Embed Size (px)
Citation preview

EV-1
Meiofauna affect the macrobenthic biodiversity–ecosystem functioning relationship
Adeline Piot, Christian Nozais and Philippe Archambault
A. Piot ([email protected]) and P. Archambault, Lab. d’écologie benthique, Inst. des sciences de la mer de Rimouski, Univ. du Québec à Rimouski, Rimouski, QC, G5L 3A1, Canada. – C. Nozais, Dépt de biologie, Univ. du Québec à Rimouski, Rimouski, QC, G5L 3A1, Canada.
The biodiversity–ecosystem functioning (B–EF) relationship has become of main interest in the last few decades, mostly because of the worldwide increase in species extinctions. However, most of these studies only consider species within single size-class or trophic level, thereby most likely underestimating ecosystem complexity. To reach more realistic scenarios, we assessed the role of meiofauna (lower size-class level) on the relationship between macrofaunal biodiversity and multiple benthic ecosystem properties. Experiments took place under controlled conditions using three macrofaunal species (Alitta (formerly Nereis) virens, Macoma balthica and Mya arenaria). A total of eight combinations of zero to three functionally different macrofaunal species were maintained in microcosms for 34 days in either the presence or absence of a different size-class grouping (a meiofaunal mixture). The organic matter content and bacterial abundance in sediments and the oxygen and nutrient (NH4
, NOx2, PO4
32) fluxes across the sediment-water interface were measured and used as proxies of ecosystem properties. Overall, macrofaunal species richness did not modify any of the measured properties; however, we observed changes associated to the presence of A. virens and M. balthica. This study also revealed a strong impact of the presence of meiofauna on ecosystem properties. They changed interactions between macrofaunal species, which led to modifications in the ecosystem properties. Thus, even if this size-class group has been poorly considered in previous studies, this experiment suggests that future studies should consider the meiofauna with greater attention, particularly in the context of B–EF.
Understanding the relationship between biodiversity and ecosystem functioning (hereafter, B–EF) has become one of the major challenges in ecology during the past few decades (Loreau et al. 2002, Naeem et al. 2009, Isbell et al. 2011, Solan et al. 2012). Indeed, human activities have led to an increase in worldwide species extinction rates and researchers have tried to understand how these biodiversity changes can affect ecosystem properties (Naeem et al. 2009). These ecosystem properties, encompassed in the concept of ‘ecosystem functioning’, have been defined by Hooper et al. (2005) as “both size of compartments (e.g. pools of materials such as carbon and organic matter) and rates of processes (e.g. fluxes of materials and energy among compartments)”.
In marine benthic ecosystems, several studies already assessed the role of biodiversity (both species composition and species interactions) on multiple processes in sediments (Waldbusser et al. 2004, Mermillod-Blondin et al. 2005, Ieno et al. 2006, Norling et al. 2007, Michaud et al. 2009). Ieno et al. (2006) showed that an increase in benthic species richness enhanced nutrient regeneration, but that each species seemed to contribute differently to nutrient regenera-tion processes depending on their bioturbation activities. In other studies, the manipulation of marine invertebrate species richness and composition led to the conclusion that
differences between measured functions were only attributed to the species-specific traits (Norling et al. 2007, Karlson et al. 2010). In their study on tidepools, O’Connor and Crowe (2005) even concluded that different species played idiosyncratic roles, which explains why no relationship between species richness and ecosystem functioning was found. These studies represent a first step in better under-standing the relationship between species richness and ecosystem functioning. However, all of these previous works only considered the influence of macrofauna on eco-system properties, without assessing the contribution of other size-class organisms to ecosystem processes.
Most B–EF studies focus only on interactions within single size-class or trophic level species (e.g. macrofauna) (horizontal biodiversity), but their observations can diffi-cultly explain functioning in real communities since they are not considering all relationships between species (Thébault and Loreau 2006, Gamfeldt et al. 2008). The integration of multi-trophic interactions (vertical biodiversity) in community studies generates non-linear models which are much more complex. The loss of one species can lead to trophic modifications (e.g. cascading extinctions or effects) which may affect ecosystem functioning (Thébault et al. 2007). Moreover, non-trophic interactions (e.g. facilitation,
Oikos 000: 001–011, 2013 doi: 10.1111/j.1600-0706.2013.00631.x
© 2013 The Authors. Oikos © 2013 Nordic Society Oikos Subject Editor: Christopher Swan. Accepted 14 July 2013

EV-2
food competition) also can play an important role in com-munity dynamics (Vasas and Jordán 2006). For example, ecosystem engineers (e.g. corals, mussel beds) can transform their environmental conditions which may affect ecological conditions for other species (Jones et al. 1994, Largaespada et al. 2012). The loss of such species can therefore change the functioning of an entire ecosystem (Coleman and Williams 2002). Thereby, studies on B–EF relationship should con-sider a large panel of species (different size-class/trophic level) to draw more realistic conclusions (Vasas and Jordán 2006, Cardinale et al. 2009).
Meiofauna, which are a pool of organisms with a size between 63 and 500 mm, have often been neglected in benthic studies (Giere 2009), even if they represent a very important component of benthic ecosystems. Indeed, these organisms are very abundant, with high reproduc-tion rates, and are very productive in sediments. Moreover, meiofauna represent an important link between smaller (e.g. bacteria, primary producers) and larger organisms (e.g. macrofauna, demersal fauna) in benthic sediments (Giere 2009), mainly through trophic interactions. Meio-fauna are dependent on environment changes, particularly those generated by macrofauna such as bioturbation and increase in dissolved oxygen in sediments (Tita et al. 2000). However, meiofauna can also modify the environ-ment and stimulate organic matter degradation, mainly because of: 1) a stimulation of the bacterial compartment (Giere 2009), 2) the production of mucus (Giere 2009) and 3) the bioturbation of sediment (biogeochemistry modifications and penetration of oxygen within deeper sediment layers) (Aller and Aller 1992). Therefore, meio-fauna could indirectly affect macrofauna present in the system, and thereby, affect ecosystem functioning.
The Macoma balthica intertidal community from the St Lawrence estuary presents great advantages when study-ing benthic communities in the context of B-EF research. This community exhibits low species diversity, reducing system complexity, with mainly three macrofaunal species (Alitta (formerly Nereis) virens, M. balthica and Mya arenaria) and a meiofaunal community dominated by nematodes (Tita et al. 2002). Moreover, the main charac-teristics and the role played by each of the macrofaunal spe-cies on multiple ecosystem properties are already well known (Mermillod-Blondin et al. 2003, Michaud et al. 2005, 2006, 2009) and contribute to classifying them as 3 functionally different species (Table 1) following their bioturbation and nutrition mode. The polychaete A. virens is an omnivorous gallery-diffuser (Nielsen et al. 1995, Piot et al. 2008) and mainly modifies the geochemical conditions of the sediment, mostly by increasing dissolved oxygen concentrations in sediments (Michaud, et al. 2005, 2006). The mollusc M. balthica is a biodiffuser and a
deposit or suspension-feeder depending on environmental conditions. It highly interacts with the surface sediment through its feeding mode (Hummel 1985). Finally, the biodiffuser M. arenaria affects mainly the overlying water due to its suspension feeding mode (Forster and Zettler 2004), but it can also modify sediment characteristics by changing biogeochemical conditions due to its presence in the denitrification zone (Michaud et al. 2006). Contrarily to macrofauna, meiofauna, which constitute a different size-class level (63 mm size 500 mm), have been less studied in the M. balthica community. These organisms are mostly epigrowth or deposit-feeders (Tita et al. 2002). Meiofaunal organisms are known to modify their environment (e.g. bacterial and benthic diatom abun-dances) (Coull 1999) and to be generally very dependent on macrofaunal species which feed on them and modify environmental conditions (Braeckman et al. 2011).
The general objective of the present study is to assess the role of meiofauna on benthic ecosystem functioning through the example of the M. balthica community. More specifi-cally, objectives of this study were: 1) to see the impact of meiofauna on multiple ecosystem properties (nutrient and oxygen fluxes, bacterial abundance and organic matter content), and 2) to assess how meiofauna can affect the mac-rofaunal biodiversity–ecosystem functioning relationship. To reach these objectives, experiments were run under con-trolled conditions, using microcosms. It was hypothesized that the presence of meiofauna: 1) induces a modification in sediment properties such as bacterial abundances and biogeochemical properties, 2) leads to changes in processes induced by the different macrofaunal species. The results of this study should enable a better understanding of the functioning of the whole benthic system in the M. balthica community. Moreover, the emphasis on meiofauna should be extremely relevant, as this component of infaunal com-munities has been little studied in the literature, particularly in the context of B–EF.
Methods
Sampling site and organisms
All organisms and sediments were randomly collected along the Pointe-au-Père intertidal area (48°31′N, 68°28′W), located on the south shore of the St Lawrence maritime estu-ary. About 1.2 m3 of sediments sampled with a grab (to a depth of 10 cm) were sieved on a 1 mm mesh (to exclude the largest particles and macrofauna) and frozen for one month (220°C) to exclude most of the meiofauna (Quaresma et al. 2004), by significantly decreasing the abundance of nematodes (Table 2, meiofauna effect: F(4,32) 4.23,
Table 1. Traits of the functionally different species used in this experiment. Burrowing depth and size where those measured in microcosms, feeding and bioturbation modes were extracted from literature.
Burrowingdepth (cm)
Size (length, cm) Feeding mode Bioturbation mode
A. virens max. 15 6–7 omnivorous (Nielsen et al. 1995) gallery-diffuser (Michaud, et al. 2006)M. balthica max. 3 0.5–1 mainly deposit-feeder (Ólafsson 1986) biodiffuser (Michaud et al. 2006)M. arenaria max. 6 2–3 suspension-feeder (Forster and Zettler 2004) biodiffuser (Michaud et al. 2006)
EV-3
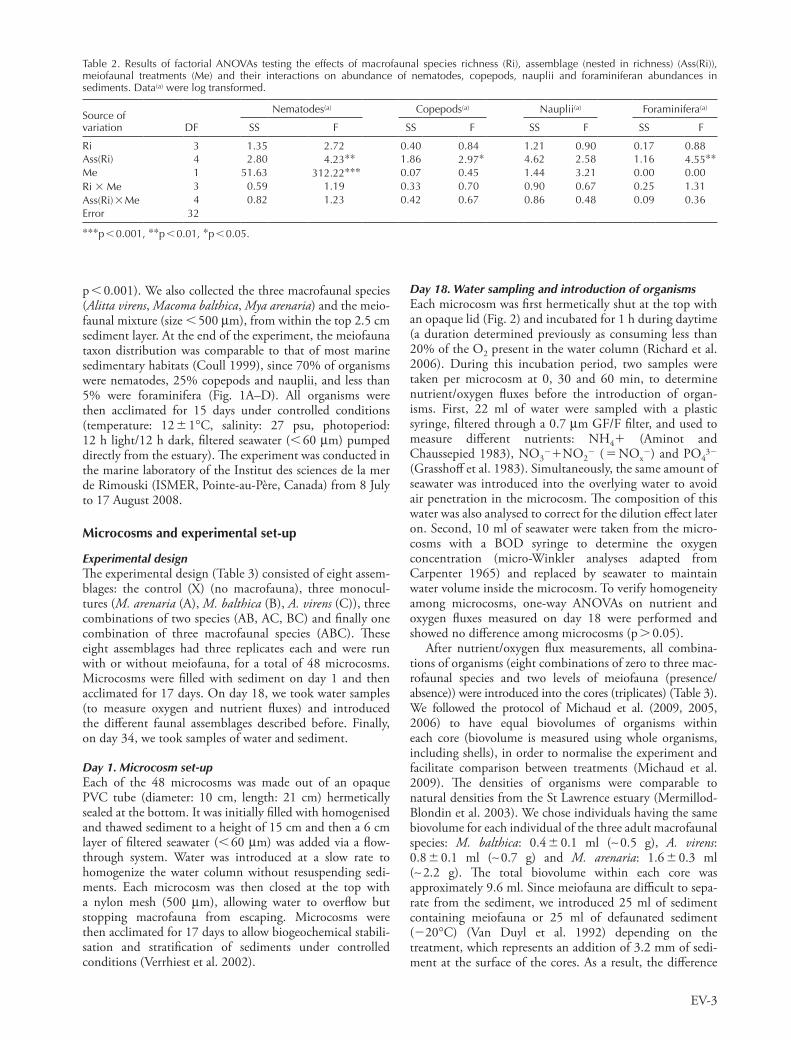
p 0.001). We also collected the three macrofaunal species (Alitta virens, Macoma balthica, Mya arenaria) and the meio-faunal mixture (size 500 mm), from within the top 2.5 cm sediment layer. At the end of the experiment, the meiofauna taxon distribution was comparable to that of most marine sedimentary habitats (Coull 1999), since 70% of organisms were nematodes, 25% copepods and nauplii, and less than 5% were foraminifera (Fig. 1A–D). All organisms were then acclimated for 15 days under controlled conditions (temperature: 12 1°C, salinity: 27 psu, photoperiod: 12 h light/12 h dark, filtered seawater ( 60 mm) pumped directly from the estuary). The experiment was conducted in the marine laboratory of the Institut des sciences de la mer de Rimouski (ISMER, Pointe-au-Père, Canada) from 8 July to 17 August 2008.
Microcosms and experimental set-up
Experimental designThe experimental design (Table 3) consisted of eight assem-blages: the control (X) (no macrofauna), three monocul-tures (M. arenaria (A), M. balthica (B), A. virens (C)), three combinations of two species (AB, AC, BC) and finally one combination of three macrofaunal species (ABC). These eight assemblages had three replicates each and were run with or without meiofauna, for a total of 48 microcosms. Microcosms were filled with sediment on day 1 and then acclimated for 17 days. On day 18, we took water samples (to measure oxygen and nutrient fluxes) and introduced the different faunal assemblages described before. Finally, on day 34, we took samples of water and sediment.
Day 1. Microcosm set-upEach of the 48 microcosms was made out of an opaque PVC tube (diameter: 10 cm, length: 21 cm) hermetically sealed at the bottom. It was initially filled with homogenised and thawed sediment to a height of 15 cm and then a 6 cm layer of filtered seawater ( 60 mm) was added via a flow-through system. Water was introduced at a slow rate to homogenize the water column without resuspending sedi-ments. Each microcosm was then closed at the top with a nylon mesh (500 mm), allowing water to overflow but stopping macrofauna from escaping. Microcosms were then acclimated for 17 days to allow biogeochemical stabili-sation and stratification of sediments under controlled conditions (Verrhiest et al. 2002).
Day 18. Water sampling and introduction of organismsEach microcosm was first hermetically shut at the top with an opaque lid (Fig. 2) and incubated for 1 h during daytime (a duration determined previously as consuming less than 20% of the O2 present in the water column (Richard et al. 2006). During this incubation period, two samples were taken per microcosm at 0, 30 and 60 min, to determine nutrient/oxygen fluxes before the introduction of organ-isms. First, 22 ml of water were sampled with a plastic syringe, filtered through a 0.7 mm GF/F filter, and used to measure different nutrients: NH4 (Aminot and Chaussepied 1983), NO3
2NO22 ( NOx
2) and PO432
(Grasshoff et al. 1983). Simultaneously, the same amount of seawater was introduced into the overlying water to avoid air penetration in the microcosm. The composition of this water was also analysed to correct for the dilution effect later on. Second, 10 ml of seawater were taken from the micro-cosms with a BOD syringe to determine the oxygen concentration (micro-Winkler analyses adapted from Carpenter 1965) and replaced by seawater to maintain water volume inside the microcosm. To verify homogeneity among microcosms, one-way ANOVAs on nutrient and oxygen fluxes measured on day 18 were performed and showed no difference among microcosms (p 0.05).
After nutrient/oxygen flux measurements, all combina-tions of organisms (eight combinations of zero to three mac-rofaunal species and two levels of meiofauna (presence/absence)) were introduced into the cores (triplicates) (Table 3). We followed the protocol of Michaud et al. (2009, 2005, 2006) to have equal biovolumes of organisms within each core (biovolume is measured using whole organisms, including shells), in order to normalise the experiment and facilitate comparison between treatments (Michaud et al. 2009). The densities of organisms were comparable to natural densities from the St Lawrence estuary (Mermillod-Blondin et al. 2003). We chose individuals having the same biovolume for each individual of the three adult macrofaunal species: M. balthica: 0.4 0.1 ml (~ 0.5 g), A. virens: 0.8 0.1 ml (~ 0.7 g) and M. arenaria: 1.6 0.3 ml (~ 2.2 g). The total biovolume within each core was approximately 9.6 ml. Since meiofauna are difficult to sepa-rate from the sediment, we introduced 25 ml of sediment containing meiofauna or 25 ml of defaunated sediment (220°C) (Van Duyl et al. 1992) depending on the treatment, which represents an addition of 3.2 mm of sedi-ment at the surface of the cores. As a result, the difference
Table 2. Results of factorial ANOVAs testing the effects of macrofaunal species richness (Ri), assemblage (nested in richness) (Ass(Ri)), meiofaunal treatments (Me) and their interactions on abundance of nematodes, copepods, nauplii and foraminiferan abundances in sediments. Data(a) were log transformed.
Source of variation
Nematodes(a) Copepods(a) Nauplii(a) Foraminifera(a)
DF SS F SS F SS F SS F
Ri 3 1.35 2.72 0.40 0.84 1.21 0.90 0.17 0.88Ass(Ri) 4 2.80 4.23** 1.86 2.97* 4.62 2.58 1.16 4.55**Me 1 51.63 312.22*** 0.07 0.45 1.44 3.21 0.00 0.00Ri Me 3 0.59 1.19 0.33 0.70 0.90 0.67 0.25 1.31Ass(Ri) Me 4 0.82 1.23 0.42 0.67 0.86 0.48 0.09 0.36Error 32
***p 0.001, **p 0.01, *p 0.05.
EV-4
Figure 1. Mean ( SE) abundances of each taxon of meiofauna: (A) nematodes, (B) foraminifera, (C) nauplii and (D) copepods, and sediment content of (E) organic matter and bacterial abundances in (F) surface (0–0.5 cm), (G) subsurface (0.5–5 cm), and (H) deep (5–12 cm) sediments among assemblages (X control, A Mya arenaria, B Macoma balthica, C Alitta virens). Significant Tukey post hoc results (p 0.05) among groups (Ass(Ri) or Ass(Ri) Me) are indicated by letters. Different letters in Fig. 1A, B, D and H indicate difference among Ass(Ri), whereas letters in Fig. 1F present differences among Ass(Ri) Me interactions. Finally, ‘*Meiofauna’ indicates a significant effect of the presence of the meiofaunal mixture in Fig. 1A, F.
between cores was not due to the introduction of sediment (including labile OM) but to the occurrence or not of meio-fauna. After the introduction of fauna, the cores were recon-nected to the seawater flow-through system for 16 days.
Day 34. Water and sediment samplingAt the end of the experiment, nutrients and oxygen measure-ments (same protocol as before) were done at time 0, 30 and 60 min to measure fluxes with the presence of

EV-5
first one. Samples were then analysed by flow cytometry (using SYBR Green for staining bacterial cells) following the method of Belzile et al. (2008) to provide bacterial abundances per gram of wet weight sediment.
Statistical analyses
Variation in oxygen and nutrient fluxes, OM content, and bacterial and meiofaunal abundances on day 34 were anal-ysed using partly nested ANOVAs based on type 1 (sequen-tial) sums of squares (Perkins et al. 2011) including the effect of species richness (four levels), assemblage nested in richness (eight levels), meiofauna (two levels) and interac-tions. Since type 1 ANOVA can be sensitive to the order that factors are fitted, analyses were run with different ordering of factors to ensure a robust interpretation of results (Perkins et al. 2011). Post hoc power analyses were also performed to support our findings (Quinn and Keough 2002). By convention, a was left at 0.05 and power set to 0.80. Effect sizes were smaller than 0.3 for all performed ANOVAs, which is considered as a small/medium effect size (Quinn and Keough 2002). Homoge-neity of variances was determined visually on the residuals as recommended by Quinn and Keough (2002), and nor-mality was tested using Shapiro–Wilk’s test. Data were transformed to satisfy both assumptions when necessary (details given on Tables where appropriate). Finally, we used a Tukey post hoc test when a source of variation was significant in order to identify differences between tested groups (Day and Quinn 1989). Even if ANOVA detected a significant difference between tested groups for O2 and NOx
2 fluxes and copepod abundance, Tukey post hoc test did not detect such significant differences. Underwood (1997) argued that such post hoc tests focus too much on type 1 error rates at the expense of power consider-ation. The ANOVA is the most powerful analysis, as com-pared to Tukey post hoc test, and therefore, we considered that at least the lowest and highest mean were significantly different (Quinn and Keough 2002).
To test the effect of additivity between macrofaunal species on oxygen and nutrient fluxes, we calculated pre-dicted performance from observed values issued from mon-oculture of each species. For example, predicted performance of AB was (A B)/2 (Michaud et al. 2009). Both values (predicted vs observed) were then compared using the Snedecor–Fisher test. Data were transformed to satisfy the assumption of normality when necessary (details given when appropriate).
Results
Macrofaunal biodiversity and measured properties
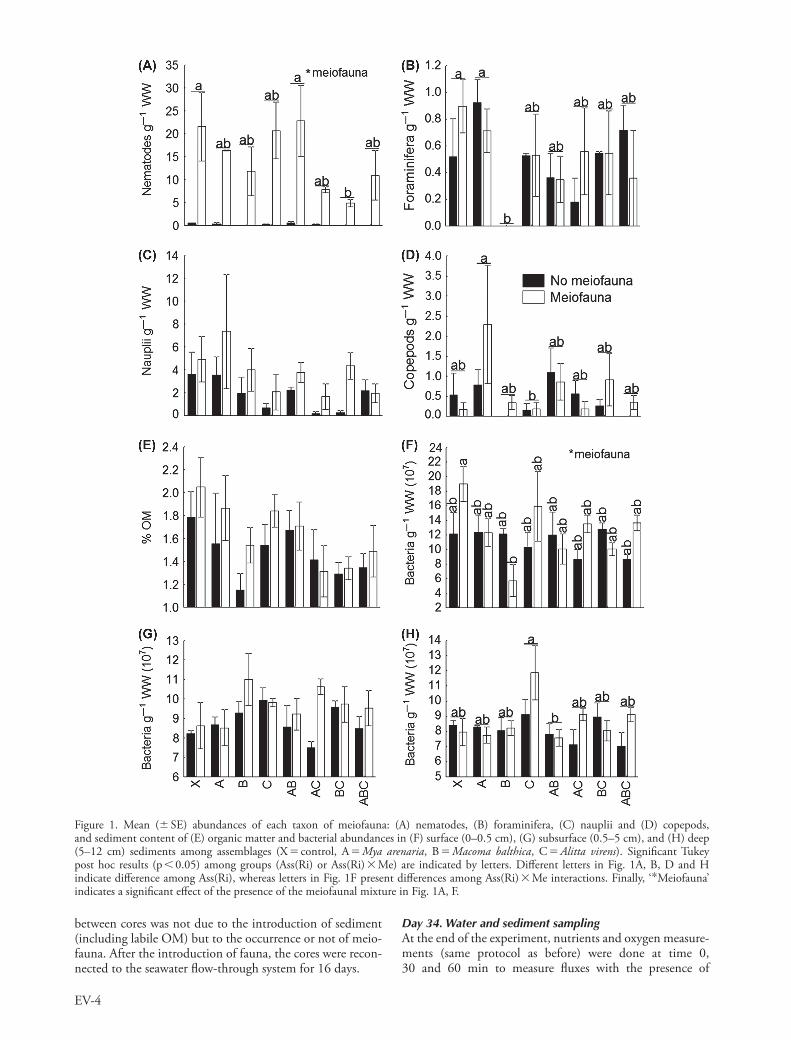
Measured O2 and nutrient fluxes, OM content and bacterial abundances were not affected by macrofaunal species rich-ness (Table 4). However, we observed that O2 flux at the water-sediment interface, ranging from 2281 to 213 mmol h21 m22 (Fig. 3A), and deep sediment bacterial abundances (Fig. 1H, Table 4) were significantly modified by assem-blages, mainly because of the occurrence of Alitta virens alone.
different combinations of organisms. Sediments from the 0–0.5 cm layer were then sampled with a corer made of a ~ 1 cm diameter cut-off syringe for different analyses: OM con-tent by loss upon ignition at 500°C for 2 h (Lyons et al. 2002) and meiofaunal composition and abundances after fixation by 4% formaldehyde for the main taxa (nematodes, foraminifera, nauplii and copepods) under a binocular microscope (Higgins and Thiel 1988). We also counted abundance of macrofaunal organisms and observed that all organisms were alive, and wet mass did not differ from the mass before the introduction into microcosms. Finally, abun-dances of extractible bacteria from 0–0.5, 0.5–5 cm and 5–12 cm sediment layers were analysed by flow cytometry. One ml of sediment was fixed with 4 ml of a solution of fil-tered (0.2 mm) seawater with 2% formaldehyde. We then used an extraction protocol to separate bacteria from the sediment. First, tubes were filled to 10 ml with filtered sea-water (0.2 mm) and we added 500 ml of 100 mM sodium pyrophosphate. Tubes were then submitted three times to vortex (1 min) and sonication (10 min), and were finally centrifuged during 5 min (3000 rpm, 20°C). The superna-tant liquid was removed and kept in a 50 ml Corning tube. A second extraction was then applied to sediments (same protocol) and the second supernatant was mixed with the
Table 3. Number of each organism and total (theoretical) biovolume per assemblage. X: control, Ma: Mya arenaria (biovol: ~ 1.6 ml each), Mb: Macoma balthica (biovol: ~ 0.4 ml each), Av: Alitta virens (biovol: ~ 0.8 ml each).
Assemblage Abbreviation Ma Mb Av Biovolume (ml)
Control X 0 0 0 XM. arenaria (Ma) A 6 0 0 9.6M. balthica (Mb) B 0 24 0 9.6A. virens (Av) C 0 0 12 9.6Ma Mb AB 3 12 0 9.6Ma Av AC 3 0 6 9.6Mb Av BC 0 12 6 9.6Ma Mb Av ABC 2 8 4 9.6
Figure 2. Schematic of a microcosm closed by a tight lid on top allowing water sampling.
EV-6
Nauplii abundances were not modified by macrofaunal assemblages present in microcosms (Fig. 1C).
Meiofauna and measured properties
The occurrence of the meiofaunal mixture into micro-cosms significantly modified PO4
32 fluxes (Table 4, Fig. 3D) (240 to 50 mmol h21 m22). It also significantly increased or decreased bacterial abundances from surface (depending on assemblages), but not from subsurface and deep sediment (Table 4, Fig. 1F–H). The OM con-tent and O2, NOx
2, NH4 fluxes were not significantly
Macrofaunal biodiversity and meiofauna
There was no effect of macrofaunal richness on meiofau-nal abundances, but the assemblage factor (nested in rich-ness) significantly affected the abundance of nematodes, copepods and foraminiferans (Table 2). Indeed, nema-todes were significantly less abundant in presence of the ‘Macoma balthica A. virens’ assemblage than in con-trols. Moreover, copepod abundances increased in presence of Mya arenaria in monoculture (Fig. 1D), whereas the monoculture of M. balthica significantly decreased the abundance of foraminiferans (Fig. 1B).
Figure 3. Mean ( SE) fluxes of (A) O2, (B) NH4, (C) NOx
2 and (D) PO432 across the sediment-water interface for eight assemblages
(X control, A Mya arenaria, B Macoma balthica, C Alitta virens). Significant Tukey post hoc results (p 0.05) among groups (Ass(Ri) or Ass(Ri) Me) are indicated by different letters. Different letters in Fig. 3A indicate difference among Ass(Ri), whereas letters in Fig. 3C present differences among Ass(Ri) Me interactions. Finally, ‘*Meiofauna’ indicates a significant effect of the addition of the meiofaunal mixture in Fig. 3D.
Table 4. Results of factorial ANOVAs testing the effects of macrofaunal species richness (Ri), assemblage (nested in richness) (Ass(Ri)), meiofaunal treatments (Me) and their interactions for O2, NH4
, NOx2 and PO4
32 fluxes across the water sediment interface, organic matter (OM) content and bacterial abundances from surface (0–0.5 cm), subsurface (0.5–5 cm) and deep (5–12 cm) sediments. Data were (a) square and (b) square root transformed.
Source of variation
O2(a) NH4
(b) NOx2 PO4
32 OM Bact.(surf.) Bact.(subsurf.) Bact.(deep)
DF SS ( 108) F SS F SS ( 103) F SS ( 102) F SS F SS ( 1015) F SS ( 1015) F SS ( 1015) F
Ri 3 6.01 0.85 150.77 1.35 4.03 0.91 6.57 2.14 1.09 2.50 9.76 2.28 0.59 1.13 0.67 1.13Ass(Ri) 4 32.28 3.43* 205.69 1.38 6.05 1.02 6.34 1.55 1.00 1.73 5.91 1.04 1.02 1.46 2.49 3.14*Me 1 1.61 0.68 5.04 0.14 0.06 0.04 7.21 7.05* 0.30 2.10 7.55 5.29* 0.59 3.41 0.41 2.04Ri Me 3 0.37 0.05 48.94 0.44 2.03 0.46 8.46 2.76 0.27 0.63 8.35 1.95 0.20 0.37 0.56 0.94Ass(Ri) Me
4 8.20 0.87 137.70 0.92 18.25 3.10* 5.96 1.46 0.03 0.05 16.20 2.83* 1.10 1.58 1.59 2.01
Error 32
***p 0.001, **p 0.01, *p 0.05.
EV-7
of meiofauna. More specifically, oxygen fluxes did not differ between observed and predicted values (Table 5), except for the assemblage ‘M. balthica A. virens’ in presence of meio-fauna. NH4
fluxes were higher than predicted in assem-blages ‘M. arenaria M. balthica’ and ‘M. arenaria A. virens’ without meiofauna. Observed NOx
2 fluxes were significantly different from predicted values in assem-blages ‘M. arenaria M. balthica without meiofauna’ and ‘M. arenaria A. virens’ with and without meiofauna. More-over, observed PO4
32 fluxes were significantly higher than expected in assemblages ‘M. arenaria M. balthica without meiofauna’ and ‘M. balthica A. virens with meiofauna’, whereas they were lower in assemblage ‘M. arenaria M. balthica A. virens without meiofauna’.
Discussion
Macrofaunal biodiversity (species richness and composition) and benthic properties
Most studies attempting to understand the effects of changes in macrofaunal biodiversity on ecosystem functions and services (B–EF) focus on direct measures of properties or ecosystem functions and test how biodiversity modified
affected by the addition of meiofauna (Table 4, Fig. 1E, 3A–C).
Macro–meiofauna interaction and measured properties
Results revealed significant interactions between macrofau-nal assemblages (nested in richness) and meiofauna. Indeed, NOx
2 fluxes (Fig. 3C) (2100 to 140 mmol h21 m22) were modified following the interaction ‘Assemblage(richness) meiofauna’ (Table 4, Fig. 3C). We also observed a significant effect of the interaction ‘Assemblage(richness) meiofauna’ on bacteria from surface sediment (Table 4), since bacterial abundance was significantly higher in control than in the ‘M. balthica’ assemblage in presence of the meiofaunal mixture (Fig. 1F).
Interactions between macrofaunal species vs meiofaunal presence
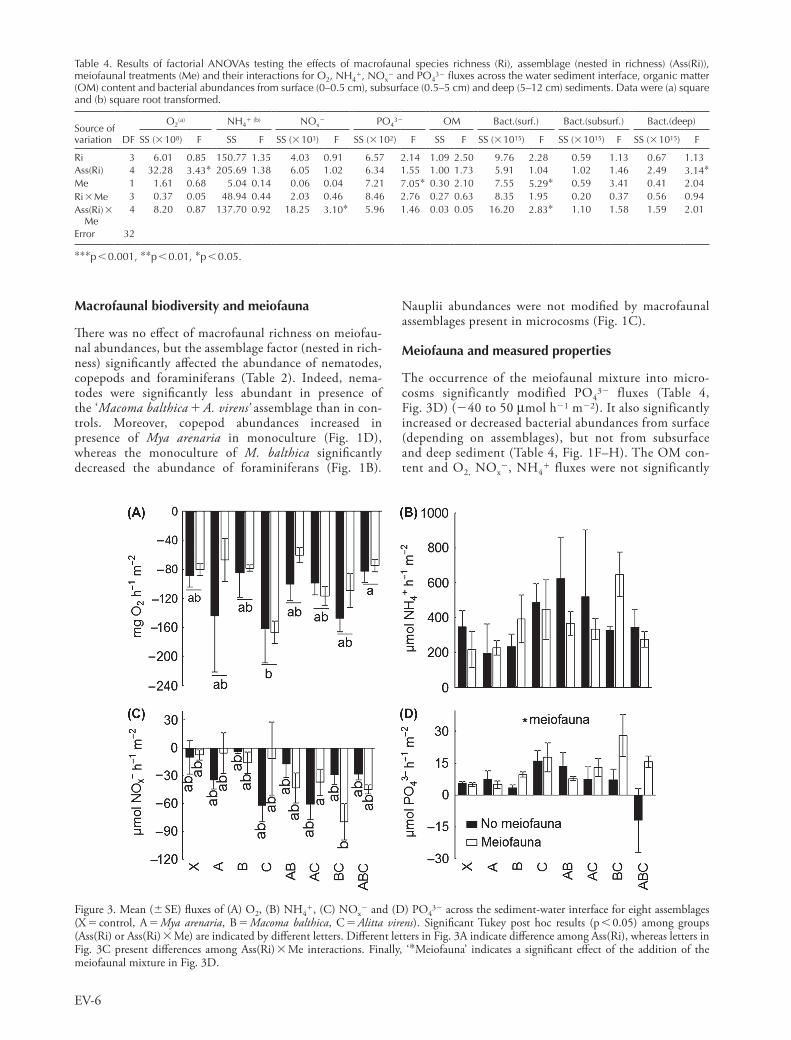
Most oxygen and nutrient fluxes measured in polyculture assemblages could be predicted from the additive model (Fig. 4). However, when the influence of macrofaunal species in polyculture assemblages was not additive, these species interactions were significantly influenced by the occurrence
Figure 4. Predicted and observed mean ( SE) fluxes of (A) O2, (B) NH4, (C) NOx
2 and (D) PO432 across the sediment-water interface
for each combination (AB, AC, BC and ABC) of macrofaunal species (A Mya arenaria, B Macoma balthica, C Alitta virens). ‘‘and ‘2’ indicate respectively the presence or absence of the meiofaunal mixture. *indicates a significant difference (p 0.05) between predicted and observed values. Circles represent outlier values.
EV-8
meiofauna) and foraminiferans abundance in surface sedi-ments. Moreover, M. balthica can keep the sediment surface clean of microalgal mats and bacteria (significant decrease of abundance) by grazing, which probably induced a reduc-tion in the abundance of the major meiofaunal taxa, i.e. nematodes (Ólafsson et al. 2005).
In this study, the biodiffuser M. arenaria did not induce measurable changes in biogeochemical fluxes or other sedi-ment properties. However, we know that M. arenaria is strongly linked to the overlying water column through its suspension feeding mode (Forster and Zettler 2004). Measures of other properties in the water column (e.g. plankton abundance) would be of main interest in future studies. We also suspect the increase of copepod abundance in surface sediment in presence of M. arenaria to be due to this strong link with the overlying water, since copepods can accumulate inside and around siphons (Piot unpubl.). This would constitute a non-trophic interaction between these two groups of species, since M. arenaria facilitates the presence of copepods in surface sediment because of its high water filtration rate (Forster and Zettler 2004).
Meiofauna and measured properties
Marine meiofauna are often neglected in benthic studies and this is more visible in B–EF studies even though they repre-sent an important faunal component in terms of abundance (105 to 106 individuals m22) (Giere 2009). In this study, only nematodes can be considered as an added group because copepods, nauplii and foraminiferans also occurred in treatments without meiofaunal mixture introduction. The addition of meiofauna also probably modified the bacterial compartment at the beginning of the experiment. Indeed, meiofauna were added through the introduction of 25 ml of sediment at the top of sediment core (thickness: 3 mm), whereas cores without meiofauna received 3 mm of defaunated sediment (frozen). The freezing method has already been tested with regard to bacterial characteristics (abundance, composition and activity) (Stocum and Plante 2006). These authors observed that bacterial abundances and activities were recovered after 10 days, whereas compo-sitional recovery took a bit longer. Since we waited 17 days between the introduction of fauna and the measurement of properties (nutrient and oxygen fluxes, organic matter con-tent), we expect that the bacterial compartment in the top 3 mm layer was no longer affected by the freezing method. For these reasons, we consider that our method of intro-duction of meiofauna induced mainly an addition of nema-todes in cores and did not significantly modify the results due to the modification of the bacterial compartment.
Meiofauna can create biogeochemical modifications in sediments (Aller and Aller 1992). Indeed, we observed an increase in PO4
32 release at the water-sediment interface in treatments where the meiofaunal mixture was introduced. Nematodes are known to have a great effect on solute fluxes and aerobic decomposition reaction rates (Rysgaard et al. 2000), partly because meiofaunal grazing stimulates bacte-rial growth (Gerlach 1978) and also because their presence enhances sediment porosity and oxygenation (Aller and Aller 1992), as observed in deep sediments. The OM is then decomposed by bacteria under aerobic conditions and could
these functions (Loreau et al. 2002, Naeem et al. 2009). Fol-lowing the definition of Hooper et al. (2005), we used oxy-gen and nutrient fluxes (rates of processes), and OM content and bacterial abundance (size of compartment) as proxies of ecosystem functioning. Since all microcosms were homogeneous at the beginning of the experiment, the varia-tions in these properties should directly result from the introduction of different combinations of faunal species. However, we did not observe a direct relationship between the number of macrofaunal species and measured responses at the end of the experiment, but other interesting patterns have been revealed.
As reported by Michaud et al. (2005, 2006), Alitta virens was the species most responsible for the variation in biogeochemical fluxes, especially because it induced a higher O2 consumption from the overlying water to the sediment column. This results from the high respiration rate of A. virens (Nielsen et al. 1995) and the creation of galleries in sediments. This contributes to the oxygenation of deep sediments by bioirrigation and generates a decrease of O2 concentration in the water column (Kristensen 1985, Michaud et al. 2005). Mya arenaria and Macoma balthica are also known to contribute to O2 consumption in the water column, but to a lesser extent (Michaud et al. 2005). The occurrence of A. virens also increased the bacterial abundance in deep sediment, as compared to control and other assemblages. This suggests that the worm activities (bioturbation, bioirrigation, food storage, etc.) strongly modified deep sediment properties, inducing changes to the bacterial compartment.
The occurrence of M. balthica, a deposit or suspension feeder depending on environmental conditions (Ólafsson 1986), can significantly affect surface sediments (Hummel 1985, Ólafsson et al. 2005). In laboratory conditions, M. balthica mainly feeds at the water-sediment interface (Hummel 1985), which is also what we observed visually during this study. This feeding mode likely contributed to the significant decrease in bacterial (in presence of
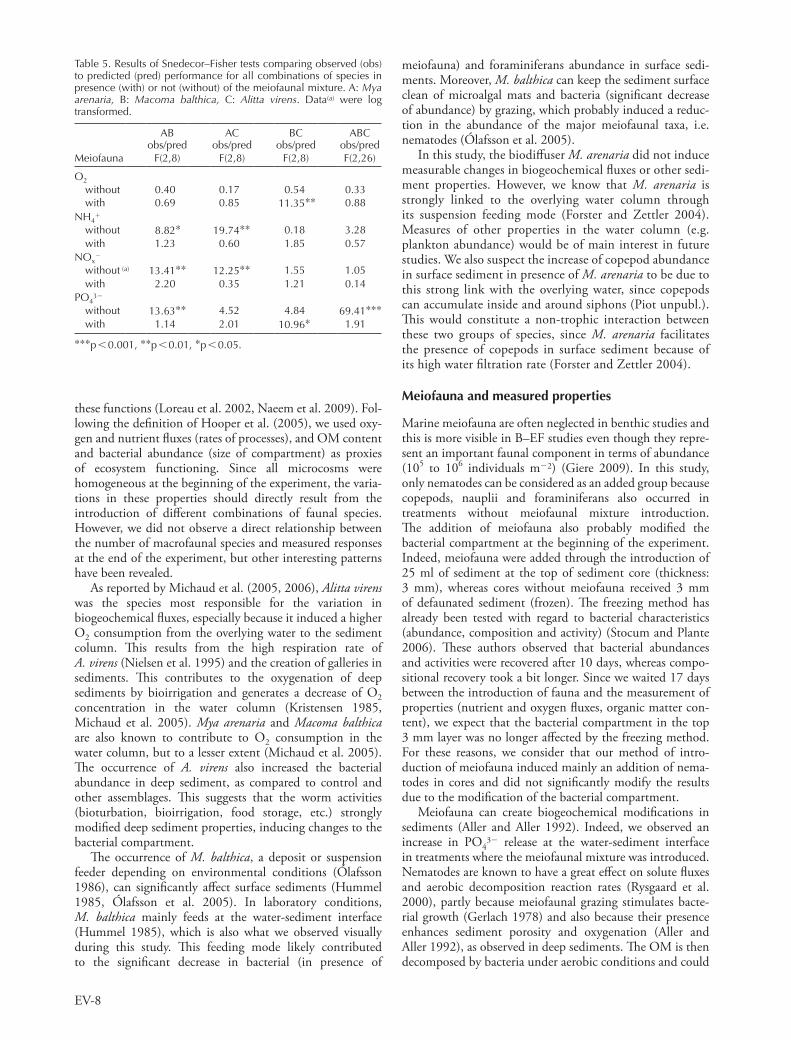
Table 5. Results of Snedecor–Fisher tests comparing observed (obs) to predicted (pred) performance for all combinations of species in presence (with) or not (without) of the meiofaunal mixture. A: Mya arenaria, B: Macoma balthica, C: Alitta virens. Data(a) were log transformed.
AB obs/pred
AC obs/pred
BC obs/pred
ABC obs/pred
Meiofauna F(2,8) F(2,8) F(2,8) F(2,26)
O2
without 0.40 0.17 0.54 0.33with 0.69 0.85 11.35** 0.88
NH4
without 8.82* 19.74** 0.18 3.28with 1.23 0.60 1.85 0.57
NOx2
without (a) 13.41** 12.25** 1.55 1.05with 2.20 0.35 1.21 0.14
PO432
without 13.63** 4.52 4.84 69.41***with 1.14 2.01 10.96* 1.91
***p 0.001, **p 0.01, *p 0.05.

EV-9
Our extraction method which consists of freezing sedi-ments at 220°C for one month (longer than most of previ-ous studies, Tolhurst et al. 2012) was not as efficient as what we had expected, since live meiofauna were found in treatments without meiofauna (Fig. 1). Other methods such as air drying, oven-heating, spraying with formalin and spraying with hydrogen peroxide (Tolhurst et al. 2012) or decantation (Higgins and Thiel 1988) can be used, but also present certain disadvantages. For example, the decantation method (commonly used to extract meio-fauna from sediment for counting, Higgins and Thiel 1988) presents great advantages, since it physically extracts meiofauna from sediment by density differences. However, this method can also extract certain sediment grain sizes and organic matter particles with similar density as the organisms you target to remove. This can induce changes in sediment properties and therefore modify biogeochemi-cal processes and strongly influencing the results. The choice of the meiofaunal extraction method needs to be considered in future studies to efficiently extract meio-fauna, without modifying sediment properties.
Copepods dwelling at the surface of the sediment have the ability and the habit of swimming up to several tens of cm upwards in the water column (Giere 2009). An open water circulation system was used during our experiment and could induce the introduction of copepods in micro-cosms, even if water was filtered through a 60 mm sieve. Because the whole experiment was 34 days long, we did not want to keep organisms without food supply for such a long time, but future studies should consider using a closed system with food addition, preventing meiofauna entrance.
Finally, as already mentioned, sediments used to inocu-late meiofauna in microcoms were sampled in the top 2.5 cm sediment layer. However, meiofaunal species and communities from surface sediment are different from those dwelling in deeper sediment layers (Joint et al. 1982). Even if we observed meiofauna in deeper sediment layers (unpubl.), it is likely that deep sediment layers were not col-onised by same meiofaunal communities than in the nature. Biogeochemical processes could then have been affected by this change. A challenge for future experiments is to find a way to introduce meiofauna from both surface and deep sediment layers, while keeping microcosms comparable (i.e. comparable sediment in all microcosms).
Conclusion
This study underlines the high complexity of the Macoma balthica community. Macrofaunal species richness did not lead to the modification of ecosystem properties, but mac-rofaunal species played an important role in these modifi-cations, as observed previously. However, the addition of fauna of another size-class (i.e. meiofauna) was shown to have a strong (though inconsistent) direct impact on ben-thic properties. Moreover, the occurrence of the meiofau-nal mixture modified the interactions between macrofaunal species, and therefore their impact on ecosystem proper-ties. The often poorly studied meiofaunal compartment represented the most important input in this study. It should therefore strongly be considered in future studies for better understanding the benthic system as a whole.
contribute significantly to a release of PO432 in the water
column (Sundby et al. 1992).The presence of nematodes in microcosms did not gen-
erally lead to a decrease in bacterial abundance, even if they feed on bacteria. This relationship seems rather closely linked to macrofaunal assemblages. Accordingly, many studies have already shown that bacterial abundances are generally not correlated with meiofauna (Montagna et al. 1983, Epstein and Shiaris 1992, Pascal et al. 2008). Van Oevelen et al. (2006) suggested that only 3% of the total bacterial production is grazed by meiofauna and 24% by macrofauna; the primary fate of bacterial pro-duction being mortality (65%). Moreover, non-trophic activities of meiofauna like bioturbation and excretion of products (faeces, mucus, etc.) induce a bacterial pro-ductivity which would not exist without them (Gerlach 1978). In our study, all of this contributes to a significant effect of the addition of the meiofaunal mixture (mainly nematodes) on bacterial abundances.
Effects of meiofauna on the macrofaunal biodiversity – benthic properties relationship
The role of meiofauna on the relationship between macro-faunal biodiversity and benthic properties has been poorly considered before. Nonetheless, our results suggest that the influence of macrofaunal diversity depends on meiofauna, since the interaction between Assemblage(richness) Meiofauna was significant for NOx
2 fluxes and the abun-dance of surface bacteria. These results are very interesting and clearly show that meiofauna must be taken into account in works studying the role of macrofaunal species at the marine water–sediment interface, mainly because meio-fauna can modify the relationship between macrofauna and benthic properties.
In our study, the performance (i.e. oxygen and nutrient fluxes) in polyculture (combinations of macrofaunal species) can mostly be predicted from monocultures. Contrarily to Michaud et al. (2009), the effects of species in polyculture were additive. This means that even if one macrofaunal species was in combination with another species, they were influencing the fluxes the same way, suggesting that non-trophic interactions between species did not modify biogeochemical fluxes. However, certain combinations of macrofaunal species led to weaker fluxes than predicted from the sum of monocultures and this was different following the occurrence of the meiofaunal mix-ture or not. Moreover, the influence of interactions among macrofaunal species on biogeochemical fluxes depended on the occurrence of meiofauna in 50% of cases (Table 5). This suggests that meiofauna could play a very important role when considering non-trophic interactions between macrofaunal species and their influence on ecosystem func-tioning. Therefore, the meiofaunal compartment needs to be taken in deep consideration to really identify its contri-bution to ecosystem functioning.
Improvement for future studies on meiofauna
Meiofauna are generally poorly studied, mainly because they are difficult to use and manipulate (Giere 2009).

EV-10
Hummel, H. 1985. Food intake of Macoma balthica (Mollusca) in relation to seasonal changes in its potential food on a tidal flat in the dutch Wadden Sea. – Neth.J. Sea Res. 19: 52–76.
Ieno, E. N. et al. 2006. How biodiversity affects ecosystem functioning: roles of infaunal species richness, identity and density in the marine benthos. – Mar. Ecol. Progr. Ser. 311: 263–271.
Isbell, F. et al. 2011. High plant diversity is needed to maintain ecosystem services. – Nature 477: 199–202.
Joint, I. R. et al. 1982. Determination of fine-scale vertical distribution of microbes and meiofauna in an intertidal sediment. – Mar. Biol. 72: 157–164.
Jones, C. G. et al. 1994. Organisms as ecosystem engineers. – Oikos 69: 373–386.
Karlson, A. M. L. et al. 2010. Higher diversity of deposit-feeding macrofauna enhances phytodetritus processing. – Ecology 91: 1414–1423.
Kristensen, E. 1985. Oxygen and inorganic nitrogen exchange in a “Nereis virens” (Polychaeta) bioturbated sediment-water system. – J. Coastal Res. 1: 109–116.
Largaespada, C. et al. 2012. Meta-ecosystem engineering: nutrient fluxes reveal intraspecific and interspecific feedbacks in fragmented mussel beds. – Ecology 93: 324–333.
Loreau, M. et al. 2002. Biodiversity and ecosystems functioning: synthesis and perspectives. – Oxford Press.
Lyons, W. B. et al. 2002. Organic carbon fluxes to the ocean from high-standing islands. – Geology 30: 443–446.
Mermillod-Blondin, F. et al. 2003. Assessment of the spatial variability of intertidal benthic communities by axial tomodensitometry: importance of fine-scale heterogeneity. – J. Exp. Mar. Biol. Ecol. 287: 193–208.
Mermillod-Blondin, F. et al. 2005. Biodiversity of benthic invertebrates and organic matter processing in shallow marine sediments: an experimental study. – J. Exp. Mar. Biol. Ecol. 315: 187–209.
Michaud, E. et al. 2005. The functional group approach to bioturbation: the effects of biodiffusers and gallery-diffusers of the Macoma balthica community on sediment oxygen uptake. – J. Exp. Mar. Biol. Ecol. 326: 77–88.
Michaud, E. et al. 2006. The functional group approach to bioturbation: II. The effects of the Macoma balthica community on fluxes of nutrients and dissolved organic carbon across the sediment–water interface. – J. Exp. Mar. Biol. Ecol. 337: 178–189.
Michaud, E. et al. 2009. Spatial interactions in the Macoma balthica community control biogeochemical fluxes at the sediment-water interface and microbial abundances. – J. Mar. Res. 67: 43–70.
Montagna, P. A. et al. 1983. The relationship between abundances of meiofauna and their suspected microbial food (diatoms and bacteria). – Estuarine Coastal Shelf Sci. 17: 381–394.
Naeem, S. et al. 2009. Biodiversity, ecosystem functioning, and human wellbeing: an ecological and economic perspective. – Oxford Press.
Nielsen, A. M. et al. 1995. Feeding, growth and respiration in the polychaetes Nereis diversicolor (facultative filter-feeder) and N. virens (omnivorous)-a comparative study. – Mar. Ecol. Progr. Ser. 125: 149–158.
Norling, K. et al. 2007. Importance of functional biodiversity and species-specific traits of benthic fauna for ecosystem functions in marine sediment. – Mar. Ecol. Progr. Ser. 332: 11–23.
O’Connor, N. and Crowe, T. P. 2005. Biodiversity loss and ecosystem functioning: distinguish between number and identity of species. – Ecology 86: 1783–1796.
Ólafsson, E. B. 1986. Density dependence in suspension-feeding and deposit-feeding populations of the bivalve Macoma balthica: a field experiment. – J. Anim. Ecol. 55: 517–526.
Multifunctional and multitrophic studies such as this one are essential if we are to understand the ecological impacts of biodiversity change.
Acknowledgements – We thank K. Coustaut for his assistance during this study, C. Belzile for helping to analyse bacterial abundances, P. Rioux for nutrient analyses and M. Huot for the revision of the English version. Research was financed by the NSERC Canadian Healthy Oceans Network-a university–government partnership dedicated to biodiversity science for the sustainability of Canada’s three oceans- and by FQRNT network Ressources Aquatiques Québec.
References
Aller, R. C. and Aller, Y. 1992. Meiofauna and solute transport in marine muds. – Limnol. Oceanogr. 37: 1018–1033.
Aminot, A. and Chaussepied, M. 1983. Manuel des analyses chimiques en milieu marin. – Centre national pour l’exploitation des océans.
Belzile, C. et al. 2008. Variations of the abundance and nucleic acid content of heterotrophic bacteria in Beaufort Shelf waters during winter and spring. – J. Mar. Systems 74: 946–956.
Braeckman, U. et al. 2011. Contrasting macrobenthic activities differentially affect nematode density and diversity in a shallow subtidal marine sediment. – Mar. Ecol. Progr. Ser. 422: 179–191.
Cardinale, B. et al. 2009. Towards a food web perspective on biodiversity and ecosystem functioning. – In: Naeem S. et al. (eds), Biodiversity, ecosystem functioning, and human wellbeing: an ecological and economic perspective. Oxford Press, pp. 105–120.
Carpenter, J. H. 1965. The Chesapeake Bay Institute technique for the Winkler dissolved oxygen method. – Limnol. Oceanogr. 10: 141–143.
Coleman, F. C. and Williams, S. L. 2002. Overexploiting marine ecosystem engineers: potential consequences for biodiversity. – Trends Ecol. Evol. 17: 40–44.
Coull, B. C. 1999. Role of meiofauna in estuarine soft-bottom habitats. – Austral Ecol. 24: 327–343.
Day, R. W. and Quinn, G. P. 1989. Comparisons of treatments after an analysis of variance in ecology. – Ecol. Monogr. 59: 433–463.
Epstein, S. S. and Shiaris, M. P. 1992. Rates of microbenthic and meiobenthic bacterivory in a temperate muddy tidal flat community. – Appl. Environ. Microbiol. 58: 2426–2431.
Forster, S. and Zettler, M. L. 2004. The capacity of the filter-feeding bivalve Mya arenaria L. to affect water transport in sandy beds. – Mar. Biol. 144: 1183–1189.
Gamfeldt, L. et al. 2008. Multiple functions increase the importance of biodiversity for overall ecosystem functioning. – Ecology 89: 1223–1231.
Gerlach, S. A. 1978. Food-chain relationships in subtidal silty sand marine sediments and the role of meiofauna in stimu-lating bacterial productivity. – Oecologia 33: 55–69.
Giere, O. 2009. Meiobenthology: the microscopic motile fauna of aquatic sediments, 2nd edn. – Springer.
Grasshoff, K. et al. 1983. Methods of seawater analysis, 2nd edn. – Verlag Chemie.
Higgins, R. P. and Thiel, H. 1988. Introduction to the study of meiofauna. – Smithsonian Inst. Press.
Hooper, D. U. et al. 2005. Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. – Ecol. Monogr. 75: 3–35.

EV-11
Thébault, E. and Loreau, M. 2006. The relationship between biodiversity and ecosystem functioning in food webs. – Ecol. Res. 21: 17–25.
Thébault, E. et al. 2007. Cascading extinctions and ecosystem functioning: contrasting effects of diversity depending on food web structure. – Oikos 116: 163–173.
Tita, G. et al. 2000. Predation and sediment disturbance effects of the intertidal polychaete Nereis virens (Sars) on associated meiofaunal assemblages. – J. Exp. Mar. Biol. Ecol. 243: 261–282.
Tita, G. et al. 2002. Intertidal meiofauna of the St Lawrence estuary (Quebec, Canada): diversity, biomass and feeding structure of nematode assemblages. – J. Mar. Biol. Ass. UK 82: 779–791.
Tolhurst, T. et al. 2012. Technical note: the effects of five different defaunation methods on biogeochemical properties of intertidal sediment. – Biogeosciences 9: 3647–3661.
Underwood, A. J. 1997. Experiments in ecology: their logical design and interpretation using analysis of variance. – Cambridge Univ. Press
Van Duyl, F. C. et al. 1992. The impact of organic matter and macrozoobenthos on bacterial and oxygen variables in marine sediment boxcosms. – Neth. J. Sea Res. 29: 343–355.
Van Oevelen, D. et al. 2006. The fate of bacterial carbon in an intertidal sediment: modeling an in situ isotope tracer experiment. – Limnol. Oceanogr. 51: 1302–1314.
Vasas, V. and Jordán, F. 2006. Topological keystone species in ecological interaction networks: Considering link quality and non-trophic effects. – Ecol. Modell. 196: 365–378.
Verrhiest, G. J. et al. 2002. Chemical and bacterial changes during laboratory conditioning of formulated and natural sediments. – Chemosphere 46: 961–974.
Waldbusser, G. G. et al. 2004. The effects of infaunal biodiversity on biogeochemistry of coastal marine sediments. – Limnol. Oceanogr. 49: 1482–1492.
Ólafsson, E. et al. 2005. The clam Macoma balthica prevents in situ growth of microalgal mats: implications for meiofaunal assemblages. – Mar. Ecol. Progr. Ser. 298: 179–188.
Pascal, P.-Y. et al. 2008. Bacterivory in the common foraminifer Ammonia tepida: isotope tracer experiment and the controlling factors. – J. Exp. Mar. Biol. Ecol. 359: 55–61.
Perkins, D. M. et al. 2011. Environmental warming and biodiversity–ecosystem functioning in freshwater microcosms: partitioning the effects of species identity, richness and metabolism. – Integr. Ecol. – Molecules Ecosyst. 43: 177–209.
Piot, A. et al. 2008. Experimental study on the influence of bioturbation performed by Nephtys caeca (Fabricius) and Nereis virens (Sars) annelidae on the distribution of dino-flagellate cysts in the sediment. – J. Exp. Mar. Biol. Ecol. 359: 92–101.
Quaresma, V. d. S. et al. 2004. The influences of biological activity and consolidation time on laboratory cohesive beds. – J. Sedimentary Res. 74: 184–190.
Quinn, G. P. and Keough, M. J. 2002. Experimental design and data analysis for biologists. – Cambridge Univ Press.
Richard, M. et al. 2006. Influence of suspended mussel lines on the biogeochemical fluxes in adjacent water in the Îles- de-la-Madeleine (Quebec, Canada). – Can. J. Fish. Aquat. Sci. 63: 1198–1213.
Rysgaard, S. et al. 2000. Marine meiofauna, carbon and nitrogen mineralization in sandy and soft sediments of Disko Bay, West Greenland. – Aquat. Microbial Ecol. 21: 59–71.
Solan, M. et al. 2012. Marine biodiversity and ecosystem functioning: frameworks, methodologies and integration. – Oxford Univ. Press.
Stocum, E. T. and Plante, C. J. 2006. The effect of artificial defaunation on bacterial assemblages of intertidal sediments. – J. Exp. Mar. Biol. Ecol. 337: 147–158.
Sundby, B. et al. 1992. The phosphorus cycle in coastal marine sediments. – Limnol. Oceanogr. 37: 1129–1145.





























