Embed Size (px)
Citation preview

Middle Devensian deposits beneath the 'Upper Floodplain' terrace ofthe River Thames at Kempton Park, Sunbury, England
P. L. Gibbard, G. R. Coope, A. R. Hall, R. C. Preece and J. E. Robinson
GIBBARD, P. L., G. R. COOPE, A. R. HALL, R. C. PREECE & J. E. ROBINSON. 1981.Middle Devensian deposits beneath the 'Upper Floodplain' terrace of the River Thames atKempton Park, Sunbury, England. Proc. Geol. Ass., 93 (3), 275-289. Gravel pit sections in the'Upper Floodplain' terrace exposed current-bedded gravel and sand containing a grey clayey siltlens. This silt has yielded fossil macroscopic plant remains, molluscs, ostracods and insects.Sedimentological and palaeontological results indicate that the silts were deposited in a slowlyflowing river channel. The gravel and sand accumulated under cool, seasonally contrasting climaticconditions and represent braided stream deposits. A new lithostratigraphical unit, the KemptonPark Gravel, is proposed and its relation to the local terrace stratigraphy of the Thames isdiscussed. A radio-carbon date of 35,230 ± 185 BP indicates that the Kempton Park Gravel unitaggraded during the Middle Devensian substage.
P. L. Gibbard, Subdepartment of Quaternary Research. Botany School. Downing Street,Cambridge CB23EA.G. R. Coope, Department of Geological Sciences. University of Birmingham. P.O. Box 363,Birmingham B152TT.A. R. Hall. Environmental Archaeology Unit, University of York. Heslington, York YO] 5DD.R. C. Preece. Department of Geology. Imperial College. Prince Consort Road, London SW72BP.J. E. Robinson, Department of Geology. University College, Gower Street, London WC] E 6BT.
1. INTRODUCTION
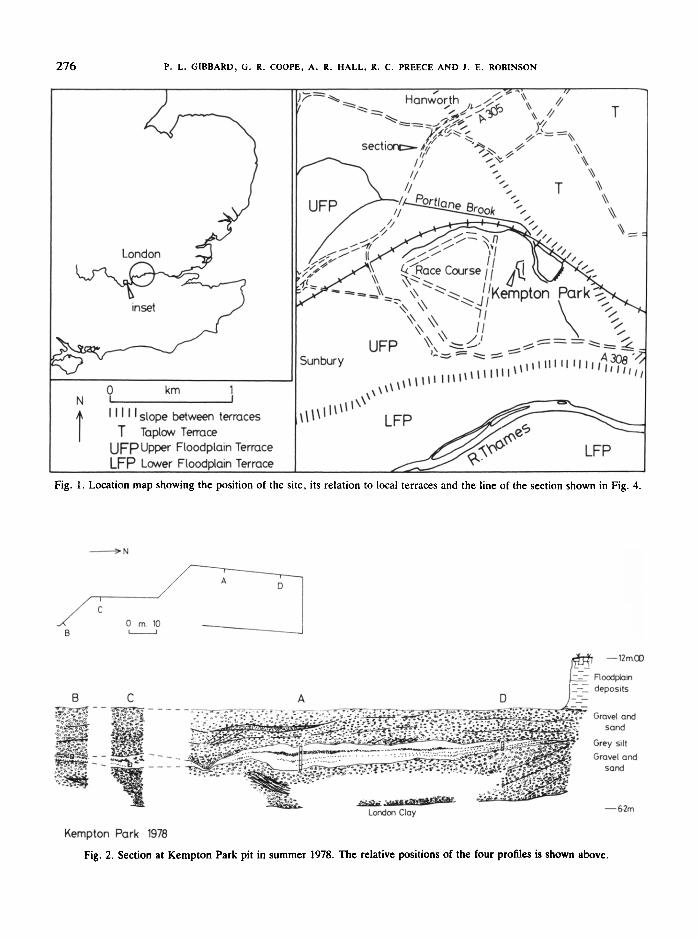
During summer 1978, at Kempton Park, Sunbury,Surrey, a bed of grey clayey silt containing fossilplant and animal remains was discovered in a gravel pit(TQ 118703). The pit was excavated in the RiverThames 'Upper Floodplain' terrace deposits (Dewey &Bromehead, 1921) which have been extensively workedin the district. The pit is located immediately east of theKempton Park Race Course and south of the HamptonShepperton railway line (Fig. 1). The ground surface atthe site is at 12 mOD.
The local geology and internal terrace stratigraphy ofthe area have been very little studied. Dewey & Bro-nehead (1915, 1921) mapped the surfaces and deposits ofterraces of the district, including the 'Upper Floodplain'terrace, the existence of which was supported by Zeuner(1959). More recently, Coope & Angus (1975) describedMiddle Devensian organic deposits containing faunaland floral remains downstream at Isleworth. These deposits were previously reported from nearby Twickenham by Leeson & Laffan (1894). The dating of the'Upper Floodplain' terrace aggradation is complicatedby the occurrence of Ipswichian interglacial deposits atBrentford (Zeuner 1959) and Trafalgar Square (Franks,Sutcliffe, Kerney & Coope, 1958; Franks 1960) beneaththe terrace. The aim ofthis paper is to present the resultsof investigations of the Kempton Park deposits andtheir bearing on the geological history of the depositsbeneath the 'Upper Floodplain' terrace of the RiverThames.
2. SITE DESCRIPTION
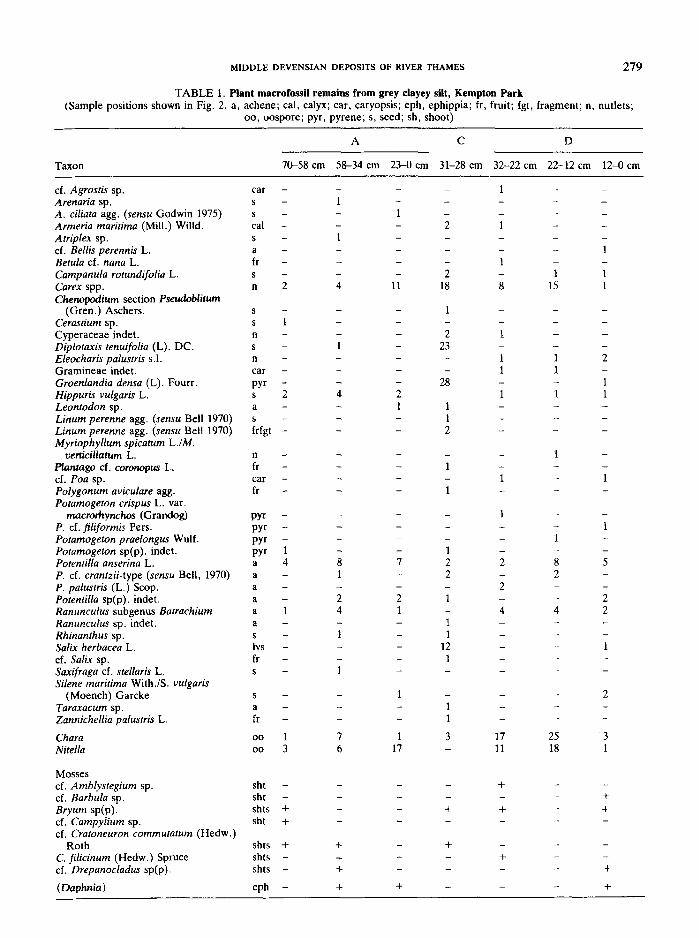
The deposits exposed at the Kempton Park site in summer 1978 are summarised in Fig. 2. The sequence consists of up to 4 m of current-bedded gravel and sand thatlies on an irregularly channelled surface of London Clay.The sediments comprise predominantly horizontallybedded clast-supported gravels that in places containnarrow, laterally impersistent lenses of current-beddedsand up to 20 em in thickness. They are almost exclusively traction load sediments laid down in broad, shallow, frequently cross-cutting channel and bar-like structures under highly variable flow conditions. Thesedimentary structures and facies relationships suggestthat the gravel and sand was probably deposited by abraided river (Williams & Rust, 1969; Rust, 1972) thatflowed towards the ENE, as shown by palaeocurrentmeasurements. A series of gravel samples for pebblecounting in the 32-8 mm size range gave a uniformcomposition of 93 per cent flint, 5 per cent vein quartz, 1per cent quartzite and 1 per cent Lower Greensand chert(using 300-600 pebbles in each of four samples, cf. Gibbard 1974). Over most ofthe western face of the quarry,2·2 m above the base (at point A), a grey clayey silt bedcontaining macroscopic plant remains, mollusc shells,ostracod valves and insects is intercalated in the gravel.The bed rests on the gravel beneath with a sharplydefined base and is in turn succeeded by further graveland sand strongly channelled into the clayey silt. Thiserosional base of the overlying gravel results in theclayey silt bed being of variable thickness; in places it is
275
276 P. L. GIBBARD, G. R. COOPE, A. R. HALL, R. C. PREECE AND J. E. ROBINSON
I II I Islope between terraces
T Taplow TerraceUFPUpper Floodplain TerraceLFP Lower Floodplain Terrace
T
LFP
LFP
/~::: ~n,,"// .'/
!?~ce CCXJrse!I J~\\ W" :--... a
- ... ,.. =\\_ \\ -....:~ .::::- " Kempton Park:"- ::-- ~ \\ .::::- J, \ ~'.:: 1 / '/
~ ~ "~ Ii ~
UFP ~\\\:=. -!--' --'~ :::= = = ~ = ~J_.::::;. = .:::::: ::::. -- A308 '
\ \ 1111111 1 11111 I" I I11\1\\111111 I I
Sunbury
\ \ \ \ I III\\\
\\\\ \ \ \ , 1\ \ \ ,
kmoIN
tFig. 1. Location map showing the position of the site, its relation 10 local terraces and the line of the section shown in Fig. 4.
B
/r-r------',~ J- 12mCD
B C A
~ -'I.rtI.,.,...P4WLondon Cloy
o
Grey silt
Gravel andsand
- &2m
Kempton Park 1978
Fig. 2. Section at Kempton Park pit in summer 1978. The relative positions of the four profiles is shown above .

MIDDLE DEVENSIAN DEPOSITS OF RIVER THAMES 277
cut out entirely, so that the overlying gravel comes torest directly on that beneath. The two gravel sub-unitsare identical in structure, palaeocurrent direction andcomposition. There is therefore no reason to supposefrom the sedimentary evidence that the clayey silt bedrepresents a major hiatus in deposition, i.e. the entiresequence represents a single aggradational event.
The section is capped by up to 2·6 m of floodplainalluvial deposits that appear to fill a channel, since theythin towards the west. Dark grey silty clay 20 em thickoccurs at the base and rests directly on the gravelbeneath. This is overlain by 1·1 m of brown clayey silt,which is in turn succeeded by 1·3 m of light brown clayeysilt. Beneath the ground surface the clayey silt is weathered and is mottled light grey and brown in colour.
3. GREY CLAYEY SILT BED: COMPOSITIONAND STRUCTURE
Detailed investigation of the fossiliferous grey clayey siltbed in the gravels shows that it represents a channel-fillthat accumulated predominantly under slow flowingwater conditions. The silt is laterally variable in thickness reaching a maximum of 70 em at profile point A,16 em at profile point B, 40 em at profile point C and32 em at profile point D. The composition of the sediments analysed from this unit is shown below using thenotation system of Troels-Smith (1955). Because of thelateral variability and irregular distribution of fossil remains four localities were sampled, the relative positionsof which are shown in Fig. 2.
The sequences are as follows:
Profile Aem
Orange gravel and sand
Depth 0-23 Light grey mottled black clayey silt withisolated pebbles and plant fragments(AsH, Ag2, DI~
23-34 Grey current bedded sand
34-66 Light grey clayey silt with narrow sandbands, with a few shell fragments andplant remains (As1, Ag2!, DI~)
66-70 Brown oxidised clayey silt (As1, Ag3)
Orange gravel and sand
Profile Bem
Orange sand
0-16 Light grey clayey silt with a number oflight grey silty sand band, plant fragmentsat 8-9 em (As1, Ag2, Dll)
Gravelly sand
Profile Cem
Orange sand and gravel
0-15 Light grey clayey silt (As1, Ag3, Dl+)
15-24 Orange gravel and sand much disturbedby loading to form 'ball-and-pillow' structures
24-40 Light grey clayey silt with narrow sandbands and bands of plant and shell fragments 1-2 mm thick (As1, Ag2!, DI!)
Orange gravel and sand
Profile Dem
Orange gravel and sand
0-32 Light grey clayey silt with narrow sandbands disturbed near base by possiblebioturbation. Plant detritus present inisolated band near top of unit 0-5 em.(As1, Ag2, Dll)
Orange gravel and sand
From these sections it emerges that the grey clayeysilt fines upwards becoming slightly more clayey andricher in plant remains in the upper parts. A series ofsand or sandy silt bands ranging from less than 0·5 toover 3 em in thickness subdivide the bed. The bandsconsist of a basal coarse member resting on an erodedbase. This member fines upwards into a grey clayey siltwhich contains an irregular distribution of plant andanimal remains. At least three prominent and sevenminor bands were identified at the northern end of thesection. In places, the sand bands become thicker andcoarser, passing laterally into pebbly sand or fine gravel.One such band reached a maximum thickness of 10 ern(Fig. 2). Where this band consisted of gravel, load casting had occurred resulting from the silt beneath havingbeen injected into the gravel above forming small 'balland-pillow' type structures. Load casting of this typemust have occurred during emplacement of the mainoverlying gravel unit and indicates that the sedimentswere not frozen at the time. The presence of high energysediments in the otherwise low energy sequence indicates that periodic flooding of the channel took placethroughout the time represented. In at least one casewater flowwas rapid enough to deposit gravel and coarsesand. Isolated pebbles occurred in the grey clayey silthorizons either singularly or in groups of two or three. Itis difficult to explain the occurrence of such pebblesunless they were rafted either by river ice or possiblyfloating vegetation and subsequently dropped intothe sediments or they were rolled across the smooth

278 P. L. GIBBARD, G. R. COOPE, A. R. HALL, R. C. PREECE AND J. E. ROBINSON
silt surface as saltation by current flow in shallowwater.
4. PALAEONTOLOGY
Samples of the grey clayey silts from the four localitiesmentioned above were subdivided for palaeobiologicalanalyses. Representative numbers of macroscopic plantremains, mollusc shells, ostracod valves and insect remains were obtained and each of these is discussedbelow. In addition, samples were prepared for palynological investigation , but no contemporary microfossilswere found .
Vertebrate remains have been collected from thegravel and sand beneath the grey clayey silt bed. Severaldecayed, rolled and broken, curved tusk fragments ofpresumed Mammuthus primigenius were found in thegravel by workmen during excavation , and a rolled fragment of an elephant ulna, identified by Dr. A. J. Stuart,was found by PLG and RCP from gravel immediatelyabove the grey clayey silt.
(a) Macroscopic plant remains
Plant macrofossils were extracted from each of sevensample residues. The samples were sieved to O·3 mmand in no case was the volume of residue more than20 cc. The results of the analyses are presented in Table1. Nomenclature used follows Clapham, Tutin and Warburg (1962) except where otherwise stated.
The samples yielded only a limited flora, the richestbeing those from profiles C (with a minimum of25 taxa)and D (29 taxa) . Most taxa were recorded as only one ortwo specimens (or shoots , in the case of mosses) and theassemblages will thus support no more than a tentativeinterpretation. Moreover , within the two profiles forwhich there are sequences of samples (A and D) , thereare no systematic differences observed which might contradict or confirm evidence for changes observed forother fossil groups: consequently, the 7 assemblagesare here considered as a single flora.
As the list shows, there is a mixture of taxa representing aquatic, marsh and better-drained habitats.Apart from leaves of Salix herbacea and two fruits determined respectively as Betula ct. nana and ct. Salix, thereare no woody taxa. The flora, limited though it is, thussuggests a treeless landscape with vegetation dominatedby grasses and sedges with heliophyte herbs such asArmeria, Campanula rotundifolia, Unum perenne agg.,Potentilla anserina, Rhinanthus sp. and Silene maritimavulgaris. A variety of water and marsh plants (such asGroenlandia densa, Potamogeton spp ., Ranunculussubgenus Batrachium, Zannichellia) indicates areas ofslowly flowing water , although with some locally staticwater in places. The upward increase in pond vegetationrecorded by the Ostracoda (section 4c) and suggested bythe sediments is to some extent borne out by the numbers of Carex spp. remains found in the upper part ofprofile A.
With regard to climatic indicators, there are a numberof taxa that are tolerant of 'arctic' conditions and havebeen described from 'arctic' floras by Bell (1970) andothers (subsumed by Godwin, 1975). In particular,Arenaria ciliata agg., Diplotaxis tenuifolia, Unumperenne agg., Rhinanthus sp. , Salix herbacea and Sileneherbacea and Silene maritima/vulgaris have Quaternaryrecords primarily, if not solely, from deposits assigned tocold stages . To these may be added the less securelyidentified Betula cf. nana and Potentilla cf. crantzii-type .
Although there are several Late Devensian cold stagefloras known from deposits in the Thames valley areae.g . Colney Heath, Herts. (Godwin, 1964), Lea Valley(Reid & Chandler, 1923) and Colney Street, Herts.(Gibbard , 1977), the paucity of the present material,and the limited extent to which many fossils have beenidentified , precludes further comparison. There is certainly no possibility of using the flora to date the depositsat Kempton Park, other than to a cold stage .
(b) Molluscs
Five samples of grey clayey silt from profiles A, BandC indicated on Fig. 2 were dried and 0.5 kg fractionswere wet sieved. All fossils retained on a 0·5 mm sievewere extracted and the results are shown in Table 2.Shells were frequent and well-preserved, the periostracum being present on many specimens , while the valvesof several bivalves were still united.
The overall nature of the molluscan assemblagesuggests a fairly clean , well-vegetated stream. There islittle stratigraphical change in the fauna within profileA. Sample C, however, contained considerably moreLymnaea peregra than profile A, while Pisidium amnicum and P. henslowanum, species characteristic of moving water , were absent. This suggests rather slackerflow, although there were no species (e.g. P. obtusale)that would indicate stagnant conditions. The relativeaburidance of the stream plant aroenlandia densa insample C supports this interpretation (Table 1).
The fauna is small, with only ten species of freshwatermollusc. All these species have wide modern rangesextending from southern Europe to the Arctic, so thereare no strictly arctic-alpine species present . The majority of the species represented are fairly catholic in theirecological requirements, although P. amnicum and P.henslowanum often prefer substantial hard-waterstreams. These two species become scarcer northwardsin Scandinavia, but this may be due to the absence ofsuitable habitats rather than resulting from climatic orother factors .
It is noteworthy that the Kempton Park P. henslowanum lack the umbonal appendicula normally characteristic of this species (form inappendiculata , or form nucleus auctt.). Kuiper (1968) figures this form from Pleistocene sediments from central Germany. In the BritishIsles this form is uncommon today in lowland habitats,but becomes more frequent towards the northern mar-
MIDDLE DEVENSIAN DEPOSITS OF RIVER THAMES 279
TABLE 1. Plant macrofossil remains from grey clayey silt, Kempton Park(Sample positions shown in Fig. 2. a, achene; cal, calyx; car, caryopsis; eph, ephippia; fr, fruit; fgt, fragment; n, nutlets;
00, oospore; pyr, pyrene; s, seed; sh, shoot)
A C D
Taxon 70-58 em 58-34 cm 23-0 em 31-28 em 32-22 em 22-12 em 12-D em
cf, Agrostis sp. carArenaria sp. sA. ciliata agg. (sensu Godwin 1975) sArmeria maritima (Mill.) Willd. cal 2Atriplex sp. scf. Bellis perennis L. aBetula cf. nana L. frCampanula rotundifolia L. s 2 1Carex spp. n 2 4 11 18 8 15Chenopodium section Pseudoblitum
(Gren.) Aschers. sCerastium sp. sCyperaceae indet. n 2Diplotaxis tenuifolia (L). DC. s 23Eleocharis palustris s.l. n 2Gramineae indet. carGroenlandia densa (L). Fourr. pyr 28Hippuris vulgaris L. s 2 4 2Leontodon sp. a 1 1Linum perenne agg. (sensu Bell 1970) s 1Linum perenne agg. (sensu Bell 1970) frfgt - 2Myriophyllum spicatum L./M.
verticillatum L. nPlantago cf. caranopus L. frcf. Poa sp. carPolygonurn aviculare agg. frPotamogeton crispus L. var.
macrorhynchos (Grandog) pyr 1P. cf. filiformis Pers. pyrPotamogeton praelongus Wulf. pyrPotamogeton sp(p). indet. pyr 1 1Potentilla anserina L. a 4 8 7 2 2 8 5P. cf. crantzii-type (sensu Bell, 1970) a 1 2 2P. palustris (L.) Scop. a 2Potentilla sp(p). indet. a 2 2 2Ranunculus subgenus Batrachium a 4 1 4 4 2Ranunculus sp. indet. a 1Rhinanthus sp. s 1Salix herbacea L. lvs 12cf. Salix sp. fr 1Saxifraga cf. stellaris L. sSilene maritima With.lS. vulgaris
(Moench) Garcke s 2Taraxacum sp. a 1Zannichellia palustris L. fr 1
Chara 00 1 7 1 3 17 25 3Nitella 00 3 6 17 11 18 1
Mossescf. Amblystegium sp. sht +cf. Barbula sp. sht +Bryum sp(p). shts + + + +d. Campylium sp. sht +cf. Cratoneuron commutatum (Hedw.)
Roth shts + + +C. filicinum (Hedw.) Spruce shts + +cf. Drepanocladus sp(p). shts + +(Daphnia) eph + + +
280 P. L. GIBBARD, G. R. COOPE, A. R. HALL, R. C. PREECE AND J. E. ROBINSON
70-58 58-34 23-0 8-0 31-28
TABLE 2. Mollusc numbers in grey clayey silt, Kempton Park(Sample positions shown in Fig. 2. The numbers forSphaerium and Pisidium refer to individual valves)
em em emFreshwater species em
Valvata piscinalis(Muller) 4
Lymnaea peregra(Muller) 2
Anisus leucostoma(Millet)
Sphaerium corneum(L.) 2
Pisidium amnicum(Muller) 2
Pisidium casertanum(Poli)
Pisidium milium HeldPisidium subtruncatum
MaIm 18Pisidium henslowanum
(Sheppard) forminappendiculataauett.
Pisidium nitidumJenyns 22
Land speciesPupilla muscorum (L.) 2
A
em
9
?1
)[
2
31
65
4
65
B
2 10
55
5 24
3
7
34 6
46 10
c
preference is possible and, in some cases, the limitingphysical factors which determine their occurrence inparticular deposits can clearly be ascertained. Some areactive swimmers well able to flourish in moving water ofappreciable current strength. Other species, however,lack the necessary strong swimming appendages andindicate still, or barely moving, water such as lakes orponds. Yet other species are dependent upon weedgrowth among which they clamber and browse.Although these facts have long been established, it islargely through the summaries of Absolon (1973) andDiebel and Pietrzeniuk (1975, 1977) that ostracodassemblages reflecting small ponds, shallow lakes, deeplakes and streams have been recognised. Broadlycategorised, the following groupings prove useful:
Springs (cool temperatures): Eucypris pigra, Ilyodromus olivaceus
Streams: Potamocypris wolji, Ilyocypris bradyiPonds: Nannocandona [aba, Eucypris virens, Cy
clocypris laevisLakes: Limnocythere inopinata, L. sanctipatricii,
Herpetocypris reptans, Darwinula stevensoni, Cytherissalacustris
On these grounds, the basal sample (70-58 em) ofsample A, suggests a flowing water open environment,
TABLE 3. Ostracod valve numbers in grey clayey silt,Kempton Park
(Samplepositionsshownin Fig. 2. Valvesfrom bulk samples.)
A B C
gins of the species range in northern England, Scotland 70-58 58-34 23-0 8-0 31-28and parts of Ireland. This was the only form present in em em em em eminterstadial deposits of Last Glaciation (Weichselian)age in the Harz Mountains (Kuiper, 1968). It also occurs Candona candida
in deposits of similar age from Lake Geneva (Favre, (0. F. Muller) II 26 6 9 20
1935). It is not clear whether the high frequency of this Candona d. lozekiAbsolon 22 49 II 17 40inappendiculate form in these deposits is due to eco-
Candona protzilogical or climatic factors. Hartwig 2 8
Pupilla muscorum was the only land mollusc re- Cyclocypris laeviscovered. This species indicates open country. Since it (0. F. Muller) 6 6often lives on the bare ground of river floodplains it Cypria ophthalmicaoccurs in stream sediments of all ages. However, it is a (Jurine) 2characteristic fossil in glacial contexts and is frequently Eucypris pigra
the only mollusc present. (Fischer) 3 35Herpetocypris reptans
(Baird) 6 19 40 6(c) Ostracods lIyocypris bradyi Sars 2 2 10
Rich assemblages of ostracods were obtained from thelIyocypris gibba
(Ramdohr) 2same samples which were analysed for molluscs. In addi- lIyodromus olivaceustion a further series of samples from identical levels was (Brady and Norman) 20processed and analysed to 120 mesh (1251J) in order to Limnocythere inopinatarecover the small species. The results are presented in (Baird) 27Table 3. Nannocandona [aba
Because British Pleistocene ostracod species are EkmanPotamocypris wolfi
members of the present day European fauna, which Brehm 15has been well studied, precise recognition of their

MIDDLE DEVENSIAN DEPOSITS OF RIVER THAMES 281
the fauna is dominated by species of Candona (75 percent). There are no species indicative of rich vegetationor still-water conditions. Similar conditions continued inthe middle sample (58-34 ern) where Candona wasjoined by Cypria ophthalmica, Eucypris pigra andllyocypris bradyi. In the uppermost sample (23-0 em)the fauna is dominated by Herpetocypris reptans, whichprefers well-vegetated, still waters such as canals orlakes. For profile A, the faunal evidence would suggestinitially a channel occupied by flowing water which, withtime, saw a reduction in the rate of flow and whichgradually infilled and transformed into a vegetated lake.This interpretation is further strengthened by thescarcity of small instars and small species in the base ofprofile A and their occurrence in the uppermost horizon,where there were no currents capable of sweepingthem away from association with the full-grown adultvalves.
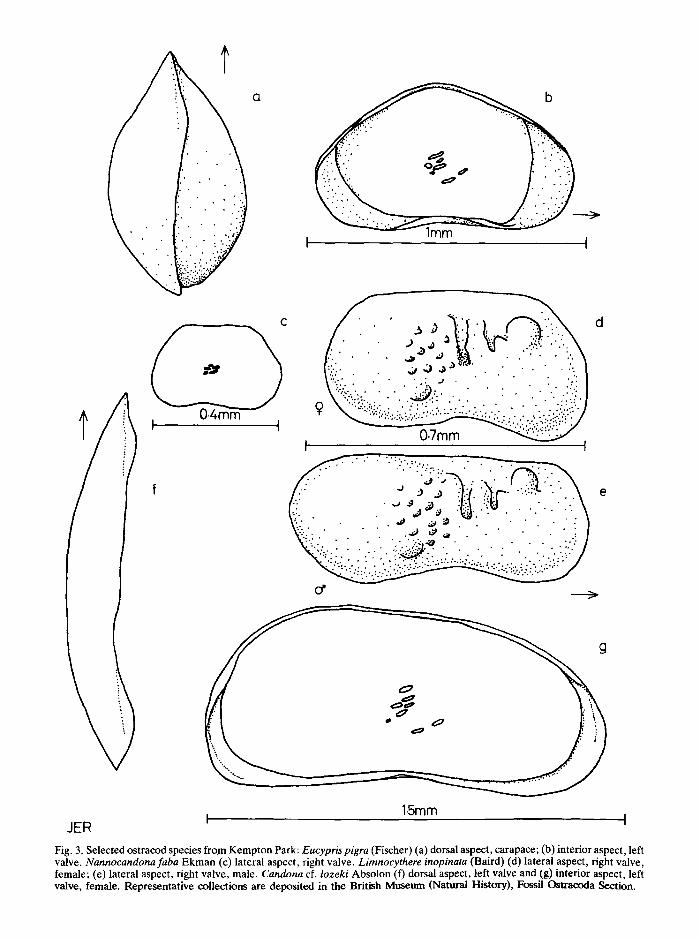
In contrast, sample C, lateral to A, has a fauna dominated by Eucypris pigra and Limnocythere inopinatawhich together with Nannocandona faba and Cyclocypris laevis suggests a quieter marginal environment.At the same time, the scarcity of Herpetocypris, as in thebase of profile A, would indicate an absence of thickweed. Sample B, again lateral to C and A, has a muchsmaller fauna, including Candona ci.lozeki, but also thespring species Eucypris pigra. The association couldrepresent a part of the pool in which a current wasflowing. Four of the species found at Kempton Park areillustrated in Fig. 3.
The presence of C. d. lozeki, a species first describedby Absolon (1973) from Bavaria and Southern Moravia,is its first record in Britain, although it is necessary tocheck with authenticated types to confirm the issue.Candona lozeki was described as a fossil species and isnot known alive today, but its significance as a coldclimate indicator will be appreciated when it is notedthat Absolon (1973, p. 70) described it from deposits ofa Wurm lake, formed by retreat of a glacier. Candona candida is a much more widely recorded species,present in many different settings.
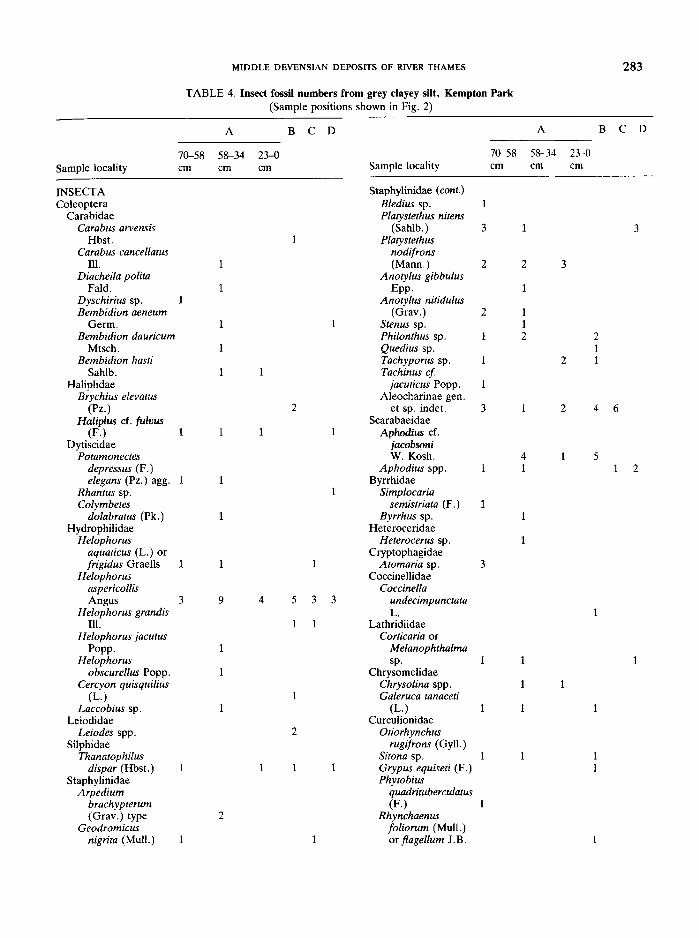
(d) Insects
Six samples of the Kempton Park silt were processedfor insect fossils and the results are shown in Table 4.The figures refer to the minimum numbers of individualsof each species per sample, but because the sampleswere of different sizes, these figures are not a validindication of changing abundance. Because, for technical reasons, the samples had to be processed wet,sample weights are given only to the nearest 0·5 kg.They are as follows: A 70 em~ 58 ern: 1·5 kg, A 58 cm~ 34 em: 1·5 kg, A23 cm~ 0 em: 1 kg, B: 1 kg, C andD: 0·5 kg. Sixty-four taxa have been recognised of whichforty-two can be assigned to species or species groups.Although these must represent a small fraction of thetotal insect fauna of the times, nevertheless they provide
enough data to reconstruct the outlines of thepalaeoecology.
The fossil insect assemblage consists of both aquaticand terrestrial species, the former living close to thesampling site and the latter probably washed in fromfurther afield. Because of the small number of speciesinvolved, it seems safest to treat the local environmentas a whole.
It is easiest to begin with the aquatic larvae of theTrichoptera (caddis flies) which are indicators of thehydrological regime and may thus be compared with theostracods. The most abundant species Anabolia nervosafeeds upon diatoms and filamentous algae in slowly flowing water. Lepneva (1966) gives its habitat as follows'species occurs in slow clear water with a current speedof 0·05-0·02 m/sec, rarely 0·5 m/sec, often in spring-fedbrooks and canals or in more sunlit streams with a summer temperature of 18°-20°C; usually on a bottom ofstones or sand with detritus'. Hiley (1976) points out thatthis species is absent from temporary water bodies. Hydropsyche contubernalis spins nets across the current ofslowly moving water to catch animals or plants. Agrypnia, Apatania and Brachycentrus subnubilus are alsospecies of slowly moving water. Ecclesopteryx guttulataprefers cool streams but the larvae avoid areas of rapidcurrent and live amongst vegetation. The Limnephilusspp. are most typically found in still water amongst thelittoral vegetation. Agrypnia crassicornis is ecologicallyenigmatic today since it lives in both shallow fresh waterlakes (the only British locality is Malham Tarn where it isfound in the wave zone: B. Wilkinson,pers. comm.) andalso brackish water. There is, however, no definitiveindication of saline conditions in the Kempton Park silt.Frequent remains of the aquatic larva of the alder flySialis lutaria group also support this interpretation ofslowly flowing water.
The aquatic coleoptera lead to similar environmentalconclusions. Brychius elevatus is almost exclusively arunning water species (Balfour-Brown, 1940). Haliplusfulvus is most frequently found in large bodies of water;both in lakes and rivers. Potamonectes depressus-elegans is also found in lakes and streams. Colymbetesdolabratus and Rhantus live amongst water weeds in stillor slowly moving water. Species of Helophorus andLaccobius are frequently found in grassy pools. Thediminution in the numbers of aquatic insect species inthe top sample at site A supports the view that thechannel had probably ceased to function as a watercourse by this time.
Along the margins of the channel, mud banks musthave been available for the fossorial beetles Bledius andHeterocerus that are often associated with the predatorDyschirius. The surface of these clay banks would havebeen the habitat of Bembidion aeneum. Bare shingle atthe water's edge is the preferred environment forBembidion hasti where it feeds upon dead or drowninginsects. The staphylinid species Arpedium brachypterumand Geodromicus nigrita feed upon small arthropods in
e
d
O·7mm
a
O·4mm
l5mmJER
Fig. 3. Selected ostracod species from Kempton Park: Eucypris pigra (Fischer) (a) dorsal aspect, carapace; (b) interior aspect, leftvalve. Nannocandona faba Ekman (c) lateral aspect, right valve. Limnocythere inopinata (Baird) (d) lateral aspect, right valve,female; (e) lateral aspect, right valve, male. Candona cf. lozeki Absolon (f) dorsal aspect, left valve and (g) interior aspect, leftvalve, female. Representative collections are deposited in the British Museum (Natural History), Fossil Ostracoda Section.
MIDDLE DEVENSIAN DEPOSITS OF RIVER THAMES 283
TABLE 4. Insect fossil numbers from grey clayey silt, Kempton Park(Sample positions shown in Fig. 2)
A B C D A B C D
70-58 58-34 23-0 70--58 58-34 23--0Sample locality em em em Sample locality em em em
INSECTA Staphylinidae (cont.)Coleoptera Bledius sp.
Carabidae Platystethus nitensCarabus arvensis (Sahib.) 3 3
Hbst. PlatystethusCarabus cancellatus nodifrons
Ill. (Mann.) 2 2 3Diacheila polita Anotylus gibbulus
Fald. Epp.Dyschirius sp. Anotylus nitidulusBembidion aeneum (Grav.) 2 I
Germ. Stenus sp. IBembidion dauricum Philonthus sp. 2 2
Mtseh. Quedius sp. IBembidion hasti Tachyporus sp. 2 I
Sahib. Tachinus cf.Haliplidae jacuticus Popp.
Brychius elevatus Aleoeharinae gen.(Pz.) 2 et sp. indet. 3 2 4 6
Haliplus d. fulvus Searabaeidae(F.) Aphodius d.
Dytiscidae jacobsoniPotamonectes W. Kosh. 4 5
depressus (F.) Aphodius spp. I 2elegans (Pz.) agg. Byrrhidae
Rhantus sp. SimplocariaColymbetes semistriata (F.)
dolabratus (Pk.) Byrrhus sp.Hydrophilidae Heteroeeridae
Helophorus Heterocerus sp.aquaticus (L.) or Cryptophagidaefrigidus Graells Atomaria sp. 3
Helophorus Coeeinellidaeaspericollis CoccinellaAngus 3 9 4 5 3 3 undecimpunctata
Helophorus grandis L.Ill. Lathridiidae
Helophorus jacutus Corticaria orPopp. Melanophthalma
Helophorus sp.obscurellus Popp. Chrysomelidae
Cercyon quisquilius Chrysolina spp.(L.) Galeruca tanaceti
Laccobius sp. (L.)Leiodidae Cureulionidae
Leiodes spp. 2 OtiorhynchusSilphidae rugifrons (Gyll.)
Thanatophilus Sitona sp.dispar (Hbst.) Grypus equiseti (F.)
Staphylinidae PhytobiusArpedium quadrituberculatus
brachypterum (F.)(Grav.) type 2 Rhynchaenus
Geodromicus foliorum (Mull.)nigrita (Mull.) or flagellum J.B.

284 P. L. GIBBARD, G. R. COOPE, A. R. HALL, R. C. PREECE AND 1. E. ROBINSON
damp moss and leaf litter . Further away from the river,sparse meadow-like vegetation must have occurred withclay substrates for Carabus cancellatus, and sand orgravel for Carabus arvensis and Bembidion dauricum.Diacheila polita is a species of open country often withpeaty soils in both moist and dry places.
There are few phytophagous species in this fossil
70-58 58-34 23-0
A
TABLE 4. Continued assemblage . Simplocaria semistriata and Byrrhus feedon various species of moss. Galeruca tanaceti and theweevil Otiorhynchus rugifrons are polyphagons. Grypusequiseti feeds exclusively on Equisetum (horse-tails).Phytobius quadrituberculatus eats various species ofPolygonaceae including Polygonum aviculare whosefruits also occurred in the silt. Rhynchaenus [oliorumand flagellum feed on Salix and are small enough toutilise Salix herbacea.
Certain specialist species of beetle do not fit into theabove categories but add their own pecularities to thegeneral environmental picture. Thus Coccinella undecimpunctata preys specifically upon aphids . Thanatophilus dispar is found under carcasses, often under deadfish. Species of Aphodius are most frequently found inthe dung of large herbivorous mammals, a habitat alsoutilised by the staphylinid beetles Platystethus andTachinus . Anotylus species of the gibbulus group inpresent-day Africa have mostly been found living in thedung of elephants (Hammond, Morgan & Morgan 1979)and it is tempting to see a similar relationship betweenthe Mid Devensian A. gibbulus and the mammothswhose tusks were recorded from various horizons atKempton Park.
It is possible to make some climatic inferences fromthe Kempton Park insect assemblage in spite of itssmallness and the coarseness of the sampling intervals. Ifthe fauna is viewed as a whole it presents a bewilderingmixture of 'northern', 'southern' and even 'asiatic'species. However this confusing picture resolves itself tosome extent when the insect assemblages from eachsample are considered separately. It is then clear that theonly obligatory cold adapted species occur in the uppertwo samples from locality A , though the uppermost ofthese samples is really too meagre to permit furtheranalysis. In the account that follows these species will beconsidered first.
The following species occur only in the upper twosamples from locality A:Diacheila polita: circumpolar in the high north almost
exclusively on the tundra. The nearest occurrence toBritain is the Kola peninsula; also occurs in the highmountains of central Asia.
Bembidion dauricum: almost circumpolar in the highnorth and in the high parts of central Asia. Its nearestlocality to Britain is in the Hardanger mountainswhere it is found in the middle alpine region at 1200 m(Fjelberg, 1972).
Bembidion hasti: circumpolar in the high north . Itsnearest occurrence to Britain is the mountains ofsouthern Norway.
Colymbetes dolabratus : circumpolar, arctic mostly athigh altitudes in Fennoscandia. Its nearest locality toBritain is in the mountains of southern Norway.
Helophorus obscurellus: Eurasia in the high northand on the high cold steppes of central Asia . Itsnearest occurrence to Britain is on the Kaninpeninsula.
+
2
BCD
+
+ +
3 2
++ +
+
+
cm
++
+
+
em
++
4
cmSample locality
Trichoptera (all larvae)Hydropsychidae
HydropsychecontubernalisMcLachlan
PhryganeidaeAgrypnia
crassicornisMcLachlan
Agrypnia sp. 2Limnephilidae
Apatania sp.Ecclesopteryx
guttulata (Pict.)Limnephilus d.
borealis (Zett.)Limnephilus
flavicornis (F.)or stigma Curt.
Limnephilusrhombicus (L.)or politusMcLachlan
Limnephilus spp.Anabolia nervosa
(Curtis) 7ScricostomatidaeBrachycentrus
subnubilus CurtisNeuroptera
SialidaeSialis lutaria L.
(type) +Hymenoptera parasitica
gen . et sp. indet. +Diptera
ChironomidaeChironomus sp. +
BibionidaeDilophus sp. +
Tipulidae indet.Hemiptera-Heteroptera
SaldidaeSalda littoralis (L.)Saldula sp.
CorixidaeCorixa sp.

MIDDLE DEVENSIAN DEPOSITS OF RIVER THAMES 285
Helophorus jacutus: Exclusively asiatic from Taymyr inthe north to the Altai mountains in the south and fromYenisey river in the west to the Lena river in the east(Angus , 1973).
Anotylus gibbulus: Disjunct distribution today, restricted to the Caucasus mountains and a singlelocality north of Vladivostock (a single specimen)(P . Hammond, pers. com.) .The occurrence of these species exclusively in the
upper two samples at locality A leave little doubt thatthe climate at that time was very much colder and morecontinental than it is in Britain today. The inference ofclimatic cont inentality is supported by the exclusivelyasiatic species, not confined to these two samples,namely Helophotus aspericollis from eastern Siberiaand Mongolia (Angus, 1973), Tachinus jacuticus fromnorth east Russia, Central Siberia and North America(Ullrich, 1975) and Aphodius ct. jacobsoni which isknown today from Turkestan and Mongolia .
Relatively southern species (i.e. species that totallyavoid arctic and alpine regions of Europe) occur in allsamples: these include Carabus cancellatus, Brychiuselevatus , Platystethus nitens , Hydropsyche contubernalisand Anabolia nervosa . In the lowest sample from locality A and in sample B, the presence of such species andthe absence of any obligate cold adapted species allsuggests that the climate when these samples were deposited was warm with summers probably as warm aspresent day but winters that may have been rathercolder. These two samples also yielded the egg cocoons ofthe leech Herpobdella, which was very common in thetemperate fossil fauna at Isleworth but which appears tobe absent from the later cold/continental faunas of theUpton Warren Interstadial complex. Samples C and Dhad restricted faunas about which little can be concluded. The fact that the middle sample from locality Acontains both northern and southern species may beexplained if the 24 em of silt spanned the episode ofclimatic deterioration. It is hardly likely that the arbitrary subdivision between two adjacent samples wouldhave precisely coincided with the faunal break. Thereremains the possibility, however, that post-depositionaldisturbance of the silt may have contributed to the mixing at this level ; by animals, by loading or by erosion andredeposition.
5. SUMMARY OF THE PALAEOECOLOGY OFTHE KEMPTON PARK SILTS
On the evidence of both sedimentation and palaeontology we conclude that the grey clayey silt accumulatedin the channel of an active river. Periodic floodingbrought in sand and gravel and may have been responsible for the inclusion in the silts of terrestrial species ofthe flora and fauna. The flow of water in the channel wasnormally slow (less than 0·5 mlsec), but the water wasfor the most part clear and well aerated. There wereboth mud and gravel banks. In places along the water's
edge there were probably small stagnant pools . Thediminution of active aquatic species in the upper parts ofthe silt and the increased abundance of sedges suggestthat the channel became silted up prior to the inundationof the area by the overlying gravel spreads.
Evidence from both insects and ostracods suggeststhat at the start of sedimentation , the climate was by nomeans cold but rapidly deteriorated, becoming cold andcontinental. The date of 35,230± 185 BP for the top ofthe silts and the presence of temperate insect species atthe bottom suggest that the silts accumulated during theperiod of climatic cooling that immediately postdatedthe temperate episode represented by the Isleworthfauna dated at 43,140 + 1520/- 1280BP , namely duringthe early part of the Upton Warren Interstadial Complex.
6. DEFINITION OF THE KEMPTON PARKGRAVEL
The deposits described lie beneath the northern marginof the Floodplain terrace surface as described by Deweyand Bromehead (1921) 'South London' (Fig. 1). A twofold division of the Floodplain terrace downstream ofTwickenham was first proposed by Dewey and Bromehead (1921). Zeuner (1959) suggested that an 'UpperFloodplain' terrace could be distinguished as far upstream as Chertsey and this is confirmed by the presentwork by PLG . It does not seem possible to recognise thisterrace subdivision upstream of Staines at present .
The nomenclature of the deposits underlying the lowlevel terrace surfaces in the Thames Valley is in a state ofsome confusion. The term 'terrace' has been used in twosenses: first to refer to the morphological feature andsecond, the deposits upon which that surface is developed (Frye & Willman, 1962). The second usage isclearly inapplicable when a surface is conventionallymapped with little or no attention to the depositsbeneath. The latter may represent more than oneaggradation and in such cases it is clearly incorrect torefer a surface to a certain geological date obtained frommaterial within the deposits beneath that surface. Asurface, unless exhumed, must post-date, or be contemporaneous with the deposits on which it is developed.Furthermore, modification of a surface at a later datecannot always be excluded.
In view of these problems , it is preferable to definedeposits using standard stratigraphical methods (ef.Hedberg, 1976) and then to restrict the term terrace tothe surface developed on the deposits thus defined . Theterrace surface should ideall y be defined from a typelocality, following morphostratigraphical principles. Inthis way ambiguity will be avoided.
No formal stratigraphical nomenclature for the deposits underlying the Floodplain terraces has ever beenestablished. It is therefore proposed that the successiondescribed here be used as the type for a new lithostratigraphical unit of member status, the Kempton Park
286 P. L. GIBBARD , G. R. COOPE, A. R. HALL , R. C. PREECE AND 1. E. ROBINSON
Gravel. Organic or fine sediment, such as that present atthe type locality, are included in this predominantlygravel and sand member as sub-units or beds, as recommended by Hedberg (1976). Because the lithostratigraphical unit defined here rests upon an eroded andchannelled surface, it is likely that in places chronologically earlier strata exist than those recorded from thetype section , but which for geological reasons should bemapped as the Kempton Park Gravel. In the presentstudy of Middle Thames terrace stratigraphy by PLG ,further definitions of other such units will likewise bepresented.
7. RELATION OF THE KEMPTON PARK GRAVELTO THE TERRACE STRATIGRAPHY
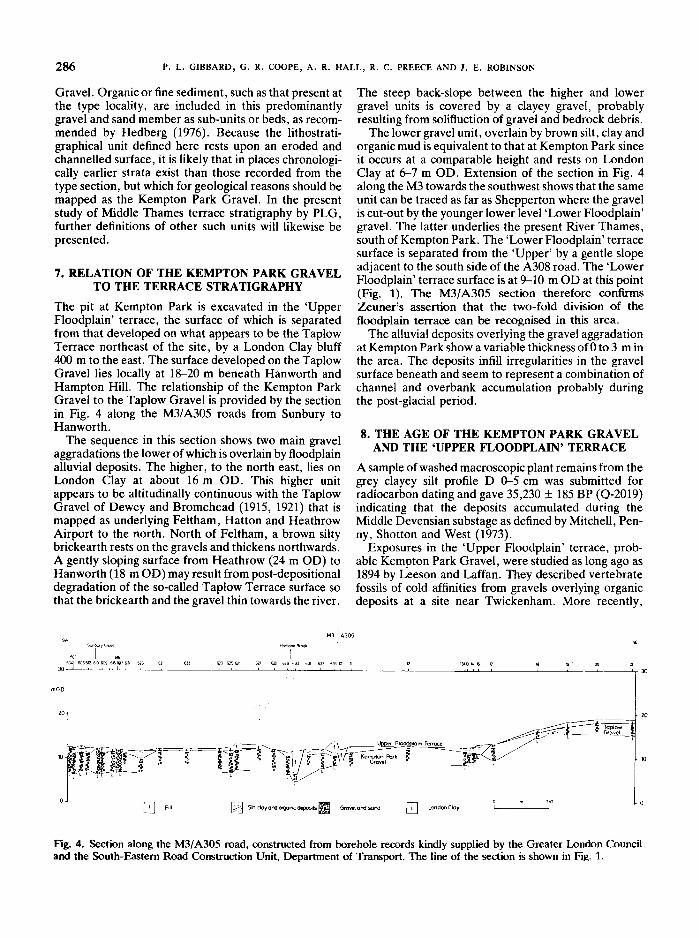
The pit at Kempton Park is excavated in the 'UpperFloodplain' terrace, the surface of which is separatedfrom that developed on what appears to be the TaplowTerrace northeast of the site , by a London Clay bluff400 m to the east. The surface developed on the TaplowGravel lies locally at 18-20 m beneath Hanworth andHampton Hill. The relationship of the Kempton ParkGravel to the Taplow Gravel is provided by the sectionin Fig. 4 along the M3/A305 roads from Sunbury toHanworth.
The sequence in this section shows two main gravelaggradations the lower of which is overlain by floodplainalluvial deposits . The higher , to the north east , lies onLondon Clay at about 16 mOD. This higher unitappears to be altitudinally continuous with the TaplowGravel of Dewey and Bromehead (1915, 1921) that ismapped as underlying Feltham, Hatton and HeathrowAirport to the north . North of Feltham, a brown siltybrickearth rests on the gravels and thickens northwards.A gently sloping surface from Heathrow (24 mOD) toHanworth (18 mOD) may result from post-depositionaldegradation of the so-called Taplow Terrace surface sothat the brickearth and the gravel thin towards the river.
M3] A305
l'o<!i!l<ooI<
I621 6dl 629 t):J 63'1
The steep back-slope between the higher and lowergravel units is covered by a clayey gravel , probablyresulting from solifluction of gravel and bedrock debris.
The lower gravel unit, overlain by brown silt, clay andorganic mud is equivalent to that at Kempton Park sinceit occurs at a comparable height and rests on LondonClay at 6-7 mOD. Extension of the section in Fig. 4along the M3 towards the southwest shows that the sameunit can be traced as far as Shepperton where the gravelis cut-out by the younger lower level 'Lower Floodplain'gravel. The latter underlies the present River Thames,south of Kempton Park . The 'Lower Floodplain' terracesurface is separated from the 'Upper' by a gentle slopeadjacent to the south side of the A308 road. The 'LowerFloodplain ' terrace surface is at 9-10 mOD at this point(Fig. 1). The M3/A305 section therefore confirmsZeuner's assertion that the two-fold division of thefloodplain terrace can be recognised in this area.
The alluvial deposits overlying the gravel aggradationat Kempton Park show a variable thickness of 0 to 3 minthe area. The deposits infill irregularities in the gravelsurface beneath and seem to represent a combination ofchannel and overbank accumulation probably duringthe post-glacial period.
8. THE AGE OF THE KEMPTON PARK GRAVELAND THE 'UPPER FLOODPLAIN ' TERRACE
A sample of washed macroscopic plant remains from thegrey clayey silt profile D 0-5 em was submitted forradiocarbon dating and gave 35,230 ± 185 BP (0 -2019)indicating that the deposits accumulated during theMiddle Devensian substage as defined by Mitchell, Penny, Shotton and West (1973).
Exposures in the 'Upper Floodplain' terrace, probable Kempton Park Gravel , were studied as long ago as1894 by Leeson and Laffan. They described vertebratefossils of cold affinities from gravels overlying organicdeposits at a site near Twickenham. More recently,
10
o london Cloy "",
Fig. 4. Section along the M3/ A305 road, constructed from borehole records kindly supplied by the Greater London Counciland the South-Eastern Road Construction Unit, Department of Transport. The line of the section is shown in Fig. 1.

MIDDLE DEVENSIAN DEPOSITS OF RIVER THAMES 287
Coope and Angus (1975) showed that equivalent organic deposits in also what is almost certainly KemptonPark Gravel at Isleworth date from the Middle Devensian Upton Warren Interstadial as indicated by aradiocarbon date of 43,143+ 1520/- 1280 BP (Birm319) and by the coleopteran fauna. Upstream at Marlow, Buckinghamshire, organic silts from Thamesfloodplain gravels were described by Bell (1968, 1969)who recorded both pollen and plant macrofossils indicating a treeless , herb-dominated environment. Athermophilous insect fauna present (Coope, unpublished data) suggests an early Upton Warren Interstadialage although too little material was available to give afine radiocarbon data (Birm-184: - 31,OOOPB,Shottonand Williams 1971). The present work confirms a MiddleDevensian age for the aggradation of the Kempton ParkGravel. The 'Upper Floodplain' terrace is developeddownstream of Staines on these gravels or upon siltsresting on these gravels.
However, the aggradation of deposits below the'Upper Floodplain' terrace is complex because it includes Ipswichian interglacial sands and sandy gravel atBrentford (Zeuner, 1959) and organic deposits at Trafalgar Square (Franks, Sutcliffe, Kerney & Coope 1958;Franks , 1960). It is not proposed to include these deposts in the Kempton Park Gravel. After a period ofpost-Taplow downcutting, deposits must therefore haveaccumulated during the Ipswichian interglacial. Subsequently , following a second period of downcutting, theKempton Park Gravel must have been deposited overthe Ipswichian sediments leaving the later preserved aseroded remnants. The Kempton Park Gravel aggradation began somewhat before and ended somewhat afterthe Upton Warren Interstadial in the Middle Devensian,thus embracing a number of contrasting sedimentaryenvironments.
The gravels upon which the 'Lower Floodplain' terrace is developed comprise a distinct, completely separate aggradational phase resting on bedrock and abut ting against and in places resting upon eroded remnantsof the Kempton Park Gravels. A period of erosion anddowncutting must have intervened between theaggradation of the two units. Dating of the 'LowerFloodplain' gravel aggradation is still unsatisfactorysince no radiocarbon-datable deposits relating to thisstage have yet been found in the Middle or LowerThames Valley. However, laterally equivalent braidedriver gravel and sand deposits which have been consistently dated to the Late Devensian substage (Reid ,1949, Godwin, 1964; 1975, Shotton & Williams, 1971,Gibbard 1974, 1977; Gibbard & Hall, 1982) occur intributary valleys, for example the Colne and the Lea. Ittherefore appears likely that the 'Lower Floodplain'gravels represent a Late Devensian aggradation distinctfrom the Middle Devensian Kempton Park Gravel. Upstream from Staines the latter are present as erodedremnants beneath these Late Devensian gravels.
It is interesting to note that equivalent Floodplain
gravel deposits from the Upper Thames Valley haveyielded two separate, but comparable series of radiocarbon dates . Both Late Devensian and Middle Devensiandeposits occur , moreover both are associated with coldfaunas (Shotton, 1973; Coope 1976; Briggs, Gilbertson& Goudie, 1976; Briggs & Gilbertson, 1980). It is tentatively suggested that these may be the respective lateralcorrelatives of the 'Lower Floodplain' gravel and theKempton Park Gravel of the Middle Thames Valley asdefined here .
9. CONCLUSIONS
Gravel excavations at Kempton Park revealed a sequence of gravel and sand deposits containing a lens offossiliferous grey clayey silt. The gravels and sands occurin facies typical of braided river deposition, representingthe predominant high energy flow, bed load channel andbar accumulation . Braided river sedimentation characteristically occurs under a strongly seasonal climate suchas is found under cold, possibly periglacial climaticconditions. The grey clayey silt represents a minor flowchannel fill. Here fine suspended-load sedimentation ,interrupted periodically by flood-deposited sand andgravel accumulation occurred.
The grey clayey silt bed contained plant macrofossilsindicative of a regional herb-dominated vegetation withareas of water supporting varied communities of waterand marsh plants. A restricted molluscan assemblagesuggests that the channel was clean and well vegetatedwith slight flow. Treeless vegetation around the rivershown by the plant macrofossil assemblages is supportedby the recovery of Pupilla muscorum, Furthermore, theostracod assemblages also indicate that the channel contained flowing water , the effect of which decreased withtime. Silting-up of the channel was accompanied by anincrease in aquatic vegetation. Evidence from the insectassemblages supports this interpretation. Local mudbanks , gravel surfaces and sparse, meadow-like vegetation was also present in the vicinity.
The insect and ostracod faunas indicate that the greyclayey silt accumulated during a period of climatic deterioration from an initially warm to a later cold continental climate.
The River Thames deposits at Kempton Park exposedbeneath the 'Upper Floodplain' terrace have beentermed the Kempton Park Gravel. The relation of thismember to the neighbouring higher member, probablythe Taplow Gravel and the lower 'Lower Floodplain 'gravel is demonstrated. Laterally equivalent deposits ofthe Kempton Park Gravel containing a vertebrate faunaof cold aspect occur at Twickenham and Isleworth . Atthe latter, an organic bed in the gravels has been shownby radiocarbon dating and coleopteran fauna to datefrom the Upton Warren Interstadial (Coope and Angus1975). Eroded remnants of the Kempton Park Gravelmay be present beneath 'Lower Floodplain' gravel upstream, for example at Marlow . Aggradation of the

288 P. L. GIBBARD , G. R. COOPE, A. R. HALL , R. C. PREECE AND 1. E. ROBINSON
Kempton Park Gravel seems therefore to have begunsomewhat before the Upton Warren event and continued until somewhat after i.e . post-35,OOO BP.Ipswichian interglacial deposits also present beneath the'U pper Floodplain' terrace surface occur as eroded remnants. The higher Taplow Gravel must pre-date theIpswichian in view of its higher position. Conversely , the'Lower Floodplain' gravel that underlies the presentRiver Thames and abutts against eroded remnants of thehigher Kempton Park Gravel, is probably of Late Devensian age .
ACKNOWLEDGEMENTS
The authors wish to thank Dr. V. R. Switsur for theradiocarbon date, the workmen at Kempton Park gravelpit , Dr. A. J. Stuart for identification of the bone fragment , Ms. B. Wilkinson for identification of theTrichoptera fossils and especially Professor R. G. WestF .R .S. for encouragement and for reading the manuscript the original of which was typed by Ms. A. Jennison.
The work was carried out whilst one of us (PLG) wassupported by a N.E.R.C. research associateship which isgratefully acknowledged.
References
ABSOLON, A. 1973. Ostracoden aus einigen Profilen spatund postglazialer Karbonatablagerungen in Mitteleuropa.Milt . Bayer Staatssamml. Palaont. hist . Geol., 13, 47-94.
ANGUS, R. B. 1973. Pleistocene HeloplwlUS (Coleoptera,Hydrophilidae) from Borislav and Starunia in the WesternUkraine with a reinterpretation of M. Lomnicki's species ,description of a new Siberian species and comparisons withBrit ish Weichselian fauna s. Phil . Trans . R . Soc . Land. B.,265, 299-326 .
BALFOUR BROWNE, F. 1940. British Water Beetles Vol. 1.Ray . Soc., London.
BELL, F. G. 1968. Weichselian glacial floras in Britain . Ph.D.thesis , University of Cambridge .
--. 1969. The occurrence of southern , steppe and halophyteelements in Weichselian (last glacial) floras from southernBritain. New Phytol. , 68, 913-22.
--. 1970. Late Pleistocene floras from Earith, Huntingdonshire. Phil. Trans. R . Soc . Lond. , 8258, 347-78 .
BRIGGS, D . J . & D. D. GILBERTSON. 1980. Quaternaryprocesses and environments in the upper Thames valley.Trans . Inst. Brit. Geogr. , 5, 53-65.
- , GILBERTSON, D. D . & GOUDIE, A. S. 1976. TheOxford Region. In: Field guide to the Oxford region .Quaternary Research Association . 1-16 .
CLAPHAM,A. R., T. G. TUTIN &E. F. WARBURG.1962.Flora of the British Isles. University Press: Cambridge.
COOPE, G . R. 1976. Assemblages of fossil Coleoptera fromterraces of the Upper Thames near Oxford. In: Field Guideto the Oxford Region. Quaternary Research Association,20-3 .
--& R. B. ANGUS. 1975. An ecological study of a temperate interlude in the middle of the Last Glaciation, based onfossil Coleoptera from Isleworth, Middlesex. J. Anim. Eco!.44,365-91.
DEWEY, H. & C. E . N. BROMEHEAD. 1915. The geologyof the country around Windsor and Chertsey. Mem. geol.Surv. U.K.
-- &--. 1921. The geology of South London. Mem . geol.Surv. U.K.
DIEBEL, K. & E . PIETRZENIUK. 1975. Mittel- und jungpleistozane Ostracodenfaune des Raums PotsdamBrandenburg-Rathenow in stratigraphischer und okologischer Sieht . Z geol. WISS. Berlin., 3, 1197-233.
FAVRE, J. 1935 . Histoire malacologique du lac de Geneve,Mem. Soc. Phys. et Hist. nat. Geneve ., 41, 295-414 .
FJELBERG, A. 1972. Coleoptera from Hardangervidda.
Fauna of the Hardangervidda No. I , 1-74 ZoologicalMuseum, University of Bergen.
FRANKS, J. W. 1960. Interglacial deposits at TrafalgarSquare, London. New Phytol., 59, 145-52 .
-, A. J. SUTCLIFFE, M. P. KERNEY & G . R. COOPE.1958. Haunt of the elephant and rhinoceros : the TrafalgarSquare of 100,000 years ago-new discoveries . IllustratedLondon News , 14 June, 1011-13.
FRYE, J . C. & H. B. WILLMAN. 1962. Morphostratigraphicunits in Pleistocene stratigraphy. Bull . Am. Ass. Petrol.CeDI., 46, 112-13.
GIBBARD, P. L. 1974. Pleistocene stratigraphy and vegetational history of Hertfordshire. Ph.D. thesis. University ofCambridge .
--. 1977. Pleistocene history of the Vale of St. Albans. Phil.Trans. R. Soc. Lond., 8280, 445-83.
-- & A. R. HALL. 1982. Late Devensian river deposits inthe Lower Colne valley , West London. Proc. Geol. Ass . , 93,291-9.
GODWIN , H. 1964. Late Weichselian conditions in southeastern Britain: organic deposits at Colney Heath , Hertfordshire. Proc. R. Soc. Lond., 8160, 258-75.
--. 1975. History of the British Flora. University Press :Cambridge .
HAMMOND, P., A. MORGAN & A. V. MORGAN. 1979.On the gibbulus Group of Anotylus and fossil occurrence ofAnorylus gibbulus (Staphylinidae) Systematic Entomalogy,4,215-21.
HEDBERG , H. B. 1976. Intemational stratigraphic guide.John Wiley and Sons: New York.
HILEY, D. D. 1976. The identification of British Limnephilidlarvac. Trichoptera. Systematic Entomology, 1, 147-67.
KUIPER, J . G . J. 1968. Die spiitpleistoziinen Pisidien desehemaligen Ascherslebener Sees. Arch. Molluskenk. , 98,23-38 .
LEESON, J . R. & G . B. LAFFAN. 1894. On the geologyof the Pleistocene deposits in the valley of the Thames atTwickenham. Q. JI. geol. Soc. Lond., 50, 453-62.
LEPNEVA, S. G . 1966. Fauna of the U.S.S.R. TrichopteraVol. 11 Number 2 Larvae and Pupae of Integripatpia. IsraelPrograms for Scientific Translations , Jerusalem 1970.
MITCHELL, C. F. , L. F. PENNY, F. W. SHOrrON & R. G .WEST. 1973. A correlation of Quaternary deposits in the
British Isles. Geol. Soc. Lond. Special Report , 4.REID , E. M. 1949. The late-glacial flora of the Lea Valley .
New Phytol., 48, 245-52.

MIDDLE DEVENSIAN DEPOSITS OF RIVER THAMES 289
REID, E. M. & CHANDLER, M. E. J. 1923. TheBarrowell (Lea Valley) arctic flora. Q . 11. geol. Soc. Lond.,29,604-5.
RUST, B. R. 1972. Structure and process in a braided river.Sedimentology 18, 221-45.
SHOrrON , F. W. 1973. English Midlands. In: A correlationof Quaternary deposits in the British Isles. (ed. G. F.Mitchell, et al.) Geol. Soc. Lond . Special Report, 4.
-- 1977. Chronology, climate and marine record. The Devensian stage: its development, limits and substages. In: Thechanging environmental conditions in Great Britain and Ireland during the Devensian (Last) cold stage . (Eds . G. F.Mitchell & R. G. West) Phil Trans . R. Soc . Lond., 8280,107-18 .
-- & R. E. G. WILLIAMS. 1971. Birmingham Universityradiocarbon dates V. Radiocarbon, 13, 141-56.
-- & --. 1973. Birmingham University radiocarbon datesVII . Radiocarbon , 15,451--68.
TROELS·SMITH, J. 1955. Karakterising of lese jordarter.Danmarks Geol. Undersegelse . IV Raekke 3, No. 10.
ULLRICH, W. G. 1975. Monographie det Gattung TachinusGravenhurst (Coleoptera, Staphylinidae) mit Bemerkungenzur phylogenie und verbreitung der arten . Doctoral thesis.Christian-Albrecht, University, Keil.
WILLIAMS , P. F. & B. R. RUST . 1969. The sedimentology ofa braided river . JI. Sedim . Petrol., 39, 649-79.
ZEUNER, F. E. 1959. The Pleistocene period. Hutchinson:London .





























