Embed Size (px)
Citation preview

Part IINTRODUCTION TO
CIRCADIAN RHYTHMS
RHY1 2/6/04 3:51 PM Page 1

� BASIC ATTRIBUTES OF CIRCADIANRHYTHMS
The word circadian derives from the Greekwords circa (about) and dian (day). Circa-dian rhythms then are cycles that occurwith a periodicity of ~24 hours. Given thatwe live in a cyclic environment created bythe rotation of the Earth, it is not surpris-ing that adaptation has involved the circa-dian cycling of physiological and behavioralactivities. What is amazing is the extent towhich circadian rhythms pervade life onEarth. Previously thought to be a charac-teristic of eukaryotic organisms, circadianrhythms are now known to occur inprokaryotes such as cyanobacteria. Indeed,as we shall see in Chapter 6, much of thecyanobacteria genome, and by inferencemuch of its physiology, is expressed in acyclic fashion. Clearly, almost all organismshave evolved ways of coordinating theirphysiology such that different functions
occur at different times of day. As we arebecoming more aware of temporal aspectsof regulation, we are realizing that there isscarcely a physiological process that doesnot involve some cycling component.
Example of Circadian Rhythm:Rest–Activity Cycle
Human sleep–wake cycles constituteperhaps the best known example of abehavior that occurs with a ~24-hour peri-odicity. Many other organisms displaysimilar behavior, which is commonlyreferred to as rest–activity cycles, takinginto account the fact that “rest” is notalways “sleep.” A record depicting rest–activity cycles in the fruitfly, Drosophilamelanogaster, is shown in Figure 1.1.Each horizontal line in this record represents a 24-hour day, and the verticallines correspond to the activity of a singlefly. The activity of this fly was monitored
1
GENERAL CONCEPTSAmita Sehgal
Molecular Biology of Circadian Rhythms. Edited by Amita SehgalISBN 0-471-41824-2 Copyright © 2004 John Wiley & Sons, Inc.
3
RHY1 2/6/04 3:51 PM Page 3
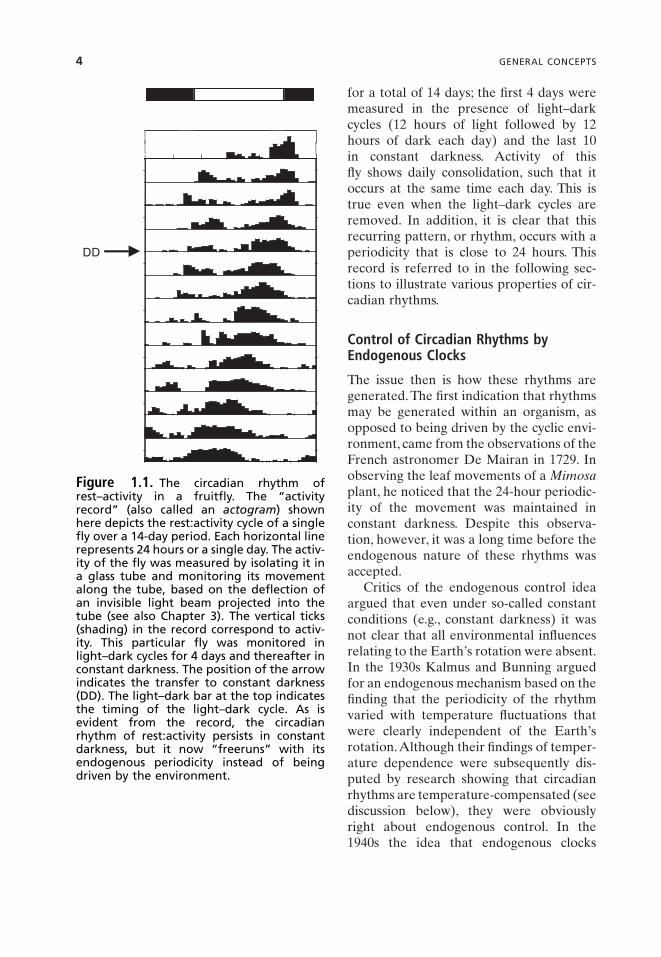
for a total of 14 days; the first 4 days weremeasured in the presence of light–darkcycles (12 hours of light followed by 12hours of dark each day) and the last 10 in constant darkness. Activity of this fly shows daily consolidation, such that itoccurs at the same time each day. This istrue even when the light–dark cycles areremoved. In addition, it is clear that thisrecurring pattern, or rhythm, occurs with aperiodicity that is close to 24 hours. Thisrecord is referred to in the following sec-tions to illustrate various properties of cir-cadian rhythms.
Control of Circadian Rhythms byEndogenous Clocks
The issue then is how these rhythms aregenerated. The first indication that rhythmsmay be generated within an organism, asopposed to being driven by the cyclic envi-ronment, came from the observations of theFrench astronomer De Mairan in 1729. Inobserving the leaf movements of a Mimosaplant, he noticed that the 24-hour periodic-ity of the movement was maintained in constant darkness. Despite this observa-tion, however, it was a long time before theendogenous nature of these rhythms wasaccepted.
Critics of the endogenous control ideaargued that even under so-called constantconditions (e.g., constant darkness) it wasnot clear that all environmental influencesrelating to the Earth’s rotation were absent.In the 1930s Kalmus and Bunning arguedfor an endogenous mechanism based on thefinding that the periodicity of the rhythmvaried with temperature fluctuations thatwere clearly independent of the Earth’srotation.Although their findings of temper-ature dependence were subsequently dis-puted by research showing that circadianrhythms are temperature-compensated (seediscussion below), they were obviously right about endogenous control. In the1940s the idea that endogenous clocks
4 GENERAL CONCEPTS
DD
Figure 1.1. The circadian rhythm ofrest–activity in a fruitfly. The “activityrecord” (also called an actogram) shownhere depicts the rest:activity cycle of a singlefly over a 14-day period. Each horizontal linerepresents 24 hours or a single day. The activ-ity of the fly was measured by isolating it ina glass tube and monitoring its movementalong the tube, based on the deflection ofan invisible light beam projected into thetube (see also Chapter 3). The vertical ticks(shading) in the record correspond to activ-ity. This particular fly was monitored inlight–dark cycles for 4 days and thereafter inconstant darkness. The position of the arrowindicates the transfer to constant darkness(DD). The light–dark bar at the top indicatesthe timing of the light–dark cycle. As isevident from the record, the circadianrhythm of rest:activity persists in constantdarkness, but it now “freeruns” with itsendogenous periodicity instead of beingdriven by the environment.
RHY1 2/6/04 3:51 PM Page 4

control circadian rhythms gained wideracceptance. What helped to reinforce thiswas the finding that the periodicity of circa-dian rhythms under noncycling environ-mental conditions varies from species tospecies. More importantly, the period inmost cases did not precisely match that ofany environmental cycle, indicating that itwas most likely endogenously generated.For instance, in the record shown in Figure1.2, note how, in constant darkness, activityeach day begins and ends a little earlier thanit did the day before. This drift occursbecause the periodicity of this rhythm, andthereby that of the underlying clock, is alittle less than 24 hours. In the presence oflight–dark cycles, the drift does not occurbecause, as discussed below, light resets theclock.
Despite the general acceptance of theidea, some skeptics remained, and thisskepticism inspired Hamner and others toconduct an interesting experiment in theearly 1960s.This experiment was performedat the South Pole with a number of organ-isms, all of which were maintained on arotating turntable. Although the turntablerotated with the same periodicity as theEarth, it did so in the opposite direction.Under these conditions, the circadianrhythm of eclosion in Drosophila pseud-oobscura persisted, as did a number ofother physiological rhythms.
It should be noted that not all rhythmicfunctions are controlled by an endogenousclock. Some are driven by the day–nightcycle and therefore do not persist in con-stant conditions. These are referred to asdiurnal rhythms, to distinguish them fromcircadian rhythms, which, by definition,persist in noncycling environmental con-ditions. In humans, rhythmic processesinclude sleep–wake cycles, release ofseveral hormones, regulation of body tem-perature, blood pressure, urine production,cholesterol metabolism, respiratory func-tion, and incidence of asthma and heartattacks. While some of these are clearlyunder circadian control, others may turnout to be dependent on other factors (e.g.,behavioral state). For instance, the releaseof some hormones appears to be rhythmicbecause it is influenced by sleep, which nor-mally occurs in a cyclic fashion, rather thanby clock function (see Chapter 12).
Circadian Rhythm Properties
This periodicity of ~24 hours is one attrib-ute of circadian rhythms. A second is thatthese rhythms can be synchronized or reset by environmental cues. The dominantenvironmental signal that affects rhythms is light, with most rhythms synchronized to the day–night cycle. Jurgen Aschoff used the term zeitgeber (“timegiver” in
BASIC ATTRIBUTES OF CIRCADIAN RHYTHMS 5
Environmentalsignal (e.g., light)
Input pathway Output pathway
Overt rhythm(e.g., sleep-wake cycle)
Figure 1.2. A simple circadian system. The very basic circadian system is thought to containa clock, an input pathway that transmits environmental signals to the clock, and an outputpathway that carries signals away from the clock and results in the manifestation of overtrhythms. In practice, there can be multiple input and output pathways to serve a single clock.In addition, the system most likely does not work in the linear fashion depicted here. Outputscan feedback on the clock and perhaps even on input and the clock can feedback on theinput pathway.
RHY1 2/6/04 3:51 PM Page 5

German) to denote a periodic environmen-tal signal that synchronizes or sets the time of a circadian rhythm. The process ofsynchronization itself is called entrain-ment. Since most rhythms are synchronizedto environmental cycles, chronobiologistsrefer to time as defined by these environ-mental cycles, thereby calling it zeitgebertime (ZT).When an organism is maintainedin a 24-hour cycle that consists of 12 hoursof light, followed by 12 hours of darkness,zeitgeber time or ZT0 corresponds to“lights on” and ZT12 is “lights off.” Thus,all timepoints between 0 and 12 refer todaytime hours, while those between 12 and24 (which is the same as “0”) refer to night-time hours. This terminology will be usedthroughout this book.
Light then is a major zeitgeber thatentrains circadian rhythms. The time atwhich a rhythmic activity/process occursdefines the phase of that particular rhythm.In the record shown in Figure 1.1, the timeat which the fly is active indicates the phaseof its rest–activity rhythm. Likewise, thetime at which it sleeps, or rests, can also beused as a measure of phase. Typically, theparameter that is most constant from day to day (whether wakeup time or sleep time)is used to define the phase of a rhythm. Itis clear that nocturnal and diurnal animalsdisplay opposite phases in their sleep–wakerhythms. A change in the timing of the zeitgeber, as might be caused by travel to adifferent time zone, results in a shift inthe phase of the rhythm. However, as we allwell know, this shift does not occur imme-diately, thereby resulting in the phenome-non commonly known as “jet lag.” Duringthe process of adjustment to a new cycle arhythm displays “transient” behavior, con-sisting of erratic phase and periodicity.
To reiterate an important property ofcircadian rhythms, they can persist in the absence of cyclic environmental cues.Under these conditions they are said to be freerunning. The endogenous nature of a rhythm and its periodicity have to be
determined under freerunning conditions.Again, note how in Figure 1.1, the patternof the rhythm in constant darkness(freerun) is somewhat different from that in the presence of light–dark cycles. This isbecause in constant darkness it runs with itsendogenous periodicity. Under these condi-tions, we speak in terms of circadian time(CT), which is the time of the internalclock. Since the periodicity of the internalclock is frequently a little different from 24hours, CT is not the same as ZT. However,on the first and second days of constantdarkness, when the endogenous rhythm hasnot drifted significantly from the environ-mental cycle, CT may approximate ZT foran organism that has a periodicity close to24 hours, and researchers may use it assuch.
Finally, the periods of circadian rhythmsare temperature compensated. This wasdemonstrated in some classic experimentsdone by Colin Pittendrigh in the 1950s. Hedetermined the effects of temperature onthe eclosion (hatching of adult flies frompupae) rhythm in Drosophila pseudoob-scura flies. In his first series of experimentshe improvised “incubators”—a pressurecooker maintained in a cold-water creekand an outhouse darkroom, to maintain the flies at cold and warm temperatures re-spectively. He found that the periodicity ofthe rhythm was relatively the same (withinan hour) at the two temperatures. In subsequent experiments done at PrincetonUniversity, he confirmed these findings,showing that the periodicity was essentiallythe same at 26 and 16°C.
Thus, the circadian period remains constant over a wide temperature range.Assuming that the underlying basis of overtrhythms is some biochemical mechanism,as indeed must be the case, then normallythe changes in protein function at differenttemperatures would alter the rate of thereaction and thereby the periodicity of therhythm. The rate of most physiologicalprocesses doubles with a 10°C rise in
6 GENERAL CONCEPTS
RHY1 2/6/04 3:51 PM Page 6

temperature. The fact that this does nothappen to circadian periodicity indicatesthat there is some mechanism that com-pensates for these temperature-inducedchanges. Clearly, it is important for organ-isms to maintain a periodicity that approx-imates that of the day–night cycle despitechanges in environmental temperature.While homeotherms can obviously do thiswith no problem, organisms that do notmaintain a constant body temperature hadto evolve mechanisms for this purpose.
Circadian Clock Input/Output
Circadian clocks are part of a system thatincludes input and output components (Fig. 2.2). If rhythms are generated by anendogenous clock—and, indeed, at thispoint we can say that they are, based notonly upon the investigations mentionedabove, but also upon a vast amount ofgenetic and molecular data that are thesubject of this book—then, if follows thatthe properties of rhythms arise from in-herent characteristics of the clocks thatproduce them. In fact, clocks are entrainedby external/environmental signals throughwhat is termed an input or entrainmentpathway. The implication is that the clockitself may not be in contact with the envi-ronment, but can communicate with itthrough a distinct pathway(s). Since thepredominant entraining cue is light, theinput pathway is usually perceived as con-taining a photoreceptor. As we shall see insubsequent chapters, the identification ofthe photic input pathway is an intense areaof research in every organism used for thestudy of circadian biology.
Time-of-day signals generated by theclock must be transmitted to other parts ofthe organism to drive physiology/behaviorin a rhythmic fashion. The pathway thatcarries signals away from the clock and produces overt rhythms is called the output pathway. Since outputs can varygreatly, even within the same organism, it
follows that there must be more than oneoutput pathway. In fact, we now know (as described below) that many of theseoutputs are actually generated not just byindependent output pathways, but by sepa-rate clocks within an organism.
Together, the clock, input pathway andoutput pathway are termed a circadiansystem. Although the components of thissystem are typically depicted in a lineardiagram, this is an oversimplification of themechanisms involved. In many cases, theoutputs can feed back and affect the clock(e.g., forced activity can reset the clock),and the clock may feedback to regulateinput, resulting in the response to lightbeing stronger at certain times of day. Theresponse to light is said to be “gated” by theclock.
Clock Entrainment to Input Components (Light)
As mentioned above, most biological clocks have periodicities that are not pre-cisely 24 hours. Aschoff suggested that theperiodicity of the same organism may bedifferent under different conditions,depending on the functional state of theorganism or even the nature of the “con-stant” environment (e.g., constant light vs.constant dark). He was intrigued by the factthat light-active (diurnal) animals show anincrease in spontaneous frequency, as meas-ured by increased bouts of activity, withincreasing intensity of light. This increasedfrequency of the oscillation results in anoverall shorter period. Night-active (noc-turnal) animals, on the other hand, decreasetheir spontaneous frequency with increas-ing intensity of light, thereby displayinglonger periods. On the basis of these obser-vations, Aschoff proposed a circadian rule according to which diurnal animalshave shorter than 24-hour rhythms in LL(constant light) and longer than 24-hourrhythms in DD (constant darkness). Thereverse would be true in nocturnal animals.
BASIC ATTRIBUTES OF CIRCADIAN RHYTHMS 7
RHY1 2/6/04 3:51 PM Page 7

However, as Aschoff himself recognized,there are exceptions to this rule. Drosophilaare diurnal and yet display periodicitiesthat are slightly less than 24 hours (~23.8hours) in DD.
Regardless of the periodicity of theendogenous rhythm all organisms are synchronized to the 24-hour environmentalcycle. This means that they must be resetevery day because otherwise they wouldeventually drift out of synchrony with theday–night cycle.Take, for instance, a rhythmthat has a period of 23.8 hours. If it were to freerun, then on each successive day itwould be an additional 0.2 hour out ofphase with the environment. In 2 months itwould have a reverse phase, relative to theenvironment, from the one with which itstarted. A rhythm such as this, which has a shorter period than the environmentalcycle, is delayed by 0.2 hour each day. Thisis illustrated by the example shown inFigure 1.1, where the endogenous periodic-ity of the fly is ~23.5 hours, but it synchro-nizes to a 12–12 light–dark cycle, because itis reset each day. A longer period rhythmmust advance each day by the amount oftime its periodicity differs from that of the day–night cycle. The manner of theresetting can differ between rhythms. Arest–activity rhythm could advance eachday by initiating either the rest or the activephase earlier than dictated by the internalclock. These earlier onsets are typicallydriven by something in the environment(e.g., sunrise or sunset).
As will be seen in subsequent chapters,even organisms that lack clock function,through genetic manipulation, are driven bythe environmental cycle to display 24-hourrhythms. The difference between theserhythms and those in clock-containingorganisms is that they cannot persist whenthe rhythmic signals from the environmentare removed. In addition, they cannot antic-ipate the daily transitions in the environ-mental cycles. For instance, in a diurnalanimal that lacks a clock, activity is usually
triggered by the light signal.The animal doesnot become active in anticipation of thedark–light transition. In a nocturnal animallight would have the opposite effect. Thiseffect of light that is independent of theendogenous clock is termed “masking,” socalled because it can mask the manifestationof the endogenous rhythm. Thus, even in ananimal that has a clock, masking effects oflight can confound analysis of the endoge-nous rhythms, which are therefore bestassayed under freerunning conditions (i.e.,constant darkness and constant tempera-ture). It should be noted that althoughmasking, as described here, denotes clock-independent effects of light on the activityrhythm, it can, in principle, apply to anyinfluence that obscures a rhythmic function.
Entrainment to Light Pulses
In addition to being entrained to day–night cycles, circadian rhythms can be en-trained by pulses of light. This type ofentrainment is called nonparametric, distin-guishing it from parametric entrainmentwhich involves synchronizing to cycles of astimulus. In general, a pulse of light deliv-ered when an organism expects to see light(“subjective day”, as it is called in constantdarkness) does not reset the rhythm.However, pulses at night reset the rhythmwith those in the first part of the nighthaving effects opposite those in the secondhalf of the night. A pulse in the early nightdelays the phase of the rhythm. It can bethought of as analogous to extended day-light, which has the effect of delaying theonset of the night function (e.g., in a diurnalanimal it might delay the onset of sleep orrest). Conversely, a pulse in the second halfof the night advances the phase of therhythm. Again, the best analogy perhaps iswith a shift in the natural day–night cycle.In this case, it would be like dawn occurringearlier, which would lead to earlier onset ofthe next day’s activities. The idea is that apulse in the early night resets the cycle to
8 GENERAL CONCEPTS
RHY1 2/6/04 3:51 PM Page 8

dusk, while one in the late night resets it todawn. In practice, of course, the resetting isnot quite so perfect. Delays are usually oflarger magnitude and easier to achieve thanadvances.
A graph that plots the magnitude of theshift relative to the time of day that theshifting stimulus is delivered is called aphase-response curve (PRC). A typical suchcurve is shown in Figure 1.3a.As mentionedabove, there is no shift during the subjec-tive day, which is thereby called the “deadzone.” In the middle of the night, the curvecrosses over from delays to advances. In this type of PRC, also called a “Type 1PRC,” pulses of light at the crossover pointdo not produce any shift in the rhythm.
Some organisms are more sensitive to lightand display a Type 0 PRC (Fig. 1.3b). AType 0 PRC is characterized by larger mag-nitude shifts and the absence of anycrossover point in the middle of the night.This lack of a crossover typically gives afalse impression of discontinuity in thePRC; in a rhythm with a 24-hour period, a12-hour delay is indistinguishable from a12-hour advance, so a shift from plottingstrong delays to weaker advances producesthe apparent discontinuity in the plot.Although previously regarded as very dis-tinct entities, there is now increasing evi-dence that with strong light pulses (eitherof higher intensity or increased duration) aType I PRC can be turned into a Type 0.
BASIC ATTRIBUTES OF CIRCADIAN RHYTHMS 9
12 12 2424Circadian time
2
4
0
-2
-4
Shi
ft (in
hou
rs)
(a) (b)
Figure 1.3. Entrainment to pulses of light. Based on the response of an organism to reset-ting stimuli delivered at different times of day, phase response curves (PRCs) can be con-structed. This type of curve basically demonstrates the effect of time of day on the magnitudeof the phase shift produced by a specific stimulus. The best documented PRCs are those inresponse to light stimuli and these are of two major kinds, shown in (a) and (b). The light–darkcycle used to entrain the organism is indicated at the bottom of each plot. The number ofhours by which the phase of the clock shifts (y axis) in response to a light pulse delivered atdifferent times of day (x axis) is plotted. In a Type 1 PRC (a), pulses of light delivered duringa time when an organism would expect to see light (daytime hours) have no effect on thephase of the rhythm. A pulse of light in the early part of the night delays the phase (indi-cated by negative numbers), and a pulse in the second half of the night advances the rhythm(indicated by positive numbers), with a gradual transition between phase delays and phaseadvances. In a Type 0 PRC, the clock is very sensitive to light and shows large shifts, in responseto pulses, at all times of the cycle, with no gradual change from phase delays to phaseadvances.
RHY1 2/6/04 3:51 PM Page 9

Multiple Clocks
An organism can have more than one cir-cadian clock. It has now become clear thatmost multicellular organisms have morethan one clock. It appears that with increas-ing complexity of physiological function,there is increasing need to have clocks specific for different organs/tissue systems.The question then becomes: Is there oneclock that controls all others in an organism—a master clock, if you will—or are allclocks equal? Among the animal systems,
the master or central clock (if there is one)is usually located in the brain (Fig. 1.4).Clocks in other parts of the body arereferred to as peripheral oscillators to dis-tinguish them from the central clock. Thedegree of autonomy that these oscillatorspossess varies from species to species andalso from oscillator to oscillator. Insectscontain many clocks that are autonomouswith respect to timekeeping as well as withrespect to photoreception. However, thereis at least one oscillator in Drosophila,the insect of choice for molecular analysis
10 GENERAL CONCEPTS
Light
Light
Central clock(brain)
Central clock(brain)
Peripheralclock
Peripheralclock
Peripheralclock
Peripheralclock
Peripheralclock
Peripheralclock
Output
Output
Output
Output
Output
Output
Figure 1.4. Organization of clocks in complex organisms. In mammals, a light-responsivecentral clock in the brain regulates clock activity in clocks found in peripheral tissues (top).The peripheral clocks are not directly photosensitive although they can entrained by othersignals (e.g., hormones). In other organisms, peripheral clocks can be photosensitive and varyin their dependence upon the central clock.
RHY1 2/6/04 3:51 PM Page 10

of clock function, that is controlled by the central clock although it may also becapable of direct photoreception. Likewise,most oscillators in the nonmammalian vertebrate, zebrafish, are autonomous and have their own photoreceptors,although interactions with the central clock likely occur. Other nonmamalian vertebrates, that do not have the transpar-ent skin of the zebrafish, may not show this extent of autonomy in peripheral oscillators.
In mammals, there is a master clock thatis housed in the hypothalamus and is calledthe suprachiasmatic nucleus (SCN). Thus,clock functions appear to have become cen-tralized over the course of evolution ashave other body functions (Fig. 1.4). TheSCN can sustain clock activity for extendedperiods of time in culture. It is also the onlyclock in the body, with the exception of the eye clock, which receives photic infor-mation. Details of this can be found inChapter 4.
Although peripheral oscillators inmammals cannot perceive light, they can be entrained by other physiological stimuli.Most notable are hormones, which might be controlled by the SCN, but may also actindependently on peripheral oscillators. Inaddition, retinoids will reset a clock in thevasculature, and restricted feeding canentrain the clock in the liver without affect-ing the SCN. Clearly, the organization ofcircadian systems in terms of central andperipheral oscillators, the interactionsbetween the different components and themechanisms that entrain them are allimportant aspects of physiology. Theseissues are discussed in the chapters onperipheral oscillators and hormonalrhythms. Specific details that pertain toindividual organisms can be found in thechapters on these respective organisms.
� CLOCK MECHANISMS
Since the general acceptance of the ideathat clocks are endogenous, a major ques-tion has been: How, within an organism,do you make a clock? Over the years, anumber of mathematical models have beendeveloped to explain how this might occur.One aspect that these models have incommon is the presence of a feedback loop.Overt cycles must have their basis in anendogenous cycle or loop that can sustainoscillations indefinitely in the absence ofenvironmental cues. The question then ishow these loops are generated. Do theyoperate at the level of the organism, thetissue, or the cell? As outlined below anddescribed in detail in other chapters in thisbook, in all organisms examined, the basicclock mechanism (i.e., the oscillatory loop)is, in fact, intracellular. While an intracellu-lar clock might be expected of a cyanobac-terium or of the bread mold Neurospora, itcomes as a surprise when it turns out to bethe case in a mammal.This is not to say thattimekeeping is performed by a single cell.In multicellular organisms, a clock tissue(central or peripheral) is likely to be com-posed of a cluster of cells, each of whichcontains a loop that oscillates in synchronywith the loops in neighboring cells.An SCNis an example of such a cluster. Rhythmicsignals generated by these clusters driveoutput rhythms.
As one might have gathered from thediscussion above, the general nature of the clock mechanism is conserved fromcyanobacteria to mammals. However,since circadian rhythms are present inwidely separated phyla and the actual molecules involved are different in thediverse phyla, the prevailing notion is thatclocks evolved several times. The fact thatthe same mechanism evolved in each caseattests to the efficacy of this particularmechanism. In addition, the availability ofmultiple, biological systems that use thesame mechanism has allowed a powerful,
CLOCK MECHANISMS 11
RHY1 2/6/04 3:51 PM Page 11

multidisciplinary approach to the study ofclocks. Particularly important have beenstudies in model organisms, which offerpowerful genetic approaches and simplersystems than more complex vertebrates.A general model for the clock mechanismwas first worked out in Drosophila and then in Neurospora. Mammalian studies ofclock function built largely on the infor-mation that was already available from these other systems, in particular on theframework provided by the Drosophilamodel. This book highlights this multiple-species approach to the molecular analysisof rhythms by describing the salient fea-tures of the clock, input and output path-ways in different species and comparing,wherever applicable, the similarities in thesystems.
Clock Mechanism at a Molecular Level
The basic clock mechanism is a feedbackloop in which oscillating products of spe-
cific genes regulate their own expression.These genes are called clock genes, and theautoregulation consists of the proteins negatively regulating synthesis of their ownmRNAs (Fig. 1.5). Each complete turn ofthe loop takes ~24 hours to complete,resulting in circadian oscillations of RNAand protein levels. Rhythmic expression of the proteins ensures that the negativefeedback on transcription is rhythmic,which, in turn, ensures rhythmic expressionof the mRNA. The precise molecules thatfunction in this fashion may differ from one species to another although certainstructural features are conserved fromcyanobacteria to mammals. Between Dro-sophila and mammals, the molecules arealso conserved.
Given that this is a cyclic process, it is,of course, difficult to ascertain what comesfirst—the RNA cycling or the proteincycling. It is clear that the mRNAs do notcycle in the absence of rhythmic feedbackfrom the proteins. However, the most
12 GENERAL CONCEPTS
Gene X
RNA
Protein
Negativefeedback
Figure 1.5. The basic model for a 24-hour oscillator. The model is based on work done in many different species ranging from cyanobacteria to mammals and consists of an autor-egulatory feedback loop in which a clock gene encodes cycling RNA and protein, and theprotein negatively regulates synthesis of its own mRNA. The loop takes ~24 hours to com-plete (shown within a 24-hour clock).
RHY1 2/6/04 3:51 PM Page 12

recent data indicate that cycling of specificclock proteins can be maintained underconditions where their RNAs do not cycle.Posttranslational mechanisms, such as cyclicphosphorylation, appear to contribute torhythmic expression of clock proteins andmay account for oscillations of these pro-teins under conditions where cycling of thecorresponding mRNAs is blocked. It isimportant to note also that these experi-ments have not eliminated the cycling of allclock gene mRNAs in an organism, so thecycling of one may be able to compensatefor loss of cycling of the others.
Although we have refrained from theuse of chronobiology jargon in this book, itis worth introducing some of the terminol-ogy used in the field here because it mayappear in other readings of the circadian lit-erature. A molecule that is part of the clockis frequently referred to as a clock compo-nent. A clock component can be a statevariable, which is a molecule that impartstime-of-day cues through rhythmic changesin its levels or activity, or a state parameter,which does not itself cycle, but is requiredfor the appropriate regulation of the statevariable. Researchers have developed crite-ria for state variables, which include arequirement that its levels or activity cycle,that it regulate itself through a feedbackmechanism and that mutations in itproduce effects on overt rhythms.
Gene Expression
We are starting to also gain an understand-ing of how it is that clocks communicatetheir temporal signals to the rest of theorganism, although these “output pathway”mechanisms represent the least understoodaspect of circadian rhythms. The diversity of overt rhythms, and thereby that of theoutput pathways that lead to these rhythms,may account, in part, for the dearth ofknowledge in this area. Clearly, the mole-cules and mechanisms that produce rhyth-mic rest–activity will be very different from
those that drive a rhythm in cholesterolmetabolism in the liver.
What is clear, however, is that one of theways clock genes transmit output signals is by driving rhythms of gene expression.Although the number varies from speciesto species, we now know that a significantproportion of the genome is expressedrhythmically in all organisms that possessclocks. The number as well as the nature ofthe genes expressed in a rhythmic fashionvary not only from species to species butalso from one organ or tissue to another in the same organism. A gene expressedrhythmically in one tissue may not beexpressed rhythmically in another tissue,because it serves a different function in theother tissue or because the process withwhich it is involved does not cycle in the other tissue. The rhythms in geneexpression are translated into rhythms inprotein expression and function that ulti-mately give rise to rhythms in physiologyand behavior.
The most direct mechanism by which theclock can drive a rhythm in gene expressionis by rhythmically regulating promoter activ-ity. Promoters of many genes contain ele-ments recognized by transcription factorsthat are part of the clock. Rhythmic activityof these factors results in rhythmic expres-sion not only of clock genes but also of genesthat are downstream of the clock (Fig. 1.6).These downstream genes may encode othertranscription factors or other proteins thataffect a cell property or function.
However, it is important to note thatclocks can also regulate downstream com-ponents through transcription-independentmechanisms. Although the mechanisms are not known yet, there are examples ofcircadian components that cycle at theprotein, but not the RNA level. In addition,in Drosophila, a neuropeptide that isreleased specifically by brain clock cellscycles at axon terminals. Since expressionof this protein does not cycle in the cellbody, the interpretation is that the levels do
CLOCK MECHANISMS 13
RHY1 2/6/04 3:51 PM Page 13

not cycle, but the release from axon termi-nals does. A simplistic model for circadiancontrol of a complex function, such asrest–activity behavior, is that oscillations inclock genes result in oscillations in somecellular (neuronal) property. This oscilla-tion in turn drives rhythmic expression ofan output (perhaps a hormone) from thecell, and that output mechanism then actsrhythmically on cells controlling rest–activ-ity (Fig. 1.7). This results in the manifesta-tion of rest–activity rhythms. In a simplecircadian system the final output rhythmmay be produced directly by the clock cell.
Thus the process is more complicatedthan would appear at first glance. Thesecomplexities, most of which are not com-pletely understood yet, are acknowledgedhere and elsewhere in this book. However,at the same time, it is clear that we havelearned a lot about how clocks operate inmany different species. Amazingly, most ofthis progress has occurred very recently (asof 2003). In the late 1990s a book focusedon molecular mechanisms underlying clockfunction might have seemed premature.Now it is perceived as something that isneeded. It is important to note, though, thatthis is still a rapidly moving and growing
field and so the material presented hereprovides the basis, but by no means thecomplete picture, of molecular clock function.
� ADAPTIVE SIGNIFICANCE OFCIRCADIAN RHYTHMS
An introduction to circadian rhythmswould, of course, be incomplete withoutsome discussion of their adaptive signifi-cance. Several possible explanations havebeen proposed. In 1987 Rapp argued thatbiological systems are periodic becausetemporal organization allows for bettersynchrony or coordination between differ-ent processes in the same organism, itallows incompatible processes to be sepa-rated from each other in time (this is particularly advantageous in unicellularorganisms; see Chapter 6) and it allows fora more precise, efficient system.
Perhaps it is easy to see how adaptationto a cyclic environment would be facilitatedby cyclic physiology or behavior. However,most rhythms can be driven by the envi-ronmental cycle. As mentioned above, evenorganisms that lack clocks, or contain
14 GENERAL CONCEPTS
Clock gene Clock-controlledgene
Clock-controlledgene
Transcription factors, hormones,ion channels
Figure 1.6. Control of downstream genes by the clock. A molecular clock (such as the oneshown in Fig. 1.7) can regulate the output pathway by driving cyclic expression of down-stream genes. One mechanism by which this occurs is shown here. While regulating its owntranscription, the clock protein also regulates the transcription of downstream genes at a spe-cific time of day. This leads to rhythmic expression of those genes.
RHY1 2/6/04 3:51 PM Page 14

mutant clocks with aberrant periodicity, canbe driven to display rhythmic activity in a12-hour light–12-hour dark cycle. Giventhis, why do we then need endogenousclocks? The most commonly cited explana-tion for endogenous rhythmicity is that it enables organisms to anticipate dailytransitions in the environment. It is statedabove that changes in the activity patternsof clockless animals are triggered by theenvironmental stimulus (say, light). Ananimal with a clock becomes active in antic-ipation of the dark–light transition, whichmay confer a selective advantage on it. It isa case of the early bird getting the worm.In addition, a clock allows finer temporalcontrol over physiological processes.
The advent of cellular and genetic toolsallowed researchers to experimentally testthe importance of clocks. The basic experi-ment is to remove the clock and assay sur-vival or fitness. Cellularly, this has beendone by lesioning the SCN. Although wenow know that clocks are found in manytissues outside the SCN, the master clock inmammals is in the SCN. In addition,genetic tools have allowed the generation
of animals that lack one or more clockgenes in all tissues and so are arrhythmic.Interestingly, early attempts to identify via-bility or fertility deficits in these animalsfailed. The advantage conferred by clocksmay not be evident under laboratory con-ditions where the animals are supplied withample food and water and do not have tofend for themselves. However, moredetailed analysis is now revealing deficitsassociated with loss of clock function. Fur-thermore, researchers found a small reduc-tion in lifespan in some of the Drosophilamutants that affect circadian period.The differences in lifespan between wildtype and mutants were altered somewhat in light–dark cycles of different lengths.However, the reduction in lifespan wasseen even when the mutants were main-tained in a light–dark cycle that approxi-mated the periodicity of their internalclock. Thus, the advantage of having a ~24-hour rhythm in Drosophila may extendbeyond the need to match the periodicity ofthe environmental cycle. In cyanobacteria,synchrony with the environmental cycleconfers a selective advantage. When cyan-
ADAPTIVE SIGNIFICANCE OF CIRCADIAN RHYTHMS 15
Cyclic molecule Cyclic membranepotential
Cyclic hormone release
Cyclic behavior
Rhythmic rest-activity
Molecular Cellular Physiological Behavioral
Figure 1.7. Control of an overt, behavioral rhythm by the clock. A simple model is shown inwhich the clock drives rhythmic expression of a gene that regulates membrane potential.Cycling membrane potential results in rhythmic release of a secreted factor that has an effecton levels of rest and activity.
RHY1 2/6/04 3:51 PM Page 15

obacterial strains of different periodicitiesare grown in the same culture, the ones thatgrow best, and thereby outcompete theirneighbors, have periodicities that matchthat of the environment. Finally, studies ofSCN-lesioned mammals released into thewild have also revealed advantages ofhaving clocks. The lack of a clock appearsto make these animals more susceptible topredators.
FURTHER READING
Aschoff, J (1960): Exogenous and endogenouscomponents in circadian rhythms. Cold SprHarb SympQuant Biol 25: 11–28.
Bunning, E (1973): The Physiological Clock. Circadian Rhythms and BiologicalChronometry,The English Universities Press.
DeCoursey, PJ, Walker, JK, Smith, SA (2000):A circadian pacemaker in free-living
chipmunks: Essential for survival? J CompPhysiol [A] 186: 169–180.
Dunlap, JC, Loros, JJ, DeCoursey, P (eds) (2003): Chronobiology: Biological Timekeep-ing, Sinauer Assoc.
Klarsfeld, A, Rouyer, F (1998): Effect of circa-dian rhythms and LD periodicity on the lifespan of Drosophila melanogaster. J BiolRhythms 13: 471–478.
Mori, T, Johnson, CH (2001): Circadian pro-gramming in cyanobacteria. Semin Cell DevBiol 12: 271–280.
Pittendrigh, CS (1954): On temperature inde-pendence in the clock system controllingemergence time in Drosophila. Proc NatlAcad Sci (USA) 40: 1018–1029.
Pittendrigh, CS (1993): Temporal organization:Reflections of a Darwinian clock-watcher.Ann Rev Physiol 55: 17–54.
Rapp, PE (1987): Why are so many biologicalsystems periodic? Prog Neurobiol 29: 261–273.
Saunders, DS (1982): Insect Clocks, PergamonPress, Oxford (Oxfordshire); New York.
16 GENERAL CONCEPTS
RHY1 2/6/04 3:51 PM Page 16





























