Embed Size (px)
Citation preview

ARTICLE
Mineralization of Pentachlorophenol WithEnhanced Degradation and Power Generation FromAir Cathode Microbial Fuel Cells
Liping Huang,1 Linlin Gan,1 Ning Wang,1 Xie Quan,1 Bruce E. Logan,2 Guohua Chen3
1Key Laboratory of Industrial Ecology and Environmental Engineering, Ministry of Education
(MOE), School of Environmental Science and Technology, Dalian University of Technology,
Dalian 116024, China; telephone: 86 411 84708546; fax: 86 411 84708546 (L. Huang),
telephone: 86 411 84706140; fax: 86 411 84706140 (X. Quan);
e-mail: [email protected], [email protected] of Civil and Environmental Engineering, The Pennsylvania State University,
University Park, Pennsylvania, 168023Department of Chemical and Biomolecular Engineering, Kowloon, Hong Kong University of
Science and Technology, Hong Kong, China
ABSTRACT: The combined anaerobic–aerobic conditions inair-cathode single-chamber MFCs were used to completelymineralize pentachlorophenol (PCP; 5mg/L), in the pres-ence of acetate or glucose. Degradation rates of 0.140�0.011mg/L-h (acetate) and 0.117� 0.009mg/L-h (glucose)were obtained with maximum power densities of 7.7�1.1W/m3 (264� 39W/m2, acetate) and 5.1� 0.1W/m3
(175� 5W/m2, glucose). At a higher PCP concentrationof 15mg/L, PCP degradation rates increased to0.171� 0.01mg/L-h (acetate) and 0.159� 0.011mg/L-h(glucose). However, power was inversely proportional toinitial PCP concentration, with decreases of 0.255W/mgPCP (acetate) and 0.184W/mg PCP (glucose). High pH(9.0, acetate; 8.0, glucose) was beneficial to exoelectrogenicactivities and power generation, whereas an acidic pH¼ 5.0decreased power but increased PCP degradation rates(0.195� 0.002mg/L-h, acetate; 0.173� 0.005mg/L-h, glu-cose). Increasing temperature from 22 to 358C enhancedpower production by 37% (glucose) to 70% (acetate), andPCP degradation rates (0.188� 0.01mg/L-h, acetate;0.172� 0.009mg/L-h, glucose). Dominant exoelectrogensof Pseudomonas (acetate) and Klebsiella (glucose) wereidentified in the biofilms. These results demonstrate thatPCP degradation using air-cathode single-chamber MFCsmay be a promising process for remediation of water
contaminated with PCP as well as for power generation.
Biotechnol. Bioeng. 2012;109: 2211–2221.
� 2012 Wiley Periodicals, Inc.
KEYWORDS: microbial fuel cell; PCP degradation rate;power production; mineralization
Introduction
Pentachlorophenol (PCP) is one of many recalcitrant andtoxic compounds found in water that are used for variouspurposes, such as herbicides, insecticides, fungicides, woodpreservatives, resins, and lubricants. The existence of thesechemicals in groundwater, industrial wastewater effluents,sediments, and surface soils poses great challenges fortreatment and remediation (Field and Sierra-Alvarez, 2008).Microbial co-metabolism is considered to be an effectiveroute of PCP degradation, in which PCP is used as a secondsubstrate and may serve as a carbon source for the growthof microorganisms (Field and Sierra-Alvarez, 2008; Shenet al., 2005). As the degradation of PCP occurs throughco-metabolism, the type of carbon source can affect theeffectiveness of this approach. Different carbon sources havebeen used in conventional biological processes for PCPdegradation, including lactate (Yang et al., 2005), glucose(Banerji and Bajpai, 1994; Shen et al., 2005; Visvanathanet al., 2005), acetate (Stuart and Woods, 1998), sucrose(Shen et al., 2005), mixtures of glucose, acetic acid, formicacid, and yeast extract (Damianovic et al., 2009), as well as acombination of peptone, sucrose and meat extract (Ye et al.,2004). It has been found that creating combined anaerobic–aerobic conditions is a particularly efficient strategy for PCP
Correspondence to: L. Huang and X. Quan
Contract grant sponsor: National Basic Research Program of China
Contract grant number: 2011CB936002
Contract grant sponsor: Natural Science Foundation of China
Contract grant number: 21077017; 51178077
Additional supporting information may be found in the online version of this article.
Received: 10 November 2011; Revision received: 3 February 2012;
Accepted: 22 February 2012
Accepted manuscript online 5 March 2012;
Article first published online 22 March 2012 in Wiley Online Library
(http://onlinelibrary.wiley.com/doi/10.1002/bit.24489/abstract)
DOI 10.1002/bit.24489
� 2012 Wiley Periodicals, Inc. Biotechnology and Bioengineering, Vol. 109, No. 9, September, 2012 2211

mineralization (Chen et al., 2010; Field and Sierra-Alvarez,2008). However, there are additional remaining challengesfor using this approach, such as increasing PCP degradationrates, reducing sludge generation, and minimizing energydemands of treatment processes.
One new promising method for more efficient and cost-effective PCP degradation is the use of microbial fuel cells(MFCs) (Huang et al., 2011a,b). An MFC is a device thatuses microbes to convert the chemical energy stored inorganic and inorganic compounds into electricity (Loganand Regan, 2006; Logan, 2009; Pant et al., 2010). It has beenshown that PCP can be degraded in the anaerobic bioanodeof a two-chamber MFC (Huang et al., 2011c), although thePCP degradation rate was slow (0.061�0.120mg/L-h) andpower production was relatively low (1.3�2.0W/m3). Inaddition, there was incomplete de-chlorination of PCP andthe accumulation of undesirable degradation products suchas 2,3,4,5-tetrachlorophenol and tetrachlorohydroquinone(Huang et al., 2011c). Themain disadvantage of using a two-chamber MFC for PCP degradation is that the anodechamber is anaerobic, and thus the benefit of a combinedanaerobic–aerobic condition for PCP degradation is notproduced. In addition, it is well known that powerproduction in two-chamber MFCs is low due to its highinternal resistance, and that the water in the cathode must beaerated, which is an energy-demanding process (Logan,2010; Ren et al., 2007).
Single-chamber, air-cathode MFCs can be used to avoidthe need to aerate water, and generally have lower internalresistances and higher power densities than two-chamberMFCs (Liu et al., 2005a; Logan et al., 2007; Logan, 2009,2010). One of the disadvantages of an air-cathode MFCfor power generation is oxygen crossover from the cathodeinto the liquid anode chamber due to diffusion of oxygenthrough the cathode. Oxygen crossover leads to growth ofaerobic bacteria on the cathode, which can lead to a decreasein power production (Kiely et al., 2011; Watson et al., 2011).The use of diffusion layers, such as carbon/polytetrafluor-oethylene on the air-side of the cathode, or a separatorbetween the anode and cathode, can reduce oxygen diffusioninto the anode chamber and improve power production(Cheng et al., 2006; Watson et al., 2011; Zhang et al., 2011b).In the case of PCP degradation, however, the micro-aerobicenvironment near the cathode may provide improvedconditions for PCP mineralization due to the combinationof anaerobic and aerobic conditions in the same chamber(Chen et al., 2010; Field and Sierra-Alvarez, 2008).
In this study, PCP degradation in single-chamber, air-cathode MFCs was investigated in the presence of eithera nonfermentable (acetate) or fermentable substrate(glucose). The degradation of PCP in the anode chamberwas examined as a function of the number of diffusion layers(in order to alter oxygen transfer rates into the anodechamber), initial COD, initial PCP concentration, pH,and temperature. The performance of the system wasevaluated in terms of PCP degradation rate, powerproduction, and recovery of electrons in the substrates in
terms of coulomic efficiency (CE). The microbial commu-nity on the anode was also analyzed.
Materials and Methods
Fuel Cell Assembly
Single-chamber air-cathode MFCs were similar to thoseused by Cheng et al. (2006). Graphite felt (2.0 cm�2.0 cm� 3.0 cm) (Sanye Co., Beijing, China) was used as theanode instead of carbon paper to increase available anodesurface area. A graphite rod was inserted into the graphitefelt anode to conduct electrons to the external circuit.Cathodes were made by applying platinum (0.5mg Pt/cm2)to the water side of the cathode, and diffusion layers to theair side of 30wt% wet-proofed carbon cloth (type B-1B,E-TEK) as described by Cheng et al. (2006). The number ofcathode diffusion layers was varied (1, 3, 5, or 7) in order tovary oxygen crossover and examine the effect of this on PCPdegradation rate and power generation. Diffusion layerswere made by brushing a polytetrafluoroethylene (PTFE)solution (60wt%) onto one side of the cathode, followed bydrying at room temperature and heating at 3708C for 10min(Cheng et al., 2006).
The net working volume of the cell was 24mL, and theelectrode packing was around 5,000m2/m3-graphite feltvolume. A reference electrode (Ag/AgCl electrode, 195mVvs. standard hydrogen electrode, SHE) was used to obtaincathode and anode potentials, with all voltages reported hereversus SHE. Two controls (duplicate reactors) were alsooperated: one was used as an abiotic control (no inoculum);the other was run in open circuit mode to examine changesin PCP and co-substrates in the absence of currentgeneration. All of the reactors were wrapped with aluminumfoil to exclude light.
Inoculation and Operation
Domestic wastewater was collected from the primary sedi-mentation tank of the Lingshui Wastewater Treatment Plantin Dalian, China, and used to inoculate the anode. For theinitial acclimation, wastewater was added into a solution(V/V: 50/50) containing (per liter) (NH4)2SO4 0.386 g,K2SO4 0.149 g, NaH2PO4�2H2O (3.31 g), Na2HPO4�12H2O(10.31 g), vitamins (12.5mL/L), and minerals (12.5mL/L)(pH 7.0, conductivity 6.5mS/cm) (Huang and Logan, 2008).Acetate or glucose was added at identical concentrationson a chemical oxygen demand (COD) basis. After theformation of stable and repeatable peaks in power, analyticalgrade PCP (Sigma, St. Louis, MO, 99.8%) dissolved in 0.2MNaOH, together with acetate or glucose in the nutrientsolution, was used as the medium. The replacement ofanodic solution was done at the end of each fed-batch cycle(<20mV). All reactors were operated at room temperature(22� 28C) except as indicated, where the temperature wasvaried to be 4, 35, and 508C. In the investigation of the
2212 Biotechnology and Bioengineering, Vol. 109, No. 9, September, 2012
effects of pH, the medium pH was adjusted using0.2N H2SO4 or NaOH to the indicated values while thesolution conductivity was kept the same at different pHvalues (conductivity change <5%).
Analyses
PCP was analyzed using a high performance liquid chro-matograph (HPLC Agilent 1100) (Supporting Materials,SM), with the intermediates quantitatively determined byHPLC after a qualitative analysis using an APCI (�) ion trapmass spectrometer coupled to an LC (Agilent HPLC-MS/MS6410).
The voltage across an external resistor was recorded(30min intervals) using a data acquisition board (PISO813,Taiwan). Power density was normalized by the volume ofthe liquid medium (24mL) and the projected surface area ofthe cathode (7 cm2). These parameters, as well as CE, werecalculated as previously reported (Huang and Logan, 2008).The maximum power densities were calculated frompolarization curves obtained by varying the externalresistance (20 kV–100V) for 60–90min (single-cyclemethod). The bioelectrochemical behavior of anodicbiofilms was examined using cyclic voltammetry (CV)and a three-electrode configuration with a potentiostat(CHI 650A, Chenhua, Shanghai). The scanned potentialwas between �0.6 and þ0.6 V (vs. SHE), at a scan rate of1.0mV/s (Huang et al., 2011d).
Community analysis was performed using a polymerasechain reaction (PCR) and denaturing gradient gel electro-phoresis (DGGE). Samples were collected from MFCs at theend of a cycle when the PCP (15mg/L) was fully depleted(pH 7.0 and 228C). Electrodes were fragmented using sterilescissors. Cells attached on the electrodes were removed byrinsing three times with sterile water, and concentrated bycentrifugation. Genomic DNA extraction, PCR amplifica-tion, and DGGE analyses were performed as described inSupplementary Materials.
Results and Discussion
Effect of the Number of Diffusion Layers
The number of diffusion layers affected both PCP degra-dation rates and power densities. Three diffusion layersproduced the highest PCP degradation rates (0.140�0.011mg/L-h, acetate; 0.117� 0.009mg/L-h, glucose) andpower densities (6.0� 0.9W/m3, 206� 30mW/m2, acetate;5.1� 0.9W/m3, 173� 32mW/m2, glucose), with CEs of57.6� 0.9% (acetate) and 40.4� 3.7% (glucose) for MFCsfed 5mg/L of PCP (SM; Table S1). These PCP degradationrates were similar to results with five diffusion layers, butpower densities were slightly lower. PCP degradation ratesand power densities were reduced using one or sevendiffusion layers. The application of additional diffusionlayers is known to reduce the oxygen flux into the anode
chamber (Cheng et al., 2006). This is consistent with resultshere, as shown by an increase in the CEs for both substrates(SM, Table S1). Based on these results, it appears that the useof three to five diffusion layers provided oxygen transfer intothe anode chamber that was optimal for maximizing PCPdegradation by anaerobic and aerobic pathways. As a resultof these findings, three diffusion layers were used in MFCsin all subsequent tests.
PCP Degradation Pathways
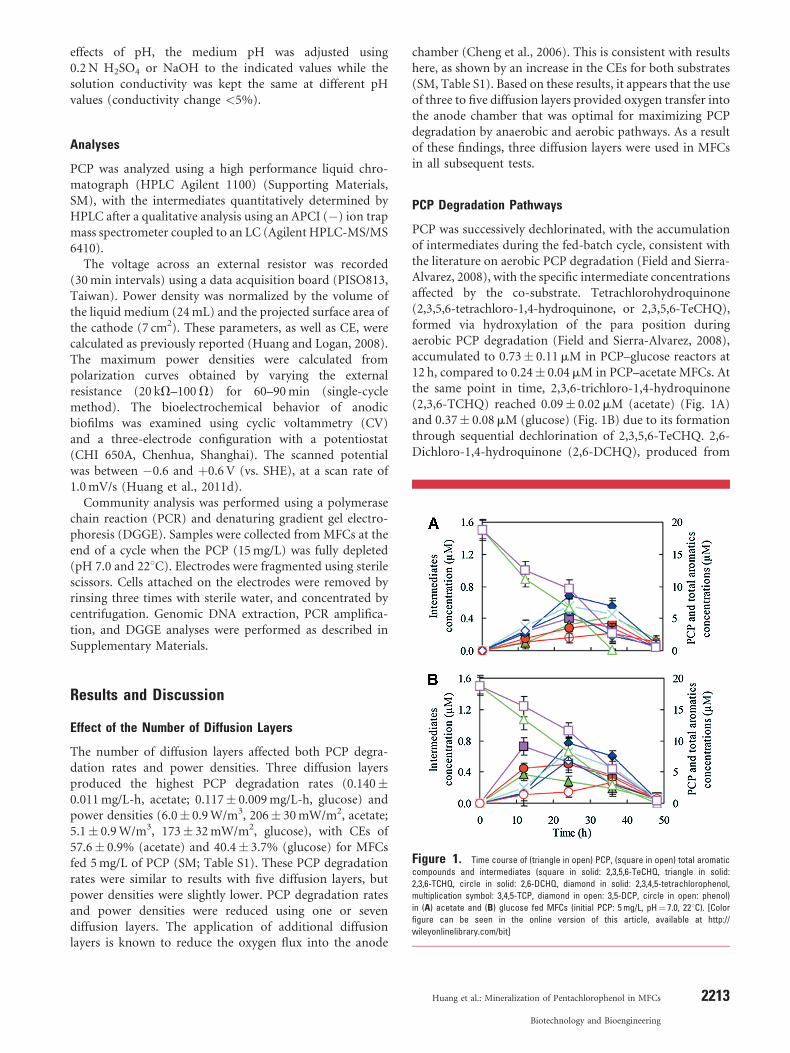
PCP was successively dechlorinated, with the accumulationof intermediates during the fed-batch cycle, consistent withthe literature on aerobic PCP degradation (Field and Sierra-Alvarez, 2008), with the specific intermediate concentrationsaffected by the co-substrate. Tetrachlorohydroquinone(2,3,5,6-tetrachloro-1,4-hydroquinone, or 2,3,5,6-TeCHQ),formed via hydroxylation of the para position duringaerobic PCP degradation (Field and Sierra-Alvarez, 2008),accumulated to 0.73� 0.11mM in PCP–glucose reactors at12 h, compared to 0.24� 0.04mM in PCP–acetate MFCs. Atthe same point in time, 2,3,6-trichloro-1,4-hydroquinone(2,3,6-TCHQ) reached 0.09� 0.02mM (acetate) (Fig. 1A)and 0.37� 0.08mM (glucose) (Fig. 1B) due to its formationthrough sequential dechlorination of 2,3,5,6-TeCHQ. 2,6-Dichloro-1,4-hydroquinone (2,6-DCHQ), produced from
Figure 1. Time course of (triangle in open) PCP, (square in open) total aromatic
compounds and intermediates (square in solid: 2,3,5,6-TeCHQ, triangle in solid:
2,3,6-TCHQ, circle in solid: 2,6-DCHQ, diamond in solid: 2,3,4,5-tetrachlorophenol,
multiplication symbol: 3,4,5-TCP, diamond in open: 3,5-DCP, circle in open: phenol)
in (A) acetate and (B) glucose fed MFCs (initial PCP: 5mg/L, pH¼ 7.0, 228C). [Colorfigure can be seen in the online version of this article, available at http://
wileyonlinelibrary.com/bit]
Huang et al.: Mineralization of Pentachlorophenol in MFCs 2213
Biotechnology and Bioengineering

2,3,6-TCHQ, increased to 0.35� 0.11mM (acetate) at 36 hand 0.50� 0.08mM (glucose) at 24 h, and then graduallydecreased in concentration.
2,3,4,5-Tetrachlorophenol, which is anaerobically pro-duced through the removal of ortho-chlorines, reachedmaximum concentrations of 0.69� 0.11mM (acetate)(Fig. 1A) and 0.78� 0.08mM (glucose) (Fig. 1B) at 24 h.Successive dechlorination of these compounds was shownby the production of expected intermediates (3,4,5-TCP,3,5-DCP, and phenol) (Chen et al., 2010; Field andSierra-Alvarez, 2008). Total aromatics decreased from aninitial concentration of 18.8� 1.6mM to a concentrationof 2.67� 1.6mM (acetate) (Fig. 1A) and 5.46� 1.8mM(glucose) at 36 h (Fig. 1B). Around 98% of all theintermediates were removed at 48 h, implying theeffective degradation and mineralization of PCP in thisreactor.
PCP was completely mineralized in the air-cathodesingle-chamber MFCs, overcoming the accumulation ofpartial de-chlorination products of 2,3,4,5-tetrachlorophe-nol and tetrachlorohydroquinone observed in previous two-chamber, hexacyanoferrate-cathode MFCs (Huang et al.,2011c). Here oxygen was transferred into the anode chamberthrough the cathode, in contrast to the two-chamberconditions where the anode is kept separated from thehexacyanoferrate solution and thus there is little oxygencrossover. The oxygen permeation through the cathode insingle-chamber MFCs created a micro-aerobic environmentthat allowed a combination of anaerobic–aerobic processesin the same chamber (Cheng et al., 2006; Cheng and Logan,2011), and thus led to more efficient PCP degradation.Based on the combination of aerobic and anaerobic PCPdegradation, possible PCP degradation pathways wereproposed as outlined in the supporting materials (SMFig. S1).
Effect of Initial COD Concentration on PowerProduction and PCP Degradation
Power increased from 3.1 to 6.8W/m3 (105–231mW/m2,acetate) and from 2.2 to 5.4W/m3 (74–185mW/m2,glucose) with an increase in the initial COD from 390 to780mg/L in the presence of 5mg/L of PCP (Fig. 2A and B).Higher initial CODs ranging from 1,170 to 1,560mg/Lgenerated similar powers of around 18.0W/m3 (617mW/m2,acetate) and 9.0W/m3 (315mW/m2, glucose). However,power generation was adversely affected by the presence ofPCP in the medium, as power was higher in the controlslacking PCP (19.4W/m3, 665mW/m2 with acetate; 15.0W/m3,513mW/m2 with glucose; COD¼ 780mg/L) (Fig. 2A andB). CEs decreased with an increase in COD, 74� 3% to24� 4% (acetate) and 51� 3% to 17� 3% (glucose), dueto the increase in the length of a fed-batch cycle (Fig. 2C).These substrate concentrations are equivalent to 226C(780mg/L COD) and 452C (1,560mg/L COD), comparedto only 0.4 C for PCP (5mg/L), assuming complete
dechlorination. Thus, the coulombs provided in thesubstrates were much higher than that in PCP.
The PCP degradation rate increased with initial CODconcentrations (Fig. 2D), reaching the maximum of0.140� 0.011mg/L-h (acetate) and 0.117� 0.009mg/L-h(glucose). These PCP degradation rates were both higherthan that previously achieved in two-chamber MFCs, with a35% increase for acetate (0.104mg/L-h) and a 92% increasefor glucose (0.061mg/L-h) at the same COD and PCPconcentrations (Huang et al., 2011c).
Current generation enhanced PCP degradation rates,demonstrating the importance of exoelectrogenic activityon PCP removal. In the open circuit controls, PCPdegradation rates were reduced by up to 31–54% withacetate (0.062� 0.006 to 0.104� 0.005mg/L-h), and 27–43% with glucose (0.052� 0.006 to 0.082� 0.007mg/L-h)for CODs ranging from 390 to 1,560mg/L (Fig. 2D). It hassimilarly been found that the degradation rates of otherpollutants (chloroethane and diesel) are improved withcurrent generation (Morris et al., 2009; Pham et al., 2009).In addition, these PCP degradation rates were higher thanthose achieved in the conventional biological processes, withrange from 0.038 to 0.051mg/L-h for PCP concentrations of5–13mg/L (Li et al., 2010; Mun et al., 2008; Szewczyk andDługonski, 2009).
PCP degradation in air-cathode single-chamber MFCsis more complex than that occurring in a single chambersystem, as a result of additional oxygen diffusion into thesystem. PCP is thought to be primarily degraded throughchemical reduction using electrons from the cathode,similar to that reported by others (Sun et al., 2009), orco-metabolized by microorganisms on the anode andelsewhere in the reactor using acetate or glucose. Whenusing an MFC, the enhanced growth of anaerobic andexoelectrogenic microorganisms in the reactor may help toincrease the microbial population capable of PCP degrada-tion compared to systems lacking current generation (Luoet al., 2010; Morris et al., 2009). These positive effects fromboth the anode and cathode, and the addition of someoxygen into the system through the air-cathode, may explainthe enhanced PCP degradation rates with current generationin theseMFCs compared to open circuit conditions inMFCsand in conventional biological processes.
Polarization Curves
Polarization data were obtained to determine the maximumpower production in these reactors in the presence andabsence of PCP. The maximum power densities were 7.7�1.1W/m3 (264� 39mW/m2, acetate) and 5.1� 0.1W/m3
(175� 5mW/m2, glucose) at 780mg/L of COD and 5mg/Lof PCP (SM, Fig. S2). Higher power (17.7� 0.9W/m3,605� 32mW/m2, acetate; 8.6� 0.4W/m3, 293� 12mW/m2,glucose) was obtained at a higher initial COD concentrationof 1,560mg/L, while a lower substrate concentration of390mg/L reduced maximum power (4.7� 0.6W/m3,
2214 Biotechnology and Bioengineering, Vol. 109, No. 9, September, 2012

161� 19mW/m2, acetate; 4.0� 0.6W/m3, 137� 19mW/m2, glucose) (SM, Fig. S2). These power densities were alllower than those produced by the controls with 780mg/LCOD and lacking PCP (19.4W/m3, 665mW/m2, for acetate;and 15.0W/m3, 513mW/m2 for glucose) (SM Fig. S2),demonstrating an adverse effect of PCP on systemperformance. These power densities using the graphitefelt anodes in the controls (19.4W/m3, 665mW/m2, withacetate) are lower than those previously reported (27W/m3,1,100mW/m2) using a similar reactor with ammonia-treated graphite fiber brush anodes (Cheng and Logan, 2007;Cheng et al., 2011). This suggests that power densities couldbe further increased. However, these maximum powerdensities in the presence of PCP were four times (acetate)and three times (glucose) as much as those previouslyobtained in two-chamber MFCs (Huang et al., 2011c),illustrating the importance of reactor architecture forimproving power generation and PCP degradation rates.
Effect of Initial PCP Concentration
PCP degradation rate increased with its concentration over arange of 5–15mg/L to maximum rates of 0.171� 0.01mg/L-h(acetate) and 0.159� 0.011mg/L-h (glucose), but the ratedecreased slightly at 20mg/L (Fig. 3A). PCP degradation
rates with current generation were always larger than thoseof the open circuit controls (Fig. 3A).
Power production was inversely proportional to initialPCP concentration, with changes of 0.255W/mg PCP(acetate, R2¼ 0.9977) and 0.184W/mg PCP (glucose,R2¼ 0.9934) over the concentration range of 5–20mg/Lof PCP (Fig. 3B). This effect of initial PCP concentration onpower production was primarily due to reduced anodeperformance, as shown by an increase in the anode potentialwith initial PCP concentration increase (Fig. 3C).
Effect of Initial pH
A lower pH was beneficial to PCP degradation, but notpower production. PCP degradation rates with acetatedecreased from amaximum of 0.195� 0.002mg/L-h at pH 5to 0.154� 0.003mg/L-h at pH 10 (Fig. 4A). Glucose fedreactors followed the same trend, with PCP degradation ratedecreasing between a pH of 5 (0.173� 0.005mg/L-h) to 10(0.136� 0.004mg/L-h) (Fig. 4A). PCP degradation rateswere lower in the open circuit controls (Fig. 4A), showingthe benefit of current generation for PCP degradation atall pH values. These results are consistent with conventionalbiological processes, where it was found that a pH of 6–7 wasbeneficial to PCP degradation due to the requirement of five
Figure 2. Power generation from (A) PCP–acetate and (B) PCP–glucose MFCs, (C) CE, and (D) PCP degradation rate in closed circuit (solid symbol) and open circuit (open
symbol) conditions at various initial COD concentrations (Rex¼ 200V, initial PCP: 5 mg/L, pH 7.0, temperature: 228C, diamond: PCP–acetate, circle: PCP–glucose). Arrows indicatewhen reactors were fed with fresh medium; two repeated cycles were shown here. COD values with peaks in (A) and (B) from left to right: 780mg/L control (no PCP); 390mg/L;
780mg/L; 1,170 mg/L; 1,560 mg/L. [Color figure can be seen in the online version of this article, available at http://wileyonlinelibrary.com/bit]
Huang et al.: Mineralization of Pentachlorophenol in MFCs 2215
Biotechnology and Bioengineering

protons for PCP de-chlorination (Mun et al., 2008; Singhet al., 2009). Power production increased with pH, to amaximum of 5.5� 0.3W/m3 (189� 10mW/m2) at pH 9(acetate), and 3.3� 0.3W/m3 (113� 10mW/m2) at pH 8(glucose) (Fig. 4B). An optimal pH¼ 9 for acetate isconsistent with previous reports using this substrate in theabsence of PCP (He et al., 2008).
Changes in power production were due to changes inanode potentials. There was no appreciable change ofcathode potential with the change of pH from 5 to 10 ineither PCP–acetate or PCP–glucose MFCs. Anode potentialsdecreased inversely with pH over the range of 5 to 9 foracetate, and from 5 to 8 for glucose (Fig. 4C). More negativeanode potentials accounted for the increase in powergeneration with pH, showing that the higher pH values werein general beneficial for exoelectrogenesis (Fig. 4C).
The effect of pH on the biocatalytic activities of the anodebiofilms was further assessed using CV. Only a single set of
oxidation–reduction peaks were observed across the pHrange of 5–10 for all biofilms, with no peaks measured inabiotic controls (Fig. 5A and B). The peaks using acetatewere in the range of �0.26 to �0.32 V (Fig. 5A), consistentwith previous reports using this substrate at similar pH andin the absence of PCP (Liu et al., 2005b), indicating the lesseffect of PCP and pH on the potentials of the oxidation–reduction peaks. In contrast, the peaks using glucoseappeared at the range of þ0.36 to þ0.40 V (Fig. 5B), morepositive than the reported�0.01 toþ0.36 V at similar pH inthe absence of PCP (Rabaey et al., 2004), demonstrating theimportance of PCP on the potentials of the oxidation–reduction peaks. These results show that substrate, muchmore than pH, had a substantial impact on the potentials ofthe oxidation–reduction peaks. The sizes of oxidation–reduction peaks were significantly affected by pH, withmaximums at pH 9 (acetate) and 8 (glucose), consistentwith the highest power densities at these pHs (Fig. 4B).
Figure 3. Effect of initial PCP concentration on (A) PCP degradation rate (symbol
in solid: closed circuit condition, symbol in open: open circuit condition), (B) power
production, and (C) cathode (open) and anode (solid) potentials (Rex¼ 200V, pH 7.0,
temperature: 228C, initial COD: 780mg/L, diamond: PCP–acetate, circle: PCP–glucose).[Color figure can be seen in the online version of this article, available at http://
wileyonlinelibrary.com/bit]
Figure 4. Effect of initial pH on (A) PCP degradation rate (solid: closed circuit
condition; open: open circuit condition), (B) power production and (C) cathode (open)
and anode (solid) potentials (Rex¼ 200V, temperature: 228C, initial COD: 780 mg/L,initial PCP: 15mg/L, diamond: PCP–acetate, circle: PCP–glucose). [Color figure can be
seen in the online version of this article, available at http://wileyonlinelibrary.com/bit]
2216 Biotechnology and Bioengineering, Vol. 109, No. 9, September, 2012

Effect of Temperature
Temperature affected all aspects of the MFC, includingmaximum power densities, open circuit potentials, CEs, andPCP degradation rates. The maximum power density wasobtained at 358C, with 6.3� 0.3W/m3 (216� 10mW/m2,acetate) (Fig. 6A) and 4.1� 0.4W/m3 (141� 15mW/m2,glucose) (Fig. 6C), with lower power densities at the othertemperatures in the order: 228C> 48C> 508C. This generaltrend with performance was similar with the changesin open circuit potentials, with the highest open circuitpotentials achieved at 358C (0.73� 0.001V, acetate;0.65� 0.002 V, glucose) (Fig. 6A and C). These resultsof power versus temperature are consistent with thoseobtained inMFCs in the absence of PCP, where it was shownthat lower temperatures reduced performance (Cheng et al.,2011). Here it was also observed that a much highertemperature of 508C also substantially reduced performance(1.0� 0.04W/m3, 34� 2mW/m2, acetate; 0.8� 0.1W/m3,27� 5mW/m2, glucose), implying the absence of thermo-philic exoelectrogenic bacteria (Liu et al., 2011; Mathis et al.,2008).
The changes in the electrode potentials (Fig. 6B and D)indicated that the anode potential was more affected by
temperature than the cathode. Anode potentials for MFCsfed glucose changed more than those with acetate(Fig. 6B and D). It has been agreed that, in the absenceof PCP and using acetate as a fuel, reducing temperaturefrom 30 to 48C led to an increase in the anode potential by34% and a decrease of the cathode potential by 37% due tothe decrease of cathodic reaction rates and exoelectrogenicbacteria activities, although the solubility of dissolvedoxygen in water increased (Cheng et al., 2011; Oh et al.,2004). Thus, the presence of PCP here mainly contributed tothe greater sensitivity of anodic exoelectrogenic bacteria totemperature, which resulted in greater changes in anodepotential with temperature.
CEs increased with a temperature change from 22 to358C, resulting in CE¼ 38.3� 1.5% (acetate) and 31.3�1.7% (glucose) at 358C (SM Fig. S3). These CEs are lowerthan those obtained in the absence of PCP (Liu et al., 2008;Logan et al., 2007), mainly due to the increased oxygentransfer into the reactor with less diffusion layers here,as well as the longer cycles due to the adverse effect of PCPon exoelectrogenic bacteria. At 4 or 508C, the CEs weresubstantially reduced. For example, for acetate the CEsdecreased to 4.2� 2.1% at 508C and 10.8� 1.3% at 48C.These values were much lower than a CE¼ 31% at 48C in theabsence of PCP (Cheng et al., 2011), suggesting that the lowCE was resulted from effects of PCP on the anodic biofilm.
PCP degradation rates were consistent with other changesin performance, with the maximum degradation ratesobtained at 358C (0.188� 0.01mg/L-h, acetate; 0.172�0.01mg/L-h, glucose) (SM, Fig. S3). Temperatures of 50 or48C substantially decreased PCP degradation rates (SM, Fig.S3). In all the cases, PCP degradation rates under closedcircuit conditions were higher than those under open circuitconditions, demonstrating that current generation increasedPCP degradation rates.
While PCP mineralization under combined anaerobic–aerobic conditions in air-cathode single-chamber MFCsprovides a promising and efficient process for remediationof water contaminated with PCP, there are still manychallenges to enable practical applications. The platinumcatalyst used here on the cathode should be replaced byeither nonprecious metal catalysts (Logan, 2010) or bio-cathodes (Huang et al., 2011a; Logan, 2010) to reducecapital costs. In addition, oxygen transport throughthe cathode could be adjusted by modifying the cathodematerial or using different diffusion layers on the outsideof the cathode (Cheng et al., 2006; Luo et al., 2011; Zhanget al., 2011a) in order to better stimulate PCP degradation.Further investigations in these directions are warranted.
Bacterial Morphologies and Community
Both PCP–acetate (Fig. 7A) and PCP–glucose (Fig. 7C)MFCs had anodes sparsely populated with bacteria. This wasin contrast to anodes in acetate (Fig. 7B) or glucose-fed(Fig. 7D) MFCs in the absence of PCP, where dense biofilms
Figure 5. Cyclic voltammetry tests carried out on the anode fed with (A) PCP–
acetate and (B) PCP–glucose (triangle in solid: abiotic control, diamond in solid: pH 5.0,
circle in open: pH 6.0, diamond in open: pH 7.0, square in open: pH 8.0, triangle in
open: pH 9.0, circle in solid: pH 10.0, temperature: 228C, initial COD: 780mg/L, initialPCP: 15mg/L). [Color figure can be seen in the online version of this article, available at
http://wileyonlinelibrary.com/bit]
Huang et al.: Mineralization of Pentachlorophenol in MFCs 2217
Biotechnology and Bioengineering

Figure 6. A, C: Voltage (solid) and power (open), and (B and D) cathode (open) and anode (solid) potentials as a function of current density in PCP–acetate (A and B) or
PCP–glucose (C and D) fedMFCs under temperature of 508C (circle), 358C (triangle), 228C (diamond) and 48C (square) (pH: 7.0, initial COD: 780mg/L, initial PCP: 15mg/L). [Color figurecan be seen in the online version of this article, available at http://wileyonlinelibrary.com/bit]
Figure 7. Morphological features of biofilms on the anode in (A) PCP–acetate or (C) PCP–glucose MFCs compared to the controls of (B) acetate or (D) glucose MFCs (pH: 7.0,
initial COD: 780mg/L, initial PCP: 15mg/L).
2218 Biotechnology and Bioengineering, Vol. 109, No. 9, September, 2012

covered the anodes. The different performance and bacterialabundances in these systems were presumably due to thechanges induced by the presence of PCP.
Bacterial communities by DGGE indicated that PCP–acetate and PCP–glucose anodes had several commonand prominent bands, but also some different bands fromacetate and glucose controls lacking PCP (Fig. 8). Bands ofA2 from acetate controls and G2 from glucose controlsshared sequences belonging to the well-known exoelectro-genic Geobacter sulfurreducens (Kiely et al., 2011; Logan,2009), which were absent in PCP–glucose or faint in PCP–acetate reactors (Fig. 8), implying the negative effect ofPCP on this strain. In addition, predominant bands wereidentified to be most similar to exoelectrogenic Bacteroidetessp. (A1) and Comamonadaceae sp. (A3) in acetate MFCs,
and Desulfovibrio desulfuricans (G1), Clostridium sp. (G3)and Burkholderiales sp. (G4) in glucose reactors (Boroleet al., 2011; Kan et al., 2011; Logan, 2009; Prasad et al., 2006).In the case of PCP–acetate and PCP–glucose MFCs,however, the prominent strains experienced apparent shifts(Fig. 8). The sequences of PA1 from PCP–acetateMFCs weremost similar to Pseudomonas putida and Pseudomonas sp.,both of which were frequently found to cometabolize PCP,1,3-dichlorobenzene, alpha-halocarboxylic acid, or crude-oil (Field and Sierra-Alvarez, 2008; Marchesi andWeightman, 2003; Mulet et al., 2011). Pseudomonas putidahas been found to be exoelectrogenic (Juang et al., 2011).The sequences of band PA2 from PCP–acetate anode weremost similar to uncultured Alicycliphilus sp., and BacteriumrA3 and rP5, which were found to be present in
Figure 8. Neighbor-joining tree based on 16S rRNA gene sequences derived from the DGGE band using Clustal X 2.0 (Un indicates uncultured, and C nitrativorans indicates
Comamonas nitrativorans). Inset figure: anodic bacterial community profiles revealed by DGGE. (From left to right: acetate without PCP; glucose without PCP; PCP–glucose; PCP–
acetate. Bands A1–A3, G1–G4, PG1–PG5 and PA1–PA2 represented selected DGGE bands that were excised and sequenced from acetate, glucose, PCP–glucose, and PCP–acetate
fed MFCs, respectively).
Huang et al.: Mineralization of Pentachlorophenol in MFCs 2219
Biotechnology and Bioengineering

chlorophenol degrading aerobic granular sludge, or respon-sible for the breakdown of a phenol-digesting activated-sludge process (Watanabe et al., 1998, 1999). The band PA2was most similar to Comamonas nitrativorans, dominant ina microbial community for treating carbazole-containingwastewater or landfill leachate (Etchebehere et al., 2001; Tanand Ji, 2010), but its exoelectrogenic activity is unknown.The presence of these diverse bacteria that are capable ofdegrading multiple recalcitrant compounds, as well as thepresence of bacteria with exoelectrogenic activities, cantherefore explain the successful dechlorination of PCPand mineralization with simultaneous power generation inacetate fed MFCs.
In the case of PCP–glucose anode, band PG1 shared a highsimilarity to phytase-producing Shigella dysenteriae, whereassequences of PG2 were similar to Klebsiella sp., Klebsiellavariicola, and Klebsiella pneumoniae (Fig. 8). Klebsiellapneumoniae has been shown to be exoelectrogenic withglucose (Logan, 2009; Zhang et al., 2008) and it can degradethe recalcitrant compound citrinin (Chen et al., 2011),suggesting that this microorganism plays a role in both PCPdegradation and power generation. The band PG4 present inthe PCP–glucose MFC, but not in the control, belonged toan uncultured gamma proteobacterium, which was presum-ably developed due to the presence of PCP. These results forboth the acetate and glucose fed MFCs demonstrate thatthere is a large diversity of bacteria in these reactors, and thatthe communities are altered by the presence of PCP.
Conclusions
PCP was shown for the first time here to be completelymineralized under a combined anaerobic–aerobic conditioncreated in air-cathode, single-chamber MFCs. PCP degra-dation rates were improved by 35% (acetate) and 92%(glucose), and power production by 300% (acetate) and200% (glucose) compared to previous results using two-chamberedMFCs (Huang et al., 2011c). Amore acidic pH of5 improved PCP degradation rates, but power productionwas maximized at higher pHs of 9 for acetate, and 8for glucose. For both acetate and glucose fed MFCs, atemperature of 358C was optimal for not only PCPdegradation but also power production. Dominant bacteriathat are known exoelectrogens and that were identified inPCP–acetate and PCP–glucose fed biofilms include thePseudomonas and Klebsiella species. These results show thatMFCs can be used for successful bioremediation of PCP-contaminated water using co-substrates such as acetateor glucose. The use of single chamber MFCs in particularcan result in complete mineralization of PCP underoptimum operational conditions of PCP concentrationsbelow 20mg/L.
We gratefully acknowledge financial support from the National Basic
Research Program of China (No. 2011CB936002) and the Natural
Science Foundation of China (Nos. 21077017 and 51178077).
References
Banerji SK, Bajpai RK. 1994. Cometabolism of pentachlorophenol by
microbial species. J Hazard Mater 39:19–31.
Borole AP, Hamilton CY, Vishnivetskaya TA. 2011. Enhancement in
current density and energy conversion efficiency of 3-dimensional
MFC anodes using pre-enriched consortium and continuous supply
of electron donors. Biores Technol 102:5098–5104.
Chen Y, Lin CJ, Lan H, Fu S, Zhan H. 2010. Changes in pentachlorophenol
(PCP) metabolism and physicochemical characteristics by granules
responding to different oxygen availability. Environ Prog Sustainable
Energy 29:307–312.
Chen YH, Sheu SC, Mau JL, Hsieh PC. 2011. Isolation and characterization
of a strain of Klebsiella pneumoniae with citrinin-degrading activity.
World J Microbiol Biotechnol 27:487–493.
Cheng SA, Logan BE. 2007. Ammonia treatment of carbon cloth anodes
to enhance power generation of microbial fuel cells. Electrochem
Commun 9:492–496.
Cheng SA, Logan BE. 2011. Increasing power generation for scaling up
single-chamber air cathode microbial fuel cells. Biores Technol
102:4468–4473.
Cheng SA, Liu H, Logan BE. 2006. Increased power generation in
a continuous flow MFC with advective flow through the porous
anode and reduced electrode spacing. Environ Sci Technol 40:2426–
2432.
Cheng SA, Xing DF, Logan BE. 2011. Electricity generation of single-
chamber microbial fuel cells at low temperatures. Biosens Bioelectron
26:1913–1917.
Damianovic MHRZ, Moraes EM, Foresti MZE. 2009. Pentachlorophenol
(PCP) dechlorination in horizontal-flow anaerobic immobilized bio-
mass (HAIB) reactors. Biores Technol 100:4361–4367.
Etchebehere C, Errazquin MI, Dabert P, Moletta R, Muxi L. 2001.
Comamonas nitrativorans sp. nov., a novel denitrifier isolated from a
denitrifying reactor treating landfill leachate. Int J Syst Evol Microbiol
51:977–983.
Field JA, Sierra-Alvarez R. 2008. Microbial degradation of chlorinated
phenols. Rev Environ Sci Biotechnol 7:211–241.
He Z, Huang Y, Manohar AK, Mansfeld F. 2008. Effect of electrolyte pH on
the rate of the anodic and cathodic reactions in an air-cathode
microbial fuel cell. Bioelectrochem 74:78–82.
Huang LP, Logan BE. 2008. Electricity production from xylose in fed-batch
and continuous-flow microbial fuel cells. Appl Microbiol Biotechnol
80:655–664.
Huang LP, Regan JM, Quan X. 2011a. Electron transfer mechanisms, new
applications, and performance of biocathodemicrobial fuel cells. Biores
Technol 102:316–323.
Huang LP, Cheng SA, Chen GH. 2011b. Bioelectrochemical systems for
efficient recalcitrant wastes treatment. J Chem Technol Biotechnol
86:481–491.
Huang LP, Gan LL, Zhao QL, Logan BE, Lu H, Chen GH. 2011c.
Degradation of pentachlorophenol with the presence of fermentable
and non-fermentable co-substrates in a microbial fuel cell. Biores
Technol 102:8762–8768.
Huang LP, Chai XL, Cheng SA, Chen GH. 2011d. Evaluation of carbon-
based materials in tubular biocathode microbial fuel cells in terms of
hexavalent chromium reduction and electricity generation. Chem Eng J
166:652–661.
Juang DF, Yang PC, Chou HY, Chiu LJ. 2011. Effects of microbial species,
organic loading and substrate degradation rate on the power generation
capability of microbial fuel cells. Biotechnol Lett 33:2147–2160.
Kan J, Hsu L, Cheung AM, Pirbazari M, Nealson K. 2011. Current
production by bacterial communities in microbial fuel cells enriched
from wastewater sludge with different electron donors. Environ Sci
Technol 45:1139–1146.
Kiely PD, Rader GK, Regan JM, Logan BE. 2011. Long-term cathode
performance and the microbial communities that develop in microbial
fuel cells fed different fermentation endproducts. Biores Technol
102:361–366.
2220 Biotechnology and Bioengineering, Vol. 109, No. 9, September, 2012

Li Z, Yang S, Inoue Y, Yoshida N, Katayama A. 2010. Complete anaerobic
mineralization of pentachlorophenol (PCP) under continuous flow
conditions by sequential combination of PCP-dechlorinating and
phenol-degrading consortia. Biotechnol Bioeng 107:775–785.
Liu H, Cheng SA, Logan BE. 2005a. Power generation in fed-batch
microbial fuel cells as a function of ionic strength, temperature, and
reactor configuration. Environ Sci Technol 39:5488–5493.
Liu H, Cheng SA, Logan BE. 2005b. Production of electricity from acetate
or butyrate using a single-chamber microbial fuel cell. Environ Sci
Technol 39:658–662.
Liu H, Cheng SA, Huang LP, Logan BE. 2008. Scale-up of membrane-free
single-chamber microbial fuel cells. J Power Sources 179:274–279.
Liu Y, Climent V, Berna A, Feliu JM. 2011. Effect of temperature on the
catalytic ability of electrochemically active biofilm as anode catalyst in
microbial fuel cells. Electroanalysis 23:387–394.
Logan BE. 2009. Exoelectrogenic bacteria that power microbial fuel cells.
Nat Rev Microbiol 7:375–381.
Logan BE. 2010. Scaling upmicrobial fuel cells and other bioelectrochemical
systems. Appl Microbiol Biotechnol 85:1665–1671.
Logan BE, Regan JM. 2006. Microbial fuel cells, challenges and applications.
Environ Sci Technol 40:5172–5180.
Logan BE, Cheng S, Watson V, Estadt G. 2007. Graphite fiber brush anodes
for increased power production in air-cathode microbial fuel cells.
Environ Sci Technol 41:3341–3346.
Luo Y, Zhang R, Liu G, Li J, Li M, Zhang C. 2010. Electricity generation
from indole and microbial community analysis in the microbial fuel
cell. J Hazard Mater 176:759–764.
Luo Y, Zhang F, Wei B, Liu G, Zhang R, Logan BE. 2011. Power generation
using carbon mesh cathodes with different diffusion layers in microbial
fuel cells. J Power Sources 196:9317–9321.
Marchesi JR, Weightman AJ. 2003. Comparing the dehalogenase gene pool
in cultivated alpha-halocarboxylic acid-degrading bacteria with the
environmental metagene pool. Appl Environ Microbiol 69:4375–4382.
Mathis BJ, Marshall CW, Milliken CE, Makkar RS, Creager SE, May HD.
2008. Electricity generation by thermophilic microorganisms from
marine sediment. Appl Microbiol Biotechnol 78:147–155.
Morris JM, Jin S, Pruden A. 2009.Microbial fuel cell in enhancing anaerobic
biodegradation of diesel. Chem Eng J 146:161–167.
Mulet M, David Z, Nogales B, Bosch R, Lalucat J, Garcia-Valdes E. 2011.
Pseudomonas diversity in crude-oil-contaminated intertidal sand sam-
ples obtained after the prestige oil spill. Appl Environ Microbiol
77:1076–1085.
Mun CH, He J, Ng WJ. 2008. Pentachlorophenol dechlorination by an
acidogenic sludge. Water Res 42:3789–3798.
Oh S, Min B, Logan BE. 2004. Cathode performance as a factor in electricity
generation in microbial fuel cells. Environ Sci Technol 38:4900–4904.
Pant D, Bogaert GV, Diels L, Vanbroekhoven K. 2010. A review of the
substrates used in microbial fuel cells (MFCs) for sustainable energy
production. Biores Technol 101:1533–1543.
Pham H, Boon N, Marzorati M, Verstraete W. 2009. Enhanced removal of
1,2-dichloroethane by anodophilic microbial consortia. Water Res
43:2936–2946.
Prasad D, Sivaram TK, Berchmans S, Yegnaraman V. 2006. Microbial fuel
cell constructed with a micro-organism isolated from sugar industry
effluent. J Power Sources 160:991–996.
Rabaey K, Boon N, Siciliano SD, Verhaege M, Verstraete W. 2004. Biofuel
cells select for microbial consortia that self-mediate electron transfer.
Appl Environ Microbiol 70:5373–5382.
Ren ZY, Ward TE, Regan JM. 2007. Electricity production from cellulose in
a microbial fuel cell using a defined binary culture. Environ Sci Technol
41:4781–4786.
Shen DS, Liu XW, Feng HJ. 2005. Effect of easily degradable substrate on
anaerobic degradation of pentachlorophenol in an upflow anaerobic
sludge blanket (UASB) reactor. J Hazardous Materials B119:239–243.
Singh S, Singh BB, Chandra R, Patel DK, Rai V. 2009. Synergistic biodeg-
radation of pentachlorophenol by Bacillus cereus (DQ002384), Serratia
marcescens (AY927692) and Serratia marcescens (DQ002385). World J
Microbiol Biotechnol 25:1821–1828.
Stuart SL, Woods SL. 1998. Kinetic evidence for pentachlorophenol-de-
pendent growth of a dehalogenating population in a pentachlorophe-
nol- and acetate-fed methanogenic culture. Biotechnol Bioeng 57:420–
429.
Sun J, Hu Y, Bi Z, Cao Y. 2009. Simultaneous decolorization of azo dye and
bioelectricity generation using a microfiltration membrane air-cathode
single-chamber microbial fuel cell. Biores Technol 100:3185–3192.
Szewczyk R, Długonski J. 2009. Pentachlorophenol and spent engine oil
degradation by Mucor ramosissimus. Int Biodeter Biodegr 63:123–129.
Tan Y, Ji G. 2010. Bacterial community structure and dominant bacteria in
activated sludge from a 70 degrees ultrasound-enhanced anaerobic
reactor for treating carbazole-containing wastewater. Biores Technol
101:174–180.
Visvanathan C, Thu LN, Jegatheesan V, Anotai J. 2005. Biodegradation of
pentachlorophenol in a membrane bioreactor. Desalination 183:455–
464.
Watanabe K, Teramoto M, Futamata H, Harayama S. 1998. Molecular
detection, isolation, and physiological characterization of functionally
dominant phenol-degrading bacteria in activated sludge. Appl Environ
Microbiol 64:4396–4402.
Watanabe K, Teramoto M, Harayama S. 1999. An outbreak of nonfloccu-
lating catabolic populations caused the breakdown of a phenol-digest-
ing activated-sludge process. Appl Environ Microbiol 65:2813–2819.
Watson VJ, Saito T, Hickner MA, Logan BE. 2011. Polymer coatings as
separator layers for microbial fuel cell cathodes. J Power Sources
196:3009–3014.
Yang S, Shibata A, Yoshida N, Katayama A. 2005. Anaerobic mineralization
of pentachlorophenol (PCP) by combining PCP-dechlorinating and
phenol-degrading cultures. Biotechnol Bioeng 102:81–90.
Ye F, Shen D, Feng X. 2004. Anaerobic granule development for removal of
pentachlorophenol in an upflow anaerobic sludge blanket (UASB)
reactor. Process Biochem 39:1249–1256.
Zhang L, Zhou S, Zhuang L, Li W, Zhang J, Lu N, Deng L. 2008. Microbial
fuel cell based on Klebsiella pneumoniae biofilm. ElectrochemCommun
10:1641–1643.
Zhang F, Pant D, Logan BE. 2011a. Long-term performance of activated
carbon air cathodes with different diffusion layer porosities in micro-
bial fuel cells. Biosens Bioelectron 30:49–55.
Zhang XY, Cheng SA, Liang P, Huang X, Logan BE. 2011b. Scalable air
cathode microbial fuel cells using glass fiber separators, plastic mesh
supporters, and graphite fiber brush anodes. Biores Technol 102:372–
375.
Huang et al.: Mineralization of Pentachlorophenol in MFCs 2221
Biotechnology and Bioengineering





























