Embed Size (px)
Citation preview

Ž .Applied Animal Behaviour Science 65 1999 29–42
Maternal behaviour of domestic sows and crossesbetween domestic sows and wild boar
Maria Gustafsson a,), Per Jensen a, Francien H. de Jonge b,Gudrun Illmann c, Marek Spinka c
a Swedish UniÕersity of Agricultural Sciences, Faculty of Veterinary Medicine, Department of AnimalEnÕironment and Health, Section of Ethology, POB 234, S-532 23 Skara, Sweden
b Department of Ecological Agriculture, Wageningen Agricultural UniÕersity, Haarweg 333, 6709 RZWageningen, Netherlands
c Group of Ethology, Research Institute of Animal Production POB 1, 10401 Prague, UhrineÕes, CzechRepublic
Accepted 16 April 1999
Abstract
Ž .We compared maternal behaviour in domestic sows and wild boar crosses wild-type sows toassess whether pre-parturient behaviour andror the ability to care for offspring has been reducedandror modified through domestication. Seven domestic sows and seven wild-type sows wereused. Nest building behaviour, nursing behaviour during week 1–3 and activity measures wererecorded in an outdoor environment with farrowing huts. There were no differences between thetwo genotypes of sows in the frequency or pattern of nest building behaviour. The number ofnutritive nursings was reduced in the second week post partum for both wild-type and domesticsows. Domestic sows allowed their piglets to massage the udder for a longer time after milkejection than the wild-type sows the first week of observation and tended to do the same thesecond week of observation. Both wild-type and domestic sows tended to terminate more nursingsthe second week post partum compared to the first week and wild-type sows terminated a largerproportion of nursings compared to the domestic sows. During the third week after parturition,domestic sows tended to lie down more and move less than wild-type sows. The frequency ofnose-contacts between the sow and her piglets was higher in the wild-type sows than domesticsows. This study demonstrated large similarities in the maternal behaviour of wild-type anddomestic sows. The few observed differences might indicate that domestic sows have adapted tosome extent to a life in human protection, where it may be more beneficial from a life-timereproductive aspect to invest more in the current litter. The results suggest that the pre-parturient
) Corresponding author. Tel.: q 46-0-511-67227; Fax: q 46-0-511-67204; e-m ail:[email protected]
0168-1591r99r$ - see front matter q 1999 Elsevier Science B.V. All rights reserved.Ž .PII: S0168-1591 99 00048-9

( )M. Gustafsson et al.rApplied Animal BehaÕiour Science 65 1999 29–4230
need of sows and the ability to care for the offspring have not been substantially modified bydomestication. q 1999 Elsevier Science B.V. All rights reserved.
Keywords: Maternal behaviour; Pigs; Domestication; Nursing; Pre-parturient behaviour
1. Introduction
Maternal behaviour of pigs has been studied in wild boar, feral and domestic pigsŽ .Hanson and Karstad, 1959; Gundlach, 1968; Jensen and Recen, 1989; Horrell, 1997 .´
Ž .There seem to be large similarities between different types of sows Jensen, 1986 , butsystematic comparisons of the behaviour of pigs, differing in degree of domestication isimportant in order to understand the biological background of maternal behaviour in thisspecies and to provide a basis for improving animal welfare and production in pigs.
In this study, we address three specific questions in relation to welfare and thematernal behaviour of sows. First, has motivation for nest building before parturitionbeen altered by domestication? Wild, feral and free-range domestic pigs build nests in
Žwhich they farrow Hanson and Karstad, 1959; Gundlach, 1968; Jensen and Recen,´.1989 , and when sows are confined in farrowing crates under commercial conditions,
and thus inhibited from nest building behaviour, there is ample evidence of welfareŽ .problems Jensen et al., 1993; Lawrence et al., 1994 . Confined sows are limited in their
selection of nest-site, unable to gather material for nest building and are forced to farrowin an uncompleted nest, which seems to be associated with both physiological and
Ž .ethological signs of reduced welfare Cronin et al., 1994; Lawrence et al., 1994 .Second, has domestication reduced or modified maternal behaviour in early lactation? Incommercial systems, pig crushing is one of the major causes of neonatal piglet mortalityŽ .Fraser, 1990 . However, crushing of piglets is often the believed cause of death when
Žmalnutrition of piglets is involved English and Smith, 1975; Dyck and Swierstra, 1987;.Fraser, 1990 . Prevention of neonatal mortality thus requires a well functioning maternal
Ž .behaviour in which the sow can provide satisfactory for the piglets Fraser, 1990 , and itis important to understand how domestication may have modified the capacity of sowsfor nursing and caring for piglets. Third, has the general pattern of maternal behaviourchanged with domestication? During domestication, pigs have mainly been selected forrapid growth and meat quality and quantity, with selection on number of weaned pigletsper year but little direct selection on maternal abilities of sows. It is possible thatselection during domestication either intentionally or unintentionally might have influ-enced maternal behaviour. The fact that some domestic pigs fail to provide adequatematernal care can be explained either by the unnatural living conditions not providingnecessary stimuli and opportunity for maternal behaviour or by modified maternalabilities due to domestication. It is also possible that selection of characters of economic
Ž .importance have had a deleterious effect on maternal behaviour Hafez, 1962 , forexample, by selecting sows which mainly allocate energy to growth and increased littersize rather than to milk or maternal care.
In summary, some piglet mortality may be associated with domestication inducedinsufficient maternal care, and some welfare problems of sows may arise from be-

( )M. Gustafsson et al.rApplied Animal BehaÕiour Science 65 1999 29–42 31
havioural needs, which have evolved in wild boars, but still prevail among domesticpigs.
In this experiment, we compared the maternal behaviour of domestic sows with thebehaviour of first generation crosses between domestic sows and wild boar, the ancestor
Ž .of domestic pigs Hemmer, 1990 , in an outdoor environment with farrowing huts. AllŽ .sows were born and raised by the same breed of mothers all domestic sows and kept
throughout life in the same type of environment, and the litters of the sows werestandardised with respect to breeds, weight, etc. Hence, we have controlled for otherthan genetic factors, such as environmental and maternal effects. Our goal was to assesswhether the abilities of sows to care for offspring were reduced or modified during theprocess of domestication.
2. Material and methods
2.1. Animals
This study was carried out at the Wageningen Agricultural University, The Nether-lands. The experimental animals were 14 2-year old primiparous sows, which were born,
Žreared and housed in an outdoor family system with half-open farrowing pens as.previously described in detail by de Jonge et al., 1996 . Half of the sows were bred from
Ž .a Yorkshire father and Yorkshire X Dutch landrace mother ‘domestic’, ns7 . TheŽother half were bred from similar mothers, but with a wild boar as father ‘wild-type’,
.ns7 . The boar originated from a wild boar farm which only bred with pigs caught inthe wild. In JanuaryrFebruary, all experimental sows were inseminated with Yorkshiresperm. Hence, the offspring of the experimental animals were either pure domestic pigor 25% wild boar.
2.2. Housing conditions
Ž 2 .All sows farrowed in half open farrowing pens 9 m with a 30-cm high barrierseparating the farrowing pens from a 0.5-ha open enclosure, common to all sows. Halfof the farrowing pen was covered with about 5 kg of straw, the other half was used asdunging area. At the back of the farrowing pens, half-open doors allowed behaviouralobservations, without disturbances to sow or litter. Two days before farrowing, sowswere transferred from gestation enclosure to the farrowing pens and they were subse-quently fed twice daily in separate feeding crates in the outdoor pasture, in order toallow the experimenter to carry out essential manipulations in the farrowing pensŽ .weighing and numbering of the piglets, etc. . Sows were fed with 3 kg commercialconcentrate twice daily, and water was always available ad libitum.
2.3. Procedure
Farrowings took place in two batches, 4 weeks apart, during April and May, 1997. Inorder to allow a controlled observation schedule, all sows were injected with 2 ml

( )M. Gustafsson et al.rApplied Animal BehaÕiour Science 65 1999 29–4232
Ž .prostaglandin analogue PGF2 alpha at day 112 of gestation. Within batches, farrow-ings then took place within a 48-h period.
ŽOne hour after injection of PGF2 alpha, nest building material 20 1–1.5-m long.branches, 20 0.5-m long twigs and 4 kg of straw was supplied. Additional nest building
Ž .material 4 kg of straw was supplied 12 h after injection of PGF2 alpha, in case pigletswere not born at that time. Starting 1 h after injection of PGF2 alpha, nest buildingbehaviour was recorded during 15 min every third hour for each sow. This observationschedule continued for each sow until the first piglet was born. This resulted in moreobservation periods for those sows who took longer time to farrow.
Within 24 h after parturition, all piglets were weighed, ear tagged and cross-fosteredin such a way that for each sow, litter composition was balanced with respect to number
Ž .of piglets, number of piglets of each genotype pure domestic or 25% wild boar ,Ž .average piglet weight and sex ratio in order of priority .
Ž .Between 4 and 7 days after parturition week 1 , nursing behaviour was observedŽ .during 4 h for each sow, either after morning feeding 8:00 AM or after afternoon
Ž .feeding 1:30 PM . Each sow and litter was observed once for 4 h. Observations startedat first nursing, after feeding and continued for 4 h on the same sow. This observation
Ž . Ž .schedule was repeated from day 8 to 11 week 2 . From day 12 to 22 week 3–4 ,farrowing pens were open so that piglets were able to leave the farrowing pen. Samplingsessions were similar in week 3–4, but were extended by activity measures andmeasures of proximity between sow and piglets in the open enclosure. Piglets weremarked on their backs for identification.
Order of observation sessions was always balanced between batches and genotypes.
2.4. ObserÕation methods
Behaviour was recorded by means of direct observations and with PSION hand heldŽ .computer, and the Observer software Noldus Information Technology .
2.4.1. Nest building obserÕationsŽThe following variables were recorded for nest building adopted from Jensen et al.,
.1993 , with one–zero sampling during 15 consecutive 60 s periods.1. Lying; lying or sitting.2. Changing position; from lyingrsitting to standing or vice versa.3. Standing.4. Walking; the sow crossed the boundary between pen halves.5. Nosing; nose-contact with floor, walls or equipment in the pen.6. Rooting; digging movements with the snout, directed at the floor.7. Pawing; scratching movements with fore legs.8. Carrying material; fetching straw or twigs and taking at least two steps with the
material in the mouth.9. Arranging; manipulating material through rooting movements, nibbling, lifting,
followed by dropping and scattering of the material.10. Bitingrchewing; biting or chewing on equipment in the farrowing pen.

( )M. Gustafsson et al.rApplied Animal BehaÕiour Science 65 1999 29–42 33
( )2.4.2. Nursing obserÕations week 1, 2The following variables were recorded:Ž .1 Duration, frequencies and inter-nursing interval of all nutritive and non-nutritive
nursings. Nutritive nursings were defined as nursings with milk ejection, while non-nutritive nursings were those without milk ejection. Nutritive nursings were identified by
Ž .increased grunt rates by the sow and rapid sucking of the piglets Fraser, 1980 . Thefollowing variables were additionally recorded in nutritive nursings only.
Ž .2 Time of pre-massage. Pre-massage was considered to start when at least half thelitter started to massage the udder and to end when milk was ejected.
Ž .3 Time of post massage. Post massage was considered to start when milk ejectionwas over and continue until at least half the litter had ceased to massage the udder,instead participating in other activities, or if the sow rolled over.
Ž .4 Time sows allowed post-massage. The time sows were lying in nursing positionafter a nursing, exposing both udder rows, even if no actual massage was seen.
Ž .5 For each nursing, it was also recorded if the sow or piglets initiated andterminated the nursing. Piglets were regarded as initiators if at least half the litter startedto massage the udder without previous grunting of the sow. The sow was regarded asterminator if she rolled over or walked away from the piglets, before post-massage hadended.
Ž .6 Duration and frequency of the sow leaving the farrowing pen.
( )2.4.3. ActiÕity and proximity between sow and piglets week 3–4The sow’s activity, frequency of nose-contact between sow and piglets and the
proximity between sow and piglets, were recorded during 10 days in the outdoorenclosure during the third and fourth week after parturition. Each day, all sows in thebatch were continuously observed for 30 min each, one after the other; domestic andwild-type sows were observed alternately. The order of observation of the individualsows was changed in a balanced way between observation days. The following activity
Ž .states of the sows were continuously recorded adopted from Jensen, 1988 :1. Lying — lying or sitting.2. Foraging — rooting or grazing.3. Locomotion — moving without doing anything else.4. Nursing — sow was lying down exposing both udder rows and grunting regularly
and at least half the litter was at the udder.In addition, proximity measures were recorded every 30 s. For that purpose, it was
Žnoted how many of the sows’ own piglets were within one sow length approximately.1.5 m from the sow. All nose-contacts with own piglets initiated by the sow were also
recorded for each observation period. For each nursing observed, it was noted if the sowŽ Ž ..or piglets initiated and terminated the nursings see above Section 2.4.2 5 .
2.5. Statistical analyses
If not otherwise stated, the data were analysed by analysis of variance with a repeatedŽmeasures design involving the independent factors genotype two levels, i.e., domesti-
. Ž .cated and wild-type and age two levels, i.e., week 1, 2 and the interaction between
( )M. Gustafsson et al.rApplied Animal BehaÕiour Science 65 1999 29–4234
( )M. Gustafsson et al.rApplied Animal BehaÕiour Science 65 1999 29–42 35
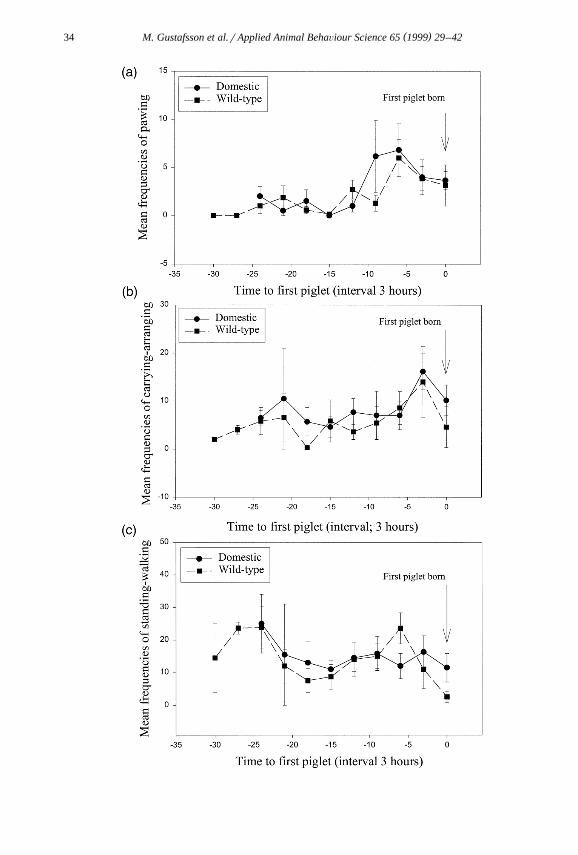
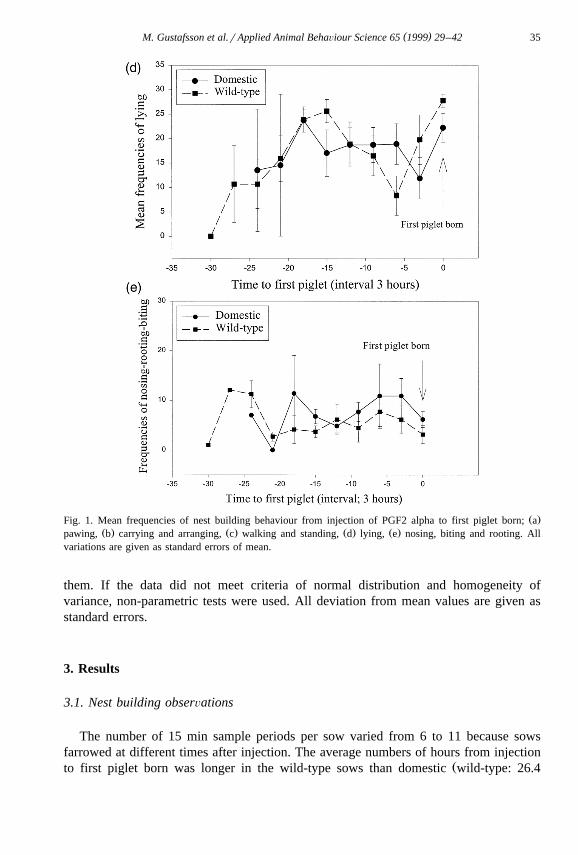
Ž .Fig. 1. Mean frequencies of nest building behaviour from injection of PGF2 alpha to first piglet born; aŽ . Ž . Ž . Ž .pawing, b carrying and arranging, c walking and standing, d lying, e nosing, biting and rooting. All
variations are given as standard errors of mean.
them. If the data did not meet criteria of normal distribution and homogeneity ofvariance, non-parametric tests were used. All deviation from mean values are given asstandard errors.
3. Results
3.1. Nest building obserÕations
The number of 15 min sample periods per sow varied from 6 to 11 because sowsfarrowed at different times after injection. The average numbers of hours from injection
Žto first piglet born was longer in the wild-type sows than domestic wild-type: 26.4
( )M. Gustafsson et al.rApplied Animal BehaÕiour Science 65 1999 29–4236
h"1.8, domestic: 19.0 h"1.6, Mann–Whitney Rank sum test: Ts27, N s6, N s7,1 2.p-0.05 . There were no significant differences in the frequency of pawing between the
Ž .genotypes over the period from injection to first piglet born Fig. 1a . The frequency ofcarrying and arranging, which are behaviours belonging to the last phase of nestbuilding, did also not differ between the genotypes over the period from injection to first
Ž . Ž .piglet born Fig. 1b and neither the frequencies of walking–standing Fig. 1c , lyingŽ . Ž .Fig. 1d or nosing–biting–rooting Fig. 1e .
3.2. Nursing obserÕations
A total of 155 nursings during week 1 and 2 were included in the study. Averageproportion of nursings that were non-nutritive was similar in both genotypes in the first
Ž . Žweek wild-types11.3%"5%, domestics10.8%"4% and in the second wild-types26.7%"7%, domestics14.4%"5%, no significant difference between either the
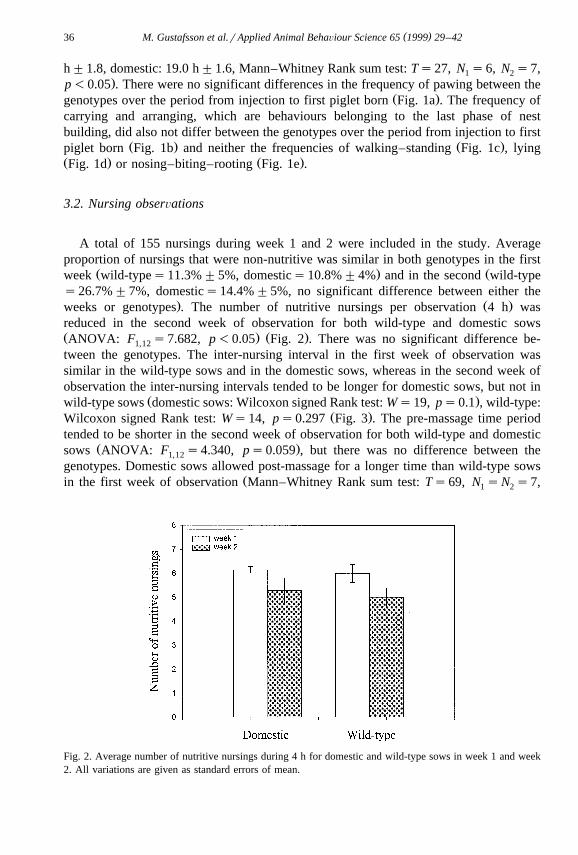
. Ž .weeks or genotypes . The number of nutritive nursings per observation 4 h wasreduced in the second week of observation for both wild-type and domestic sowsŽ . Ž .ANOVA: F s7.682, p-0.05 Fig. 2 . There was no significant difference be-1,12
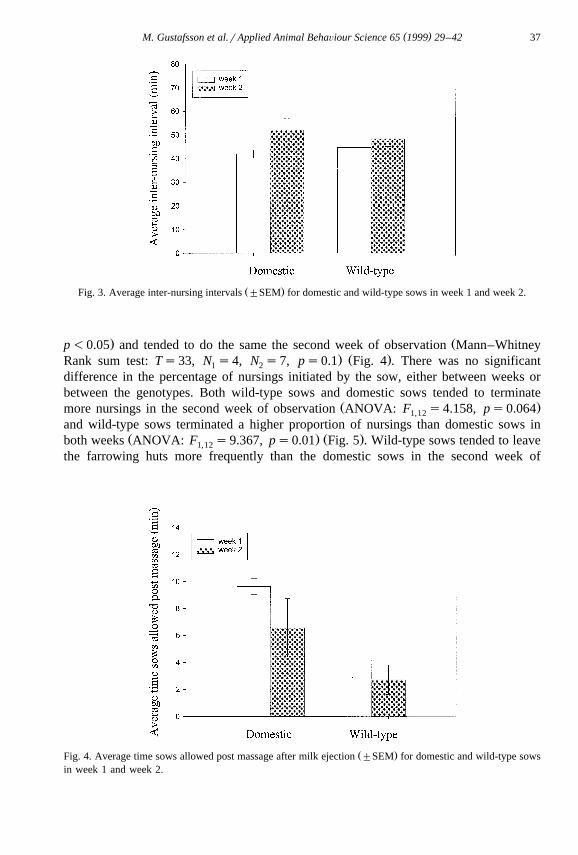
tween the genotypes. The inter-nursing interval in the first week of observation wassimilar in the wild-type sows and in the domestic sows, whereas in the second week ofobservation the inter-nursing intervals tended to be longer for domestic sows, but not in
Ž .wild-type sows domestic sows: Wilcoxon signed Rank test: Ws19, ps0.1 , wild-type:Ž .Wilcoxon signed Rank test: Ws14, ps0.297 Fig. 3 . The pre-massage time period
tended to be shorter in the second week of observation for both wild-type and domesticŽ .sows ANOVA: F s4.340, ps0.059 , but there was no difference between the1,12
genotypes. Domestic sows allowed post-massage for a longer time than wild-type sowsŽin the first week of observation Mann–Whitney Rank sum test: Ts69, N sN s7,1 2
Fig. 2. Average number of nutritive nursings during 4 h for domestic and wild-type sows in week 1 and week2. All variations are given as standard errors of mean.
( )M. Gustafsson et al.rApplied Animal BehaÕiour Science 65 1999 29–42 37
Ž .Fig. 3. Average inter-nursing intervals "SEM for domestic and wild-type sows in week 1 and week 2.
. Žp-0.05 and tended to do the same the second week of observation Mann–Whitney. Ž .Rank sum test: Ts33, N s4, N s7, ps0.1 Fig. 4 . There was no significant1 2
difference in the percentage of nursings initiated by the sow, either between weeks orbetween the genotypes. Both wild-type sows and domestic sows tended to terminate
Ž .more nursings in the second week of observation ANOVA: F s4.158, ps0.0641,12
and wild-type sows terminated a higher proportion of nursings than domestic sows inŽ . Ž .both weeks ANOVA: F s9.367, ps0.01 Fig. 5 . Wild-type sows tended to leave1,12
the farrowing huts more frequently than the domestic sows in the second week of
Ž .Fig. 4. Average time sows allowed post massage after milk ejection "SEM for domestic and wild-type sowsin week 1 and week 2.

( )M. Gustafsson et al.rApplied Animal BehaÕiour Science 65 1999 29–4238
Ž .Fig. 5. Average proportion of nursings terminated "SEM by domestic and wild-type sows in week 1 andweek 2.
Ž . Ž .observation Mann–Whitney Rank sum test: Ts39, N sN s7, p-0.1 Fig. 6 .1 2
However, there were no differences between genotypes in the first week.
3.3. Mother–young interactions
On average 189 min of observations per sow were included in the study during week3–4, including 58 nursings, 29 with domestic sows and 29 with wild-type sows.
The time spent lying tended to be longer for domestic sows compared to theŽ .wild-type sows Mann–Whitney Rank sum test: Ts67, N sN s7, ps0.0731 2
Ž .Fig. 6. Average frequencies of leaving pen "SEM for domestic and wild-type sows in week 1 and week 2.
( )M. Gustafsson et al.rApplied Animal BehaÕiour Science 65 1999 29–42 39
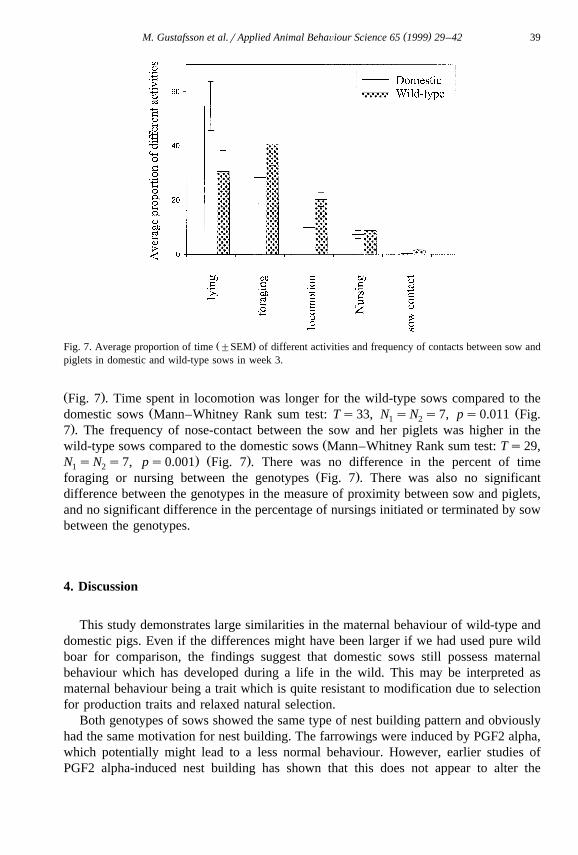
Ž .Fig. 7. Average proportion of time "SEM of different activities and frequency of contacts between sow andpiglets in domestic and wild-type sows in week 3.
Ž .Fig. 7 . Time spent in locomotion was longer for the wild-type sows compared to theŽ Ždomestic sows Mann–Whitney Rank sum test: Ts33, N sN s7, ps0.011 Fig.1 2
.7 . The frequency of nose-contact between the sow and her piglets was higher in theŽwild-type sows compared to the domestic sows Mann–Whitney Rank sum test: Ts29,
. Ž .N sN s7, ps0.001 Fig. 7 . There was no difference in the percent of time1 2Ž .foraging or nursing between the genotypes Fig. 7 . There was also no significant
difference between the genotypes in the measure of proximity between sow and piglets,and no significant difference in the percentage of nursings initiated or terminated by sowbetween the genotypes.
4. Discussion
This study demonstrates large similarities in the maternal behaviour of wild-type anddomestic pigs. Even if the differences might have been larger if we had used pure wildboar for comparison, the findings suggest that domestic sows still possess maternalbehaviour which has developed during a life in the wild. This may be interpreted asmaternal behaviour being a trait which is quite resistant to modification due to selectionfor production traits and relaxed natural selection.
Both genotypes of sows showed the same type of nest building pattern and obviouslyhad the same motivation for nest building. The farrowings were induced by PGF2 alpha,which potentially might lead to a less normal behaviour. However, earlier studies ofPGF2 alpha-induced nest building has shown that this does not appear to alter the

( )M. Gustafsson et al.rApplied Animal BehaÕiour Science 65 1999 29–4240
Ž .behaviour in any noticeable way Widowski et al., 1990; Boulton et al., 1998 . Bothtypes of sows also showed a similar nursing pattern, in both its qualitative and
Ž .quantitative aspects which also has been reported by Horrell 1997 for wild boars keptin an outdoor paddock. Apparently, selection has not altered these central aspects ofmaternal behaviour in pigs.
With respect to nest building, the fitness benefits of this behaviour may have beenŽ .large until very recently in history through increased survival of the piglets , whereas
the life-time reproductive costs involved in performing the behaviour may not be veryhigh. Therefore, there may not have been a strong selective pressure to remove thisbehaviour from the repertoire or to decrease its frequency, even though the selectiveadvantages have been removed through human care during domestication. Furthermore,
Žnest building only occurs within a 24-h period before farrowing at the most two times.per year , which means that a sow does not perform this behaviour very often during her
life-time. It seems likely that such an infrequent but essential behaviour is hardwiredinto the nervous system and may be altered only with an intensive selection program. Aninteresting observation was that the interval between injection of PGF2 alpha and onsetof parturition was almost 8 h longer in the wild-type sows. This indicates a domestica-tion induced difference in physiological responsiveness to parturition related hormonalchanges. Even though this is less important for the present study, it may be aninteresting finding to follow up in future research.
As the piglets in this study grew older, sows terminated more nursings, the number ofnutritive nursings decreased and inter-nursing interval increased. This pattern is in
Žaccordance with general predictions from parent–offspring conflict theory Trivers,. Ž1974 , and has also been reported in other studies of maternal behaviour of pigs Jensen,
.1988; Jensen and Recen, 1989; Jensen et al., 1991 . Both genotypes had a similar´Ž .proportion of non-nutritive nursings, which supports suggestions of Jensen et al. 1991
Ž .and Illmann et al. 1999 that non-nutritive nursings probably constitute a part of thenormal behaviour repertoire of pigs. However, their function still remains unknownŽ .Illmann et al., 1999 .
Ž .According to Price 1984, 1997 , there are three main processes involved in theŽ .selection during domestication. 1 Relaxed natural selection of certain selection factors,
Ž .such as predation and food shortage Price, 1997 . This process might have affectedmaternal abilities, since human protection and feeding may have decreased the reproduc-
Ž .tive advantages of some costly behaviour strategies Jensen and Gustafsson, 1997 , suchas intense protection of piglets and nest defence, which will be discussed further in
Ž .another paper. 2 Artificial selection; this active, conscious selection of preferred traitsŽ .is a historically rather late phenomenon Clutton-Brock, 1981 . Artificial selection is
Ž .always accompanied by 3 selection of linked traits, because of genetic or functionalŽ .linkage to the preferred trait. For example, according to Ruzzante 1994 , selection on
growth may at the same time cause selection for increased aggression in fish. Since allthree processes act simultaneously, it is difficult to assess their relative importance inaffecting maternal behaviour. However, this study suggests that even though theselection processes during domestication have acted over a long period of time, there arevery small differences in the maternal behaviour of wild boar crossings and domesticpigs.

( )M. Gustafsson et al.rApplied Animal BehaÕiour Science 65 1999 29–42 41
Although the similarities between the genotypes were striking, there were a fewnoticeable differences in the maternal behaviour. Domestic sows were less active and itappeared as if they had less nose-contact with their piglets. Other differences weremostly attributed to the nursing behaviour, i.e., wild-type sows terminated morenursings, stayed in the recumbent nursing position for shorter time after milk ejectionand tended to leave their piglets more frequently in the first 2 weeks; these arebehaviours that are related to the parental investment in the litters. We might speculatethat the wild-type sows are genetically adapted to a situation where it is more beneficialfrom a life-time reproductive success prospective to save resources for future offspringand reduce the care for the present. If so, the domestic sows may be better adapted to alife in human protection where they do not benefit from considering future offspring tothe same extent, since future reproduction is safe-guarded to a larger degree by humaninteractions and provision of food and protection. Should this be the case, domestic sowsmay be adapted to a situation where a strategy to invest more in present offspring bringshigher life-time reproductive success. One alternative possibility could be that wild-typesows were to a larger extent compared with the domestic sows stressed by theconfinement. However, it is not likely that this is the case since all sows were born andraised together with their mothers in this environment.
5. Conclusion
Ž .1 The motivation to perform pre-parturient nest building was largely unaffected bydegree of domestication, as was the actual pattern of nest building behaviour.
Ž .2 The early maternal behaviour and nursing behaviour was largely unaffected bydegree of domestication, even though some differences between domestic and wild-typesows may be interpreted as an adaptation to a life in human protection.
Ž .3 At 3 weeks of age of the piglets, the similarities between the genotypes in nursingand keeping in proximity with the young prevailed. However, domestic sows tended tobe less active and had less nose-contact with the offspring.
These findings suggest that the pre-parturient needs of sows and the ability to care foroffspring have not been substantially modified through domestication. Results suggestthat domestic sows still possess maternal behaviour similar to the behaviour of the wildboar.
Acknowledgements
Financial support was received from the Swedish Council for Forestry and Agricul-tural Research. Maria Gustafsson received travel grants from the Royal SwedishAcademy of Agriculture and Forestry and Swedish University of Agricultural Sciences.Participation of Gudrun Illmann and Marek Spinka in the experiment was supported bythe grant No. 524r96r0958 from the Grant Agency of the Czech Republic. Thanks toAnite Mulder for helping out with some of the behavioural observations.

( )M. Gustafsson et al.rApplied Animal BehaÕiour Science 65 1999 29–4242
References
Boulton, B.I., Wickens, A., Goode, J.A., Lawrence, A.B., Gilbert, C.L., 1998. Does prolactin mediate inducednest-building behaviour in pseudopregnant gilts treated with PGF2a? J. Neuroendocrinol. 10, 601–609.
Clutton-Brock, J., 1981. Domesticated animals from early times. University of Texas Press, Austin.Cronin, G.M., Smith, J.A., Hodge, F.M., Hemsworth, P.H., 1994. The behaviour of primiparous sows around
farrowing in response to restraint and straw bedding. Appl. Anim. Behav. Sci. 39, 269–280.de Jonge, F.H., Bokkers, E.A.M., Schouten, W.G.P., Helmond, F.A., 1996. Rearing piglets in a poor
environment: developmental aspects on social stress in pigs. Physiol. Behav. 60, 1–7.Dyck, G.W., Swierstra, E.E., 1987. Causes of piglet death from birth to weaning. Can. J. Anim. Sci. 67,
543–547.English, P.R., Smith, W.J., 1975. Some Causes of Death in Neonatal Piglets, 15, 95-104.Fraser, D., 1980. A review of the behavioural mechanism of milk ejection of the domestic pig. Appl. Anim.
Ethol. 6, 247–255.Fraser, D., 1990. Behavioural perspectives on piglet survival. J. Reprod. Fertil. 40, 355–370.Gundlach, H., 1968. Brutfursorge, Brutpflege, Verhaltensontogenese und Tagesperiodik beim Europaischen¨ ¨
Ž .Wildschwein Sus scrofa L. . Z. Tierpsychol. 25, 955–995.Hafez, E.S.E., 1962. The behaviour of domestic animals. Williams and Wilkins, Baltimore.Hanson, R.P., Karstad, L., 1959. Feral swine in the south eastern United States. J. Wildl. Manage. 23, 64–79.Hemmer, H., 1990. Domestication — the decline of environmental appreciation. Cambridge Univ. Press,
Cambridge.Horrell, I., 1997. The characterisation of suckling in wild boar. Appl. Anim. Behav. Sci. 53, 271–277.Illmann, G., Spinka, M., Stetkova, Z., 1999. Predictability of nursings without milk ejection in domestic pigs.
Appl. Anim. Behav. Sci. 61, 303–311.Jensen, P., 1986. Observations on the maternal behaviour of free ranging domestic pigs. Appl. Anim. Behav.
Sci. 16, 131–142.Jensen, P., 1988. Maternal behaviour and mother–young interactions during lactation in free ranging domestic
pigs. Appl. Anim. Behav. Sci. 20, 297–308.Jensen, P., Recen, B., 1989. When to wean — observations from free ranging domestic pigs. Appl. Anim.´
Behav. Sci. 23, 49–60.Jensen, P., Gustafsson, M., 1997. Towards a functional view on domestication. Proc. 31st Intern. Congress of
the ISAE, 13–16 of August, Prague, Czech Republik. Polygravia SAV, pp. 43–44.Jensen, P., Stangel, G., Algers, B., 1991. Nursing and suckling behaviour of semi-naturally kept pigs during
the first 10 days postpartum. Appl. Anim. Behav. Sci. 31, 195–210.Jensen, P., Vestergaard, K., Algers, B., 1993. Nest building in free-ranging domestic sows. Appl. Anim.
Behav. Sci. 38, 245–255.Lawrence, A.B., Petherick, J.C., McLean, K.A., Deans, L.A., Chirnside, J., Vaughan, A., Clutton, E.,
Terlouw, E.M.C., 1994. The effect of environment on behaviour, plasma cortisol and prolactin in parturientsows. Appl. Anim. Behav. Sci. 39, 313–330.
Noldus Information Technology. Wageningen, The Netherlands.Price, E.O., 1984. Behavioral aspects of animal domestication. Q. Rev. Biol. 59, 1–32.
Ž .Price, E.O., 1997. Behavioural genetics and the process of animal domestication. In: Grandin, T. Ed. ,Genetics and the behaviour of domestic animals, Academic Press, New York, pp. 31–65.
Ruzzante, D.E., 1994. Domestication effects on aggressive and schooling behavior in fish. Aquaculture 120,1–24.
Trivers, R.L., 1974. Parent–offspring conflict. Am. Zool. 14, 249–264.Widowski, T.M., Curtis, S.E., Dzuik, P.J., Wagner, W.C., Sherwood, O.D., 1990. Behavioural and endocrine
responses of sows to prostaglandin F2a and cloprostenol. Biol. Reprod. 43, 290–297.





























