Embed Size (px)
Citation preview

.
UNIVERSITE DE OUAGADOUGOU ---------------
UNITE DE FORMATION ET DE RECHERCHE
SCIENCE DE LA VIE ET DE LA TERRE
(UFR/SVT)
Mémoire Présenté
Par : Moumouni Aissatou
Pour l’obtention du Master II
de Biologie Moléculaire et de Génétique Moléculaire Appliquées
de l’Université de Ouagadougou
SUR LE THEME:
CARACTERISATION DES GENES DE RESISTANCE AUX QUINOLONES
CHEZ LES ENTEROBACTERIES PRODUCTRICES DE
BETALACTAMASES A SPECTRE ELARGI ISOLEES CHEZ LES ENFANTS
DE 6 A 59 MOIS, DANS LA REGION DE MARADI AU NIGER
NNNNNNNNNNIGERNNNIGERNIGER..…………………………………
………………………………………………………..……………
…………………………………………………………………….
………………………………
Soutenu le 11 Novembre 2015 devant le jury composé de :
Président : Pr Aboubakar Sidiki OUATTARA, Professeur Titulaire, Université de
Ouagadougou
Membres : Pr Jacques SIMPORE, Professeur Titulaire, Université de Ouagadougou
Dr Serge DIAGBOUGA, Maitre de Recherche, IRSS
Dr Christelle M.W. NADEMBEGA, Maître Assistant, Université de
Ouagadougou
LABORATOIRE DE BIOLOGIE MOLECULAIRE
ET DE GÉNÉTIQUE MOLÉCULAIRE
(LABIOGENE)
N° d’Ordre .............................../LABIOGENE

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page i
Préface du Coordonnateur du Master BIOGEMA
De nos jours, les connaissances avancées en génétique et biologie moléculaires sont
incontournables pour conduire des études de hautes valeurs ajoutées en sciences
biologiques. Les outils de la Biologie moléculaire ont permis d’accomplir de grands
progrès dans le domaine du diagnostic, de la pharmacie, de la thérapeutique, de
l’agriculture et même dans l’aide à la justice par l’identification humaine. Les Universités
et les laboratoires de recherche des Pays membres de l’UEMOA dans leur grande
majorité, restent arrimés aux pays et laboratoires de recherche du Nord pour leurs
besoins en recherches et activités en génétique et biologie moléculaires. Cet état de fait
est lié au déficit en personnel qualifié et le manque de ressources financières et matériel
pour conduire les recherches en local in situ. Cela a pour conséquence, une non maîtrise
de la finalité ainsi que de l’utilisation des résultats et produits des recherches que nous
conduisons, une surenchère du coût des examens et des études en biologie et génétique
moléculaires. Un autre corollaire et non des moindres de cet état de fait est la fuite de
capitaux mais également la fuite des cerveaux car les étudiants les plus compétents
envoyés dans les pays du Nord ont tendance à y rester.
Le master en Biologie Moléculaire et en Génétique Moléculaire Appliquées (BioGeMA) a
pour but de combler le vide constaté dans l’expertise en génétique et biologie
moléculaires par la mise à disposition des pays de l’espace UEMOA, de personnels
qualifiés, de haut niveau de compétences pour conduire des études et recherches en
génétique et biologie moléculaires.
Le master BioGeMA est :
Un Master à dimension sous-régionale
Géré par un réseau de chercheurs et praticiens en génétique et biologie
moléculaires
Soutenu par une plateforme technologique sous-régionale à LABIOGENE
Ce Master a pour objectif de former des biologistes, des pharmaciens, des vétérinaires et
des médecins biologistes capables d’effectuer des diagnostics biomoléculaires dans des
centres hospitaliers et d’élaborer des études d’investigations dans des structures de
recherches. En outre, il ouvrira la porte d’études doctorales aux meilleurs étudiants pour
permettre la formation de chercheurs et d’enseignants-chercheurs afin d’assurer la relève
du corps enseignants, la constitution d’une masse critique d’experts africains et la mise en
place d’un véritable réseau africain de recherche dans le domaine ci-dessus cité.
Professeur Jacques SIMPORE
Professeur Titulaire de Biologie Moléculaire et
de Génétique Moléculaire
UFR/SVT - École Doctorale Sciences et
Technologie
Université de Ouagadougou – Burkina Faso

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page ii
DEDICACES
Je dédie ce travail:
A mon défunt Père Moumouni Harouna
A ma Mère Haoua Harouna, pour votre amour, votre confiance, votre soutien, et vos
encouragements.
A mes Frères, Sœurs, Parents et Amis pour votre soutien moral, matériels et pour vos
encouragements.
A tous ceux, qui, d’une manière ou d’une autre, ont contribué à l’aboutissement de ce
travail.

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page iii
REMERCIEMENTS
Ce travail a été réalisé au laboratoire d’Epicentre à Maradi au Niger, à l’Hôpital Saint
Camille de Ouagadougou au Burkina Faso (HOSCO) et au Centre de Recherche
Biomoléculaire Pietro ANNIGONI (CERBA).
Nous tenons à exprimer notre profonde gratitude :
o Au Professeur Jacques SIMPORE, Professeur titulaire, Responsable du Laboratoire
de Biologie Moléculaire et de Génétique (LABIOGENE); Coordonnateur du Master
de Biologie Moléculaire et de Génétique Moléculaire Appliquées (BioGeMA),
Université de Ouagadougou; Directeur du Centre de Recherche Biomoléculaire Pietro
Annigoni (CERBA); Recteur de l’Université Saint Thomas d’Aquin (USTA);
Membre de l’Académie Nationale des Sciences du Burkina (ANSB), notre Directeur
de Mémoire pour son encadrement et pour son soutien dans la réussite de notre
Master de recherche ;
o Au Professeur Aboubakar Sidiki OUATTARA, Professeur titulaire à l’université de
Ouagadougou, notre président du jury pour avoir accepté de juger notre travail ;
o Au Docteur Serge DIAGBOUGA, Maitre de Recherche à l’Institut de Recherche en
Sciences de la Santé (IRSS), notre co-directeur de mémoire pour avoir accepté de
juger notre travail et pour le temps qu’il a consacré à ce document ;
o Au Docteur Christelle Marie Wendyam NADEMBEGA, Maître Assistant, Université
de Ouagadougou pour son aide, son soutien et pour avoir accepté de juger notre
travail ;
o Au Docteur Florencia DJIGMA, assistante à l’Université de Ouagadougou pour son
encadrement et ses conseils si précieux tout au long de ce master ;
o Au Docteur Céline LANGENDORF, référente Bactériologie à Epicentre Paris,
France, pour sa collaboration, son aide, son soutien ;
o Au Docteur Metuor Dabire Amana, Docteur en Biologie Moléculaire option
Enzymologie , Laboratoire de Biologie et de Génétique moléculaire (LABIOGENE),
pour son soutien et son aide ;
o A Epicentre de m’avoir donné l’autorisation de travailler sur les souches résistantes
aux bétalactamases isolées, pendant l’étude ancillaire ;
o A tous les doctorants du CERBA/LABIOGENE pour leur participation à la réalisation
de ce travail ; particulièrement OUATTARA Abdoul Karim pour son aide ;

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page iv
o Au personnel du laboratoire d’Epicentre, d’HOSCO et du CERBA pour leur bonne
collaboration ;
o A toute la promotion du Master 2014, particulièrement Zongo Arsène pour son aide;
o A tous nos enseignants du Master BioGéMA pour leur encadrement de qualité;
o A l’Union Economique et Monétaire Ouest Africaine (UEMOA) pour le soutien
financier du master BioGéMA à travers le programme PACER2 ;
o A toute autre personne qui de près ou de loin a participé à la réalisation et à la qualité
de notre travail ;
o A mes parents, frères et sœurs pour leurs soutiens et encouragements

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page v
RESUME
Objectifs : L’objectif de cette étude était de décrire les gènes plasmidiques de résistance aux
quinolones chez E.coli et Klebsiella spp productrices de bétalactamase à spectre élargi
(BLSE) isolées chez les enfants de 6 à 59 mois inclus dans une étude menée par le centre de
recherche « Epicentre » dans la région de Maradi au Niger.
Méthodologie : 78 souches d’entérobactéries productrices de BLSE et résistantes aux
quinolones nous ont été fournies par le laboratoire de l’EPICENTRE Maradi-Niger. La
sélection des souches a été effectuée en fonction des couleurs des colonies sur milieu de
culture chromagar-BLSE: roses, et bleues suspectant respectivement E. Coli et Klebsiella.
L’identification a été confirmée par la galérie API 20E. La recherche des gènes qnr et aac(6’)
a été effectuée respectivement par PCR multiplex et simplex au CERBA/LABIOGENE,
Université de Ouagadougou, Burkina Faso
Résultats : 5,1 % des souches (03 E. coli et 01 Klebsiella pneumoniae) présentaient une
résistance au qnrA; le gène qnrB a été retrouvé chez 11 souches (02 E.coli, 02 Klebsiella
oxytoca et 07 Klebsiella pneumoniae) soit 14,1 % ; 27 souches (34,6 %) dont 23 E. coli et 04
Klebsiella pneumoniae ont montré une résistance au qnrS ; le gène aac(6’) était présent chez
53 souches (38 E. coli, 2 K. oxytoca et 13 K. pneumoniae) soit 67,9 %. Les souches BLSE
contenant le gène aac(6’) sont les plus nombreuses. La résistance associée aux quinolones
dans le cas de BLSE a été observée dans 88,46 % (69/78) soit 22 % de l’ensemble des
souches de Klebsiella. Aucune des souches résistantes aux bétalactamases n’héberge à la fois
les gènes qnrA, qnrB et qnrS mais il existe une association des gènes de résistance BLSE, qnr
et aac(6’) chez 26 souches répartis comme suit 12 E.coli, 02 Klebsiella oxytoca et 12
Klebsiella pneumoniae. Parmi les 21 souches (16 E. coli, 04 Klebsiella pneumoniae et 01
Klebsiella oxytoca) sensibles à l’acide nalidixique mais résistante aux fluoroquinolones (soit
26,9 %), on observe la présence du gène qnrA chez 01 souche, le gène qnrB est retrouvé chez
03 souches, 16 souches présentent une résistance au qnrS, 09 souches montrent une résistance
au gène aac(6’).
Conclusion : Nous avons obtenu des données sur les génotypes des déterminants qnr et sur
leur proportion chez les enfants dans la région de Maradi, au Niger. Nous avons aussi eu une
idée sur la répartition de ces gènes en fonction des espèces étudiées.

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page vi
Mots clés : Entérobactéries ; bétalactamases à spectre élargi, quinolones ; Klebsiella spp ; E.
coli
Abstract
Aim : the aim of this study is to describe the plasmid resistance to quinolone of wide
spectrum betalactamases producing E. coli and Klebsiella spp isolated from 6 to 59 months
children included in a study conducted by the research center ‘Epicentre ’at Maradi in Niger
Republic.
Methodology: 78 strains of ESBL producing E. coli and Klebsiella spp resistant to the
quinolone group were provided by the Epicentre laboratory of Maradi-Niger. The strain
selection was based on the colony color on the medium Chromagar ESBL: pink and blue
suspecting E. coli and Klebsiella respectively. The identification was confirmed the ‘’galerie
20E’’. The search for qnr and aac (6’) genes was carried out respectively by PCR multiplex
and simplex.
Results: 5.1 % of strains (3 E. coli and 1 Klebsiella pneumonia) were resistant to qnrA; the
qnrB gene was found in 11 strains (2 E.coli, 2 Klebsiella oxytoca et 7 Klebsiella pneumoniae)
making 14.1 % ; 27 strains (34,6 %) among which 23 E. coli and 4 Klebsiella showed a
resistance to qnrS; the and aac (6’) gene was present in 53 strains (67.9 %). The ESBL strains
containing the aac (6’) gene were the most important. The resistance to quinolone associated
with ESBL was observed in 88.46 % (69/78) making 22 % the whole Klebsiella strains.
None of the ESBL strains carried simultaneously qnrA, qnrB and qnrS genes but there may
be and association of ESBL, qnr and aac(6’) genes in the 26 strains as follow: 12 E. coli, 2
Klebsiella oxytoca and 12 Klebsiella pneumoniae.
Among the 21 strains (16 E. coli, 4 K. pneumoniae and 1K. oxytoca) sensible to nalidixic acid
but resistant to fluoroquinolones making 26.9 %, we noticed the presence of qnrA gene in
one (1) strain; the qnrB gene was found in 3 strains; the qnrS gene was found in 16 strains
and finally 9 strains showed a resistance to aac(6’) gene.
Conclusion: we have obtained data concerning the genotype of QNR determinant and their
proportion in Maradi region in Niger republic and we’ve gotten an idea on the repartition of
these genes based on the species studied.
Key words: Enterobacteria; wide spectrum betalactamase; quinolone, Klebsiella spp;

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page vii
TABLE DES MATIERES
Préface du Coordonnateur du Master BIOGEMA ..................................................................................................... i
DEDICACES ............................................................................................................................................................. ii
REMERCIEMENTS ..................................................................................................................................................iii
RESUME ................................................................................................................................................................... v
ABSTRACT ............................................................................................................................................................... vi
TABLE DES MATIERES ........................................................................................................................................ vii
LISTE DES FIGURES .............................................................................................................................................. x
LISTE DES TABLEAUX ...........................................................................................................................................x
LISTE DES ABREVIATIONS ................................................................................................................................. xii
INTRODUCTION ...................................................................................................................................................... i
I. Revue Bibliographie .......................................................................................................................................... 3
I.1 Généralités sur les bactéries.............................................................................................................................. 3
I.1.1 Définition ................................................................................................................................................... 3
I.1.2 Morphologie .............................................................................................................................................. 3
I.1.3 Matériel génétique ..................................................................................................................................... 4
I.1.4 Paroi cellulaire ........................................................................................................................................... 5
I-2. Les entérobactéries .......................................................................................................................................... 5
I-2-1 Définition .................................................................................................................................................. 5
I-2-2 Historique .................................................................................................................................................. 6
I-2-3 Epidémiologie ........................................................................................................................................... 7
I.2.4 Les groupes d'entérobactéries .................................................................................................................... 7
I.2.5 Les caractères généraux et biochimiques................................................................................................... 8
I.2.6 Identification .............................................................................................................................................. 9
I.3 Antibiotiques .................................................................................................................................................. 10
I.3.1 Généralités ............................................................................................................................................... 10
I.3.1.1 Définition .............................................................................................................................................. 10

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page viii
I.3.1.2 Historique ............................................................................................................................................. 10
I.3.1.3 Bêta-lactamines..................................................................................................................................... 12
I.3.1.4 les quinolones ...................................................................................................................................... 13
I.4 Résistance bactérienne aux antibiotiques ....................................................................................................... 15
I.4.1 Résistance aux β-lactamines .................................................................................................................... 16
I.4.2 Résistance aux quinolones ....................................................................................................................... 17
II. Objectifs de l’étude ............................................................................................................................................. 19
II.1 Objectif principal ........................................................................................................................................... 19
II-2 Objectifs spécifiques .................................................................................................................................... 19
III Matériel et méthodes .......................................................................................................................................... 20
III.1 Cadre de l’étude ........................................................................................................................................... 20
III.2 Type et période d’étude ............................................................................................................................... 21
III.3 Echantillonnage ............................................................................................................................................ 21
III.3.1 - Mode de sélection ............................................................................................................................... 22
III.3.2 - Critère d’inclusion .............................................................................................................................. 22
III.3.3 -Critères d’exclusion ............................................................................................................................. 22
III.4 Matériel ........................................................................................................................................................ 23
III.4.1 Réactifs ................................................................................................................................................. 23
III.4.2 Appareillage .......................................................................................................................................... 23
III.4.3 Autres matériels .................................................................................................................................... 23
III.5 Méthodes ...................................................................................................................................................... 23
III.5.1 Confirmation de l’identité des souches ................................................................................................. 23
III.5.2. Sensibilité aux antibiotiques ................................................................................................................ 23
III.5.3 Condition de cultures des souches ........................................................................................................ 24
III.5.4 Extraction des ADNs ............................................................................................................................ 24
III.5.5 Amplification des gènes de quinolones................................................................................................. 24
III.5.6 Electrophorèse sur gel d’agarose .......................................................................................................... 26

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page ix
III.5.7 Analyse des données ............................................................................................................................. 27
III.5.8 Considérations éthiques ........................................................................................................................ 27
IV Résultats .............................................................................................................................................................. 28
IV.1 Caractéristiques et répartition des souches d’entérobactéries en fonction des espèces
bactériennes.......................................................................................................................................................... 28
IV.2 Répartition des gènes en fonction de l’espèce ............................................................................................. 28
IV.3 Répartition de l’association des gènes en fonction des espèces bactériennes .............................................. 29
IV.4 Répartition de gènes chez les souches sensibles à l’acide nalidixique et résistantes aux
fluoroquinolones .................................................................................................................................................. 29
IV.5 Caractérisation des gènes codant pour la résistance aux quinolones .......................................................... 30
V. Discussion ........................................................................................................................................................... 37
Conclusion ............................................................................................................................................................... 39
PERSPECTIVES ..................................................................................................................................................... 39
REFERENCES BIBLIOGRAPHIQUES ................................................................................................................. 40

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page x
LISTE DES FIGURES
Figure 1: Les trois types de bactéries ......................................................................................................................... 4
Figure 2: Structure d’une bactérie .............................................................................................................................. 5
Figure 3: Structure des quinolones, A : noyau commun, B : acide nalidixique, C : fluoroquinolone. .................... 14
Figure 4: Inactivation enzymatique de l’antibiotique .............................................................................................. 16
Figure 5: Mécanisme d’action des quinolones (Dentan, 2015) ................................................................................ 18
Figure 6: Gel d’agarose des produits PCR des gènes qnrA et qnrB qnrS ................................................................ 36
Figure 7: Gel d’agarose des produits PCR du gène aac(6’)-Ib. ............................................................................... 36
LISTE DES TABLEAUX
Tableau I : Protocoles de PCR du gène qnr ............................................................................................................. 25
Tableau II: Séquences des amorces utilisées au niveau de cette étude .................................................................... 26
Tableau III: Répartition des souches d’entérobactéries en fonction de l’espèce ..................................................... 28
Tableau IV: Répartition des gènes en fonction de l’espèce bactérienne .................................................................. 28
Tableau V: Répartition de l’association des gènes en fonction des espèces bactériennes ....................................... 29
Tableau VI : Répartition de gènes chez les souches sensibles à l’acide nalidixique et résistantes aux
fluoroquinolones ...................................................................................................................................................... 29
Tableau VII: Données des antibiogrammes et résultats de la PCR .......................................................................... 31

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page xi
LISTE DES ABREVIATIONS
BLSE : Bétalactamase à Spectre Elargi
Qnr : Quinolone resistant
PCR : Polymerase Chain Reaction
CERBA : Centre de Recherche Biomoléculaire Pietro Annigoni
LABIOGENE : Laboratoire de Biologie Moléculaire et de Génétique
E : Escherichia
K : Klebsiella
Aac : Aminoglycoside acétyl-transférase
Cr : Ciprofloxacine résistant
Β : Béta
CTX : Cefotaxime
OXA : Oxacilline
ADN : Acide désoxyribonucléique
µl : Microlitre
mm : Millimètre
% : Pourcentage
C : Degré Celsius
GC : Guanine – cytosine
Cint : Concentration interne
Cext : Concentration externe
Zn : Zinc
HOSCO : Hôpital Saint Camille de Ouagadougou
MSF : Médecins Sans Frontières
OMS : Organisation Mondial de la Santé

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page xii
C3G : Céphalosporine troisième génération
MH : Muller-Hinton
LB : Luria-Bertani
TCS : Trypticase Soja
CA-SFM : Comité de l’antibiogramme de la Société française de microbiologie
CIP : Collecte de l’Institut Pasteur
H : Heure
UV : Ultra-violet
Pb : Paire de base
CCNE : Comité Consultatif National d’Ethique
PM : Poids Moléculaire
CMI : Concentration Minimale Inhibitrice

INTRODUCTION

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 1
INTRODUCTION
La résistance aux antibiotiques est un problème majeur de santé publique, en particulier dans
les pays où l'utilisation des antibiotiques n’est souvent pas contrôlée (Pons et al, 2014).
Les antibiotiques sont des substances chimiques élaborées par des micro-organismes capables
d'inhiber la multiplication (action bactériostatique) ou de tuer (action bactéricide) les
bactéries. Il existe de nombreux antibiotiques, qui peuvent être classés en familles selon leurs
modes d’action ou leur structure moléculaire. Parmi ceux-ci, les bêta-lactamines (les
pénames, les pénèmes, les céphèmes, les monobactames et les oxapènames) inhibent la
synthèse de la paroi bactérienne et les quinolones (quinolones et fluoroquinolones.)
empêchent la réplication de l'ADN bactérien en bloquant l'ADN gyrase ou topoisomérase II.
Au cours de la dernière décennie, il a été observé une augmentation importante de la
résistance des entérobactéries aux antibactériens et surtout l’émergence des bactéries
multirésistantes par hyperproduction de céphalosporinase ou production de bétalactamase à
spectre élargie (BLSE) ou encore de carbapénèmases (Bradford, 2001b, Rodriguez-villalobos
and Struelens, 2006).
Découvertes au début des années 1980 (Ambler, 1980), les BLSE constituent une grande
famille très hétérogène d’enzymes bactériennes appartenant à la classe A ou D de la
classification d’Ambler (Ambler, 1980, Hall and Barlow, 2005). Elles sont soit induites par
des plasmides soit dérivées de mutations ponctuelles dans la séquence génétique codant pour
le site actif des β-lactamases (Vora and Auckenthaler, 2009). Les gènes de types TEM, SHV,
CTX-M, OXA ont été décrits dans plusieurs études en Europe (Giraud-Morin and Fosse,
2008) et en Afrique (Pitout et al, 1998 ; Woerther et al, 2011).
Le développement des quinolones à partir de l’acide nalidixique a débuté en 1962 (Takahashi
et al, 2003). Le premier gène plasmidique de résistance aux quinolones (qnr) a été découvert
en 1994 isolé d’une Klebsiella pneumoniae à Birmingham, Alabama et rapportée en 1998
(Martınez-Martınez et al, 1998). Depuis la découverte du gène de résistance plasmidique
(qnr) aux quinolones, un grand nombre d'allèles qnr ont été découvertes sur des plasmides ou
le chromosome bactérien (Nordmann and Poirel, 2005 ; Robicsek et al, 2006). Les gènes qnr
d’origine plasmidique comprennent actuellement cinq familles, qnrA, qnrB, qnrC, qnrD et
qnrS, différentes l'une de l'autre (Jacoby et al, 2008). En effet les mécanismes de résistance
bactérienne contre les quinolones proviennent généralement de mutations de l'ADN gyrase
(topoisomérase II) ou la topoisomérase IV, ou à partir de l'expression d'efflux de pompes
agissant seul ou en combinaison avec des taux réduits d'expression de porines de la

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 2
membrane externe. (Piddock et al, 1999 ; Oethinger et al, 2000 ; Wang et al, 2001 ; Oktem et
al, 2008). Récemment, un nouveau mécanisme de résistance transférable aux quinolones a été
signalé : il s’agit de la variante aac(6') qui code pour une aminoglycoside acétyltransférase
conférant une sensibilité réduite à la ciprofloxacine et norfloxacine par N-acétylation de
l’amine pipérazinyle (Robicsek et al, 2006).
Des études antérieures ont montré que les souches de qnr-positif expriment fréquemment des
BLSE, tels que CTX-M-15 et SHV-12(Jacoby et al, 2006 ; Robicsek et al, 2006).
Les quinolones et les β-lactamines sont largement utilisés en tant que antibiotiques à large
spectre pour traiter diverses infections bactériennes (Oktem et al, 2008). Cette utilisation
intensive des antibiotiques en médecine humaine et vétérinaire, mais aussi comme
supplément alimentaire (Martel and Chaslus-Dancla, 2001) a sélectionné les bactéries
possédant des mécanismes de résistance naturelles ou ayant la capacité de les acquérir
(Schwarz and Chaslus-Dancla, 2001).
Bien que de nombreux isolats d’entérobactéries positif en qnrA, qnrB ou qnrS aient été
signalés en Europe (Oktem et al, 2008), au Niger les différents types de gènes de résistance
aux quinolones circulants sont inconnus. Ainsi la problématique se rapporte à la
caractérisation moléculaire des gènes plasmidiques codant pour la résistance aux quinolones
au sein d’enterobactéries productrices de bétalactamase à spectre élargi afin d’identifier les
déterminants qnr et les gènes aac (6´) qui y circulent.
Le choix s’est porté sur les gènes plasmidiques de résistance car ils ont beaucoup plus
d’intérêt scientifique du point de vue épidémiologique du fait de l’importance de la
transmission inter-espèce. Généralement les résistances liées aux plasmides sont toujours
associées c'est-à-dire le plasmide porte le plus souvent plusieurs gènes de résistance à la fois.
Cette étude va situer du point de vue moléculaire, les gènes plasmidiques de résistance aux
quinolones qui circulent au Niger et qui sont plus répandus du fait de leur transfert facile
entre bactéries. Elle sera une contribution à la mise en place d’une meilleure surveillance du
point de vue épidémiologique et la mise en évidence des nouvelles résistances.

REVUE BIBLIOGRAPHIQUE

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 3
I. Revue Bibliographie
I.1 Généralités sur les bactéries
I.1.1.Définition
La bactérie est une cellule procaryote (dépourvue d’un véritable noyau) : absence de
membrane nucléaire, chromosome unique en général.
La bactérie possède une paroi rigide faite d’un constituant spécifique : peptidoglycane ou
mureine (les bactéries, 2014).
Leur classification est fondée sur les caractères :
- morphologiques,
- tinctoriaux,
- biochimiques,
- antigéniques,
- génétiques.
I.1.2 Morphologie
Une bactérie, est un micro-organisme (organisme de très petite taille) unicellulaire (formé
d’une seule cellule). En fait, une bactérie est une entité complète, entourée d’une paroi
cellulaire complexe, et possédant à l’intérieur de cette paroi, dans son cytoplasme, toute la
machinerie nécessaire à son autonomie structurale et fonctionnelle. Les bactéries sont des
organismes procaryotes, c’est-à-dire qu’ils ne contiennent pas de noyau. Le matériel
génétique n’est pas séparé du reste du cytoplasme. Ce dernier contient également des
ribosomes nécessaires à la fabrication des protéines suite à l’interprétation du message
contenu dans le code génétique, ainsi que divers organites responsables du maintien des
fonctions métaboliques de base indispensables à la survie cellulaire. La taille des bactéries
peut varier mais elle est généralement de l’ordre du micromètre. Les bactéries peuvent être de
différentes formes : bacilles ou bâtonnets (de forme allongée), coques (de forme arrondie) et
spiralée. La figure (1) montre les trois types de bactéries : des bacilles (orange), des coques
(vert) et des spirilles (jaune) (Acar et al, 1998)
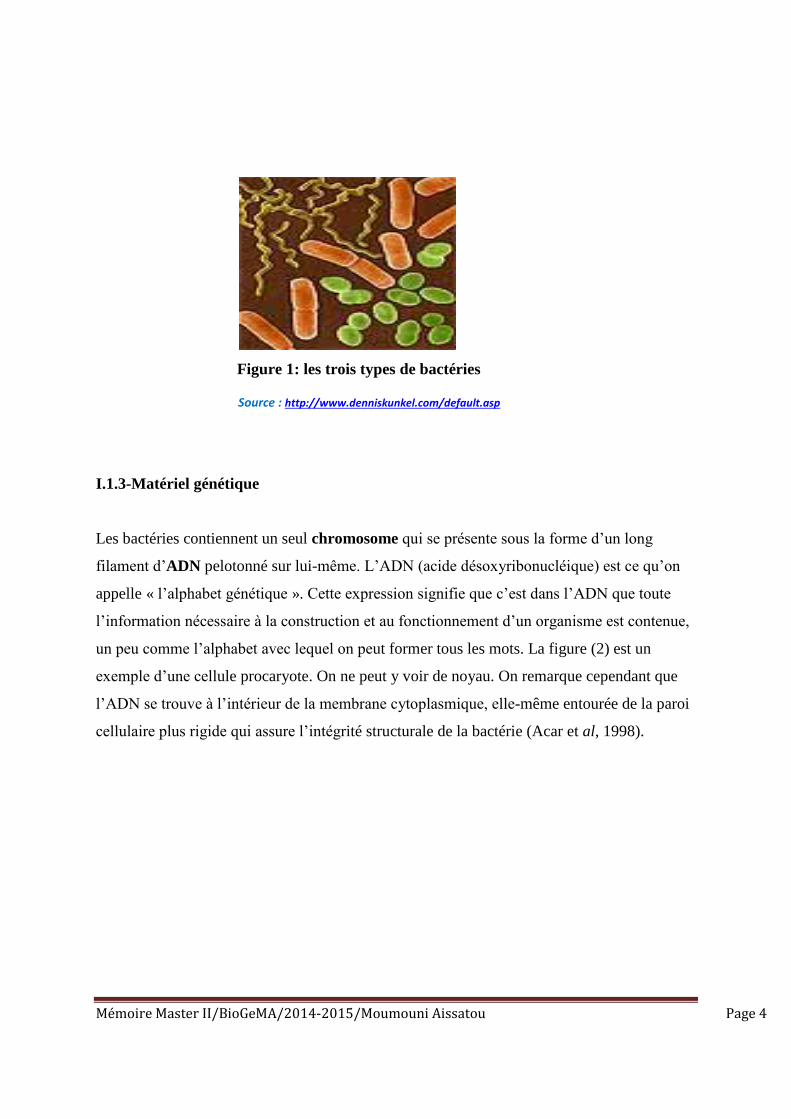
Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 4
Figure 1: les trois types de bactéries
Source : http://www.denniskunkel.com/default.asp
I.1.3-Matériel génétique
Les bactéries contiennent un seul chromosome qui se présente sous la forme d’un long
filament d’ADN pelotonné sur lui-même. L’ADN (acide désoxyribonucléique) est ce qu’on
appelle « l’alphabet génétique ». Cette expression signifie que c’est dans l’ADN que toute
l’information nécessaire à la construction et au fonctionnement d’un organisme est contenue,
un peu comme l’alphabet avec lequel on peut former tous les mots. La figure (2) est un
exemple d’une cellule procaryote. On ne peut y voir de noyau. On remarque cependant que
l’ADN se trouve à l’intérieur de la membrane cytoplasmique, elle-même entourée de la paroi
cellulaire plus rigide qui assure l’intégrité structurale de la bactérie (Acar et al, 1998).

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 5
Figure 2: structure d’une bactérie
Source : www.nirgal.net/ori_intro.html
I.1.4- Paroi cellulaire
On peut distinguer deux grandes classes de bactéries, caractérisées par une architecture
distincte de la paroi cellulaire qui les entoure. Ainsi, chez les bactéries Gram(+), la paroi
cellulaire est constituée principalement de peptidoglycane. C’est la couche principale.
L’épaisseur de cette couche est beaucoup plus importante que pour les bactéries à Gram
négatif, chez les bactéries à Gram(-), la paroi cellulaire est constituée de trois couches. Les
deux premières couches, les plus externes, sont composées de phospholipides et de protéines.
Elles forment la membrane externe. Sous cette membrane se trouve la troisième couche de la
paroi, constituée d’un mince feuillet de peptidoglycane, qui ressemble beaucoup au
peptidoglycane de la bactérie à Gram positif, sauf que la nature de son lien peptidique n’est
pas le même. La paroi peut être identifiée à l’aide de la coloration de Gram (Acar et al,
1998).
I-2. Les entérobactéries
I-2-1 Définition
Les entérobactéries sont des bacilles à Gram négatif, le plus souvent courts (1 à 6 µl), droits,
immobiles ou mobiles par une ciliature péritriche (rares, exceptions) de culture aisée, aéro-
anaérobies facultatifs, fermentaires, oxydase négative, catalase positive (rares, exceptions),
nitrate réductase positive (rares, exceptions) (Bidet et Bingen, 2007).

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 6
Les entérobactéries sont retrouvées partout dans le sol, dans l'eau, et surtout dans l'intestin de
l'homme et des animaux. Elles comprennent un nombre très élevé de genres et d'espèces.
Leur abondance dans l'intestin, leur mobilité, la rapidité de leur multiplication, l'acquisition
fréquente de mécanismes de résistance aux antibiotiques expliquent qu'elles soient les
bactéries les plus souvent impliquées en pathologie infectieuse humaine, surtout en milieu
hospitalier.
Outre une résistance naturelle aux antibiotiques actifs sur les germes à Gram positif
(pénicillines, macrolides), les entérobactéries présentent fréquemment une résistance acquise
aux antibiotiques à large spectre. Cette résistance est souvent conditionnée par la présence de
plasmides porteurs de déterminants de résistances multiples et transférables à d'autres
bactéries à Gram négatif (Delmée, 2004).
I-2-2 Historique
La période de naissance de la famille des Enterobacteriaceae se situe entre 1937 lorsque Otto
RAHN proposa le genre Enterobacter pour regrouper les microorganismes présentant des
propriétés biochimiques et morphologiques communes et parmi lesquelles on trouvait déjà
des noms tels que Escherichia, Salmonella, Klebsiella, Proteus, Serratia ou Shigella.
Deux années après cette description qui concernait 112 espèces, ce nombre fut ramené à 67.
Avec les travaux de Don Brenner et de Patrick Grimont (Jean Freney et al, 2007), cette
famille a connu un essor et beaucoup de nouveaux genres et espèces furent découverts.
En 1972, Edwards et Ewing rapportaient 11 genres et 26 espèces dans la famille des
Enterobacteriaceae.
En 1985, Farmer et al décrivaient 22 genres comprenant 69 espèces et 29 groupes entériques
(Farmer et al, 1985).
Aujourd’hui, plus de 40 genres différents et près de 200 espèces ont été décrits depuis la
Serratia découverte par Bartelomeo Bizio en 1983 jusqu’aux derniers genres Samsonia en
2001 et Dickeya en 2004. (Jean Freney et al, 2007).

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 7
I-2-3 Epidémiologie
Les entérobactéries représentent la première cause d’infection des voies urinaires. Elles font
partie des bactéries les plus souvent isolées dans les hémocultures : Environ 35 % des
bactériémies d’origine nosocomiale et plus de 50 % des bactériémies d’origine
communautaire, recensées en France en 2002 étaient dues à des entérobactéries.
Dix à 35 % des entérobactéries nosocomiales sont capables de développer des résistances à
plusieurs familles d’antibiotiques par production de béta-lactamases à spectre large.
Cependant, depuis quelques années, ces entérobactéries multirésistantes ont également pu
être isolées en dehors de l’hôpital : en 2006, elles représentaient moins de 5 % des
entérobactéries communautaires (E. Pilly, 21ème édition, 2008).
Partout dans le monde, on constate des fréquences élevées de résistances aux quinolones.
Chez les entérobactéries d’origine communautaires et nosocomiales, le taux est supérieur à
50 %, particulièrement en Asie. Cependant, cette résistance est plus fréquente chez les
entérobactéries résistantes aux C3G (Dalhoff, 2012).
En Afrique, le rapport de l’OMS fait état de lacunes majeures dans le suivi de la résistance
aux antibiotiques, des données n’étant fournies que dans un nombre limité de pays. Bien qu’il
ne soit pas possible d’évaluer la véritable ampleur du problème, compte tenu du manque de
données, celles dont on dispose sont inquiétantes (Premier rapport de l’OMS sur la résistance
aux antibiotiques à Genève : une menace grave d’ampleur mondiale, 2014).
Au Niger, nous n’avons trouvé aucune donnée par rapport aux enterobactéries productrices
de béta-lactamases à spectre élargi résistantes aux quinolones.
I.2.4 Les groupes d'entérobactéries
On peut schématiquement subdiviser l'ensemble des entérobactéries en deux groupes :
- d’une part les entérobactéries qui font partie des flores fécales commensales habituelles de
l'homme et des animaux; ce groupe comprend principalement Escherichia coli, Klebsiella,
Enterobacter, Proteus, Morganella, Providencia, Serratia, Citrobacter...

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 8
- d'autre part les espèces pathogènes pour l'intestin, dont l'ingestion provoque une infection
intestinale (Salmonella enteritidis, Yersinia, Shigella et certaines souches d'E.coli dites
"pathogènes") ou un syndrome septicémique (Salmonella typhi) (Delmée, 2004).
I.2.5 Les caractères généraux et biochimiques
La plupart des bactéries appartenant à la famille des Enterobacteriaceae partagent les
caractères généraux suivants :
Ce sont des bacilles à Gram négatif, ne formant pas de spores ;
Lorsqu’elles sont mobiles (Klebsiella, Shigella et Yersinia pestis sont immobiles), ces
bactéries présentent une ciliature péritriche ;
Elles se développent aussi bien en aérobiose qu’en anaérobiose sur des milieux «
ordinaires » (Janda et Abbott, 2006). La température optimale de croissance est
généralement de 35 à 37 °C à l’exception des Yersinia (30 à 37 °C), des Pantoea et
des Erwinia (27 à 30 °C);
L’aspect général des colonies de ces bactéries sur gélose nutritive est florissant,
colonie de 1 à 3 mm de diamètre généralement bombées, lisses et brillantes (excepté
pour Shigella, Yersinia, certaines espèces de Salmonella, Proteus);
Le plus souvent, ces colonies sont opaques et blanchâtres, mais il en est de plus
transparentes telles les Salmonella et celles présentant des pigments rouge et jaune
respectivement chez les Serratia et les Erwinia.
Les Klebsiella forment des colonies souvent très muqueuses, larges et luisantes (Denis
et al, 2007).
La famille des Enterobacteriaceae partage les caractères biochimiques suivants :
Elles utilisent le D-glucose et les sucres par fermentation plutôt que par oxydation
souvent avec production de gaz ;
Les entérobactéries sont catalase-positives (à l’exception de Shigella dysenteriae
sérotype 1) et oxydase-négatives ;
Elles réduisent les nitrates en nitrites (à l’exception de certaines souches d’Erwinia et
de très rares mutants)

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 9
Elles contiennent un antigène commun et possèdent un GC % compris entre 39 et 59
% (Janda et Abbott, 2006)
I.2.6 Identification
L’identification par des techniques issues de la biologie moléculaire n’est pas encore à la
portée de tous les laboratoires. Cependant, la recherche des caractères généraux et
biochimiques de la famille demeurent les moyens d’identification couramment mis en œuvre
(Denis et al, 2007).
C’est dans le domaine des Enterobacteriaceae que l’évolution technologique a été la plus
importante en bactériologie médicale. L’ère des galeries d’identification en tubes est quasi
révolue en pratique quotidienne pour faire place à celles des systèmes prêts à l’emploi.
Quelques remarques doivent néanmoins être apportées quant à l’utilisation de ces systèmes
prêts à l’emploi :
- Le mode opératoire doit être rigoureusement suivi ;
- Les caractères déterminés n’ont de signification que confrontés aux tableaux fournis
par le fabricant.
Bien souvent d’ailleurs, le principe d’identification proposé est établi à partir d’une méthode
probabiliste qui consiste à traduire numériquement la séquence des caractères positifs trouvés
et à confronter le profil trouvé au catalogue des profils recensés par le fabricant. Globalement
plus de 90 % des souches sont correctement identifiées d’emblée par ces systèmes prêts à
l’emploi.
Les difficultés surgissent quand le profil numérique ne figure pas dans le catalogue ou
conduit à une discrimination insuffisante entre plusieurs espèces ou genres. (Denis et al,
2007).

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 10
I.3 Antibiotiques
I.3.1 Généralités
I.3.1.1 Définition
Un antibiotique (du grec anti : contre, et bios : la vie) est une substance d'origine naturelle ou
synthétique, ayant la capacité d'arrêter la multiplication des bactéries, mais également d'autres
agents infectieux. Certains sont également capables de détruire les microbes (regroupe tous
les organismes microscopiques ne possédant qu'une seule cellule : virus, bactéries,
champignons).
Les antibiotiques, contrairement aux antiseptiques, agissent sur les bactéries en inhibant des
fonctions physiologiques précises telles que la synthèse de la paroi (β-lactamines,
glycopeptides, fosfomycine), la réplication/transcription de l’ADN (fluoroquinolones,
rifampicine, sulfamides, triméthoprime), la synthèse protéique (aminosides, tétracyclines,
macrolides et apparentés, chloramphénicol) ou encore la respiration cellulaire (polymixines).
Pour exercer leur action, ils doivent se lier à des cibles moléculaires spécifiques le plus
souvent intracellulaires.
Des facteurs complexes déterminent l’activité intrinsèque d’un antibiotique sur une espèce
donnée, notamment l’affinité de l’inhibiteur pour sa cible, le nombre de molécules cibles à
inactiver et, enfin la concentration de l’inhibiteur au voisinage de la cible (Cint). Ce dernier
paramètre dépend évidemment de la concentration du produit dans le milieu où se trouve la
bactérie (Cext), mais également d’autres éléments comme le niveau de perméabilité des
membranes à l’agent considéré (diffusion simple ou transport actif), l’existence éventuelle de
mécanismes d’inactivation ou d’activation, ou encore la présence de systèmes d’efflux actif
naturels pouvant s’opposer à son accumulation intracellulaire. (Plesiat, 2012).
I.3.1.2 Historique
Toutes les découvertes médicales effectuées au cours du XXe siècle sont importantes.
Néanmoins l'antibiotique, dans la mesure où il a fait reculer le taux de mortalité en permettant
de guérir certaines maladies infectieuses, est sans doute le médicament dont la découverte a

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 11
le plus bouleversé la médecine et la démographie. Bien entendu, ces maladies n'ont pas
disparu et sont encore la principale cause de mortalité. Généralement, quand on pense aux
antibiotiques et à leur histoire, le premier nom qui vient à l'esprit est celui du britannique
Alexander Fleming. Pourtant, à partir de 1874, Roberts, puis Tyndall (en 1876), Pasteur et de
Joubert (en 1877), enfin Duchesne (en 1897-1898), amorcent la découverte de Sir Alexander
Fleming par leurs réflexions sur la question des produits susceptibles d'entraver la
multiplication des germes comme les moisissures (qui sont des champignons microscopiques
se développant à l'humidité sur des milieux organiques).
En fait, l'histoire des antibiotiques commence en 1929, date à laquelle on constate qu'une
moisissure se développant naturellement sur des fruits ou des fromages empêche la
prolifération du bacille de la diphtérie et de celui du charbon dans les boîtes où l'on cultive
ces microbes, en laboratoire. On baptise le liquide de culture de cette moisissure Penicillium
notatum, et on constate qu'elle n'est pas toxique pour la souris à laquelle on l'injecte. Malgré
tout, cette découverte n'attire pas vraiment l'attention des chercheurs et reste sans lendemain.
(http://www.vulgaris-medical.com/encyclopedie-medicale/antibiotique-generalites, 2015).
En 1935, l'Allemand Domagk reprend les idées d'Ehrlich sur l'effet anti-infectieux de certains
colorants qu'il avait mis au point en 1905, en se servant d'un colorant pour traiter certaines
infections par une bactérie appelée streptocoque. Le Français J. Tréfouël et sa femme, à
l'Institut Pasteur, montrent que le produit actif appartient à une famille appelée sulfamide. A
partir de là, et pendant une quinzaine d'années, cette variété de médicaments sera employée
contre les germes, reléguant du même coup à une seconde place les moisissures et autres
produits susceptibles de posséder des capacités antibiotiques. Ce n'est qu'en 1939 que le
français R. Dubos découvre qu'une bactérie appelée Bacillus brevis est capable de produire
une substance empêchant la multiplication de certaines bactéries. Grâce à un système de
coloration qualifiée de «gram +» (d'après une méthode de coloration due au biologiste danois
Hans Gram), il met en évidence cette bactérie. Cependant, le premier antibiotique identifié fut
la pénicilline découverte par hasard (Fleming, 1929). Ce premier antibiotique a ouvert une
voie nouvelle dans la lutte contre de nombreuses maladies qui étaient considérées comme
incurables auparavant. Suite à la découverte de la pénicilline, de nombreuses autres
générations d’antibiotiques (nature synthétique) ont vu le jour et continuent d’apparaître dans
le but de combattre. (Plesiat, 2012) Leur nombre deviendra tel qu'il faudra établir

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 12
progressivement des règles de prescription que l'on appelle antibiothérapie
(http://www.vulgaris-medical.com/encyclopedie-medicale/antibiotique-generalites, 2015).
I.3.1.3 Bêta-lactamines
Les β-lactamines sont des antibiotiques qui possèdent un noyau β-lactame qui est la partie
efficace de la molécule. Des variations au niveau de la chaîne latérale permettent de modifier
les propriétés de la molécule antibiotique. Par exemple, une modification des chaînes
latérales entraîne une résistance aux acides (suc gastrique) et permet l'administration de
l'antibiotique par voie orale. Les β-lactamines ou antibiotiques à noyau β-lactame sont une
large classe d'antibiotiques qui comprennent les pénames, les pénèmes, les céphèmes, les
monobactames et les oxapènames, bref, toute substance qui contient un noyau β-lactame dans
sa structure moléculaire (Glupczynski et al, 2002).
Du fait de leur diversité, faible toxicité, activité bactéricide et large spectre d’action, les β-
lactamines sont parmi les antibiotiques les plus utilisés dans le traitement des infections
causées par les Enterobacteriaceae. L’efficacité des β-lactamines dépend d’au moins trois
facteurs : la quantité d’antibiotique au contact de la cible, l’affinité de l’antibiotique pour la
cible et la production de β-lactamase inactivant l’antibiotique.
Ces facteurs sont responsables, soit d’une résistance naturelle (intrinsèque), et donc présents
chez toutes les souches de l’espèce, soit d’une résistance acquise par certaines souches, suite
à l’apparition de mutations ou à l’acquisition de matériel génétique tels que des plasmides,
des transposons ou des intégrons. Pour chaque espèce, on distingue donc un «phénotype
sauvage» de résistance aux β-lactamines ou un «phénotype sensible» et des phénotypes de
résistance acquise ou «phénotypes résistants». La résistance peut s’exprimer à bas niveau in
vitro pour certaines molécules mais être responsables d’échecs thérapeutiques. La
connaissance des phénotypes de résistance permet :
d’établir la liste des β-lactamines à étudier en routine,
de contrôler la cohérence entre l’identification et le phénotype de résistance
de pallier les imperfections des tests de sensibilité in vitro par l’application de règles
de lecture interprétative,
de guider le choix thérapeutique (Bonnet, 2012).

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 13
I.3.1.4 les quinolones
Les quinolones sont des composés antibactériens de synthèse qui dérivent d'acides
carboxyliques hétérocycliques diversement substitués. Toutes les quinolones actuelles
présentent une structure bicyclique, avec un azote en position 1, un carboxylate en position 3
et un carbonyle en position 4. Les quinolones dont le chef de file, l’acide nalidixique, a été
décrit en 1962 par Lesher et al. (Hiramatsu et al, 1997). Cependant, l’acide nalidixique fut la
première quinolone à être introduite pour un usage clinique au début des années soixante
(Ball, 2000).
Les quinolones (Figure 3-B) sont réparties en deux groupes : les quinolones (spectre utile
limité aux entérobactéries) et les fluoroquinolones (Figure 3-C), quinolones comportant un
atome de fluor en position 6 (antibiotiques à spectre large dont l’activité intrinsèque est très
supérieure à celle des premières quinolones) (Hiramatsu et al, 1997). L'apparition sur le
marché dans les années 1980 de la norfloxacine, ofloxacine, ciprofloxacine, péfloxacine et
loméfloxacine a permis aux fluoroquinolones de devenir des antibiotiques de référence pour
de nombreuses infections.
La présence d'une fonction acide carboxylique en position 3, et un cycle pyridone dont la
fonction aminée en position 1 est substituée par une chaine aliphatique ou par un cycle sont
indispensables à l'activité antibiotique, tandis que l'addition d'un fluor en 6 et d'un cycle
diamine en 7 accroit très significativement l'activité par rapport aux dérivés originaux (acide
nalidixique).
Lorsqu’elles ont diffusé dans le cytoplasme, les quinolones exercent une inhibition sélective
de la synthèse de l’ADN bactérien en agissant sur deux enzymes impliquées dans cette
synthèse, l’ADN topo-isomérase de type II ou ADN gyrase et l’ADN topo-isomérase IV
(Hiramatsu et al, 1997). Elles ont une diffusion tissulaire et cellulaire excellente,
Les fluoroquinolones sont rapidement bactéricides, elles présentent aussi un effet
postantibiotique important et prolongé.
Les quinolones sont des antibactériens puissants, massivement utilisées pour le traitement
d’une grande variété d’infections bactériennes chez l’homme.

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 14
A : Noyau commun (Dentan, 2015)
B C
Fluor: augmentation de l’activité antibactérienne et
élargissement du spectre !
Figure 3: Structure des quinolones, A : noyau commun, B : acide nalidixique, C :
fluoroquinolone.
Fluoroquinolone

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 15
I.4 Résistance bactérienne aux antibiotiques
Le phénotype observé à l’antibiogramme n’est rien d’autre que la somme des effets liés au
contenu génétique de la souche étudiée. Comme la sensibilité ou la résistance aux
antibiotiques est génétiquement définie, l’émergence et la diffusion de la résistance chez les
bactéries n’est pas étonnante étant donné leur extraordinaire flexibilité génétique. La
résistance aux antibiotiques peut être naturelle ou acquise. La résistance naturelle est
déterminée dès le début du développement d’un antibiotique et ne pose pas de problème en
pratique clinique. Aussi, les bactéries peuvent acquérir la résistance via des mutations dans
les gènes endogènes, l’acquisition de gènes de résistance exogènes ou des mutations dans les
gènes acquis (Rice, 2012). Cependant, la résistance aux antibiotiques résulte principalement
de l’acquisition de gènes de résistance exogènes. Les progrès importants en génétique ont
permis de mieux comprendre les mécanismes par lesquels les bactéries échangent des
séquences d’ADN. Pendant de nombreuses années, il était convenu que les échangent
génétiques se faisaient uniquement à partir de structures bien connues : les plasmides et les
transposons. Ces éléments portant le plus souvent des déterminants de résistance aux
antibiotiques étaient en effet facilement caractérisables au laboratoire. L’avènement de
l’analyse génomique comparative a permis de montrer que ces vecteurs de résistance et leurs
mécanismes de transfert étaient probablement mineurs et, qu’en réalité, les échanges
génétiques entre bactéries étaient le plus souvent complexes. Ainsi, pendant des années, il
était admis que certaines souches de E. coli pouvaient être responsables d’infections urinaires
et d’autres de diarrhées. Désormais, il est évident que ces différences sont génétiquement
déterminées par l’acquisition de différents ilôts de pathogénicité de grande taille qui portent
les déterminants responsables de chacun des processus pathologiques (Welch et al, 2002).
Sans surprise les bactéries peuvent utiliser des associations de déterminants génétiques pour
évoluer vers la résistance dans des situations où les mutations ponctuelles par elles-mêmes ne
confèrent pas des niveaux de résistance suffisamment élevés sélectionnables in vivo.
L’utilisation de mécanismes auxiliaires afin d’amplifier la résistance aux fluoroquinolones
associée aux mutations dans les gènes des topoisomérases est un bon exemple. Chez les
bactéries à Gram négatif, les fluoroquinolones ont pour cible primaire l’ADN gyrase et pour
cible secondaire la topoisomérase IV (Hooper, 2001).

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 16
I.4.1 Résistance aux β-lactamines
La production d’enzyme inactivatrice (Figure 4) est le principal mécanisme de
résistance des entérobactéries aux β-lactamines. Le processus repose sur un résidu
sérine actif chez les enzymes les plus fréquentes ou sur un ion métallique Zn2+. Dans
les deux cas, l’inactivation des β-lactamines est due à l’ouverture du cycle β-lactame
au niveau de la liaison amide, selon une réaction d’hydrolyse suite à l’activation d’une
molécule d’eau (Bonnet, 2012).
Inactivation enzymatique de l’antibiotique
Bêta-lactamines / Béta-lactamases (Archambaud, 2009)
Figure 4: Inactivation enzymatique de l’antibiotique

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 17
I.4.2 Résistance aux quinolones
Pendant plus de 30 ans, les mécanismes de résistance aux quinolones connus avaient un
support chromosomique. Depuis 1998, des déterminants d’origine plasmidique ont été
découverts (Nordmann al, 2007 ; Andremont et al, 2006).
Les quinolones ont pour cible les topoisomérases de type II, ADN gyrase et topoisomérase IV
(Figure 5). Les deux grands mécanismes responsables de la résistance aux quinolones sont
l’altération de la cible avec des mutations dans les gènes des topoisomérases et l’efflux.
Ainsi, La résistance aux quinolones peut être facilitée par l’acquisition des gènes. Par
exemple, les gènes qnr, codant des protéines qui protègent les topoisomérases de l’action des
quinolones, peuvent supplémenter la résistance. Cependant plusieurs types de gènes qnr ont
été décrits (qnrA, qnrB, qnrC, qnrD et qnrS) avec différents variants. Le niveau de résistance
aux quinolones peut aussi être augmenté par l’acquisition du gène aac(6’)-Ib-cr codant pour
un variant d’une enzyme inactivatrice des aminosides qui acétyle spécifiquement la
ciprofloxacine (Rice. 2012).
L’utilisation abusive des quinolones, a conduit à l’apparition d’une résistance augmentée à
ces antibiotiques.
Chez les bactéries à Gram négatif, les mécanismes de résistance acquise par :
- mutations chromosomiques, induisent l’imperméabilité de la paroi ;
- support plasmidique, induit la protection des topoisomérases de liaison à fluoroquinolone
(ADN bactérien) et l’acétylation des quinolones donc l’inactivation enzymatique (aac(6’)-lb-
cr enzyme) (Dentan, 2015) .
Qu
in
ol
on
es

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 18
Figure 5: Mécanisme d’action des quinolones (Dentan, 2015)
Quinolones

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 19
II. Objectifs de l’étude
II.1 Objectif principal
Décrire les gènes plasmidiques de résistance aux quinolones chez les E.coli et Klebsiella spp
productrices de bétalactamase à spectre élargi isolées chez les enfants de 6 à 59 mois inclus
dans une étude menée par le centre de recherche « Epicentre » dans la région de Maradi au
Niger.
II-2 Objectifs spécifiques
Déterminer la proportion respective de chaque gène de résistance aux quinolones chez
les bactéries productrices de BLSE ;
Rechercher les gènes de résistance aux quinolones chez les souches sensibles à l’acide
nalidixique mais résistante aux fluoroquinolones sur l’antibiogramme standard, par
des techniques approfondies (PCR).

MATERIEL ET METHODES

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 20
III Matériel et méthodes
III.1 Cadre de l’étude
La collecte de nos échantillons a été effectuée au laboratoire du centre de Recherche
Epicentre à Maradi, au Niger, l’identification des souches a été confirmée par Galerie
API20E au laboratoire de l’Hôpital Saint Camille de Ouagadougou (HOSCO). Les analyses
moléculaires ont été réalisées au Laboratoire de Biologie moléculaire et de Génétique
(LABIOGENE) au Centre de Recherche Biomoléculaire Pietro Annigoni (CERBA).
EPICENTRE est une association à but non lucratif créée par Médecins Sans Frontières
(MSF) en 1986 qui mène des activités de recherche et de formation sur les programmes de
MSF ainsi que dans ses propres centres de recherche au Niger et en Ouganda. Epicentre est
un centre collaborateur de l’OMS.
CERBA/LABIOGENE, relève du district sanitaire de Bogodogo. Créé par la délégation
Camillienne, ce centre a pour missions premières la formation et la promotion de jeunes
chercheurs en particulier dans le domaine biomoléculaire où il dispose d’un plateau technique
moderne (appareils de PCR en temps réel, HPLC, séquenceur, etc.). Les activités de
recherche s’articulent autour de plusieurs axes principaux. Le premier axe qui concerne la
recherche fondamentale et les sciences du génome se décompose en sous-axes sur les
pathologies génétiques (hémoglobinopathies, ostéoporose, déficiences génétiques), les
marqueurs de résistance génétique (résistance bactérienne aux antimicrobiens), le diagnostic
moléculaire de diverses pathologies en particulier des pathologies émergentes (VIH, HPV,
VHC, VHB) ainsi que sur la vaccinologie. Le deuxième axe fait intervenir des études de la
médecine traditionnelle (travaux sur des recettes traditionnelles fournies par des tradi-
praticiens). Les deux derniers axes comprennent la recherche clinique (suivi biologique et
clinique des personnes vivant avec le VIH, activités portant sur la PTME, expérimentations
pharmaco-cliniques) et la recherche en nutrition (réhabilitation nutritionnelle des enfants
malnutris).

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 21
III.2 Type et période d’étude
Il s’agit d’une étude rétrospective descriptive réalisée sur diverses espèces d’entérobactéries
productrices de BLSE et résistantes aux quinolones, isolées de selles d’enfants de 6 à 59 mois
inclus dans un essai clinique mené par Epicentre dans la région de Maradi au Niger, entre
Février et Novembre 2013. Les analyses moléculaires ont été effectuées au
CERBA/LABIOGENE, Université de Ouagadougou, Burkina Faso entre Juillet et Aout 2015.
III.3 Echantillonnage
Epicentre a mené un essai clinique randomisé dont l’objectif était d’évaluer l’effet de
l’utilisation systématique de l’amoxicilline pour le traitement en ambulatoire des enfants
sévèrement malnutris sans complication versus placebo à Maradi au Niger de 2012 à 2013.
Une étude ancillaire a également été menée sur un sous-échantillon d’enfants inclus dans
l’étude générale afin de décrire l’émergence des résistances aux C3G dans la flore intestinale.
Durant les visites régulières à l’inclusion dans l’étude, au 7ème et au 28ème jour, les enfants
sélectionnés dans l’étude ancillaire ont fourni un échantillon de selles (écouvillon ou selle en
pot) prélevé sur les sites et conservés à température de +4 ºC jusqu’à leur acheminement au
laboratoire d’Epicentre.
Tous les échantillons de selles collectés dans le cadre de cette étude ancillaire ont été
transportés au laboratoire Epicentre de Maradi sous condition de température de +4 ºC le
jour même pour analyse bactériologique. Dans ce laboratoire, la présence de BLSE a été
déterminée par la mise en culture sur le milieu chromagar BLSE qui est un milieu sélectif. Ce
milieu a permis d’isoler les colonies roses, bleues foncées, blanches suspectant
respectivement E. coli, Klebsiella et Salmonella. Une colonie de chaque espèce a été placé
dans un tube contenant un milieu de conservation et stocké à +4 ºC.
Les souches ont été collectées sur les 04 sites de l’étude (Madarounfa, Gabi, Dan-issa et
Tofa) du centre de recherche Epicentre à Maradi dans la période allant du 18 février au 11
Novembre 2013 pour les trois premiers sites et Juillet 2013 pour le site de Tofa.
Des études approfondies sur les différents isolats récoltés ont été réalisées au laboratoire de
biologie moléculaire CERBA (Centre de Recherche Biomoléculaire Pietro
Annigoni)/LABIOGENE (laboratoire de biologie moléculaire et de génétique) du
département de Biochimie-Microbiologie situé dans l’Unité de Formation et de Recherche en
Science de la Vie et de la Terre (URF/SVT) à l’Université de Ouagadougou.

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 22
Soixante dix huit (78) souches d’entérobactéries productrices de BLSE et résistantes aux
quinolones nous ont été fournies par le laboratoire de l’EPICENTRE Maradi-Niger. Ces
souches sont issues des prélèvements de selles provenant des enfants de six (06) à cinquante-
neuf (59) mois inclus dans l’étude Epicentre entre Février et Novembre 2013.
III.3.1 - Mode de sélection
A partir de la base de données de l’étude ancillaire nous avions sélectionné de façon aléatoire
en respectant la proportion de la répartition de la population d’étude les 78 souches en
fonction de la couleur des colonies : roses, et bleues suspectant respectivement E. coli, et
Klebsiella.
III.3.2 - Critère d’inclusion
Les souches bactériennes inclus dans cette étude répondent aux critères d’inclusion suivants:
- Isolées d’un prélèvement de selles au moment de l’inclusion des enfants dans l’étude
générale;
- Collectées entre le 18 février et le 11Novembre 2013;
- BLSE isolées et conservées au laboratoire d’Epicentre à Maradi;
- Repoussent sur Hektoen;
- Résistantes aux quinolones et aux fluoroquinolones selon l’antibiogramme standard
du laboratoire de l’Epicentre à Maradi;
- Les souches de couleur ne concordant pas aux deux espèces cités ont été remplacés
par une liste randomisée;
- Les souches sensibles à l’acide nalidixique mais résistantes aux fluoroquinolones sur
l’antibiogramme standard.
III.3.3 -Critères d’exclusion
Les souches isolées dans des prélèvements collectés à la semaine 01 et 04 du suivi n’ont pas
été inclues dans l’étude.

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 23
III.4 Matériel
III.4.1 Réactifs
Les amorces et le Master Mix universel, KIT.
III.4.2 Appareillage
La PCR a été réalisée à l’aide du Thermocycleur GeneAmp PCR System 9700 (Applied
Biosystems, Courtabœuf, France).
III.4.3 Autres matériels
D’autres matériels ont été utilisés à savoir les milieux de culture, la galérie API 20E, les
micropipettes, le vortex, les cônes ou embouts, des incubateurs, une centrifugeuse, une hotte
de sécurité, des tubes stériles, des congélateurs, et divers autres matériels de laboratoire.
III.5 Méthodes
III.5.1 Confirmation de l’identité des souches
Après repiquage sur milieu Hektoen, l’identification des souches a été confirmée par la
galerie API 20E au laboratoire de l’Hôpital Saint Camille de Ouagadougou (HOSCO).
III.5.2. Sensibilité aux antibiotiques
La sensibilité aux antibiotiques a été déterminée par antibiogramme qui a été réalisé par la
méthode de diffusion en milieu gélosé de Muller-Hinton (MH) selon les recommandations du
CA-SFM 2013 (Comité de l’antibiogramme de la Société française de microbiologie
(microbiologie 2013).
Les soixante-dix-huit (78) isolats (E. coli, Klebsiella) ont été testés par le laboratoire
d’Epicentre pour leur sensibilité à 16 antibiotiques appartenant à 05 familles. Les familles
d’antibiotiques utilisées sont les suivantes : les bétalactamines (Amoxicilline, Amoxicilline +
Acide clavulanique, Tircarcilline Cefalotine, Cefoxitine, Cefotaxime, Ceftazidime, Cefipime,
Imipenème, Ertapenème), les quinolones (Acide nalidixique, Ofloxacine), les aminosides
(Amikacine, Gentamicine) les cyclines (Tétracycline) et les sulfamides (Cotrimoxazole).
E.coli CIP 7624 a été utilisé comme souche de référence.

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 24
III.5.3 Condition de cultures des souches
Pour les identifications et les purifications, les bactéries ont été repiquées sur des milieux
Chromagar Orientation et Hektoen en boites de pétri et incubées à 37 ºC pendant 24h .Pour
les extractions des ADN,
les bactéries ont été cultivées dans un bouillon de culture Luria-Bertani (LB) tamponné à pH
7 et incubées à 37 ºC pendant 24h.
III.5.4 Extraction des ADNs
Les souches conservées sur milieu Leminor ont été repiquées sur milieu TCS puis incubées à
37 °C pendant 24h. Quelques colonies bactériennes issues de cette culture ont été ensuite
cultivées dans 2 ml de bouillon de culture Luria-Bertani (LB) tamponné à pH 7 et incubées à
37 ºC pendant 24h.
Les culots bactériens obtenus par centrifugation à 10000 tours par minute pendant 5 minutes
à la centrifugeuse (eppendorf Centrifuge 5417C) ont été mis en suspension par 500 µl de
tampon phosphate 100Mm pH 7 puis porter à 100 °C (eau bouillante) pendant 15 minutes et
centrifugés à 10000 tours par minute pendant 10 minutes. Les surnageants recueillis ont été
mis en suspension dans 250 µl d’alcool absolu et laissés à température ambiante pendant 5
minutes ensuite centrifugés à 10000 tours par minute pendant 10 minutes ; les surnageants
ont été rejetés et les culots ont été recueillis.
Les culots obtenus ont été mis en suspension dans 1000 µl d’alcool 75 °C, vortexés puis
centrifugés à 5000 tours pendant 5 minutes. Les lavages ont été effectués deux fois et le culot
recueilli a été séché pendant 24h sous hotte couverte de papier essuie-tout. L’ADN est repris
dans 100 µl d’eau distillé stérile puis conservé à -80 °C.
III.5.5 Amplification des gènes de quinolones
La PCR a été réalisée dans un volume réactionnel de 50 µl. Deux programmes
d’amplification ont été utilisés pour la recherche des quatre gènes (Tableau I) (Dallenne et al,
2010 ; Robicsek et al, and Park et al, 2006). Les gènes qnrA, qnrB et qnrS ont été amplifiés
par PCR multiplex.
Les gènes aac (6´) ont été amplifiés par PCR simplex (Park et al, 2006).

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 25
Tableau I : Protocoles de PCR du gène qnr
Condition/durée
Paramètres qnrA qnrB qnrS aac
Dénaturation initiale 94 °C/10 mn 94 °C/10 mn 94 °C/10 mn 94 °C/10 mn
Dénaturation 94 °C/45 s 94 °C/45 s 94 °C/45 s 94 °C/45 s
Appariement 53 °C/45 s 53 °C/45 s 53 °C/45 s 55 °C/45 s
Elongation 72 °C/1 mn 72 °C/1 mn 72 °C/1 mn 72 °C/45 s
Elongation Finale 72 °C/7 mn 72 °C/7 mn 72 °C/7 mn 72 °C/7 mn
Nombre de cycles 32 32 32 34
Les séquences des amorces utilisées au niveau de cette étude ont été mentionnées dans le
tableau II. Ces amorces ont été fournies par la maison ABI. L’amplification a été effectuée à
l’aide du thermocycleur GeneAmp 9700 PCR system de Applied Biosystems.

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 26
Tableau II: Séquences des amorces utilisées au niveau de cette étude
Gènes recherchés Séquences des amorces (5'-3') paires de bases références
qnrA for 5′-ATTTCTCACGCCAGGATTTG 516 ( Robicsek et al, 2006)
Rev 5′-GATCGGCAAAGGTTAGGTCA
qnrB For 5′-GATCGTGAAAGCCAGAAAGG 469 ( Robicsek et al, 2006)
Rev 5′-ACGATGCCTGGTAGTTGTCC
qnrS For 5′-ACGACATTCGTCAACTGCAA 417 (Robicsek et al, 2006)
Rev 5′-TAAATTGGCACCCTGTAGGC
aac(6) For 5′-TTGCGATGCTCTATGAGTGGCTA 482 ( Park et al, 2006)
Rev 5′-CTCGAATGCCTGGCGTGTTT
III.5.6 Electrophorèse sur gel d’agarose
Les fragments d’ADN amplifiés par PCR étaient séparés par électrophorèse sur un gel
d’agarose à 2 % préparé dans une solution de tris base - borate – EDTA 1X et contenant 16 µl
de bromure d’éthidium. La migration s’est réalisée sous une tension de 100 volts (V) pendant
1H.
Le tampon de migration utilisé a été le Tris base- borate – EDTA 1X. La migration a été
suivie grâce à une solution de dépôt (bleu de charge) ajoutée à la solution d’ADN à environ
20 % du volume. Les tailles des différents fragments ont été déterminées en déposant à côté
des puits contenant l’ADN des marqueurs de masses moléculaires. Ces fragments étaient
ensuite visualisés sous lumière UV. Des fragments d’ADN de 516 paires de base (pb) étaient
obtenus en cas de présence de qnrA, de 482 paires de base en cas de présence de aac(6’) , de
469 paires de base en cas de présence de qnrB et de 417 paires de base en cas de présence de
qnrS.

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 27
III.5.7 Analyse des données
Les données ont été traitées à l’aide du logiciel SPSS Statistics 21.0.
III.5.8 Considérations éthiques
Cette étude a obtenu l’accord du comité Consultatif National d’Ethique du Niger (CCNE),
notamment pour le transfert des souches du laboratoire d’Epicentre de Maradi, Niger au
laboratoire de biologie moléculaire CERBA/LABIOGENE, Ouagadougou, Burkina-Faso.

RESULTATS
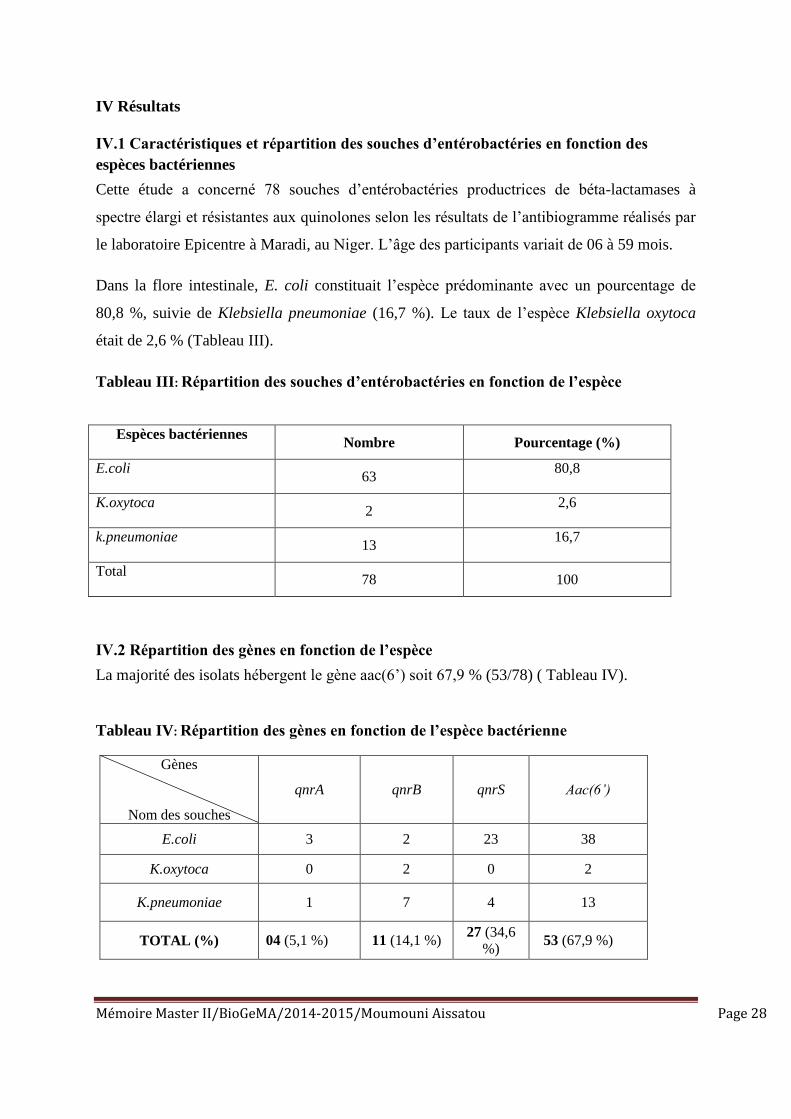
Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 28
IV Résultats
IV.1 Caractéristiques et répartition des souches d’entérobactéries en fonction des
espèces bactériennes
Cette étude a concerné 78 souches d’entérobactéries productrices de béta-lactamases à
spectre élargi et résistantes aux quinolones selon les résultats de l’antibiogramme réalisés par
le laboratoire Epicentre à Maradi, au Niger. L’âge des participants variait de 06 à 59 mois.
Dans la flore intestinale, E. coli constituait l’espèce prédominante avec un pourcentage de
80,8 %, suivie de Klebsiella pneumoniae (16,7 %). Le taux de l’espèce Klebsiella oxytoca
était de 2,6 % (Tableau III).
Tableau III: Répartition des souches d’entérobactéries en fonction de l’espèce
Espèces bactériennes
Nombre Pourcentage (%)
E.coli
63
80,8
K.oxytoca
2
2,6
k.pneumoniae
13
16,7
Total
78 100
IV.2 Répartition des gènes en fonction de l’espèce
La majorité des isolats hébergent le gène aac(6’) soit 67,9 % (53/78) ( Tableau IV).
Tableau IV: Répartition des gènes en fonction de l’espèce bactérienne
Gènes
Nom des souches
qnrA qnrB qnrS Aac(6’)
E.coli 3 2 23 38
K.oxytoca 0 2 0 2
K.pneumoniae 1 7 4 13
TOTAL (%) 04 (5,1 %) 11 (14,1 %) 27 (34,6
%) 53 (67,9 %)

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 29
IV.3 Répartition de l’association des gènes en fonction des espèces bactériennes
La résistance associée aux quinolones dans le cas de BLSE a été observée dans 88,46 %
(69/78) soit 22 % de l’ensemble des souches de Klebsiella. Aucune des souches résistantes
aux béta-lactamases n’héberge à la fois les gènes qnrA, qnrB et qnrS mais il existe une
association des gènes de résistance BLSE, qnr et aac(6’) chez 26 souches (33,32 %) répartis
comme suit 12 E. coli, 02 Klebsiella oxytoca et 12 Klebsiella pneumoniae ( Tableau V).
Tableau V: Répartition de l’association des gènes en fonction des espèces bactériennes
Espèces bactériennes
Association Qnr-BLSE-aac Pourcentage ( % )
E.coli
12 15,38
K.oxytoca
2 2,56
k.pneumoniae
12 15,38
Total 26 33, 32
IV.4 Répartition de gènes chez les souches sensibles à l’acide nalidixique et résistantes
aux fluoroquinolones
Parmi les 21 souches (16 E. coli, 04 K. pneumoniae et 01 K. oxytoca) sensibles à l’acide
nalidixique mais résistante aux fluoroquinolones soit 26,9 %, on observe la présence du gène
qnrA chez 01 souche, le gène qnrB est retrouvé chez 03 souches, 16 souches présentent une
résistance au qnrS, 09 souches montrent une résistance au gène aac(6’) (Tableau VI).
Tableau VI : Répartition de gènes chez les souches sensibles à l’acide nalidixique et
résistantes aux fluoroquinolones
Gènes
Nom des souches
qnrA qnrB qnrS Aac(6’)
E.coli 0 0 15 4
K.oxytoca 0 1 0 1
K.pneumoniae 1 2 1 4
TOTAL 01 03 16 09

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 30
IV.5 Caractérisation des gènes codant pour la résistance aux quinolones
Les ADN des 57 souches répondant positives à la résistance aux quinolones et de 21 souches
sensibles aux quinolones mais résistantes aux fluoroquinolones ont été soumis à la technique
de PCR pour la recherche du type de gène qnr et aac(6’) en utilisant les amorces spécifiques
pour qnrA, qnrB, qnrS et aac(6’). L’examen des produits PCR après électrophorèse sur gel
d’agarose a permis de noter que 69 isolats se sont révélés porteurs d’au moins un des gènes
recherchés et 09 isolats ne portent aucun des gènes recherchés. 26 isolats hébergent à la fois
le gène qnr et aac(6’), 04 isolats hébergent le gène qnrA, 11 isolats hébergent le gène qnrB,
27 isolats hébergent le gène qnrS, 53 isolats hébergent le gène aac(6’)
(Tableau VII)

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 31
Tableau VII: Données des antibiogrammes et Résultats de la PCR
Age(mois) sexe Nom souche AMX TIC CF FOX TE AMC CTX fep IMP CAZ ETP NA OFX GM AN SXT qnr A qnr B qnr S aac(6)
9 M
E.coli R R R S R R R R S R S R R R S R neg neg neg pos
6 M
E.coli N R R S R I R R S R S R R R S R neg neg neg pos
11 F
E.coli R R R S R I R I S R S S R S S R neg neg pos pos
7 F
E.coli R R R S R I R R S R S R R R S R neg pos neg pos
20 F
E.coli R R R S R S R R S R S S R S S R neg neg pos neg
9 F
E.coli R R R S R I R R S R S R R R S R neg neg neg pos
29 M
E.coli R R R S R S R R S R S S R S S R neg neg pos pos
7 M
E.coli R R R S R S R R S R S R R R S R neg neg neg pos
26 F
E.coli R R R S R I R R S R S R R R S R neg neg neg pos
22 F
E.coli R R R S R I R R S R S R R R S R neg neg neg pos
30 M
E.coli R R R S R I R R S R S R R S S R neg neg pos pos
19 F
E.coli R R R S R S R R S R S R R S S R pos neg neg pos
11 M
E.coli N R R S R S R R S R S R R S S R neg neg pos neg
14 F
E.coli R R R S R I R R S R S R R R S R neg neg neg pos
29 M
E.coli R R R S R S R I S R S S R S S R neg neg pos neg
19 M
E.coli R R R S R I R R S R S R R S S R neg neg neg neg
12 M
k.pneumoniae R R R S R I R R S R S I R R I R neg pos neg pos
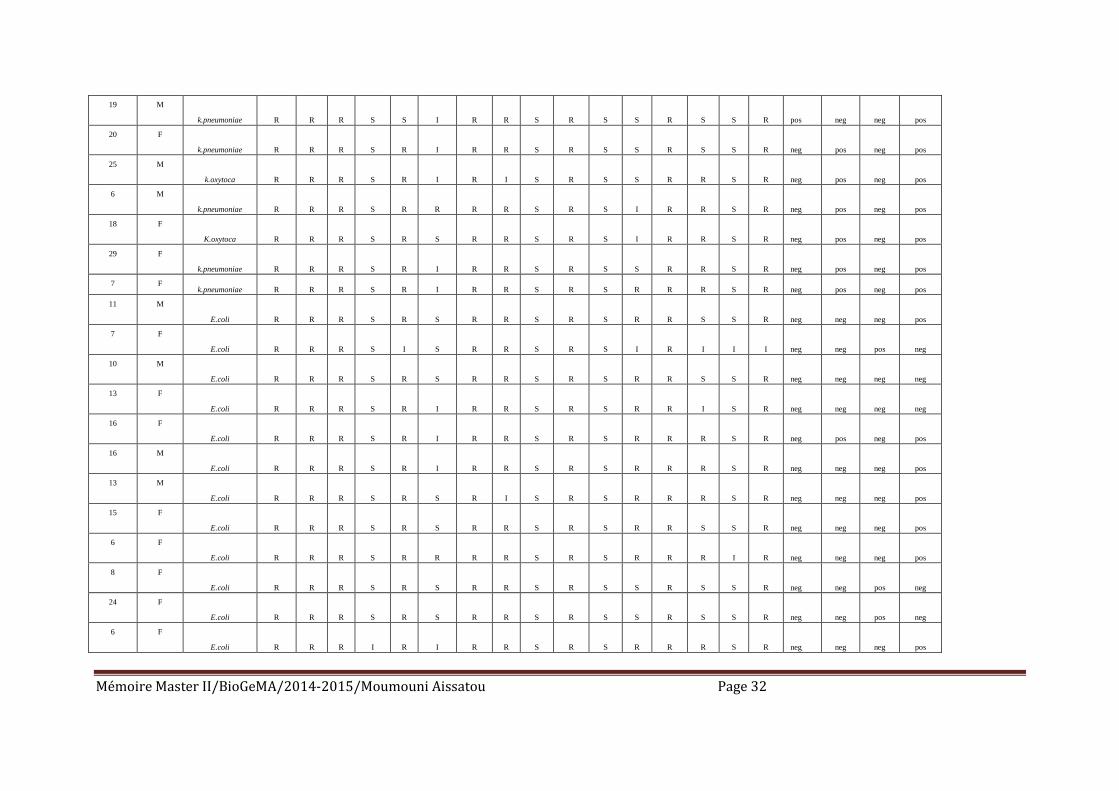
Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 32
19 M
k.pneumoniae R R R S S I R R S R S S R S S R pos neg neg pos
20 F
k.pneumoniae R R R S R I R R S R S S R S S R neg pos neg pos
25 M
k.oxytoca R R R S R I R I S R S S R R S R neg pos neg pos
6 M
k.pneumoniae R R R S R R R R S R S I R R S R neg pos neg pos
18 F
K.oxytoca R R R S R S R R S R S I R R S R neg pos neg pos
29 F
k.pneumoniae R R R S R I R R S R S S R R S R neg pos neg pos
7 F k.pneumoniae R R R S R I R R S R S R R R S R neg pos neg pos
11 M
E.coli R R R S R S R R S R S R R S S R neg neg neg pos
7 F
E.coli R R R S I S R R S R S I R I I I neg neg pos neg
10 M
E.coli R R R S R S R R S R S R R S S R neg neg neg neg
13 F
E.coli R R R S R I R R S R S R R I S R neg neg neg neg
16 F
E.coli R R R S R I R R S R S R R R S R neg pos neg pos
16 M
E.coli R R R S R I R R S R S R R R S R neg neg neg pos
13 M
E.coli R R R S R S R I S R S R R R S R neg neg neg pos
15 F
E.coli R R R S R S R R S R S R R S S R neg neg neg pos
6 F
E.coli R R R S R R R R S R S R R R I R neg neg neg pos
8 F
E.coli R R R S R S R R S R S S R S S R neg neg pos neg
24 F
E.coli R R R S R S R R S R S S R S S R neg neg pos neg
6 F
E.coli R R R I R I R R S R S R R R S R neg neg neg pos

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 33
9 F
E.coli R R R S S I R R S R S R R R S R neg neg neg pos
29 M
E.coli R R R S R S R R S R S R R S S R neg neg neg neg
9 M
E.coli R R R S R S R R S R S S R S S R neg neg pos neg
8 M
E.coli R R R S R S R R S R S R R R S R neg neg neg neg
7 F
E.coli R R R S R S R R S R S S R S S R neg neg pos neg
32 M
E.coli R R R S R S R R S R S S R S S R neg neg pos neg
34 M
E.coli R R R S R S R R S R S R R S S R neg neg neg neg
24 F
E.coli R R R S R S R R S R S S R S S R neg neg pos neg
23 M
E.coli R R R S R I R R S R S R R R S R neg neg neg pos
6 M
E.coli R R R S R S R R S R S S R S S S neg neg neg pos
10 F
E.coli R R R S R I R R S R S R R R S R neg neg neg pos
12 F
E.coli R R R S R I R R S R S R R R S R neg neg neg pos
8 M
E.coli R R R S R I R R S R S R R S S R neg neg neg pos
22 F
E.coli R R R S R S R R S R S S R S S S neg neg pos neg
24 M
E.coli R R R S R S R R S R S S R S S R neg neg pos neg
18 F
E.coli R R R S R I R R S R S R R R S R neg neg pos neg
14 M
E.coli R R R S R R R R S R S R R R S R neg neg neg pos
8 M
E.coli N R R S R S R R S R S R R S S R neg neg pos pos
7 F
E.coli R R R S R S R R S R S R R R S R neg neg neg neg

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 34
11 F
E.coli R R R S R S R R S R S R R S S R neg neg neg pos
22 F
k.pneumoniae R R R S R I R I S R S I R R S R neg neg pos pos
32 M
k.pneumoniae R R R S R I R R S R S R R R S R neg neg pos pos
12 F
k.pneumoniae R R R S S I R R S R S R R R S R neg neg neg pos
24 M
k.pneumoniae N R R S R I R R S R S I R R S R neg neg pos pos
8 M
k.pneumoniae N R R S R I R R S R S R R R S R neg pos neg pos
34 M
k.pneumoniae R R R S S I R R R R S S R R S R neg neg pos pos
22 F
k.pneumoniae R R R S R S R R S R S I R S S R neg pos neg pos
21 F
E.coli R R R S R S R R S R S S R S S R neg neg pos neg
7 F
E.coli R R R R R R R R S R S R R S S R neg neg neg pos
6 M
E.coli R R R S R S R R S R S R R R I R neg neg neg neg
12 M
E.coli R R R S R S R S S R S S R S S R neg neg pos neg
6 F
E.coli R R R S R I R R S R S R R S S R pos neg neg pos
22 F
E.coli R R R S R I R I S R S I R S S R neg neg pos neg
21 M
E.coli R R R R R R R I S R S R R R S R neg neg neg pos
22 M
E.coli R R R S R I R R S R S I R I I R neg neg pos pos
6 F
E.coli R R R S R I R R S R S R R R S R neg neg neg neg
9 M
E.coli R R R S R I R R S R S I R I S R neg neg pos pos
12 M
E.coli R R R S R I R I S R S S R S S R neg neg pos pos

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 35
11 M
E.coli R R R S R I R R S R S R R R S R neg neg neg pos
11 M
E.coli R R R S R I R R S R S R R R S R neg neg neg pos
22 M
E.coli R R R S R I R R S R S R R S S R pos neg neg pos
24 M
E.coli R R R S R I R R S R S R R R S R neg neg neg pos
NB: les qnr A-B-S sont les gènes plasmidiques de resistance aux quinolones ; aac(6’) est le gène de resistance aux fluoroquinolone ; Légende : Neg : négatif ; Pos : positif

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 36
Des amplicons de 516 paires de base (pb) étaient obtenus en cas de présence de qnrA, de 469
paires de base en cas de présence de qnrB et de 417 paires de base en cas de présence de qnrS
( Figure 6) et des amplicons de 482 paires de base étaient obtenus en cas de présence de
aac(6’) (Figure 7)
Figure 6: Gel d’agarose des produits PCR des, qnrB et qnrS qnrA
PM: Marqueur de taille (100pb); 1,2,4,8: Souches qnr(S+); 3,15: Souches qnr(A+); 5,6,7,9:
Souches qnr (B+)
Figure 7: Gel d’agarose des produits PCR du aac(6’)-Ib.
PM: Marqueur de taille (100pb) ; 1 à 15 : Souches aac(6’) (+)
1 2 3 1 4 5 6 7 8 9 15 PM
500pb
1 15
500 pb
PM

DISCUSSION

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 37
V. Discussion
Selon la présente étude les gènes qnr et aac(6’) ont une prévalence de 88,45 %, avec une
prédominance du variant aac(6’) (67,9 %), suivi du qnrS (34,6 %) ,de qnrB (14,1 %) et
qnrA (5,1 %).
Ces fréquences sont supérieures à celles rapportée en Turquie, où la prévalence de gènes de
résistance à médiation plasmidique de déterminantes quinolones a été rapportée à 30 % des
souches (Mammeri et al, 2005 ; Corkill et al, 2005) et le gène qnrA a été retrouvé chez 6,3 %
(Mehmet et al, 2008).
Par ailleurs, il existe une fréquente association entre les déterminants géniques de type qnr et
ceux des BLSE, ce qui souligne la possibilité d’une co-sélection de ces deux mécanismes de
résistance plasmidique (Lavilla et al, 2008).
Une étude menée en Espagne a évalué la prévalence des gènes qnr chez 305 souches
d’entérobactéries productrices de BLSE (Lavilla et al, 2008). Les résultats de cette étude ont
révélé une prévalence de
4,9 % de l’association « qnr–BLSE » Une autre étude menée en Côte d’Ivoire a évalué la
prévalence des gènes qnr chez 151 souches d’entérobactéries productrices de BLSE
(Guessennd et al, 2007). La prévalence de l’association « qnr-BLSE » au cours de cette étude
était de 27 % en moyenne pour l’ensemble des entérobactéries alors que dans notre étude, 23
souches multirésistantes ont été identifiées (BLSE + résistance aux qnr).
Ces souches représentent 29,49 % (23/78). Cela pourrait s’expliquer par la petite taille de
notre échantillon.
Aux états unis, (Park et al, 2006) ont rapporté que les gènes responsables de la résistance aux
quinolones (qnr) ont été trouvés chez les entérobactéries ; la même étude révèle la prévalence
de l’allèle du gène aac(6’) de 14%. Cette donnée est confirmée par (Karah.N et al, 2010) en
Tunisie alors que dans notre étude le gène aac(6’) est majoritaire avec une prévalence de 67,9
% .
Dans notre étude, la distribution de la variante aac(6’) diffère entre les espèces. Elle est
détectée le plus souvent chez les E. coli suivies des K. pneumoniae, donnée confirmée par
l’étude de Park et al, 2006.

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 38
Bien que la variante de gène aac(6’) n’a pas été signalée jusqu’en 2006, elle était déjà
présente dans plus de la moitié des isolats multirésistants de E. coli à Shanghai, en Chine, en
2000 et 2001 ( Wang et al, 2003).
Les différents antibiogrammes des espèces étudiés suggèrent que la diffusion de l'aac (6’) -
Ib-Cr ne se produit pas à travers la propagation clonale ou la propagation d'un plasmide
unique. La diversité des plasmides sur laquelle circule ce gène n’est pas encore connue, mais
sa présence dans une cassette d'intégrons (Casin et al, 1998 ; Robicsek et al, 2006) suggère
qu'il pourrait être largement mobile parmi les plasmides.
L’association des gènes de résistance qnr et aac(6’) confère un haut niveau de resistance aux
bactéries. Notre étude révèle l’existence d’une association des gènes de résistance qnr et
aac(6’) chez 26 souches répartis comme suit 12 E. coli, 2 Klebsiella oxytoca et 12 Klebsiella
pneumoniae mais ces données sont différentes de celles rapportées par (Robicsek et al, 2006 ;
Wang et al, 2003) où il n’existe aucune association entre l’aac(6’) et les gènes qnr.
Les résultats de la sensibilité à l’acide nalidixique et résistante aux fluoroquinolones ont
montré en général que les bactéries présentent un taux de résistance aux antibiotiques. En
effet, 21 souches (16 E. coli, 04 K. pneumoniae et 01 K.oxytoca) sensibles à l’acide
nalidixique mais résistante aux fluoroquinolones ont révélés la présence des gènes
plasmidiques de résistance aux quinolones avec une prévalence de 26,9 %. Cependant, on
observe la présence du gène qnrA chez 01 souche, le gène qnrB est retrouvé chez 03 souches,
16 souches présentent une résistance au qnrS, 09 souches montrent une résistance au gène
aac(6’).
Une des limites de notre étude est que nous n’avons pas disposé des informations sur les CMI
des antibiotiques ciblés comme par exemple les céphalosporines de 3ème génération
(ceftazidime, cefotaxime), les quinolones (acide nalidixique) et fluoroquinolones (ofloxacine
et ciprofloxacine) pouvant confirmer la résistance de ces antibiotiques avant la PCR.
Une autre limite à notre étude est le manque des caractéristiques moléculaires des gènes de
résistance aux béta-lactamases, nous nous sommes contentés seulement de l’aspect
phénotypique révélé par l’antibiogramme classique.
Malgré ces limites, les résultats de la présente étude nous paraissent importants et pertinents
en termes de recherche sur les gènes qnr, aac(6’) en santé publique.

CONCLUSION ET PERSPECTIVES

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 39
Conclusion
Les bactéries ont accès à un nombre important d’éléments génétiques pour développer la
résistance aux antibiotiques auxquels elles sont confrontées. Si la résistance ne peut être
acquise par mutation(s) de gènes domestiques, les nombreuses possibilités de transfert
d’information génétique dont les bactéries sont pourvues leur permettent d’acquérir la
résistance à partir d’autres espèces. L’utilisation fréquente des antibiotiques à l’hôpital et
dans la communauté est responsable d’une pression de sélection permanente favorisant
l’émergence de la résistance parmi les souches cliniques, la multi-résistance est le plus
souvent le résultat d’une accumulation régulière de gènes exogènes.
Les résultats de notre étude nous ont permis de révéler les types de gènes plasmidiques de
résistance aux quinolones circulant dans les milieux pédiatriques de notre pays, cela nous
permettra aussi d’envisager à l’image d’autres pays un réseau de surveillance de gènes de
résistance aux quinolones.
Cependant, la caractérisation des gènes de résistance aux quinolones nous a permis une
meilleure compréhension de la transmission inter-espèce et la mise en évidence des nouvelles
résistances.
Perspectives
Améliorer la détection des bas niveaux de résistance avant qu’ils n’aient un impact clinique
pourrait aider à prévenir les résistances de plus haut niveau.
Promouvoir l’instauration d’une surveillance épidémiologique afin de vérifier la validité des
protocoles thérapeutiques de première intention et de proposer d’éventuelles mesures
adaptatives.

REFERENCES BIBLIOGRAPHIQUES

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 40
REFERENCES BIBLIOGRAPHIQUES
CA-SFM, Comité d’antibiogramme de la Société Française de Microbiologie, 2013.
ACAR, J. E. C., PATRICE 1998. La fin de l’âge d’or des antibiotiques. La Recherche, 314,
50-53.
AMBLER, R. P. 1980. The structure of Béta-lactamases. Philos Trans R Soc Lond B Biol Sci,
289, 321-31.
BALL P. 2000. Quinolone generations: natural history or natural selection? J. Antimicrob.
Chemother, 46, 17-24.
BONNET, R. 2012. Β-LACTAMINES ET ENTEROBACTERIES. ANTIBIOGRAMME, 16,
165-187.
CASIN, I., F. BORDON, P. BERTIN, A. COUTROT, I. PODGLAJEN, R. BRASSEUR,
AND E. COLLATZ 1998. Aminoglycoside 6_-N-acetyltransferase variants of the Ib
type
with altered substrate profile in clinical isolates of Enterobacter cloacae and Citrobacter
freundii. Antimicrob. Agents Chemother, 42, 209–215.
CATTOIR, V., POIREL, L., ROTIMI, V., SOUSSY, C. J. & NORDMANN, P. 2007.
Multiplex PCR for detection of plasmid-mediated quinolone resistance qnr genes in
ESBL-producing enterobacterial isolates. J Antimicrob Chemother, 60, 394-7.
CHI HYE PARK, A. R., GEORGE A. JACOBY, DANIEL SAHM, AND DAVID C.
HOOPER 2006. Prevalence in the United States of aac(6_)-Ib-cr Encoding a
Ciprofloxacin-Modifying Enzyme_. ANTIMICROBIAL AGENTS AND
CHEMOTHERAPY, 50, 3953–3955.
CORKILL, J. E., ANSON, J.J AND HART. 2005: High prevalence of the plasmid-mediated
quinolone resistance determinant (qnrA) in multidrug resistant Enterobacteriaceae
from blood cultures in Liverpool, UK. J Antimicrob.chemother, 56, 1115-1117.
DALHOFF, A. 2012. Interdisciplinary Perspectives on Infectious Diseases
DALLENNE, C., DA COSTA, A., DECRE, D., FAVIER, C. & ARLET, G. 2010.
Development of a set of multiplex PCR assays for the detection of genes encoding
important beta-lactamases in Enterobacteriaceae. J Antimicrob Chemother, 65, 490-5.
DELMÉE, P. M. 2004. Microbiologie médicale.
DENTAN, C. 2015. Interne. Service des Maladies infectieuses
FRANÇOIS DENIS, M.-C. P. L., CHRISTIAN MARTIN, EDOUARD BINGEN, ROLAND

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 41
QUENTIN. 2007. Bactériologie Médicale, Techniques Usuelles. Elsevier Masson
SAS.
GENÈVE, Premier rapport de l’OMS sur la résistance aux antibiotiques.2014. Une menace
grave d’ampleur mondiale.
GIRAUD-MORIN, C. & FOSSE, T. 2008. [Recent evolution and characterization of
extended-spectrum beta-lactamase producing enterobacteria in the CHU of Nice
(2005-2007)]. Pathol Biol (Paris), 56, 417-23.
GLUPCZYNSKI , Y., LAMBERT, D., SPINEWINE, A., TULKENS, P. M., VAN
BAMBEKE, F.,VAN LAETHEM, Y., VAN LANDUYT ,H. 2002. Pharmacologie et
pharmacothérapie des anti-infectieux. Antimicrob Agents Chemother, 42, 13-26.
GUESSENND N, B. S., GBONON V, KAKOU-NDOUBA A, LAMBERT T, DOSSO P
2007. Qnr-type quinolones resistance in extended-spectrum β-lactamases producing
enterobacteria in Abidjan, Ivory Coast. 27e réunion interdisciplinaire de
chimiothérapie anti-infectieuse.
HALL, B. G. & BARLOW, M. 2005. Revised Ambler classification of {beta}-lactamases. J
Antimicrob Chemother, 55, 1050-1.
HIRAMATSU, K., H. HANAKI, T. INO, K. YABUTA, T. OGURI, AND F.C.TENOVER
1997. Methicillin-resistant staphylococcus aureus clinical strain with reduced
vancomycin susceptibility. J. Antimicrob. Chemother, 40, 135-136.
HOOPER, D. C. 2001. Emerging mechanisms of fluoroquinolone resistance. Emerg. Infect.
Dis, 7, 337- 341.
MEHMET ALI OKTEM, Z. G., MERAL BIÇMEN, DENIS GUR AND HITIT PROJECT
STUDY GROUP 2008. qnrA prevalence in extended spectrum β-lactamase-positive
Enterobacteriaceae isolates from Turkey. Jpn.J.Infect.Dis, 61, 13-17.
JACOBY, G., CATTOIR, V., HOOPER, D., MARTINEZ-MARTINEZ, L., NORDMANN,
P., PASCUAL, A., POIREL, L. & WANG, M. 2008. qnr Gene nomenclature.
Antimicrob Agents Chemother, 52, 2297-9.
JACOBY, G. A., WALSH, K. E., MILLS, D. M., WALKER, V. J., OH, H., ROBICSEK, A.
& HOOPER, D. C. 2006. qnrB, another plasmid-mediated gene for quinolone
resistance. Antimicrob Agents Chemother, 50, 1178-82.
JANDA, J. M. A. S. L. A. 2006. The family Enterobacteriaceae: taxonomie considerations.
In J.M. Janda and S. L. Abbott (ed).The enterobacteria, p7-13.
JEAN FRENEY, F. R., ROLAND LECLERCQ ET PHILIPPE RIEGEL 2007. Précis de

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 42
Bactériologie Clinique.
LAVILLA S, G.-L. J., SABATÉ M, GARCÍA-FERNÁNDEZ A, LARROSA MN,
BARTOLOMÉ RM. 2008. Prevalence of qnr genes among extended spectrum β-
lactamase-producing enterobacterial isolates in Barcelona. Spain J Antimicrob
Chemother 61, 291-5...
MAMMERI. H., V. D. L., M., POIREL, L. 2005. Emergence of plasmid-mediated quinolone
resistance in Escherichia coli in Europe
MARTEL, J. L. & CHASLUS-DANCLA, E. 2001. [Antibiotic use in animal breeding]. Rev
Prat, 51, 9-12.
MARTINEZ-MARTINEZ, L., PASCUAL, A. & JACOBY, G. A. 1998. Quinolone resistance
from a transferable plasmid. Lancet, 351, 797-9.
N, K. 2010. Diagn Microbiol Infect Dis.
NORDMANN, P. & POIREL, L. 2005. Emergence of plasmid-mediated resistance to
quinolones in Enterobacteriaceae. J Antimicrob Chemother, 56, 463-9.
NORDMANN P, M. H. 2007. Résistance plasmidique aux quinolones. Antibiotiques, 9, 246-
53.
OETHINGER, M., KERN, W. V., JELLEN-RITTER, A. S., MCMURRY, L. M. & LEVY,
S. B. 2000. Ineffectiveness of topoisomerase mutations in mediating clinically
significant fluoroquinolone resistance in Escherichia coli in the absence of the AcrAB
efflux pump. Antimicrob Agents Chemother, 44, 10-3.
OKTEM, I. M., GULAY, Z., BICMEN, M., GUR, D. & GROUP, H. P. S. 2008. qnrA
prevalence in extended-spectrum beta-lactamase-positive Enterobacteriaceae isolates
from Turkey. Jpn J Infect Dis, 61, 13-7.
P.BIDET, E. B. 2007. Bacilles à Gram négatif aéro-anaérobies, Enterobacteriaceae (à
l’exception de Yersinia).
PARK, C. H., ROBICSEK, A., JACOBY, G. A., SAHM, D. & HOOPER, D. C. 2006.
Prevalence in the United States of aac(6')-Ib-cr encoding a ciprofloxacin-modifying
enzyme. Antimicrob Agents Chemother, 50, 3953-5.
PIDDOCK, L. J. 1999. Mechanisms of fluoroquinolone resistance: an update 1994-1998.
Drugs, 58 Suppl 2, 11-8.
PILLY, E. 2008. Maladies Infectieuses et Tropicales. Page 340.
PITOUT, J. D., THOMSON, K. S., HANSON, N. D., EHRHARDT, A. F., MOLAND, E. S.
& SANDERS, C. C. 1998. Beta-Lactamases responsible for resistance to expanded-

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 43
spectrum cephalosporins in Klebsiella pneumoniae, Escherichia coli, and Proteus
mirabilis isolates recovered in South Africa. Antimicrob Agents Chemother, 42, 1350-
4.
PLESIAT, P. 2012. Biochimie de la résistance. ANTIBIOGRAMME, 2, 17-35.
PONS, M. J., MOSQUITO, S., GOMES, C., DEL VALLE, L. J., OCHOA, T. J. & RUIZ, J.
2014. Analysis of quinolone-resistance in commensal and diarrheagenic Escherichia
coli isolates from infants in Lima, Peru. Trans R Soc Trop Med Hyg, 108, 22-8.
RICE, L. B. 2012. GENETIQUE DE LA RESISTANCE. ANTIBIOGRAMME, 3, 25-37.
ROBICSEK, A., JACOBY, G. A. & HOOPER, D. C. 2006. The worldwide emergence of
plasmid-mediated quinolone resistance. Lancet Infect Dis, 6, 629-40.
ROBICSEK, A., STRAHILEVITZ, J., JACOBY, G. A., MACIELAG, M., ABBANAT, D.,
PARK, C. H., BUSH, K. & HOOPER, D. C. 2006. Fluoroquinolone-modifying
enzyme: a new adaptation of a common aminoglycoside acetyltransferase. Nat Med,
12, 83-8.
ROBICSEK, A., STRAHILEVITZ, J., SAHM, D. F., JACOBY, G. A. & HOOPER, D. C.
2006. qnr prevalence in ceftazidime-resistant Enterobacteriaceae isolates from the
United States. Antimicrob Agents Chemother, 50, 2872-4.
RODRIGUEZ-VILLALOBOS, H. S., M.-J 2006. Résistance bactérienne par Béta-lactamases
à spectre étendu: implications pour le réanimateur. Reanimations, 15, 205-213.
SCHWARZ, S. & CHASLUS-DANCLA, E. 2001. Use of antimicrobials in veterinary
medicine and mechanisms of resistance. Vet Res, 32, 201-25.
SKURNIK D, A. A. Antibiothérapie sélectionnante : de la théorie à la pratique. Reanim 2006,
15, 198-204.
TAKAHASHI, H., HAYAKAWA, I. & AKIMOTO, T. 2003. [The history of the
development and changes of quinolone antibacterial agents]. Yakushigaku Zasshi, 38,
161-79.
VORA, S. & AUCKENTHALER, R. 2009. [What is the significance of extended spectrum
betalactamases in clinical practice?]. Rev Med Suisse, 5, 1991-4.
WANG, H., DZINK-FOX, J. L., CHEN, M. & LEVY, S. B. 2001. Genetic characterization
of highly fluoroquinolone-resistant clinical Escherichia coli strains from China: role
of acrR mutations. Antimicrob Agents Chemother, 45, 1515-21.
WANG, M., J. H. TRAN, G. A. JACOBY, Y. ZHANG, F. WANG, AND D. C. HOOPER
2003. Plasmid-mediated quinolone resistance in clinical isolates of Escherichia coli

Mémoire Master II/BioGeMA/2014-2015/Moumouni Aissatou Page 44
from Shanghai, China. . Antimicrob. Agents Chemother. , 47, 2242–2248.
WELCH, R. A., V.BURLAND, G.PLUNKETT, 3RD, P.REDFORD, P.ROESCH,
D.RASKO, E.L.BUCKLES, S. R.LIOU, A.BOUTIN, J.HACKETT, D.STROUD,
G.F.MAYHEW, D.J.ROSE, S.ZHOU, D.C.SCHWARTZ, N.T.PERNA,
H.L.MOBLEY, M.S.DONNENBERG, AND F.R.BLATTNER. 2002. Extensive
mosaic structure revealed by the complete genome sequence of uropathogenic
Escherichia coli. Proc. Natl. Acad. Sci USA 99, 17020-17024.
WOERTHER, P. L., ANGEBAULT, C., JACQUIER, H., HUGEDE, H. C., JANSSENS, A.
C., SAYADI, S., EL MNIAI, A., ARMAND-LEFEVRE, L., RUPPE, E., BARBIER,
F., RASKINE, L., PAGE, A. L., DE REKENEIRE, N. & ANDREMONT, A. 2011.
Massive increase, spread, and exchange of extended spectrum beta-lactamase-
encoding genes among intestinal Enterobacteriaceae in hospitalized children with
severe acute malnutrition in Niger. Clin Infect Dis, 53, 677-85.
Internet
- http://www.denniskunkel.com/default.asp
- www.nirgal.net/ori_intro.html
- Vulgaris Médical. La connaissance médicale dans un langage accessible à tous,
http://www.vulgaris-medical.com/encyclopedie-medicale/antibiotique-generalites





























