Embed Size (px)
Citation preview

UNIVERSITE DE OUAGADOUGOU
---------------
UNITE DE FORMATION ET DE
SCIENCES DE LA VIE ET DE LA TERRE
(UFR/SVT)
Mémoire Présenté
Par : Zakaria GAMSONRE
Pour l’obtention du Master II
de Biologie Moléculaire et de Génétique Moléculaire Appliquées
de l’Université de Ouagadougou
SUR LE THEME:
Diagnostic des gastro-entérites astrovirales et parasitaires chez les
enfants de moins de 5 ans de Ouagadougou (Burkina Faso)
Soutenu le 16 Novembre devant le jury composé de :
Président : Prof Nicola Barro, Professeur Titulaire, Université de Ouagadougou
Membres: Prof Jacques Simporé, Professeur Titulaire, Université de Ouagadougou
Dr Virginio Pietra, Chargé de recherche, Université de Brescia, Italie
LABORATOIRE DE BIOLOGIE MOLECULAIRE
ET DE GÉNÉTIQUE MOLÉCULAIRE
(LABIOGENE)
N° d’Ordre .............................../LABIOGENE

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page i
Préface du Coordonnateur du Master BIOGEMA
De nos jours, les connaissances avancées en génétique et biologie moléculaires sont
incontournables pour conduire des études de hautes valeurs ajoutées en sciences biologiques.
Les outils de la Biologie moléculaire ont permis d’accomplir de grands progrès dans le
domaine du diagnostic, de la pharmacie, de la thérapeutique, de l’agriculture et même dans
l’aide à la justice par l’identification humaine. Les Universités et les laboratoires de recherche
des Pays membres de l’UEMOA dans leur grande majorité, restent arrimés aux pays et
laboratoires de recherche du Nord pour leurs besoins en recherches et activités en génétique et
biologie moléculaires. Cet état de fait est lié au déficit en personnel qualifié et le manque de
ressources financières et matériel pour conduire les recherches en local in situ. Cela a pour
conséquence, une non maîtrise de la finalité ainsi que de l’utilisation des résultats et produits
des recherches que nous conduisons, une surenchère du coût des examens et des études en
biologie et génétique moléculaires. Un autre corollaire et non des moindres de cet état de fait
est la fuite de capitaux mais également la fuite des cerveaux car les étudiants les plus
compétents envoyés dans les pays du Nord ont tendance à y rester.
Le master en Biologie Moléculaire et en Génétique Moléculaire Appliquées (BioGeMA) a
pour but de combler le vide constaté dans l’expertise en génétique et biologie moléculaires
par la mise à disposition des pays de l’espace UEMOA, de personnels qualifiés, de haut
niveau de compétences pour conduire des études et recherches en génétique et biologie
moléculaires.
Le master BioGeMA est :
Un Master à dimension sous-régionale
Géré par un réseau de chercheurs et praticiens en génétique et biologie
moléculaires
Soutenu par une plateforme technologique sous-régionale à LABIOGENE
Ce Master a pour objectif de former des biologistes, des pharmaciens, des vétérinaires et des
médecins biologistes capables d’effectuer des diagnostics biomoléculaires dans des centres
hospitaliers et d’élaborer des études d’investigations dans des structures de recherches. En
outre, il ouvrira la porte d’études doctorales aux meilleurs étudiants pour permettre la
formation de chercheurs et d’enseignants-chercheurs afin d’assurer la relève du corps
enseignants, la constitution d’une masse critique d’experts africains et la mise en place d’un
véritable réseau africain de recherche dans le domaine ci-dessus cité.
Professeur Jacques SIMPORE
Professeur Titulaire de Biologie Moléculaire et de
Génétique Moléculaire
UFR/SVT-École Doctorale Sciences et
Technologie
Université de Ouagadougou – Burkina Faso

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page ii
Dédicaces
Je dédie ce Mémoire:
A ma famille notamment ma mère Koussoubé Aminata, mon père Gamsonré Yéwaya
Oumarou et sa deuxième épouse Sarambé Nathalie Marceline kalidiata pour
leurs assistances continues et tous les sacrifices consenties.
A mes frères et sœurs: Abasse, Maliki, Mahamadi, Arouna, Rihanata,
Roukiétou, Ramata, Neimatou, Boukari, Fatimata, Mamoudou, Ladifatou,
Salifou pour leurs conseils,
A ma chérie Salimata Sana pour tout le soutien et les encouragements durant
cette aventure du 3è cycle;
A toute la première promotion du Master de Biologie et de Génétique
Moléculaires Appliquées (BioGeMA), Qu’elle trouve ici l’expression de mes
profondes considérations.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page iii
Remerciements
Le présent mémoire qui sanctionne notre formation au Master est aussi le fruit de la collaboration et des sacrifices
consenties par des personnes physiques et /ou morales de bonne volonté auxquelles nous traduisons toutes nos
reconnaissances. Mes remerciements vont:
A l’endroit des autorités de l’université de Ouagadougou notamment celles de l’UFR/SVT pour avoir accepté
notre inscription au Master de BIOGEMA.
Au professeur Jacques Simporé, professeur titulaire de génétique et de Biologie Moléculaires à l’université
de Ouagadougou, Directeur du Centre de Recherche Biomoléculaire Pietro Annigoni (CERBA), Directeur du Centre
Médical Saint Camille (CMSC) et Recteur de l’Université Saint Thomas d’Aquin (USTA) pour nous avoir permis de nous
initier à la recherche en Biologie Moléculaire, pour avoir accepté de diriger cette étude et pour ses précieux conseils
ainsi que ses encouragements;
Au Profession Nicola Barro, professeur titulaire de virologie générale, vice-président chargé des
Enseignements et des innovations pédagogiques pour avoir accepté présider ce Jury;
Au Docteur Piétra Virginio, chargé de recherche à l’université de Bréscia, Italie, enseignant du Master pour
avoir accepté de juger ce travail;
Au Docteur Cyrille BISSEYE, Enseignant chercheur à l’Université de Franceville au Gabon, Enseignant du Master II
BioGeMA, pour l’encadrement au laboratoire et pendant la rédaction du présent mémoire, pour la qualité de son
encadrement et sa grande disponibilité;
A l’endroit de tout le personnel du Centre de Recherche de Biomoléculaire Piétro Annigoni (CERBA/LABIOGENE)
et du Centre Médical Saint Camille (SMSC) pour les services rendus, les conseils prodigués tout au long de ce stage et
principalement le père Yonli Directeur adjoint du CERBA/LABIOGENE;
Aux docteurs : Djenéba Ouermi, Florencia Djigma, Linda Sagnan et Léon Nitiéma, Soyez assurés de ma
profonde gratitude et de toute ma reconnaissance;
A l’endroit de Monsieur Youssouf Tao pour son aimable implication à l’amélioration de la qualité de ce
mémoire et son soutien moral;
A mes parents, mes frères et sœurs pour leurs soutiens multiformes et tous ceux qui d’une manière ou d’une
autre ont contribué à la réalisation de ce travail;
A tous les enseignants du master de BioGeMA et de l’UFR/SVT, pour la qualité des enseignements reçu au
cours de notre formation;
A toutes les mères qui ont acceptées participer à cette étude.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page iv
Résumé
Introduction: Les gastro-entérites constituent une des principales causes de mortalité et de
morbidité dans le monde entier notamment dans les pays en développement. La malnutrition
est un facteur favorisant l’aggravation des maladies diarrhéiques chez les enfants de 0 à 5 ans.
L’incidence des maladies diarrhéiques s’observe durant toute l’année avec un pic élevé en
saison sèche. La prévalence des astrovirus dans le monde varie de 2 à 16% (Hamkar et al,
2010) et celle parasitaire reste toujours élevée dans la plus part des pays pauvres.
L’objectif de notre étude est d’établir la prévalence des infections astrovirales et parasitaires
chez les enfants diarrhéiques de moins de 5 ans de la ville de Ouagadougou (Burkina Faso).
Méthode: Nous avons évalué le statut nutritionnel des 213 enfants admis en consultation au
Centre Médical Saint Camille de Mai 2012 à Avril 2013. Les astrovirus ont été diagnostiqués
par la RT-PCR grâce à l’amplification de leur région de capside (ORF2). Pour cela les ARNs
viraux ont été extraits à l’aide du kit DNA Sobent B de Sacaces biotechnologies à partir des
selles des enfants. Après la rétro-transcription et l’amplification, les produits PCR sont migrés
par électrophorèse puis revelés sur Ultra-violet. La bande de 175 Pb correspond aux
échantillons positifs. Quant aux parasites (Gliadia lamblia, les Trichomonas intestinalis,
Hyponema nana, Entamoeba histolytica), ils ont été détectés par observation directe à la
microscopie ou après coloration au lugol.
Résultats: Sur l’ensemble de notre échantillon la malnutrition touche 59,7% des enfants avec
une forte prévalence chez les enfants de moins de 24 mois. Les astrovirus ont une prévalence
de 14,60% (31/213) et celle des Parasites de 20,7% (44/213). Parmi les parasites isolés
Gliadia lamblia représentent (12,2%), Trichomonas intestinalis (8%), Entamoeba histolytica
(1,9%). Les cas de coïnfections (astrovirus et parasites) sont observés seulement dans 4,2%
(9/213) des individus de la cohorte. Les fortes prévalences des astrovirus sont observées chez
les enfants de moins de 24 mois et celles parasitaires chez les plus de 24 mois. L’allaitement
maternel au sein est un facteur protecteur contre les épisodes diarrhéiques. La saison et l’âge
serait des causes de fluctuations de l’incidence diarrhéiques chez les enfants de moins de 5
ans.
Conclusion: La connaissance de l’ensemble des étiologies impliquées dans les diarrhées est
nécessaire pour une lutte efficace des gastro-entérites infantiles.
Mots clés: Gastro-entérites, astrovirus, Parasites, Malnutrition, RT-PCR, Microscopie et
Burkina Faso.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page v
Abstract:
Introduction: gastroenteritis is the leading cause of mortality and morbidity worldwide
including in the developing world. Malnutrition is a contributing factor to the increase of
diarrheal diseases among children 0-5 years. The incidence of diarrhea was observed
throughout the year with a high peak in the dry season. The prevalence of astrovirus in the
world varies from 2 to 16% (Hamkar et al, 2010) and the parasite remains high in most poor
countries.
The aim of our study was to establish the prevalence of parasitic infections and astrovirales
diarrheal children under 5 years of Ouagadougou (Burkina Faso) .
Methods: We assessed the nutritional status of 213 children admitted in consultation Medical
Center Saint Camille May 2012 to April 2013. Astroviruses were diagnosed by RT- PCR
amplification with their capsid region (ORF2). For that viral RNAs were extracted using the
DNA Sobent B Sacaces biotechnology kit from stool of children. After reverse transcription
and amplification, the PCR products were migrated by electrophoresis and identified on
Ultraviolet. The bands 175 Pb correspond to positive samples. As for parasites (Gliadia
lamblia, Trichomonas intestinalis, Hyponema nana, Entamoeba histolytica), they were
detected by direct observation or microscopy after staining with Lugol.
Results: Of the whole sample malnutrition affects 59.7 % of children with a high prevalence
in children under 24 months. Astroviruses have a prevalence of 14.60 % (31 /213) and the
Pest 20.7% (44/ 213). Among the isolated parasites are Gliadia lamblia (12.2%),Trichomonas
intestinalis (8%), Entamoeba histolytica (1.9 %).Cases of co-infection (astrovirus and
parasites) are observed only in 4.2% (9/ 213) of individuals in the study cohort .The high
prevalence of astrovirus are seen in children less than 24 months and the parasite in more than
24 months . Breastfeeding Breastfeeding is a protective factor against diarrheal episodes.
Season and age would be the causes of fluctuations in diarrheal incidence among children
under 5 years.
Conclusion: an observation of individual and collective ownership would reduce the incidence
of diarrhea and malnutrition is a public health problem. Knowledge of all etiologies involved
in diarrhea is necessary for effective control of infantile gastroenteritis.
Keywords: Gastroenteritis, astrovirus, Parasite, Malnutrition, RT -PCR, microscopy and
Burkina Faso.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page vi
Table de Matières
Préface du Coordonnateur du Master BIOGEMA ...................................................................... i
Dédicaces ................................................................................................................................... ii
Remerciements .......................................................................................................................... iii
Résumé ...................................................................................................................................... iv
Table de Matières ...................................................................................................................... vi
Listes des figures ..................................................................................................................... viii
Liste des tableaux ...................................................................................................................... ix
Abréviation ................................................................................................................................. x
Introduction ................................................................................................................................ 2
1 Revue Bibliographique ........................................................................................................ 5
1.1. Généralités sur les astrovirus (HAstvs) ........................................................................ 5
1.1.1. Définition ......................................................................................................................... 5
1.1.2. Historique et distribution .................................................................................................. 5
1.1.3. La morphologie ................................................................................................................ 5
1.1.4. Critère de classification des astrovirus ............................................................................. 6
1.1.5. Organisation génomique .................................................................................................. 6
1.1.6. Mode de transmission des Astrovirus .............................................................................. 7
1.1.7. Les facteurs de risque ....................................................................................................... 8
1.1.8. Variabilité génétique et transmission interspécifique .................................................... 10
1.1.9. Le processus de clivage du génome des HAstVs ........................................................... 10
1.1.10. Cycle de vie et mécanisme d’invasion ......................................................................... 11
1.1.11. Réponse immunitaire .................................................................................................... 12
1.1.12. Diagnostic des astrovirus ............................................................................................. 13
1.1.13. Physiopathologie .......................................................................................................... 15
1.1.14. Traitement et prévention .............................................................................................. 16

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page vii
1.2. Les parasites intestinaux ............................................................................................ 17
1.2.1. Protozooses ..................................................................................................................... 17
1.3. Les autres gastroentérites. .......................................................................................... 20
1.3.1. Les rotavirus (Rv) ........................................................................................................... 21
1.3.2. Les norovirus .................................................................................................................. 21
1.3.3. Les adenovirus ................................................................................................................ 22
1.3.4. Les Bactéries .................................................................................................................. 22
2. Objectif du mémoire ............................................................................................................. 24
2.1. Objectif principal ........................................................................................................... 24
2.2. Objectifs spécifiques ...................................................................................................... 24
3. Matériels et Méthodes .......................................................................................................... 26
3.1. Cadre d’étude ................................................................................................................. 26
3.2. Population d’étude ......................................................................................................... 26
3.3. Détection des entéro-pathogènes ................................................................................... 27
3.3.1. Détection des astrovirus .......................................................................................... 27
3.2.6. Détection des parasites ............................................................................................ 30
3.2.6.1 Mode opératoire ........................................................................................................... 30
3.2.7. Considération éthique ............................................................................................. 30
3.2.8. Analyse statistique .................................................................................................. 30
4. Résultats ............................................................................................................................... 32
4.1. Caractéristiques sociodémographiques et cliniques des enfants ................................... 32
4.2. Prévalence des astrovirus et des parasites chez les enfants ........................................... 33
4.3. Répartition des astrovirus et des parasites en fonction des caractéristiques .................. 34
4.4. Répartition des infections astrovirales et parasitaires en fonction des signes cliniques des
enfants diarrhéiques .................................................................................................................. 36
4.5. L’influence des saisons sur les infections des entéro-pathogènes ................................. 36
5. Discussion ............................................................................................................................ 40
6. Conclusion ............................................................................................................................ 45
7. Références ............................................................................................................................ 47

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page viii
Listes des figures
Figure 1:Image d’astrovirus observée au microscope électronique en coloration négative....... 6
Figure 2: Structure du génome des astrovirus ............................................................................ 7
Figure 3: Différentes voies de transmission des HAsTVs ......................................................... 8
Figure 4: Processus de clivage du génome des astrovirus ....................................................... 11
Figure 5: Cycle de vie et mécanisme d’invasion des astrovirus ............................................. 12
Figure 6: Physiopathologie des astrovirus. .............................................................................. 16
Figure 7: Trophozoite vue de face ........................................................................................... 18
Figure 8: Kyste de giardia lamblia .......................................................................................... 18
Figure 9: Morphologie de strongyloides stercoralis ................................................................ 20
Figure 10: Structure d’un rotavirus .......................................................................................... 21
Figure 11: Carte administrative du burkina faso ...................................................................... 26
Figure 12:Extraction de l’ARN des astrovirus au CERBA ...................................................... 29
Figure 13:Electrophoregramme des produits PCR ................................................................... 34
Figure 14: Influence des saisons sur les infections des entero-pathogènes .............................. 38
Figure 15 : Prévalence saisonnière des astrovirus et des parasites .......................................... 38

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page ix
Liste des tableaux
Tableau 1 : Répartition des paramètres anthropométriques en fonction des tranches d’ âge. .. 32
Tableau 2: Répartition de l’état nutritionnel des enfants diarrhéiques en fonction des tranches
d’ âge ........................................................................................................................................ 33
Tableau 3: Caractéristiques pédiatriques en fonction des entéro-pathogènes .......................... 35
Tableau 4: Fréquence des astrovirus et des parasites en fonction des signes cliniques des
enfants ...................................................................................................................................... 37

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page x
Abréviation
ADNc
AdV
AGE
ARN
ARNg
ARNsg
BioGeMA
BET
CERBA
CMSC
EDTA
ELISA
HAstV
HAZ
Kb
LABioGene
MML-V
mn
nm
Nov
Nsp
OMS
ORF
PCR
PBS
PV/VIH
RGPH
RHO
RT
Rv
UEMOA
UTR
:
:
:
:
:
:
:
:
:
:
:
:
:
:
:
:
:
:
:
:
:
:
:
:
:
:
:
:
:
:
:
:
Acide Désoxyribonucléique Complémentaire;
Adenovirus;
Gastroentérite Aigue;
Acide Ribonucléique;
Acide Ribonucléique génomique;
Acide Ribonucléique sous génomique;
Biologie et Génétique Moléculaires Appliquées;
bromure d’éthidium;
Centre de Recherche Biomoléculaire Pietro Anigonni;
Centre Médical Saint Camille;
Acide éthylène diamine tétra-acétique;
Enzyme Linked Immuno Sorbent Assay;
Human Astrovirus;
Heigth for Age (taille/ âges);
Kilo-base;
Laboratoire de Biologie et de Génétique;
Virus de la leucémie murine de Moloney;
Minute;
Nanomètre;
Norovirus;
Protéine non structurale;
Organisation Mondiale de la Santé;
(Open Reading Frame) cadre de lecture ouverte;
Polymérase Chain Réaction (Réaction de Polymérase en Chaine);
Phosphate Buffered Saline;
Personne Vivant avec le VIH
Recensement Général de la Population et des Habitats;
Réhydratation par voie orale;
Reverse Transcription (Reverse transcriptase);
Rotavirus;
Union Economique et Monétaire Ouest Africain;
Untranslated Region (Région non traduite);

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page xi
UV
V
VLP
VPg
WAZ
WAH
:
:
:
:
:
:
Ultra-Violet;
Volt;
Virus-Like Particule (particule(s) virale(s) de synthèse);
Protéine Virale liée au Génome;
Weight for Age (poids/ âges);
Weigth for Heigth (Poids/ Taille).

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 1
Introduction

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 2
Introduction
Les gastroentérites constituent une des principales causes de mortalité et de morbidité chez les
enfants de moins de 5 ans et sont aussi une cause majeure de la malnutrition et du retard de
croissance (Hamkar et al., 2010). De par leur répartition saisonnière, les maladies diarrhéiques
causent annuellement 2,5 millions de décès parmi les enfants de moins de 5 ans et
représentent 21% de tous les décès dans cette tranche d’âge (Kosek et al., 2003, Glass et al.,
2006). Voir aussi données de mortalité infantile sur le site de l’OMS
Au Burkina Faso les diarrhées chez les enfants de moins de 5 ans sont causées par les virus
(Rotavirus, Astrovirus, Norovirus, Adenovirus), les parasites (Giardia lamblia, Trichomonas
intestinalis, Hyponelepis nana, Entamoeba histolytica, Cryptospridium, amibes), les bactéries
(Salmonella, Shigella, Campylobacter, Escherichia coli, Yersinia, Vibrio cholerae,
Helicobacter pylori) et les toxines (champignons, coquillages infectés par des algues toxiques
lesquels ?) (Sanou et al., 1999, Nitiema et al., 2011). Les coïnfections sont les plus fréquentes
dans presque la totalité des épisodes de diarrhées chez les moins de 5ans (Ouermi et al., 2007;
Nitiema et al., 2011).
Les astrovirus représentent la deuxième cause des gastro-entérites virales après les rotavirus
(Finkbeiner. et al., 2009, De Benedictis et al., 2011) chez les nourrissons, les adultes, les
vieillards (Mitchell et al., 2002), les immunodéprimées (Coppo et al., 2000, Wunderli et al.,
2011) y compris les personnes vivant avec le virus de l’immunodéficience humaine (Pv/VIH)
(Pollok et al., 2001).
Les astrovirus (HAstVs) sont des virus à ARN appartenant à la famille des Astroviridae qui
comprend le genre Mamastrovirus et le genre Avastrovirus (Chu et al., 2008, Bosch et al.,
2011). Les HAstVs sont reconnus comme une cause endémique des gastro-entérites aiguës
dans le monde entier, des études sérologiques démontrant que 90% des individus dans
certaines populations ont des anticorps dirigés contre les HAstV (Koopmans et al., 1998;
Glass et al., 2001).
Cependant, tous les sujets porteurs d’HAstV ne développent pas une maladie ulcéreuse; ceci
pourrait s’expliquer par les facteurs génétiques de l’hôte, des facteurs environnementaux et/ou
la virulence des sous types d’HAstVs. Les HAstVs ont un tropisme pour les microvillosités,
la lamina propria, les entérocytes matures, les macrophages sous-épithéliaux, les lysosomes,
les cryptes et les parenchymes (Sebire et al., 2004).
Ils induisent une diarrhée modérée ou sévère, des fièvres, la perte d’appétit, des douleurs
abdominales et des vomissements (Cubitt et al., 1996; Mitchell et al., 2002).

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 3
La transmission du virus se fait par la voie féco-orale, interhumaine et de l’animal à l’homme
(Pinto et al., 1996; Matsui et al., 2001). Les astrovirus constituent donc un problème majeur
de santé publique dans les pays développés et en voie de développements (Bon et al., 1999).
Les études menées dans le monde et en Afrique ont montré des prévalences très variées. Des
prévalences de 8,6%, 4,3% et de 3% ont été rapportées respectivement en Thailande, en
Australie et en Iran (Hermann et al, 1991); Schnagl et al., 2002; Hamkar et al, 2010). Au
Brésil 33% des enfants de moins de 5 ans présentaient des diarrhées à astrovirus (Silva et al.,
2001).
En Afrique, les rares études menées sur les HAstVs ont rapporté prévalences de 37,0% en
Afrique du Sud (Taylor et al., 1997), 4,0% en Côte d’Ivoire (Bini et al., 2007), et de 6,7% au
Nigeria et au Zaïre (Pennap et al., 2002). Au Burkina Faso la prévalence des astrovirus
jusque-là inconnue et leur rôle dans les gastroentérites chez les enfants n’a encore fait l’objet
d’étude.
Quant aux parasitoses, elles sont causées principalement par les helminthes et les
protozooses. Ces infections constituent un problème de santé publique dans les pays en
développement. En dépit des efforts entrepris pour leurs éliminations, les prévalences des
parasitoses restent toujours presque stables. Des prévalences de 10,7%, 18,2% ont été
rapportées respectivement au Burkina Faso (Sanou et al., 1999, Ouermi et al., 2007 chercher
autres références pour le BF) et en Ethiopie (Amare et al., 2012).
Au regard des fortes prévalences des infections astrovirales et parasitaires, il s’avère donc
nécessaire d’approfondir les connaissances sur le rôle de ces micro-organismes dans les
pathologies digestives. C’est donc cette nécessité qui a motivé le choix du thème:«Diagnostic
des gastro-entérites astrovirales et parasitaires chez les enfants de moins de 5 ans de
Ouagadougou (Burkina Faso)».

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 4
REVUE BIBLIOGRAPHIQUE

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 5
1 Revue Bibliographique
1.1. Généralités sur les astrovirus (HAstvs)
1.1.1. Définition
Les astrovirus (HAstV) sont des virus à ARN nu (sans enveloppe), à symétrie icosaedrale de
petite taille (28 à 30 nm de diamètre) appartenant à la famille des Astroviridae (Guix et al.,
2002; Bosch et al., 2011). Ils infectent les mammifères et certaines espèces d’oiseaux.
1.1.2. Historique et distribution
Les astrovirus humains (HAstV) ont été observés pour la première fois par Appleton et
Higgins (Appleton et al., 1975) au cours d’une épidémie de gastroentérites dans une maternité
en Ecosse. Leur appellation découle de l’observation des formes astrons (en grec étoile) en
microscopie électronique (Madeley et al., 1975).
Les astrovirus infectent les humains, les bovins, les porcins, les ovins, les visons, les chiens,
les chats, les souris, les poulets et les dindes (Jonassen et al., 2001, 2003). Ils se divisent en
huit sérotypes HAstV-1 à HAstV-8.
Les données d’épidémiologies moléculaires des infections à HAstV montrent que HAstV-1
est le sérotype le plus détecté dans de nombreuses régions du monde (Gaggero et al., 1998;
Mustafa et al., 2000; Mendez-Toss et al., 2004). Toutefois, le sérotype prédominant peut
varier dans différentes régions géographiques. Les sérotypes 1, 3, et 4 circulent en Australie
(Kurtz et Lee, et al., 1987; Schnagl et al., 2002), le sérotype 2 est le type le plus commun
trouvé au Mexique (Guerrero et al., 1998), et HAstV-8 a été trouvé en circulation dans la
population de l'Inde occidentale.
1.1.3. La morphologie
L’observation en microscopie électronique après coloration négative des HAstVs montre des
particules à symétries icosaédrales, les bords arrondis, lisses, un triangle dense aux électrons
et un centre clair (figure 1) leur conférant un aspect en étoile à cinq (5) ou six (6) pointes
(BAJOLET et al., 1998; Guix et al., 2002).

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 6
Figure 1:Image d’Astrovirus observée au microscope électronique en coloration négative
http://www.microbe-edu.org/etudiant/astroviridae.html 26/03/2013
1.1.4. Critère de classification des astrovirus
Les astrovirus appartenant à la famille des Astroviridae sont divisés en deux genres: les
Mamastrovirus et les Avastrovirus qui infectent respectivement les mammifères et les espèces
aviaires (Mendez et al., 2007; Bosch et al., 2011).
Les huit sérotypes ont été classés selon plusieurs critères. Les sept premiers sérotypes
humains ont été classés en HAstV-1 à HAstV-7 à partir de la réactivité du virus avec les
anticorps polychoraux et le huitième (HAstV-8) par génotypage (Belliot et al., 1997). La
comparaison des polyproteines nsp1a et nsp1b codées respectivement par les régions ORF1a,
ORF1b et ORF2 a permis d’isoler de nouveaux sérotypes des Astrovirus Humain:
HastrovirusMLB1, HastrovirusMLB2, Hastrovirus-SG, Hastrovirus-VA1, Hastrovirus-VA2,
HMOastrovirus-A, HMOastrovirus-B, HMOastrovirus-C (Méndez et al., 2013).
1.1.5. Organisation génomique
Le génome viral est à ARN positif d’environ 6,4 à 7,3 Kb (Guix et al., 2002). Il renferme trois
régions de fragments ouvertes (ORFs) désignés par ORF1a, ORF1b et ORF2. Les régions
ORF1a et ORF1b représentent la protéine non structurale codante et ORF2 est la protéine de
capside.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 7
1.1.5.1. Région non structurale
Les deux segments du gène de la protéine non structurale codent chacune pour une protéine
virale. ORF1a code pour la polyprotéine nsp1a non structurale. Il s’agit d’une protéine de
2700 nucléotides (nt) (Liu et al., 2008) qui contient une sérine protéase virale et nucléaire.
ORF1b code pour la polyprotéine nsp1ab dont l'ARN polymérase dépendante de l'ARN
(RdRp) qui s'exprime à travers un code de lecture ribosomique à la jonction ORF1a/1b médié
par une séquence polyAdénine glissante. Cette région variable code pour le domaine
d'interaction du récepteur qui se lie à des récepteurs spécifiques sur la surface des cellules-
hôtes (Krishna et al., 2005; Jeong et al., 2012).
1.1.5.2. Région de capside: région ORF2
La protéine de capside formée à partir de la région ORF2 du génome des HAstVs codant les
polypeptides viraux structuraux (Sanchez et al., 1994; Monroe et al., 2005; Mende'z et al.,
2007) sont traduits à partir d'un ARN sous-génomique synthétisé lors de la réplication du
virus. La région hypervariable de la protéine d'enveloppe code pour le domaine de
l'assemblage qui est nécessaire pour la formation de la capside virale et l’encapsidation de
l'ARN viral (Krishna et al., 2005; Bosch et al., 2011; De Benedictis et al., 2011; Jong et al.,
2012). Il est situé à l’extrémité 3’ de la séquence et code pour le précurseur de la protéine de
capside qui s’exprime à partir d'un ARNm sous-génomique de 2,4 kb dans les cellules
infectées par les HAstVs (Monroe et al., 1993).
La région ORF2 est celle qui a été amplifiée par la RT-PCR pour le diagnostic des astrovirus
au cours de la présente étude.
Figure 2: Structure du génome des Astrovirus. ORF : Région de Fragment Ouverte, Pro :
Promoteur, Pol : Polymérase, Capsid : Capside, nt : nucléotides. Walter et al., 2001.
1.1.6. Mode de transmission des Astrovirus
Les astrovirus se transmettent par voie directe par voie indirecte.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 8
1.1.6.1. Transmission directe
Les HAstVs se transmettent par les voies hydriques, d'origine alimentaire et oro-fécale (Pinto
et al., 1996; Matsui et al., 2001). Cette première voie est considérée principale ou majeure.
1.1.6.2. Transmission indirecte
Les astrovirus peuvent aussi se transmettre d’une personne à une autre, d’un l’animal à
l'homme et à travers les aérosols (Cattoli et al., 2007; Kapoor et al., 2009; Levy et al., 2009;
Chu et al, 2010; Luo et al., 2010). La transmission s’effectue aussi via les aliments
contaminés ou par contact.
Figure 3: Différentes voies de transmission des HAstVs
http://www.microbe-edu.org/etudiant/astroviridae.htm 26/02/2013
1.1.7. Les facteurs de risque
Les infections astrovirales sont associées à l’âge, à l’agent étiologique et dépendent des
facteurs associés à l’hôte (système immunitaire, hypo-acidité gastrique….).
1.1.7.1. Facteurs liés à l’hôte
Les facteurs liés à l’hôte sont ceux liés directement à la personne hébergeant l’agent
infectieux. Le système immunitaire constitue le facteur lié à l’hôte le plus important. Le
système de défense contre les astrovirus comprend une immunité naturelle d’origine
muqueuse et une immunité acquise issue des tissus lymphoïdes.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 9
L’allaitement au sein interrompu avant l’âge de 2 ans, le déficit immunitaire, l’hypo-acidité
gastrique, la motricité intestinale réduite et les facteurs génétiques (par exemple type sanguin)
contribuent à une réduction de la résistance aux infections astrovirales.
1.1.7.2. L’âge
La plupart des épisodes diarrhéiques à HAstV se produisent au cours des deux premières
années de la vie. Les fortes incidences sont observées avant le deuxième anniversaire (0 à 24
mois), âge auquel le sevrage est fréquent (Hamkar et al., 2010, Nitiema et al., 2011; Maham et
al., 2012; Lal et al., 2012).
1.1.7.3. Les saisons
Des études épidémiologiques ont montré qu'il existe une forte prévalence des infections
astrovirales en hiver et dans les régions tempérées (Walter et al., 2003; Liu et al., 2006; Lin et
al., 2008) de même qu’en saison hivernale dans les régions tropicales (Nguyen et al., 2008;
Verma et al., 2010). Mais le pic d’infection est observé en période chaude pendant le mois de
Janvier (Lin et al., 2008). De faibles prévalences ont été observées au cours des autres mois de
l’année (Verma et al., 2010, Lin et al., 2008).
1.1.7.4. L’agent étiologique
Il s’agit de la virulence de l’agent étiologique, de son écologie et de sa capacité à vivre et à
s’adapter hors de l’hôte et de sa physiopathologie. La virulence de l’agents étiologique des
diarrhées à astrovirus varie selon les sérotypes (Walter et al., 2001; Kapoor et al., 2009).
Les HAstVs sont des virus qui résistent à un pH=3, aux chloroformes, aux détergents, aux
solvants lipidiques et à la chaleur.
Ils conservent leurs pouvoirs infectieuses pendant 6 à 10 ans lorsqu’ils sont gardés à -70°C.
Les mutations et les recombinaisons génétiques favorisent la résistance des astrovirus aux
antibiotiques (Cattoli et al., 2007; Luo et al., 2010).
La concentration du virus dans les selles, sa capacité à conserver sa pathogénicité et la dose
nécessaire pour permettre son développement hors de l’organisme sont également des facteurs
favorisant son infectiosité.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 10
1.1.8. Variabilité génétique et transmission interspécifique
Les mutations et les recombinaisons génétiques seraient à l’origine de la variabilité génétique
observée dans les souches recombinantes d’astrovirus (Walter et al., 2001). De nouvelles
souches d’astrovirus (HAstV, MLB1 et HMOAstV) détectés dans les échantillons de selles
humaines sont affiliées aux astrovirus d’origine animale et indiquent la transmission
zoonotique des HAstVs chez les humains (Kapoor et al., 2009). Récemment, de nouvelles
variantes des AHstVs trouvés chez les rats urbains (Rattus norvegicus) ont montré une
relation étroite avec HAstV-MLB1 et HAstV-MLB2 qui ont indiqué que ces deux souches ont
partagé un ancêtre commun (Chu et al., 2010).
1.1.9. Le processus de clivage du génome des HAstVs
La réplication des astrovirus serait liée à une réponse cellulaire de type apoptotique et les que
caspases seraient impliquées dans la maturation des capsides avant le clivage extracellulaire
(Mendez et al., 2004; Guix et al ., 2004).
Le clivage de la région VP90 se fait au niveau de la région carboxyle terminale en quatre
positions libérant ainsi des précurseurs des protéines intermédiaires de 82k, 78k, 75k et le
produit de clivage intracellulaire mature VP70 (Mendez et al., 2004). L’arginine 393
(Arg393) va d’abord cliver la protéine VP70 en Vp41 et VP28. Ces protéines sont ensuite
clivées d’une manière séquentielle au niveau des carboxyles et des amines terminales
respectivement pour donner les produits VP34 dérivés de VP41, VP27 et VP25 dérivés de
VP28 (figure 5).
Les différentes souches des HAstVs ont un processus de clivage similaire car les produits
finaux des différentes souches sont très identiques (Méndez et al., 2013; Mendez et al., 2004;
Sanchez et al.,1994).
Les particules des astrovirus sont libérées sous forme de particule VP70. Elles sont soit non
infectieuses ou moins infectieuses et l'activation de leur infectiosité nécessite que la particule
VP70 soit traitée par la trypsine. En effet ce traitement multiplie par 100 l’infectiosité dans le
cas des huit souches des HAstVs (Béliot et al., 1997; Mendez et al.,2004; Méndez et al.,
2013).

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 11
Figure 4: Processus de clivage du génome des astrovirus. VP: Particule Virale, aa: acide
aminés, P: Protéine. D’après Méndez et al., 2013.
1.1.10. Cycle de vie et mécanisme d’invasion
Il est probable que la pénétration des astrovirus dans les cellules s’effectue par le pôle apical à
travers un mécanisme d’endocytose. L’interaction du virus avec la membrane provoque
l’activation de la régulation du signal Kinase (ERK1/2). Bien que ce mécanisme d’activation
demeure inconnu, il est cependant indépendant de la réplication du virus (Moser et al., 2008).
Les endosomes précoces formées se transforment par la suite en endosomes tardifs qui se
désintègrent libérant ainsi les ARNs génomiques qui subiront une translation et puis une
réplication. L’ARN génomique est ensuite traduit en un précurseur de polyproteines nsp1a et
nsp1b qui sont ensuite clivées en protéines non structurales. Ils se fixent sur le cytosquelette et
subiront une réplication complexe pour donner deux ARNs génomiques (ARNg) négatifs et
positifs. Une première transcription de l’ARN génomique négatif produit des ARNs sous
génomiques qui subiront une translation avant d’être traduits par les ribosomes en protéines
de structures précoces qui s’assemblent avec l’ARNg+ pour donner des particules virales
(VP) de 90 Pb. Les particules VP90 libérés par lyse de la cellule doivent être clivées par les
caspases impliquées et la trypsine en particules VP70 qui seront ainsi libérées (Méndez et al.,
2013).

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 12
Figure 5: Cycle de vie et mécanisme d’invasion des Astrovirus : nsp1a: Protéine non
structurale 1a, nsp1b : Protéine non structurale 1ab, gRNA: Acide Ribonucléique génomique, sgRNA:
Acide Ribonucléique sous-génomique, VP: Particule Virale, ERK1/2: Signal de Régulateur Kinase.
Adapté de Méndez et al., 2013.
1.1.11. Réponse immunitaire
1.1.11.1. Immunité naturelle
L’immunité astrovirale n’est pas bien comprise. La défense contre les agents pathogènes se
fait à partir de l’immunité naturelle d’origine muqueuse et secondairement à partir du tissu
lymphoïde. Les jeunes enfants et les personnes âgées, (population à risques), sont
généralement ceux qui développent des infections asymptomatiques. Ce qui suggère que les
anticorps sont acquis tôt dans l’enfance. A la naissance, les anticorps maternels passent chez
l’enfant par voie transplacentaire et semble intervenir dans le système de défense pendant une
durée indéterminée.
Le lait maternel et le colostrum fournissent également des protections initiales contre les
microorganismes par le biais des anticorps qu’ils contiennent. Ces anticorps assurent la
protection au cours de l’enfance mas diminue avec l’âge (Glass et al., 2006).

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 13
La présence des HAstVs activent les macrophages qui produisent l’oxyde nitrique qui
supprime la réplication virale (Koci et al., 2004). Le système immunitaire inné en particulier
les macrophages et l’oxyde nitrique inductible synthétase (iNOs) jouent un rôle clé dans le
contrôle de la réplication. Les anticorps spécifiques du virus (Abs) sont essentiels à la
résistance aux infections à astrovirus (Koci et al., 2004).
1.1.11.2. Immunité acquise
L’immunité acquise implique l’action des anticorps synthétisés par l’hôte pour neutraliser les
antigènes des astrovirus. L’action des cytokines, des interleukines et les lymphocytes est peu
efficace à cause de l’immaturité du système immunitaire chez les nourrissons tandis que, chez
les adultes, c’est l’immunodépression qui favorise la progression de la maladie.
1.1.12. Diagnostic des astrovirus
Le diagnostic des HAstVs repose sur deux types de méthodes de détection: direct et indirect.
1.1.12.1. Les méthodes directes
Ces méthodes cherchent à isoler le virus lui-même à partir des fèces. Elles utilisent des
techniques de biologie moléculaire: RT-PCR pour amplifier l’ARN viral et la technique de
microscopie électronique.
La microscopie électronique
Principe: Elle consiste à placer un échantillon suffisamment mince sous un faisceau
d'électrons et à utiliser un système de lentilles magnétiques pour projeter l'image de
l'échantillon sur un écran phosphorescent qui transforme l'image électronique en image
optique.
Avantages: Les effets d'interactions entre les électrons et l'échantillon donnent naissance à
une image dont la résolution peut atteindre 0,08 nm. C’est ainsi que les HAstVs de 28 à 30
nm ont été observés par microscopie électronique (Appleton et al., 1975).
Inconvénients: Les inconvénients majeurs de cette technique sont le coût élevé de l’appareil
et la possibilité de confondre souvent les astrovirus à d’autres petits virus ronds entériques.
Elle est moins sensible et difficile à utiliser en routine (Glass et al., 2000).

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 14
La Transcription Reverse-Réaction de Polymérisation en Chaine (RT-PCR)
La RT-PCR est une technique qui associe une transcription reverse (RT) suivie d’une réaction
de polymérisation en chaine (PCR).
La RT-PCR est 3 fois plus sensible que les tests enzymatiques (Monroe et al., 2001; Guix et
al., 2002).Une étude de Grote et al en (2011) a révélé que 30% des échantillons fécaux
négatifs par ELISA (Enzyme–Lynked immunosorbent assay) étaient positifs par RT-PCR.
Des résultats similaires ont été obtenus en Arabie Saoudite et en Corée respectivement par
(Tayeb et al, en 2008 et Jeong et al, en 2011).
Inconvénients: L’inconvénient majeur de cette technique est le coût élevé de l’appareil.
RT-PCR one-step
Elle est une variante de la RT-PCR qui s’effectue en une seule étape associant la reverse
transcriptase et la Taq-polymerase assurant le couplage de la transcription reverse et
l’amplification génomique. Cette technique est plus avantageuse car elle réduit la possibilité
de contamination croisée. Elle a la même sensibilité que la RT-PCR classique (Jiang et al.,
1999; Vennema et al., 2002; Vinje et al., 2003).
1.1.12.2. Les méthodes indirectes
Ces types d’analyses médicales ne visent pas directement le virus mais les anticorps anti-
astrovirus synthétisés par l’organisme humain lors du contact avec l’antigène viral. Les
méthodes les plus connues et les plus utilisées sont : Les tests ELISA et le test d’agglutination
au latex (LA).
Les tests ELISA
L’ELISA est une technique de diagnostic de laboratoire qui révèle la présence des anticorps
anti-astrovirus dans les selles (Guix et al., 2002). La prévalence des astrovirus serait sous-
estimée par l’utilisation des tests enzymatiques qui sont moins sensibles que la RT-PCR
(Tayeb et al., 2008; Jeong et al., 2011).
Principe: Les tests rapides ELISA sont très simples, trop qualifié et l'anticorps qui détecte
l'antigène n'est pas marqué.
Avantage: Le test est rapide (2 heures30mn) et sensible (utilisation d'anticorps monoclonaux)
Inconvénients: La Faible sensibilité en cas de période de séroconversion.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 15
Le test d’agglutination au latex
Le test d'agglutination au latex (LA) est plus simple et plus pratique. C’est la méthode de
diagnostic rapide de référence utilisée pour la détection du virus dans les échantillons de
selles. Elle est plus rapide que la microscopie électronique (EM) ou la RT-PCR. Komorya et
al., (2003) ont rapporté une sensibilité et une spécificité de 88,5% et de 97,9% du test
d’agglutination au latex dans le diagnostic des astrovirus.
1.1.13. Physiopathologie
Contrairement aux rotavirus qui ont été largement étudiés, de nombreuses questions
concernant la pathogénicité, l’épidémiologie et l’implication des HAstVs dans la survenue des
millions de cas annuels des gastroentérites restent à évaluer de façon précise.
Les HAstVs ont une localisation très variée dans le tube digestif. Ils sont détectés dans les
selles et la maladie se manifeste par une diarrhée (chez les enfants en milieu tropical).
Une étude expérimentale faite sur la lame basale a montré que les HAstVs infectent les
entérocytes matures et les macrophages sous-épithéliaux de l’intestin grêle provoquant une
atrophie villositaire partielle ou plus rarement totale (Hall et al.,1987). Ce qui pourrait induire
un syndrome de malabsorption en cas d’infestation massive des microvillosités, le rejet de
l’eau et des nutriments avec les selles.
La multiplication intra-cytoplasmique entraîne une vacuolisation du cytoplasme avec
aplatissements des villosités qui se couvrent de cellules cuboïdes immatures et résistantes aux
l'infections. L’examen en microscopie électronique des parties de l’intestin grêle a révélé la
présence des HAstVs dans les para cristallines, le long des microvillosités et dans les
lysosomes des vacuoles autophagiques (Sebire et al., 2004). On note également la présence du
virus dans les macrophages et dans la lamina propria.
Les études antérieures menées sur les dindes ont rapporté les dommages causées par les
infections à HAstVs qui se caractérisent par une nécrose épithéliale bénigne, un infiltrat
inflammatoire de la lamina propria, une atrophie minimale des villosités et une hyperplasie
des cryptes (Sebire et al., 2004).
Une étude récente a révélé que les HAstVs seraient les agents causals de l'encéphalite chez un
garçon de 15 ans avec agammaglobulinémies (Quan et al., 2010).

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 16
Figure 6: La physiopathologie des astrovirus. (A) Photomicrographie d’une biopsie du jéjunum d’un
receveur de greffe osseuse atteint d’une infection à HAstV (grossissement×6100): microvillosités moussantes,
modification non spécifique des cellules épithéliales et un infiltrat inflammatoire de la lamina propria (B) et (C):
Photomicrographie d’une biopsie du jéjunum (B) et du duodénum (C) d’un receveur de greffe osseuse atteint
d’une infection à HAstV immunocolorée avec des anticorps anti-Astrovirus: présence d'importantes tâches sur la
surface des cellules l'épithéliales, le plus souvent près des extrémités des villosités (grossissements original,
6400 et 6100, respectivement).
Figure D: Electromicrographe d'un entérocytes du jéjunum démontrant le cytoplasmique para cristallin de
particules virales des HAstVs (grossissements originaux, 63000 et 6100 000 (en médaillon). Sebire et al.,
2004.
1.1.14. Traitement et prévention
Certains aspects concernant la réplication et les mécanismes immunitaires de l'infection à
HAstV restent à éclaircir. Une évaluation plus précise du rôle des HAstVs dans les diarrhées
d’origine virale de l'enfant est nécessaire afin d'estimer l'intérêt des différentes stratégies de
prévention et en particulier le développement d'un vaccin.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 17
1.1.14.1. Traitement
Il n’existe pas jusqu’à ce jour un vaccin contre les gastroentérites à HAstV. Le sel de
réhydratation orale (SRO) est liquide qui confère une protection contre les diarrhées modérées
et sévères dues aux HAstVs. Les enfants ayant des signes de déshydratation sévère risquent de
mourir rapidement par choc hypovolémique. Dans ce cas le traitement de choix est la
réhydratation par voie intraveineuse qui permet de restaurer plus rapidement le volume d’eau
et d’électrolytes perdus. Ajouter protocole réhydratation en cas de malnutrition sévère
(RESOMAL).
1.1.14.2. Prévention
La prévention des maladies diarrhéiques à HAstV nécessite une bonne hygiène
individuelle et collective. La multiplication des sensibilisations de la population sur l’impact
des infections gastro-entérites et les Conseils de Récupération et d’Education Nutritionnelle
(CREN) pourraient éviter les infections astrovirales.
Une mise au sein précoce les heures qui suivent la naissance protègerait les nouveau-
nés des infections astrovirales et réduirait la mortalité infantile.
La supplémentation des aliments en micronutriments (Iode, vitamines A, fer, Calcium,
Zinc, Magnesium).
1.2. Les parasites intestinaux
L’épidémiologie des maladies parasitaires (protozooses et helminthiases) constitue un
problème de santé publique dans les pays de l’Afrique subsaharienne. Les parasitoses
intestinales sont répandues pratiquement dans le monde entier et ont des prévalences élevées
dans de nombreuses régions.
1.2.1. Protozooses
1 .2.1.1. Giardia lamblia
Giardia Lamblia, protozoaire ubiquitaire de l’environnement, est l’un des parasites
intestinaux les plus courants infectants les êtres humains (Ali et al., 2003; Savioli et al., 2006).
Dans certains pays à faible revenu plus de 90% des enfants ont vécu au moins une épisode
d’infection à Giardia avant l’âge d’un an (Mondal et al., 2012). Les Giardioses surviennent
après l’ingestion de kystes de Giardia Lamblia qui subissent un réveil dans le duodénum puis

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 18
une différenciation en formes végétatives. Elles adhèrent ensuite aux cellules épithéliales de
l’intestin mais ne peuvent cependant pas envahir la muqueuse duodénale (Hill et al., 2009).
Morphologie
Giardia lamblia est un protozoaire flagellé, diplomonodida de la famille des Hexamitidés se
présentant sous deux formes: le stade végétatif ou trophozoite et le kyste. Le trophozoïte
mesure environ 15 μm de long, 10 μm de large et 3 μm d’épaisseur avec une face dorsale
convexe et ventrale concave muni de ventouse. Cette ventouse permet la fixation du parasite
aux cellules intestinales. Il possède 4 paires de flagelles très longs dirigés vers la partie
postérieure. Le kyste est ovale à contour lisse de 10-14 µm, mince, réfringent avec une
enveloppe rétractée et décollée donnant l’impression d’une double coque. Le kyste mur
renferme 4 noyaux.
http://www.infectiologie.org.tn 10/07/2013
Physiopathologie
Les Giardioses se transmettent par ingestion des kystes qui se transforment en forme
végétative une fois dans l’estomac. Les formes végétatives après division binaire vont se fixer
sur la muqueuse duodénale provoquant l’altération des entérocytes qui est à l’origine de
l’atrophie villositaire. Elles utilisent les nutriments et les acides biliaires qui seraient la cause
d’une malabsorption des graisses et des vitamines liposolubles (Vitamine B12). Le diagnostic
peut se faire par la recherche des kystes et des formes végétatives grâce à l’examen
Figure 7: Trophozoite vue de face Figure 8: kyste de Giardia Lamblia

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 19
parasitologique directe des selles ou encore la détection des antigènes par ELISA dans les
selles et les tests moléculaires.
1 .2.1.2. Les Helminthiases
Hyponelepis nana
Hyponelepis nana est un cestode cosmopolite le plus commun des infections parasitaires dans
les régions tropicales à mauvaises conditions hygiéniques.
Cycle évolutif
Hymenolepis nana est un cestode monoxène (à cycle direct) et capable d’une auto-infestation
endogène. Dans le cas de l'autoreinfestation, les œufs pondus dans l'intestin par le parasite
adultes libèrent des larves qui pénètrent les villosités et s'y transforment en larves
cysticercoides. Ces dernières se libèrent ensuite des villosités et une fois activées, évoluent en
adultes. Une transmission interhumaine est également possible et peut occasionner des
épidémies. Il existe également un mode de transmission indirecte dans lequel l'acquisition des
stades cysticercoides infestants se fait par ingestion de charançons.
Physiopathologie
Les infestations intensives par Hyponelepis nana sont observées chez des enfants malnutris ou
chez les immunodéprimés. Les larves cysticercoides lèsent la muqueuse intestinale, ce qui
pourrait expliquer les signes de pertes protéiques notés chez les sujets parasités. Cette
infection s'accompagne de diarrhée, de douleurs abdominales, d'asthénie et d'amaigrissement.
1.2.1.3. Strongyloides Stercoralis
Strongyloides stercoralis est un nématode parasite qui se développe librement dans les
cellules animales.
Morphologie
Strongyloides stercoralis est un vers intestinal de 2 mm de long provoquant une maladie
appelée strongyloïdiose.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 20
Figure 9: Morphologie de Strongyloides stercoralis
http://www.mlo-online.com/articles/200911/strongyloides-stercoralis-a-case-
study.php&docid 13/07/13
Physiopathologie
Strongyloides stercoralis est un nématode parasite qui peut aussi se reproduire dans l'hôte
animal. L’hôte s’infeste souvent par effraction et l’infection varie en gravité allant largement
de la forme asymptomatique à des complications.
1.3. Les autres gastroentérites.
Les gastroentérites sont très fréquentes. Les agents les répandus sont les rotavirus, connus
pour être la cause la plus commune de diarrhée aiguë sévère chez les enfants de moins de 5
ans dans le monde (Olesen et al., 2005), les adénovirus entériques, les calicivirus (dont le
virus de Norwalk ou norovirus), et les bactéries. Ces micro-organismes sont non spécifiques
de la muqueuse intestinale et se caractérisent par des troubles digestifs aigus (nausées,
vomissements, douleurs abdominales, diarrhées hydro-électrolytiques) accompagnés de signes
généraux souvent modérés. Les diarrhées évoluent généralement de manière bénigne en
quelques jours mais des formes sévères avec des déshydratations sont possibles notamment
chez les nourrissons (Pignatelli et al., 2000).
Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 21
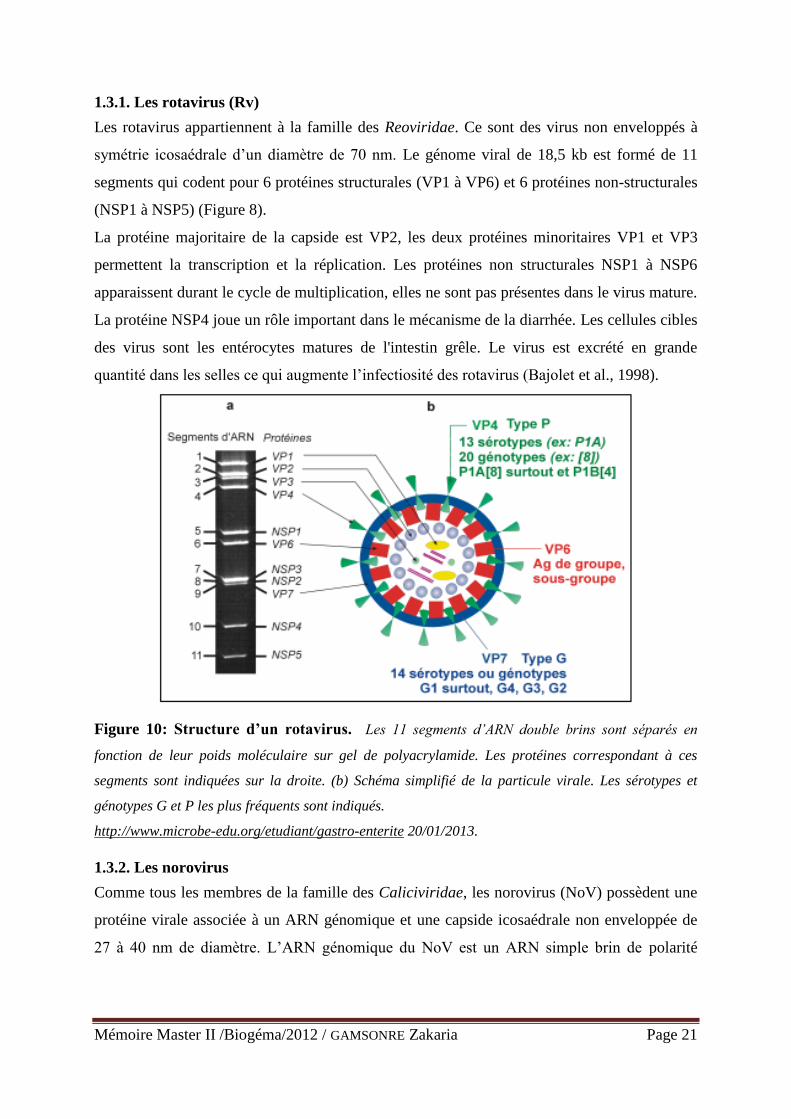
1.3.1. Les rotavirus (Rv)
Les rotavirus appartiennent à la famille des Reoviridae. Ce sont des virus non enveloppés à
symétrie icosaédrale d’un diamètre de 70 nm. Le génome viral de 18,5 kb est formé de 11
segments qui codent pour 6 protéines structurales (VP1 à VP6) et 6 protéines non-structurales
(NSP1 à NSP5) (Figure 8).
La protéine majoritaire de la capside est VP2, les deux protéines minoritaires VP1 et VP3
permettent la transcription et la réplication. Les protéines non structurales NSP1 à NSP6
apparaissent durant le cycle de multiplication, elles ne sont pas présentes dans le virus mature.
La protéine NSP4 joue un rôle important dans le mécanisme de la diarrhée. Les cellules cibles
des virus sont les entérocytes matures de l'intestin grêle. Le virus est excrété en grande
quantité dans les selles ce qui augmente l’infectiosité des rotavirus (Bajolet et al., 1998).
Figure 10: Structure d’un rotavirus. Les 11 segments d’ARN double brins sont séparés en
fonction de leur poids moléculaire sur gel de polyacrylamide. Les protéines correspondant à ces
segments sont indiquées sur la droite. (b) Schéma simplifié de la particule virale. Les sérotypes et
génotypes G et P les plus fréquents sont indiqués.
http://www.microbe-edu.org/etudiant/gastro-enterite 20/01/2013.
1.3.2. Les norovirus
Comme tous les membres de la famille des Caliciviridae, les norovirus (NoV) possèdent une
protéine virale associée à un ARN génomique et une capside icosaédrale non enveloppée de
27 à 40 nm de diamètre. L’ARN génomique du NoV est un ARN simple brin de polarité

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 22
positive, protégé par une protéine VPg jouant le rôle de coiffe à l’extrémité 5’, et polyadénylé
à l’extrémité 3’ (Green et al., 2001).
Les NoVs sont subdivisés en génogroupes I et II. Le génogroupe II est impliqué dans 90% des
gastro-entérites à NoV (Glass et al., 2000, Lopman et al.,2002). Les conséquences d’une
infection par les NoVs sont variables. Après deux à trois jours d’incubation se développent
différents symptômes dont les principaux sont la fièvre, les vomissements, les diarrhées et le
mal de tête. Les NoVs sont associés à 12% des cas de gastro-entérites sévères chez les enfants
de moins de 5 ans dans le monde (Patel et al., 2008).
1.3.3. Les adenovirus
Les adenovirus (AdV) humains classiques provoquent pour la plupart des infections
respiratoires et conjonctivales et certains sérotypes possèdent un pouvoir oncogène. Parmi les
51 sérotypes d’AdV (De jong et al., 1999), les types 40 et 41 d’origines entériques sont les
plus étudiés en tant que responsables des troubles digestifs chez l’Homme. Ils ont été liés à un
spectre de manifestations cliniques, notamment respiratoires, gastro-intestinales, oculaires,
neurologiques et infections urinaires. Ils sont le plus souvent retrouvés dans les voies
aériennes supérieures et parfois dans les selles. Les sérotypes 40 et 41, non cultivables, sont
connus comme agents des gastroentérites aiguës (GEA) infantiles (Bon et al., 1999).
Les AdV humain sont des virus à ADN non enveloppés de taille variant entre 70 et 80 nm et
ayant une symétrie icosaédrale portant des fibres hémagglutinines. Ils résistent aux conditions
environnementales défavorables.
1.3.4. Les Bactéries
La plupart des bactéries responsables des diarrhées appartiennent à une grande famille des
bactéries Gram négatif, les Enterobacteriaceae et certaines Vibrionaceae. Les entéro-
bacteriaceae sont majoritairement des commensaux de la flore bactérienne. Les diarrhées
d’origines bactériennes ont un impact sur la mortalité infantile. Ces bactéries colonisent
diverses parties du tube digestif provoquant des gastro-entérites plus ou moins sévères.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 23
OBJECTIFS DE L’ETUDE

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 24
2. Objectif du mémoire
2.1. Objectif principal
L’objectif principal de cette étude est d’évaluer les prévalences des infections astrovirales et
parasitaires chez les enfants de moins de 5 ans à Ouagadougou.
2.2. Objectifs spécifiques
Pour réaliser notre objectif principal, plusieurs objectifs spécifiques ont été définis:
1. Evaluer les caractéristiques sociodémographiques et cliniques, de même que l’état
nutritionnel des enfants.
2. Évaluer les prévalences des astrovirus et des parasites en fonction des paramètres
sociodémographiques, des saisons, des signes cliniques et du type d’allaitement.
3. Déterminer la prévalence des gastro-entérites à astrovirus et à parasites en fonction de
l’état nutritionnel des enfants.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 25
MATERIELS ET METHODES

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 26
3. Matériels et Méthodes
3.1. Cadre d’étude
L’étude s’est déroulée au Centre Médical Saint Camille (CMSC) de Ouagadougou au
Burkina Faso. Ouagadougou est située dans la zone intertropicale marquée par deux saisons :
une saison sèche d’octobre à Mai et une saison pluvieuse de Juin à Septembre. Le CMSC qui
fait partie du district sanitaire du secteur 30 est un centre médical à caractère social et
humanitaire comportant plusieurs services entre autres : le service de Santé Maternel et
Infantile (SMI) et le laboratoire d’analyses biomédicales où a eu lieu la collecte d’échantillons
de selles. Les analyses biomoléculaires ont été faites au Centre de Recherche Biomoléculaire
Piétro Annigoni (CERBA).
Figure 11: Carte administrative du Burkina Faso
http://www.1clic1planet.com/burkina%20faso.htm,1/08/2013 Ne pas « tirer » les images
par les côtés, ça le déforme. Tirer par les coins.
3.2. Population d’étude
Deux cent treize (213) enfants de moins de 5 ans ont été recrutés dans cette étude entre Mars
2012 et Mai 2013. Les enfants recrutés présentaient des diarrhées (donner critères définissant
diarrhée) avec des signes cliniques très variés (vomissements, nausées, douleurs
abdominales, déshydrations sévères, manque d’appétit et une malnutrition plus ou moins
sévère).

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 27
Les mères des enfants de moins de 5 ans inclus dans l’étude ont répondu à un questionnaire
où différentes informations sociodémographiques et médicales ont été répertoriées.
L’état nutritionnel des enfants a été évalué à partir des mesures telles le poids, la taille et l’âge
de chaque enfant. Ces mesures sont faites au niveau de la Santé Maternelle et Infantile (SMI)
après la collecte des selles. L’état nutritionnel est déterminé en utilisant la population de
référence internationale définie par le U.S. National Center for Health Statistics (standard
NCHS). Attention, le standard international et national actuel est la table bisex OMS 2006.
Les enfants sont classés selon le Z-score (ou SD-score: standard déviation score). Cette
classification selon le Z-score recommandée par l’OMS et l’UNICEF utilise les indices
nutritionnels poids/taille (WHZ), poids/âge (WAZ), taille/âge (HAZ) et se fait selon les
critères suivants:
Z-score inférieur ou égale à -3: correspond à une malnutrition sévère.
Z-score compris entre -3 et -2: correspond à une malnutrition modérée.
Z-score supérieur ou égal à -2: correspond à un état nutritionnel normal.
3.3. Détection des entéro-pathogènes
3.3.1. Détection des astrovirus
Préparations des échantillons
Nous avons dilué les selles en ajoutant 800µl de solution saline (PBS). Ensuite on a prélevé
100 µl de cette suspension qu’on ajoute 1 ml de PBS pour obtenir la dilution au 1/10 (0,1 mg
de selles/1ml de PBS) dans des tubes polypropylènes de 1,5 ml. Le mélange obtenu est
centrifugé à 12000 g pendant 5 minutes et le surnagent est utilisé pour l’extraction des ARNs
viraux. La préparation peut être conservée à température ambiante pendant 24 h ou à
-20°C pour un temps suffisamment long.
Extraction de L’ARN viral des HAstVs
Les ARNs totaux ont été extraits à l’aide du kit DNA Sorb-B (Sacace Biotechnologies, Como,
Italie) en suivant les recommandations du fabricant. Brièvement l’extraction s’est faite selon
le protocole ci-dessous :
1. Mettre dans les tubes Eppendorf 300µl de solution de lyse, 10 µl de contrôle interne et
100 µl d’échantillon;

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 28
2. Le contrôle négatif d’extraction s’obtient en mélangeant 300µl de la solution de lyse,
10 µl de contrôle interne et 100 µl de contrôle négatif;
3. Le contrôle positif d’extraction est un mélange de 300µl de la solution de lyse, 10µl de
contrôle interne, 90µl de contrôle négatif et 10µl de contrôle positif;
4. Les tubes sont Vortexés, incubé pour 5 mn à 65°C et centrifuger à 12000g pendant 30
sec;
5. Les surnageants sont récupérés dans de nouveaux tubes Eppendorf. On ajoute 20 µl
de Sorbent dans tous les tubes Eppendorf;
6. Les tubes sont Vortexés 5 à 7 sec puis incuber à température ambiante pendant 3
minutes.
7. Répéter l’étape 6;
8. Centrifuger tous les tubes pendant 30 secs à 5000 g. écarté le surnageant;
9. Ajouté 300 µl de la solution de lavage 1 dans les tubes, Vortexer vigoureusement et
centrifuger pendant 30 sec à 8000 g. écarté le surnageant des tubes;
10. Ajouter 500 µl de la solution de lavage 2. Vortexer vigoureusement et centrifuger
pendant 30 sec à 8000g. écarté le surnageant des tubes;
11. Répéter l’étape 10 et incuber tous les tubes bouchons ouvert pour 5 minutes à 65°c
pour sécher le culot;
12. Resuspendre le culot dans 50 µl d’éluant puis incubé tous les tubes pendant 5 minutes
tout en vortexant périodiquement;
13. Centrifuger tous les tubes pour 1 mn à 12000 g;
14. Le surnageant contient les ARNs totaux prêts pour l’amplification ou qui peuvent être
conservés à -20 ou -80°C.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 29
Figure 12:Extraction de l’ARN des Astrovirus au CERBA
Amplification des astrovirus
Le génome viral a été amplifié en utilisant le kit commercial astrovirus 175 PCR kit with
electrophoretic detection (Sacace Biotechnologies, Como, Italie) en suivant les instructions du
fabricant. La rétro-transcription de l’ARN a été faite dans un thermocycleur à 37°C pendant
30 minutes. Les ADNc obtenus ont servi de matrice pour la PCR. Brièvement, Nous avons
mélangé 10 µl de PCR-mix-2 et 10µl d’échantillon (ADNc dilué au ½ avec du TE-buffer)
dans les tubes contenant le PCR-Mix-1. La PCR a été réalisée dans un volume réactionnel de
20 µl dans un Thermocycleur de Gene Ampli PCR system 9700 (Applied Biosystems, USA).
Le programme d’amplification était le suivant : activation de la Taq Polymérase à 94°C
pendant 5 minutes suivi de 42 cycles composés d’une dénaturation pendant dix Secondes
(10s) à 94°C, une hybridation à 67°C pendant une minute (1mn) et une élongation à 72°C
pendant 10 s. Les 42 cycles sont suivis d’une élongation finale à 72°C pendant 1 minute.
Electrophorèse et révélation des produits PCR
La séparation des amplicons a été faite par électrophorèse sur gel d’agarose à 2% et leur
visualisation à l'aide d'un trans-illuminateur UV après coloration par le bromure d’éthidium.
Les échantillons positifs pour les astrovirus présentaient une bande caractéristique à 175 pb.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 30
3.2.6. Détection des parasites
Les préparations de selles sont observées directement entre lame et lamelle à l’état frais et
après coloration.
Principe: L’examen direct a consisté à mettre en évidence les formes végétatives, vivantes et
mobiles des protozoaires ainsi que leurs kystes, mais également les œufs ou les adultes des
helminthes. L’observation de toute la lame se fait d’abord à l’objectif 10× puis à l’objectif
40×. La coloration (au Lugol) permet d’une part de différencier les kystes d’Entamoeba
histolytica (noyaux colorés en jaune) de ceux d’Entamoeba coli, et d’autre part de bien
distinguer les kystes de Giardia (forme du kyste bien nette).
3.2.6.1 Mode opératoire
Les selles ont été prélevé à plusieurs endroits (2 à 3 endroits selon l’aspect des selles), ce
prélèvement a été étalé entre lame et lamelle dans une goutte d’eau physiologique puis
observé au microscope optique respectivement à l’objectif 10× et 40× (la préparation ne doit
pas être épaisse). Une goutte de Lugol a été utilisée pour la coloration de la préparation et
l’observation s’est faite comme précédemment.
3.2.7. Considération éthique
L’étude a été approuvée par le comité d’éthique du CMSC/ CERBA. Les enfants ont été
recrutés après le consentement écrit de leurs parents ou tuteurs.
3.2.8. Analyse statistique
L’analyse des résultats s’est faite à l’aide du logiciel SPSS Software 17.0 (SPSS, Chicago, il,
USA). L’état nutritionnel des enfants a été calculé en utilisant Nutrition StatCal de épi info
3.5.1 (US, Center for Disease Control and prévention (CDC), 2008 et le CDC/who 1978). Les
échantillons positifs ont été comparés en fonction des caractéristiques sociodémographiques
et cliniques en utilisant le test de chi-square ou le test exact de Ficher. La différence a été
considérée significative pour p< 0,05.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 31
RESULTATS

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 32
4. Résultats
4.1. Caractéristiques sociodémographiques et cliniques des enfants
Deux cent treize enfants provenant de Ouagadougou et ses environs ont été inclus dans cette
étude. L’échantillon est composé de 114 (56,5 %) garçons contre 99 (43,5 %) fille pour un
sexe ratio de 1,3. La majorité des enfants soit 198 (92,96%) provenaient de Ouagadougou.
L’âge moyen des enfants était de 15,7 ± 8,5 mois. La taille et le poids moyen des enfants
étaient respectivement de 73,9 ± 9,1 cm et de 7,8 ± 1,9 kg (Tableau 1).
Les enfants ont été réparties dans les trois catégories d’âge suivants: de 0-12 mois, >12-24
mois et de >24-60. La répartition des enfants dans les trois catégories d’âge citées ci-dessus
était respectivement de 40,4%, 44,6 et de 15%. Il aurait été plus approprié 0-11, 12-23 et 24-
59.
Tableau 1 : Répartition des paramètres anthropométriques en fonction des tranches d’âges.
Tranches
d’âge
Effectifs
N (%)
Poids moyen
(en kg)
Taille moyenne
(en cm)
Age moyen
(en mois)
0-12 86 (40,4) 6,8 ± 1,4 68,2 ± 3,8 8,6 ± 2,2
>12-24 95 (44,6) 7,8 ± 1,4 74,9 ± 8,4 17 ± 3,5
>24 30 (15,0) 10,4 ± 1,9 86,9 ± 7,6 32 ± 6,6
Total 211 7,8 ± 1,9 73,9 ± 9,1 15,7 ± 8,5
Les enfants présentaient des signes cliniques très variés en fonction de leur état nutritionnel.
Les signes cliniques les plus fréquents chez les enfants étaient les douleurs abdominales et les
pertes d’appétit avec respectivement 87,7% (186/213) et 67,5% (143) de cas. D’autres signes
cliniques ont été retrouvés chez ces derniers tels que: les vomissements (50%), la fièvre (34%)
et les nausées (16,9%).
Les cas de déshydratations ont été observés chez 139/213 (66,1%) enfants. La fréquence de la
déshydratation modérée et sévère (définir les critères des deux) était respectivement de 49% et
17,1% chez les enfants.
La majorité des mères des enfants diarrhéiques (51,2%) ont consulté quatre jours après le
début de la survenue des diarrhées.
L’état nutritionnel des enfants a aussi été évalué à leur inclusion dans l’étude.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 33
Un retard de croissance et une insuffisance pondérale ont été observés respectivement chez
32,1% (68/213) et 59,0 (125/213) d’enfants. La malnutrition est très fréquente dans toutes les
tranches d’âges (p< 0,001). Je ne comprend pas ce que tu as comparé. Une différence
statiquement significative (entre quoi et quoi ?) est observée quel que soit les paramètres
d’évaluations nutritionnels (WAZ, WAH, HAZ). Donner définition de tous les 3 paramètres.
Le tableau 2 présente la distribution de l’état nutritionnel en fonction des tranches d’âges.
Tableau 2: Répartition de l’état nutritionnel des enfants diarrhéiques en fonction des tranches
d’âges
Tranches d’âge (mois)
0-11 (I)
12-24 (II) 25-60 (III) Total
Effectifs (%)
86 (40,4) 96 (44,6) 30 (15) 212
HAZ
X< -3
-3<X<-2
X>-2
5 (19,2)
12 (28,6)
69 (47,9)
15 (57,7)
27 (64,3)
54 (37,5)
6 (23,1)
3 (7,1)
21 (14,6)
26
42
144
WHZ
X< -3
-3<X<-2
X>-2
11 (28,2)
22 (34,9)
53 (48,2)
26 (66,7)
30 (47,6)
40 (36,4)
2 (5,1)
11 (17,5)
17 (15,4)
39
63
110
WAZ
X< -3
-3<X<-2
X>-2
22 (33,3)
15 (25,5)
49 (56,3)
38 (57,6)
31 (52,5)
27 (31)
6 (9,1)
13 (22,0)
11 (12,7)
66
59
87
Légende : P(X<-3) < 0,001; P (-3<X<-2)= 0,047; P (X>-2) <0,001 ; P (0-12)= 0,003 ;
P (>12-24)= 0,0004; P (>24-60)= 0,033.
4.2. Prévalence des astrovirus et des parasites chez les enfants
Les astrovirus ont été retrouvés chez 14,6% (31/213) des enfants présentant des signes
cliniques de diarrhées. Les patients positifs ont été identifiés à partir de l’électrophore
gramme (Figure 15). La fréquence des parasites était de 20,7% (44/213) chez les enfants
diarrhéiques. Giardia lambia (12,2%) et Trichomonas intestinalis (8%) étaient les parasites
les plus fréquents chez les enfants suivis d’Entamoeba histolytica/dispar (1,9%).

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 34
Figure 13:Electrophorégramme des produits PCR
Légende: M: Marqueur de poids moléculaire, C1: Contrôle négatif d’extraction, C2:Contrôle négatif d’amplification,
C3:Contrôle Positif d’amplification, P1, P5, P6, P7, P9, P10 et P11 sont des patients négatifs, P2, P3, P4, P8 sont les
patients positifs.
4.3. Répartition des astrovirus et des parasites en fonction des caractéristiques
sociodémographiques des enfants
Les infections astrovirales et parasitaires étaient réparties de façon similaire chez les enfants
de sexe masculin et féminin (p = 0,634 et p = 0,228).
La fréquence des astrovirus a été comparée en fonction de l’âge des enfants. Elle était plus
élevée chez les enfants âgés de 0 à 12 mois (45,2%) et ceux dont l’âge est compris entre 12 et
24 mois (41,9%) comparativement aux enfants âgés de 24 à 60 mois (2,9%) (p = 0,006)
(Tableau 5).
La majorité des infections astrovirales (87,1%) était retrouvée chez les enfants âgés de 0 à 24
mois.
Une association a été retrouvée entre l’infection parasitaire et l’âge chez les enfants
diarrhéiques. L’infection parasitaires était plus retrouvée chez les enfants âgés de 12 à 24
mois (52,3%) et >24-60 mois (33,3%) comparativement aux enfants âgés de 0 à 12 mois
(11,1%) (p < 0,001) (Tableau3).
Les diarrhées astrovirales (9,7%) et parasitaires (2,3%) étaient significativement moins
fréquentes chez les enfants nourris exclusivement au sein comparativement aux enfants ayant
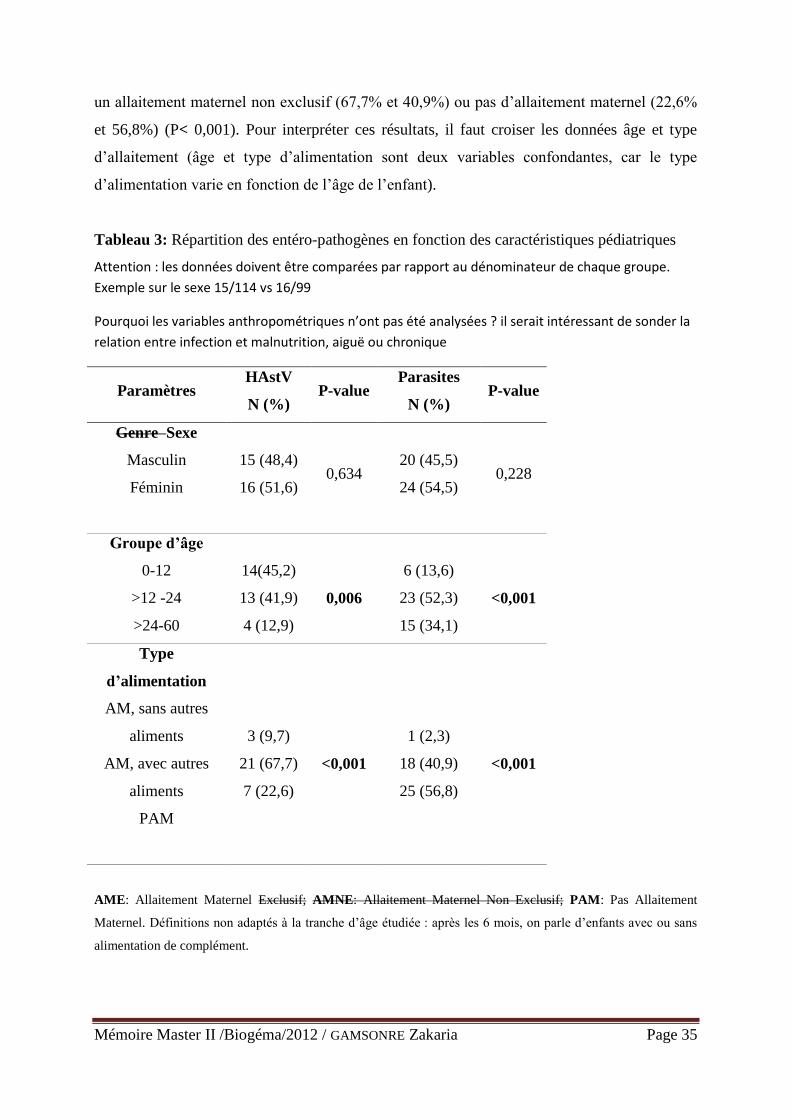
Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 35
un allaitement maternel non exclusif (67,7% et 40,9%) ou pas d’allaitement maternel (22,6%
et 56,8%) (P< 0,001). Pour interpréter ces résultats, il faut croiser les données âge et type
d’allaitement (âge et type d’alimentation sont deux variables confondantes, car le type
d’alimentation varie en fonction de l’âge de l’enfant).
Tableau 3: Répartition des entéro-pathogènes en fonction des caractéristiques pédiatriques
Attention : les données doivent être comparées par rapport au dénominateur de chaque groupe.
Exemple sur le sexe 15/114 vs 16/99
Pourquoi les variables anthropométriques n’ont pas été analysées ? il serait intéressant de sonder la
relation entre infection et malnutrition, aiguë ou chronique
Paramètres HAstV
N (%) P-value
Parasites
N (%) P-value
Genre Sexe
Masculin
Féminin
15 (48,4)
16 (51,6) 0,634
20 (45,5)
24 (54,5) 0,228
Groupe d’âge
0-12
>12 -24
>24-60
14(45,2)
13 (41,9)
4 (12,9)
0,006
6 (13,6)
23 (52,3)
15 (34,1)
<0,001
Type
d’alimentation
AM, sans autres
aliments
AM, avec autres
aliments
PAM
3 (9,7)
21 (67,7)
7 (22,6)
<0,001
1 (2,3)
18 (40,9)
25 (56,8)
<0,001
AME: Allaitement Maternel Exclusif; AMNE: Allaitement Maternel Non Exclusif; PAM: Pas Allaitement
Maternel. Définitions non adaptés à la tranche d’âge étudiée : après les 6 mois, on parle d’enfants avec ou sans
alimentation de complément.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 36
4.4. Répartition des infections astrovirales et parasitaires en fonction des signes cliniques
des enfants diarrhéiques
Les diarrhées astrovirales ne sont associées ni aux vomissements, ni à la perte d’appétit, ni
aux douleurs abdominales ou encore à la déshydratation. Elles sont par contre caractérisées
par une fièvre modérée (p <0,001), une durée moyenne de la diarrhée de 4-5 jours (p<0,001),
un nombre de selles de 1-3 par jour (p<0,001) et une flore bactérienne normale (p< 0,001)
(Tableau4).
Par contre, les diarrhées parasitaires étaient caractérisées respectivement chez 54,3%, 77,3%
et 86,4% d’enfants par la perte d’appétit (p=0,004), les douleurs abdominales (p=0,017) et la
déshydratation (p<0,001). Les diarrhées parasitaires étaient aussi significativement associées
à la forte fièvre, (fièvre >38°C) (79,5%), a un nombre de selles >6 par jours (40,9%) et à une
flore bactérienne anormale (63,6%). Elles n’étaient associées ni aux nausées, ni à la durée de
la diarrhée ou à la déshydratation (Tableau 4).
4.5. L’influence des saisons sur les infections des entéro-pathogènes
L’incidence des maladies diarrhéiques s’observe pendant toute l’année avec de fortes
prévalences observées en saison sèche. Les mois de Juin, Juillet et Août sont associés à
39,91% des incidences contre 47,88% pour les mois de Décembre, Janvier et février. La plus
forte incidence est observée durant le mois de Janvier 25,35% (54/213).Une différence
significative de l’incidence saisonnière est observée au cours de l’année (p<0,001).
La prévalence des astrovirus et des parasites varie selon les saisons. Les astrovirus sont
retrouvés aussi bien en saison sèche que pluvieuse. Un pic d’infection a été observé en saison
sèche froide au mois de Janvier (29,3%). Les infections parasitaires sont présentes tout au
long de l’année avec une fréquence plus élevée en saison pluvieuse, notamment au mois
d’Aout (22,7%).Comme le montre les figures 24 et 25, la prévalence des astrovirus a été
significativement plus élevée en saison sèche qu’en saison pluvieuse (p=0,043) alors que celle
des parasites était plus élevée en saison pluvieuse comparativement à la saison sèche
(p <0,001).

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 37
Tableau 4: Fréquence des astrovirus et des parasites en fonction des signes cliniques des enfants Comme pour le tableau 3,
comparer par rapport au dénominateur de chaque groupe
Signes cliniques Astrovirus
N (%)
P-value Parasites
N (%)
P-value
Fièvre
37,5-38°C
>38°C
21 (67,7)
10 (32,3)
<0,001
9 (20,5)
35 (79,5)
<0,001
Vomissements
Présence
Absence
11 (35,5)
20 (64,5)
0,080
10 (22,7)
34 (79,3)
<0,001
Perte d’appétit
Présence
Absence
20 (64,5)
11 (35,5)
0,706
24 (54,5)
20 (45,5)
0,040
Nausées
Présence
Absence
8 (25,8)
23 (74,2)
0,157
6 (13,6)
38 (86,4)
0,507
Douleurs abdominales
Présence
Absence
29 (93,5)
2 (6,5)
0,286
34 (77,3)
10 (22,7)
0,017
Durée de la diarrhée (jours)
1-3
4-5
>6
8 (25,8)
20 (64,5)
3 (9,7)
<0,001
11(25,0)
22 (50,0)
11(25,0)
0,218
Nbre de selles en 24 h
1-3
17 (54,8)
<0,001
18 (40,9)
0,019
4-5 11 (35,5) 8 (18,2)
> 6
3 (9,7) 18 (40,9)
Déshydratation
sévère
9 (31)
0,101
6 (34,1)
0,772
modérée 12 (41,4) 23 (52,3)
Pas de déshydratation 8 (27,6) 15 (13,6)
Flore bactérienne
Normale
Anormale
23 (74,2)
8 (25,8)
<0,001
16 (36,4)
28 (63,6)
<0,001
Légende: En gras P-values significatif

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 38
Figure 14: L’influence des saisons sur les infections des entéro-pathogènes
Figure 15 : Prévalence saisonnière des Astrovirus et des Parasites

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 39
DISCUSSION

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 40
5. Discussion
La taille de l’échantillon n’est certes pas représentatif de toute la population du Burkina Faso
mais cette étude donne des informations pertinentes sur l’implication des astrovirus et des
Parasites dans les gastro-entérites infantiles. Cette étude avait pour but d’évaluer le rôle des
astrovirus et des parasites dans les diarrhées chez les enfants de moins de 5 ans à
Ouagadougou au Burkina Faso.
La répartition des diarrhées en fonction des tranches d’âge montre une incidence plus élevée
chez les enfants de 12 à 24 mois (44%) que chez ceux âgés de 0 à 12 mois (40,6%). Pour cette
tranche d’age (0- 12) Ouermi et al.,2007 et Sanou et al., 1999 ont rapportés des incidences
diarrhéiques supérieurs aux notre respectivement mois des incidences 47 et 56,0%.
Dans cette étude nous rapportons pour la première fois au Burkina Faso, la prévalence des
astrovirus chez les enfants diarrhéiques de moins de 5 ans. Nous avons trouvé une prévalence
astrovirale de 14,6%. Cette prévalence est comprise entre les prévalences de 2 et 16%
rapportées par plusieurs études antérieures (Hermann et al, 1991; Pennap et al, 2002; Bini et
al, 2007 ; Hamkar et al, 2010). Cependant, des prévalences astrovirales supérieures à 33% et
37% ont été rapportées respectivement au Brésil (Silva et al, 2001) et en Afrique du sud
(Taylor et al, 1997). Cette grande différence relevée par Taylor pourrait s’expliquer par la
taille de notre échantillon.
Les infections astrovirales chez les enfants diarrhéiques étaient plus fréquentes (45,2%) dans
la tranche de 0 à 12 mois comparativement à la tranche d’âge >24 mois. La prévalence des
astrovirus était égale à 88,1% chez les enfants âgés d’au plus 24 mois. Nos résultats
corroborent ceux de Hamkar et al, 2010 en Iran, de Mendez-Toss et al, 2004 au Mexico , en
Thaïlande (Hermann et al 1991), au Guatemala, en France (Bon et al, 1999), en Australie
(Mustapha et al, 2000), au Chilie (Gaggero et al, 1998), en Colombie et au Venezuela.
Ces forts taux s’expliqueraient par une contamination massive à travers les aliments et les
eaux de boisson souillées du fait de la résistance des astrovirus aux facteurs
environnementaux.
La prévalence des infections parasitaires dans les cas de diarrhées chez les enfants était de
20,7%. Cette prévalence est supérieure aux prévalences de 10,7 et 18,2% rapportées dans de
précédentes études faites au Burkina Faso (Sanou et al., 1999; Ouermi et al, 2007).
Cependant, elle est inférieure à la prévalence de 22,7% récemment rapportée en Ethiopie par
Amare et al.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 41
La prévalence des infections parasitaires était particulièrement élevée (85,4%) chez les
enfants diarrhéiques âgés de plus de 12 mois. Les études antérieures ont rapportées de fortes
prévalences dans cette tranche au Burkina Faso (Ouermi et al., 2007; Nitiéma et al., 2011).
La prédominance des parasites dans ce groupe d'âge s’expliquerait par la transmission des
agents pathogènes par voie orale, par les aliments souillés, par la voie transcutanée et par la
terre souillée de larves.
Nous avons observé une forte saisonnalité dans la prévalence des parasites chez les enfants
inclus dans cette étude. Une plus forte prévalence parasitaire a été observée pendant la saison
pluvieuse de Juin à Août. Gliardia lamblia, le parasite le plus fréquent (12,2%) identifié chez
les enfants est connu pour être omniprésents dans l'eau et les aliments au Burkina Faso
(Ouermi et al., 2009).
Les infections à astrovirus sont très fréquentes tant en période chaude que froide (Lin et al.,
2008, Liu et al., 2006, Walter et al., 2003). Pendant la saison froide et sèche de Décembre à
Février nous avons observés 48,39% (15/31) des infections astrovirales, ce qui démontre la
forte saisonnalité des infections à astrovirus. En effet une prévalence 41,94% a été détectée
pendant la période chaude (Juin, Juillet et Août). Ces résultats sont comparables à ceux de
Hermann et al., 1991, Noel et al., 1994 et de Lin et al., en (2008) à Taipei. Egalement au
cours de cette période la prévalence des infections parasitaires étaient abondantes (67,9%).
Les variations saisonnières des zoonoses entériques résultent d'une complexe interaction entre
les facteurs climatiques et non climatiques saisonnières qui agissent à différentes échelles
temporo-spatiales et qui interagissent continuellement (Lal et al., 2012). Les diarrhées
d’origine virales et parasitaires sont plus fréquentes au cours des saisons sèches et froides
(Levy et al., 2009).
Cette saisonnalité est plus marquée dans les pays à climat tempéré que dans les pays
tropicaux. Ces différences observées pourraient être dues aux faibles variabilités climatiques
dans les pays tropicaux par rapport à celles des pays tempérés. La fréquence élevée
d’astrovirus pendant la saison sèche pourrait également s'expliquer par une baisse de
l'humidité et des précipitations favorisant l'assèchement du sol augmentant le transport aérien
de particules fécales sèches contaminées. Ce changement pourrait favoriser aussi la formation
de poussières, lesquelles fournissent des substrats aux particules virales (Levy et al., 2009).
La baisse de matières organiques pendant la saison sèche peut augmenter la quantité de
particules dans l'air, lesquelles peuvent contaminer l'eau ou les surfaces de l’environnement
ou être ingérées.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 42
L’allaitement maternel a été associé à une diminution des taux de la diarrhée chez les
nourrissons dans les pays développés et en développement (Dewey et al., 1995). L’allaitement
maternel confère une protection efficace aux infections astrovirales. Les enfants qui ont un
allaitement mixte sont les plus atteints 67,7% suivi de ceux qui non pas été allaités 22,6% et
enfin les enfants allaités au lait maternel exclusivement 9,7%. L’allaitement exclusif au sein
pendant une période de six mois présente bien des avantages pour le nourrisson et la mère, et
notamment l’insigne avantage de protéger contre les infections gastro-intestinales, tant dans
les pays en développement que dans les pays industrialisés.
La proportion de diarrhée à astrovirus supérieur à 3 jours est observée dans 74,2% des cas.
Ces résultats sont conformes à ceux de Tseng et al., 2011 en Taiwan. Mais il n’existe pas de
différences significatives entre la durée des diarrhées astrovirales. Les astrovirus
prolongeraient donc la durée des diarrhées qui conduirait à une déshydratation pouvant se
compliqués de choc hypovolémique.
Dans notre étude 40,8% des enfants ont subi une consultation entre 1 à 3 jours de début de
diarrhée. En effet un taux de 52,2% de consultation après les trois premiers jours de diarrhée
témoigne de la banalisation de la diarrhée et la méconnaissance de ces conséquences par les
mères. Nos résultats concordent avec ceux de Sanou et al (1999) au Burkina Faso.
Les infections astrovirales ne sont pas associées à la flore bactérienne.
Le taux de malnutrition de 59,4% est respectivement supérieur à celui de Simporé et al (2009)
au Burkina Faso (35,1%), de Sanou et al (1999) au Burkina Faso (32,5 %). Les enfants de 12
à 24 mois sont les plus atteints. De nombreuses études ont montré que la malnutrition a un
effet sur la durée des épisodes diarrhéiques, ainsi les enfants malnutris ont des épisodes
diarrhéiques plus longs (Black et al., 1984, Bairagi et al., 1987) et une maladie plus grave
(Guerrant R L et al., 2008). Une cause importante de la malnutrition et la mortalité infantile
serait le faible taux d'allaitement maternel au Burkina Faso, un taux qui décline rapidement
dans les premiers mois de la vie d'un bébé selon le rapport de l’enquête nutritionnelle
(SMART, 2009). De plus, les épisodes diarrhéiques peuvent provoquer un retard de
croissance chez les enfants (Assis et al., 2005; Chechley et al., 2003). La relation
bidirectionnelle entre malnutrition et la diarrhée constitue un problème car d’une part, les
épisodes diarrhéiques peuvent avoir un impact sur le statut nutritionnel et d’autre part la
malnutrition favorise les infections intestinales et les diarrhées. Ce cercle vicieux entre
diarrhées et malnutrition constitue un problème notamment dans les pays en développements
(PED). Il a été estimé qu’1/4 des enfants souffre de malnutrition dans les PED (Thapar et al.,

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 43
2004), un (1) enfant sur deux (2) est atteint de malnutrition protéino-énergetique (MPE) en
Asie du sud et 30% en Afrique subsaharienne (FAO., 2012).

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 44
CONCLUSION ET PERSPECTIVES

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 45
6. Conclusion
Les maladies diarrhéiques dont la principale manifestation est la diarrhée constituent un
problème de santé publique pour les pays en développements et le Burkina Faso en
particulier. Cette étude nous a permis de d’établir les fréquences du virus et de micro-
organismes responsable justement de ces maladies diarrhéique par la méthode de la RT-PCR
une prévalence de 14,6% d’astrovirus a été notée. Les parasites intestinaux isolés par la
microscopie constituent 20,7% des étiologies infectieuses avec une plus grande fréquence de
Protozoaires (20,2%) que d’Helminthes (1,9%). Les astrovirus ont été isolés principalement
chez les enfants de moins de 12 mois et les parasites intestinaux chez les enfants de plus de 12
moins. La malnutrition qui reste le principal facteur d’aggravation est associée à 59,7% aux
gastro-entérites dans notre étude. La répartition d’astrovirus est saisonnière avec une forte
prévalence en saison sèche. Une observation de la propriété individuelle et collective réduirait
la malnutrition et par conséquent la mortalité infantile. En effet une connaissance de
l’ensemble des étiologies impliquées dans les diarrhées est nécessaire pour une lutte efficace
des gastro-entérites infantiles.
PERSPECTIVES
Au regard des gastro-entérites occasionner par la malnutrition et les infections
multiples en particulier les astrovirus et les parasites en guise de perspective nous proposons
de:
Identifier les différents génotypes des astrovirus circulants au Burkina Faso à travers
des recherches moléculaires.
Élargir l’étude aux autres Centres de Récupération et d’Éducation Nutritionnelle
(CREN) du Burkina Faso pour avoir le maximum d’informations.
Effectuer une étude de mise en évidence de l’ensemble des étiologies virale,
parasitaires et bactériennes de même que tous leurs génotypes.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 46
REFERENCES

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 47
7. Références
ABAD, F. X., PINTO, R. M., VILLENA, C., GAJARDO, R. & BOSCH, A. (1997)
Astrovirus survival in drinking water. Appl Environ Microbiol, 63, 3119-22.
AHMED, S. F., SEBENY, P. J., KLENA, J. D., PIMENTEL, G., MANSOUR, A., NAGUIB,
A. M., BRUTON, J., YOUNG, S. Y., HOLTZ, L. R. & WANG, D. (2011) Novel
astroviruses in children, Egypt. Emerg Infect Dis, 17, 2391-3.
ALI, S. A. & HILL, D. R. (2003) Giardia intestinalis. Curr Opin Infect Dis, 16, 453-60.
APPLETON, H. & HIGGINS, P. G. (1975) Letter: Viruses and gastroenteritis in infants.
Lancet, 1, 1297.
BAJOLET, O. & CHIPPAUX-HYPPOLITE, C. (1998) [Rotavirus and other viruses of
diarrhea]. Bull Soc Pathol Exot, 91, 432-7.
BELLIOT, G., LAVERAN, H. & MONROE, S. S. (1997) Capsid protein composition of
reference strains and wild isolates of human astroviruses. Virus Res, 49, 49-57.
BERN, C., MARTINES, J., DE ZOYSA, I. & GLASS, R. I. (1992) The magnitude of the
global problem of diarrhoeal disease: a ten-year update. Bull World Health Organ, 70,
705-14.
BINI, J. C., EKAZA, E., FAYE-KETTE, H., VEH, K. A., NIGUE, L., BORGET-ALLOUE,
M. Y., AKRAN, A. V. & DOSSO, M. (2007) [Detection by RT-PCR of the 1st cases
of Astrovirus in human stools in Abidjan, Cote d'Ivoire]. Bull Soc Pathol Exot, 100,
243-5.
BISHOP, R. F. (1996) Natural history of human rotavirus infection. Arch Virol Suppl, 12,
119-28.
BLACK, R. E., BROWN, K. H. & BECKER, S. (1984) Malnutrition is a determining factor
in diarrheal duration, but not incidence, among young children in a longitudinal study
in rural Bangladesh. Am J Clin Nutr, 39, 87-94.
BON, F., FASCIA, P., DAUVERGNE, M., TENENBAUM, D., PLANSON, H., PETION, A.
M., POTHIER, P. & KOHLI, E. (1999) Prevalence of group A rotavirus, human
calicivirus, astrovirus, and adenovirus type 40 and 41 infections among children with
acute gastroenteritis in Dijon, France. J Clin Microbiol, 37, 3055-8.
BRANSKI, D., SHARON, P., KARMELI, F. & RACHMILEWITZ, D. (1984) Effect of
cimetidine on human gastric and duodenal prostanoid synthesis. Scand J
Gastroenterol, 19, 457-60.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 48
BRIDGER, J. C. (1980) Detection by electron microscopy of caliciviruses, astroviruses and
rotavirus-like particles in the faeces of piglets with diarrhoea. Vet Rec, 107, 532-3.
CATTOLI, G., DE BATTISTI, C., TOFFAN, A., SALVIATO, A., LAVAZZA, A.,
CERIOLI, M. & CAPUA, I. (2007) Co-circulation of distinct genetic lineages of
astroviruses in turkeys and guinea fowl. Arch Virol, 152, 595-602.
CHECKLEY, W., EPSTEIN, L. D., GILMAN, R. H., CABRERA, L. & BLACK, R. E.
(2003) Effects of acute diarrhea on linear growth in Peruvian children. Am J
Epidemiol, 157, 166-75.
CHU, D. K., CHIN, A. W., SMITH, G. J., CHAN, K. H., GUAN, Y., PEIRIS, J. S. & POON,
L. L. (2010) Detection of novel astroviruses in urban brown rats and previously known
astroviruses in humans. J Gen Virol, 91, 2457-62.
CHU, D. K., POON, L. L., GUAN, Y. & PEIRIS, J. S. (2008) Novel astroviruses in
insectivorous bats. J Virol, 82, 9107-14.
CLARK, B. & MCKENDRICK, M. (2004) A review of viral gastroenteritis. Curr Opin Infect
Dis, 17, 461-9.
COPPO, P., SCIEUX, C., FERCHAL, F., CLAUVEL, J. & LASSOUED, K. (2000)
Astrovirus enteritis in a chronic lymphocytic leukemia patient treated with fludarabine
monophosphate. Ann Hematol, 79, 43-5.
CUBITT, W. D. (1996) Historical background and classification of caliciviruses and
astroviruses. Arch Virol Suppl, 12, 225-35.
CUBITT, W. D., MITCHELL, D. K., CARTER, M. J., WILLCOCKS, M. M. & HOLZEL, H.
(1999) Application of electronmicroscopy, enzyme immunoassay, and RT-PCR to
monitor an outbreak of astrovirus type 1 in a paediatric bone marrow transplant unit. J
Med Virol, 57, 313-21.
DAY, J. M., SPACKMAN, E. & PANTIN-JACKWOOD, M. (2007) A multiplex RT-PCR
test for the differential identification of turkey astrovirus type 1, turkey astrovirus type
2, chicken astrovirus, avian nephritis virus, and avian rotavirus. Avian Dis, 51, 681-4.
DE BENEDICTIS, P., SCHULTZ-CHERRY, S., BURNHAM, A. & CATTOLI, G. (2011)
Astrovirus infections in humans and animals - molecular biology, genetic diversity,
and interspecies transmissions. Infect Genet Evol, 11, 1529-44.
DE GRAZIA, S., PLATIA, M. A., ROTOLO, V., COLOMBA, C., MARTELLA, V. &
GIAMMANCO, G. M. (2011) Surveillance of human astrovirus circulation in Italy
2002-2005: emergence of lineage 2c strains. Clin Microbiol Infect, 17, 97-101.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 49
DOMANSKA-BLICHARZ, K., SEROKA, A. & MINTA, Z. (2011) One-year molecular
survey of astrovirus infection in turkeys in Poland. Arch Virol, 156, 1065-72.
DONG, J., DONG, L., MENDEZ, E. & TAO, Y. (2011) Crystal structure of the human
astrovirus capsid spike. Proc Natl Acad Sci U S A, 108, 12681-6.
ERNESTO MÉNDEZ, A. M., RODRIGO VELÁZQUEZ, ANDREW BURNHAM,AND
CARLOS F. ARIAS (2013) Replication Cycle of Astroviruses.
FINKBEINER, S. R., LE, B. M., HOLTZ, L. R., STORCH, G. A. & WANG, D. (2009)
Detection of newly described astrovirus MLB1 in stool samples from children. Emerg
Infect Dis, 15, 441-4.
GAGGERO, A., O'RYAN, M., NOEL, J. S., GLASS, R. I., MONROE, S. S., MAMANI, N.,
PRADO, V. & AVENDANO, L. F. (1998) Prevalence of astrovirus infection among
Chilean children with acute gastroenteritis. J Clin Microbiol, 36, 3691-3.
GLASS, R. I., BRESEE, J., JIANG, B., GENTSCH, J., ANDO, T., FANKHAUSER, R.,
NOEL, J., PARASHAR, U., ROSEN, B. & MONROE, S. S. (2001) Gastroenteritis
viruses: an overview. Novartis Found Symp, 238, 5-19; discussion 19-25.
GLASS, R. I., BRESEE, J., JIANG, B., PARASHAR, U., YEE, E. & GENTSCH, J. (2006)
Rotavirus and rotavirus vaccines. Adv Exp Med Biol, 582, 45-54.
GLASS, R. I., NOEL, J., MITCHELL, D., HERRMANN, J. E., BLACKLOW, N. R.,
PICKERING, L. K., DENNEHY, P., RUIZ-PALACIOS, G., DE GUERRERO, M. L.
& MONROE, S. S. (1996) The changing epidemiology of astrovirus-associated
gastroenteritis: a review. Arch Virol Suppl, 12, 287-300.
GROHMANN, G. S., GLASS, R. I., PEREIRA, H. G., MONROE, S. S., HIGHTOWER, A.
W., WEBER, R. & BRYAN, R. T. (1993) Enteric viruses and diarrhea in HIV-
infected patients. Enteric Opportunistic Infections Working Group. N Engl J Med,
329, 14-20.
GUERRANT, R. L., SCHORLING, J. B., MCAULIFFE, J. F. & DE SOUZA, M. A. (1992)
Diarrhea as a cause and an effect of malnutrition: diarrhea prevents catch-up growth
and malnutrition increases diarrhea frequency and duration. Am J Trop Med Hyg, 47,
28-35.
GUERRERO, M. L., NOEL, J. S., MITCHELL, D. K., CALVA, J. J., MORROW, A. L.,
MARTINEZ, J., ROSALES, G., VELAZQUEZ, F. R., MONROE, S. S., GLASS, R.
I., PICKERING, L. K. & RUIZ-PALACIOS, G. M. (1998) A prospective study of
astrovirus diarrhea of infancy in Mexico City. Pediatr Infect Dis J, 17, 723-7.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 50
GUIX, S., BOSCH, A. & PINTÓ, R. M. (2007) Structure-based Study of Viral Replication.
GUIX, S., CABALLERO, S., BOSCH, A. & PINTO, R. M. (2004) C-terminal nsP1a protein
of human astrovirus colocalizes with the endoplasmic reticulum and viral RNA.
J Virol, 78, 13627-36.
GUIX, S., CABALLERO, S., FUENTES, C., BOSCH, A. & PINTO, R. M. (2008) Genetic
analysis of the hypervariable region of the human astrovirus nsP1a coding region:
design of a new RFLP typing method. J Med Virol, 80, 306-15.
GUIX, S., CABALLERO, S., VILLENA, C., BARTOLOME, R., LATORRE, C.,
RABELLA, N., SIMO, M., BOSCH, A. & PINTO, R. M. (2002) Molecular
epidemiology of astrovirus infection in Barcelona, Spain. J Clin Microbiol, 40, 133-9.
GUO, L., XU, X., SONG, J., WANG, W., WANG, J. & HUNG, T. (2010) Molecular
characterization of astrovirus infection in children with diarrhea in Beijing, 2005-
2007. J Med Virol, 82, 415-23.
HAMKAR, R., YAHYAPOUR, Y., NOROOZI, M., NOURIJELYANI, K., JALILVAND, S.,
ADIBI, L., VAZIRI, S., POOR-BABAEI, A., PAKFETRAT, A. & SAVAD-KOOHI,
R. (2010) Prevalence of Rotavirus, Adenovirus, and Astrovirus Infections among
Patients with Acute Gastroenteritis in, Northern Iran. Iran J Public Health, 39, 45-51.
JAKAB, F., MELEG, E., BANYAI, K., MELEGH, B., TIMAR, L., PETERFAI, J. &
SZUCS, G. (2004) One-year survey of astrovirus infection in children with
gastroenteritis in a large hospital in Hungary: occurrence and genetic analysis of
astroviruses. J Med Virol, 74, 71-7.
JAKAB, G., MANRIQUE, A., ZIMMERLI, L., METRAUX, J. P. & MAUCH-MANI, B.
(2003) Molecular characterization of a novel lipase-like pathogen-inducible gene
family of Arabidopsis. Plant Physiol, 132, 2230-9.
JEONG, A. Y., JEONG, H. S., JO, M. Y., JUNG, S. Y., LEE, M. S., LEE, J. S., JEE, Y. M.,
KIM, J. H. & CHEON, D. S. (2011) Molecular epidemiology and genetic diversity of
human astrovirus in South Korea from 2002 to 2007. Clin Microbiol Infect, 17, 404-8.
JEONG, H. S., JEONG, A. & CHEON, D. S. (2012) Epidemiology of astrovirus infection in
children. Korean J Pediatr, 55, 77-82.
JONASSEN, C. M., JONASSEN, T. O., SAIF, Y. M., SNODGRASS, D. R., USHIJIMA, H.,
SHIMIZU, M. & GRINDE, B. (2001) Comparison of capsid sequences from human
and animal astroviruses. J Gen Virol, 82, 1061-7.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 51
JONASSEN, C. M., JONASSEN, T. T., SVEEN, T. M. & GRINDE, B. (2003) Complete
genomic sequences of astroviruses from sheep and turkey: comparison with related
viruses. Virus Res, 91, 195-201.
KAPOOR, A., LI, L., VICTORIA, J., ODERINDE, B., MASON, C., PANDEY, P., ZAIDI, S.
Z. & DELWART, E. (2009) Multiple novel astrovirus species in human stool. J Gen
Virol, 90, 2965-72.
KHUROO, M. S. (1996) Ascariasis. Gastroenterol Clin North Am, 25, 553-77.
KOCI, M. D., KELLEY, L. A., LARSEN, D. & SCHULTZ-CHERRY, S. (2004) Astrovirus-
induced synthesis of nitric oxide contributes to virus control during infection. J Virol,
78, 1564-74.
KOCI, M. D., MOSER, L. A., KELLEY, L. A., LARSEN, D., BROWN, C. C. & SCHULTZ-
CHERRY, S. (2003) Astrovirus induces diarrhea in the absence of inflammation and
cell death. J Virol, 77, 11798-808.
KOCI, M. D., SEAL, B. S. & SCHULTZ-CHERRY, S. (2000) Molecular characterization of
an avian astrovirus. J Virol, 74, 6173-7.
KOMORIYA, T., KOHNO, H., KIMURA, A. & USHIJIMA, H. (2003) The development of
sensitive latex agglutination tests for detecting astroviruses (serotypes 1 and 3) from
clinical stool specimen. Rinsho Biseibutshu Jinsoku Shindan Kenkyukai Shi, 13, 103-
14.
KOOPMANS, M. P., BIJEN, M. H., MONROE, S. S. & VINJE, J. (1998) Age-stratified
seroprevalence of neutralizing antibodies to astrovirus types 1 to 7 in humans in The
Netherlands. Clin Diagn Lab Immunol, 5, 33-7.
KOSEK, M., BERN, C. & GUERRANT, R. L. (2003) The global burden of diarrhoeal
disease, as estimated from studies published between 1992 and 2000. Bull World
Health Organ, 81, 197-204.
KRISHNA, N. K. (2005) Identification of structural domains involved in astrovirus capsid
biology. Viral Immunol, 18, 17-26.
LAL, A., HALES, S., FRENCH, N. & BAKER, M. G. (2012) Seasonality in human zoonotic
enteric diseases: a systematic review. PLoS One, 7, e31883.
LAURIN, M. A., DASTOR, M. & L'HOMME, Y. (2011) Detection and genetic
characterization of a novel pig astrovirus: relationship to other astroviruses. Arch
Virol, 156, 2095-9.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 52
LETTERIO, J. J. & ROBERTS, A. B. (1998) Regulation of immune responses by TGF-beta.
Annu Rev Immunol, 16, 137-61.
LEVY, K., HUBBARD, A. E. & EISENBERG, J. N. (2009) Seasonality of rotavirus disease
in the tropics: a systematic review and meta-analysis. Int J Epidemiol, 38, 1487-96.
LI, L., SHAN, T., WANG, C., COTE, C., KOLMAN, J., ONIONS, D., GULLAND, F. M. &
DELWART, E. The fecal viral flora of California sea lions. J Virol, 85, 9909-17.
LIN, H. C., KAO, C. L., CHANG, L. Y., HSIEH, Y. C., SHAO, P. L., LEE, P. I., LU, C. Y.,
LEE, C. Y. & HUANG, L. M. (2008) Astrovirus gastroenteritis in children in Taipei. J
Formos Med Assoc, 107, 295-303.
LIU, C., GRILLNER, L., JONSSON, K., LINDE, A., SHEN, K., LINDELL, A. T.,
WIRGART, B. Z. & JOHANSEN, K. (2006) Identification of viral agents associated
with diarrhea in young children during a winter season in Beijing, China. J Clin Virol,
35, 69-72.
LIU, M. Q., PENG, J. S., TANG, L., ZHOU, Y., YANG, B. F., WANG, Y. H., WANG, B.,
ZHOU, D. J., HUANG, H. J. & HO, W. Z. (2008) Identification of new subtype of
astrovirus type 3 from an infant with diarrhea in Wuhan, China. Virology, 375, 301-6.
LUO, Z., ROI, S., DASTOR, M., GALLICE, E., LAURIN, M. A. & L'HOMME, Y. (2011)
Multiple novel and prevalent astroviruses in pigs. Vet Microbiol, 149, 316-23.
MADELEY, C. R. & COSGROVE, B. P. (1975) Letter: 28 nm particles in faeces in infantile
gastroenteritis. Lancet, 2, 451-2.
MATSUI, S. M., KIANG, D., GINZTON, N., CHEW, T. & GEIGENMULLER-GNIRKE, U.
(2001) Molecular biology of astroviruses: selected highlights. Novartis Found Symp,
238, 219-33; discussion 233-6.
MBAE, C. K., NOKES, J., MULINGE, E., NYAMBURA, J., WARURU, A. & KARIUKI, S.
(2013) Intestinal parasitic infections in children presenting with diarrhoea in outpatient
and inpatient settings in an informal settlement of Nairobi, Kenya. BMC Infect Dis, 13,
243.
MÉNDEZ, E., , A. M., VELÁZQUEZ, R., , A. B. & ARIAS, A. C. F. (2013) Replication
Cycle of Astroviruses, New York, Springer.
MENDEZ, E., AGUIRRE-CRESPO, G., ZAVALA, G. & ARIAS, C. F. (2007) Association
of the astrovirus structural protein VP90 with membranes plays a role in virus
morphogenesis. J Virol, 81, 10649-58.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 53
MENDEZ, E., FERNANDEZ-LUNA, T., LOPEZ, S., MENDEZ-TOSS, M. & ARIAS, C. F.
(2002) Proteolytic processing of a serotype 8 human astrovirus ORF2 polyprotein. J
Virol, 76, 7996-8002.
MENDEZ, E., SALAS-OCAMPO, E. & ARIAS, C. F. (2004) Caspases mediate processing of
the capsid precursor and cell release of human astroviruses. J Virol, 78, 8601-8.
MENDEZ-TOSS, M., GRIFFIN, D. D., CALVA, J., CONTRERAS, J. F., PUERTO, F. I.,
MOTA, F., GUISCAFRE, H., CEDILLO, R., MUNOZ, O., HERRERA, I., LOPEZ, S.
& ARIAS, C. F. (2004) Prevalence and genetic diversity of human astroviruses in
Mexican children with symptomatic and asymptomatic infections. J Clin Microbiol,
42, 151-7.
MENDEZ-TOSS, M., ROMERO-GUIDO, P., MUNGUIA, M. E., MENDEZ, E. & ARIAS,
C. F. (2000) Molecular analysis of a serotype 8 human astrovirus genome. J Gen
Virol, 81, 2891-7.
MENSENKAMP, A. R., JONG, M. C., VAN GOOR, H., VAN LUYN, M. J., BLOKS, V.,
HAVINGA, R., VOSHOL, P. J., HOFKER, M. H., VAN DIJK, K. W., HAVEKES, L.
M. & KUIPERS, F. (1999) Apolipoprotein E participates in the regulation of very low
density lipoprotein-triglyceride secretion by the liver. J Biol Chem, 274, 35711-8.
MITCHELL, D. K. (2002) Astrovirus gastroenteritis. Pediatr Infect Dis J, 21, 1067-9.
MITCHELL, D. K., MATSON, D. O., JIANG, X., BERKE, T., MONROE, S. S., CARTER,
M. J., WILLCOCKS, M. M. & PICKERING, L. K. (1999) Molecular epidemiology of
childhood astrovirus infection in child care centers. J Infect Dis, 180, 514-7.
MONDAL, D., MINAK, J., ALAM, M., LIU, Y., DAI, J., KORPE, P., LIU, L., HAQUE, R.
& PETRI, W. A., JR. (2012) Contribution of enteric infection, altered intestinal barrier
function, and maternal malnutrition to infant malnutrition in Bangladesh. Clin Infect
Dis, 54, 185-92.
MONROE, S. S., JIANG, B., STINE, S. E., KOOPMANS, M. & GLASS, R. I. (1993)
Subgenomic RNA sequence of human astrovirus supports classification of
Astroviridae as a new family of RNA viruses. J Virol, 67, 3611-4.
MOR, S. K., CHANDER, Y., MARTHALER, D., PATNAYAK, D. P. & GOYAL, S. M.
(2012) Detection and molecular characterization of Porcine astrovirus strains
associated with swine diarrhea. J Vet Diagn Invest, 24, 1064-7.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 54
MOSER, L. A., CARTER, M. & SCHULTZ-CHERRY, S. (2007) Astrovirus increases
epithelial barrier permeability independently of viral replication. J Virol, 81, 11937-
45.
MOSER, L. A. & SCHULTZ-CHERRY, S. (2008) Suppression of astrovirus replication by
an ERK1/2 inhibitor. J Virol, 82, 7475-82.
MOYO, S. J., GRO, N., KIRSTI, V., MATEE, M. I., KITUNDU, J., MASELLE, S. Y.,
LANGELAND, N. & MYRMEL, H. (2007) Prevalence of enteropathogenic viruses
and molecular characterization of group A rotavirus among children with diarrhea in
Dar es Salaam Tanzania. BMC Public Health, 7, 359.
MUSTAFA, H., PALOMBO, E. A. & BISHOP, R. F. (2000) Epidemiology of astrovirus
infection in young children hospitalized with acute gastroenteritis in Melbourne,
Australia, over a period of four consecutive years, 1995 to 1998. J Clin Microbiol, 38,
1058-62.
NAFICY, A. B., RAO, M. R., HOLMES, J. L., ABU-ELYAZEED, R., SAVARINO, S. J.,
WIERZBA, T. F., FRENCK, R. W., MONROE, S. S., GLASS, R. I. & CLEMENS, J.
D. (2000) Astrovirus diarrhea in Egyptian children. J Infect Dis, 182, 685-90.
NITIEMA, L. W., NORDGREN, J., OUERMI, D., DIANOU, D., TRAORE, A. S.,
SVENSSON, L. & SIMPORE, J. (2011) Burden of rotavirus and other
enteropathogens among children with diarrhea in Burkina Faso. Int J Infect Dis, 15,
e646-52.
NOEL, J. S., LEE, T. W., KURTZ, J. B., GLASS, R. I. & MONROE, S. S. (1995) Typing of
human astroviruses from clinical isolates by enzyme immunoassay and nucleotide
sequencing. J Clin Microbiol, 33, 797-801.
OLESEN, B., NEIMANN, J., BOTTIGER, B., ETHELBERG, S., SCHIELLERUP, P.,
JENSEN, C., HELMS, M., SCHEUTZ, F., OLSEN, K. E., KROGFELT, K.,
PETERSEN, E., MOLBAK, K. & GERNER-SMIDT, P. (2005) Etiology of diarrhea
in young children in Denmark: a case-control study. J Clin Microbiol, 43, 3636-41.
PENNAP, G., PAGER, C. T., PEENZE, I., DE BEER, M. C., KWAGA, J. K., OGALLA, W.
N., UMOH, J. U. & STEELE, A. D. (2002) Epidemiology of astrovirus infection in
Zaria, Nigeria. J Trop Pediatr, 48, 98-101.
PIGNATELLI, S., SIMPORE, J., RUGGIERI, M. & MUSUMECI, S. (2000) Effectiveness of
forced rehydration and early re-feeding in the treatment of acute diarrhoea in a tropical
area. Minerva Pediatr, 52, 357-66.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 55
PINTO, R. M., ABAD, F. X., GAJARDO, R. & BOSCH, A. (1996) Detection of infectious
astroviruses in water. Appl Environ Microbiol, 62, 3073.
POLLOK, R. C. (2001) Viruses causing diarrhoea in AIDS. Novartis Found Symp, 238, 276-
83; discussion 283-8.
QUAN, P. L., WAGNER, T. A., BRIESE, T., TORGERSON, T. R., HORNIG, M.,
TASHMUKHAMEDOVA, A., FIRTH, C., PALACIOS, G., BAISRE-DE-LEON, A.,
PADDOCK, C. D., HUTCHISON, S. K., EGHOLM, M., ZAKI, S. R., GOLDMAN,
J. E., OCHS, H. D. & LIPKIN, W. I. (2010) Astrovirus encephalitis in boy with X-
linked agammaglobulinemia. Emerg Infect Dis, 16, 918-25.
RIVERA, R., NOLLENS, H. H., VENN-WATSON, S., GULLAND, F. M. & WELLEHAN,
J. F., JR. (2010) Characterization of phylogenetically diverse astroviruses of marine
mammals. J Gen Virol, 91, 166-73.
SANCHEZ-FAUQUIER, A., CARRASCOSA, A. L., CARRASCOSA, J. L., OTERO, A.,
GLASS, R. I., LOPEZ, J. A., SAN MARTIN, C. & MELERO, J. A. (1994)
Characterization of a human astrovirus serotype 2 structural protein (VP26) that
contains an epitope involved in virus neutralization. Virology, 201, 312-20.
SANOU, I., KAM, K.L., TOUGOUMA, A., SANGARE.L.,. NIKIEMA. J.H.P., SANOU. I.,
& KOUETA. F., D. L., SAWADOGO. S.A., SOUDRE. R.B. (1999) DIARRHEES
AIGUES DE L’ENFANT : ASPECTS EPIDEMIOLOGIQUES, CLINIQUES ET
EVOLUTIFS EN MILIEU HOSPITALIER PEDIATRIQUE A OUAGADOUGOU.
Médecine d'Afrique Noire.
SANTOS, R. A., BORGES, A. M., DA COSTA, P. S., TEIXEIRA, J. M., GIUGLIANO, L.
G., LEITE, J. P. & CARDOSO, D. (2007) Astrovirus infection in children living in
the Central West region of Brazil. Mem Inst Oswaldo Cruz, 102, 209-13.
SAVIOLI, L., SMITH, H. & THOMPSON, A. (2006) Giardia and Cryptosporidium join the
'Neglected Diseases Initiative'. Trends Parasitol, 22, 203-8.
SCHNAGL, R. D., BELFRAGE, K., FARRINGTON, R., HUTCHINSON, K., LEWIS, V.,
ERLICH, J. & MOREY, F. (2002) Incidence of human astrovirus in central Australia
(1995 to 1998) and comparison of deduced serotypes detected from 1981 to 1998. J
Clin Microbiol, 40, 4114-20.
SEBIRE, N. J., MALONE, M., SHAH, N., ANDERSON, G., GASPAR, H. B. & CUBITT,
W. D. (2004) Pathology of astrovirus associated diarrhoea in a paediatric bone marrow
transplant recipient. J Clin Pathol, 57, 1001-3.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 56
SHAN, T., LI, L., SIMMONDS, P., WANG, C., MOESER, A. & DELWART, E. (2011
) The fecal virome of pigs on a high-density farm. J Virol, 85, 11697-708.
SILVA, A. M., LEITE, E. G., ASSIS, R. M., MAJEROWICZ, S. & LEITE, J. P. (2001) An
outbreak of gastroenteritis associated with astrovirus serotype 1 in a day care center, in
Rio de Janeiro, Brazil. Mem Inst Oswaldo Cruz, 96, 1069-73.
SILVA, P. A., CARDOSO, D. D. & SCHREIER, E. (2006) Molecular characterization of
human astroviruses isolated in Brazil, including the complete sequences of astrovirus
genotypes 4 and 5. Arch Virol, 151, 1405-17.
SIMPORE, J., OUERMI, D., ILBOUDO, D., KABRE, A., ZEBA, B., PIETRA, V.,
PIGNATELLI, S., NIKIEMA, J. B., KABRE, G. B., CALIGARIS, S.,
SCHUMACHER, F. & CASTELLI, F. (2009) Aetiology of acute gastro-enteritis in
children at Saint Camille Medical Centre, Ouagadougou, Burkina Faso. Pak J Biol Sci,
12, 258-63.
SIMPORE, J., ZONGO, F., KABORE, F., DANSOU, D., BERE, A., NIKIEMA, J. B.,
PIGNATELLI, S., BIONDI, D. M., RUBERTO, G. & MUSUMECI, S. (2005)
Nutrition rehabilitation of HIV-infected and HIV-negative undernourished children
utilizing spirulina. Ann Nutr Metab, 49, 373-80.
TANAKA, H., KISIELIUS, J. J., UEDA, M., GLASS, R. I. & JOAZEIRO, P. P. (1994)
Intrafamilial outbreak of astrovirus gastroenteritis in Sao Paulo, Brazil. J Diarrhoeal
Dis Res, 12, 219-21.
TAYEB, H. T., DELA CRUZ, D. M., AL-QAHTANI, A., AL-AHDAL, M. N. & CARTER,
M. J. (2008) Enteric viruses in pediatric diarrhea in Saudi Arabia. J Med Virol, 80,
1919-29.
TAYLOR, M. B., MARX, F. E. & GRABOW, W. O. (1997) Rotavirus, astrovirus and
adenovirus associated with an outbreak of gastroenteritis in a South African child care
centre. Epidemiol Infect, 119, 227-30.
TSE, H., CHAN, W. M., TSOI, H. W., FAN, R. Y., LAU, C. C., LAU, S. K., WOO, P. C. &
YUEN, K. Y. (2011) Rediscovery and genomic characterization of bovine
astroviruses. J Gen Virol, 92, 1888-98.
ULLOA, J. C. & GUTIERREZ, M. F. (2010) Genomic analysis of two ORF2 segments of
new porcine astrovirus isolates and their close relationship with human astroviruses.
Can J Microbiol, 56, 569-77.

Mémoire Master II /Biogéma/2012 / GAMSONRE Zakaria Page 57
WOLFAARDT, M., KIULIA, N. M., MWENDA, J. M. & TAYLOR, M. B. (2011) Evidence
of a recombinant wild-type human astrovirus strain from a Kenyan child with
gastroenteritis. J Clin Microbiol, 49, 728-31.
WUNDERLI, W., MEERBACH, A., GUNGOR, T., BERGER, C., GREINER, O., CADUFF,
R., TRKOLA, A., BOSSART, W., GERLACH, D., SCHIBLER, M., CORDEY, S.,
MCKEE, T. A., VAN BELLE, S., KAISER, L. & TAPPAREL, C. (2011) Astrovirus
infection in hospitalized infants with severe combined immunodeficiency after
allogeneic hematopoietic stem cell transplantation. PLoS One, 6, e27483.
YOON, P. W., BLACK, R. E., MOULTON, L. H. & BECKER, S. (1997) The effect of
malnutrition on the risk of diarrheal and respiratory mortality in children < 2 y of age
in Cebu, Philippines. Am J Clin Nutr, 65, 1070-7.





























