Embed Size (px)
Citation preview

Plant Cell Physiol. 39(11): 1119-1126 (1998)JSPP © 1998
Mini Review
Metabolic Engineering to Modify Flower Color
Yoshikazu Tanaka, Shinzo Tsuda and Takaaki KusumiInstitute for Fundamental Research, Suntory Ltd., 1-1-1 Wakayamadai, Shimamoto-cho, Mishima-gun, Osaka, 618-8503 Japan
Thanks to the rapid progress in molecular biology offlavonoid biosynthesis and plant transformation, it hasbecome feasible to modify the pathway and flower colorthrough genetic engineering. One of the advantages ofmolecular breeding is that flower color can be specificallymodified without changing the other characteristics of thetargeted variety. Novel flower color varieties such as brick-red petunias and violet carnations have been successfullymade by expression of heterologous flavonoid genes. Flavo-noid metabolic engineering has and will give new perspec-tives in plant molecular biology besides its industrial ap-plication.
Key words: Anthocyanin — Flavonoid — Flower color —Metabolic engineering — Transgenic plants.
Flower colors are mainly produced by flavonoids,carotenoids and betalains. Carotenoids are largely responsi-ble for the production of yellow and orange flowers suchas sun flower and tomato (Bartley and Scolnik 1995).Betalains are yellow to red nitrogenous compounds derivedfrom tyrosine and are distributed only in Caryophyllales(Stafford 1994). Flavonoids have a wide range of colorsfrom pale yellow to red, purple and blue (Goto 1987, Gotoand Kondo 1991). The flavonoid biosynthetic pathway hasbeen the choice to genetically engineer flower color to ob-tain novel colors. Such engineering has also helped us to un-derstand transgene expression in plants. This review sum-marizes the achievements made in engineering flavonoidbiosynthesis to modify flower color. The field has beenreviewed several times (Mol et al. 1995, Davies andSchwinn 1997, Tanaka et al. 1998).
Flavonoids and flower color—Anthocyanins are a col-ored class of flavonoids and accumulate in vacuoles. Thereare six major anthocyanidins (chromophores of anthocya-nins); pelargonidin, cyanidin, peonidin (3' O-methyl cya-nidin), delphinidin, petunidin (3' O-methy delphinidin) andmalvidin (3',5' O-dimethy delphinidin). As the number ofhydroxyl groups in the B-ring is increased, the visible ab-sorption maximum is shifted longer. The maxima in 0.01%
Abbreviations: CaMV, cauliflower mosaic virus; cv.,cultivar.
HCl-methanol solution of pelargonidin, cyanidin anddelphinidin are 520, 535 and 546 nm, respectively (Goto1987).
Modification of the anthocyanidins mainly with glyco-sylation and acylation results in a wide variety of antho-cyanins. Hundreds of anthocyanins have been purifiedand their structures determined (Strack and Wray 1994).Besides the structures of anthocyanins, changes of vacuo-lar pH, intermolecular stackings (self-association of antho-cyanins and co-pigmentation of anthocyanins with poly-phenols), intramolecular stacking of aromatically modifiedanthocyanins, metal complexation and cell shapes givealmost infinite flower colors (Goto 1987, Goto and Kondo1991, Brouillard and Dangles 1994, Mol et al. 1998). Antho-cyanins are red and stable at a low pH. They are bluer butoften unstable at a vacuolar pH which is usually weaklyacidic. The stackings and metal complexation contribute tostabilize anthocyanins and flower color, especially blue col-or (Goto 1987, Goto and Kondo 1991, Brouillard andDangles 1994). The major role of anthocyanin productionin petals is to attract pollinators. Flavonoids and anthocya-nins also play important roles in photoprotection, repro-duction, pathogenesis and symbiosis (Shirley 1996).
Flavonoid biosynthesis—The flavonoid biosyntheticpathway shown in figure 1 has been extensively reviewed(Heller and Forkmann 1994, Forkmann 1994, Holton andCornish 1995). The pathway leading to anthocyanidin 3-glucosides is generally conserved among plant species andis well understood. Anthocyanidin 3-glucosides are furthermodified with sugars and aliphatic and aromatic acids.There are both species and variety differences in the extentof modification and the types of glycosyl and acyl groups at-tached (Fig. 1).
The first enzyme committed to flavonoid production ischalcone synthase (CHS). This enzyme catalyzes the step-wise condensation of three acetate units starting frommalonyl-CoA with p-coumaroyl-CoA to yield 4,2',4',6-tetrahydroxychalcone. CHS shares homology with somecondensing enzymes such as arabidopsis FA El and /?-ketoacyl-acyl carrier protein synthase (James et al. 1995).Flavanone 3-hydroxylase (F3H), a kind of 2-oxoglutaratedependent dioxygenases, catalyzes hydroxylation of narin-genin to dihydrokaempferol (DHK). DHK is hydroxylat-ed to dihydroquercetin (DHQ) and to dihydromyricetin(DHM) by flavonoid 3-hydroxylase (F3'H) and flavonoid
1119
Dow
nloaded from https://academ
ic.oup.com/pcp/article-abstract/39/11/1119/1861440 by guest on 18 N
ovember 2018

1120 Metabolic engineering to modify flower color
COOH COS-CoA
PAL
OH OH " • OHPhenylalanine Cinnamic acid 4-Coumaric acid 4-Coumaroyl-CoA 4-Caffeoyl-CoA
3 x Malonyj^CoA
JCHS
OH 04,2',4',6'-Tetrahydroxychalcone
1 CHI
OHOH O
KaempferolFLS
FNS Hl
oNaringenin
F3H
OH ODihydroauercetin
IDFR, ANS, 3GT" PH
-OH
OHOH O
Dihydrokaempferol
OHOH b
Dihydromyricetin
I DFR, ANS, 3GT
OH
DFR, ANS, 3GT
OGIcOH
Cyanidin 3-Glucoside
OGIc-MalonateOH
A chrysanthemumanthocyanin
OGIcOH
Delphinidin 3-Glucoside
OGIc
R2
Rose anthocyanins
R1 = H or OH or OCH3OH R2 = OH or OGIc
HO.
OGIc
OCH3I
OH
OGIc-Caf
A gentian anthocyanin(gentiodelphin)
OCH3
Carnation anthocyanins
R1 = H or OHR2 = OH or OGIc
OGIc
A petunia anthocyanin
Fig. 1 Generalized pathway leading to anthocyanidin 3-glucosides (Holton and Cornish 1995). Anthocyanins of non-blue flowers(rose, chrysanthemum and carnation) and blue flowers (gentian and petunia) are also shown. Blue flowers usually have delphinidin modi-fied with one or more aromatic acyl group(s) such as caffeic acid (Caf) and coumaric acid. PAL, phenylalanine ammonia-lyase; C4H,cinnamate4-hydroxylase; 4CL, 4-coumarate:CoA ligase; CHS, chalcone synthase; CHI, chalcone isomerase; F3H, flavanone 3-hydroxy-lase; Fill, flavonoid-3'-hydroxylase; F3'5"H, flavonoid 3\5'-hydroxylase; DFR, dihydroflavonol 4-reductase; ANS, anthocyanidin syn-thase; 3GT, flavonoid 3-glucosyl transferase; CC3H, 4-coumaroyl-CoA 3-hydroxylase; FNS, flavone synthase; FLS, flavonol synthase;Glc, glucose; Rha, rhamnose.
Dow
nloaded from https://academ
ic.oup.com/pcp/article-abstract/39/11/1119/1861440 by guest on 18 N
ovember 2018

Metabolic engineering to modify flower color 1121
3',5'-hydroxylase (F3'5'H), respectively (Fig. 1). These twohydroxylases belong to the same cytochrome P-450 family(CYP 75, Holton et al. 1993a, Brugliera et al. 1997). Bothare key enzymes in determining flower color because theydetermine the hydroxylation patterns of the B-ring of thedihydroflavonols and this hydroxylation eventually deter-mines the structure of the anthocyanidin leading to flowercolor (Holton and Tanaka 1994). F3'5"H can also convertDHQ to DHM. Dihydroflavonol 4-reductase (DFR) cata-lyzes the reduction of dihydroflavonols to leucoanthocya-nidins and is proposed to belong to the 3/?-hydroxysteroiddehydrogenase/DFR superfamily (Baker and Blasco 1992).Leucoanthocyanidins are converted to anthocyanidins byanother 2-oxoglutarate-dependent dioxygenase, anthocya-nidin synthase (ANS), and then to the anthocyanidin 3-glu-cosides by UDP-glucose: flavonoid 3-O-glucosyltransferase(3GT). UDP-glucose: anthocyanin 5-O-glucosyltransferase(5GT) genes have been recently cloned (Yamazaki et al.1998). Both 3GT and 5GT belong to the distinct families ofglycosyltransferase superfamily. Petunia UDP-rhamnose:anthocyanin rhamnosyltransferase also has homology toglycosyltransferases (Brugliera et al. 1994, Kroon et al.1994). Genes of aromatic acyl transferase (AAT) catalyzingthe transfer of aromatic group to 3- or 5-glucose of antho-cyanins have been cloned (Tanaka et al. 1997). Many ofthese enzymes have been biochemically characterized by ex-pressing cloned cDNAs in heterologous systems (Holton etal. 1993b, Tanaka et al. 1996, 1997, Yamazaki et al. 1998).Peonidin-type anthocyanins are synthesized from the cya-nidin-type anthocyanins and petunidin- and malvidin-typeanthocyanins are from the delphinidin-type anthocyaninsby the transfer of a methyl group from S-adenosylmethio-nine catalyzed by anthocyanin methyltransferases (Hellerand Forkmann 1994).
Anthocyanins are reversibly modified with glutathione(Marrs et al. 1995, Alfenito et al. 1998) and postulated tobe transported into vacuoles by ATP-binding cassette(ABC) transporters (Lu et al. 1997). A 24 kDa vacuolar pro-tein has been suggested to be involved in intravacuolar pig-mented structures in sweet potato (Nozue et al. 1997).These processes are not clearly understood.
Flavonols are derived from dihydroflavonols by a2-oxoglutarate-dependent dioxygenase, flavonol synthase(FLS, Holton et al. 1993b). Apigenin (a flavone) is synthe-sized from naringenin by flavone synthase (FNS). Interest-ingly there are two kinds of FNS, a cytochrome P-450 anda dioxygenase (Heller and Forkmann 1994).
Regulation of flavonoid biosynthetic pathway—Spa-tial and/or temporal expressions of structural genes orenzymes of flavonoid biosynthesis in flowers, leaves andseedlings have been well studied in many plants such aspetunia (Brugliera et al. 1994), snapdragon (Jackson et al.1992), gerbera (Helariutta et al. 1993), carnation (Stich etal. 1992), rose (Tanaka et al. 1995), eggplant (Toguri et al.
1993), Arabidopsis (Pelletier et al. 1997), grape (Boss et al.1996), perilla (Gong et al. 1997) and lisianthus (Nielsenand Podivinsky 1997). Transcription of these genes is spati-ally and developmentally regulated in a well-coordinatedway paralleling flavonoid synthesis.
In maize, two gene families, R (Myc type) and Cl(Myb type) encoding transcriptional factors, regulate theexpression of the structural genes in the pathway. Snap-dragon and petunia have similar transcriptional factorsthat regulate the structural genes (Holton and Cornish1995, Mol et al. 1998). Regulatory genes controlling an-thocyanin biosynthesis are functionally conserved amongplant species and some of them function in heterologousspecies. They have distinct sets of target genes (Quattroccioet al. 1993, Martin 1996), which allows regulatory diversityin the pathway depending on species. In petunia flowers,Anl (a Myc gene), An2 (a Myb gene) and Anil are regula-tory genes of anthocyanin biosynthesis in the flower. Anilencodes a novel Trp-Asp repeat protein and is a componentof a signal transduction cascade that modulates An2 func-tion (de Vetten et al. 1997).
Molecular tools for flower color modification—Thefirst step in metabolic engineering is to clone the structuralgenes. Cloning structural genes in the flavonoid pathway isnot difficult because all but a few structural genes in thepathway (Fig. 1) have been cloned. The homologous genescan be isolated by the hybridization approach at least fromclosely related species. This approach is very effective forconserved genes such as CHS and is less effective forvariable genes such as CHI (Gutterson 1995). In our ex-perience, most structural genes can be readily cloned bylow stringent screening with heterologous counterpartgenes within dicots. For less conserved genes such as CHIand sometimes DFR, a polymerase chain reaction ap-proach using conserved amino acid sequences has been suc-cessfully used (Gutterson 1995, Tanaka et al. 1996). Clonedgenes in the pathway from various flowers are listed(Davies and Schwinn 1997).
Constitutive promoters including cawliflower mosaicvirus (CaMV) 35S promoter, an enhanced CaMV 35S pro-moter (Mistuhara et al. 1996) and Mac promoter (Comai etal. 1990) have been successfully used to express genes inpetals. Constitutive expression of the structural genes doesnot have deleterious effect on the transgenic plant. Promot-ers derived from genes in the flavonoid pathway should beuseful for temporal and spatial expression.
Suppression of a gene is more challenging than expres-sion of a gene in transgenic plants. Plant gene expressioncan be suppressed by introducing either an antisense (Molet al. 1990) or sense gene of interest or its close homologs.The mechanism of sense suppression (cosuppression) is notyet fully understood (Taylor 1997, Gallie 1998) althoughthe phenomenon is a post-transcriptional event. Double-stranded RNA is proposed to be a mediator in sequence-
Dow
nloaded from https://academ
ic.oup.com/pcp/article-abstract/39/11/1119/1861440 by guest on 18 N
ovember 2018

1122 Metabolic engineering to modify flower color
specific genetic silencing and cosuppression (Montgomeryand Fire 1998). The frequency and degree of cosuppressiondepend on transgene promoter strength and an intact fulllength cDNA may be preferable for efficient suppression(Que et al. 1997) although it is not essential to use fulllength cDNA to obtain phenotypes of antisense or sensesuppression (Gutterson 1995). The frequency and degreeof antisense and sense suppression also depend on thetargeted gene and species. The frequency varies from lessthan 1% to 40% in our laboratory.
Transformation of floricultural crops—Plant cellshave can detect the transgenes in the genome and suppressthe expression unless the transgenes are integrated in "cor-rect positions" (Kumpatla et al. 1998, Matzke and Matzke1998, Gallie 1998). It is, however, not possible to integratea transgene into the correct position artificially. It isnecessary to obtain many (dozens to hundreds) transgenicplants and select a few transgenic lines whose phenotypesare desirable and stable enough to be commercialized. Ther-fore, a highly efficient transformation system for the tar-geted species or variety is essential. Interestingly, a highfrequency of shutdown of transgene expression duringgrowth is often observed in chrysanthemum (Deroles et al.1997).
Transformation of floricultural crops has been review-ed (Deroles et al. 1997, Tanaka et al. 1998). Rose, carna-tion, tulip and chrysanthemum that occupy more than 50%of the cut flower market are claimed to be routinely trans-formed (Mol et al. 1995) but there is much variation in ex-tent of success among cultivars within a species. Problemsand strategies of plant transformation have already beendiscussed (Birch 1997).
Plant transformation generally consists of three steps.The first step is to transfer foreign DNA into plant cells (in-fection), the second step is to select transgenic cells (selec-tion) and the third step is to regenerate them into completeplants (regeneration). Each step needs careful optimizationof various procedures and parameters.
To deliver foreign DNA into plant cells, Agrobacte-/•/wm-mediated gene transfer has been widely used especial-ly for dicots and some monocots. Particle bombardment isoften used for monocots resistant to Agrobacterium(Deroles et al. 1997).
For selection, the neomycin phosophotransferase genethat confers kanamycin resistance has been most widelyand successfully used as a selection marker. Herbicide-re-sistant genes encoding enzymes which are insensitive to theherbicide or detoxify it can be used as selectable markers. Itis important to select only the cells with the introducedtransgenes. Weak selection often results in chimerical pro-blems.
Most dicot species regenerate well via organogenesison media containing balanced hormone supplementation,usually cytokinin and auxin. An efficient transformation
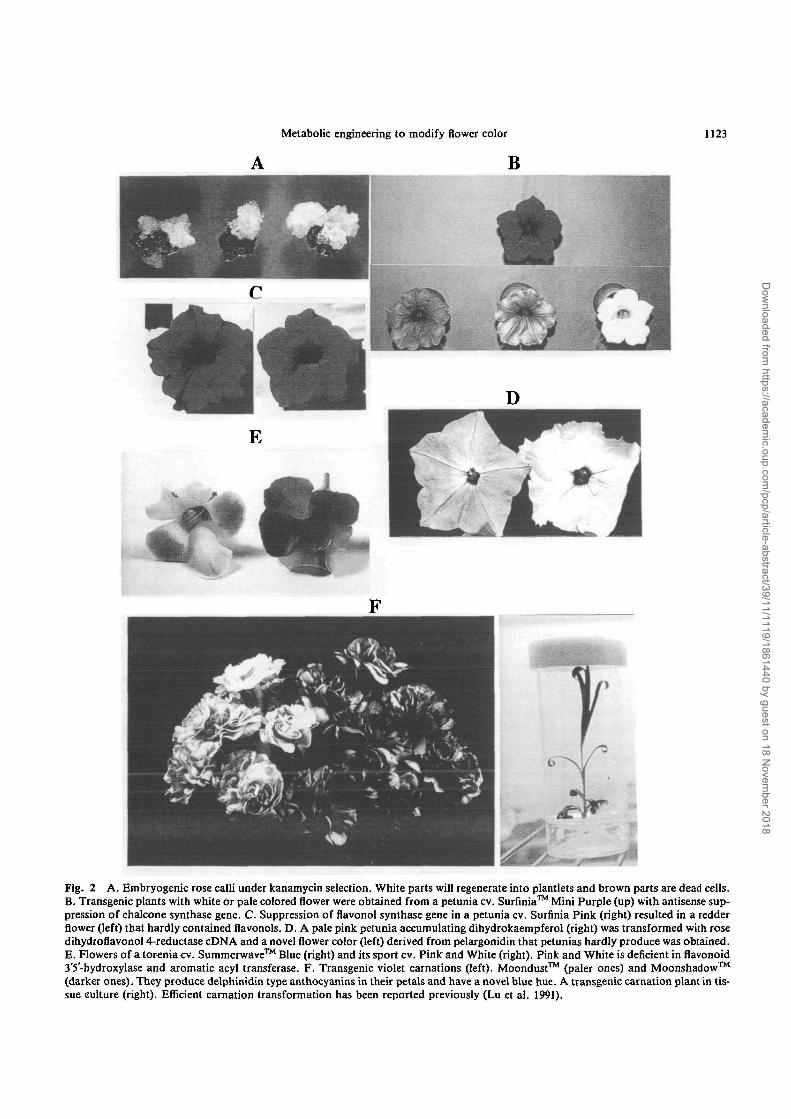
system has been established for petunias and torenias and50 to 100 independent transgenic plants per gene constructare routinely obtained in our laboratory. Embryogenesis isthe preferred route to transform roses in our laboratory(Fig. 2A, Katsumoto et al. 1995). A rose cultivar (cv.)Royalty was also transformed via embryogenic callus(Firoozababy et al. 1994).
Modification of anthocyanin amount—White is not anovel color because of abundant occurrence. Still, molecu-lar breeding of a white variety is industrially useful becauseonly flower color can be modified without sacrificing theother desirable characteristics. Such a sacrifice is oftenmade during hybridization breeding. It is possible to obtainwhite flowers from anthocyanin producing flowers by sup-pressing the expression of one of many structural or regula-tory genes in the pathway. The reported results are summa-rized in Table 1. It is interesting that novel color patternswere obtained in petunia and lisianthus while only uniformsuppression was observed in rose, gerbera, and chrysan-themum. This may be relevant to the fact that petunia andlisianthus have natural white-sectored patterns (Deroles etal. 1998).
Suntory Ltd. markets Surfinia™ (a petunia) and Sum-merwave™ (a torenia) that are superior to conventionalpetunias and torenias with their creeping and vigorous char-acters. We have been working on widening their flower col-or variation by metabolic engineering. Antisense CHS-A ina petunia cv. Surfinia Mini Purple resulted in a few plantswith pale color or pure white flowers (Katsumoto et al. un-published results, Figure 2B). White and blue/white (twoout of four petals are white and the other two petals areblue) transgenic torenia plants were successfully generatedfrom a blue torenia cv. Summerwave Blue by sense sup-pression of CHS or DFR genes (Suzuki et al. 1997). In bothcases the transgenic plants retain the superior characteris-tics of the host and only flower color was modified.
The amount of anthocyanins was increased using thetranscriptional factors of the pathway. The flower color ofthe tobacco was changed from pink to intense red by ex-pressing the maize R gene (Lc allele) with CaMV 35S pro-moter. Cl driven by the same promoter had no effect.Arabidopsis root, petal and stamen accumulate anthocya-nins by constitutive expression of both Cl and R (Lloyd etal. (1992)). Transgenic petunia plants that had a higheranthocyanin content in floral and vegetative tissues were ob-tained by constitutive expression of Lc. Leaves of the trans-genic petunia are purple due to accumulation of anthocya-nins (Bradley et al. 1998). Delila (an R homolog controllingflavonoid synthesis in snapdragon) increased anthocyaninsin tobacco flower and tomato flowers and vegetative tissues(Mooney et al. 1995).
Because dihydroflavonols are the common precursorsof anthocyanins and colorless flavonols, DFR and FLScompete with each other for dihydroflavonols. Antisense
Dow
nloaded from https://academ
ic.oup.com/pcp/article-abstract/39/11/1119/1861440 by guest on 18 N
ovember 2018
Metabolic engineering to modify flower color 1123
B
Fig. 2 A. Embryogenic rose calli under kanamycin selection. White parts will regenerate into plantlets and brown parts are dead cells.B. Transgenic plants with white or pale colored flower were obtained from a petunia cv. Surfinia™ Mini Purple (up) with antisense sup-pression of chalcone synthase gene. C. Suppression of flavonol synthase gene in a petunia cv. Surfinia Pink (right) resulted in a redderflower (left) that hardly contained flavonols. D. A pale pink petunia accumulating dihydrokaempferol (right) was transformed with rosedihydroflavonol 4-reductase cDNA and a novel flower color (left) derived from pelargonidin that petunias hardly produce was obtained.E. Flowers of a torenia cv. Summerwave™ Blue (right) and its sport cv. Pinlc and White (right). Pink and White is deficient in flavonoid3'5'-hydroxylase and aromatic acyl transferase. F. Transgenic violet carnations (left). Moondust™ (paler ones) and Moonshadow™(darker ones). They produce delphinidin type anthocyanins in their petals and have a novel blue hue. A transgenic carnation plant in tis-sue culture (right). Efficient carnation transformation has been reported previously (Lu et al. 1991).
Dow
nloaded from https://academ
ic.oup.com/pcp/article-abstract/39/11/1119/1861440 by guest on 18 N
ovember 2018

1124 Metabolic engineering to modify flower color
Table 1 Suppression of flavonoid pathway
Host species (colour) Gene construct Phenotype Reference
Petunia (purple)Petunia (violet)Petunia (purple)Chrysanthemum (pink)Gerbera (red)Rose (red)Carnation (pink)Torenia (blue)Torenia (blue)
Carnation (red)Lisianthus (purple)
Antisense CHS-ASense CHS-ASense CHS-ASense CHSAntisense CHS, DFRSense CHSSense CHSSense CHS, DFRSense CHS, DFRAntisense CHS, DFRAntisense F3HAntisense CHS
White, patternWhite, patternWhite, patternWhite, uniformPink, uniformPink, uniformPale pink, uniformWhite, patternPale blue, pattern
White, uniformWhite, pattern
van der Krol et al. (1988)Napoli et al. (1990)van der Krol et al. (1990)Courtney-Gutterson et al. (1994)Elomaa et al. (1993)Gutterson (1995)Gutterson (1995)Suzuki et al. (1997)Aida et al. (1997)
Zucker et al. (1998)Deroles et al. (1998)
Phenotypes of transgenic plants depend on each transgenic event. The strongest phenotype of each report is shown. Some species show-ed uniform suppression and some formed novel color patterns.
suppression of the FLS gene in petunia and tobaccoresulted in a higher content of anthocyanin and more in-tense flower color (Holton et al. 1993b). A similar resultwas obtained with sense suppression of FLS gene in apetunia cv. Surfinia Pink. Flower color changed from pinkto red purple and the transgenic plant flower containedmore anthocyanins and less flavonol than the host (Fig. 2C.Suzuki et al., unpublished results).
Making brick-red flowers—Petunia flowers rarely con-tain pelargonidin-type anthocyanin and are not brick-redbecause DHK can not be reduced by petunia DFR that hashigh activity for DHM (Forkmann and Ruhau 1987) andis suiable for delphinidin production. Transgenic brick-red petunias accumulating pelargonidin-type anthocyaninshave been obtained from DHK accumulating petunia (defi-cient in F3'5H, F3"H and FLS) with using maize, gerberaand rose DFR cDNAs (Meyer et al. 1987, Helariutta et al.1993, Tanaka et al. 1995 (Fig. 2D), respectively). Stabilityof the phenotype and the transgene of the petunias contain-ing maize DFR cDNA has been extensively studied (Linn etal. 1990. Meyer and Heidmann 1994). The gerbera DFRgene showed a stronger and more consistent expressionthan maize DFR. The choice of the gene source is a factorto be considered for engineering plants even if the gene en-codes the same enzyme activity (Elomaa et al. 1995). Whenthe rose DFR gene was introduced to a petunia cv. SurfiniaPurple that did not accumulate DHK due to its dominancein F3"H and FS'S'H genes was transformed, no flower colorchanges were observed (unpublished results). This indicatesthat it is difficult for a transgene to compete with endoge-nous genes.
Molecular breeding of blue flowers—Structures of an-thocyanin, presence of co-pigments and relatively high vac-uolar pH (more than about 5.0, Holton and Tanaka 1994)
are key factors for flowers to have a bluish hue (Goto andKondo 1991, Holton and Tanaka 1994, Mol et al. 1998).
Most blue flowers contain aromatically acylated delphi-nidin derivatives (Fig. 1). The absorbance maximum of an-thocyanin shifts towards longer wavelengths (bluer color)by about 10 nm with one hydroxylation of B-ring, and byabout 4 nm with one aromatic acylation in \% trifluoroacet-ic acid solution (Fujiwara et al. 1997). Figure 2E shows theeffect of hydroxylation of the B-ring and aromatic acyla-tion of anthocyanins to flower color. Blue petals of atorenia cv. Summerwave Blue mainly contain malvidin5-(p-coumaroyl)-glucoside 3-glucoside while petals of itspink counterpart contain peonidin 3,5-diglucoside. This in-dicates that F3'5'H and AAT activities are necessary for theproduction of blue flowers in vivo. There is little or nochange in color of anthocyanins with glycosylation andaliphatic acylation. Rose, chrysanthemum and carnationonly have pelargonidin- and cyanidin-type anthocyaninsnot modified with aromatic acyl groups.
Genes encoding F3'5'H have been isolated frompetunia (Holton et al. 1993a), eggplant (Toguri et al. 1993),gentian (Tanaka et al. 1996), lisianthus (Nielsen andPodivinsky 1997) and many others (unpublished results).Petunia F3'5'H cDNAs could complement their deficiencyin petunia that was also deficient in F3'H and low in pH(see below). The flower color changed from pale pink toreddish purple (Holton et al. 1993a).
Florigene Ltd, (Australia) and Suntory Ltd. successful-ly developed transgenic violet carnations by introductionof a petunia F3'5'H and DFR genes into a DFR-deficientwhite carnation (Holton et al., unpublished results). Thepetals of the carnations predominantly contain delphi-nidin, which is not produced by the native carnations. Suchbluish flowers have not been produced by traditional
Dow
nloaded from https://academ
ic.oup.com/pcp/article-abstract/39/11/1119/1861440 by guest on 18 N
ovember 2018

Metabolic engineering to modify flower color 1125
breeding of carnation (Fig. 2F). The transgenic violet carna-tions named Moondust™ have been marketed in Australiaand Japan and became the first transgenic floricultural cropto be sold. Darker versions, Moonshadow™, have been ob-tained and will be marketed soon. Application of AATgenes (Tanaka et al. 1997) to modify flower color is inter-esting.
When anthocyanins make a complex with polyphenolssuch as flavonol and flavone glycosides (co-pigmentation),their absorption is increased by up to about 35 nm in wave-length (bathocromic shift, Goto 1987). Co-pigmentationcan also result in a large increase in absorptivity. Theamount of flavonols can be modified by genetic engineeringof FLS genes (Holton et al. 1993b). Although intramolecu-lar stacking of anthocyanins may be achieved by expressionof multiple AAT and GT genes in heterologous plants,only some of the genes have been cloned.
Anthocyanins are bluer at a higher pH (Goto 1987,Goto and Kondo 1991). Even an peonidin derivative canmake blue flowers as in blue petals of morning glory whenit is highly modified with six glucose and three caffeic acidmolecules and its vacuolar pH is high, 7.7 (Yoshida et al.1995). Because non-blue flowers, however, tend to have alower vacuolar pH, it is important to increase the pH toproduce blue flowers. Petunia is known to have seven locicontrolling vacuolar pH (phl-phl, Mol et al. 1998). Whenone of them is homozygous recessive, the vacuolar pH in-creases in the petal. Only Ph6 has been cloned (Chuck et al.1993). Anl and Ph6 turned out to be alleles of the samelocus (Mol et al. 1998). Mutations in An2 and Anil alsoaffect pH (Mol et al. 1998). Cloning of Ph genes and theirfunctional analysis may be useful to elevate flower vacuolarpH (Mol et al. 1998). However, it is not certain if pH regula-tion by petunia Ph genes is applicable to other species. Fur-ther biochemical studies on the mechanism of pH controlin vacuoles or bluing phenomena of senescent flowers oftenobserved in some species may help us to engineer vacuolarpH.
The three dimensional structure of Commelina bluepigment has been determined. The complex is made of sixanthocyanins (malonylawobanin), six flavones (flavocom-melin) and two magnesium ions (Kondo et al. 1992). Thebiochemical process of formation of the complex is yet tobe revealed. A specific co-pigment and a metal ion arenecessary for a specific anthocyanin to form a stable metal-loanthocyanin.
Tulip, impatiens, cyclamen and pelargonium producedelphinidin but lack true blue colors (Davies and Schwinn1997). A species-specific strategy is required to obtain bluevarieties.
Many genes must be introduced and properly express-ed to synthesize highly modified anthocyanins or formmetal complexes and/or elevate vacuolar pH in order tomake truly blue flowers from non-blue flowers. This is a
very challenging task at the moment and substantial im-provement of plant biotechnology is essential. Furtherchemical and biological understanding of flower color andflavonoid pathway together with improved plant transfor-mation systems should make modification of flower colormore feasible.
The authors thank Florigene Ltd. for providing the photo ofFigure 2 and permission of presenting unpublished results.
References
Aida, R., Kishimoto, S., Tanaka, Y. and Shibata, M. (1997) J. Japan. Soc.Hort. Aci. 66 suppl. 2: 486-487.
Alfenito, M.R., Souer, E., Goodman, C D . , Buell, R., Mol, J., Koes, R.and Walbot, V. (1998) Plant Cell 10: 1135-1149.
Baker, M.E. and Blasco, R.E. (1992) FEBS Lett. 301: 89-93.Bartley, G.E. and Scolnik, P.A. (1995) Plant Cell 7: 1027-1038.Birch, R.G. (1997) Annu. Rev. Plant Physiol. Plant Mol. Biol. 48: 297-
326.Boss, P.K., Davies, C. and Robinson, S.P. (1996) Plant Mol. Biol. 21:
565-569.Bradley, J.M., Davies, K.M., Deroles, S.C., Bloor, S.J. and Lewis, D.H.
(1998) Plant J. 13: 381-392.Brouillard, R. and. Dangles, 0. (1994) In The Flavonoids: Advances in
Research Since 1986. Edited by Harborne, J.B. pp. 565-588. Chapmanand Hall, London.
Brugliera, F., Barri-Rewell, G., Holton, T.A. and Mason, J.G. (1997) InAbst. 5th Intl. Cong. Plant Mol. Biol. p. 118.
Brugliera, F., Holton, T.A., Stevenson, T.W., Farcy, E., Lu, C.-Y. andCornish, E. (1994) Plant J. 5: 81-92.
Chuck, G., Robbins, T., Nijjar, C , Ralston, E. and Courtney-Gutterson,N. (1993) Plant Cell 5: 371-378.
Comai, L., Moran, P. and Maslyar, D. (1990) Plant Mol. Biol. 15: 373-381.
Courtney-Gutterson, N., Napoli, C , Lemieux, C , Morgan, A.,Firoozababy, E. and Robinson, K.E.P. (1994) Bio/Technology 12: 268-271.
Davies, K.M. and Scwinn, K.E. (1997) In Biotechnology of OrnamentalPlants. Edited by Geneve, R.L., Preece, J.E. and Merkle, S.A. pp. 259-294. CAB International, Wallingford.
de Vetten, N., Quattrocchio, F., Mol. J. and Koes, R. (1997) Gene Dev.11: 1422-1434.
Deroles, S.C., Boase, M.R. and Konczak, I. (1997) In Biotechnology ofOrnamental Plants. Edited by Geneve, R.L., Preece, J.E. and Merkle,S.A. pp. 87-120. CAB International, Wallingford.
Deroles, S., Bradley, J.M., Schwinn, K.E., Markham, K.R., Bloor, S.,Manson, D.G. and Dvies K.M. (1998) Mol. Breed. 4: 59-66.
Elomaa, P., Helariutta, Y., Griesbach, R.J., Kotilainen, M., Seppanen, P.and Teeri, T.H. (1995) Mol. Gen. Genet. 248: 649-656.
Elomaa, P., Honkanen, J., Puska, R., Seppanen, P., Helariutta, Y.,Mehto, M., Kotilainen, M., Nevalainen, L. and Teeri, T.H. (1993) Bio/Technology 11: 508-511.
Firoozababy, E., Moy, Y., Courtney-Gutterson, N. and Robinson, K.(1994) Bio/Technology 12: 609-613.
Forkmann, G. (1994) In The Flavonoids: Advances in Research Since1986. Edited by Harborne, J.B. pp. 537-564. Chapman and Hall, Lon-don.
Forkmann, G. and Rehnau, B. (1987) Z. Naturforsch. 42c: 1146-1148.Fujiwara, H., Tanaka, Y., Fukui, Y., Nakao, M., Ashikari, T. and
Kusumi, T. (1997) fur. / . Biochem. 249: 45-51.Gong, Z., Yamazaki, M., Sugiyama, M., Tanaka, Y. and Saito, K. (1997)
Plant. Mol. Biol. 35: 915-927.Goto, T. (1987) Prog. Chem. Org. Nat. Prod. 52: 114-158.Goto, T. and Kondo, T. (1991) Angrew. Chem. Int. Ed. Engl. 30: 17-33.Gallie, D.R. (1998) Curr. Opin. Plant Biol. 1: 166-172.Gutterson, N. (1995) Hort Sci. 30: 964-966.
Dow
nloaded from https://academ
ic.oup.com/pcp/article-abstract/39/11/1119/1861440 by guest on 18 N
ovember 2018

1126 Metabolic engineering to modify flower color
Helariutta, Y., Elomaa, P., Kotilainen, M., Seppanen, P. and Teeri, T.H.(1993) Plant Mol. Biol. 22: 183-193.
Heller, W. and Forkmann, G. (1994) In The Flavonoids: Advances inResearch Since 1986. Edited by Harborne, J.B. pp. 499-536. Chapmanand Hall, London.
Holton, T.A., Brugliera, F., Lester, D.R., Tanaka, Y., Hyland, CD. ,Menting, J.G.T., Lu, C.-Y., Farcy, E., Stevenson, T.W. and Cornish,E.C. (1993a) Nature 366: 276-279.
Holton, T.A., Brugliera, F. and Tanaka, Y. (1993b) Plant J. 4: 1003-1010.Holton, T.A. and Cornish, E.C. (1995) Plant Cell 7: 1071-1083.Holton, T.A. and Tanaka, Y. (1994) Trends Biotech. 12: 40-42.Jackson, D., Roberts, K. and Martin, C. (1992) Plant J. 2: 425-434.James, D.W., Lim, E., Keller, J., Plooy, I., Ralston, E. and Dooner, H.K.
(1995) Plant Cell 7: 309-319.Katsumoto, Y., Tsuda, S., Nakamura, N., Kusumi, T. and Fukui, H.
(1995) In Abst. 14th Plant Tissue Culture Conference Jpn. Assoc. PlantTissue Cult. pp. 103. (in Japanese).
Kondo, T., Yoshida, Y., Nakagawa, A., Kawai, T., Tamura, H. andGoto, T. (1992) Nature 358: 515-518.
Kroon, J., Souer, E., de Graaff, A., Xue, Y., Mol, J. and Koes, R. (1994)Plant J. 5: 69-80.
Kumpatla, S., Chandrasekharan, M.B., Iyer, L.M., Li, G. and Hall, T.C.(1998) Trends Plant Sci. 3: 97-104.
Linn, F., Heidman, I., Saedler, H. and Meyer, P. (1990)Mol. Gen. Genet.222: 329-336.
Lloyd, A.M., Walbot, V. and Davis, R.W. (1992) Science258: 1773-1775.Lu, C , Nugent, G., Wardley-Richardson, T., Chandler, S.F., Young, R.
and Dalling, M.J. (1991) Bio/Technology 9: 864-868.Lu, Y-P., Li, Z-S. and Rea, P.A. (1997) Proc. Natl. Acad. Sci. USA 94:
8243-8248.Marrs, K.A., Alfenito, M.R., Lloyd, A.M. and Walbot, V. (1995) Nature
375: 397-400.Martin, C. (1996) Curr. Opin. Biotech. 7: 130-138.Matzke, A.J.M. and Matzke, M.A. (1998) Curr. Opin. Plant Biol. 1: 142-
148.Meyer, P. and Heidmann, I. (1994) Mol. Gen. Genet. 243: 390-399.Meyer, P. , Heidemann, I., Forkmann, G. and Saedler, H. (1987) Nature
330: 677-678.Mitsuhara, I., Ugaki, M., Hirochika, H., Ohshima, M., Murakami, T.,
Gotoh, Y., Katayose, Y., Nakamura, S., Honkura, R., Nishimiya, S.,Ueno, K., Mochizuki, A., Tanimoto, H., Tsugawa, H., Otsuki, Y. andOhashi, Y. (1996) Plant Cell Physiol. 37 49-59.
Mol, J., Grotewold, E. and Koes, R. (1998) Trends Plant Sci. 3: 212-217.Mol, J.N.M., Holton, T.A. and Koes, R.E. (1995) Trends Biotech. 13:
350-355.Mol, J.N.M., van der Krol, A.R., van Tunen, A.J., van Blokland, R., de
Lange, P. and Stuitje, A.R. (1990) FEBS Lett. 268: 427-430.
Montgomery, M.K. and Fire, A. (1998) Trends Genet. 14: 255-258.Mooney, M., Desnos, T., Harrison, K., Jones, J,, Carpenter, R. and
Coen, E. (1995) Plant J. 7: 333-339.Napoli, C , Lemieux, C. and Jorgensen, R. (1990) Plant Cell 2: 279-289.Nielsen, K.M. and Podivinsky, E. (1997) Plant Sci. 167-174.Nozue, M., Yamada, K., Nakamura, T., Kubo, H., Kondo, M. and Nishi-
mura, M. (1997) Plant Physiol. 115: 1065-1072.Pelletier, M.K., Murrell, J.R. and Shirley, B.W. (1997) Plant Physiol. 113:
1437-1445.Quattroccio, F., Wing, J.F., Leppen, H.T.C., Mol, J.N.M. and Koes, R.
(1993) Plant Cell 5: 1497-1512.Que, Q., Wang, H-Y., English, J.J. and Jorgensen, R.A. (1997) Plant Cell
9: 1257-1368.Shirley, B.W. (1996) Trends Plant Sci. 1: 377-381.Stafford, H.A. (1994) Plant Sci. 101: 91-98.Stich, K., Eidenberger, T., Wurst, F. and Forkmann, G. (1992) Z. Natur-
forsch. 47C: 553-560.Strack, D. and Wray, V. (1994) In The Flavonid: Advances in Research
Since 1986. Edited by Harborne, J.B. pp. 1-22. Chapman and Hall, Lon-don.
Suzuki, K., Zue, H., Tanaka, Y., Fukui, Y., Mizutani, M. and Kusumi, T.(1997) Plant Cell Physiol. 38: s38.
Tanaka, Y., Fukuchi-Mizutani, M., Sakakibara, K., Fujiwara, H., Fukui,Y., Nakao, M., Ashikari, T. and Kusumi, T. (1997) In Absrtact 5th Intl.Cong. Plant Mol. Biol. p. 736.
Tanaka, Y., Fukui, Y., Fukuchi-Mizutani, M., Holton, T.A., Higgins, E.and Kusumi, T. (1995) Plant Cell Physiol. 36: 1023-1031.
Tanaka, Y., Tsuda, S. and Kusumi, T. (1998) In Applied Plant Biotechnol-ogy. Edited by Chopra, V.L. pp. 177-231. Oxford & IBH, New Delhi.
Tanaka, Y., Yonekura, K., Fukuchi-Mizutani, M., Fukui, Y., Fujiwara,H., Ashikari, T. and Kusumi, T. (1996) Plant Cell Physiol. 37: 711-716.
Taylor, C.B. (1997) Plant Cell 9: 1245-1249.Toguri, T., Umemoto, N., Kobayashi, O. and Ohtani, T. (1993) Plant
Mol. Biol. 23: 933-946.van der Krol, A.R., Lenting, P.E., Veenstra, J., van der Meer, I.M., Koes,
R.E., Gerats, A.G.M., Mol, J.N.M. and Stuitje, A.R. (1988) Nature333: 866-869.
van der Krol, A.R., Mur, L.A., Beld, M., Mol, J.N.M. and Stuitje, A.R.(1990) Plant Cell 2: 291-299.
Yamazaki, M., Gong, Z., Mizutani, M., Fujiwara, H., Tanaka, Y.,Kusumi, T. and Saito, K. (1998) In Abst. Japanese Society for Plant Celland Molecular Biology.
Yoshida, K., Kondo, T., Okazaki, Y. and Katou, K. (1995) Nature 373:291.
Zuker, A., Ahroni, A., Tzfira, T., Ovadis, M., Itzhaki, H., Scklarman, E.,Ben-Meir, H. and Vainstein, A. (1998) In Abst. IX. International Con-gress on Plant Tissue and Cell Culture 35.
(Received August 26, 1998; Accepted October 9, 1998)
Dow
nloaded from https://academ
ic.oup.com/pcp/article-abstract/39/11/1119/1861440 by guest on 18 N
ovember 2018





























