Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY (0 1992 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 267, No. 19, Issue of July 5, pp. 13229-13238,1992 Printed in U.S A .
Molecular Cloning of Human Mevalonate Kinase and Identification of a Missense Mutation in the Genetic Disease Mevalonic Aciduria*
(Received for publication, January 28, 1992)
Beverly L. SchaferS, Richard W. Bishop, Valerie J. Kratunis, Stephen S. Kalinowski, Stephen T. Mosley, K. Michael Gibson§, and Richard D. Tanakan From the Department of Metabolic Diseases, the Bristol-Myers Squibb Pharmaceutical Research Institute, Princeton, New Jersey 08543-4000 and the §Kimberly H. Courtwright and Joseph W. Summers Metabolic Disease Center, Baylor University Medical Center, Dallas, Texas 75246
Mevalonic aciduria is the first proposed inherited disorder of the cholesterol/isoprene biosynthetic path- way in humans, and it is presumed to be caused by a mutation in the gene coding for mevalonate kinase. To elucidate the molecular basis of this inherited disorder, a 2.0-kilobase human mevalonate kinase cDNA clone was isolated and sequenced. The 1188-base pair open reading frame coded for a 396-amino acid polypeptide with a deduced M, of 42,450. The predicted protein sequence displayed similarity to those of galactokinase and the yeast RARl protein, indicating that they may belong to a common gene family. Southern hybridiza- tion studies demonstrated that the mevalonate kinase gene is located on human chomosome 12 and is a single copy gene. No major rearrangements were detected in the mevalonic aciduria subject. The relative size (2 kilobases) and amounts of human mevalonate kinase mRNA were not changed in mevalonic aciduria fibro- blasts. Approximately half of the mevalonic aciduria cDNA clones encoding mevalonate kinase contained a single base substitution (A to C) in the coding region at nucleotide 902 that changed an asparagine residue to a threonine residue. The presence of this missense mu- tation was confirmed by polymerase chain reaction amplification and allele-specific hybridization of the genomic DNAs from the proband and the proband’s father and brother. Similar analysis failed to detect this mutation in the proband’s mother, seven normal subjects, or four additional mevalonic aciduria sub- jects, indicating that the mutation does not represent a common gene polymorphism. Functional analysis of the defect by transient expression confirmed that the mutation produced an enzyme with diminished activ- ity. Our data suggest that the index case is a compound heterozygote for a mutation in the mevalonate kinase gene.
Cholesterol is an essential component of cellular mem-
* The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The nucleotide sequence(s) reported in thispaper has been submitted to the GenBankTM/EMBL Data Bank with accession number(s) “ 3 4 6 8 .
$ To whom correspondence should be addressed Dept. of Metabolic Diseases, the Bristol-Myers Squibb Pharmaceutical Research Inst., P . 0. Box 4000, Princeton, NJ 08543-4000. Tel.: 609-252-6347; Fax:
B Present address: Dept. of Molecular Pharmacology, Arris Phar- 609-252-6802.
maceutical Corp., South San Francisco, CA 94080.
branes as well as a precursor for steroid hormones, vitamin D, and bile acids. Regulation of cholesterol biosynthesis oc- curs principally at the step catalyzed by 3-hydroxy-3-meth- ylglutaryl-CoA reductase (1). However, other enzymes in the pathway are also involved in regulation of cholesterol biosyn- thesis. Recent evidence indicates that mevalonate kinase (EC 2.7.1.36; ATP(R)-mevalonate 5-phosphotransferase), the first enzyme in the pathway to follow 3-hydroxy-3-methylglutaryl- CoA reductase, can play an important regulatory role because its activity may be controlled by feedback inhibition (2-5). Geranyl pyrophosphate and farnesyl pyrophosphate (FPP)’ are distal intermediates in the cholesterol biosynthetic path- way that inhibit mevalonate kinase activity by binding com- petitively at the ATP-binding site on the enzyme (2, 3). This feedback inhibition is thought to function in the regulation of the intracellular concentration of FPP. When intracellular levels of FPP are elevated, feedback inhibition would block mevalonate kinase activity and curtail further FPP synthesis. Regulation of the intracellular concentration of FPP is im- portant because FPP is a key intermediate at a branch point in the pathway. In mammalian cells, FPP is a precursor for the biosynthesis of many vital cellular components such as cholesterol, dolichols, ubiquinones, farnesylated proteins, and heme A (1, 6). Therefore, feedback regulation of mevalonate kinase activity may serve to maintain the size of one or more intracellular pools of FPP that are used by multiple biosyn- thetic pathways.
A mutation in the gene coding for mevalonate kinase is presumed to cause the genetic disease mevalonic aciduria, which is the first proposed inherited disorder of the choles- terol biosynthetic pathway in humans (7, 8). Mevalonic aci- duria is transmitted as an autosomal recessive trait, and there are at least seven reported cases of this genetic disease (7, 9, 10). Cells taken from subjects with mevalonic aciduria have <lo% of the normal levels of mevalonate kinase activity (9, l l ) , and all subjects have massive accumulation of mevalonate in both their serum and urine (7,8,10). In spite of the marked reduction in mevalonate kinase activity, subjects with meva- lonic aciduria have nearly normal levels of serum cholesterol (7,8). Cells taken from these subjects also appear to synthe- size nearly normal amounts of cholesterol (11,12). These data suggest that either the small residual amount of mevalonate kinase activity in these cells is capable of maintaining normal rates of cholesterol biosynthesis or that a mutation in the enzyme causes rapid denaturation and loss of enzyme activity when cells are disrupted for enzymatic analysis. The molecu-
’ The abbreviations used are: FPP, farnesyl pyrophosphate; SDS, sodium dodecyl sulfate; bp, base pair(s); kb, kilobase(s); PCRs, polym- erase chain reactions.
13229

13230 Identification of a Missense Mutation in Mevalonic Aciduria
lar defect causing this genetic disease is still unknown. As a first step in elucidating the molecular basis of this inherited disorder, we have isolated a cDNA clone coding for human mevalonate kinase, and we have used the DNA se- quence to identify a mutation in the mevalonate kinase gene from a subject with mevalonic aciduria.
EXPERIMENTAL PROCEDURES
Materials-A human skin fibroblast Xgtll cDNA library, a human B-lymphocyte Xgtll library, and Escherichia coli Y1090 were pur- chased from Clontech. A Xgtll cDNA library (MKlT) derived from mRNA purified from fibroblasts of the mevalonic aciduria index case (Z. W.) was prepared by Invitrogen. Other materials were purchased from the manufacturers as follows: A.S.A.P. genomic DNA isolation kit, Boehringer Mannheim; oligo(dT)-cellulose, human genomic DNA, and M13mp18 and M13mp19 sequencing vectors, Pharmacia LKB Biotechnology Inc.; chromosomal test panel blots and Speed- Hyb solution, Bios Corp.; Sequenase kit, U. S. Biochemical Corp.; GeneAmp DNA hybridization reagent kit, Perkin-Elmer Cetus In- struments; custom-synthesized gel-purified oligonucleotides, The Midland Certified Reagent Co.; pGEM vectors and T7 DNA polym- erase sequencing system kits, Promega Biotec; minimal essential medium, Opti-MEM, lipofectin reagent, and DH5aF' competent cells, Life Technologies Inc., fetal calf serum, HyClone Laboratories; [a-'"PIdCTP (3000 Ci/mmol), [ T - ~ ' P ] ~ A T P (7000 Ci/mmol), 35S- dATP (3000 Ci/mmol), and (R)-[5-'4C]mevalonic acid 5-phosphate (58 mCi/mmol), Amersham Corp.; and [3H]mevalonolactone (27.5 Ci/mmol), DuPont-New England Nuclear. The SV40-Ad2 construct was supplied by Yakov Gluzman (Cold Spring Harbor Laboratory), and the pCMV5 vector was a gift from David W. Russell (University of Texas Southwestern Medical Center, Dallas).
Cell Lines-Normal fibroblasts and fibroblasts from the mevalonic aciduria index case (Z. W.) and the proband's family (mother, father, and brother) were transformed with an SV40-Ad2 construct (13). Cells were grown (at 37 "C in humidified air containing 5% CO,) in minimal essential medium supplemented with 10% fetal calf serum, 100 units/ml penicillin, 100 pg/ml streptomycin, and 2 mM L-gluta- mine. Confluent monolayers of cells were rinsed twice with 5 ml of ice-cold phosphate-buffered saline, scraped into 2 ml of ice-cold phosphate-buffered saline, and collected by centrifugation at 1200 X g for 10 min at 4 "C. Cell pellets were frozen in dry ice/ethanol and stored a t -70 "C. COS-7 cells were grown under the same conditions in Dulbecco's modified Eagle's medium supplemented with 10% fetal calf serum, 100 units/ml penicillin, and 100 pg/ml streptomycin.
Isolation of cDNA Clones-Approximately 1 X lo6 recombinants from both a human skin fibroblast cDNA library and B-lymphocyte cDNA library were screened with a full-length rat mevalonate kinase cDNA clone (14) radiolabeled by random priming (15) to a specific activity of 5 X 10' cpmlpg. Plaque hybridization was performed as described (14). The nitrocellulose filters were washed twice a t 68 "C with 2 X SSC (1 X SSC = 0.15 M NaCI, 0.015 M sodium citrate (pH 7.0)) containing 0.1% SDS and washed twice with 0.1 X SSC contain- ing 0.1% SDS. Nine cDNA clones were isolated from the human skin fibroblast library, and six cDNA clones were isolated from the B- lymphocyte library. Approximately 1 X lo6 recombinants from the M K l T (mevalonic aciduria) cDNA library were screened by the same procedure, and 12 cDNA clones were isolated. All cDNA inserts were subcloned into the pGEM-3Z vector for restriction enzyme mapping or into M13mp18 and M13mp19 for DNA sequencing.
DNA Sequencing of cDNA Clones-The DNA sequence was deter- mined by the dideoxynucleotide chain termination method (16). Se- quencing reactions using Sequenase's modified T7 DNA polymerase were performed following the manufacturer's protocol. The DNA and protein sequences were aligned and compared to rat mevalonate kinase cDNA and protein sequences using the Intelligenetics and PCGene computer programs. The Swiss Protein and PIR data bases were also searched for other proteins that show amino acid sequence similarity to human mevalonate kinase.
Southern Blot Analysis-Normal human genomic DNA (10 pg) and genomic DNA isolated from fibroblasts from the mevalonic aciduria index case (Z. W.) were digested with restriction endonucleases. DNA fragments were separated by electrophoresis through a 1.0% agarose gel, transferred to a BioTrans nylon membrane, and covalently linked using a UV Stratalinker. Each membrane was hybridized overnight at 42 "C with a radiolabeled EcoRIIPstI cDNA fragment (specific
activity = 1 X 10' cpm/pg) that spans the entire human mevalonate kinase coding region. The hybridization solution contained 50% form- amide, 5 X SSPE (1 X SSPE = 0.15 M NaC1,0.25 M NaH2P04, 1 mM EDTA), 5 X Denhardt's solution (1 X Denhardt's = 0.2% Ficoll, 0.2% polyvinylpyrrolidone, 0.2% bovine serum albumin fraction V), 1% SDS, 1 mM NaPPi, and 100 pg/ml denatured salmon sperm DNA. The membranes were washed twice at 65 "C in 1 X SSC, 1% SDS and then twice a t 65 "C in 0.1 X SSC, 1% SDS. The BioTrans nylon membranes were dried, and the radiolabeled bands were visualized by autoradiography.
RNA Blot Analysis-Poly(A)' RNAs (15 pg) were isolated (17) from normal human fibroblasts and fibroblasts from the mevalonic aciduria index case, electrophoresed on a 1.2% agarose gel containing 1.1% formaldehyde, and transferred to BioTrans nylon membranes. The RNA was cross-linked to the membrane using a UV Stratalinker; prehybridized for 4 h at 42 "C in a solution containing 50% (v/v) formamide, 7% SDS, 0.25 M NaHP04 (pH 7.2), 0.25 M NaC1, 1 mM EDTA, and 100 pg/ml salmon sperm DNA; and then hybridized overnight at 42 "C with a radiolabeled 350-bp EcoRVIBglI DNA fragment (specific activity = 2 X lo8 cpm/pg) of the human mevalon- ate kinase cDNA clone. The membrane was washed twice a t 65 "C in 0.25 M NaHPO, (pH 7.2), 0.5% SDS, and 1 mM EDTA; followed by two washes in 0.05 M NaHP04 (pH 7.2), 0.5% SDS, and 1 mM EDTA a t 65 "C. The radiolabeled bands were visualized by autoradiography. The nitrocellulose membrane was stripped in boiling water and then rehybridized by the same method to an actin cDNA probe radiolabeled to a specific activity of 2 X 10' cpm/pg.
Chromosomal Localization-Chromosomal test panel blots were obtained containing 5 pg each of genomic DNAs from 25 different hamster-human somatic cell hybrid lines. The samples had been digested with EcoRI restriction endonuclease, and the DNA frag- ments were separated by electrophoresis through a 1.0% agarose gel before transfer to nitrocellulose membranes. Following the manufac- turer's protocol, the membranes were hybridized overnight a t 65 "C with a random-primed radiolabeled 430-bp PstIIEcoRI DNA frag- ment (specific activity - 1.1 X lo9 cpm/pg) from the full-length human mevalonate kinase cDNA clone, washed at 65 "C, and autoradi- ographed.
Polymerase Chain Reactions-Genomic DNAs from normal sub- jects and the mevalonic aciduria index case were prepared from cultured cell lines. A 208-bp region surrounding the putative mutation was amplified in these genomic DNA samples using oligonucleotide primers based on the sequence obtained from the human cDNA clones. A sense primer (5'-TGGGCATAGGACCTTGGCCT-3') and an antisense primer (5'-GAGGAGTGTGATGCCACAGCCAC-3') were synthesized and gel-purified. The polymerase chain reactions (PCRs) were performed on a Perkin-Elmer Cetus DNA thermal cycler using conditions described by the manufacturer (GeneAmp kit) with a 1 p~ concentration of each primer and 200 ng of genomic DNA template in a 100-pl reaction volume. Initially, the reaction was heated for 5 min at 95 "C for complete denaturation prior to cycling; 35 cycles were then carried out with a cycle profile of 2 min a t 95 "C, 1 min at 55 "C, and 2 min at 72 'C, with a final extension step at 72 "C for 5 min. For direct DNA sequencing of the PCR product, five identical reactions were pooled, and the reaction product was precip- itated with ethanol. The DNA was resuspended in 10 mM Tris (pH 7.4), 1 mM EDTA and electrophoresed on a preparative 2% NuSieve agarose gel. The DNA was recovered with NA45 paper following the manufacturer's protocol. The purified DNA was sequenced directly with Promega's T7 DNA polymerase sequencing system kit using an internal primer to the sense strand (5"CAGCTTGCTGT- GAAGTCCGC-3'). Approximately 500 ng of template DNA was added to 4 pmol of primer and heated to 98 "C for 8 min and then cooled a t room temperature for 1 min. Extensions were carried out on ice with the addition of 0.5 pl of 35S-dATP, 1.0 pl of 0.1 M dithiothreitol, 2.0 p1 of diluted (1: lO) labeling mixture, 2.0 pl of 5 X sequencing buffer, and 2.0 pl of a dilution (1%) of T7 DNA polymer- ase. 3.5 pl of this mixture was removed and added to a mixture containing 2.5 pl of each dideoxynucleotide triphosphate mixture. The reaction was incubated a t 37 "C for 3 min and was stopped with 4 pl of the formamide dye mixture. The reactions were heated a t 95 "C for 5 min prior to loading on a 6% sequencing gel. The radiolabeled bands were visualized by autoradiography.
Allele-specific Oligonucleotide Hybridization-Genomic DNA sam- ples were prepared from both normal and mevalonic aciduria cell lines. PCR amplification of the DNA region containing the putative mutation was performed as described above. Approximately 200 ng

Identification of a Missense Mutation in Mevalonic Aciduria 13231
of the DNA was dried down to a volume of 10 pl, and 1 pl of 5 N NaOH was added to denature each sample. The samples were then spotted onto BioTrans nylon membranes and dried. Duplicate mem- branes were prepared for hybridization with either the radiolabeled normal or mutant oligonucleotides. The membranes were hybridized for 4 h at 55 ‘C in 0.9 M NaCl, 0.2 M Tris (pH 8.01, 5 mM EDTA, 0.5% dried milk, and 1% SDS with a 1.3 X 10’ cpm/ml concentration of either the normal oligonucleotide (5”ATTGACATGAAC- CAGCACCAT-3’) or the mutant (mevalonic aciduria) oligonucleotide (5’-ATTGACATGACCCAGCACCAT-3’) radiolabeled at the 5’-ter- minus with 32P. The blots were washed in 1 X SSC, 1% SDS at room temperature to remove background. The wash temperature was then increased to 60 or 65 ‘C for the normal and mutant probes, respec- tively. The radiolabeled bands were visualized by autoradiography.
Transfection Analysis-Standard methods were used to construct the normal and mutant mevalonate kinase expression plasmids. Briefly, a 1.3-kb EcoRIIPstI fragment containing the coding sequence of the normal human mevalonate kinase cDNA was inserted into the eukaryotic expression vector pCMV5, a derivative of pCMV4 (18). The mutant expression construct was made by substituting into the normal expression construct a 821-bp EcoRVIPstI fragment from one of the mutant cDNAs obtained through the cDNA library screening. The sequence of the mutant insert was verified by automated sequenc- ing using fluorescence-labeled primers (19) and a DNA Sequencer (Model 373A, Applied Biosystems, Inc.). Liposome-mediated trans- fection of COS-7 cells was performed according to the manufacturer. Subconfluent cultures in 100-mm dishes were transfected with 26 pg of the vector alone, the normal mevalonate kinase construct, or the mutant construct for 16 h in 7 ml of serum-free media. Following 40 h of expression in minimal essential medium and 10% fetal calf serum, the cells were harvested in 250 pl of 100 mM KH,PO,, 10 mM dithiothreitol, 10 mM KF and lysed by sonication. Cellular debris was removed by centrifugation at 1000 X g for 5 min, and supernatant protein concentrations were determined (20). 20 pg of protein was used to measure mevalonate kinase activity by radiochemical assay (21). SDS-polyacrylamide gel electrophoresis and immunoblotting were as described (2) using 300 pg of protein and a polyclonal antibody generated against purified rat mevalonate kinase. The rat mevalonate kinase used as a standard was purified as described (2).
RESULTS
Isolation and DNA Sequence Analysis of cDNA Clone Coding for Human Mevalonate Kinase-Six human mevalonate ki- nase cDNA clones (pHMK1-pHMK6) were initially isolated after screening 1 X IO6 recombinants from a human B-lym- phocyte cDNA library with the radiolabeled rat mevalonate kinase cDNA. The sizes of the cDNA inserts in these clones ranged from 3.0 to 1.6 kb. All of the cDNAs were subcloned into sequencing vectors, and the DNA sequence was deter- mined. The resulting DNA sequences were compared to the known nucleotide sequence for rat mevalonate kinase (14); and although there was strong conservation of the DNA sequence, each of the isolated clones contained a deletion ranging in size from 150 to 300 bp within the 5‘-coding region.
To obtain a full-length cDNA clone, a 1.6-kb cDNA insert from one of these clones was used to screen a human skin fibroblast cDNA library. Nine clones were isolated after screening 1 X lo6 recombinants, and the sizes of the cDNA inserts in these clones ranged from 2.0 kb to 500 bp. The DNA sequences of two different 2.0-kb cDNA inserts (pHMK101 and pHMK104) were determined (Fig. 1). The cDNA contained a 1188-bp open reading frame and coded for a 396-amino acid polypeptide with a predicted M, of 42,450 assuming translational initiation at the first methionine co- don. The 689-bp 3”untranslated region contained a consensus polyadenylation signal (AATAAA), and a poly(A) tail was located 17 bp downstream from this site.
One of the 3.0-kb cDNA clones (pHMK4) isolated from the B-lymphocyte cDNA library was also subcloned into a se- quencing vector, and the nucleotide sequence was determined. There was no sequence conservation to the human mevalon- ate kinase cDNA sequence observed for the first 2101 nucle-
otides until a SacI restriction site was reached. The DNA sequence of the remaining 950 nucleotides from the SacI site to the poly(A) tail of the 3.0-kb clone was identical to the full-length human cDNA sequence from nucleotides 886 to 1972. The presence of a consensus splice site signal immedi- ately 5’ of the SacI site suggests that the first 2101 nucleotides are within the intron sequence derived from incomplete splic- ing of the mRNA (see Fig. 7). The DNA sequence of the intron region was used in later studies to produce primers for PCR amplification.
Analysis of Deduced Amino Acid Sequence of Human Mev- alonate Kinase-With a single residue gap inserted in the deduced amino acid sequence of rat mevalonate kinase, there was an 82% homology between the human and rat amino acid sequences (Fig. 1). The deduced molecular weight of human mevalonate kinase (42,450) is only slightly larger than the molecular weight of the rat enzyme (41,990). Based on the hydropathy profile, human mevalonate kinase, like rat mev- alonate kinase, is a very hydrophobic protein, with an overall index of hydropathy of +0.06 (22); and hydrophobic amino acids compose 45% of the total amino acids. The consensus sequence for a putative ATP-binding site previously identified in rat mevalonate kinase is also present in the human enzyme (14).
Protein Sequence Similarities-Computer-assisted searches of the Swiss Protein and PIR data bases identified three regions in the protein sequence of human mevalonate kinase that showed strong identity to the amino acid sequences reported for galactokinase (23-26), the yeast RARl protein (27), the yeast GAL3 protein (28), and the yeast phospho- mevalonate kinase protein (29). Region A spans 27 amino acids, region B spans 22 amino acids, and region C spans 12 amino acids in length (Fig. 2). Consensus sequences were identified in each region. When compared over the length of the protein, the relative locations of these motifs were also found to be spatially conserved (Fig. 3), suggestive of a close evolutionary linkage for these proteins.
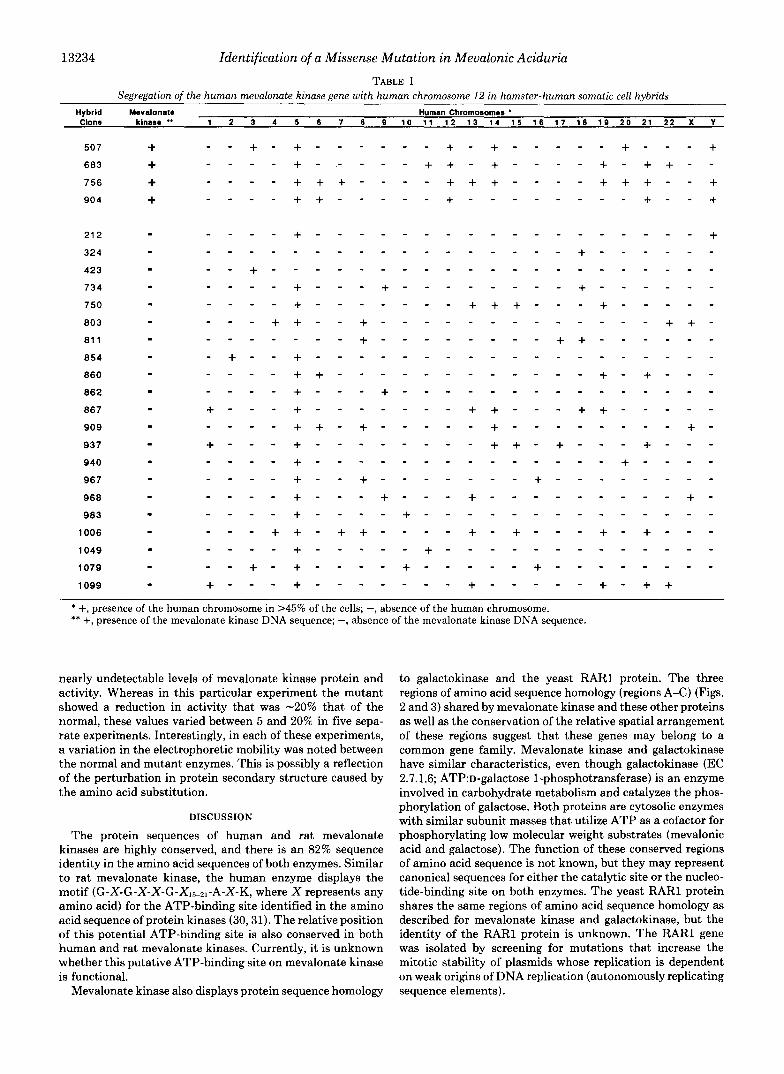
Chromosomal Localization of Mevalonate Kinase-The chromosomal location of the mevalonate kinase gene was determined by hybridizing a radiolabeled PstIIEcoRI DNA fragment of the human mevalonate kinase cDNA to a South- ern blot containing DNA purified from 25 different hamster- human somatic cell hybrids. The PstIIEcoRI DNA fragment that derived from the 3”untranslated region of the cDNA did not cross-hybridize to the hamster gene homolog. The radio- labeled cDNA hybridized to an -3-kb DNA fragment on Southern blots from four hamster-human hybrid cell lines. Comparison of the content of human chromosomes in these hybrid cell lines versus cell lines that did not hybridize to the radiolabeled probe indicated that the mevalonate kinase gene is located on human chromosome 12 (Table I). There were no discordant hybrid cell lines.
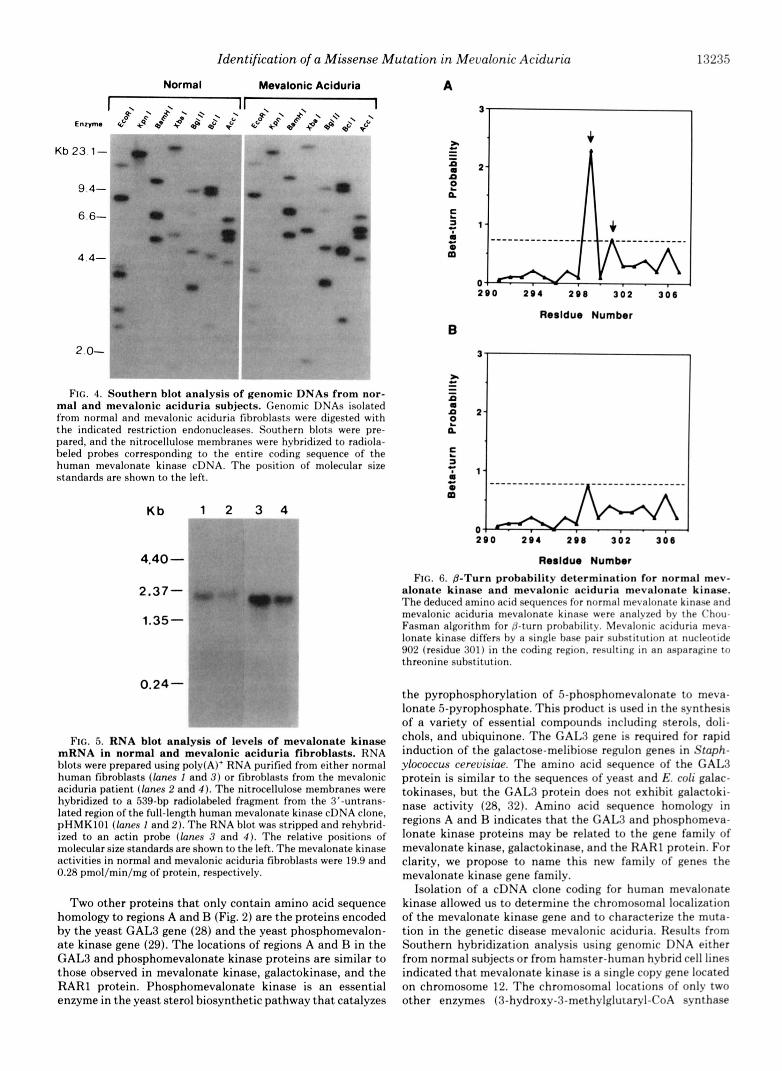
Southern Hybridization Analysis of Genomic DNAs from Normal and Mevalonic Aciduria Subjects-To determine if the mevalonic aciduria index case has a major rearrangement of the mevalonate kinase gene, genomic DNA was isolated from either normal fibroblasts or fibroblasts from the proband and digested with eight separate restriction endonucleases. Southern blot hybridization studies showed identical restric- tion fragments in both DNA samples, indicating that the reduced level of mevalonate kinase activity in the index case does not result from a major rearrangement of the mevalonate kinase gene (Fig. 4). Data using a smaller (122-bp) fragment of radiolabeled cDNA for Southern blot analysis also suggests that the genomic DNA coding for mevalonate kinase is a single copy gene (data not shown).

13232 Identification of a Missense Mutation in Mevalonic Aciduria
FIG. 1. Nucleotide and predicted amino acid sequences of human mevalonate kinase and comparison to rat mevalonate kinase. Nucleotides and amino acids are numbered to the left and right, respectively, with position 1 being assigned to the first nucleotide and residue of the coding sequence. The con- sensus polyadenylation signal (AA- TAAA) is underlined. The site of the missense mutation is indicated (6) a t nucleotide 902. The protein sequences are shown in single letter code. The hu- man and rat mevalonate kinase protein sequences have been aligned with the introduction of a single gap a t residue 394 to maximize identity. Asterisks de- note identical amino acids. The two se- quences shared an overall identity of 82%.
-91
1
91
181
271
361
4 5 1
54 1
6 3 1
121
811
901
991
1081
1111
1261
1351
1441
1531
1621
1711
1801
CAAAACAAAAGGTAGTGGGGAGCTGCTCCGGCTTCGGCGCGGAGGGGCGGCGGCCGGGGAGGCGGCGGCGGCGGCAGGATTCCCAGGAGCC
ATGTTGTCAGAAGTCCTACTGGTGTCTGCTCCGGGGAAAGTCATCCTTCATGGAGAACATGCCGTGGTACATGGCAAGGTAGCACTGGCT METLeuSerGluValLeuLeuValSerRlaProGlyLysValIleLeuHi~GlyGluHisAlaValValH~~GlyLy~V~lAl~LeuAl~ . . f * l ~ ~ * + ~ * . . + f . * . ~ * ~ . . * . * , * ~ *
GTATCCTTGAACTTGAGAACATTCCTCCGGCTTCAACCCCACAGCAATGGGAAAGTGGACCTCAGCTTACCCAACATTGGTATCAAGCGG V~lSerLeUA9nLe~A~qThrPheLeuArgLeuArqLeuGl~P~oHi~S~rAsnGlyLy~V~lA~pLe~SerLeuPr~A~~lleGlyIleLy~A~q
* Ala * * Val f Arg f G l n * * * * Ser * Asn * Val f f * Gln
GCCT~GGATGTGGCCAGGCTTCAGTCACTGGACACAAGCTTTCTGGAGC~GGTGATGTCACAACACCCACCTCAGAGCAAGTGGAGAAG AlaTrpAspValAlaArgLeuGlnSerLeuAspThrSerPheLeuGluGlnGlyAspValThrTh~P~oTh~SerGluGlnValGl~Lys Val * * f Thr Leu * Gly * f * * ProAla * Leu * * Leu *
CTAAAGGAGGTTGCAGGCTTGCCTGACGACTGTGCTGTCACCGAGCGCCTGGCTGTGCTGGCCTTTCTTTACTTATACCTGTCCATCTGC LeuLy~GluValAlaGlyLeuProA~pA~pCysAlaValTh~GluArqLeuAlaValLeuAlaPheLeuTy~LeuTy~LeuSerIleCys
* Lys * * Arg * ValGlyAsn Gly * SerLeU * * f * * Ala * *
CGGAAGCAGAGGGCCCTGCCGAGCCTGGATATCGTAGTGTGGTCGGAGCTGCCCCCCGGGGCGGGCTTGGGCTCCAGCGCCGCCTACTCG ArgLySGlnArgAlaLeuProSerLeuA~pIleValValTr~Se~GluLeuP~oProGlyAlaGlyLeuGlySe~Se~AlaAlaTy~Ser + ~ ~ ' ~ ~ ~ ' * ' " " e ~ , ~ ' * ' * . * . * * * ~ * ~ ~ * *
GTGTGTCTGGCAGCAGCCCTCCTGACTGTGTGCGAGGAGATCCCAAACCCGCTGAAGGACGGGGATTGCGTCAACAGGTGGACCAAGGAG ValCysLeuAlaAlaAlaLeuLeuThrValCysGluGluIleProAsnProLeuLysA~pGlyA~pCysValAsnA~qT~pThrLysGl~
Val * * * * * Ala ValThr * * ArgGlySerIleGlySer * ProGlu
GATTTGGAGCTAATTAACAAGTGGGCCTTCCAAGGGGAGAGAATGATTCACGGGAACCCCTCCGGAGTGGACAATGCTGTCAGCACCTGG AspLeuGlULeUIleAsnLysTrpAlaPheGlnClyGlyGl~ArgMetIleHi~GlyA~~P~~Se~GlyValA~pA~~Al~V~lSerTh~T~p
f t LysSer * * I t . TyrGIU * * Val . 1 f * f f . Ser f 1 . 1
GGAGGAGCCCTCCGATACCATCAAGGGAAGATTTCATCCTTAAAGAGGTCGCCAGCTCTCCAGATCCTGCTGACCAACACCAAAGTCCCT GlyGlyAlaLeuArqTyrHisGlnGlyLysIleSerSerLe~Ly~ArqSerProAlaLeuGlnIleLeuLeuThrAsnThrLysValP~o
II * t +Gin+ * "et' II f t * L e u ' t f * f 1/ II * * t II *
CGCAATACCAGGGCCCTTGTGGCTGGCGTCAGAAACA~CTGCTCAAGTTCCCAGAGATCGTGGCCCCCCTCCTGACCTCAATAGATGCC ArgAsnThrArqAlaLeuValAlaGlyValArgAsnA~qLeuLeuLysPheP~oGluIleValAl~P~oLeuLeuTh~SerIleAspAla ' S e r * L y s * I f f + ' S e r i * I l e + * f + M e t ' * * * f f
ATCTCCCTGGAGTGTGAGCGCGTGCTGGGAGAGATGGGGGAAGCCCCAGCCCCGGAGCAGTACCTCGTGCTGGAAGAGCTCATTGACATG IleSerLeuGlUCysGluArgValLeuGlyGluMetGlyGluAlaProAlaProGluGlnTyrLeuValLeuG~uGl~LeuIleAspMet . * f * * .AlaAla* 'Val* * * t f f * 'Met. *
AACCAGCACCATCTGAATGCCCTCGGCGTGGGCCACGCCTCTCTGGACCAGCTCTGCCAGGTGACCAGGGCCCGCGGACTTCACAGCAAG AsnGlnHisHisLeuAsnAlaLeuGlyValGlyHisAlaSerLeuA~pGl~LeuCy~GlnValThrArqAlaArqGlyLeuHisSe~Lys
* * . . " ' * " * * + * " * 1 * ' * , A l = , H ~ ~ * ~ * * *
CTGACTGGCGCAGGCGGTGGTGGCTGTGGCATCACACTCCTCAAGCCAGGGCTGGAGCAGCCAGAAGTGGA~CCACGAAGCAGGCCCTG LeuThrGlyAlaGlyGlyGlyGlyCysGlyIleThrLeuLeuLy~P~oGlyLeuGluGl~ProGl~V~lGluAlaThrLysGlnAlaLeu * * * * t I II I t I 1 L t f II f +ArgAlaLys* + A l a * .I t
ACCAGCTGTGGCTTTGACTGCTTGGAAACCAGCATCGGTGCCCCCGGCGTCTCCATCCACTCAGCCACCTCCCTGGACAGCCGAGTCCAG ThrSe~CysGlyPheASpCysLeuGluThrSerIleGlyAlaProGlyValSerIleHlsSerAlaTh~SerLeuAspSerAr~ValGln
* Gly * * * Trp * * * * * * * * Met * * IleGluAspPro Arq
CAAGCCCTGGATGGCCTCTGAGAGGAGCCCACGACACTGCAGCCCCACCCAGATGCCCCTTTCTGGATTATTCTGGGGGCTGCAGTTCGA GlnAlaLeuAspGlyLeu... * . * _ , *
CTCTGTGCTGGCCAGCGAGCGCCCAGCTCCTGACACTGCTGGAGAGGCCCCAGCCGCTTGGCGATGCCAGCCAAGCTCTGCAGTCCCAGC
GGTGGGACCTAGGGAGGCATGGTCTGCCCTCTGCATCCTCTGGAGCCAGCCGAGCAGGAGGCCTAGGAGGGTCCTCTGAGACTCCAGACC
TGAGGCGAGAAGGGCTGCTTCCCTGAAGCTCCCACAGTCCCATCTGCTTCAGGCCCCCGCCTTGGCCTGTGTTCTTCCTGGCCGCCTGGG
TCCAATGCTCAGGTGCTGGGGCCTGGTTCCCGGAGAAGTGTGCCTTCTCTCTCCCTTTTCAGGGACGGCCCCCTGTCTCTCAGGGCCAGG
CCTCTCCCTCCTCCAGGAAGCCTTCCCCTACCCCTTGTCGCCCCTCCCTCCCAGAGCACCTGCTGTCTGGGTGGC~CACTCAGCACTTGG
CCCTTCTACCTAGCGGGATGGGGCTCCCCCAGGGGCTGTCCCGGAGGCGGTG~CCTGGTTAAATAAGGCAGTGTGGCCTTGGTTTATA
GCACTTTCTTCCGATCTGTACCTGAGAGGTTTGTGGAAAAGATGGCAAATGGGG~AGATTTTGTGTcAAC(A)n
Meualonate Kinase mRNA-To determine the size and relative levels of mevalonate kinase mRNA, a Northern blot was prepared using poly(A)+ RNA isolated from either normal fibroblasts or fibroblasts from the mevalonic aciduria index case. The size of human mevalonate kinase mRNA from normal and mevalonic aciduria samples (Fig. 5, lanes I and 2, respectively) is -2.0 kb, which is identical to the size reported for rat mevalonate kinase mRNA (14). Normalization to the levels of actin mRNA (lanes 3 and 4) showed that the relative abundance of mevalonate kinase mRNA was similar in both normal and mevalonic aciduria cells. In contrast, mevalonate kinase activities measured by a radiochemical assay (21) were 19.9 and 0.28 pmol/min/mg of protein ( n = 2) in the normal and mevalonic aciduria cells, respectively.
Isolation and D N A Sequence Analysis of Meualonate Kinase cDNA Clone from Subject with Mevalonic Aciduria-A cDNA library was prepared from mRNA purified from fibroblasts from the mevalonic aciduria index case. Twelve cDNA clones (pMKlT1-pMKlT12) were obtained after screening 1 x lo6 recombinants with a radiolabeled normal human mevalonate kinase cDNA insert (pHMK101). The cDNA inserts in these clones ranged in size from 880 bp to 2.0 kb. The DNA sequences were aligned with the normal human mevalonate kinase cDNA sequence. Two of the cDNA inserts contained
30
60
90
120
150
180
210
2 4 0
270
30 0
330
360
390
396
full-length sequence corresponding to the normal mevalonate kinase. However, 4 of the 12 cDNA inserts contained a single base pair substitution in the coding region (nucleotide 902) that replaced an adenine with a cytosine (the remaining clones contained partial sequences only). The effect of this transver- sion was to change an asparagine residue to a threonine residue, which resulted in the elimination of a predicted p- turn in the secondary structure of the protein (Fig. 6).
Sequence Analysis of Normal Human Genomic DNA and Fibroblasts from Mevalonic Aciduria Patient-To confirm the A to C nucleotide substitution and to ensure that its presence in the cDNA clones was not the result of an artifact produced during construction of the MKlT cDNA library, normal human genomic DNA as well as genomic DNA isolated from fibroblasts from the mevalonic aciduria index case were se- quenced. The 208-bp region surrounding the putative muta- tion was amplified from genomic DNA by PCR, and the nucleotide sequence was determined (Fig. 7). The sequence obtained from the mevalonic aciduria genomic DNA confirms the presence of the missense mutation at nucleotide 902 and also demonstrates that the patient is a heterozygote for the allele containing the A to C mutation.
Identification of Missense Mutation by Allele-specific Oligo- nucleotide Hybridization-To determine whether the A to C

Identification of a Missense Mutation in Mevalonic Aciduria
REGION A_
RAT MEV KINASE : ( 1 ) HUMAN MEV KINASE : ( 1 ) EgjzK E . c o l i GAL KINASE : ( 1 6 ) GYPATHTI S. c e r e v i s i a e GAL KINASE (GAL 1) : ( 4 1 ) DAKPDFVA Streptomyces S p . GAL KINASE : ( 1 8 G R ~ R G C G R K. marxianus GAL KINASE : ( 3 1 ) DSRKFFITR S. c e r e v i s i a e RARl : ( 1 ) "HLPFLTS S. c e r e v i s i a e GAL3 S. c e r e v i s i a e P-MEV KINASE : ( 1 ) -@LRAFS
: ( 3 5 ) HVNLILSLG DTKYEAFVVGL
CONSENSUS - -0- - - - - - APGRVNLIGEH-DY----VL-----
D F S ~ L A I D
- RAT MEV KINASE
E . c o l i GAL KINASE S. c e r e v i s i a e GAL KINASE (GAL
X. marxianus GAL KINASE Streptomyces S p . GAL KINASE
S. c e r e v i s i a e RARl S. c e r e v i s i a e GAL3 S. c e r e v i s i a e P - m v KINASE
n u w MEV KINASE
1)
CONSENSUS ""_ S--P---GLGSSAA--V----A-- S
BEGION C
RAT MEV KINASE HUMAN MEV KINASE
: (323) AAHGLHSK : (323) RARGLHSK
E . c o l i GAL KINASE : ( 3 2 3 ) IGDKGGVR S. c e r e v i s i a e GAL KINASE (GAL 1) : (456) SNGSYGSR Streptomyces sp. GAL KINASE : ( 3 2 3 ) GLRGPRRR S. c e r e v i s i a e RARl : ( 3 4 1 ) DLRIGSTK
CONSENSUS ---------TG-G-GGC---L--------
13233
: ( 3 4 ) : ( 3 4 ) : ( 4 9 ) : (74) : ( 5 1 ) : ( 6 4 ) : ( 3 3 )
: ( 3 3 ) : (68)
: (158) : ( 1 5 8 )
: (142) : (183)
: (170) : (142)
: ( 1 6 1 ) : ( 1 7 6 ) : (170)
: ( 3 5 1 ) : (351) : (351) : ( 4 8 4 ) : (351) : ( 3 6 9 )
FIG. 2. Protein sequence conservation found in the three regions of human mevalonate kinase. Shown are the three regions of protein sequence homology, regions A-C. Amino acids are shown in single letter code, and the numbers in parentheses represent the positions in the amino acid sequence for each protein. Amino acids present a t a given position in 50% or more of the sequences are boxed and shaded, and the consensus amino acid sequence is shown below each region. MEV, mevalonate; GAL, galactokinase; P-MEV, phosphomevalonate. The amino acid sequence shown for Kluyueromyces marxianus represents only a fragment of the galactokinase.
XESlDCE hfl
H M K
R M K
EGK
YGK
SGK
R A R l
YGAL3
YPMK
FIG. 3. Spatial arrangement of three conserved regions of protein sequence. Regions A-C of conserved protein sequence are represented by solid, hatched, and checkered boxes, respectively. The approximate residue number for each protein is shown at the top. HMK, human mevalonate kinase; RMK, rat mevalonate kinase; EGK, E. coli galactokinase; YGK, yeast galactokinase; SGK, Streptomyces galactokinase; RARI, yeast RARl protein; YGAW, yeast GALS; YPMK, yeast phosphomevalonate kinase.
nucleotide substitution was present as a common polymor- phism in normal human genomic DNA and whether it was present in other mevalonic aciduria subjects, an allele-specific oligonucleotide hybridization screen was employed. The anal- ogous 208-bp region surrounding the mutation was amplified from genomic DNA by the PCR technique, and the product
was spotted onto duplicate BioTrans nylon membranes. The membranes were then hybridized with allele-specific end- labeled oligonucleotide probes to discriminate between the normal sequence or the sequence containing the A to C substitution. As shown in Fig. 8, seven different samples of normal genomic DNA samples were negative for the mutation in addition to genomic DNA samples from four different subjects with mevalonic aciduria. However, hybridization of the oligonucleotide with the mutation to genomic DNAs from the proband (Z. W.) and the proband's father and brother confirmed the presence of the point mutation in these sub- jects, whereas the proband's mother was negative. Hybridi- zation of the oligonucleotide with the normal sequence to genomic DNA from the proband indicated that the index case is a compound heterozygote.
Transient Expression of Normal and Mutant Mevalonate Kinases-Although the mutation identified in the patient with mevalonic aciduria displayed mendelian inheritance, ver- ification of this defect as the causative agent required expres- sional analysis. To this end, mutant and normal cDNAs were inserted into the expression vector pCMV5 and expressed transiently in COS-7 cells (18). Fig. 9 shows the typical enzyme activity and protein levels (inset) expressed from the two constructs. Using a polyclonal antibody generated against purified rat mevalonate kinase, immunoblot analysis was performed on equivalent amounts of protein from cell lysates of each transfection. Human mevalonate kinase enzyme ac- tivity was determined by a radiochemical assay of the same cell lysates. It is clear that whereas each construct expressed an equal amount of mevalonate kinase protein, the activity associated with the mevalonic aciduria defect was greatly diminished. Control and mock-transfected cells displayed
13234 Identification of a Missense Mutation in Mevalonic Aciduria
TABLE I Segregation of the human mevalonate kinasegene with human chromosome 12 in hamster-human somatic cell hybrids
Hybrid Mevalonate Clone kinase ** 1 2 3 4 5 6 7 8 0 1 0 1 1 1 2 1 3 1 4 1 5 1 6 1 7 1 8 1 9 2 0 2 1 2 2 X Y
Human Chromosomes *
507 + " + - + " " " + - + " " - + - - - + 683 + " " + " " _ + + - + " " + - + + - - 756 + " " + + + " " + + + " " + + + " + 904 + " " + + " " _ + - - - - - - - - + - - +
21 2
324
423
734
750
803
81 1
854
860
862
867
909
937
940
967
968
983
1006
1049
1079
1099
* +, presence of the human chromosome in >45% of the cells; -, absence of the human chromosome. ** +, presence of the mevalonate kinase DNA sequence; -, absence of the mevalonate kinase DNA sequence.
nearly undetectable levels of mevalonate kinase protein and activity. Whereas in this particular experiment the mutant showed a reduction in activity that was -20% that of the normal, these values varied between 5 and 20% in five sepa- rate experiments. Interestingly, in each of these experiments, a variation in the electrophoretic mobility was noted between the normal and mutant enzymes. This is possibly a reflection of the perturbation in protein secondary structure caused by the amino acid substitution.
DISCUSSION
The protein sequences of human and rat mevalonate kinases are highly conserved, and there is an 82% sequence identity in the amino acid sequences of both enzymes. Similar to rat mevalonate kinase, the human enzyme displays the motif (G-X-G-X-X-G-X,,-,,-A-X-K, where X represents any amino acid) for the ATP-binding site identified in the amino acid sequence of protein kinases (30,31). The relative position of this potential ATP-binding site is also conserved in both human and rat mevalonate kinases. Currently, it is unknown whether this putative ATP-binding site on mevalonate kinase is functional.
Mevalonate kinase also displays protein sequence homology
to galactokinase and the yeast RARl protein. The three regions of amino acid sequence homology (regions A-C) (Figs. 2 and 3) shared by mevalonate kinase and these other proteins as well as the conservation of the relative spatial arrangement of these regions suggest that these genes may belong to a common gene family. Mevalonate kinase and galactokinase have similar characteristics, even though galactokinase (EC 2.7.1.6; ATP:D-galactose 1-phosphotransferase) is an enzyme involved in carbohydrate metabolism and catalyzes the phos- phorylation of galactose. Both proteins are cytosolic enzymes with similar subunit masses that utilize ATP as a cofactor for phosphorylating low molecular weight substrates (mevalonic acid and galactose). The function of these conserved regions of amino acid sequence is not known, but they may represent canonical sequences for either the catalytic site or the nucleo- tide-binding site on both enzymes. The yeast RARl protein shares the same regions of amino acid sequence homology as described for mevalonate kinase and galactokinase, but the identity of the RARl protein is unknown. The RARl gene was isolated by screening for mutations that increase the mitotic stability of plasmids whose replication is dependent on weak origins of DNA replication (autonomously replicating sequence elements).
Identification of a Missense Mutation in Mevalonic Aciduria
Normal Mevalonic Aciduria t
"" ,"n_ -7, - Kb 23.1 -
9.4-
6.6-
4.4-
2.0-
FIG. 4. Southern blot analysis of genomic DNAs from nor- mal and mevalonic aciduria subjects. Genomic DNAs isolated from normal and mevalonic aciduria fihroblasts were digested with the indicated restriction endonucleases. Southern blots were pre- pared, and the nitrocellulose memhranes were hybridized to radiola- beled probes corresponding to the entire coding sequence of the human mevalonate kinase cDNA. The position of molecular size standards are shown to the left.
Kb 1 2 3 4
4.40 - 2.37-
1.35-
0.24-
FIG. 5. RNA blot analysis of levels of mevalonate kinase mRNA in normal and mevalonic aciduria fibroblasts. RNA blots were prepared using poly(A)' RNA purified from either normal human fibroblasts (lanes I and 3 ) or fibrohlasts from the mevalonic aciduria patient ([anes 2 and 4 ) . The nitrocellulose membranes were hybridized to a 539-bp radiolabeled fragment from the 3"untrans- lated region of the full-length human mevalonate kinase cDNA clone, pHMKlOl (lanes 1 and 2). The RNA blot was stripped and rehyhrid- ized to an actin probe (lanes 3 and 4 ) . The relative positions of molecular size standards are shown to the left. The mevalonate kinase activities in normal and mevalonic aciduria fibroblasts were 19.9 and 0.28 pmol/min/mg of protein, respectively.
Two other proteins that only contain amino acid sequence homology to regions A and B (Fig. 2) are the proteins encoded by the yeast GAL3 gene (28) and the yeast phosphomevalon- ate kinase gene (29). The locations of regions A and B in the GAL3 and phosphomevalonate kinase proteins are similar to those observed in mevalonate kinase, galactokinase, and the RARl protein. Phosphomevalonate kinase is an essential enzyme in the yeast sterol biosynthetic pathway that catalyzes
I
2 9 0 2 9 4 2 9 0 3 0 2 306
Rerldue Number
2-
1- w """"""""_ """"""""_
I
1323.5
"
2 9 0 2 9 4 2 9 8 3 0 2 306
Rerldue Number FIG. 6. &Turn probability determination for normal mev-
donate kinase and mevalonic aciduria mevalonate k i n a . The deduced amino acid sequences for normal mevalonate kinase and mevalonic aciduria mevalonate kinase were analyzed hv the Chou- Fasman algorithm for &turn prohahility. Mevalonic aciduria rneva- lonate kinase differs by a single bane pair suhstitution at nucleotide 902 (residue 301) in the coding region. resulting in an asparagine to threonine substitution.
the pyrophosphorylation of 5-phosphomevalonate to meva- lonate 5-pyrophosphate. This product is used in the synthesis of a variety of essential compounds including sterols, doli- chols, and ubiquinone. The GAL3 gene is required for rapid induction of the galactose-melibiose regulon genes in Staph- ylococcus cereukiae. The amino acid sequence of the GAL3 protein is similar to the sequences of yeast and E. coli galac- tokinases, but the GAL3 protein does not exhibit galactoki- nase activity (28, 32). Amino acid sequence homology in regions A and B indicates that the GAL3 and phosphomeva- lonate kinase proteins may be related to the gene family of mevalonate kinase, galactokinase, and the RARl protein. For clarity, we propose to name this new family of genes the mevalonate kinase gene family.
Isolation of a cDNA clone coding for human mevalonate kinase allowed us to determine the chromosomal localization of the mevalonate kinase gene and to characterize the muta- tion in the genetic disease mevalonic aciduria. Results from Southern hybridization analysis using genomic DNA either from normal subjects or from hamster-human hybrid cell lines indicated that mevalonate kinase is a single copy gene located on chromosome 12. The chromosomal locations of only two other enzymes (3-hydroxy-3-methylglutaryl-CoA synthase

13236 Identification of a Missense Mutation in Mevalonic Aciduria
A (; C ‘I’
b A A/C C C A G
N O R M A I ,
h
_tacracataaaacctta~cctcaggcaggccaagtggga~cagatggaaccttctccccta~~~TTGACAT 8 8 6 *
GAACCAGCACCATCTGAATGCCCTCGGCGTGGGCCACGCCTCTCTGGACCAGCTCTGCCAGGTGACCAGGGCCCG 906 326 946 966
CGGACTTCACAGCAAGCTGACTGGCGCAGGCGGTGGTGGC~TGGC~TCCTC 986 1006 1026
FIG. 7. Nucleotide sequence of genomic DNA from patient with mevalonic aciduria. Genomic DNA from either normal human fibroblasts or fibroblasts from a mevalonic ( M E V . ) aciduria patient (Z. W.) was amplified by PCR, and the nucleotide sequence was determined. Upper, a portion of the autoradiograph is shown for the sequences from hp 899 to 910, reading in a 5’ to 3‘ direction from the t.op of the gel, of the normal DNA and of the patient’s DNA. The heterozygous state of the patient’s DNA is shown at the arrowhead. with one normal allele and one allele containing the A to C nucleotide substitution. Lower, the site of the mutation in reference to the gene structure is shown. The intron-exon junction immediately 5’ of the SacI site was determined by comparison to the cDNA sequence and the identification of a consensus splice site. The intron sequence is shown in lower-case letters, and the exon sequence is shown in uppr-case letters. Numbering is based on the cDNA sequence. The SacI site is boxed. The 5’- and 3’-PCR primers used to amplify this region are underlined, and the position of the missense mutation is indicated by an asterisk.
1 7 3 4 5 6 7 8
NORMAL ALLELE s 0 b I 0 0
MEV. ACIDURIA ALLELE
9 10 I 1 1;’ 13 1 4 15 16
NORMAL ALLELE b * . @ e * ‘ m
MEV. ACIDUAIA ALLELE
FIG. 8. Identification of mevalonic aciduria mutation by allele-specific hybridization. Genomic DNAs were isolated from control lymphoblasts and fibroblasts as well as fibroblasts from a number of mevalonic aciduria patients, including the patient (Z. W.) and the patient’s mother, father, and brother. The region surrounding the A to C nucleotide suhstitution was amplified by PCR, and the product was spotted onto duplicate filters. The filters were hybridized either to an oligonucleotide corresponding to the normal sequence in the region of the A to C change (normal allele) or to an oligonucleotide from the same region incorporating the A to C change (mevalonic ( M E V . ) aciduria allele). Lane 1, human mevalonate kinase clone DNA; lanes 2 and 3, purchased normal human genomic DNA; lanes 4-8, normal genomic DNA from control fibrohlasts/lymphoblasts; lane 9, patient Z. W.; lane IO, Z. W.’s mother; lane 11, Z. W.’s father; lane 12. Z. W.’s brother; lanes 13-16, four individual mevalonic aciduria patients.
and reductase) in the cholesterol biosynthetic pathway are known, and both genes are located on human chromosome 5 (33-36). In situ hybridization studies are needed to determine
the regional mapping of human mevalonate kinase on chro- mosome 12.
Mevalonic aciduria is postulated to be caused by a mutation in the gene coding for mevalonate kinase. This hypothesis was based on the observation that there is a marked reduction in mevalonate kinase activity in cells from mevalonic aciduria subjects, and there is a large increase in the levels of meva- lonate in the serum and urine of these subjects. These data are consistent with having a mutation in the mevalonate kinase gene, but they are not conclusive. By isolating a cDNA clone coding for human mevalonate kinase, we were able to identify a missense mutation in the mevalonate kinase gene from a subject with mevalonic aciduria. The missense muta- tion changed the protein sequence of mevalonate kinase by replacing an asparagine residue with a threonine. This sub- stitution caused a significant alteration in the predicted sec- ondary structure of the enzyme. Analysis of the protein se- quence of mevalonate kinase using the Chou-Fasman algo- rithm indicated that this amino acid substitution eliminated a @-turn in the secondary structure of the enzyme. The marked effect of this amino acid substitution on protein structure may explain the loss of mevalonate kinase activity associated with this genetic disease. The transfection studies support this conclusion. COS-7 cells transfected with the mutant cDNA construct exhibited a severe reduction in the level of mevalonate kinase activity when compared to the normal expression construct, even though equivalent amounts of pro- tein were produced from the two vectors. Thus, the missense mutation resulted in a substantial inactivation of enzyme activity.
This mutation was identified in the DNA sequences of both a cDNA clone and a PCR-amplified region of genomic DNAs

Identification of a Missense Mutation in Mevalonic Aciduria 1.7237
T 1
\ I C \ ,\c l c lu r l . 1 \ o r n u 1 \lh \',x 11jr L f i t r ~ n ~ r ~ ~ ~ 1 $ , ( 1
FIG. 9. Transient expression of mutant and normal meValonate kinases in COS-7 cells. Expression constructs containing either normal (normal M K ) or mutant ( M e u Aciduria, MA) mevalonate kinase were transfected into COS-; cells and expressed for 4 0 h. Cell lysates were analyzed for enzyme activity by a radiochemical assay (histogram) and for mevalonate kinase expressional levels hy immunohlotting (inset). Equivalent amounts of protein were used from each sample in both the enzyme assay (21) p g ) and in each lane of the SI)S gel ( : ( 0 1 )
pa). Molecular mass standards ( S T D ) are indicated to the left of the autoradiograph.
from the proband and the proband's father and brother. The missense mutation does not represent a common gene poly- morphism because it was not found in genomic DNA samples from seven normal subjects and from four other subjects with mevalonic aciduria. Allele-specific oligonucleotide hybridiza- tion studies indicated that the proband inherited this muta- tion from the father and that the index case is a compound heterozygote. The absence of the missense mutation in the four other mevalonic aciduria subjects suggests that this ge- netic disease may be caused by a variety of mutations in the mevalonate kinase gene.
A genetic defect in the proband's mother has not yet been identified. Two full-length cDNA clones isolated from the mevalonic aciduria cDNA library contained the normal coding sequence for mevalonate kinase; and based on results from allele-specific oligonucleotide hybridization, this normal DNA sequence was inherited from the proband's mother. The pres- ence of a normal coding sequence for mevalonate kinase seems incompatible with the marked reduction in mevalonate kinase activity measured in the proband and, to a lesser degree, in the proband's mother. However, both of the cDNA clones with the normal coding sequence for mevalonate kinase did not contain the complete 5"untranslated region. Therefore, i t is possible that a mutation in the 5"untranslated region of the mevalonate kinase mRNA may inhibit translation. It is unlikely that this allele contains a mutation that alters gene transcription since cells from the proband contained normal levels of mevalonate kinase mRNA (Fig. 5 ) . To date, the molecular basis for the mevalonate kinase deficiency in the proband's mother is not known, and additional studies are needed to determine the complete nucleotide sequence of the 5"untranslated region for the mevalonate kinase mRNA.
Our study provides the first evidence that a mutation in the gene coding for mevalonate kinase can cause the genetic
disease mevalonic aciduria, and this is the first identified mutation in the cholesterol biosynthetic pathway in humans. Moreover, the fact that the missense mutation identified in our study was not detected in the prohand's mother as well as four other subjects with mevalonic aciduria indicates that this genetic disease may he caused hy different types of mutations in the gene coding for mevalonate kinase.
Acknowledgments-We thank Drs. Gordon Rohinson. Todd Kirrh- gessner, and Richard Cregg for useful discussions. Excellent technical assistance was provided hv Bernadette Kienzle.
REFERENCES 1. Brown. M. S.. and Goldstein. -1. I,. (19ROI J . 1.i id Rrs 21, 505-517 2. Tanaka. R. D.. Schafer. H. I... Lee. I,. Y.. Freucfhwrrer. .I. 5. . nnd 5!oslev.

13238 Identification of a Missense Mutation in Mevalonic Aciduria 19. Smith, L. M., Sanders, J. Z., Kaiser, R. J., Hughes, P., Dodd, C., Connell, 28. Bhat, P. J., Oh, D., and Hopper, J. E. (1990) Genetics 125,281-291
674-679 C. R., Heiner, C., Kent, S. B. H., and Hood, L. E. (1986) Nature 321 , 29. Tsay, Y. H., and Robinson, G. W. (1991) Mol. Cell. Biol. 11,620-631
20. Bradford, M. M. (1976) Anal. Biochem. 72 , 248-254 31. Argos, P., and Leberman, R. (1985) Eur. J. Biochem. 152,651-656
22. Kyte, J., and Doolittle, R. F. (1982) J. Mol. Biol. 157 , 105-132 21. Faust, J., and Krieger, M. (1987) J. Biol. Chem. 2 6 2 , 1996-2004 32. Bajwa, W., Torchia, T. E., and Hopper, J. E. (1988) Mol. Cell. Biol. 8 ,
23. Debouck, C., Riccio, A,, Schumperli, D., McKenny, K., Jeffers, J., Hughes, 33. Lindgren, V., Luskey, K. L., Russel, D. W., and Francke, U. (1985) Proc. C., Rosenberg, M., Heusterspreute, M., Brunel, F., and Davison, J. (1985) Natl. Acad. Sci. (I. S . A. 82,8567-8571
24. Citron, B. A,, and Donelson, J. E. (1984) J. Bacteriol. 158 , 269-278 Nuclerc Acrds Res. 13, 1841-1853 34. Mohandas, R., Heinzmann, C., Sparkes, R. S., Wasrnuth, J., Edwards, P.
25. Webster, T. D., and Dickson, R. C. (1988) Nucleic Acids Res. 16 , 8192- 35. Mehrabian, M., Callaway, K. A., Clarke, C. F., Tanaka, R. D., Greenspan, 8194 M., Lusis, A. J., Sparkes, R. S., Mohandas, T., Edmond, J., Fogelrnan,
26. Adarns, C. W., Fornwald, J. A,, Schmidt, F. J., Rosenberg, M., and Brawner, A. M., and Edwards, P. A. (1986) J. Biol. Chem. 261 , 16249-16255 M. E. (1988) J. Bacteriol. 170,203-212 36. Leonard, S., Arbogast, D., Geyer, D., Jones, C., and Sinensky, M. (1986)
27. Kearsey, S. E., and Edwards, J. (1987) Mol. Gen. Genet. 210, 509-517 Proc. Natl. Acad. Sci. U. S. A. 83, 2187-2189
30. Hanks, S. K., Quinn, A. M., and Hunter, T. (1988) Science 241,42-52
3439-3447
A,, and Lusis, A. J. (1986) Somatic Cell Mol. Genet. 12 , 89-94





























