Embed Size (px)
Citation preview

415
Mol. Biol. Evol. 15(4):415–426. 1998q 1998 by the Society for Molecular Biology and Evolution. ISSN: 0737-4038
Molecular Characterization and Phylogenetic Relationships of a Proteinwith Potential Oxygen-Binding Capabilities in the Grasshopper Embryo. AHemocyanin in Insects?
Diego Sanchez,* Marıa D. Ganfornina,* Gabriel Gutierrez,† and Michael J. Bastiani**Biology Department, University of Utah; and †Departamento de Genetica, Universidad de Sevilla, Sevilla, Spain
Arthropodan hemocyanins, prophenoloxidases (PPOs), and insect hexamerins form a superfamily of hemolymphproteins that we propose to call the AHPH superfamily. The evolutionary and functional relationships of theseproteins are illuminated by a new embryonic hemolymph protein (EHP) that is expressed during early stages ofdevelopment in the grasshopper embryo. EHP is a 78-kDa soluble protein present initially in the yolk sac content,and later in the embryonic hemolymph. Protein purification and peptide sequencing were used to identify an em-bryonic cDNA clone coding for EHP. In situ hybridization identifies hemocytes as EHP-expressing cells. As deducedfrom the cDNA clone, EHP is a secreted protein with two potential glycosylation sites. Sequence analysis definesEHP as a member of the AHPH superfamily. Phylogenetic analyses with all the currently available AHPH proteins,including EHP, were performed to ascertain the evolutionary history of this protein superfamily. We used both theentire protein sequence and each of the three domains present in the AHPH proteins. The phylogenies inferred foreach of the domains suggest a mosaic evolution of these protein modules. Phylogenetic and multivariate analysesconsistently group EHP with crustacean hemocyanins and, less closely, with insect hexamerins, relative to cheli-ceratan hemocyanins and PPOs. The grasshopper protein rigorously preserves the residues involved in oxygenbinding, oligomerization, and allosteric regulation of the oxygen transport proteins. Although insects were thoughtnot to have hemocyanins, we propose that EHP functions as an oxygen transport or storage protein during embryonicdevelopment.
Introduction
Some arthropods rely on proteins called hemocya-nins to transport the molecular oxygen needed for res-piration. In hemocyanins, oxygen binds to a pair of cop-per atoms located between four antiparallel a-helices.Each copper atom is coordinated to three histidine res-idues and is surrounded by a strong hydrophobic envi-ronment. Arthropodan hemocyanins are formed by sub-units of ;75 kDa arranged in hexamers or multihex-amers (for a review, see Van Holde and Miller 1995),with each subunit containing a dinuclear copper site ableto bind one oxygen molecule.
Arthropodan hemocyanins share significant globalsequence similarity with two other protein families: hex-amerins and prophenoloxidases (PPOs). All these pro-teins are presumably related by common ancestry (Bein-tema et al. 1994; Burmester and Scheller 1996), andthey might be considered to form a superfamily that wepropose to name the AHPH superfamily, an acronymfrom arthropodan hemocyanins, PPOs, and hexamerins.Hexamerins are proteins used for amino acid storage inlarval stages of insects (Telfer and Kunkel 1991). Somehexamerins can be grouped into separate classes accord-ing to their content of methionine or aromatic residuesand are therefore called methionine-rich proteins and ar-
Abbreviations: DIC, differential interference optics; EHP, embry-onic hemolymph protein of the grasshopper; LHP, larval hemolymphprotein of the locust; ORF, open reading frame; PPO, prophenoloxi-dase; SDS-PAGE, sodium dodecylsulphate polyacrylamide gel electro-phoresis; UTR, untranslated region.
Key words: grasshopper, hemocyanin, hexamerin, prophenoloxi-dase.
Address for correspondence and reprints: Diego Sanchez, BiologyDepartment, 229 South Biology, University of Utah, Salt Lake City,Utah 84112. E-mail: [email protected].
ylphorins, respectively. They are hexamers with sub-units in the 80-kDa range synthesized in the fat bodyand secreted to the hemolymph, where they reach highconcentrations just before metamorphosis (Telfer andKunkel 1991). Hexamerins do not bind oxygen, sincethe copper-binding histidine residues have been re-placed. They are very common and abundant in insects(Scheller, Fischer, and Schenkel 1990), and hexamerin-like proteins might exist in crustaceans (Markl et al.1979a). However, no copper-free hemocyanin-relatedprotein has been found in chelicerates (Markl et al.1979b). Phenoloxidase activity is ubiquitously found ininvertebrates, and PPOs from a crustacean and severalinsects have recently been molecularly characterized(Aspan et al. 1995; Fujimoto et al. 1995; Hall et al.1995; Kawabata et al. 1995). As copper-containing pro-teins, PPOs preserve the dinuclear copper sites of he-mocyanins. PPOs are synthesized in the cytoplasms ofhemocytes without a signal peptide, released to the he-molymph upon cellular rupture, and activated by pro-teolysis. In arthropods, the PPO active form (PO) is in-volved in defense reactions, wound healing, and cutic-ular sclerotization (Johansson and Soderhall 1996; Sug-umaran et al. 1992).
Hemocyanins have been found in three arthropodgroups: crustaceans, chelicerates, and a single order ofmyriapods. However, hemocyanin has not been reportedin insects. This finding has been explained by the ex-istence of a tracheal respiratory system in this subphy-lum. In this paper, we describe the finding of a solubleprotein present in the hemolymph of the grasshopper(Schistocerca americana) embryo that we call EHP (forembryonic hemolymph protein), whose sequence showshigh similarity to hemocyanins, and particularly pre-serves the residues involved, both directly and indirect-ly, in oxygen binding. We also study the EHP expression
Downloaded from https://academic.oup.com/mbe/article-abstract/15/4/415/1021391by gueston 04 April 2018

416 Sanchez et al.
pattern and the phylogenetic relationships of this insectprotein with the AHPH proteins. These data support amonophyletic origin of crustacean hemocyanins andhexamerins but also suggest that hexamerins haveevolved from hemocyanins present in the insect lineage.Moreover, the separate phylogenetic analyses of theAHPH domains suggest an independent evolution forthese protein domains. Our phylogenetic and multivar-iate analyses strongly suggest that EHP is an insect he-mocyanin, likely functioning in oxygen transport or stor-age during embryonic development.
Materials and MethodsProtein Purification, Microsequencing, and MolecularCloning of EHP cDNA
The EHP protein was accidentally purified by af-finity chromatography with the mAb 7D2 against theprotein Conulin (Sanchez, Ganfornina, and Bastiani1996) from the soluble fraction of embryonic grasshop-per (Schistocerca americana) lysates. After purification,the EHP protein was either N-terminally sequenced byautomated Edman degradation (Applied Biosystems477A) or digested with Endolysine-C (Lys-C; Boehrin-ger-Mannheim). The resulting peptides were then sepa-rated by reverse-phase HPLC and sequenced.
Degenerate oligonucleotides were designed fromthe peptide sequences to amplify fragments from grass-hopper cDNA using PCR with Taq DNA polymerase(Saiki et al. 1988). Embryos at 45% of development(staged by percentage of embryonic development;hatching in 20 days at 308C) were used to obtain thecDNA. A nested PCR strategy was used, and only thebands that reamplified with the expected size were stud-ied further. PCR was conducted in a thermal cycler (Per-kin Elmer Cetus), and cycling conditions were as fol-lows: one cycle of 948C for 2 min; 35 cycles of 948Cfor 30 s, 478C for 30 s, and 728C for 45 s; and a finalcycle of 728C for 5 min. A PCR product of 550 bp wasamplified, cloned into the pCR-II vector using the TAsystem (Invitrogen), and sequenced.
The 550-bp fragment was radiolabeled by randompriming (Prime-It II kit, Stratagene) and used as a probeto screen a cDNA library made from nerve cords dis-sected from 55% embryos. This library was constructedusing cDNA primed with oligo(dT) and directionallycloned using the lZAP system (Stratagene). Wescreened 0.5 3 106 plaque-forming units from the am-plified cDNA library. Several positive clones were ob-tained, and two were studied further. The clone EHP-8contains an insert of 2,310 bp and codes for the full-length EHP protein. The clone EHP-1 contains an insertof 2,067 bp and is identical to EHP-8, but is truncatedat the 59 end.
Both strands of the cDNA inserts were sequencedusing Sequenase (version 2.0, U.S. Biochemicals) andcustom primers. DNA and protein sequences were an-alyzed with the BLAST service (Altschul et al. 1990)and the GCG programs (Devereux, Haeberli, and Smith-ies 1984). To study the codon preference in the EHP-8open reading frame (ORF), a codon usage table was
made from available Schistocerca and Locusta codingsequences.
Analysis of EHP RNA Expression
In situ hybridizations were carried out according toa protocol for grasshopper whole-mount embryos (Gan-fornina, Sanchez, and Bastiani 1995). A digoxigenin-11-dUTP labeled (Genius-4 kit, Boehringer-Mannheim)RNA probe was synthesized using the EHP 550-bp frag-ment as template. Embryos were dissected and fixed inPEM-formaldehyde (37% formaldehyde 1:9 in 0.1 MPIPES, 2 mM EGTA, 1 mM MgSO4, pH 6.9) for 50min. After washing, they were incubated in hybridiza-tion solution (50% deionized formamide, 4 3 SSC, 250mg/ml yeast tRNA, 500 mg/ml salmon sperm DNA, 50mg/ml heparin, 0.1% Tween-20, 1 3 Denhardt’s solu-tion, 5% dextran sulfate) at 558C. The labeled RNAprobe was added at 0.5 mg/ml, and incubation proceededfor 36–48 h. After washes, the labeled RNA was de-tected with an alkaline phosphatase-conjugated anti-di-goxigenin antibody (Boehringer-Mannheim).
Phylogenetic Analysis
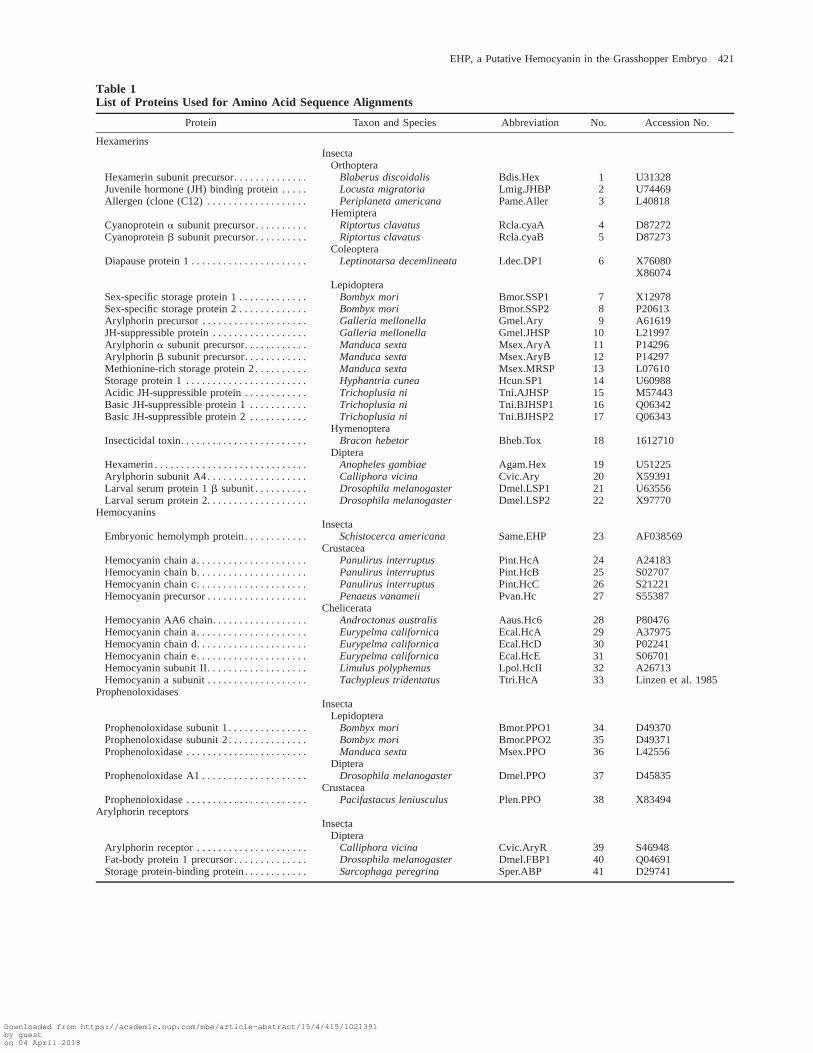
Sequences were retrieved from the databasesSwissProt, PIR, PDB, EMBL, and GenBank at their re-leases by February 13, 1997. The most recent entry wasconsidered when several sequences were available for agiven protein. Only the mature protein sequences wereused, either deduced from the known mature N-terminalamino acid or predicted by von Heijne’s (1990) method.Partial sequence entries were excluded. According toBeintema et al. (1994), proteins were named using anabbreviated species name followed by a functional label.Protein sequences were aligned with CLUSTAL W (1.6)(Thompson, Higgins, and Gibson 1994) using a PAMseries scoring matrix and a gap penalty mask based onthe aligned secondary structures of Pint.HcA andLpol.HcII. The alignment has been deposited in EMBL(accession number DS32507). Phylogenetic analyseswere carried out using the PHYLIP (3.5) (Felsenstein1993) and PAUP (3.0) (Swofford 1991) software pack-ages. Alignment gaps were considered missing data andnot used for phylogenetic inference. Corrected distanceswere calculated with the program PROTDIST using thePAM 001 matrix. The neighbor-joining (Saitou and Nei1987) method was used to reconstruct distance trees.Parsimony analyses were performed with the PROT-PARS program of the PHYLIP package, and with thePAUP software under a heuristic search option. Analysiswith PAUP was performed for comparison but is notshown in figures.
Multivariate Analysis
Principal-component analysis (PCA) on centeredvariables was performed with the program NetMul(Thioulouse and Chevenet 1996). The input for thisanalysis consisted of the mole percentage amino acidcompositions of the mature AHPH proteins used in thephylogenetic analyses, along with the compositions ob-tained by amino acid analysis of the locust LHP (de Kortand Koopmanschap 1987), Pint.HcA, Ecal.HcD, and
Downloaded from https://academic.oup.com/mbe/article-abstract/15/4/415/1021391by gueston 04 April 2018

EHP, a Putative Hemocyanin in the Grasshopper Embryo 417
FIG. 1.—Expression pattern of grasshopper EHP. In situ hybridi-zations in whole-mount grasshopper embryos with an antisense (A) orsense (B) RNA probe made from a 550-bp DNA fragment of the EHPcDNA. View from the dorsal surface with DIC optics. Anterior is up.Hemocytes attached to the basal membrane appear labeled, while nolabeling is observed in muscle (arrow) and neural (asterisk) cells. Scalebar 25 mm.
two hemocyanins from centipedes (Mangun et al. 1985).Given the difficulty for the amino acid analysis to dis-tinguish between residues Q-E, and N-D, the mole per-centage values of these pairs estimated from the proteinsequences were combined. Seventeen variables, includ-ing 15 residues and the combinations Q 1 E and N 1D, were considered for the analysis.
Results and DiscussionIdentification of Grasshopper EHP as a Soluble ProteinSynthesized by Hemocytes During EmbryonicDevelopment
The grasshopper EHP was identified during puri-fication of the neural protein Conulin (Sanchez, Gan-fornina, and Bastiani 1996). After purification and Lys-C digestion, the sequences of three EHP peptides wereobtained. These sequences were used to design degen-erate oligonucleotides to PCR-amplify the embryoniccDNA coding for the protein. A 550-bp DNA fragmentwas obtained that shows a unique ORF and containedregions coding for the N-terminal peptide and an inter-nal peptide. We studied the spatial localization of EHPmRNA using in situ hybridization to whole-mountgrasshopper embryos. Sense and antisense digoxigenin-labeled RNA probes were generated using the 550-bpPCR fragment. EHP mRNA expression is detected at45% of development in sessile hemocytes, shown in fig-ure 1A over the basal membrane of the ventral nervecord. Neurons and muscle cells are highlighted with as-terisks and arrows, respectively, to show the absence oflabeling in those cell types. No labeling was observedin the cells of the fat body (not shown), the place ofsynthesis of insect storage proteins (Telfer and Kunkel1991). Hybridization is absent in embryos exposed tothe sense RNA probe (fig. 1B), indicating that the signalobserved with the antisense probe is specific for the en-dogenous mRNA. These results indicate that EHPmRNA is present during embryonic development andthat EHP is synthesized by hemocytes.
The purification of the protein from the solublefraction of embryonic lysates, its hemocyte origin, andthe presence of a signal sequence in the molecule (seebelow) suggest that EHP is exported into the hemo-lymph. It was consequently named embryonic hemo-lymph protein. Intriguingly, EHP is present as a solubleprotein at 45% of development, long before dorsal clo-sure and heart functioning (which occurs at 65%–70%of development), and thus before a true hemolymph isformed. Therefore, at early developmental stages, EHPis probably free in the yolk sac.
EHP-8, the full-length cDNA coding for EHP, hasa 2,310-bp insert that includes the three sequenced pep-tides (underlined in fig. 2A). It comprises an ORF of2,025 bp after a 59 untranslated region (UTR) of 27 bp.The 550-bp probe hybridizes to bp 91–664. The firstmethionine codon encountered in frame in the EHP-8clone is in an appropriate context to be the translationinitiation site according to Cavener and Ray (1991) andby comparison with the nucleotide flanking sequencesfound in other grasshopper genes. The correctness of theORF defined above is further supported by the GC biasat every third base and the codon preference analysis.Two potential polyadenylation signals are found in the257-bp 39 UTR, which ends with a poly-A sequence.
The N-terminal end of EHP predicted by this ORFis highly hydrophobic (fig. 2B) and may represent aleader signal peptide. The predicted cleavage site for thesignal peptide (von Heijne 1990) (arrowhead in fig. 2A)agrees with the N-terminal protein sequence obtained.This and the lack of other suspected membrane-anchor-ing regions indicates that EHP is a secreted protein. Thepredicted mature EHP would have a molecular mass of76.2 kDa and an isoelectric point of 6.38. EHP has twopotential N-glycosylation sites (residues N172 andN317, circled in fig. 2A), and five cysteines (boxed infig. 2A) that may form disulfide bonds.
Similarity of EHP to Crustacean Hemocyanins;Phylogenetic and Multivariate Analysis of the AHPHSuperfamily
When the sequence of the mature EHP is comparedwith those of other known proteins, it shows the highestsimilarity with subunit A of the lobster Panulirus inter-ruptus hemocyanin (Pint.HcA; Bak and Beintema 1987).The Pint.HcA and EHP sequences align with minorgaps, and the overall pairwise similarity (43% identity;64% when considering conservative substitutions) ishigher than that between crustacean and cheliceratan he-mocyanins (31%–33% identity; Beintema et al. 1994).The grasshopper EHP showed significant global simi-larity with hemocyanins, insect hexamerins, and PPOs.Therefore, we studied the phylogenetic relationships ofthese protein families (see fig. 3 and table 1), includingthe grasshopper EHP. After aligning the amino acid se-quences, two methods were used to find phylogeneticrelationships (corrected distances and maximum parsi-mony, see Materials and Methods). Trees inferred withboth methods showed identical topologies for the mainbranches (fig. 3). Numbers at nodes indicate bootstrappercentage from 100 replicates. A majority rule of 50%
Downloaded from https://academic.oup.com/mbe/article-abstract/15/4/415/1021391by gueston 04 April 2018

418 Sanchez et al.
→
FIG. 2.—A, cDNA and deduced protein sequence of EHP. Nucleotide numbers are on the left, and amino acid numbers are on the right(referred to the first methionine). The cleavage site of the signal peptide is marked by an arrowhead. Two potential N-linked glycosylation sitesare circled. Cysteine residues are boxed. The N-terminal peptide and the peptides obtained by Lys-C digestion are underlined. The polyadenylationsites are shown in italics. B, Hydropathy plot of the predicted protein sequence determined by Kyte and Doolittle’s (1982) method using awindow of nine residues. The hydrophobic domain at the N-terminal region of the protein represents the signal peptide.
was established, unsupported nodes were excluded, andtheir branches were forced to yield polytomies. We usedPPOs as an outgroup due to their assumed presence inall arthropod groups and their closer relationship withcheliceratan hemocyanins, which suggest a common an-cestry with hemocyanins before the separation of themain arthropod groups. Three main protein groups aredetermined. The PPOs and cheliceratan hemocyaninsboth appear as different clades, while a single mono-phyletic group is formed by crustacean hemocyaninsand insect hexamerins. The grasshopper EHP groupswith the crustacean hemocyanins with a well supportednode, and consistently separates from the monophyletichexamerins. The internal branching pattern of crusta-cean hemocyanins and PPOs is well supported, but thatof cheliceratan hemocyanins shows some unsupportednodes. A number of nodes organizing the hexamerin treehave low bootstrap values and vary when parsimony ordistance methods are considered, as previously reported(Beintema et al. 1994; Burmester and Scheller 1996).However, some groups, like the methionine-rich hex-amerins (sequences 7, 13, 14, 16, and 17 of table 1),and the dipteran (sequences 19–21 of table 1) and lep-idopteran (sequences 8, 9, 11, and 12 of table 1) aryl-phorins consistently separate in distinct clusters. Otherhexamerins that group together are the juvenile hor-mone-suppressible proteins (sequences 10 and 15 of ta-ble 1), the hexamerins from two cockroaches (sequences1 and 3 of table 1), and the Rcla.cyaA-B cyanoproteins(sequences 4 and 5 of table 1). The poorly supportedtopology and the longer overall branch length of thehexamerin distance tree (not shown) imply a higher di-vergence in the sequences of these insect proteins, whichprobably relates to a release of their sequences fromfunctional constraints, a conclusion also reached by Bur-mester and Scheller (1996).
A PCA of the amino acid content of the AHPHproteins under study (fig. 4) was used as a different ap-proach to establish groups based on biochemical fea-tures other than protein sequence. This analysis also al-lowed us to include the locust LHP (de Kort and Koop-manschap 1987) and two centipede hemocyanins (Man-gun et al. 1985) whose amino acid sequences are notavailable yet. PCA shows hemocyanins and PPOs ascompact and close groups, while hexamerins form ascattered group with three to four main clusters. One ofthese clusters (I in fig. 4A) clearly segregates the me-thionine-rich hexamerins, while others (II, III, and IV infig. 4A and C) group three dipteran arylphorins, lepi-dopteran arylphorins and the Bdis.Hex, and two juvenilehormone-suppressible proteins, respectively. An inde-pendent PCA (fig. 4C) was carried out with the proteinsgrouped in the lower left quadrant of the plot in figure4A. PPOs and hemocyanins clearly become apart, and
EHP and the centipede proteins group with hemocya-nins. LHP segregates between EHP and Lmig.JHBP (ar-rowhead in fig. 4C). Although EHP is recognized by aserum anti-LHP (kindly provided by S. C. de Kort; datanot shown), PCA analysis questions an orthologous re-lationship between these two proteins. The validity ofPCA for including proteins with amino acid compositionobtained biochemically was demonstrated by the inclu-sion in the hemocyanin cluster of the experimental res-idue composition of Pint.HcA and Ecal.HcD (dotted cir-cles in fig. 4A and C).
Analysis of the Evolutionary History and Functions ofthe Three AHPH Protein Domains
Arthropodan hemocyanins are structurally com-posed of three domains (Volbeda and Hol 1989a; Hazeset al. 1993), with domain 2 being directly involved inoxygen transport. This oxygen-binding domain, in par-ticular the pair of helices forming each copper site, hasbeen proposed to be the ancestral structural charactershared by otherwise unrelated oxygen-binding proteins(Volbeda and Hol 1989b). Arthropodan hemocyanins in-herited this domain, but two new protein domains wereapparently added during evolution. Domain 1 is veryimportant for the allosteric regulation of oxygen binding(Magnus et al. 1994) and seems to determine the extentof the oligomerization (Hazes et al. 1993). Domain 3 isstructurally very different from the others, as it containsa seven-stranded Greek key b barrel. It contains a pu-tative calcium-binding site that can play a structural andregulatory role (Hazes et al. 1993).
The overall sequence conservation exhibited byAHPH proteins suggests a preserved tertiary structurewith three distinct domains that were probably presentin the ancestor of these proteins. This prompted us toinvestigate the existence of different evolutionary trendsin each individual domain. If they have followed similarpathways, one may expect congruency between the to-pology of the tree inferred from the whole sequence (fig.3) and that of those reconstructed from the separate do-mains. The initial alignment was subdivided accordingto the domains of Pint.HcA (Volbeda and Hol 1989a).Corrected distances were used to infer phylogenetictrees of the three domains separately (fig. 5). The overalltree topologies of the different domains are similar tothose inferred from the entire sequence (separating theclades of PPOs, cheliceratan hemocyanins, and a groupthat joins crustacean hemocyanins, grasshopper EHP,and insect hexamerins), as was observed in previous re-ports (Beintema et al. 1994; Burmester and Scheller1996). However, important differences emerge from ouranalysis.
Downloaded from https://academic.oup.com/mbe/article-abstract/15/4/415/1021391by gueston 04 April 2018
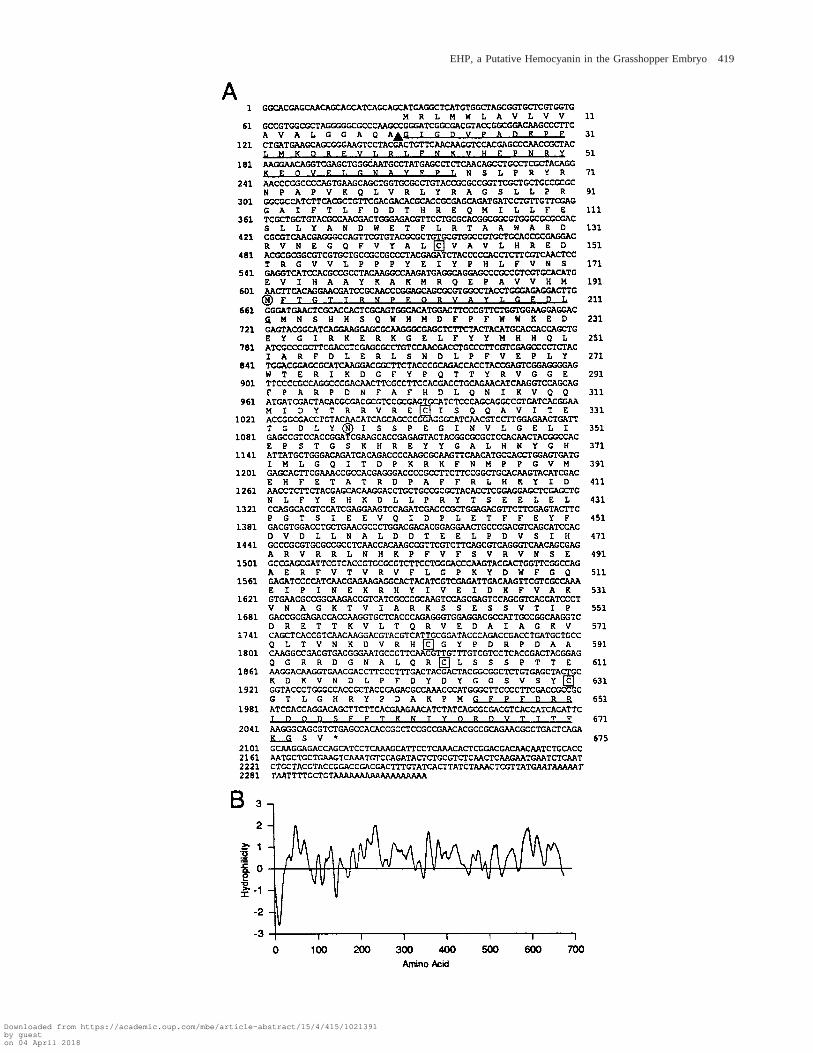
EHP, a Putative Hemocyanin in the Grasshopper Embryo 419
Downloaded from https://academic.oup.com/mbe/article-abstract/15/4/415/1021391by gueston 04 April 2018

420 Sanchez et al.
FIG. 3.—Phylogenetic analysis of the AHPH proteins listed in table 1 derived from a protein sequence alignment conducted with CLUSTALW. The secondary structures of two hemocyanins (Lpol.HcII and Pint.HcA) were used to guide the alignment. The topologies of trees inferredwith distance matrix (left side) or maximum parsimony (right side) methods are shown. The trees were rooted with PPOs as outgroup. Branchlengths are not shown. Numbers at nodes are bootstrap values from 100 replicates. A 50% majority rule was used to yield polytomies.
Position of Grasshopper EHP in the Domain Trees
The grasshopper EHP and crustacean hemocyaninsform a monophyletic group for domains 1 and 2. Theseresults add support to the hypothesis that EHP is aninsect hemocyanin. However, EHP becomes groupedwith hexamerins when analyzing domain 3. Both topol-ogies support the existence of a common ancestor forhexamerins and crustacean hemocyanins as proposed byBurmester and Scheller (1996), but the existence of amonophyletic association of EHP and the hexamerinclade for domain 3 refines this proposal. This result sug-gests that unless convergence is invoked, hexamerinshave evolved from insect hemocyanins, with which theystill share higher similarity in domain 3, and not directlyfrom crustacean hemocyanins. Taken together, these datareinforce the current view of a monophyletic origin ofinsects and crustacea (Averof and Akam 1995).
Domainal Analysis of Different AHPH Clades
The tree topologies observed in the PPO clade ap-pear to be well maintained. The branch lengths are lon-ger for domain 1, which might be explained by the pro-
teolytic separation of domain 1 from the active PO form(Aspan et al. 1995). This would liberate domain 1 fromfunctional constraints, enabling this domain to evolvewith higher rates of change in different arthropods.
The branching patterns of the cheliceratan hemo-cyanins broadly vary depending on the domain consid-ered, but are generally weakly supported. This variationcan be due to the presence of hemocyanins from severalclasses and orders of chelicerates in the sample of se-quences available. These hemocyanins show differencesin their quaternary structures (Van Holde and Miller1995) that could be reflected in their sequences. Thebranch lengths in this clade also differ depending on thedomain under study. The longest are those of domain 1,and the shortest are those of the oxygen-binding do-main 2.
Crustacean hemocyanins show a well-supportedbranching pattern with only a slight change in domain2. The branch lengths show the same tendency as inother hemocyanins and PPOs. The shorter distances ob-served in domain 2 suggest a higher divergence in thedomains not directly related to the oxygen-binding func-
Downloaded from https://academic.oup.com/mbe/article-abstract/15/4/415/1021391by gueston 04 April 2018
EHP, a Putative Hemocyanin in the Grasshopper Embryo 421
Table 1List of Proteins Used for Amino Acid Sequence Alignments
Protein Taxon and Species Abbreviation No. Accession No.
Hexamerins
Hexamerin subunit precursor. . . . . . . . . . . . . .Juvenile hormone (JH) binding protein . . . . .Allergen (clone (C12) . . . . . . . . . . . . . . . . . . .
InsectaOrthoptera
Blaberus discoidalisLocusta migratoriaPeriplaneta americana
Hemiptera
Bdis.HexLmig.JHBPPame.Aller
123
U31328U74469L40818
Cyanoprotein a subunit precursor. . . . . . . . . .Cyanoprotein b subunit precursor. . . . . . . . . .
Diapause protein 1 . . . . . . . . . . . . . . . . . . . . . .
Riptortus clavatusRiptortus clavatus
ColeopteraLeptinotarsa decemlineata
Rcla.cyaARcla.cyaB
Ldec.DP1
45
6
D87272D87273
X76080X86074
Sex-specific storage protein 1 . . . . . . . . . . . . .Sex-specific storage protein 2 . . . . . . . . . . . . .Arylphorin precursor . . . . . . . . . . . . . . . . . . . .JH-suppressible protein . . . . . . . . . . . . . . . . . .
LepidopteraBombyx moriBombyx moriGalleria mellonellaGalleria mellonella
Bmor.SSP1Bmor.SSP2Gmel.AryGmel.JHSP
789
10
X12978P20613A61619L21997
Arylphorin a subunit precursor. . . . . . . . . . . .Arylphorin b subunit precursor. . . . . . . . . . . .Methionine-rich storage protein 2 . . . . . . . . . .Storage protein 1 . . . . . . . . . . . . . . . . . . . . . . .Acidic JH-suppressible protein . . . . . . . . . . . .
Manduca sextaManduca sextaManduca sextaHyphantria cuneaTrichoplusia ni
Msex.AryAMsex.AryBMsex.MRSPHcun.SP1Tni.AJHSP
1112131415
P14296P14297L07610U60988M57443
Basic JH-suppressible protein 1 . . . . . . . . . . .Basic JH-suppressible protein 2 . . . . . . . . . . .
Insecticidal toxin. . . . . . . . . . . . . . . . . . . . . . . .
Trichoplusia niTrichoplusia ni
HymenopteraBracon hebetor
Tni.BJHSP1Tni.BJHSP2
Bheb.Tox
1617
18
Q06342Q06343
1612710
Hexamerin . . . . . . . . . . . . . . . . . . . . . . . . . . . . .Arylphorin subunit A4. . . . . . . . . . . . . . . . . . .Larval serum protein 1 b subunit . . . . . . . . . .Larval serum protein 2. . . . . . . . . . . . . . . . . . .
DipteraAnopheles gambiaeCalliphora vicinaDrosophila melanogasterDrosophila melanogaster
Agam.HexCvic.AryDmel.LSP1Dmel.LSP2
19202122
U51225X59391U63556X97770
Hemocyanins
Embryonic hemolymph protein. . . . . . . . . . . .Insecta
Schistocerca americana Same.EHP 23 AF038569
Hemocyanin chain a. . . . . . . . . . . . . . . . . . . . .Hemocyanin chain b. . . . . . . . . . . . . . . . . . . . .Hemocyanin chain c. . . . . . . . . . . . . . . . . . . . .Hemocyanin precursor . . . . . . . . . . . . . . . . . . .
CrustaceaPanulirus interruptusPanulirus interruptusPanulirus interruptusPenaeus vanameii
Pint.HcAPint.HcBPint.HcCPvan.Hc
24252627
A24183S02707S21221S55387
Hemocyanin AA6 chain. . . . . . . . . . . . . . . . . .Hemocyanin chain a. . . . . . . . . . . . . . . . . . . . .Hemocyanin chain d. . . . . . . . . . . . . . . . . . . . .Hemocyanin chain e. . . . . . . . . . . . . . . . . . . . .Hemocyanin subunit II. . . . . . . . . . . . . . . . . . .Hemocyanin a subunit . . . . . . . . . . . . . . . . . . .
ChelicerataAndroctonus australisEurypelma californicaEurypelma californicaEurypelma californicaLimulus polyphemusTachypleus tridentatus
Aaus.Hc6Ecal.HcAEcal.HcDEcal.HcELpol.HcIITtri.HcA
282930313233
P80476A37975P02241S06701A26713Linzen et al. 1985
Prophenoloxidases
Prophenoloxidase subunit 1 . . . . . . . . . . . . . . .Prophenoloxidase subunit 2 . . . . . . . . . . . . . . .Prophenoloxidase . . . . . . . . . . . . . . . . . . . . . . .
InsectaLepidoptera
Bombyx moriBombyx moriManduca sexta
Bmor.PPO1Bmor.PPO2Msex.PPO
343536
D49370D49371L42556
Prophenoloxidase A1 . . . . . . . . . . . . . . . . . . . .
Prophenoloxidase . . . . . . . . . . . . . . . . . . . . . . .
DipteraDrosophila melanogaster
CrustaceaPacifastacus leniusculus
Dmel.PPO
Plen.PPO
37
38
D45835
X83494Arylphorin receptors
Arylphorin receptor . . . . . . . . . . . . . . . . . . . . .Fat-body protein 1 precursor . . . . . . . . . . . . . .Storage protein-binding protein. . . . . . . . . . . .
InsectaDiptera
Calliphora vicinaDrosophila melanogasterSarcophaga peregrina
Cvic.AryRDmel.FBP1Sper.ABP
394041
S46948Q04691D29741
Downloaded from https://academic.oup.com/mbe/article-abstract/15/4/415/1021391by gueston 04 April 2018

422 Sanchez et al.
FIG. 4.—Principal-component analyses on centered variables ofthe amino acid compositions of selected AHPH proteins. A, Projectionof the two most significant components (1 5 36% of the total variance;2 5 19% of the total variance) of all the AHPH proteins (names de-tailed in table 1). C, Projection of the two most significant components(1 5 28% of the total variance; 2 5 18% of the total variance) whenonly the proteins segregated in the lower left quadrant of plot A wereconsidered. B and D, Amino acid (single-letter code) contribution tothe two first components obtained in A and C, respectively. Symbolidentification is shown on the right. Hollow symbols show amino acidcompositions calculated from the mature protein sequence. Crossedhollow circles are the Pint.HcA and Ecal.HcD compositions calculatedfrom the mature protein sequences. Dotted hollow circles are thePint.HcA and Ecal.HcD compositions obtained by amino acid analysis.These and the centipede hemocyanin compositions are reported in(Mangun et al. 1985). Locust LHP composition obtained from (de Kortand Koopmanschap 1987). Arrows point at the juvenile hormone-sup-pressible hexamerins. Arrowhead indicates Lmig.JHBP. The clusters ofhemocyanins, PPOs, and hexamerins are enclosed by continuous, shortdashed, and long dashed lines, respectively. Roman numbers representdifferent hexamerin clades (see Results for details).
tion. However, we might expect this clade topology tochange substantially when hemocyanins from othercrustacean classes are described.
The same difficulties in assigning hexamerin rela-tionships obtained from the entire protein sequences areobserved when individual domains are considered. Al-though the internal branching patterns of hexamerins aredisparate, several clades are relatively well supported,such as the methionine-rich hexamerins, the juvenilehormone-suppressible proteins, and the two groups ofarylphorins. It is important to emphasize the lightly sus-tained topology in domain 1, where even the monophy-letic nature of hexamerins is challenged with a 38%bootstrap node. Distances are larger for domains 2 and1, which can be explained by the missing oxygen-bind-ing function reported in these insect proteins. In con-trast, domain 3 displays shorter and more homogeneousdistances.
With respect to domain 3, we did separate distanceand parsimony analyses (not shown) including three ar-ylphorin receptors that seem to be phylogenetically re-lated to their arylphorin ligands (Burmester and Scheller1996). Only domain 3 was considered, as the other twodomains show nonsignificant sequence similarities whenall the AHPH proteins are included in the analysis. Thearylphorin receptors form a monophyletic clade withinthe hexamerins in distance and parsimony phylogenies,in contrast to a previous report (Burmester and Scheller1996) in which the arylphorin receptors and hexamerinsseparate in different clades. The different grouping can-not be explained by substantial alignment differences,but instead might be the result of the larger number ofsequences included in our analysis.
Functional and Structural Conservation of Domain 3
The divergence among the distinct clades of AHPHproteins, as derived from the corrected distances of themain branches of the trees, also indicate that domain 1is the most variable, while domains 2 and 3 are wellpreserved. The conservation of domain 2 in hemocya-nins and PPOs is not surprising, given the functionalsimilarities between those proteins. However, the pres-ervation of domain 3, not only in hemocyanins andPPOs, where it could play an important role in the reg-ulation of copper-binding, but also in the highly diver-gent hexamerins and their receptors, is interesting.
The secondary structure of domain 3 appears tohave convergently evolved in a number of other proteins(Hazes and Hol 1992), and it could be argued that itspreservation is merely due to a structural requirementfor the AHPH proteins. However, it is clear from otherprotein families (e.g., the lipocalins) that a strongly con-served b barrel topology can be achieved without thenecessity of a well-preserved primary structure. Ouranalysis shows a highly conserved sequence for the do-main 3 of the AHPH proteins and leads us to proposeits further preservation for functional reasons. Given itsouter location in the hexamer (Volbeda and Hol 1989a;Hazes et al. 1993), we hypothesize a role in protein–protein interactions such as those involving receptor-me-diated endocytosis of hexamerins (Wang and Hauner-land 1994) and those of PPOs with other proteins par-ticipating in cellular defense reactions (Aspan et al.1995). The inner cavity of the barrel could also be usedfor binding ligands such as biliverdin (found in the hex-amerins Rcla.cyaA-B [Chinzei et al. 1990] and in a lo-cust hexamerin [De Bruyn, Koopmanschap, and de Kort1986]) or riboflavin (found in several hexamerins; Mil-ler and Silhacek 1995). Moreover, domain 3 has beenproposed to be conserved between the AHPH proteinsand the yeast and vertebrate tyrosinases (Kupper et al.1989), where this domain is proteolytically separatedfrom the active form of the enzyme. The tyrosinases andmolluscan hemocyanins are thought to be ancestrally re-lated to some AHPH proteins due to the similarity ex-hibited in the pair of a-helices of copper site B (VanHolde and Miller 1995). We propose as a working hy-pothesis that domain 3 was part, together with these a-
Downloaded from https://academic.oup.com/mbe/article-abstract/15/4/415/1021391by gueston 04 April 2018

EHP, a Putative Hemocyanin in the Grasshopper Embryo 423
FIG. 5.—Phylogenetic relationships of the three separate domains of the AHPH proteins estimated by a distance matrix method. Domainboundaries are taken from Pint.HcA (Volbeda and Hol 1989a). Protein sequences are numbered as shown in table 1. Boxes, circles, and diamondsrepresent hexamerins, hemocyanins, and PPOs, respectively. EHP is represented as a black circle. Bootstrap values (100 replicates) are shownwith a 50% majority rule.
helices, of the ancestral protein that originated hemo-cyanins and tyrosinases.
In summary, our results support the view that thethree domains present in the AHPH proteins are evolv-ing as independent modules, under different selectivepressures and functional constraints.
Sequence Analysis Strongly Supports a FunctionalRelationship of EHP with Hemocyanins
The results above suggest a well-supported rela-tionship of EHP with crustacean hemocyanins and,therefore, the existence of hemocyanins in insects. Tobetter substantiate this proposal, we studied in detail theresidues conserved between EHP and Pint.HcA. Theoverall similarity between these two proteins becomeseven more significant when the oxygen-binding domain2 (56% identity) is considered separately. An alignmentof this domain of EHP with two hemocyanins and se-
lected PPOs and hexamerins is shown in figure 6. EHPshares with hemocyanins and PPOs the six copper-bind-ing histidine residues. Moreover, the grasshopper proteinpreserves every residue involved in the hydrogen-bond-ing network of the dinuclear copper site and 80% of theresidues forming the hydrophobic core close to the cop-per atoms of Pint.HcA. The residue F49 in domain 1(not shown), key in the allosteric regulation of oxygenbinding, and E309, which controls the entrance of ox-ygen to the coppers, are also conserved in EHP (Hazeset al. 1993; Magnus et al. 1994). Finally, two out of thefive cysteine residues of EHP (C562, C611) are sharedwith Pint.HcA, where they form a disulfide bond in-volved in the formation of cavities surrounding the cop-per sites. This disulfide link is conserved in hemocya-nins and PPOs but is present only in some hexamerins.The extensive sequence similarity suggests that EHPprobably contains copper that would allow it to bind
Downloaded from https://academic.oup.com/mbe/article-abstract/15/4/415/1021391by gueston 04 April 2018

424 Sanchez et al.
FIG. 6.—Alignment of the oxygen-binding domain of hemocyanins with EHP and selected hexamerins and prophenoloxidases (see table 1for protein identification). Domain 2 boundaries are taken from Pint.HcA (Volbeda and Hol 1989a). The sequences were aligned with CLUSTALW. Gray boxes show residue identities of EHP either with Pint.HcA or with more than three of the selected proteins. Important residues involvedin oxygen binding and oligomerization of Pint.HcA are highlighted as detailed in the figure.
molecular oxygen. Moreover, residues involved in thequaternary structure of Pint.HcA (dimer and trimer con-tact sites in the hexamer) are also significantly con-served in EHP (fig. 6). These residues are also con-served in hexamerins and suggest a common hexamericstructure for the whole clade.
Several other features of EHP appear to be relatedto this evolutionary history. EHP is a secreted protein,a character shared with hexamerins and crustacean he-mocyanins, and not with cheliceratan hemocyanins andPPOs, which lack a signal peptide. The secreted natureof crustacean hemocyanins is based on the signal pep-tide present in Pvan.Hc (Sellos, Lemoine, and VanWormhoudt 1997). However, EHP is synthesized by he-mocytes, while crustacean hemocyanins and insect hex-amerins are synthesized by the hepatopancreas (VanHolde and Miller 1995) and the fat body (Telfer andKunkel 1991), respectively. Like EHP, PPOs are synthe-sized by hemocytes (Kawabata et al. 1995), and theseenzymes are expected to be present in grasshoppers.However, the sequence similarity values, the numerousand distinctly situated gaps present in the alignment ofEHP with insect PPOs, and the differences in their bio-synthetic pathways and posttranslational modifications
of these proteins do not support a functional relationshipbetween EHP and PPOs.
How do we interpret the sequence relationships ofEHP with hemocyanins? One could consider EHP to bea nonfunctional relic of hemocyanins in insects. Wethink, however, that this is an unlikely hypothesis, giventhe relative abundance of this embryonic protein and thehigh divergence rates that it should have experiencedwhen released from a functional constraint, as is clearfor hexamerins (see above). A second alternative is thatEHP represents an ancestral state in the evolution ofhexamerins. Orthopteran hexamerins are expected to bemore closely related to crustacean hemocyanins, giventhat orthopteroids are primitive insects with a long, in-dependent evolution. This view is supported by the dis-position of Lmig.JHBP, Bdis.Hex, and Pame.Aller closeto the base of the hexamerin clade in our trees. However,EHP never groups with these orthopteran proteins,which eliminates the possibility that EHP is an ancestralinsect hexamerin.
In summary, sequence similarity and other molec-ular features suggest a close relationship of EHP andhemocyanins. This is surprising, given the apparent lackof hemocyanins in arthropods equipped with an efficient
Downloaded from https://academic.oup.com/mbe/article-abstract/15/4/415/1021391by gueston 04 April 2018

EHP, a Putative Hemocyanin in the Grasshopper Embryo 425
tracheal respiratory system. However, EHP is presentduring developmental stages in which the tracheal sys-tem is not fully developed. We do not know whetherEHP expression is restricted to embryonic life, whichmight be a reason why EHP-like proteins have not beenfound in other insects in which only postembryonic de-velopment has been explored. Based on the currentlyavailable data on grasshopper EHP, a question arises:What is the role of this protein during the embryogen-esis of this orthopteran? Given its presence as a solubleprotein in the yolk sac, we propose that EHP might betransporting or storing molecular oxygen in the egg. Ox-ygen is presumably transported in the egg by simplediffusion (Chapman 1971), but an ancillary pathway in-volving an oxygen transport protein might be a safetymechanism during periods of low oxygen supply to theegg or periods of accelerated embryonic growth withhigh metabolic rates. A similar function has been dem-onstrated for hemoglobins found in solution in the he-molymph of some aquatic insects (Chapman 1971;Markl and Decker 1992). In spite of the plausibility ofthis hypothesis, a definitive demonstration for a role ofEHP as an oxygen transport protein and its prevalencein insects will require further investigations. However,the existence of insect proteins with strong sequencesimilarity to hemocyanins raises interesting questionsabout their physiological role and their evolutionary re-lationships within the AHPH protein superfamily.
During the review process of this article, a newcrustacean hemocyanin from the Dungeness crab Can-cer magister was reported (Durstewitz and Terwilliger1997), and its phylogenetic relationship with somemembers of the AHPH family was analyzed. A parsi-mony analysis groups the crab hemocyanin withPint.HcC.
Acknowledgments
We want to thank S. C. de Kort for his gift of thelocust LHP anti-serum, and E. Ball, J. Law, R. Doolittle,J. Seger, and two anonymous reviewers for helpful com-ments on the manuscript. We also thank M. Herrera forhelp with protein purification. L. Giuli, a high schoolstudent sponsored by a Howard Hughes fellowship, en-thusiastically helped us with the molecular cloning. Pep-tide sequencing and synthesis of oligonucleotides wereperformed at the Protein/DNA Core Facility of the UtahCancer Center under the direction of R. W. Schackmann.This work was supported by an NIH grant to M.J.B.(NS25387). G.G. held an HFSP fellowship (SF-474/96).
LITERATURE CITED
ALTSCHUL, S. F., W. GISH, W. MILLER, E. W. MYERS, and D.J. LIPMAN. 1990. Basic local alignment search tool. J. Mol.Biol. 215:403–410.
ASPAN, A., T. HUANG, L. CERENIUS, and K. SODERHALL. 1995.cDNA cloning of prophenoloxidase from the freshwatercrayfish Pacifastacus leniusculus and its activation. Proc.Natl. Acad. Sci. USA 92:939–943.
AVEROF, M., and M. AKAM. 1995. Insect-crustacean relation-ships: insights from comparative developmental and molec-
ular studies. Philos. Trans. R. Soc. Lond. B Biol. Sci. 347:293–303.
BAK, H. J., and J. J. BEINTEMA. 1987. Panulirus interruptushemocyanin: the elucidation of the complete amino acidsequence of subunit A. Eur. J. Biochem. 169:333–348.
BEINTEMA, J. J., W. T. STAM, B. HAZES, and M. P. SMIDT.1994. Evolution of arthropod hemocyanins and insect stor-age proteins (hexamerins). Mol. Biol. Evol. 11:493–503.
BURMESTER, T., and K. SCHELLER. 1996. Common origin ofarthropod tyrosinase, arthropod hemocyanin, insect hex-amerin, and dipteran arylphorin receptor. J. Mol. Evol. 42:713–728.
CAVENER, D. R., and S. C. RAY. 1991. Eukaryotic start andstop translation sites. Nucleic Acids Res. 19:3185–3192.
CHAPMAN, R. F. 1971. The insects. Structure and function.Elsevier, New York.
CHINZEI, Y., T. HARUNA, K. MIURA, H. NUMATA, and S. NA-KAYAMA. 1990. Purification and characterization of bili-verdin-associated cyanoprotein from eggs and hemolymphof the bean bug, Riptortus clavatus (Heteroptera: Alydidae).Insect Biochem. 20:545–555.
DE BRUYN, S. M., A. B. KOOPMANSCHAP, and C. A. D. DE
KORT. 1986. High-molecular-weight serum proteins fromLocusta migratoria: identification of a protein specificallybinding juvenile hormone III. Physiol. Entomol. 11:7–17.
DE KORT, C. A. D., and A. B. KOOPMANSCHAP. 1987. Isolationand characterization of a larval hemolymph protein in Lo-custa migratoria. Arch. Insect Biochem. Physiol. 4:191–202.
DEVEREUX, J., P. HAEBERLI, and O. SMITHIES. 1984. A com-prehensive set of sequence analysis programs for the VAX.Nucleic Acids Res. 12:387–395.
DURSTEWITZ, G., and N. B. TERWILLIGER. 1997. cDNA clon-ing of a developmentally regulated hemocyanin subunit inthe crustacean Cancer magister and phylogenetic analysisof the hemocyanin gene family. Mol. Biol. Evol. 14:266–276.
FELSENSTEIN, J. 1993. PHYLIP: phylogeny inference package.Distributed by the author, Department of Genetics, Univer-sity of Washington, Seattle.
FUJIMOTO, K., N. OKINO, S. KAWABATA, S. IWANAGA, and E.OHNISHI. 1995. Nucleotide sequence of the cDNA encod-ing the proenzyme of phenol oxidase A1 of Drosophila mel-anogaster. Proc. Natl. Acad. Sci. USA 92:7769–7773.
GANFORNINA, M. D., D. SANCHEZ, and M. J. BASTIANI. 1995.Lazarillo, a new GPI-linked surface lipocalin, is restrictedto a subset of neurons in the grasshopper embryo. Devel-opment 121:123–134.
HALL, M., T. SCOTT, M. SUGUMARAN, K. SODERHALL, and J.H. LAW. 1995. Proenzyme of Manduca sexta phenol oxi-dase: purification, activation, substrate specificity of the ac-tive enzyme, and molecular cloning. Proc. Natl. Acad. Sci.USA 92:7764–7768.
HAZES, B., and W. G. J. HOL. 1992. Comparison of the he-mocyanin b-barrel with other Greek key b-barrels: possibleimportance of the ‘‘b-zipper’’ in protein structure and fold-ing. Proteins Struct. Funct. Genet. 12:278–298.
HAZES, B., K. A. MAGNUS, C. BONAVENTURA, J. BONAVEN-TURA, Z. DAUTER, K. H. KALK, and W. G. J. HOL. 1993.Crystal structure of deoxygenated Limulus polyphemussubunit II hemocyanin at 2.18 A resolution: clues for amechanism for allosteric regulation. Protein Sci. 2:597–619.
JOHANSSON, M. W., and K. SODERHALL. 1996. The prophe-noloxidase activating system and associated proteins in in-vertebrates. Prog. Mol. Subcell. Biol. 15:46–66.
KAWABATA, T., Y. YASUHARA, O. MASANORI, S. MATSUURA,and M. ASHIDA. 1995. Molecular cloning of insect pro-
Downloaded from https://academic.oup.com/mbe/article-abstract/15/4/415/1021391by gueston 04 April 2018

426 Sanchez et al.
phenol oxidase: a copper-containing protein homologous toarthropod hemocyanin. Proc. Natl. Acad. Sci. USA 92:7774–7778.
KUPPER, U., D. M. NIEDERMANN, G. TRAVAGLINI, and K.LERCH. 1989. Isolation and characterization of the tyrosi-nase gene from Neurospora crassa. J. Biol. Chem. 264:17250–17258.
KYTE, J., and R. F. DOOLITTLE. 1982. A simple method fordisplaying the hydropatic character of a protein. J. Mol.Biol. 157:105–132.
LINZEN, B., N. M. SOETER, A. F. RIGGS et al. (17 co-authors).1985. The structure of arthropod hemocyanin. Science 229:519–524.
MAGNUS, K. A., B. HAZES, H. TON-THAT, C. BONAVENTURA,J. BONAVENTURA, and W. G. J. HOL. 1994. Crystallograph-ic analysis of oxygenated states of arthropod hemocyaninshows unusual differences. Proteins 19:302–309.
MANGUN, C. P., J. L. SCOTT, R. E. L. BLACK, K. I. MILLER,and K. E. VAN HOLDE. 1985. Centipedal hemocyanin: itsstructure and its implications for arthropod phylogeny. Proc.Natl. Acad. Sci. USA 82:3721–3725.
MARKL, J., and H. DECKER. 1992. Molecular structure of thearthropod hemocyanins. Adv. Comp. Environ. Physiol. 13:325–376.
MARKL, J., A. HOFER, G. BAUER, A. MARKL, B. KEMPTNER,M. BRENZINGER, and B. LINZEN. 1979a. Subunit hetero-geneity in arthropod hemocyanins: II. Crustacea. J. Comp.Physiol. 133:167–175.
MARKL, J., A. MARKL, W. SCHARTAU, and B. LINZEN. 1979b.Subunit heterogeneity in arthropod hemocyanins: I. Cheli-cerata. J. Comp. Physiol. 130:283–292.
MILLER, S. G., and D. L. SILHACEK. 1995. Riboflavin bindingproteins and flavin assimilation in insects. Comp. Biochem.Physiol. 110:446–475.
SAIKI, R. K., D. H. GELFAND, S. STOFFEL, S. J. SCHARF, R.HIGUCHI, G. T. HORN, K. B. MULLIS, and H. A. EHRLICH.1988. Primer-directed enzymatic amplification of DNAwith a thermostable DNA polymerase. Science 239:487–494.
SAITOU, N., and M. NEI. 1987. The neighbor-joining method:a new method for reconstructing phylogenetic trees. Mol.Biol. Evol. 4:406–425.
SANCHEZ, D., M. D. GANFORNINA, and M. J. BASTIANI. 1996.Developmental expression and biochemical analysis ofConulin, a protein secreted from a subset of nerve growthcones. J. Neurosci. 16:663–674.
SCHELLER, K., B. FISCHER, and H. SCHENKEL. 1990. Molec-ular properties, functions and developmentally regulatedbiosynthesis of arylphorin in Calliphora vicina. Pp. 155–162 in H. H. HAGEDORN, J. G. HILDEBRAND, M. G. KID-WELL, and J. H. LAW, eds. Molecular insect science. PlenumPress, New York.
SELLOS, D. Y., S. LEMOINE, and A. VAN WORMHOUDT. 1997.Molecular cloning of hemocyanin cDNA from Penaeusvannamei (Crustacea, Decapoda): structure, evolution andphysiological aspects. FEBS Lett. 407:153–158.
SUGUMARAN, M., L. B. GIGLIO, H. KUNDZICZ, S. J. SAUL, andV. SEMENSI. 1992. Studies on the enzymes involved in pu-parial cuticle sclerotization in Drosophila melanogaster.Arch. Insect Biochem. 19:271–283.
SWOFFORD, D. L. 1991. PAUP: phylogenetic analysis usingparsimony. Version 3.0s. Illinois Natural History Survey,Champaign.
TELFER, W. H., and J. G. KUNKEL. 1991. The function andevolution of insect storage hexamers. Annu. Rev. Entomol.36:205–228.
THIOULOUSE, J., and F. CHEVENET. 1996. NetMul, a World-Wide Web user interface for multivariate analysis software.Comp. Stat. Data Anal. 21:369–372.
THOMPSON, J. D., D. G. HIGGINS, and T. J. GIBSON. 1994.CLUSTAL W: improving the sensitivity of progressive mul-tiple sequence alignment through sequence weighting, po-sitions-specific gap penalties and weight matrix choice. Nu-cleic Acids Res. 22:4673–4680.
VAN HOLDE, K. E., and K. I. MILLER. 1995. Hemocyanins.Adv. Protein Chem. 47:1–81.
VOLBEDA, A., and W. G. J. HOL. 1989a. Crystal structure ofhexameric haemocyanin from Panulirus interruptus refinedat 3.2 A resolution. J. Mol. Biol. 209:249–279.
. 1989b. Pseudo 2-fold symmetry in the copper-bind-ing domain of arthropodan haemocyanins. J. Mol. Biol.206:531–546.
VON HEIJNE, G. 1990. The signal peptide. J. Membr. Biol. 115:195–201.
WANG, Z., and N. HAUNERLAND. 1994. Receptor-mediated en-docytosis of storage proteins by the fat body of Helicoverpazea. Cell Tissue Res. 278:15–26.
MANOLO GOUY, reviewing editor
Accepted December 30, 1997
Downloaded from https://academic.oup.com/mbe/article-abstract/15/4/415/1021391by gueston 04 April 2018





























