers) and chemicals (bulky DNAadducts) such as cisplatin, 4 NQO, ben-zpyrene, and alfatoxin (Lindahl andWood, 1999; Balajee and Bohr, 2000).
The major difference between NERand BER is the way the DNA damage isremoved. Basically, NER cuts out thedamage as a part of an oligonucleotidefragment, while BER excises only onenucleotide. From 24 to 32 nucleotidescan be removed by NER in eukaryote(Huang et al., 1992; Moggs et al., 1996).The repair of lesions over the entire
genome is called as global genome NER,while the repair of transcription blockedlesions present in transcribed DNAstrands is called as transcription coupled-NER (TC-NER) (Mu and Sancar, 1997).
NER pathway in mammalian sys-tem involves product of at least 30 genesand most of them have been cloned (deLaat et al., 1999). In bacteria, mainlyfour proteins (UvrA, B, C, and D) carryout NER process. The UvrA, B, and Care required for incision reaction. TheUvrA first dimerizes and then binds toUvrB, which recognize the damage onthe DNA and causes local melting at 3¢to the damage (Mazur and Grossman,1991; Myles and Sancar, 1991). UvrBthen makes a 3¢ incision (Lin et al., 1992),preceding the 5¢ incision by UvrC (Linand Sancar, 1992). Incision is made atseventh nucleotide from 3¢ side and atfourth or fifth positions form 5¢ side ofthe damaged DNA (Sancar and Rupp,1983). The UvrD helicase releases ~12or ~13 bases long oligonucleotide contain-ing the damage, DNA polymerase I fills thegap, and finally DNA ligase seals the repairpatch. The NER processes in yeast andmammalian cells are very much alike. Themolecular mechanisms of NER in a mam-malian system have been described in de-tail in several reviews (Lehman, 1998; deLaat et al., 1999; Lindahl and Wood, 1999;Balajee and Bohr, 2000).
Most of the NER genes have beenisolated through transfection of repair-deficient rodent mutant cell lines and therecovery of the excision repair crosscomplementing (ERCC) human genes.Nearly all ERCC proteins subsequentlyappeared to be involved in the humanrepair disorders Xeroderma pigmentosum(XP) and Cockayne syndrome (CS) andtherefore are called XP and CS factors,respectively. The XPA, XPB, XPC, XPD,XPF, and XPG have been cloned from
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.
357
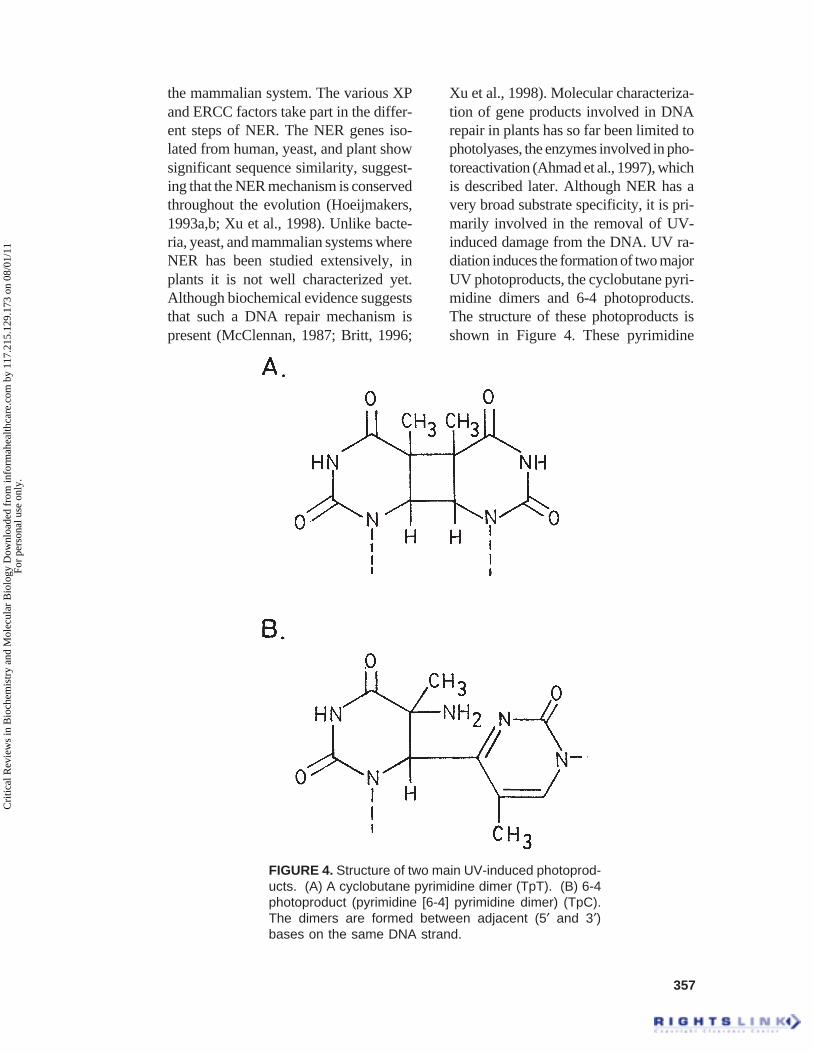
the mammalian system. The various XPand ERCC factors take part in the differ-ent steps of NER. The NER genes iso-lated from human, yeast, and plant showsignificant sequence similarity, suggest-ing that the NER mechanism is conservedthroughout the evolution (Hoeijmakers,1993a,b; Xu et al., 1998). Unlike bacte-ria, yeast, and mammalian systems whereNER has been studied extensively, inplants it is not well characterized yet.Although biochemical evidence suggeststhat such a DNA repair mechanism ispresent (McClennan, 1987; Britt, 1996;
Xu et al., 1998). Molecular characteriza-tion of gene products involved in DNArepair in plants has so far been limited tophotolyases, the enzymes involved in pho-toreactivation (Ahmad et al., 1997), whichis described later. Although NER has avery broad substrate specificity, it is pri-marily involved in the removal of UV-induced damage from the DNA. UV ra-diation induces the formation of two majorUV photoproducts, the cyclobutane pyri-midine dimers and 6-4 photoproducts.The structure of these photoproducts isshown in Figure 4. These pyrimidine
FIGURE 4. Structure of two main UV-induced photoprod-ucts. (A) A cyclobutane pyrimidine dimer (TpT). (B) 6-4photoproduct (pyrimidine [6-4] pyrimidine dimer) (TpC).The dimers are formed between adjacent (5¢ and 3¢)bases on the same DNA strand.
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

358
dimers are highly toxic because they haveeffect on transcription. Their efficientremoval is an essential function for anyliving cells that is exposed to sunlight.The cyclobutane pyrimidine dimers canbe formed between any two adjacent py-rimidines (Figure 4A) (Protic-Sabljic etal., 1986; Doetsch, 1995). The 6-4 photo-products are formed by covalent bondbetween the carbon 6 and carbon 4 ofadjacent pyrimidines (Figure 4B). Theseare the most frequent occurring UV pho-toproducts and occur at 5¢-T-C-3’ (Fig-ure 4B), 5¢-C-C-3¢, and 5¢-T-T-3’ but notat 5¢-C-T-3¢ sites in DNA (Doetsch,1995). These photoproducts can be re-paired by NER; enzymatic photoreacti-vation, recombination repairs, and bypostreplications repair (Friedberg et al.,1995).
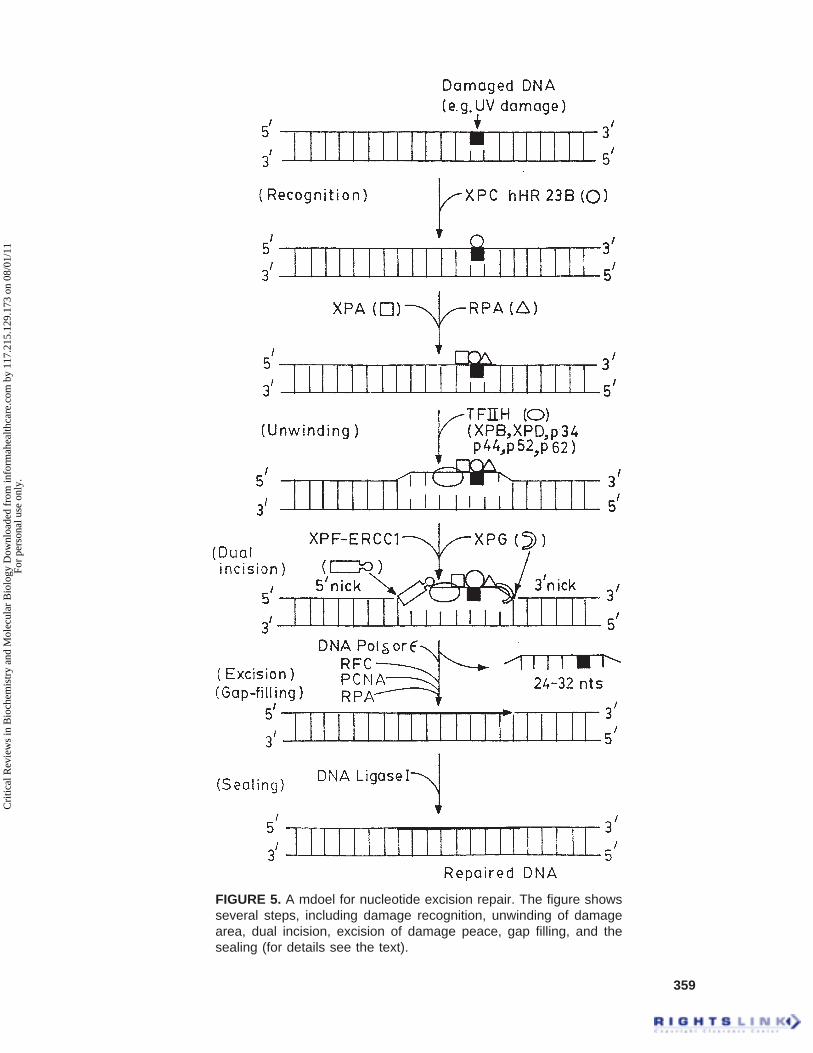
The molecular model for NER inmammalian system is shown in Figure 5.The NER sequentially involves recogni-tion of DNA damage, dual incision ondamaged strand, excision of damagecontaining oligonucleotide, DNA syn-thesis (gap-filling), and ligation to re-place an excised oligonucleotide (Fig-ure 5) (Wood, 1997; de Laat et al., 1999).The initiation of DNA repair is the firststep in the NER. The XPC recognizesthe damage and binds strongly to thedamage and then recruits the entire re-pair protein apparatus to the damage(Drapkin et al., 1994; Masutani et al.,1994; Reardon et al., 1996; Sugasawa etal., 1998). The XPC protein (125 kDa)exists in a complex with hHR23B (58kDa), a human homolog of the Saccha-romyces cerevisiae repair protein Rad23that enhances XPC-dependent excisionrepair in vitro (Sugasawa et al., 1998).The DNA damage is also recognized bythe XPA protein in association with rep-lication protein A (RPA) and forms aXPA-XPC-RPA complex on the dam-
age (Figure 5) (Matsuda et al., 1995;Saijo et al., 1996). The RPA is aheterotrimeric single-stranded DNA-binding protein that is required for boththe incision reaction as well as repairsynthesis (Coverley et al., 1992;Aboussekhra et al., 1995; He et al., 1995;Mu et al., 1995). RPA also binds to thenondamaged strand to prevent it fromincision of the strand (which serves as atemplate for repair synthesis) and stabi-lizes the open structure (Kim et al., 1992;Seroussi and Lavi, 1993). RPA is alsoknown to stimulate the interaction ofXPA and ERCC1 (Li et al., 1994; Parkand Sancar, 1994; Saijo et al., 1996),and on the other hand can bind to theendonuclease XPG (He et al., 1995) andpossible XPF (Bessho et al., 1997). TheXPA binds to basal transcription factor,TFIIH, and thereby recruits it to thedamage area (Figure 5). The TFIIH is a6-9 subunit complex containing XPB(ERCC3) and XPD (ERCC2) DNAhelicases (Winkler et al., 1998; de Laatet al., 1999; Lindahl and Wood, 1999).
The action of TFIIH complex re-sulted in the formation of a open du-plex structure around the damaged sitethat enables the structure-specific nu-cleases (XPG and ERCC1-XPF) for thedual incision (Evans et al., 1997; Mu etal., 1997a,b; de Laat et al., 1999;Lindahl and Wood, 1999). TFIIH andRPA can both bind to XPG, whichmakes the first incision while it cutsthe damaged DNA strand on the 3¢ sideof the damage (Figure 5) (O’Donovanet al., 1994; Matsunaga et al., 1995; Muet al., 1996). The incision by XPG ismade at 5 to 6 phosphodiester bondsaway from the 3¢ side of the damage(Figure 5) (Svoboda et al., 1993). TheXPG protein is a member of the FEN-1family of structure-specific endonu-cleases and cuts at the junctions of du-
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.
359
FIGURE 5. A mdoel for nucleotide excision repair. The figure showsseveral steps, including damage recognition, unwinding of damagearea, dual incision, excision of damage peace, gap filling, and thesealing (for details see the text).
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

360
plex and unpaired DNA (Lieber, 1997;de Laat et al., 1999). XPG is also knownto interact with proliferating cell nuclearantigen (PCNA) (Gary et al., 1997),which is involved in DNA repair syn-thesis but dispensable for the incisionstage of NER (Shivji et al., 1992). Next,the ERCC1-XPF complex makes the in-cision at 22 to 24 phosphodiester bondsaway from the 5¢ side of the damage(Figure 5) (Svoboda et al., 1993; Mu etal., 1996). The dual incision is abso-lutely dependent on ATP hydrolysis(Svoboda et al., 1993). The specificitiesof these two nucleases make them ide-ally suited to cleave the opened up dam-aged site on either side of the damage.This results in the removal of a 24- to32-residue oligonucleotides in length(Figure 5) (Huang et al., 1992; Moggs etal., 1996; Mu et al., 1997a,b; de Laat etal., 1999).
The final step in the NER pathwayis the gap-filling or repair synthesis ofthe excised patch, which is performedby common DNA replication factors.An in vitro reconstituted repair reactionshowed that efficient repair synthesisrequired the mammalian replication fac-tors RPA, RFC, PCNA, and DNA poly-merase d or e (Hubscher and Thommes,1992; Shivji et al., 1995). RFC is neededto load PCNA onto the DNA (Podust etal., 1994), and PCNA is required for theinitiation of repair synthesis. PCNA alsostimulates polymerase activity. Thenewly synthesized repair patch, whichexactly matches the excision patch, isthen sealed by ATP-dependent DNA li-gase, most likely ligase I (Figure 5)(Lindahl and Barnes, 1992; Petrini etal., 1995).
In plant, the NER system is still notwell characterized, although biochemicalevidence suggests that such a DNA re-pair mechanism is present (McCleannan,
1987; Britt, 1996; Vonarx et al., 1998;Britt, 1999). An UV-specific endonu-clease resembling UVrABC nuclease,which is known to be involved in NERin bacteria, was reported and character-ized from spinach (Doetsch et al., 1989).Molecular characterization of gene prod-ucts involved in DNA repair in plants sofar has been limited to photolyases(Ahmad et al., 1997). The UV productsCPDs and (6-4) photoproducts are alsoknown to be repaired in plant through alight-independent repair pathway thatsupports the existence of a pathwayequivalent to NER mechanism in yeastand mammal (Howland, 1975; Quaite etal., 1994; Taylor et al., 1996). NER wasalso suggested to occur in carrot proto-plasts (Howland, 1975, Eastwood andMcLennan, 1985), Arabidopsis and al-falfa seedlings (Pang and Hays, 1991;Quaite et al., 1994), soybean chloroplastsand leaves (Cannon et al., 1995;Sutherland et al., 1996), and in differentrice cultivars (Hidema et al., 1997). Itwas also reported in wheat leaf tissue(Taylor et al., 1996). The Rad23 gene ofyeast is involved in NER and transcrip-tion coupled repair (Mueller andSmerdon, 1996). Mutation of the yeastRad23 gene resulted in moderate UVsensitivity that was due to the defect inNER pathway (Verhage et al., 1996). In1997 a structural homolog of Rad23,OSRad23, was reported from rice but itsrole in NER has not been determined(Shultz and Quatrano, 1997). Severalputative Rad23 homolog in ArabidopsisEST databases have been observed.
Until recently, the only evidence ofthe existence of light-independent DNArepair pathways in plants came fromphysiological studies based on dark re-pair of UV-induced pyrimidine dimers(Eastwood and McLennan, 1985; Quaiteet al., 1994; Taylor et al., 1996). In ad-
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

361
dition, UV-sensitive Arabidopsis mu-tants defective in dark repair have beendescribed (Britt et al., 1993; Harlow etal., 1994; Jiang et al., 1997). Recently, afew plant homologs of some of the com-ponents involved in mammalian NERpathway have been reported. The mo-lecular evidences for the conservationof the NER pathway came from the clon-ing of Arabidopsis XPB (araXPB)(Ribeiro et al., 1998) and the lily ho-molog of human ERCC1 (Xu et al.,1998). The araXPB protein shared 50%identity and 70% conserved amino ac-ids with yeast and human homologs.The plant XPB contained all the func-tional domains found in the other pro-teins, including nuclear localization sig-nal, DNA-binding domain, and helicasemotifs, suggesting that it might be play-ing a role in NER in plant cells (Ribeiroet al., 1998). However, the DNA un-winding activity of plant XPB has notbeen determined yet.
The plant ERCC1 gene was isolatedfrom male germline cells of lily, and thededuced amino acid sequence containedstriking homology with human ERCC1and yeast Rad10 (Xu et al., 1998). Thelily ERCC1 homologs were present inthe genomes of taxonomically diverseplant species, including Arabidopsisthaliana, Brassica napus, Zea mays,Oriza sativa, Nicotiana tobacum, andLycopersicon esculentum, as shown bySouthern blot analysis of genomic DNAs(Xu et al., 1998). These findings sug-gested that the NER role of the ERCC1gene might be conserved in higher plants.The lily ERCC1 expression was alsoreported to be upregulated in the malegermline cells of plants (Xu et al., 1998).Pollen, as part of its developmental pro-cess, is exposed to solar UV radiationand other environmental mutagens afterbeing released from the anther. This ex-
posure could inevitably lead to DNAdamage in both vegetative and genera-tive cell nuclei of pollen. The up-regula-tion of ERCC1 homolog in lily genera-tive cells suggested that a highly activeDNA repair mechanism existed in malegermline cells in order to protectgermline DNA from heritable mutationsresulting from damaged DNA (Xu et al.,1998). The protein encoded by the lilyERCC1 gene was also able to correct thesensitivity to the cross-linking agentmitomycin C in ERCC1-deficient Chi-nese hamster over cells that further sug-gested that the NER mechanism is con-served in yeast, animals, and higherplants (Xu et al., 1998).
Recently, two different groups havereported the cloning of plant (Arabidopsis)homolog of human XPF and S. cerevisiaeRAD1 and they gave the name of the geneas UVH1, Rad1 (Liu et al., 2000), andAtRAD1 (Gallego et al., 2000). The UVH1/Rad1 gene was cloned from Arabidopsisby positional cloning methods and hasbeen mapped to chromosome 5 and alsoshown to be the product of a repair endo-nuclease (Liu et al., 2000). The UVH1/Rad1 gene was shown to be strongly ex-pressed in flower tissue as well as in othertissues (Liu et al., 2000). The AtRAD1gene from the other group is also shown tobe involved in NER in the dark and is alsoshown to be ubiquitously expressed, whichsuggested that plant NER has some otherroles (Gallego et al., 2000). Recently, theXPF gene was knocked out in tobacco byintroducing AtXPF antisense in tobacco,which resulted in the increased UV sensi-tivity (Crockett et al., unpublished data).The transgenic tobacco plant carrying onAtXPF promoter GUS gene fusion showedrelatively higher levels of GUS expres-sion in generative cells of pollen and seeds.These findings suggested a key role ofNER pathway in maintaining germline
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

362
DNA integrity and also in the removal ofendogenous DNA lesions in seed, whichaccumulated during the seed aging pro-cess (Crockett et al., unpublished data).
D. Photoreactivation (PR) andPhotolyases
It is clear now that UV radiation candamage plants, decreasing growth, andproductivity (Teramura, 1983). The UV-induced DNA damages (CPDs and 6-4pps) are removed or repaired by twodifferent repair pathways. One is NER,which is described above, and other isPR which is a direct reversal phenom-enon. PR is performed by the combinedaction of one or more proteins termed‘photolyases’ and visible light in an er-ror-free fashion (Sancar, 1994; Britt,1999; Thoma, 1999). The photolyasesspecifically recognize and bind to thepyrimidine dimers form a complex thatis stable in the absence of light. Afterabsorbing a blue light photon, the py-rimidine dimers are reversed to pyri-midine monomers without excision ofthe damaged bases (Sancar, 1996;Thoma, 1999) as shown in Figure 6.Photorepair of CPDs have been re-ported in several plant species, includ-ing gingko (Trosko and Mansour,1969), Arabidopsis (Pang and Hays,1991; Britt et al., 1993), alfalfa (Quaiteet al., 1994), soybean (Sutherland etal., 1996), cucumber (Takeuchi et al.,1996), rice (Hidema et al., 1997,Hidema et al., 2000), maize (Stapleton,1992; Stapleton et al.,1997), and wheat(Taylor et al., 1996a).
Photolyases are known to be very spe-cialized in terms of their substrate speci-ficity (Britt, 1999). CPD photolyases werereported in bacteria, fungi, plants, inverte-
brates, and many vertebrates, while 6-4photolyases were identified in silkworm,frog, fly, and rattlesnakes (Yasui et al.,1994, Sancar, 1996; Todo, 1999). The firsthigher eukaryotic photolyase was CPDspecific and cloned from goldfish (Britt,1999). Two distinct photolyases, one spe-cific for CPDs and other specific for 6-4products, were reported from Arabidopsisseedling (Chen et al., 1994). The twoclasses of CPD photolyases share only 10to 15% sequence identity (Yasui et al.,1994). Class I is a microbial CPDphotolyase and Class II is metazoan CPDphotolyase. The Arabidopsis homolog ofclass II photolyase sequence (PHR1) cor-responds to a gene (UVR2), which wasidentified via classic genetic analysis(Ahmad et al., 1997; Jiang et al., 1997).The phylogenetic analysis of photolyasehomologs are described in Nakajima et al.(1998) and Britt (1999). Photolyases con-tain two prosthetic chromophores: FADH2
and either methenyl tetrahydrofolate or8-hydroxy-5-deazaflavin (Sancar, 1994).The second chromophore works as a light-harvesting antenna. The wavelength oflight for photoreactivation ranges from thevisible (500 nm) to the UV-B (300 nm).The existence of PR in plant was firstreported by Ikenaga and Mabuchi (1966),who demonstrated that the frequency ofendosperm mutations generated by UVradiation of maize pollen dropped sub-stantially if the pollen was exposed to vis-ible light after UV irradiation. A photolyasecDNA was reported earlier from whitemustard (Batschauer, 1993). The crystalstructures of CPD photolyase of E. coliand Anacystis nidulans have been resolved.This suggested that the photolyases flipthe pyrimidine dimer out of the duplexinto the hole that contains the catalyticcofactor (Park et al., 1995; Tamada et al.,1997) followed by splitting the cyclobutanering after light absorption.
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

363
The plant blue light photoreceptorgenes were discovered during thephotolyase research and named CRY af-ter cryptochrome. The CRY of plant andanimal have some sequence homologywith DNA photolyase (Todo, 1999), butthey do not contain any repair activity.In yeast it is known that repair byphotolyase is faster than NER especiallyin nucleosome-free regions, for example,promoters and origin of replication re-
gions (Thoma, 1999). The photolyaseand NER can also play complementaryroles in the genes that are transcribedby RNA polymerase II; they removethe damage in the nontranscribedand transcribed strands, respectively(Livingstone-Zatchej et al., 1997; Suteret al., 1997; Thoma, 1999). Aboussekhraand Thoma (1999) have shown that NERcan remove CPDs, which are not acces-sible to photolayase, such as CPDs gen-
FIGURE 6. Schematic illustration of photoreactivation. Photolyase bindsto pyrimidine dimer (T = T), split the dimer and restore it to nativepyrimidine monomer in a light-dependent reaction.
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

364
erated in the TATA-box of the yeastSNR6 gene.
Increased sensitivity to UV radia-tion may result from failure to repairphotodamage in DNA. The UVradiation-sensitive UVR1 mutant ofArabidopsis cannot photorepair 6-4 pho-toproducts (Britt et al., 1993). Landryet al. (1997) showed that anotherArabidopsis UV radiation-sensitivemutant, UVR2-1, was deficient inphotorepair of CPDs and cloning andsequencing of the photolyase genes fromUV radiation-sensitive and wild-typestrains indicated alterations of DNA se-quence that led to defective photolyaseprotein (Ahmad et al., 1997). These datasuggested that such deficiencies mightbe ameliorated by restoration of photo-lyase function in photorepair-deficientstrains. However, for such restorationexact knowledge of the molecular ori-gin of their photorepair deficiency isessential, as suggested by Hidema et al.(2000). It is important to know whetherthe deficiency is in the photolyase pro-tein itself, or it has resulted from othercause, such as a regulatory mutation thatcauses the production of fewer normalphotolyase molecules per cell. TheUV-B radiation-sensitive rice cultivarNorin 1, a progenitor of many Japanesecommercial rice strain, has been reportedto be deficient in photorepair of CPDS(Hidema et al., 1997). Recently, the samegroup, Hidema et al. (2000), reportedthat the photorepair deficiency in Norin1 is due to the alteration in the structureof the photolyase enzyme rather than aregulatory mutation. Their findings in-dicated that a strategy of increasing theactivity of photolyase through selectivebreeding or engineering could increaseresistance to UV radiation and perhapsthe productivity of such cultivars(Hidema et al., 2000).
E. Bypass Damage Repairand DNA Polymerases
It is known that the pyrimidinedimers can act as blocks to DNA repli-cation, so cells have to possess special-ized low fidelity and often error-proneDNA polymerases that can bypass suchlesions and promote replication of dam-aged DNA (Johnson et al., 1999c). Usu-ally at cell-cycle checkpoints, these dam-ages are repaired before replication orcell division proceeds. If the damage isnot repaired properly, then the cell cyclecheckpoints can lead to the inhibition ofcell cycle progression (Costanzo et al.,2000). In yeast S. cerevisiae the cata-lytic subunit (Rev3) of DNA polymerasez together with an accessory factor Rev7can bypass pyrimidine dimers and otheradducts in DNA (Lawrence and Hinkle,1996). Some other polymerases, for ex-ample, yeast, Rev1 (hREV1 in human),E. coli UmuD’2C (E. coli Pol V), E. coliDinB (Pol IV), are also known for by-passing the damage before the action ofreplicative polymerases (Nelson et al.,1996; Gerlach et al., 1999; Lin et al.,1999; McDonald et al., 1999; Tang etal., 1999; Wagner et al., 1999). The DNApolymerase encoded by yeast RAD30and human hRAD30A also bypasses acis-syn thymine-thymine dimer effi-ciently and accurately (Johnson et al.,1999 a,b; Masutani et al., 1999; Wash-ington et al., 1999; Johnson et al., 2000b,Washington et al., 2000). The damagebypassed by polymerases that insertwrong residues result in mutations,which can create more problems.
The human gene hRAD30B (McDonaldet al., 1999), which is a analog of yeastRAD30B, encodes the ninth eukaryoticDNA polymerase named Pol i, whichmisincorporates deoxynucleotides at a high
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

365
rate (Johnson et al., 2000 a). In general,DNA polymerases extend from mispairedbases at about the same frequency at whichthey insert the respective mispair (Goodmanet al., 1993). A new model for mutagenicbypass of DNA lesions in eukaryotesshowed that two DNA polymerases (i andz) act sequentially (Johnson et al., 2000 a).Pol i functions in damage bypass by insert-ing deoxynucleotides opposite DNA lesions,whereas Pol z acts at the subsequent step ofextending from them (Gibbs et al., 1998;Johnson et al., 2000a).
Whether the bypass repair is respon-sible for mutagenesis in plants remainsto be studied in detail. However, in maizepollen the UV-induced mutations werefound to be nontransmissible, whichsuggested that the mutations are of dele-tion type rather then point mutation(Nuffer, 1957).
F. Double-Strand Break (DSB)Repair
As early as in 1931, by using cyto-logical tools, Barbara McClintockshowed that the outcome of radiation-induced chromosome breaks is translo-cations, deficiencies, ring chromosomes,and end fusions (McClintock, 1931). Shesuggested that the broken ends can fusewith the same or other broken ends orchromosomes, we now call this DSBrepair. DSBs are usually created byionizing radiation (Dizdaroglu, 1992),chemical agents, during meiosis (Fried-berg et al., 1995) or transposition reac-tion (Lieber, 1991) and also as a resultof a nick in the single-stranded regiondue to mechanical stress of a chromo-somes (Britt, 1999). The unprotectedDSB is a dangerously cytotoxic lesion.If not repaired immediately, DSBs can
be attached to by unwelcome recombi-nation events, which can lead to loss ofportions of chromosomes or to rearrange-ments (error prone). Usually DSBs arerepaired by either homologous recombi-nation or by nonhomologous end-join-ing (NHEJ), which involves the joiningof two broken ends (Britt, 1999). Ho-mologous recombination is particularlyeffective in S phase when the break canbe repaired using genetic informationfrom a sister chromatid, whereas NHEJis thought to be effective at all times inthe cell cycle (Takata et al., 1998; Esserset al., 2000). NHEJ also plays an impor-tant role in DSB repair during V(D)Jrecombination in antibody coding frag-ments (Blunt et al., 1995). The model ofDSB is shown in Figure 7. In general,the organisms with relatively compactgenomes (yeast, bacteria, etc.) repair theDSB via homologous recombination,provided some homology is availableand organisms with larger genomes fol-low NHEJ (Britt, 1999). In plants theDSB repair products have been charac-terized as excision products of transpos-able elements, or insertion products ofAgrobacterium T-DNA (Ohba et al.,1995; Scott et al., 1996).
Recombination is initiated by a 5’to 3¢ exonuclease (Figure 7) that gen-erates long single-stranded 3¢ tails(White and Haber, 1990; Sugawara andHaber, 1992). In homologous recom-bination, one of these tails invades ahomologous DNA duplex. Overall, thisresulted in a formation of HollidayJunction, which resolved and formstwo recombinant DNA molecules withor without crossing over. The DSBrepair by homologous recombinationwith another allele can be achievedwith high fidelity, whereas repair byNHEJ may result in lost or changedgenetic information.
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

366
FIG
UR
E 7
. M
odel
s fo
r do
uble
-str
and
brea
k re
pair.
In
the
se s
chem
atic
mod
els,
dou
ble-
stra
nd b
reak
sca
used
by
eith
er ir
radi
atio
n or
che
mic
al a
ssau
lt or
dur
ing
mei
osis
and
tran
spos
ition
rea
ctio
ns o
r du
e to
mec
hani
cal s
tres
s ca
n be
rep
aire
d by
non
hom
olog
ous
end
join
ing
or n
onho
mol
ogou
s re
com
bina
tion
orho
mol
ogou
s re
com
bina
tion.
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

367
In plants, the DSB repair is stillpoorly understood. Evidence from trans-formed plants suggests that the majorityof transformed DNAs are integrated viaNHEJ. However, plants are capable ofrepairing DSB via either pathway. Inplants a few genes that are involved inDSB repair have been isolated. The ho-mologs of RecA genes (DMC/LIM15/RAD51) have been reported from plants(Kobayashi et al., 1993; Klimyuk andJones, 1996; Doutriauk et al., 1998). TheRecA-like recombination protein wasshown to be localized on chromosomesof the lily at various meiotic stages(Terasawa et al., 1995). The DSBs inplant somatic tissues can be repaired byhomologous recombination. Greater in-duction of homologous recombinationwas observed when site-specific DSBswere created using transposon or rare-cutting endonucleases. The first molecu-lar data on end-joining in plants camefrom the analysis of footprints left be-hind after transposable element excision(Rinehart et al., 1997). These footprintsusually consist of minor changes (i.e., afew base pair deletions, or inversion),and the exact configuration of DNA endsproduced by transposon excision remainsunknown. Recently, a Rad50 homologuegene and the encoded protein have beenisolated from Arabidopsis and shown tobe involved in meiosis and DSB repairin plants (Gallego et al., 2001).
The repair of DSBs by NHEJ re-quires the products of the XRCC4,XRCC5, XRCC6, and XRCC7 genes inmammalian system (reviewed by Chu,1997; Critochlow and Jackson, 1998).XRCC4 encodes a protein (XRCC4) thatforms a heterodimer with ligase IV thatrejoins two DNA ends in the last stepof V(D)J recombination and NHEJ torepair DSBs. XRCC4-defective cells areextrememly sensitive to ionizing radia-
tion and disruption of the XRCC4 geneor ligase IV results in embryonic le-thality in mice (Barnes et al., 1998).Recently, the crystal strucutre ofXRCC4 has been resolved at 2.7 A,which suggests a dumbbell-like tetramerstructure (Junop et al., 2000). In vitro,XRCC4 stimulates adenylation of li-gase IV, the first chemical step in DNAligation (Modesti et al., 1999). XRCC4may also bridge two ends for ligation,perhaps by interacting with Ku proteinalready present at the ends (Chen et al.,2000). XRCC5 and XRCC6 encode the70- and 80-kDa subunit of Ku, andXRCC7 encodes the catalytic subunitof DNA-dependent protein kinase(DNA-PKcs). In mammalian system, ithas been shown that DNA-PK is in-volved in the major pathway for repair-ing DSB (Jeggo et al., 1995). DNA-PKis a heterotrimer consisting of a 450-kDa catalytic subunit (DNA-PKcs) andKu80 and Ku70 regulatory subunits(Anderson, 1993; Tuteja et al., 1994,Tuteja and Tuteja, 2000). The Ku80and Ku70 of DNA-PK bind to DNAends (broken ends), which activate thekinase activity of DNA-PKcs. TheDNA-PKcs is a serine/threonine pro-tein kinase and can phosphorylate avariety of substrates in vitro (Tutejaand Tuteja, 2000). Finally, the Ku80and Ku70 bring the DNA broken endstogether and then DNA ligase joinedthe ends (Figure 7). The end-bindingactivity of Ku serves as an entry site forother proteins and the phosphorylationactivity of DNA-PKcs activate otherrepair factors (Tuteja and Tuteja, 2000).DNA-PK is the only factor known toparticipate in NHEJ. DNA-PKcs is amember of the phosphatidylinositol3-kinase (PI3K)-related kinase family,as are ATM and ATR. All three pro-teins exhibit a strong sequence homol-
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

368
ogy to the PI3Ks, especially in the cata-lytic core domain that binds and phos-phorylates the phosphoinositol headgroup of phosphtidylinositol. However,no phosphorylation of lipid substrateshas been observed by DNA-PKcs or byother members of this PI3K-relatedfamily of kinases (Carpenter andCantley, 1996; Wymann and Pirola,1998).
The mutants deficient in Ku orDNA-PKcs are deficient in the repairof DSBs and also sensitive to lethaleffects of ionizing radiation. A newactivity of Ku as an unwinding enzymewas also reported (Tuteja et al., 1994;Tuteja et al., 1997). The analog of Kuin plant has not been reported yet. How-ever, we have shown through Westernblotting that the Ku or Ku-like proteinare also present in plant (Tuteja andTuteja, 2000). However, by using hu-man Ku antibodies and pea cDNA li-brary, we got the positive clone thatlater turned out to be nonspecific andactually was the clone of calnexin, aCa2+-binding molecular chapareon pro-tein (Ehtesham et al., 1999).
In plants NHEJ has been analyzedin a more direct and thorough man-ner. Linearized plasmid DNA hasbeen introduced into tobacco cells,and novel joints that formed betweenthe plasmid ends have been sequenced(Gorbunova and Levy, 1997). In an-other study, DSBs were induced at anI-SceI recognition site, within a nega-tive selectable marker genome on achromosome, and new junctions weresequenced (Salomon and Puchta,1998). Gorbunova and Levy (1997)summarized the results of these stud-ies as follows:
∑ End joining by simple ligation with nosequence alteration is rare.
∑ End-joining is usually associated withdeletions ranging from 1 bp up to > 1kb.
∑ Rejoining frequently occurs at shortrepeats.
∑ A distinct feature of the junctions is thepresence of relatively large insertions,up to 1.2 kb long.
The filler DNA could consist of asimple insertion corresponding to aplasmid or genomic sequence. Inser-tions might also have a complex struc-ture consisting of a patchwork of dif-ferent plasmid and tobacco genomicsequences.
In response to DSB, different repairroutes can be taken. The break can alsobe simply ligated, without any modifi-cation of the ends or different events canhappen at the broken site, such asexonuclear degradation or exposure of3¢-ss end, which was proposed to be thefirst step of DSB repair pathway inplants. When annealing of the exposedends occurs at sites of microhomology,as often found in plants (Gorbunova andLevy, 1997; Salomon and Puchta, 1998),this results in a deletion of the regionbetween the microrepeats. This processis reminiscent of the single strand an-nealing (SSA) mechanism, which wasalso proposed for extrachromosomal ho-mologous recombination events in plants(Puchta and Hohn, 1991). The outcomeof DSB repair is also the presence oftemplated filler DNA at the repair site(Gorbunova and Levy, 1997). Formationof such filler DNA can be best explainedby the synthesis-dependent strand anneal-ing (SDSA) mechanism that involves tem-plate invasion and DNA synthesis(Gorbunova and Levy, 1999). The SSAand SDSA appear to be the two prominentmechanisms for DSBs repair in plants.They can be used either for homologousor nonhomologous recombination.
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

369
G. Mismatch Repair (MMR)
Because of the errors of replica-tion or homologous recombination,mismatched bases can be produced(Friedberg et al., 1995; Friedberg andWood, 1996). The MMR basically dis-criminates between the correct and in-correct bases and finally corrects theerror after DNA synthesis. Most of thepresent understanding of eukaryoticMMR has come from studies of theeukaryotic homologs of the bacterialMutS and MutL proteins (Kolodner,1996; Kolodner and Marsischky,1999). The model of MMR in E. coliis shown in Figure 8. In E. coli theMutS dimer recognize the mispairs andthen bind on it (Grilley et al., 1989)followed by MutL binding, which ac-tivates MutH (endonuclease) thatmakes a single-strand incision. Thisincision by MutH can be on either sideof the mismatch (Au et al., 1992). Sub-sequently, the excision initiates at theincision (nick) and proceeds towardthe mismatch (Grilly et al., 1993). Tofill the gap (100 to 1000 nucleotidegap) the original template strand canthen be replicated and finally sealedby ligation (Figure 8). The proteinsinvolved in the last step of eukaryoticMMR are DNA pol d, RP-A, PCNA,and RFC (for a review see Kolodner,1996; Kolodner and Marsischky,1999).
The homologs of MutS and MutLhave been reported in yeast and human(Modrich, 1994; Drummond et al., 1995;Palombo et al., 1995; Hunter et al., 1996;Johnson et al., 1996; Palombo et al.,1996). In plant, the homolog of MutSwas reported from Arabidopsis (Culliganand Hays, 1997), but the function hasnot been determined yet.
IV. ROLE OF DNA HELICASESIN DAMAGE AND REPAIR
The DNA helicases are ATP-depen-dent DNA unwinding enzymes and thusplay an important role in repair as wellas in replication, recombination, andtranscription (Tuteja and Tuteja, 1996;Tuteja, 1997). The unrepaired DNA le-sions can affect the outcome of all theseprocesses. The helicases are among thefirst proteins that would encounter DNAdamage during all the processes above.The interstrand or intrastrand DNA le-sions can block or impair the DNAhelicase action. The XPB and XPD arerepair helicases that are required for lo-cal unwinding of the damage area ofduplex DNA in NER in the mammaliansystem (Schaeffer et al., 1993; Schaefferet al., 1994). During DNA repair, espe-cially in NER, the damaged area on DNAhas to unwind first in order to get properrepair, because most of the DNA repairmachinery enters through single-stranded DNA. This single-strandedDNA is provided transiently by the DNArepair helicase(s). This unwinding pro-cess by DNA helicases is an energy-requiring process that comes through thehydrolysis of ATP by the intrinsic DNA-dependent ATPase activity of thehelicase protein itself (Tuteja, 2000).
DNA helicases generally bind tossDNA or ss-/ds-DNA junctions andtranslocate unidirectionally along thebound DNA into the duplex region, dis-rupting the hydrogen bonds linking thetwo strands (Tuteja and Tuteja, 1996).The XPB can unwind DNA in the 3¢ to5¢ direction (Schaeffer et al., 1993; Royet al., 1994) similar to previously re-ported human DNA helicases (HDH) I,II, III, V, and VI (Tuteja et al., 1990;Tuteja et al., 1992 to 1995), pea chloro-
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

370
plast DNA helicases I and II (Tuteja etal., 1996; Tuteja and Phan, 1998), peanuclear DNA helicase 45 (Pham et al.,2000), and pea nuclear DNA helicase65 (Tuteja et al., 2001). However, XPDunwinds the DNA opposite to XPB po-larity, that is, in 5¢ to 3¢ direction (Sunget al., 1993), which is similar to HDHIV (Tuteja et al., 1991). In order to bet-ter understand NER in plant, it is impor-tant to isolate the DNA repair helicases
along with other components. Theserepair helicases could be analogs of XPBand XPD that need to be cloned in anexpression vector for purification of theencoded proteins. Furthermore, theseproteins need to be characterized in de-tail. XPC-hHR23B may assist this strandseparation by interacting with TFIIH(Drapkin et al., 1994). The XPB andXPD both contain the seven conserveddomains (helicase motifs) of the super-
FIGURE 8. A model for mismatch repair in E. coli. The eukaryotic process is believed toshare many of the features shown here.
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

371
family of DNA and RNA helicases(Gorbalenya and Koonin, 1993; Tuteja,2000). However, the sole presence ofhelicase motifs in the protein may notprove that it is a helicase. For example,an E. coli Mfd protein (a strand-specificrepair protein) (Selby and Sancr, 1993)and a human Rad54 protein (Swagemakerset al., 1998) both contain conservedhelicase motifs but lack the unwindingactivity. So a direct biochemical demon-stration of ATP-dependent DNA or RNAunwinding activity is essential before aprotein is classified as helicase (Tuteja,2000).
In E. coli the Uvr2B and UvrD arethe repair DNA helicases involved inNER. The DNA unwinding activity ofUvrD helicase was shown to be exquis-itely sensitive to major groove DNAintercalators (George et al., 1992), whileit was moderately sensitive to UV radia-tion. In general, the UV-induced lesionsor other helix-distorting intrastrand ad-ducts are better tolerated by DNAhelicases than those produced by inter-calating agents. However, in yeastS. cerevisiae the Rad3 protein, a repairhelicase, is absolutely required in NER.It is found to be inhibited by UV dam-age and intrastrand cisplatin adductslocated on the strand along which theenzyme translocates, whereas lesions onthe opposite strand had no effect (Villaniand Tanguy Le Gac, 2000). Calf thymusDNA helicase E is a 3¢ to 5¢ helicase thatcan unwind DNA duplex from nicks andhas been suggested to play a role inDNA repair (Turchi et al., 1992).
Ku protein is another DNA helicase(human DNA helicase II) that plays animportant role in double-strand breakrepairs (Tuteja et al., 1994; Tuteja andTuteja 2000). Ku is a heterodimeric pro-tein composed of ~70 and ~80 kDa sub-units (Ku 70 and Ku 80) originally iden-
tified as an autoantigen recognized bythe sera of patients with autoimmunediseases. It is an abundant nuclear pro-tein found in vertebrates, insects, yeastworms, and also in plant (Tuteja andTuteja, 2000). Ku contains ssDNA-de-pendent ATPase, DNA end binding, andATP-dependent DNA unwinding activi-ties (Tuteja et al., 1994). It prefers tounwind the partially, unwound DNAsubstrates and moves in the 3¢ to 5¢ di-rection on the bound strand (Tuteja etal., 1994). Ku also functions as a regu-latory subunit of DNA-PK that phos-phorylates many proteins, such as p53,SV-40 large T antigen, RP-A, hsp90, c-jun, C-Fos, Sp-1, C-Myc, TFIID, etc.(Tuteja and Tuteja, 2000). The DNAunwinding activity of Ku was found tobe severely inhibited by DNA interca-lating agents such as nogalamycin,daunorubicin, actinomycin, and ethidiumbromide (Tuteja et al., 1997).
In general, intrastrand DNA dam-age seems to affect DNA unwinding ac-tivity of the helicase in vitro when placedon the strand to which the protein trans-locates with exception to the SV-40 Tantigen and RecA helicases (Villani andTanguy Le Gac, 2000). The DNAhelicases involved in initial stages ofNER are more sensitive to DNA dam-age (e.g., UvrAB, Rad3, or XPD) whencompared with the helicases that are in-volved in the subsequent removal of thedamaged DNA (e.g., UvrD).
V. DESSICATION-RELATEDDAMAGE, TOLERANCE, ANDDNA REPAIR
Plant seed, the product of fertiliza-tion, during its final stages of develop-ment and maturation undergoes desic-
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

372
cation, resulting in a significant reduc-tion in its water content. In addition,metabolic activities are also reduced toundetectable levels, leading seed em-bryo in quiescent state. Under low tem-perature and moisture, seed can now bestored for long periods until it is reacti-vated by favorable seed germinationconditions. Seed viability loss has beenshown to result in a loss of membraneintegrity, impaired RNA, and proteinsynthesis and ATP production (Priestleyet al., 1985; Bailly et al., 1996). Ultra-structural, biochemical, and biophysicalstudies have shown that the loss of des-iccation tolerance is correlated with anirreversible alteration to the physicalproperties of membrane lipid bilayers,increased liquid crystalline-gel phasetransition temperature, deesterificationof phospholipids, and an accumulationof several products of peroxidized lip-ids. Observations that lipid-rich seedstend to have a limited longevity tend tosuggest that seed aging based on lipidperoxidation is important (Priestley etal., 1985).
Free radicals and lipid peroxidationare considered to be major cause of seeddeterioration (Leprince et al., 1996).Ungerminated seeds and pollen areknown to contain and accumulate freeradicals during artificial aging (Priestleyet al., 1985) and germinating seeds afterdehydration (Senaratna and McKersie,1986). Changes in lipid peroxidation andin the molecular defenses against theactivated forms of oxygen have also beenobserved in dry stored seeds and seedafter accelerated aging. The loss of seedviability has been correlated with theMDA (malondialdehyde) content, aproduct of lipid oxidation. Reuzeau andCavalie (1995) found a significant dif-ference in the MDA content of seedsable or unable to germinate. A higher
lipid peroxidation was found in seedsunable to germinate. It has been sug-gested that secondary toxic products pro-duced as a result of oxygen-dependentreactions in imbibed seeds by the accu-mulation of oxygenated fatty acids couldparticipate in altered cellular componentsleading to a decrease in seed viability(Gidrol et al., 1989; Reuzeau andCavalie, 1995). When such seeds arehydrated, toxic oxygen products, if theyare not efficiently detoxified, can allowdamaging reactions and prevent seed ger-mination. Because dry seeds are affectedin their ability to detoxify O2-relatedtoxic products, it seems reasonable tosuggest that important changes in solubleand membrane entities might interferein the signal transduction necessary forthe initiation of germination process(Reuzeau and Cavalie, 1995). Similarconclusions on the role of lipid peroxid-ation were drawn when accelerated ag-ing of seed was studied (Bailly et al.,1996). Nonviable seeds with acceleratedaging induced the accumulation ofmalondialdehyde, indicating that seeddeterioration was associated with lipidperoxidation. No direct correlation be-tween lipid peroxidation and deteriora-tion in membrane integrity was found.However, loss of seed viability was as-sociated with a decrease in superoxidasedismutase, catalase and glutathione re-ductase (GR) activities. This study es-tablished a clear relationship betweenthe loss of detoxifying enzymes such ascatalase, and GR, lipid peroxidation, andseed deterioration during acceleratedaging. Whether oxidative damage anddecrease in SOD, catalase, and GR ac-tivities are the cause or consequence ofthe loss of seed viability was not estab-lished (Bailly et al., 1996).
The desiccation tolerance mecha-nisms allow cells to withstand the loss
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

373
of the major percentage of its water con-tent and to survive in a dry state forprolonged periods. The seeds, spores,wind-dispersed pollen, and specialized‘resurrection’ species can withstand des-iccation without incurring cellular dam-age, and they naturally dehydrate tomoisture contents that will result in celldeath in other plant tissues.
The role of free radicals in loss ofdesiccation tolerance in germinatingseed has also been studied (Leprince etal., 1990). Desiccation of 24 h duringgermination resulted in a significant in-crease in lipid peroxidation in the de-hydrated tissues, and rehydration notonly fails to reduce lipid peroxidationbut resulted in a 15-fold increase inMDA accumulation, indicating thatdesiccation treatment results in irrevers-ible peroxidative damage to lipids. It isnot clear whether oxygen radicals arethe cause of or a result of the damagefollowing desiccation, but it is well es-tablished that desiccation alters theproperties of membranes (Senaratna andMcKersie, 1986). A desiccation-in-duced rise in lipid peroxid-ation origi-nates from oxygen radicals and a lossof desiccation tolerance is a conse-quence of increased formation of oneor more forms of activated oxygencoupled with decreased protection fromSOD and peroxidases. It is now clearthat damages to DNA are harmful,which is why the repair of DNA dam-age is of vital importance to the cell inorder to keep its normal function.
A. In Bacterial Spores
Desiccation tolerance of bacterialspores depends on the conversion ofB-form DNA to a more condensed A-
form (Setlow, 1992a,b). The DNA indormant spores of Bacillus species isassociated with alpha/beta-type, smallacid-soluble proteins (SASP), which aredouble-stranded DNA-binding proteinswhose amino acid sequence has beenhighly conserved in evolution. The ex-pression of these proteins occurs at aspecific stage of spore maturation whenthe B-form DNA duplexes lose waterand are converted to A-form conforma-tion (Setlow, 1992a,b). These proteinsbind to A-form, conferring stability onthe DNA during the final stages of de-hydration and through the dehydratedstage. These proteins are released by theaction of the sequence-specific protease(termed GPR) that degrades SASP dur-ing germination. These spore proteaseof Bacillus are synthesized during sporu-lation as an inactive precursor termedP46. The activation of this proteaseduring rehydration leads to the re-moval of SASPs and the return of theDNA to the B-form (Illadas-Aguiarand Setlow, 1994). The saturation ofspore DNA with SASP provides pro-tection against DNA base loss due towet heat (i.e., de-purination) andbreakage of DNA single-strands bydesiccation and hydrogen peroxide.The spores lacking a/b SASP are muchmore sensitive to desiccation, dry andwet heat, and hydrogen peroxide thanare wild-type spores. All these treat-ments have been shown to kill sporesin large part by damaging DNA(Setlow and Setlow, 1996). Bindingof a/b type SASP also helps in con-ferring resistance to UV damage(Setlow and Setlow, 1996).
The major deleterious DNA photo-product generated by UV irradiation ofgrowing cells is a cyclobutane-typedimer between adjacent thymine resi-dues (termed TT), but TT is not formed
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

374
in spores. The major UV photoproductformed in spore DNA is a thymine-thym-ine adduct termed the spore photoprod-uct (SP), which is also formed betweenadjacent thymine residues. SASP defi-cient spores are ~30 times more UVsensitive than the wild-type spores, andUV generates a large amount of TT indeficient spores. According to Setlowand Setlow (1996), the generation of TTis the cause for the increased UV sensi-tivity of mutant spores. These authorsexamined the expression of genes in-volved in DNA repair during germina-tion and outgrowth of both wild typeand a-b-spore, which were exposed to avariety of treatments. This study hasshown that the repair of DNA damageduring spore outgrowth is an importantcomponent of spore resistance to a num-ber of DNA-damaging treatments. TheSASP-type proteins have not been iden-tified in plants as such. However, otherproteins may also play an important rolein survival of dehydrated seeds and pol-len. Dehydrinds and LEA-type proteinsmay have some role in desiccation toler-ance.
The Deinococcus radiodurans is anon-spore-forming bacterium that exhibita remarkable capacity to resist the lethaleffects of ionizing radiation. It has beenreported that D-radiodurans ionizing ra-diation resistance is incidental, a conse-quence of this bacterium’s adaptation todehydration (Mattimore and Battista,1996). The authors showed thatD. radiodurans’s cellular responses to ion-izing radiation and dehydration signifi-cantly overlap by showing that 41 ioniz-ing radiation-sensitive strains ofD. radiodurans were also sensitive to des-iccation. It was also inferred that loss ofDNA repair ability in the ionizing radia-tion-sensitive strains was responsible forthe desiccation sensitivity of these strains.
B. In Seeds
It has been known for more than 2decades that a feature of loss of viabilityin stored dry seed is the increasing lossof integrity of nuclear DNA with in-creasing levels of low-molecular-weightoligonucleotide fragments (Cheah andOsborne, 1978, Elder et al., 1987; Elderand Osborne 1993; Leprince et al., 1996).When rye embryos that are nonviableare imbibed for 2.5 h the extent of frag-mentation of DNA shows a significantincrease pointing toward the continuousactivity of endonuclease cleavage in theabsence of DNA repair (Cheah andOsborne, 1978; Elder et al., 1987). Em-bryos of imbibed dormant A. fatua alsoshowed an efficient repair of radiation-induced lesions in the DNA of dry seeds,and this DNA repair also remained stablewhen embryos were dried back. On thebasis of results from various experi-ments, Boubriak et al. (1997) have sug-gested that the failure to restore geneticintegrity to the genome on rehydrationof an embryo whose DNA is desicca-tion damaged could be a major factorin determining desiccation tolerance.The ability to repair DNA when em-bryos are rehydrated is critical to main-taining a transcriptionally functionalgenome.
Elder et al. (1987) have shown ac-cumulation of single- and double-strandbreaks in the embryos of seeds duringthe loss of viability in the dry state. Incontrast to hydrated cell, which can ef-fectively repair DNA damage, the nu-clei of dry cells have to accumulate DNAdamage until rehydration activates DNArepair machinery. The stability of theDNA on desiccation and the capacityfor repair of DNA on rehydration havebeen suggested as integral components
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

375
of a total desiccation tolerance mecha-nism.
C. DNA Integrity in Pollens
Osborne and Boubriak (1994) haveshown that dry birch pollen (14% mois-ture content) can be fully hydrated inmoist air and dehydrated back to theoriginal dry weight without the loss ofgerminability and without the fragmen-tation damage detectable in the extractedDNA. At the time of anther dehiscence,birch pollen is haploid and bi-nucleateand arrested at the G2 stage of the cellcycle. The authors addressed the ques-tion whether there is evidence for DNArepair during the air hydration of birchpollen. Pollen that has been exposed toincreasing durations of hydration inmoist air prior to the transfer to the liq-uid germination medium showed simi-lar levels of H3-methyl thymidine. Itwas suggested that an excision repairmeasured as unscheduled DNA synthe-sis does not occur during the relativelylow level of hydration possible in moistair. As pollen gets exposed naturally toUV in solar radiation following releasefrom the anther, it is the likely candidatefor UV-induced heritable mutations.
The role of DNA repair in the desic-cation tolerance of pollen and spores isnot unlike that ascribed to DNA repairsin bacterial spores (Setlow, 1992a,b). Intheir dehydrated state, pollen and seedcontinue to accumulate DNA damagethat cannot be repaired until the rehy-dration occurs. The low water content inthe desiccated stage prevents enzymaticactivity and therefore DNA repair. Noevidence of DNA repair has been ob-served in dry pollen or seed. Thus DNAdamage cannot be repaired until pollen
and seed are rehydrated. Given the DNAdamage that may accumulate in pollenand spores during the dormant period,they may contain high levels of enzymesneeded for DNA repair.
Jackson and Linskens (1978) madecomprehensive analysis of DNA repairsystem in Petunia pollen. The maturepollen of Petunia carries generativenucleus blocked at G2 stage of cell cycle.The addition of 4-nitroquinolide 1-ox-ide to the liquid germination mediumled to unscheduled DNA synthesis inpollen. Mature Petunia pollen is bicellu-lar and thus has two nuclei, that is, gen-erative and vegetative nuclei. In thesestudies on Petunia pollen it was not fea-sible to discern whether the repair syn-thesis takes place in either or both nucleiof pollen. It was shown that unsched-uled DNA synthesis occurs after ultra-violet radiation. As this unscheduledDNA synthesis was not affected by hy-droxyurea, an agent used to discrimi-nate between replicative and repair syn-thesis, the authors suggested that darkrepair of DNA takes place in germinat-ing pollen of Petunia (Jackson, 1987).
VI. DNA REPAIR GENESIDENTIFIED BY SEQUENCEHOMOLOGY
Recently, Wood et al. (2001) havedescribed a catalog of 130 known hu-man repair genes on the basis of func-tion or sequence homology to knownrepair genes. Here we are mainly de-scribing the plant repair genes found onthe basis of sequence homology. Thecompletion of the first plant genomesequence of Arabidopsis thaliana is alandmark of twentieth century in plantresearch (The Arabidopsis Genome Ini-
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

376
tiative, 2000). The information it con-tains will certainly give a boost to everyarea of plant science, including DNArepair in near future. A. thaliana is amodest little flowering brassica plantchoosen first for plant genome sequencebecause it has many advantages for ge-nome analysis, including a small size,large number of offspring, a short gen-eration time, and a relatively smallnuclear genome. In 1977 William Curtis,a British botanist and apothecary,described A. thaliana in his floralandinensis as a plant of “no particularvirtues or uses”, but now with the reportof its genome sequence it becomes themost significant plant in the kingdom(Dennis and Surridge, 2000). The sizesof the Arabidopsis genome of nuclear,plastid and mitochondrial are 125 Mb,560 kb, and 367 kb, respectivley. It hasfive chromosomes (chromosomes 1 to5) with respective sizes of 29.1, 19.6,23.2, 17.5, and 26.0 Mb. The evolutionof Arabidopsis involved a whole-genomeduplication, followed by subsequentgene loss and extensive local gene du-plications. The nuclear genome contains25,498 gene encoding proteins from11,000 families, which is the largest geneset published to date (The ArabidopsisGenome Initiative, 2000).
Comparing the Arabidopsis genomewith other species (Eisen and Hanawalt,1999) indicated that Arabidopsis has aset of DNA repair and recombination(RAR) genes similar to most other eu-karyotes. The pathways represented in-clude PR, DNA ligation, NHEJ, BER,MR, NER, and many aspects of DNArecombination. The Arabidopsis RARgenes include homologues of many DNArepair genes that are defective in differ-ent human diseases. For example, he-reditary breast cancer and nonpolyposiscolon cancer, Xeroderma pigmentosum
(D-XPD, F-XPF, B-ERCC3), Ataxia te-langiectasia (ATM), and Cockayne’ssyndrome (CSA, CSB) (The ArabidopsisGenome Initiative, 2000). The presenceof additional homologues of many RARgenes was found in Arabidopsis genome.This is seen for almost every major classof DNA repair, including recombina-tion (four RecA), DNA ligation (fourDNA ligase I), PR (five class I photolyaseand one class II photolyase), and NER(six RPA1, two RPA2, two Rad25, threeTFB1, and four Rad23). Regarding BER,Arabidopsis contianed 16 homologuesof DNA base glycosylates, which is themaximum number known for any spe-cies. It also contains three homologuesof AP endonuclease. Some of the extracopies of RAR genes originated throughrelatively recent gene duplication be-cause many of the sets of genes weremore closely related to each other thanto their homologues in any otherspecies. The presence of duplicate(paralogous) genes might be responsiblefor greater repair capacity.
The multiplicity of RAR genes couldalso be partly due to the transfer of genesfrom the organellar genomes to thenucleus. Repair gene homologues thatappear to be of plastid origin include therecombination protein RecA, RecG, andSMS, two class I photolyase homo-logues, Fpg, two MutS2 proteins, andthe transcription repair coupling factorMfd. The homologues of E. coli repairgene UvrABC was found to be absent inArabidopsis. As Arabidopsis showedmany similarities to the set of DNA re-pair genes found in other eukaryotes, itwill not be difficult to determine thefunctions of these gene-encoded proteinsin the plant system (The ArabidopsisGenome Inititative, 2000). Recently, theA. thaliana Rad50 homologue (AtRad50)has been isolated and shown to be lo-
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

377
cated on chromosome 2 (Gallego et al.,2001). The Rad50 protein is involved inthe cellular response to DNA double-strand break (DSBs), including the de-tection of damage, activation of cell-cylce checkpoints, and DSB repair viarecombination. Gallego et al. (2001) haveshown that Rad50 homologue protein isinvolved in meiosis and DNA repair inplants.
VII. CONCLUSIONS ANDFUTURE PROSPECTS
The survival of organisms dependson the accurate transmission of geneticinformation from one cell to its daugh-ters. Such faithful transmission requiresnot only extreme accuracy in the repli-cation of DNA and the precision in chro-mosome distribution, but also the abilityto survive spontaneous and inducedDNA damage while minimizing thenumber of heritable mutations. Toachieve this fidelity, cells have evolvedseveral DNA repair mechanisms that canmonitor the structure of chromosomeand coordinate repair and cell-cycle pro-gression. These repair mechanisms alsohelp in maintaining the genome integ-rity and in regulating mutation rates. Ifthe DNA damage is not repaired prop-erly, it can lead to various complica-tions, including cell death or mutations.The mutagenicity may be caused by per-sistent, unrepaired adducts that are sub-sequently converted to mutations throughDNA replication or to mutation fixationthrough misrepair.
In general, the damaged proteins andRNA molecules can be replaced usinginformation encoded in the DNA, butthe DNA molecules themselves are irre-placeable. Therefore, the damaged DNA
has to be repaired. DNA repair is pos-sible largely because the DNA moleculeconsists of two complementary strands.DNA damage in one strand can be re-moved and accurately replaced by usingthe undamaged complementary strandas a template. DNA can become dam-aged by a variety of means, some spon-taneous, others catalyzed by environ-mental agents or defect in replication,which can leave mispaired bases. Thechemistry of DNA damage is diverseand complicated. Consequently, the cel-lular response to this damage includes awide range of enzymatic systems thatcatalyze some of the most interestingchemical transformations in DNA me-tabolism.
During the last several years, con-siderable progress has been made inelucidating the components of the eu-karyotic DNA damage response andDNA repair pathways. How damagedDNA activated DNA damage responseproteins is a question of central impor-tance to be addressed in near future. Inprinciple, a sensor protein should havethe ability to interact with damagedDNA. As there are many ways to dam-age DNA, it is important to determinewhether there is a sensor for each typeof damage. If the damage itself is sensedthrough the interaction with a sensorprotein, this sensor protein may poten-tially interfere with repair proteins thatalso need access to damaged DNA. Thecell must be aware not only of damage,but also of when the damage is repaired,because the completion of repair shouldbe the signal for termination of the dam-age response. Further work is needed todefine the activation of damage-signal-ing kinases. Another area ripe for futureexploration is the relationship betweenthe DNA damage signaling pathway andtelomeric DNA structures. Any DNA
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

378
lesions that block the movement of DNAand RNA polymerases may kill cells byinterfering with replication in activelygrowing and dividing cells or by block-ing transcription and thus depriving non-dividing cells of an essential protein.Current models dealing with replicationof damaged DNA postulate the initialdissociation of replicative DNA poly-merases when encountering sites of DNAdamage. Replication can then restart be-yond leisons, or DNA polymerases spe-cialized in translesion synthesis can tem-porarily replace replicative polymerasesto synthesize across the lesion; replica-tive enzymes then take over again. TheDNA helicase can help in this bypassreaction; however, this helicase-medi-ated bypass hypothesis is still specula-tion.
In modern biology the DNA repairstudy occupies a central place. The DNArepair field in eukaryotes has a strongconnection with replication, transcrip-tion, recombination, and cell cycle con-trol and checkpoints (G1/S and G2/M).Checkpoints are mechanisms that allowcells to deal with DNA damage or otherinsults. A major role of checkpoints is todelay cell cycle transitions in order toallow time for the damage to be repaired.Therefore, it is important to understandhow checkpoints regulate the basic cellcycle machinery. The overall progressof research on DNA repair in plant cellhas been very slow when compared withother systems. However, now this fieldhas started progressing fast in plants es-pecially after the isolation of several re-pair protein and genes. However, thetwentieth century began with the redis-covery of Mendel’s rules of inheritancein pea (De Vries, 1900), and it ends withthe elucidation of the complete sequenceof Arabidopsis. The analysis of com-pleted sequence provides insights into
the genetic basis of the similarities anddifferences of diverse multicellular or-ganisms. Regarding DNA repair genes,it appears that they are highly conservedbetween plants and mammals than withinthe animal kingdom, perhaps reflectingcommon factors such as DNA methyla-tion.
DSB repair in plants is error prone,with the main pathway of repair beingillegitimate recombination. Such an er-ror-prone DNA repair mechanism mighthave an important impact on plant evo-lution. In plant the DSB repair productshave largley been characterized as exci-sion products of transposable elementsor insertion products of T-DNAs. Theseproducts are similar to mammalian DSBrepair products, which also suggests thatplant DSB repair is similar to mammals.There is an urgent need to study the Kuand DNA-PK in plants in order to un-derstand the mechanism of DSB repair.Still there is a need to identify all therepair genes in plants as reported in othersystems. One way of identifying thegenes involved in DNA repair is to studymutants that are sensitive (or resistant)to DNA-damaging agents. The identifi-cation of mutants that are selectivleyaltered in only one type of DSB repairwill shed light on the interrelations be-tween the different DSB repair path-ways. It might also allow the selectiveactivation of a homologous pathway toachieve gene targeting.
There is still much to be done. Vari-ous models proposed for repairs arelargely hypothetical and require furtherexperimental support. Cloning of all therepair genes and studying knockouts oroverexpression of these genes could pro-vide a solution for a longstanding prob-lem of gene targeting in plants. Further,the crystallographic studies of DNA re-pair enzymes will reveal a wide range of
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

379
structural motifs used to recognize spe-cific lesions in DNA. These studies willalso help to unravel the catalytic mecha-nism by which these lesions are repaired.We hope that in future new technolo-gies, such as gene machines for reversegenetics, sequence analysis of wholeplant genomes, and utilization of DNAchips, will give a boost to unravel themystery behind DNA repair mecha-nisms. Also in the near future proteomicswill provide a wealth of protein-proteininteraction data, which will add to theknowledge of repair proteins function.The challenge for future research is touncover the exact molecular mechanismsin plant DNA repair system throughwhich these repair proteins repair theDNA damage in cooperation with otherproteins. More research is needed inplants to answer the many questions thatstill surround the repair proteins and theirmany targets.
ACKNOWLEDGMENT
The writing of this review was initi-ated during the visit of the senior author(NT) to the University of Melbourne un-der the Australia India Council program.
REFERENCES
Aboussekhra, A., Biggerstaff, M., Shivji,M. K. K., Vilpo, J. A., Moncollin, V.,Podust, V. N., Protic, M., Hubscher,U., Egly, J.-M., and Wood, R. D. 1995.Mammalian DNA nucleotide excisionrepair reconstituted with purified com-ponents. Cell 80:859–868.
Aboussekhra, A. and Thoma, F. 1999.TATA-binding protein promotes the
selective formation of UV-induced (6-4)-photoproducts and modulates DNArepair in the TATA box. EMBO J.18:433–443.
Agrawal, A. and Kale, R. K. 2001. Radia-tion induced peroxidative damage:mechanism and significance. IndianJ. Exp. Biol. 39:291–309.
Ahmad, M., Jarillo, J. A., Klimczak, L. J.,Landry, L. G., Peng, T., Last, R. L.,and Cashmore, A. R. 1997. An en-zyme similar to animal type IIphotolyases mediates photoreactiva-tion in Arabidopsis. Plant Cell 9:199–207.
Anderson, C. W. 1993. DNA damage andthe DNA-activated protein kinase.Trends Biochem. Sci. 18:433–437.
Au, K. G., Welsh, K., and Modrich, P. 1992.Initiation of methyl-directed mismatchrepair. J. Biol. Chem. 267:12142–12148.
Bailly, C., Benamar, A., Corbineau, F.,and Cume, D. 1996. Changes inmalondialdehyde content and in su-peroxide dismutase, catalase andglutathione reductase activities insunflower seeds as related to dete-rioration during accelerated aging.Physiologia Plantarum 97:104–110.
Balajee, A. A. and Bohr, V. A. 2000. Ge-nomic heterogeneity of nucleotideexcision repair. Gene 250:15–30.
Barb, W. G., Baxendale, J. H., George, P.,and Hargrave, K. R. 1957. Hydroxylradical generation by Fenton reaction.Trans. Faraday Soc. 47:462–470.
Barber, A. A. and Berheim, F. 1967. Loss offluidity and break down of cell mem-brane: functions and transmembraneionic gradient by lipid peroxidation.Adv. Gerontol. Res. 2:355–363.
Barnes, D. E., Stamp, G., Rosewell, I.,Denzel, A., and Lindahl, T. 1998.
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

380
Targeted disruption of the gene en-coding DNA ligase IV leads to lethal-ity in embryonic mice. Curr. Biol.8:1395–1398.
Batschauer, A. 1993. A plant gene forphotolyase: An enzyme catalyzing therepair of UV-light induced DNA dam-age. Plant J. 4:705–709.
Beauchamp, C. O. and Fridovich, I. 1973.Isozymes of superoxide dismutasefrom wheat germ. Biochem. Biophys.Acta. 317:50–56.
Bennett, R. A. O., Wilson, D. M., Wong,D., and Demple, B. 1997. Interactionof human apurinic endonuclease andDNA polymerase beta in the base ex-cision repair pathway. Proc. Natl.Acad. Sci. USA, 94:7166–7169.
Beranek, D. T. 1990. Distribution of methyland ethyl adducts following alkyla-tion with monofunctional alkylatingagents. Mutat. Res. 231:11–30.
Bessho, T., Sancar, A., Thompson, L. H.,and Thelen, M. P. 1997. Reconstitu-tion of human excision nuclease withrecombinant XPF-ERCC1 complex. J.Biol. Chem. 272:3833–3837.
Bethke, P. C. and Jones, R. L. 2001. Celldeath of barley aleurone protoplasts ismediated by reactive oxygen secies.Plant J. 25:19–29.
Bialkowski, K. and Olinski, R. 1999. Oxi-dative damage to plant DNA in rela-tion to growth conditions. ActaBiochim Pol. 46:43–49.
Blunt, T., Finnie, N. J., Tacciolo, G. E.,Smith, G. C. M., Demengeot, J.,Gottlieb, T. M., Mizuta, R., Vrghese,A. J., Alt, F. W., Jeggo, P. A., andJackson, S.P. 1995. Defective DNA-dependent protein kinase activity islinked to V(D)J recombination andDNA repair defects associated withthe murine scid mutation. Cell 80:813–823.
Bormman, J. F. 1989. Target sites of UV-Bradiation in photosynthesis of higherplants. J. Photochem Photobiol. B4:145–158.
Bornman, J. F., Reuber, S., Chen, Y-P., andWeissenboeck, G. 1997. Ultravioletradiation as stress factor and the roleof protective pigments. In: Plants andUV-B. Responses to EnvironmentalChange (Lumsden, P. J., Ed.). Cam-bridge: Cambridge University Press,pp. 157–168.
Bornman, J. F. and Teramura, A. H. 1993.Effects of UV-B radiation on terres-trial plants. In: Young, A. R., Bjorn,L. O., Moan, J., and Nultsch, W. (Eds.)Environmental UV Photobiology. pp.427–471. Plenum Press, New York.
Boubriak, I., Kargiolaki, H., Lyne, L., andOsborne, D. J. 1997. The requirementfor DNA repair in desiccation toler-ance of germinating embryos. SeedScience Research 7:97–105.
Britt, A. B. 1996. DNA damage and repairin plants. Annu. Rev. Plant Physiol.Plant Mol. Biol. 47:75–100.
Britt, A. B. 1999. Molecular genetics ofDNA repair in higher plants. TIPS4:20–25.
Britt, A. B., Chen, J.-J., Wykoff, D., andMitchell, D. 1993. A UV-sensitivemutant of Arabidopsis defective in therepair of pyrimidine-pyrimidinone (6–4) dimers. Science 261:1571–1574.
Cannon, G. C., Hedrick, L. A., andHeinhorst, S. 1995. Repair mecha-nisms of UV-induced DNA damagein soybean chloroplasts. Plant Mol.Biol. 29:1267–1277.
Carpenter, C. L. and Cantley, L. C. 1996.Phosphoinositide kinases. Curr. Opin.Cell Biol. 2:153–158.
Cheah, K. S. E. and Osborne, D. J. 1978.DNA lesions occur with loss of viabil-
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

381
ity in embryos of ageing rye seeds.Nature 272:593–599.
Chen, J-J., Mitchell, D. L., and Britt, A. B.1994. A light-dependent pathway forthe elimination of UV-induced pyri-midine (6–4) pyrimidine photoprod-ucts in Arabidopsis. Plant Cell 6:1311–1317.
Chen, L., Trujillo, K., Sung, P., andTomkinson, A. E. 2000. Interactionsof the DNA ligase IV-Xrcc4 complexwith DNA ends and the DNA-depen-dent protein kinase. J. Biol. Chem. 275:26196–26205.
Chu, G. 1997. Double-strand break repair.J. Biol. Chem. 272:24097–24100.
Costanzo, V., Robertson, K., Ying, C.Y., Kim, E., Avvedimento, E.,Gottesman, M., Grieco, D., andGautier, J. 2000. Reconstitution ofan ATP-dependent checkpoint thatinhibits chromosomal DNA repli-cation following DNA damage. Mo-lecular Cell 6:649–659.
Coverley, D., Kenny, M. K., Lane, D. P.,and Wood, R. D. 1992. A role for thehuman single-stranded DNA bindingprotein HSSB/RPA in an early stageof nucleotide excision repair. NucleicAcids Res. 20:3873–3880.
Critchlow, S. E. and Jackson, S. P. 1998.DNA end-joining; from yeast to man.Trends Biochem. Sci. 23:394–498.
Culligan, K. M. and Hays, J. B. 1997.DNA mismatch repair in plants. AnArabidopsis thaliana gene that pre-dicts a protein belonging to theMSH2 subfamily of eukaryoticmutS homologs. Plant Physiol.115:833–839.
Dandoy, E., Schyns, R., Deltour, R., andVerly, W. G. 1987. Appearance andrepair of apurine/apyrimidinic sites inDNA during early germination of Zeamays. Mutation Res. 181:57–60.
De Laat, W. L., Jaspers, G. J., andHoeijmakers, J. H. J. 1999. Molecularmechanism of nucleotide excision re-pair. Gene Dev. 13:768–785.
Del Maestro, R. F., Bjork, J., and Arfors, K.E. 1981a. Increase in microvascularpermeability induced by enzymaticallygenerated free radicals. I. In vivo study.Microvasc. Res. 22:239–254.
Del Maestro, R. F., Bjork, J., and Arfors, K.E. 1981b. Increase in microvascularpermeability induced by enzymaticallygenerated free radicals. II. Role ofsuperoxide anion radical, hydrogenperoxide, and hydroxyl radical.Microvasc Res. 22:255–270
Demple, B. and Harrison, L., 1994. Repairof oxidative damage to DNA — enzy-mology and biology. Annu. Rev.Biochem. 63:915–948.
Dennis, C. and Surridge, C. 2000. A. thalianagenome. Nature 408:791.
De Vries, H. 1900. Sur la loi de disjuncitondes hybrides. C. R. Acad. Sc. Paris.130:845–847.
Dianov, G. L., Prasad, R., Wilson, S. H.,and Bohr, V. A. 1999. Role of DNApolymerase beta in the excision stepof long patch mammalian base exci-sion repair. J. Biol. Chem. 274:13741–13743.
Dizdaroglu, M. 1992. Measurement of ra-diation-induced damage to DNA at themolecular level. Int. J. Radiat. Biol.61:175–183.
Doetsch, P. W. 1995. What’s old is new: analternative DNA excision repair path-way. Trends Biochem. Sci. 20:384–386.
Doetsch, P. W., McCray, W. H., Jr., andValenzuela, M. R. 1989. Partial purifi-cation and characterisation of an endo-nuclease from spinach that cleaves ul-traviolet light-damaged duplex DNA.Biochim. Biophys. Acta 1007:309–317.
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

382
Dormandy, T.L. 1980. Free radical reactionin biological systems. Ann. Rev. Coll.Surg. Engl. 62:188–194.
Doutriaux, M-P., Couteau, F., Bergounioux,C., and White, C. 1998. Isolation andcharacterisation of the RAD51 andDMC1 homologs from Arabidopsisthaliana. Mol. Gen. Genet. 257:283–291.
Drapkin, R., Reardon, J. T., Ansari, A.,Huang, J. C., Zawel, L., Ahn, K.,Sancar, A., and Reinberg, D. 1994.Dual role of TFIIH in DNA exci-sion repair and in transcriptionby RNA polymerases II. Nature368:769–772.
Drew, M. C., He, C.-J., and Morgan,P. W.2000. Programmed cell death and aer-enchyma formaiton in roots. TrendsPlant Sci. 5:123–127.
Drummond, J. T., Li, G. M., Longley, M. J.,and Modrich, P. 1995. Isolation of anhMSH2–p160 heterodimer that re-stores DNA mismatch repair to tumorcells. Science 268:1909–1912.
Eastwood, A. C. and McLennan, A. G. 1985.Repair replication in ultraviolet light-irradiated protoplasts of Daucuscarota. Biochem. Biophys. Acta826:13–19.
Ehtesham, N. Z., Tuan-Nghia, P., Gaikwad,A., Sopory, S., and Tuteja, N. 1999.Calnexin from Pisum sativum: molecu-lar cloning of the cDNA and charac-terization of the encoded protein. DNACell Biol. 18:865–874.
Eisen, J. A. and Hanawalt, P. C. 1999. Aphylogenomic study of DNA repairgenes, proteins and processes. Mutat.Res. 435:171–213.
Elder, R. H., Dell’Aquila, A., Mezzina, M.,Sarasin, A., and Osborne, D. J. 1987.DNA ligase in repair and replicationin the embryos of rye, Secale cereale.Mutat. Res. 181:61–71.
Elder, R. H., and Osborne, D. J. 1993. Func-tion of DNA synthesis and DNA re-pair in the survival of embryos duringearly germination and in dormancy.Seed Sci. Res. 3:43–53.
Essrs, J., van Steeg, H., de Wit, J.,Swgagemaekrs, S. M. A., Vermeij, M.,Hoeijmkers, J. H. J., and Kanaar, R.2000. Homologous and non-homolo-gous recombination differentially af-fect DNA damage repair in mice.EMBO J. 19:1703–1710.
Esterbauer, H. and Cheeseman, K. H. 1990a.Determination of aldehydic lipidperoxidation products: malonaldehydeand 4–hydroxynonenal. MethodsEnzymol. 186:407–421.
Esterbauer, H., Dieber-Rotheneder, M.,Waeg, G., Puhl, H., and Tatzber, R.1990b. Endogenous antioxidants andlipoprotein oxidation. Biochem. Soc.Trans. 18:1059–1061.
Evans, E., Moggs, J. G., Hwang, J. R., Egly,J.-M., and Wood, R. D. 1997. Mecha-nisms of open complex and dual inci-sion formation by human nucleotideexcision repair factors. EMBO J.16:6559–6573.
Fee, J. A. 1982. In: Oxidases and RelatedRedox Systems (King, T.E., Hanson,T.S., and Morrison, M., Eds). PergamonPress, Oxford, pp. 101–132.
Fink, S. P., Reddy, G. R., and Marnett, L. J.1997. Mutagenicity in E. coli of the majorDNA adduct derived from the endog-enous mutagen malondialdehyde. Proc.Natl. Acad. Sci. USA 94:8652–8657.
Fortini, P., Pascucci, B., Parlanti, E., Sobol,R. W., Wilson, S. H., and Dogliotti, E.1998. Different DNA polymerases areinvolved in the short- and long-patchbase excision repair in mammaliancells. Biochemistry 37:3575–3580.
Fridovich, I. 1976. Oxygen radicals, hydro-gen peroxide and oxygen toxicity. In:
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

383
Free Radical in Biology (Pryor, W.A.,Ed), 1, Academic Press, NY, pp. 239–277.
Friedberg, E. C. 1985. DNA Repair. NewYork, W.H. Freeman.
Friedberg, E. C., Walker, G. C., and Siede,W. 1995. In: DNA Repair and Mu-tagenesis. ASM Press, WashingtonDC.
Friedberg, E. C. and Wood, R. D. 1996.DNA excision repair pathways. In:DNA Replication in Eukaryotic Cells.Cold Spring Harbor Laboratory Press,pp. 249–266.
Frosina, G., Fortini, P., Rossi, O., Carrozzino,F., Raspaglio, G., Cox, L. S., Lane, D.P., Abbondandolo, A., and Dogliotti,E. 1996. Two pathways for base exci-sion repair in mammalian cells. J. Biol.Chem. 271:9573–9578.
Gallego, F., Fleck, O., Li, A., Wyrzykowska,J., and Tinland, B. 2000. At RAD1, aplant homologue of human and yeastnucleotide excision repair endonu-cleases, is involved in dark repair ofUV damages and recombination. PlantJ. 21:507–518.
Gallego, M. E., Jeanneau, M., Granier, F.,Bouchez, D., Bechtold, N., and White,C. I. 2001. Disruption of the ArabidopsisRad50 gene leads to plant sterility andMMS sensitivity. Plant J. 25:31–41.
Gary, R., Ludwig, D., Cornelius, H.,MacInnes, M., and Park M. 1997. TheDNA repair endonuclease XPG bindsto proliferating cell nuclear antigen(PCNA) and shares sequence elementswith the PCNA-binding regions ofFEN-1 and cyclin-dependent kinase in-hibitor P21. J. Biol. Chem. 272:24522–24529.
George, J. W., Ghate, S., Matson, S. W.,and Besterman, J. M. 1992. Inhibitionof DNA helicase II unwinding andATPase activities by DNA-interact-
ing ligands. J. Biol. Chem. 267:10683–10689.
Gerlach, V. L., Aravind, L., Gotway, G.,Schultz, R. A., Koonin, E. V. andFriedberg, E. C. 1999. Human andmouse homologs of E. coli DinB(DNA polymerase IV), members ofthe UmuC/DinB superfamily. Proc.Natl. Acad. Sci. USA 21:11922–11927.
Gibbs, P. E., McGregor, W. G., Maer, V.M., Nisson, P., and Lawrence, C. W.1998. A human homolog of the Sac-charomyces cerevisiae REV3 gene,which encodes the catalytic subunit ofDNA polymerase zeta. Proc. Natl.Acad. Sci. USA 94:6876–6880.
Gidrol, X., Serghini, H., Noubhani, A.,Mocquot, B., and Pradet, P. 1989.Biochemical changes induced by ac-celerated aging in sunflower seeds. I.Lipid peroxidation and membranedamage. Physiologia Plantarum76:591–597.
Goodman, B. A., McPhail, D. B., andLnehan, D. J. 1986. Oxygen-inducedfree radical in wheat roots. Free Rad.Res. Commun. 2:173–178.
Goodman, M. E., Creighton, S., Bloom, L.B., and Petruska, J. 1993. Biochemi-cal basis of DNA replication fidelity.Crit. Rev. Biochem. Mol. Biol. 28:83–126.
Gorbalenya, A. E. and Koonin, E. V. 1993.Helicases: amino-acid sequence com-parisons and structure-function rela-tionships. Curr. Biol. 3:419–429.
Gorbunova, V. and Levy, A. A. 1997. Non-homologous DNA end-joining in plantcells is associated with deletions andfiller DNA insertions. Nucleic AcidsRes. 25:4650–4675.
Gorbunova, V. and Levy, A. A. 1999. Howplants make ends meet: DNA double-strand break repair. Trends Plant Sci.4:263–269.
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

384
Gorman, M. A., Morera, S., Rothwell, D.G., de La Fortelle, E., Mol, C. D.,Tainer, J. A., Hickson, I. D., andFreemon, P. S. 1997. The crystal struc-ture of the human DNA repair endo-nuclease HAP1 suggests the recogni-tion of extra-helical deoxyribose atDNA abasic sites. EMBO J. 16:6548–6558.
Grilley, M., Griffith, J., and Modrich, P.1993. Bidirectional excision in me-thyl-directed mismatch repair. J. Biol.Chem. 268:11830–11837.
Grilley, M., Welsh, K. M., Su, S. S., andModrich, P. 1989. Isolation and char-acterization of the E. coli mutL geneproduct. J. Biol. Chem. 264:1000–1004.
Gu, H., Marth, J. D., Orban, P. C.,Mossmann, H., and Rajewsky, K.1994. Deletion of a DNA polymerasebeta gene segment in T cells using celltype-specific gene targeting. Science265:103–106.
Halliwell, B. 1987. Oxidative damage, lipidperoxidation and antioxidant protec-tion in chloroplasts. Chem. Phys. Lip-ids 44:327–340.
Halliwell, B. 1990. How to characterize abiological antioxidant. Free Radic.Res. Commun. 9:1–32.
Halliwell, B. and Gutteridge, J. M. C. 1984.Oxygen toxicity, oxygen radicals, tran-sition metals and disease. Biochem. J.219:1–4.
Halliwell, B. and Gutteridge, J. M. C. 1986.Oxygen-free radicals and iron in rela-tion to biology and medicine: someproblems and concepts. Arch. Biochem.Biophys. 246:501–514.
Hang, B., Signer, B., Mrgison, G. P., andElder, R. H. 1997. Targeted deletionof alkylpurine-DNA-N-glycosylase inmice eliminates repair of 1,N6–ethenoadenine and hypoxanthine but
not of 3,N4–ethenocytosine or 8–oxoguanine. Proc. Natl. Acad. Sci.USA 94:12869–12874.
Harber, F. and Weiss, J. 1934. The catalyticdecomposition of hydrogen peroxideby iron salt. Proc. R. Soc. Lond. A.Math. Phys. Sci. 147:337–351.
Harlow, G. R., Jenkins, M. E., Pittalwala, T.S., and Mount, D. W. 1994. Isolationof uvh1, an Arabidopsis mutant hy-persensitive to ultraviolet light andionizing radiation. Plant Cell 6:227–235.
Harrington, J. J. and Lieber, M. R. (1994)Functional domains within Fen-1 andRad2 define a family of structure-spe-cific endonucleases; implications fornucleotide excision repair. Genes Dev.8:1344–1355.
He, Z., Henricksen, L. A., Wold, M. S., andIngles, C. J. 1995. RPA involvementin the damage-recognition and inci-sion step of nucleotide excision re-pair. Nature 374:566–569.
Heath, R. L. and Taylor, G. E., Jr. 1997.Physiological processes and plant re-sponses to ozone exposure. Ecol. Stud.127:317–368.
Hemler, M. E. and Lands, W. E. 1980. Evi-dence for a peroxide-initiated free radi-cal mechanism of prostaglandin biosyn-thesis. J. Biol. Chem. 255:6253–6261.
Hendry, G. A. F., Price, A. H., andBrocklebank, K. J. 1989. Role of ironin chlorophyll destruction in stressedplants. Mol. Aspects Med. 11:132–133.
Hidema, J., Kumagai, T., and Sutherland, B.M. 2000. UV radiation-sensitive norin1 rice contains defective cyclobutanepyrimidine dimer photolyase. PlantCell, 12:1569–1578.
Hidema, J., Kumagai, T., Sutherland, J. C.,and Sutherland, B. M. 1997. Ultravio-let B-sensitive rice cultivar deficient
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

385
in cyclobutyl pyrimidine dimer repair.Plant Physiol. 113:39–44.
Hodges, D. M., Andrews, C. J., Johnson, D.A., and Hamilton, R. I. 1997. Antioxi-dant enzyme and compound responsesto chilling stress and their combiningabilitites in differentially sensitivemaize hybrids. Crop Sci. 37:857–863.
Hoeijmakers, J. H. J. 1993a. Nucleotideexcision repair I: from E. coli to yeast.Trends Genet. 9:173–177.
Hoeijmakers, J. H. J. 1993b. Nucleotideexcision repair II. from yeast to mam-mals. Trends Genet. 9: 211–27.
Hoeijmakers, J. H. J. and Bootsma, D. 1990.Molecular genetics of eukaryotic DNAexcision repair. Cancer Cells 2:311–320.
Howland, G. P. 1975. Dark-repair of ultra-violet-induced pyrimidine dimers inthe DNA of wild carrot protoplasts.Nature 254:160–161.
Huang, J. C., Svododa, D. L., Reardon, J.T., and Sancar, A. 1992. Human nucle-otide excision nuclease removes thym-ine dimers from DNA by incising the22nd phosphodiester bond 5¢ and the6th phosphodiester bond 3¢ to thephotodimer. Proc. Natl. Acad. Sci.USA 89:3664–3668.
Hubscher, U. and Thommes, P. 1992. DNApolymerase: in search of a function.Trends Biochem. Sci. 17:55–58.
Hunter, N., Chambers, S. R., Louis, E. J.,and Borts, R. H. 1996. The mismatchrepair system contributes to meioticsterility in an interspecific yeast hy-brid. EMBO J. 15:1726–1733.
Ikenaga, M. and Mabuchi, T. 1996. Photo-reactivation of endosperm mutationsin maize. Radiat. Bot. 6:165–169.
Illades-Aguiar, B. and Setlow, P. 1994.Autoprocessing of the protease thatdegrades small, acid-soluble proteins
of spores of Bacillus species is trig-gered by low pH, dehydration, anddipicolinic acid. J. Bacteriol. 176:7032–7037.
Inze, D. and Van Montagu, M. 1995. Oxi-dative stress in plants. Curr. Opin.Biotechnol. 6:153–158.
Jabs, T. 1999. Reactive oxygen intermedi-ates as mediators of programmed celldeath in plants and animals. Biochem.Pharm. 57:231–245.
Jackson, J. F. 1987. DNA repair in pollen. Areview. Mutat. Res. 181:17–29.
Jackson, J. F. and Linskens, H. F. 1978.Evidence for DNA repair after ultra-violet irradiation of Petunia hybridapollen. Mol. Gen. Genet. 161:117–120.
Jansen, M. A., Gaba, V., and Greenberg, B.M. 1998. Higher plants and UV-B ra-diation: balancing damage, repair andacclimatation. Trends Plant Sci.3:131–135.
Jeggo, A. P., Taccioli, G. E., and Jackson,S. P. 1995. Menage a trois: doublestrand break repair, V(D)J recombina-tion and DNA-PK. Bioessays 17:949–957.
Jiang, C. Z., Yen, C.-N., Cronin, K.,Mitchell, D., and Britt, A. B. 1997.UV- and gamma-radiation sensitivemutants of Arabidopsis thaliana. Ge-netics 147:1401–1409.
Jin, H., Cominelli, E., Bailey, P., Parr, A.,Mehrtens, F., Jones, J., Tonelli, C.,Weisshaar, B., and Martin, C. 2000.Transcriptional repression by AtMyB4controls production of UV-protectingsunscreens in Arabidopsis. EMBO J.19:6150–6161.
Johnson, K. A., Fink, S. P., and Marnett, L. J.1997. Repair of propanodeoxyguanosineby nucleotide excision repair in vivo andin vitro. J. Biol. Chem. 272:11434–11438.
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

386
Johnson, R. E., Kondratick, C. M., Prakash,S., and Prakash, L. 1999a. hRAD30mutations in the variant form ofXeroderm pigmentosum. Science285:263–265.
Johnson, R. E., Kovvali, G. K., Prakash, L.,and Prakash, S. 1996. Requirement ofthe yeast Msh3 and Msh6 genes forMsh2–dependent genomic stability. J.Biol. Chem. 271:7285–7288.
Johnson, R. E., Prakash, S., and Prakash, L.1999b. Efficient bypass of a thymine-thymine dimer by yeast DNA poly-merase, Pol h. Science 283:1001–1004.
Johnson, R. E., Washington, M. T.,Haracska, L., Prakash, S., and Prakash,L. 2000a. Eukaryotic polymerases iotaand zeta act sequentially to bypassDNA lesions. Nature 406:1015–1019.
Johnson, R. E., Washington, M. T., Prakash,S., and Prakash, L. 1999c. Bridgingthe gap: a family of novel DNA poly-merases that replicate faulty DNA.Proc. Natl. Acad. Sci. USA 96:12224–12226.
Johnson, R. E., Washington, M. T., Prakash,S., and Prakash, L. 2000b. Fidelity ofhuman DNA polymerase P. J. Biol.Chem. 275:7447–7450.
Junop, M. S., Modesti, M., Guarne, A.,Ghirlando, R., Gellert, M., and Yang,W. 2000. Crystal structure of the Xrcc4DNA repair protein and implicationsfor end joining. EMBO J. 19:5962–5970.
Kaminaka, H., Morita, S., Tokumoto, M.,Masumura, T., and Tanaka, K. 1999.Differential gene expressions of ricesuperoxide dismutase isoforms to oxi-dative and environmental stresses.Free Radic. Res. 31 (Suppl.):S219–S225.
Kanofsky, J. R. and Sima, P. 1991. Singletoxygen production from the reactions
of ozone with biological molecules. J.Biol. Chem. 266:9039–9042.
Kehrer, J. P. 2000. The Haber-Weiss reac-tion and mechanisms of toxicity. Toxi-cology 149:43–50.
Kim, C., Snyder, R. O., and Wold, M. S.1992. Binding properties of replica-tion protein-A from human and yeastcells. Mol. Cell. Biol. 12:3050–3059.
Kley, D., Kleinman, M., Sandermann, H.,Jr., and Krupa, S. 1999. Photochemi-cal oxidants: State of the science.Environ. Pollut. 100:19–24.
Klimyuk, V. and Jones, J. 1996. At DMC1,the Arabidopsis homologue of theyeast DMC1 gene: characterization,transposon-induced allelic viriton andmeiosis-specific expression of apAtDMC1: GUS fusion. Plant J. 11:1–14.
Klungland, A. and Lindahl, T. 1997. Sec-ond pathway for completion of humanDNA base excision-repair; reconstitu-tion with purified proteins and require-ment for DNase IV (FEN1). EMBO J.16:3341.
Kobayashi, T., Hota, Y., and Tabata, S. 1993.Isolation and characterization of a yeastgene that is homologous with a meio-sis-specific cDNA from a plant. Mol.Gen. Genet. 237:225–232.
Kolodner, D. and Marsischky, G. T. 1999.Eukaryotic DNA mismatch repair.Curr. Opin. Genet. Dev. 9:89–96.
Kolodner, R. 1996. Biochemistry and ge-netics of eukaryotic mis-match repair.Genes Dev. 10:1433–1442.
Krokan, H. E., Standal, R., and Slupphaug,G. 1997. DNA glycosylases in the baseexcision repair of DNA. Biochem. J.325:1–16.
Kubota, Y., Nash, R. A., Klungland, A.,Schar, P., Brnes, D. E., and Lindahl,T. 1996. Reconstitution of DNA base
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

387
excision-repair with purified humanproteins: interaction between DNApolymerase beta and the XRCC1 pro-tein. EMBO J. 15:6662–6670.
Landry, L. G., Stapleton, A. E., Lim, J., Hofman,P., Hays, J., Walbot, V., and Last, R.1997. An Arabidopsis photolyase mutantis hypersensitive to ultraviolet-B radia-tion. Proc. Natl. Acad. Sci. USA 94:328–332.
Lawrence, C. W. and Hinkle, D. C. 1996DNA polymerase zeta and the controlof DNA damage-induced mutagenesisin eukaryotes. Cancer Surv. 28:21–31.
Lehmann, A. R. 1998. Dual functions ofDNA repair genes: molecular, cellularand clinical implications. BioEssays20:146–155.
Leprince, O., Deltour, R., Thorpe, P. C.,Atherton, N. M., and Hendry, G. A.1990. The role of free radicals andradical processing systems in loss ofdesiccation tolerance in germinatingmaize. New Phytol. 116:573–580.
Leprince, O., Hendry, G. A., Atherton, N.M., and Walters-Vertucci, C. W. 1996.Free radicals and metabolism associ-ated with the acquisition and loss ofdesiccation tolerance in developingseeds. Biochem. Soc. Trans. 24:451–455.
Levine, A., Tenhaken, R., Dixon, R., andLamb C. 1994. H2O2 form the oxida-tive burst orchestrates the plant hyper-sensitive response. Cell 79:583–93.
Li, L., Elledge, S. J., Peterson, C. A., Bales,E. S., and Legerski, R. J. 1994. Spe-cific association between the humanDNA repair proteins XPA and ERCC1.Proc. Natl. Acad. Sci. USA 91:5012–5016.
Lieber, M. R. 1991. Site-specific recombi-nation in the immune system. FASEBJ. 5:2934–2944.
Lieber, M. R. 1997. The FEN-1 family ofstructure-specific nucleases in eukary-otic DNA replication, recombinationand repair. BioEssays 19:233–240.
Lin, J. J., Phillips, A. M., Herst, J. E., andSancar, A. 1992. Active site of (A)BCexcinuclease. II. Binding, bending andcatalysis mutants of UvrB reveal adirect role in 3¢ and indirect role in 5¢incision. J. Biol. Chem. 267:17693–17700.
Lin, J. J. and Sancar, A. 1992. Active site of(A)BC excinuclease. I. Evidence for5’ incision by UvrC through a cata-lytic site involving Asp399, Asp438,Asp466 and His538 residues. J. Biol.Chem. 267:17688–17692.
Lin, W., Xin, H., Zhang, Y, Wu, X., andWang, Z. 1999. The human REV1gene codes for a DNA template-de-pendent dCMP transferase. Nucl. Ac-ids Res. 27:4468–4475.
Lindahl, T. 1993. Instability and decay ofthe primary structure of DNA. Nature362:709–710.
Lindahl, T. and Barnes, D. E. 1992. Mam-malian DNA ligases. Annu. Rev.Biochem. 61:251–281.
Lindahl, T. and Wood, R. D. 1999. Qualitycontrol by DNA repair. Science286:1897–1905.
Liu, L. and Wells, P. G. 1995. DNAoxidaiton as a potential molecularmechnaism mediating drug-in-duced birth defects: phenytoin andstructurally related teratogens ini-tiate the formation of 8–hydroxy-2¢-deoxyguanosine in vitro and invivo in murine maternal hepatic andembryonic tissues. Free Radic.Biol. Med. 19:639–648.
Liu, Z., Hossain, G. S., Islas-Osuna, M. A.,Mitchell, D. L., and Mount, D. W. 2000.Repair of UV damage in plants bynucleotide excision repair: Arabidopsis
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

388
UVH1 DNA repair gene is a homologof Saccharomyces cerevisiae RAD1.Plant J. 21:519–528.
Livingstone-Zatchej, M., Meier, A.,Suter, B., and Thoma, F. 1997.RNA polymerase II transcriptioninhibits DNA repair by photolyasein the transcribed strand of activeyeast genes. Nucl. Acids Res .25:3795–3800.
Luxford, C., Morin, B., Dean, R. T., andDavies, M. J. 1999. Histone H1 andother protein and amino acid hydrop-eroxides can give rise to free radicalsthat oxidize DNA. Biochem. J.344:125–134.
Marintchev, A., Mullen, M. A., Maciejewski,M. W., Pan, B., Gryk., M. R., andMullen, G. P. 1999. Solution structureof the single-strand break repair pro-tein XRCC1 N-terminal domain. Na-ture Struct. Biol. 6:884–893.
Marnett, L. J. 1999a Lipid peroxidation-DNA damage by malondialdehyde.Mutat. Res. 424:83–95.
Marnett, L. J. 1999b. Chemistry and biologyof DNA damage by malondialdehyde.IARC Sci. Publ. 150:17–27.
Masutani, C., Kusumoto, R., Yamada, A.,Dohmae, N., Yokoi, M., Yuasa, M.,Araki, M., Iwai, S., Takio, K., andHanaoko, F. 1999. The XPV (xero-derma pigmentosum variant) gene en-codes human DNA polyemrase h.Nature 399:700–704.
Masutani, C., Sugasawa, K., Yanagisawa, J.,Sonoyama, T., Ui, M., Enomoto, T.,Takio, K., Tanaka, K., van der Spek, P.J., Bootsman, D., Hoeijmakers, J. H. J.,and Hanaoka, F. 1994. Purification andcloning of a nucleotide excision repaircomplex involving the Xerodermapigmentosum gorup C protein and ahuman homolog of yeast Rad23. EMBOJ. 13:1831–1843.
Matsuda, T., Saijo, M., Kuraoka, I.,Kobayashi, T., Nahatssu, Y., Nagai,A., Enjoji, T., Masutani, C., Sugasawa,K., Hanaoka, F., Yasui, A., andTanaka, K. 1995. DNA repair proteinXPA binds to replication protein A(RPA). J. Biol. Chem. 270:4152–4157.
Matsumoto, Y. and Kim, K. 1995. Excisionof deoxyribose phosphate residues byDNA polymerase beta during DNArepair. Science 269:699–702.
Matsumoto, Y., Kim, K., and Bogenhagen,D. F. 1994. Proliferating cell nuclearantigen-dependent a basic site repairin Xenopus laevis oocytes: an alterna-tive pathway of base decision DNArepair. Mol. Cell. Biol. 14:6187–6197.
Matsunaga, T., Mu, D., Park, C. H., Reardon,J. T., and Sancar, A. 1995. HumanDNA repair excision nuclease. Analy-sis of the roles of the subunits involvedin dual incisions by using anti-XPGand anti-ERCC1 antibodies. J. Biol.Chem. 270:20862–20869.
Mattimore, V., and Battista, J. R. 1996.Radioresistance of Deinococcusradiodurans: functions necessary tosurvive ionizing radiation are alsonecessary to survive prolonged desic-cation. J. Bacteriol. 178:633–637.
Mazur, S. J. and Grossman, L. 1991. Dimer-ization of E. coli UvrA and its bindingto undamaged and ultraviolet light dam-aged DNA. Biochemistry 30:4432–4443.
McClennan, A. G. 1987. The repair of ultra-violet light-induced DNA damage inplant cells. Mutat. Res. 181:1–7.
McClintock, B. 1931. Cytological observa-tions of deficiencies involving knowngenes, translocations and an inversionin Zea mays. Mo. Agric. Exp. Res. Stn.Res. Bull. 163:4.
McDonald, J. P., Rapic-Otrin, V., Epstein,J. A., Broughton, B. C., Wang, X.,
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

389
Lehmann, A. R., Wolgemuth, D. J.,and Woodgate, R. 1999. Novel humanand mouse homologs of Saccharomy-ces cerevisiae DNA polymerase eta.Genomics 60:20–30.
Mehlhorn, H., Tabner, B., and Welburn, A.R. 1990. Electron spin resonance evi-dence for the formation of free radi-cals in plants exposed to ozone. Phys.Plant 79:377–83.
Mehlhorn, H. and Wellburn, A. R. 1987.Stress ethylene formation determinesplant sensitivity to ozone. Nature327:417–418.
Misra, H. P. and Fridovich, I. 1972. Therole of superoxide anion in theautooxidation of epinephrin and asimple asay for superoxide dismutase.J. Biol. Chem. 247:3170–3175.
Modesti, M. Hesse, J. E., and Gellert, M.1999. DNA binding of XRCC4 pro-tein is associated with V(D)J recom-bination but not with stimulation ofDNA ligase IV activity. EMBO J.18:2008–2018
Modrich, P. 1994. Mismatch repair, geneticstability, and cancer. Science 266:1959–1960.
Moggs, J. G., Yarema, K. J., Essigmann, J.M., and Wood, R. D. 1996. Analysisof incision sites produced by humancells extracts and purified proteinsduring nucleotide excision repair of a1, 3–intrastrand d(gpTpG)-C is platinadduct. J. Biol. Chem. 271:7177–7186.
Moore, M. H., Gulbis, J. M., Dodson, E. J.,Demple, B., and Moody, P. C. E. 1994.Crystal structure of a suicidal DNA re-pair protein: the Ada O6–methylguanine-DNA methyltransferase from E. coli.EMBO J. 13:1495–1501.
Mu, D., Hsu, D.S., and Sancar, A. 1996.Reaction mechanism of human DNArepair excision nuclease. J. Biol. Chem.271:8285–8294.
Mu, D., Park, C. H., Matsunaga, T., Hsu, D.S., Reardon, J. T., and Sancar, A. 1995.Reconstitution of human DNA repairexcision nuclease in a highly definedsystem. J. Biol. Chem. 270:2415–2418
Mu, D. and Sancar, A. 1997. Model for XPC-independent transcription-coupled re-pair of pyrimidine dimers in humans. J.Biol. Chem. 272:7570–7573.
Mu, D., Turson, M., Duckett, D. R.,Drummond, J. T., Modrich, P., andSancar, A. 1997a. Recognition andrepair of compound DNA lesions (basedamage and mismatch) by human mis-match repair and excision repair sys-tems. Mol Cell Biol. 17:760–769.
Mu, D., Wakasugi, M., Hsu, D. S., andSancar, A. 1997b. Characterization ofreaction intermediates of human exci-sion repair nuclease. J. Biol. Chem.272:28971–28979.
Mueller, J. P. and Smerdon, M. J. 1996.Rad23 is required for transcription-coupled repair and efficient overallrepair in Saccharomyces cerevisiae.Mol. Cell. Biol. 16:2361–2368.
Mustafa, M. G. and Tiernery, D.F. 1978.Biochemical metabolic changes in thelung and oxygen, ozone and nitrogendioxide toxicity. Ann. Rev. Resp. Dis.118:1061–1090.
Myles, G. M. and Sancar, A. 1991. Isolationand characterization of functional do-mains of uvrA. Biochemistry 30:3834–3840.
Nakajima, S., Sugiyama, M., Iwai, S.,Hitorni, K., Otoshi, E., Kim, S. T.,Jiang, C. Z., Todo, T., Britt, A. B., andYamamoto, K. 1998. Cloning andcharacterization of a gene (UVR3)required for photorepair of 6–4 prod-ucts in Arabidopsis thaliana. Nucl.Acids Res. 26:638–644.
Naqvi, A., Chance, B., and Cadenos, E. 1986.Reactive oxygen intermediates in bio-
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

390
chemistry. Ann. Rev. Biochem. 55:137–166.
Nelson, J. R., Lawrence, C. W., and Hinkle,D. C. 1996. Deoxycytidyl transferaseactivity of yeast REV1 protein. Na-ture 382:729–731.
Nuffer, M. G. 1957. Additional evidence onthe effect of X-ray and ultraviolet ra-diation on mutation in maize. Genet-ics 42:273–82.
O’Donovan, A., Davies, A. A., Moggs, J.G., West, S. C., and Wood, R. D. 1994.XPG endonuclease makes the 3¢ inci-sion in human DNA nucleotide exci-sion repair. Nature 371:432–435.
Ohba, T., Yoshioka, Y., Machida, C., andMachida, Y. 1995. DNA rearrange-ment associated with the integrationof T-DNA in tobacco: an example formultiple duplications of DNA aroundthe integration target. Plant J. 7:157–164.
Orzaez, D. and Granell, A. 1997. DNA frag-mentation is regulated by ethyleneduring carpel senescence in Pisumsativum. Plant J. 11:137–144.
Osborne, D. J. and Boubriak, I. 1994. DNAand desiccation tolerance. Seed Sci-ence Res. 4:175–185.
Overmyer, K., Tuominen, H., Kettunen, R.,Betz, C., Langebartels., C., Sundermann,H., and Kangasjarvi, J. 2000. Ozone-sensitive Arabodipsis rcd1 mutant re-veals opposite roles for ethylene andjasmonate signaling pathways in regu-lating superoxide-dependent cell death.Plant Cell 12:1849–1862.
Painter, R. B. 1985. Inhibition and recoveryof DNA synthesis in human cells afterexposure to ultraviolet light. Mutat.Res. 145:63–69.
Palombo, F., Gallinari, P., Iaccarino, I.,Lettieri, T., Hughes, M., DqArrigo,A., Truong, O., Hsuan, J. J., and
Jiricny, J. 1995. GTBP, a 160–kilodalton protein essential for mis-match-binding activity in human cells.Science 268:1912–1914.
Palombo, F., Iaccarino, I., Nakajima, E.,Ikejima, M., Shimada, T., and Jiricny,J. 1996. HMutSb, a heterodimer ofhMSH2 and hMSH3, binds to inser-tion/deletion loops in DNA. Curr. Biol.6:1181–1184.
Pang, Q. and Hays, J. B. 1991. UV-Binducible and temperature-sensitivephotorectivation of cyclobutane py-rimidine dimers in Arabidopsisthaliana. Plant Physiol. 95:536–543.
Parikh, S. S., Mol. C.D., Slupphaug, G.,Bharati, S., Krokan, H. E., and Tainer,J. A. 1998. Base excision repair initia-tion revealed by crystal structures andbinding kinetics of human uracil-DNAglycosylase with DNA. EMBO J.17:5214–5216.
Park, C-H. and Sancar, A. 1994. Formationof a ternary complex by human XPA,ERCC1 and ERCC4 (XPF) excisionrepair proteins. Proc. Natl. Acad. Sci.USA 91:5017–5021.
Park, H. W., Kim, S. T., Sancar, A., andDeisenhofer, J. 1995. Crystal struc-ture of DNA photolyase form E. coli.Science 268:1866–1872.
Pellinen, R., Palva, T., and Kangasjarvi, J.1999. Subcellular localization ofozone-induced hydrogen peroxide pro-duction in birch (Betula pendula) leafcells. Plant J. 20:349–356.
Petrini, J. H. J., Xiao, Y., and Weaver, D. T.1995. DNA ligase I mediates essentialfunctions in mammalian cells. Mol.Cell Biol. 15:4303–4308.
Pham, X. H., Reddy, M. K., Ehtesham, N.Z., Matta, B., and Tuteja, N. 2000. ADNA helicase from Pisum sativum ishomologous to translation initiation
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

391
factor and stimulates toposiomeraseinactivity. Plant J. 24:219–229.
Pieper, R. O. DNA Repair in Higher eukary-otes, In: DNA Damage and Repair, vol.2, J.A. Nickoloff and M. F. Hoekstra,Eds. (Humana Press, Totowa, NJ,1998), pp. 33–49.
Pinhero, R. G., Rao, M. V., Paliyath, G.,Murr, D. P., and Fletcher, R. A. 1997.Changes in activities of antioxidantenzymes and their relationship to ge-netic and paclobutrzol-induced chill-ing tolerance of maize seedlings. PlantPhysiol. 114:695–704.
Plooy, A. C., van Dijk, M., and Lohman, P.H. M. 1984. Induction and repair ofDNA cross links in Chinese hamsterovary cells treated with various plati-num coordination compounds in rela-tion to platinum binding to DNA, cy-totoxicity, mutagenicity and anti-tumoractivity. Cancer Res. 44:2043–2051.
Podust, L. M., Podust, V. A., Floth, C., andHubscher, U. 1994. Assembly of DNApolymerase d and e holoenzymes de-pends on the geometry of the DNAtemplate. Nucl. Acids Res. 22:2970–2975.
Price, A. H., Atherton, N. M., and Hendry,G. A. F. 1989a. Plants under drought-stress generate activated oxygen. FreeRad. Res. Commun. 8:61–66.
Price, A. H., Atherton, N. M., and Hendry,G. A. F. 1989b. Superoxide formationand enhanced transition-metal uptakein drought-damaged wheat leaves. In:Free Radicals, Diseased States andAntiradical Intervention (Ed. by C.Rice-Evans) pp. 467–479, RichelieuPress, London.
Price, A. H. and Hendry, G. A. F. 1989.Stress and the role of activated oxygenscavengers and protective enzymes inplants subjected to drought. Biochem.Soc. Trans. 17:493–494.
Priestley, D. A., Werner, B. G., Leopold, A.C., and McBride, M. G. 1985. Or-ganic free radicals in seeds and pol-len: the effect of hydration and aging.Physiol. Plant. 64:88–94.
Protic-Sabljic, M. and Kraemer, K. H. 1985.One pyrimidine dimer inactivates ex-pression of a transfected gene in Xero-derma pigmentosum cells. Proc. Natl.Acad. Sci. USA 82:6622–6626.
Protic-Sabljic, M., Tuteja, N., Munson, P.J., Hauser, J., Kraemer, K. H., andDixon, K. 1986. UV light-inducedcyclobutane pyrimidine dimers aremutageneic in mammalian cells. Mol.Cell. Biol. 6:3349–3356.
Pryor, W.A. 1986. Oxy radical related spe-cies: their formation, life time and re-actions. Ann. Rev. Physiol. 48:657–667.
Puchta, H. and Hohn, B. 1991. The mecha-nism of extrachromosomal homolo-gous DNA recombination in plantcells. Mol. Gen. Genet. 230:1–7.
Quaite, F. E., Takayananagi, S., Ruffini, J.,Sutherland, J. C., and Sutherland, B.M. 1994. DNA damage levels deter-mine cyclobutyl pyrimidine dimer re-pair mechanisms in alfalfa seedlings.Plant Cell 6:1635–1641.
Rao, M. V. and Davis, K. R. 1999. Ozone-induced cell death occurs via two dis-tinct mechanisms in Arabidopsis: therole of salicylic acid. Plant J. 17:603–614.
Reardon J. T., Mu D., and Sancar, S. 1996.Overproduction, purification andcharacterisation of the XPC subunit ofthe human DNA repair excision nu-clease. J. Biol. Chem. 270:2415–2418.
Reuzeau, C. and Cavalie, G. 1995. Activi-ties of free radical processing enzymesin dry sunflower seeds. New Phytol.130:59–66.
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

392
Ribeiro, D. T., Machdo, C. R., Costa, R. M.A., Praekelt, U. M., Van-sluys, M. A.and Menck, C. F. M. 1998. Cloning ofa cDNA from Arabidopsis thalianahomologous to the human XPB gene.Gene 208: 207–213.
Rinehart, T. A., Dean, C., and Weil, C. F.1997. Comparative analysis of non-random DNA repair following Actransposon excision in maize andArabidopsis. Plant J. 12:1419–1427.
Rosen, D. R., Siddiqui, T., and Patterson, D.1993. Mutation in Cu/Zn superoxidedismutse gene are associated with fa-milial amyotrophic lateral schlerosis.Nature 362:59–62.
Roy, R., Schaeffer, L., Humber, S.,Vermeulen, W., Weeda, G., and Egly,J.-M. 1994. The DNA-dependentATPase activity associated with theclass II basic transcription factor BTF2/TFIH. J. Biol. Chem. 269:9862–9832.
Saijo, M., Kuraoka, I., Masutani, C.,Hanaoka, F., and Tanaka, K. 1996.Sequential binding of DNA repair pro-teins RPA and ERCC1 to XPA in vitro.Nucl. Acids Res. 24:4719–4724.
Sakumi, K. and Sekiguchi, M. 1990. Struc-tures and functions of DNA glycoslases.Mutat. Res. 236:161–62.
Salomon, S. and Puchta, H. 1998. Captureof genomic and T-DNA sequencesduring double-strand break repair insomatic plant cells. EMBO J. 17:6086–6095
Sancar, A. 1994. Structure and function ofDNA photolyase. Biochemistry 33:2–9.
Sancar, A. 1996. No ‘end of history’ forphotolyases. Science 272:48–49.
Sancar, A. and Rupp, W. D. 1983. A novelrepair enzyme: UVrABC excisionnuclease of Escherichia coli cuts aDNA strand on both sides of the dam-aged region. Cell 33:349–260.
Sancar, G. B. 1990. DNA photolyases:physical properties, action mechanismand roles in dark repair. Mutat. Res.236:147–160.
Sandermann, H., Jr., Ernst, D., Heller, W.,and Langebartels, C. 1998. Ozone: anabiotic elicitor of plant defence reac-tions. Trends Plant Sci. 3:47–50.
Santerre, A. and Britt, A. B. 1994. Cloningof a 3–methyladenine-DNA glycoslasefrom Arabidopsis thaliana. Proc. Natl.Acad. Sci. USA 91:2240–2244.
Saparbaev, M. and Laval, J. 1998. 3,N4–ethenocytosine, a highly mutagenicadduct, is a primary substrate for E.coli double-stranded uracil-DNAglycosylase and human mismatch-spe-cific thymine-DNA glycosylase. Proc.Natl. Acad. Sci. USA 95:8508–8513.
Satoh, M. S. and Lindahl, T. 1992. Role ofpoly(ADP-ribose) formation in DNArepair. Nature 356:356–358.
Satoh, M. S., Poirier, G. G., and Lindahl, T.1993. NAD(+)-dependent repair ofdamaged DNA by human cell extracts.J. Biol. Chem. 268:5480–5487.
Schaeffer L., Moncollin, V., Roy, R., Staub,A., Mezzina, M., Sarasin, A., Weeda,G., Hoeijmakers, J. H. J., and Egly, J-M. 1994. The ERCC2 DNA repairprotein is associated with the class IIBTF2/TFIIH transcription facotr.EMBO J. 13:2388–2392.
Schaeffer, L., Roy, R., Humbert, S., Moncollin,V., Vermeulen, W., Hoeijmakers, J. H.J., Chambon, P., and Egly, J-M. 1993.DNA repair helicase: a component ofBTF2 (TFIIH) basic transcription fac-tor. Science 260:58–63.
Schraudner, M., Moeder, W., Wiese, C.,van Camp, W., Inze, D., Langebarels,C., and Sandermann, H., Jr. 1998.Ozone-induced oxidative burst in theozone biomonitor plant, tobacco BelW3. Plant J. 16:235–245.
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

393
Schultz, T. F. and Quatrano, R. S. 1997.Characterisation and expression of arice RAD23 gene. Plant Mol. Biol.34:557–562.
Scott, L., Lafoe, D., and Weil, C. F. 1996.Adjacent sequences influence DNArepair accompanying transposon exci-sion in maize. Genetics 142:237–246.
Seeberg, E., Eide, L., and Bjoras, M. 1995.The base excision repair pathway.Trends Biochem. Sci. 20:391–397.
Selby, C. P. and Sancr, P. 1993. Moelcularmechanism of transcription-repair cou-pling. Science 260:53–57.
Senaratna, T. and McKersie, B. D. 1986.Loss of desiccation tolerance duringseed germination: a free radical mecha-nism of injury. In: Membranes, Me-tabolism and Dry Organisms (Ed. byA. C. Leopold), pp. 85–102, CornellUniversity Press, Ithaca.
Seroussi, E. and Lavi, S. 1993. Replicationprotein A is the major single-strandedDNA binding protein detected in mam-malian cell extracts by gel retardationassays and UV cross-linking of longand short single-stranded DNA mol-ecules. J. Biol. Chem. 268:7147–7154.
Setlow, B. and Setlow, P. 1996. Role ofDNA repair in Bacillus subtilis sporeresistance. J. Bacteriol. 178:3486–3495.
Setlow, P. 1992a. I will survive: protectingand repairing spore DNA. J. Bacteriol.174:2737–2741.
Setlow, P. 1992b DNA in dormant spores ofBacillus species is in an A-like con-formation. Mol. Microbiol. 6:563–567.
Sharma, Y. K. and Davis, K. R. 1997. Theeffects of ozone on anti-oxidant re-sponses in plants. Free Radical Biol.Med. 23:480–488.
Shi, L., Kent, R., and Bence, A. B. 1997.Developmental expression of a DNA
repair gene in Arabidopsis. Mutat. Res.384:145–146.
Shiloh, Y. 1997. Ataxia-telangiectasia andthe Nijmegen breakage syndrome: re-lated disorders but genes apart. Annu.Rev. Genet. 31:635–662.
Shivji, M. K. K., Kenny, M. K., and Wood,R. D. 1992. Proliferating cell nuclearantigen is required for DNA excisionrepair. Cell 69:367–374.
Shivji, M. K. K., Podust, V. N., Hubscher,Y., and Wood, R. D. 1995. Nucleotideexcision repair DNA synthesis byDNA polymerase epsilon in the pres-ence of PCNA, RFC and RPA. Bio-chemistry 34:5011–5017.
Sies, H. 1991. Oxidative Stress: Oxidantand Antioxidants. Academic Press,London.
Sobol, R. W., Horton, J. K., Kuhn, R., Gu.H., Singhal, R. K., Prasad, R.,Rajewsky, K., and Wilson, S. H. 1996.Requirement of mammalian DNApolymerase-beta in base-excision re-pair. Nature 379:183.
Stapleton, A. E. 1992. Ultraviolet radiationand plants: burning questions. PlantCell 4:1353–1358.
Stapleton, A. E., Thorner, C. S., and Walbot, V.1997. UV-B component of sunlightcauses measurable damage in field-grownmaize (Zea mays L.): developmental andcellular heterogeneity of damage and re-pair. Plant Cell Environ. 20:279–290.
Sugasawa, K., Ng, J. M. Y., Masutani, C.,van der Spek, P. J., Eker, A. P. M.,Hanaoka, F., Bootsma, D., andHoeijmakers, J. H. J. 1998. Xerodermapigmentosum group C is the initiatorof global genome nucleotide excisionrepair. Mol. Cell 2:223–232.
Sugawara, N. and Haber, J. E. 1992. Char-acterization of double-strand break-induced recombination: homology
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

394
requirements and single-strandedDNA formation. Mol. Cell Biol.12:563–575.
Sung, P., Bailly, V., Weber, C., Prakash, L.,and Prakash, S. 1993. Human xerodermapigmentosum group D gene encodes aDNA helicase. Nature 365, 852–855.
Suter, B., Livingstone-Zatchej, M., andThoma, F. 1997. Chromatin structuremodulates DNA repair by photolyasein vivo. EMBO J. 16:2150–2160.
Sutherland, B. M., Takayanagi, S., Sullivan,J. H., and Sutherland, J. C. 1996. Plantresponses to changing environmentalstress: cylcobutyl pyrimidine dimerrepair in soybean leaves. Photochem.Photobiol. 64:464–468.
Svoboda, D. L., Taylor, J. S., Hearst, J. E., andSancar, A. 1993. DNA repair by eu-karyotic excision nuclease: removal ofthymine dimer and psoralen monoadductby HeLa cell free extract and of thyminedimer by Xenopus laevis oocytes. J. Biol.Chem. 268:1931–1936.
Swagemakers, S.M.A., Essers, J., Wit, J. De.,Hoeijmakers, J.H.J., and Kanaar, R. 1998.The human Rad 54 recombinatorial DNArepair protein is a double-stranded DNA-dependent ATPase. J. Biol. Chem.273:28292–28297.
Takata, M., Sasaki, M.S., Sonoda, E.,Morrison, C., Hashimoto, M., Utsumi,H., Yamaguchi-Iwai, Y., Shinohara,A., and Takeda, S. 1998. Homologousrecombination and non-homologousend-joining pathways of DNA double-strand break repair have overlappingroles in the maintenance of chromo-somal integrity in vertebrate cells.EMBO J. 17:5497–5508.
Takeuchi, Y., Murakami, M., Nakaima, N.,Kondo, N., and Nikaido, O. 1996. In-duction and repair of damage to DNAin cucumber cotyledons irradiated withUVB. Plant Cell Physiol. 37:181–187.
Talpaert-Borle, M. and Liuzzi, M. 1982.Base-excision repair in carrot cells.Eur. J. Biochem. 124:435–440.
Tamada, T., Kitadokoro, K., Higuchi, Y.,Inaka, K., Yasui, A., de Ruiter, P. E.,Eker, A. P., and Miki, K. 1997. Crys-tal structure of DNA photolyase fromAnacystis nidulans. Nature Struct.Biol. 4:887–891.
Tang, M., Shen, X., Frank, E. G., O’Donnell,M., Woodgate, R., and Goodman, M.F. 1999. UmuD’(2)C is an error-proneDNA polymerase, E. coli pol V. Proc.Natl. Acad. Sci. USA 96:8919–8924.
Tappel, A. L. 1973. Lipid peroxidation dam-age to cell components. Fed Proc.32:1870–1874.
Taylor, R.M. 1996. Ultraviolet B-inducedDNA lesions and their removal inwheat (Triticum aestivum L.) leaves.Plant Cell Environ. 19:171–181.
Taylor, R. M., Nikaido, O., Jordan, B. R.,Bray, C. M., and Tobin, A. K. 1996a.UV-B-induced DNA lesions and theirremoval in wheat (Triticum aestivumL.) leaves. Plant Cell Environ. 19:171–181.
Taylor, R. M., Rosamond, J., Tobin, A. K.,and Bray, C. M. 1996b. The cloningand sequence analysis of DNA ligasefrom Arabidopsis thaliana. AccesionNumber:1359495.
Teo, I., Sedgwick, B., Demple, B., Li, B.,and Lindahl, T. 1984. Induction ofresistance to alkylating agents in E.coli: the ada+ ne product serves bothas a regulatory protein and as an en-zyme for repair of mutagenic damage.EMBO J 3:2151–2157.
Teramura, A. H. 1983. Effects of ultraviolet-B radiation on the growth and yield ofcrop plants. Physiol. Plant 58:415–427.
Teramura, A. H. and Sullivan J. H. 1994.Effects of UV-B radiation on photo-
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

395
synthesis and growth of terrestrialplants. Photosynth. Res. 39:463–473.
Terasawa, M., Shinohara, A., Hotta, Y.,Ogawa, H., and Ogawa, T. 1995. Lo-calization of RecA-like recombinationproteins on chromosomes of the lily atvarious meiotic stages. Genes Dev.9:925–934.
Tevini, M. and Teramura, A. H. 1989. UV-B effects on terrestrial plants.Photochem. Photobiol. 50:479–487.
The Arabidopsis Genome Initiative. 2000.Analysis of the genome sequence ofthe flowering plant Arabidopsisthaliana. Nature 408:796–815.
Thoma, F. 1999. Light and dark in chroma-tin repair: repair of UV-induced DNAlesions by photolyase and nucelotideexcision repair. EMBO J. 18:6585–6598.
Todo, T. 1999. Functional diversity of theDNA photolyase blue light receptorfamily. Mutat. Res. 434:89–97.
Trosko, J. E., and Mansour, V. H. 1969.Photoreactivation of ultraviolet light-induced pyrimidine dimers in Ginkgocells grown in vitro. Mutat. Res. 7:120–121.
Turchi, J. J., Murante, R. S., and Bambara,R. A. 1992. DNA substrate specificityof DNA helicase E from calf thymus.Nucleic Acids Res. 20:6075–6080.
Tuteja, N. 1997. Unraveling DNA helicasesfrom plant cells. Plant Mol. Biol.33:947–952.
Tuteja, N. 2000. Plant Cell and viralhelicases: essential enzymes fornucleic acid transactions. Crit. Rev.Plant Sci. 19:449–478.
Tuteja, N., Beven, A. F., Shaw, P. J., andTuteja, R. 2001. A pea homologue ofhuman DNA helicase I is localizedwithin the dense fibrillar componentof the nucleolus and sitmulated by
phosphorylation with CK2 and cdc2protein kinases. Plant J. 25:9–17.
Tuteja, N., Ochem, A., Taneja, P., Tuteja,R., Skopac, D., and Falaschi, A. 1995.Purification properties of human DNAhelicase VI. Nucl. Acids Res. 23:2457–2463.
Tuteja, N. and Phan, T. N. 1998. A chloro-plast DNA helicase II from pea thatprefers fork-like structures. PlantPhysiol. 118:1029–1039.
Tuteja, N., Phan, T-N., and Tewari, K. K.1996. Purification and characterizationof a DNA helicase from pea chloro-plast that translocates in the 3’ to 5’direction. Eur. J. Biochem. 238:54–63.
Tuteja, N., Phan, T-N., Tuteja, R., Ochem,A., and Falaschi, A. 1997. Inhibitionof DNA unwinding and ATPase ac-tivities of human DNA helicase II bychemotherapeutic agents. Biochem.Biophys. Res. Commun. 236:636–640.
Tuteja, N., Rahman, K., Tuteja, R., andFalaschi, A. 1991. DNA helicase IVfrom HeLa cells. Nucl. Acids Res.19:3613–3618.
Tuteja, N., Rahman, K., Tuteja, R., andFalaschi, A. 1993. Human DNAhelicase V, a novel DNA unwindingenzyme form HeLa cells. Nucl. AcidsRes. 21:2323–23329.
Tuteja, N. Rahman, K., Tuteja, R., Ochem,A., Skopac, D., and Falaschi, A. 1992.DNA helicase III from HeLa cells: anenzyme that acts preferentially on par-tially unwound DNA duplexes. Nucl.Acids Res. 20:5329–5337.
Tuteja, N., and Tuteja, R. 1996. DNAhelicases: the long unwinding road.Nature Genetics 13:11–12.
Tuteja, N. and Tuteja, R., 2001. UnravelingDNA repair in human: molecularmechanisms and consequences of
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

396
repair defect. Crit. Rev. Biochem. Mol.Biol. (in press).
Tuteja, N., Tuteja, R., Ochem, A., Taneja, P.,Huang, N. W., Simoncsits, A., Susic, S.,Rahman, R., Marusic, L., Chen, J.,Zhang, J., Wang, S., Pongor, S., andFalaschi, A. 1994. Human DNA helciaseII: a novel DNA unwinding enzymeidentified as the Ku autoantigen. EMBOJ. 13:4991–5001.
Tuteja, N., Tuteja, R., Rahman, K., Kang,L.-Y., and Falaschi, A. 1990. A DNAhelicase from human cells. Nucl. Ac-ids Res. 18:6788–6792.
Tuteja, R. and Tuteja, N. 2000. Kuautoantigen: a multifunctional DNAbinding protein. Crit. Rev. Biochem.Mol. Biol. 35:1–33.
Urdea, M. S., Warner, B. D., Running, J. A.,Stempien, M., Clyne, J., and Horn, T.1988. A comparison of nonradioisotopichybridization assay methods usingfluorescen, chemiluminescent and en-zyme labeled synthetic oligonucleotideprobes. Nucl. Acids Res. 16:4937–4955.
Van Camp, W., Van Montagu, M., and Inze,D. 1994. Superoxide dismutases. InCausesof Photooxidative stress andamerlioration of defense systems inplants. Foyer, C. H. and Mullineaux,P. M., Eds., pp. 317–341. CRC Press,Boca Raton, FL.
Vaughan, P., Lindahl, T., and Sedgwick, B.1993. Induction of the adaptive re-sponse of E. coli to alkylation damageby the environmental mutagen, me-thyl chloride. Mutat. Res. 293:249.
Verhage, R. A., Zeeman, A. M., Lombaerts,M., van de Putte, P., and Brouwer, J.1996. Analysis of gene- and strand-spe-cific repair in the moderately UV-sensi-tive Saccharomyces cerevisiae rad23mutant. Mutat. Res. 362:155–165.
Villani, G., and Tanguy Le Gac, N. 2000.Interactions of DNA helicases with
damaged DNA: possible logical con-sequences. J. Biol. Chem. 275:33185–33188.
Vodenicharov, M. D., Sallmann, F. R.,Satoh, M. S., and Poirier, G.G. 2000.Base excision repair is efficient in cellslacking poly(ADP-ribose) polymerase1. Nucl. Acids Res. 28:3887–3896.
Vonarx, E. J., Mitchell, H. L., Karthikeyan,R., Chatterjee, I., and Kunz, B. A.1998. DNA repair in higher plants.Mutat. Res. 400:187–200.
Wagner, J., Gruz, P., Kim, S.R., Yamada,M., Matsui, K., Fuchs, R.P., andNohmi, T. 1999. The dinB gene en-codes a novel E. coli DNA polymerase,DNA pol IV, involved in mutagen-esis. Mol. Cell. 4:281.
Washington, M. T., Johnson, R. E., Prakash, S.,and Prakash, L. 1999. Fidelity andprocessivitiy of Saccharomyces cerevisiaeDNA polymerase n. J. Biol. Chem.274:36835–36838.
Washington, M. T., Johnson, R. E., Prakash,S., and Prakash, L. 2000. Accuracy ofthymine-thymine dimer bypass by Sac-charomyces cerevisiae DNA poly-merase h. Proc. Natl. Acad. Sci. USA97:3094–3099.
Weisinger, R.A. and Fridovich, I. 1973.Mitochondrial superoxide dismutasesite of synthesis and intermitochondriallocaliztion. J. Biol. Chem. 248:4793–4796.
White, C. I. and Haber, J. E. 1990. Interme-diates of recombination during matingtype switching in Saccharomycescerevisiae. EMBO J 9:663–673.
Wilson, D. M. and Thompson, L. H. 1997.Life without DNA repair. Proc. Natl.Acad. Sci. USA 94:12754–12757.
Winkler, G., Vermeulen, W, Coin, F., Egly,J., Hoeijmakers, J., and Weeda, G.1998. Affinity purification of human
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.

397
DNA repair transcription factor TFIIHusing epitope-tagged Xerodermapigmentosum B protein. J. Biol. Chem.273:1092–1098.
Wise, R. R. and Naylor, A. W. 1987. Chill-ing-enhanced photooxidation. Theperoxidative destruction of lipids dur-ing chilling injury to photosynthesisand ultrastructure. Plant Physiol.83:272–277.
Wood, R. D. 1997. Nucleotide excision re-pair in mammalian cells. J. Biol. Chem.272:23465–23468.
Wood, R. D., Mitchell, M., Sgouros, J., andLindahl, T. 2001. Human DNA repairgenes. Science, 291:1284–1289.
Wu, X., Li, J., Li, X., Hsieh, C. L., Burgers,P. M., and Lieber, M. R. 1996. Pro-cessing of branched DNA intermedi-ates by a complex of human FEN-1and PCNA. Nucl. Acids Res. 24:2036–2043.
Wymann, M.P. and Pirola, L. 1998. Struc-ture and function of phosphoinositide3–kinases. Biochim. Biophys. Acta1436:127–150.
Xu, H., Swobodo, I., Bhalla, P. L., Sijbers, A.M., Zhao, C., Ong, E.-K., Hoeijmakers,J. H. J., and Singh, M. B. 1998 Planthomologue of human excision repair
gene ERCC1 points to conservation ofDNA repair mechanisms. Plant J.13:823–829.
Yang, H., Clendenin, W. M., Wong, D.,Demple, B., Slupska, M. M., Chiang,J-H., and Miller, J. H. 2001. Enhancedactivity of adenine-DNA glycosylase(Myh) by AP endonuclease (Ape1) inmammalian base excision repair of anA/GO mismatch. Nucl. Acids Res.29:743–752.
Yasui, A. and Eker, A. P. M. 1998. DNAphotolyases. In: DNA Damage andRepair, vol. II (Nickoloff, J. A. andHoekstra, M. F., Eds.) Totowa:Humana Press, pp. 9–32.
Yasui, A., Eker, A. P. M., Yasuhira, S.,Yajima, H., Kobayashi, T., Takao, M.,and Oikawa, A. 1994. A new class ofDNA photolyases present in variousorganisms including aplacental mam-mals. EMBO J. 13:6143–6151.
Young, T. E., Gallie, D. R., and Demason,D. A. 1997. Ethylene-mediated pro-grammed cell death during maize en-dosperm development of wild-type andshrunken2 genotypes. Plant Physiol.115:737–751.
Zhou, B.-B. and Elledge, S. J. 2000. The DNAdamage response: putting checkpointsin perspective. Nature 408:433–439.
Cri
tical
Rev
iew
s in
Bio
chem
istr
y an
d M
olec
ular
Bio
logy
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
117.
215.
129.
173
on 0
8/01
/11
For
pers
onal
use
onl
y.
























![SES Refinery Damage Mechanisms Symposium AUG2004[1]](https://img.dokumen.tips/doc/110x75/5571fc48497959916996e6eb/ses-refinery-damage-mechanisms-symposium-aug20041.jpg)




