Embed Size (px)
Citation preview

Micromechanical study ofprotein-DNA interactions
and chromosome structure
Assemble and disassemble protein-DNA complexes, observe in real time, control by force
1. DNA looping by E. coli nucleoid protein Fis
2. Micromanipulation of mitotic chromosomes to probe higher-order chromosome structure
(3. Chromatin assembly using Xenopus egg extractsTheoretical model for experiment: Ranjith Padinhateeri)
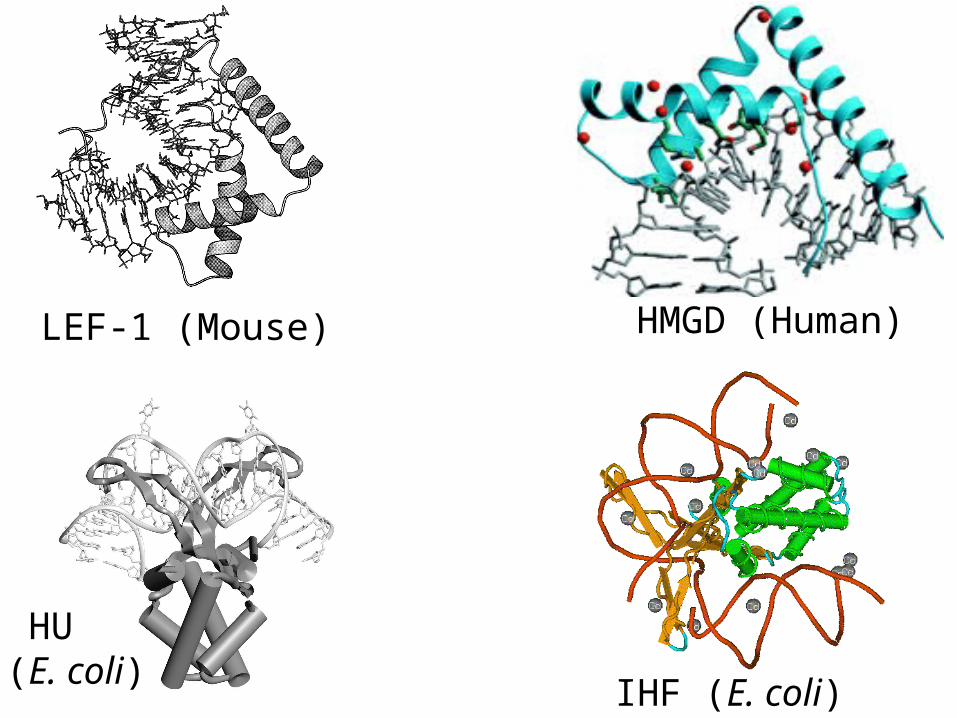
HMGD (Human)
HU(E. coli)
LEF-1 (Mouse)
IHF (E. coli)

Micromechanical study of protein-DNA interactions
naked DNA
+ sharp bends
Study the action of these proteins via mechanicalresponse of the DNA they are binding to
+ loops

E. coli is filled with DNA
E. coli chromosome = 4.5 Mb = 1.5 mm Nucleoid volume < 1 m3
Concentration of DNA ~ 10 mg/mlDNA is covered with protein
Wang, Possoz, Sherratt Genes Dev 2005 E. coli phase contrast
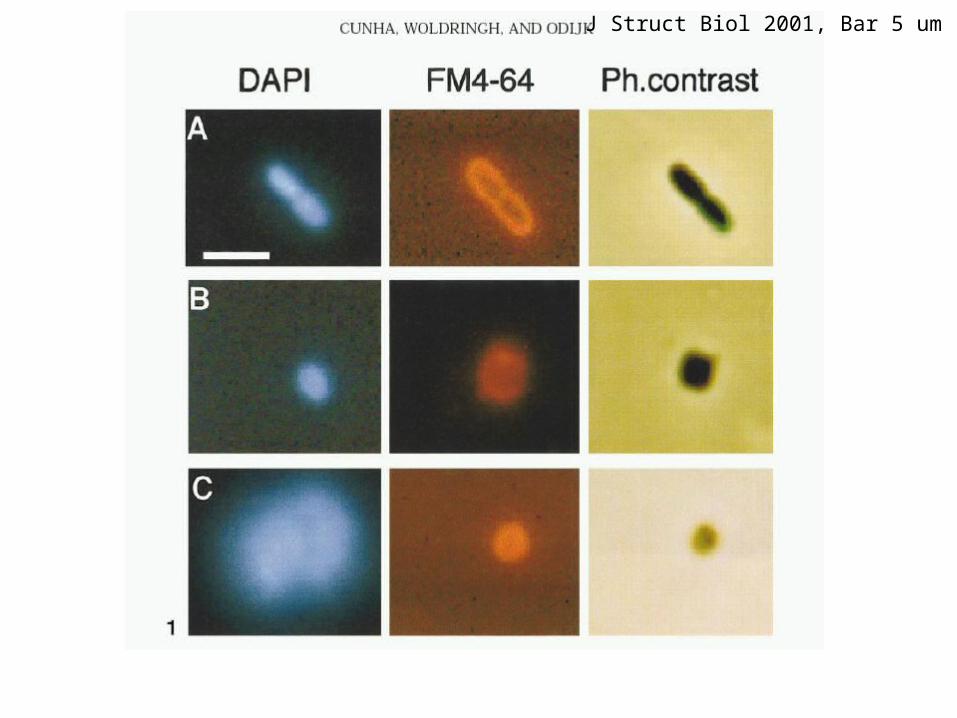
J Struct Biol 2001, Bar 5 um

E. coli loop domains: classical view
50-500 loops
Each loop10 to 100 kb3 to 30 m
+supercoiling
Loop anchors?

Postow, Hardy, Asuaga, Cozzarelli, Genes Dev 2004
Visualization ofsmall E. coli loop
domains
500 nm
100 nm

Transverse magnetic tweezer 97004 bp 32.8 m dimer of
•built on inverted microscope stage•micropipette holds left 2.8 m bead•right bead under tension applied by magnet off to right•wide range of forces (0.1 to 100 pN)•high position sensitivity (< 10 nm) Jie Yan, Ph.D. 05•Relatively easily combined w/ fluorescence John Graham, Ph.D.
Mag
net
(20
0 m
aw
ay)
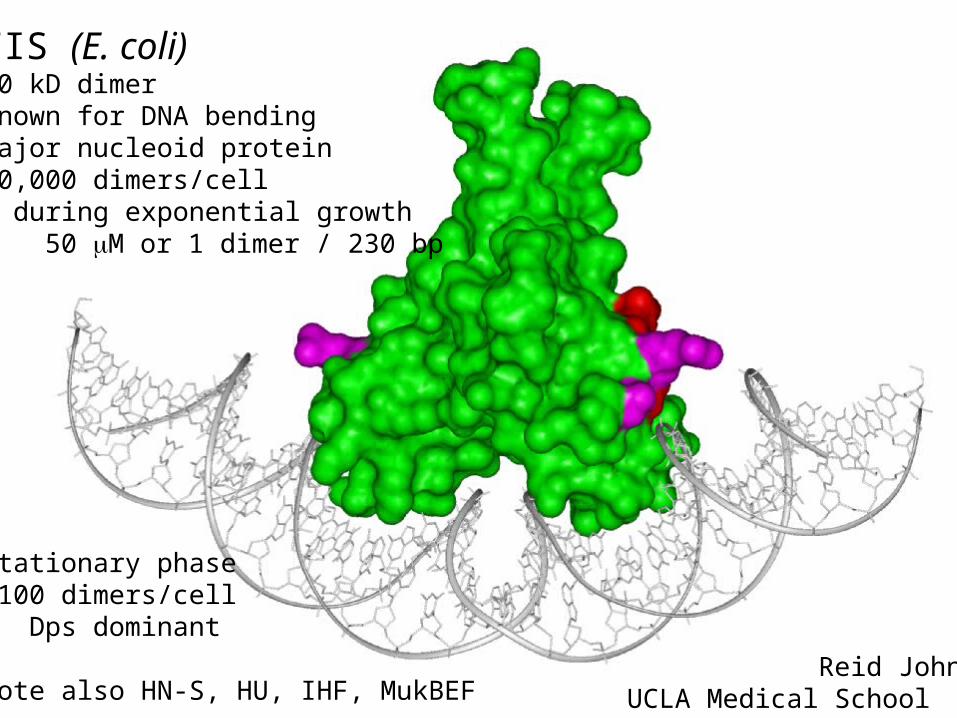
FIS (E. coli)20 kD dimerknown for DNA bendingmajor nucleoid protein40,000 dimers/cell during exponential growth 50 M or 1 dimer / 230 bp
Stationary phase~100 dimers/cell Dps dominant
note also HN-S, HU, IHF, MukBEF Reid JohnsonUCLA Medical School

Schneider et al NAR 2001
FIS observed to increase branching of scDNA binds to DNA in clusters
Fis has been suggested to generate “higher-order” DNA folding

0 nM1 M
6 M
13 M
≥1 uM FIS condenses DNA completely below a low critical force
200 nM

Hypothesis: FIS can stabilize DNA loops
Free energy cost vs binding (free) energyF = F0() + f -
Minimum F0 ~ +5 kBT for ~ 80 nm
Loop formation rate exp( – f / kBT )
Looping requires f < 10 kBT / ~ 0.5 pN
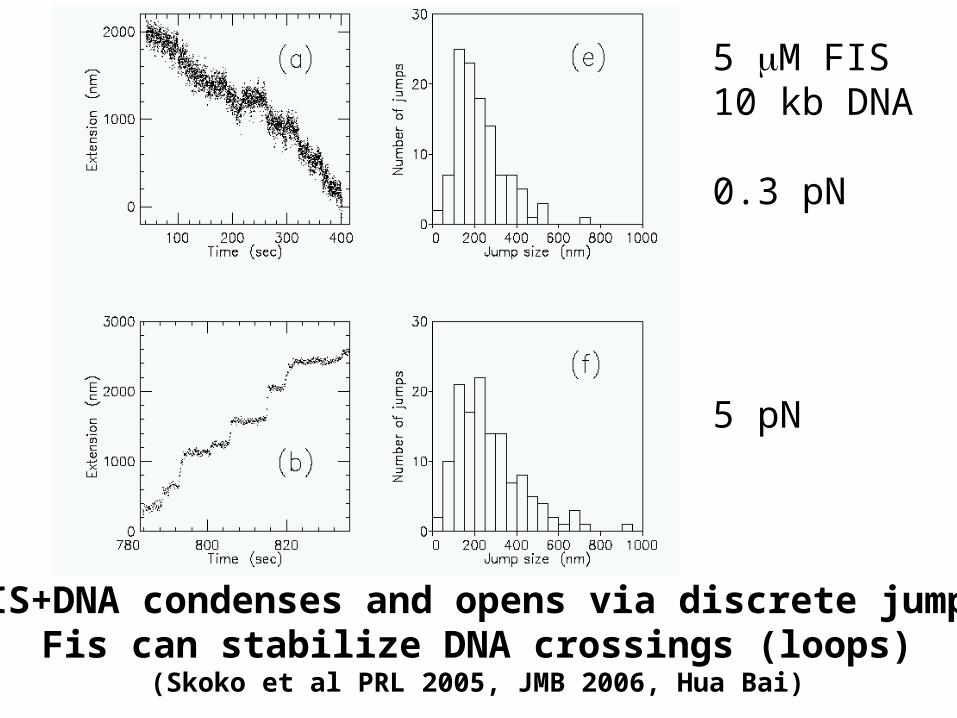
FIS+DNA condenses and opens via discrete jumpsFis can stabilize DNA crossings (loops)
(Skoko et al PRL 2005, JMB 2006, Hua Bai)
5 M FIS10 kb DNA
0.3 pN
5 pN

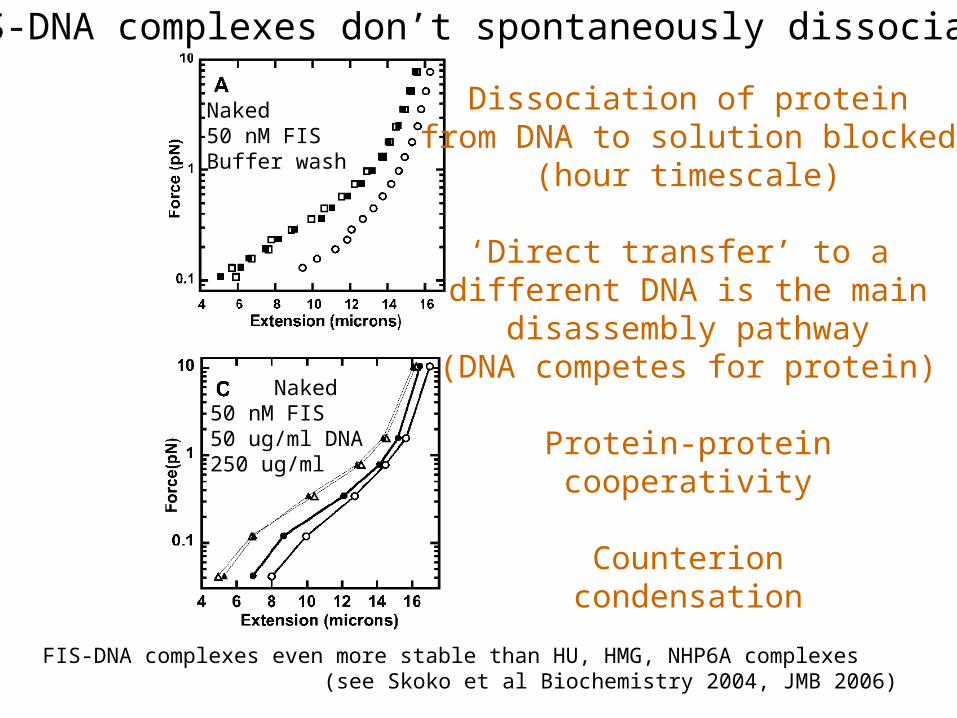
FIS-DNA complexes don’t spontaneously dissociate
Naked50 nM FISBuffer wash
Naked5 uM FISBuffer washThen reduce f
Naked50 nM FIS50 ug/ml DNA250 ug/ml
Naked5 uM FIS50 ug/ml DNAThen reduce f
FIS-DNA complexes even more stable than HU, HMG, NHP6A complexes (see Skoko et al Biochemistry 2004, JMB 2006)
Dissociation of proteinfrom DNA to solution blocked
(hour timescale)
‘Direct transfer’ to a different DNA is the main
disassembly pathway(DNA competes for protein)
Protein-proteincooperativity
Counterioncondensation


http://www.npwrc.usgs.gov/ narcam/idguide/rsnewt.htm

Metaphase
Bar: 20 m

Micromanipulation of individual mitotic chromosomes

stretch

Mitotic chromosomes are elastic “springs”


‘Microspraying’ - kinetic biochemical studiese.g. shift local ionic conditions
(100 mM MgCl2 in culture buffer)
Surprisingly reversible chromosome ‘breathing’ using ionsPoirier et al, J Cellular Biochem 2002
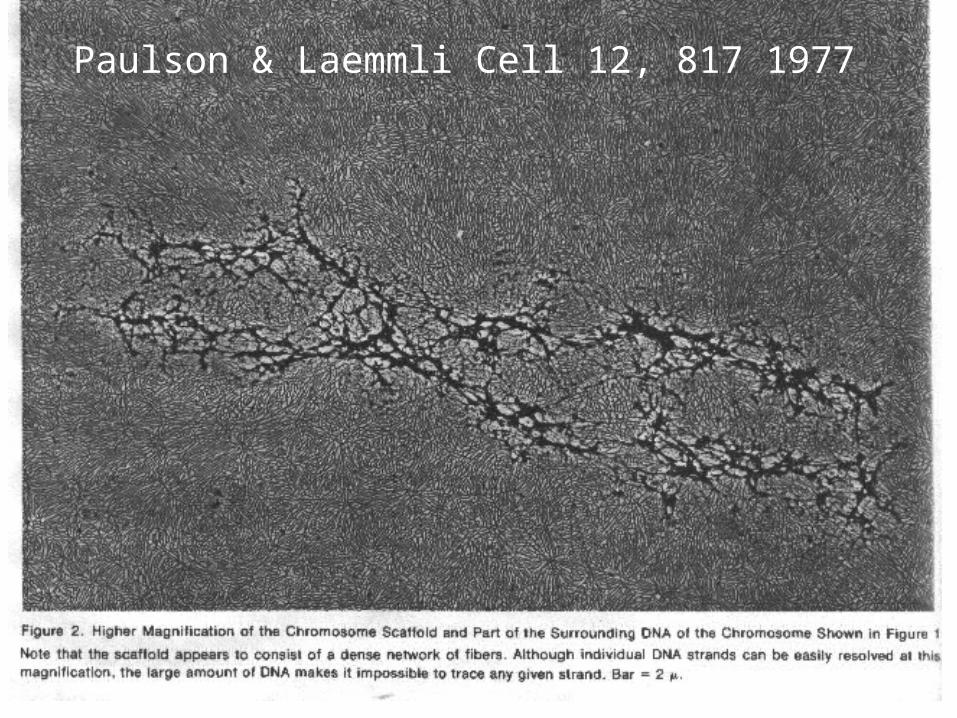
Paulson & Laemmli Cell 12, 817 1977

Stack & AndersonChromosome Res. 9, 175 (2001)

What happens when we cut DNA only?MC nuclease digestion, 0.1 nN initial tension

Cut DNA in mitotic chromosome sufficiently frequently and it falls apart;
classical ‘protein scaffold’ must be disconnected(1 nM AluI AG^CT, PNAS 2002)
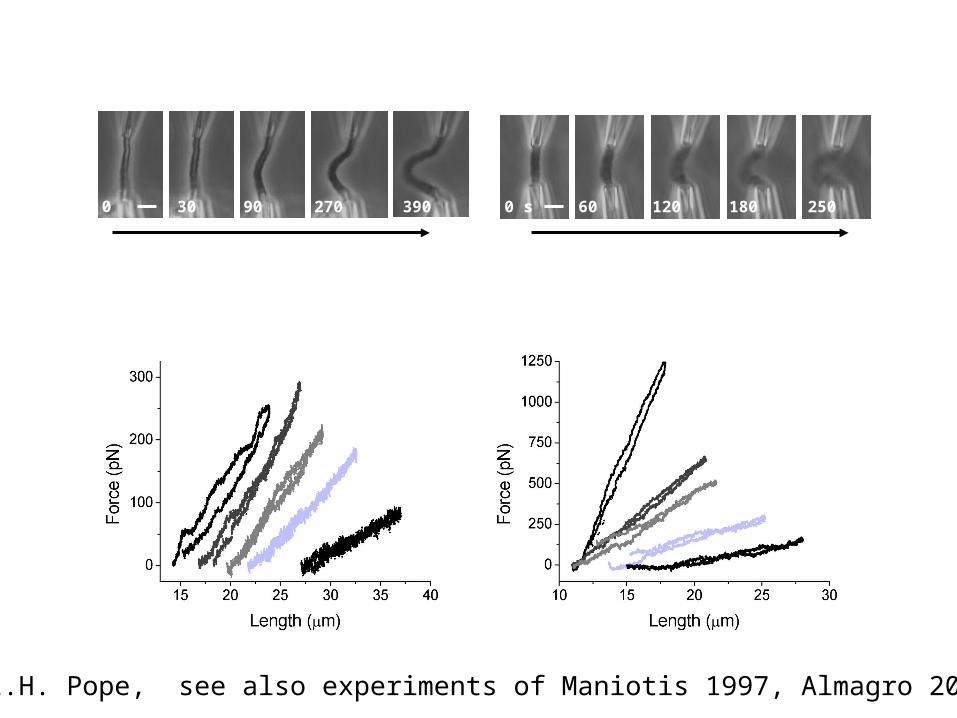
0 s 30 90 390270 0 s 60 120 180 250
c
a b
Increasing trypsin digestion Increasing proteinase K digestion
Proteolysis reduces but does not eliminate elastic response
0 s30
90
270
390
100 nM trypsin
0 s
3060
90
120
500 nM proteinase Kd
L.H. Pope, see also experiments of Maniotis 1997, Almagro 2004

Proteolysis leads to a strong swelling of the mitotic chromosome but never breaks or dissolves it
Chromosome still elastic with well-defined shape after >30 min proteolysis
0 s30
60
120 240
480840
1320
Enhanced contrast
1320
Extensive proteinase K digestion

30 nmchromatin fiber
Mitotic Chromosome
linker protein (SMCs ?) approximately 15 kb spacing
Pope: Effects of protease (MBC 2006)Kawamura: Topo II and DNA entanglements (2007)
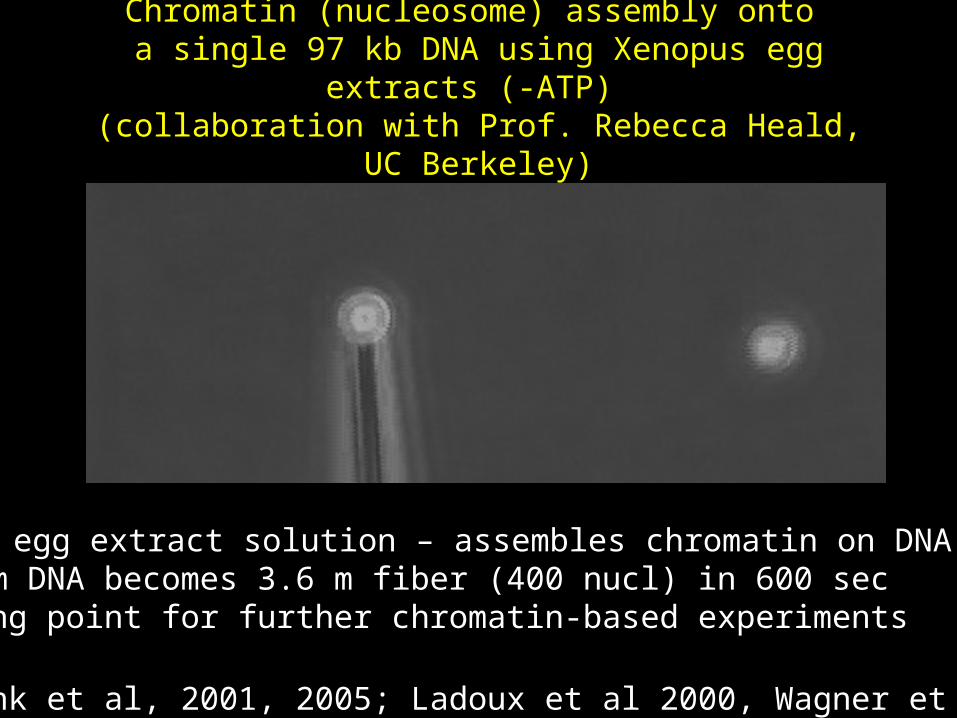
Chromatin (nucleosome) assembly onto a single 97 kb DNA using Xenopus egg extracts (-ATP) (collaboration with Prof. Rebecca Heald, UC Berkeley)
•Xenopus egg extract solution – assembles chromatin on DNA•32.8 um DNA becomes 3.6 m fiber (400 nucl) in 600 sec•Starting point for further chromatin-based experiments
(Bennink et al, 2001, 2005; Ladoux et al 2000, Wagner et al 2005)

UCLAReid Johnson
NYUDavid RothBernhard Schnurr
UCBRebecca Heald
UICMike Poirier (OSU)Dunja Skoko (NIH)Jie Yan (NUS)Lisa Pope (Cambridge)Hua BaiRyo Kawamura
NULance Min
NSF DMR BIO PHYNIH (via NYU)
Northwestern University





























