Embed Size (px)
Citation preview

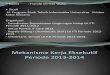
Mekanisme kerja RNAi
Studi awal menunjukkan bahwa siRNA merupakan dupleks 21-26 nukleotida RNA dengan 2-
nukleotida 3’ yang menggantung dan 5’ fosfat dan 3’ hidroksi sebagai terminal. 21-22 nt terlibat dalam
degradasi mRNA dan memiliki ukuran yang lebih panjang, 24-26 nt mengarahkan metilasi DNA dan
penghentian sistemik. Sebuah komponen RISC, domain PAZ dari Argonaute, memfasilitasi pengenalan
siRNA dengan untai tunggal 3’ yang menggantung dengan bantuan enzim Dicer. Telah dikemukakan
bahwa dupleks bergabung di prekursor RISC dan kemudian siRNA yang melepaskan ATP-dependent
mengubah prekursor RISC menjadi RISC aktif. Rasio RISC mengandung untai antisense atau sense dari
RNA yang ditentukan dengan kestabilan termodinamika dari pasangan basa 5’ terminal dari dupleks
siRNA. Basa-basa di dekat ujung 5’ menyumbangkan energi pada pelekatan RNA, sedangkan pasangan
basa yang dibentuk oleh pusat dan daerah 3’ dari siRNA menyediakan geometri berbentuk spiral yang
dibutuhkan untuk katalisis. Domain PIWI pada Argonaute di RISC mempunyai kemiripan dengan
ribonuklease H dengan sebuah motif aspartat-aspartat-glutamat yang dilindungi. Fosfat di antara
nukleotida 11 dan 12 dari ujung 5’ mRNA jatuh mendekati pusat pembelahan RISC yang aktif.
miRNA yang matang merupakan endogen 22 nt RNA yang penting untuk mengarahkan mRNA
dalam pembelahan atau represi translasi pada hewan dan tumbuhan. Molekul RNA yang pendek ini
dihasilkan di sitoplasma Rnase III Dicer dari pre-miRNA yang berbentuk jepit rambut yang diproses oleh
nuklir Rnase III Drosha. Residu 2-8 dari miRNA yang pertama berpasangan tepat dengan elemen daerah
3’ taktertranslasi (UTR) dari RNA target. Endogen dari si RNA dan miRNA mempunyai kemiripan
sehingga dua kelas dari RNA ini tidak dapat dibedakan dari komposisi kimia atau mekanisme kerjanya.
Pada mamalia, siRNA dapat berfungsi seperti miRNA melalui represi ekspresi mRNA target dengan
komplementer sebagian sampai kelipatan dari 3’ UTR. Daerah 5’ dari siRNA dan miRNA semuanya
memainkan peran analogi dalam pengenalan target dan penggabungan RISC ke target RNA. Akan tetapi,
perbedaan asal keduanya, perlindungan evolusioner, dan tipe gen yang dihentikan oleh keduanya telah
diuraikan dengan jelas.
Kemampuan RNA dalam genomic secara fungsional telah ditunjukkan. Teknologi RNAi telah
menjadi pilihan untuk pendekatan pada identifikasi target dan validasi serta pencegahan dan terapi
penyakit. Sebagai perbandingan dengan terapi yang berdasarkan basa RNA lainnya, RNAi mempunyai
banyak keuntungan. Teknologi menjadi jawaban dan memberikan pengaruh besar pada terapi kanker.