Embed Size (px)
Citation preview

Hum Genet
DOI 10.1007/s00439-008-0603-8ORIGINAL INVESTIGATION
Maternal age and risk for trisomy 21 assessed by the origin of chromosome nondisjunction: a report from the Atlanta and National Down Syndrome Projects
Emily Graves Allen · Sallie B. Freeman · Charlotte Druschel · Charlotte A. Hobbs · Leslie A. O’Leary · Paul A. Romitti · Marjorie H. Royle · Claudine P. Torfs · Stephanie L. Sherman
Received: 15 September 2008 / Accepted: 26 November 2008© Springer-Verlag 2008
Abstract We examined the association between mater-nal age and chromosome 21 nondisjunction by origin ofthe meiotic error. We analyzed data from two population-based, case–control studies: Atlanta Down Syndrome Pro-ject (1989–1999) and National Down Syndrome Project(2001–2004). Cases were live born infants with trisomy
21 and controls were infants without trisomy 21 deliveredin the same geographical regions. We enrolled 1,215 of1,881 eligible case families and 1,375 of 2,293 controls.We report four primary Wndings. First, the signiWcantassociation between advanced maternal age and chromo-some 21 nondisjunction was restricted to meiotic errors inthe egg; the association was not observed in sperm or inpost-zygotic mitotic errors. Second, advanced maternalage was signiWcantly associated with both meiosis I (MI)and meiosis II (MII). For example, compared to mothersof controls, mothers of infants with trisomy 21 due to MInondisjunction were 8.5 times more likely to be¸40 years old than 20–24 years old at the birth of theindex case (95% CI = 5.6–12.9). Where nondisjunctionoccurred in MII, mothers were 15.1 times more likely tobe ¸40 years (95% CI = 8.4–27.3). Third, the ratio of MIto MII errors diVered by maternal age. The ratio was loweramong women <19 years of age and those ¸40 years(2.1, 2.3, respectively) and higher in the middle agegroup (3.6). Lastly, we found no eVect of grand-maternalage on the risk for maternal nondisjunction. This studyemphasizes the complex association between advancedmaternal age and nondisjunction of chromosome 21 dur-ing oogenesis.
Introduction
The chromosomal basis of Down syndrome, trisomy 21,has been recognized for nearly half a century (Book et al.1959; Ford et al. 1959; Jacobs et al. 1959; Lejeune 1959),and the link between Down syndrome and advanced mater-nal age predates that discovery by at least another half cen-tury (Penrose 1933; Penrose 1934). In the United States,over 5,000 infants are born with Down syndrome (DS) each
The Wndings and conclusions in this report are those of the authors and do not necessarily represent the oYcial position of the Centers for Disease Control and Prevention.
E. G. Allen (&) · S. B. Freeman · S. L. ShermanDepartment of Human Genetics, Emory University, 2165 North Decatur Rd, Decatur, Atlanta, GA 30030, USAe-mail: [email protected]
C. DruschelNew York State Department of Health , Troy, NY, USA
C. A. HobbsDepartment of Pediatrics, College of Medicine, University of Arkansas for Medical Sciences, Little Rock, AR, USA
L. A. O’LearyCenters for Disease Control and Prevention, National Center on Birth Defects and Developmental Disabilities, Atlanta, GA, USA
P. A. RomittiDepartment of Epidemiology, College of Public Health, The University of Iowa, Iowa City, IA, USA
M. H. RoyleNew Jersey Department of Health and Senior Services, Trenton, NJ, USA
C. P. TorfsBirth Defects Studies, Public Health Institute, Emeryville, CA, USA
123

Hum Genet
year with an estimated prevalence of 13.65/10,000 births (1in 733) (CanWeld et al. 2006). We now know that the extrachromosome 21 is the result of nondisjunction during meio-sis in either the egg or the sperm (standard trisomy 21) inapproximately 95% of individuals (e.g., Mutton et al.1996).
In addition to Down syndrome and other trisomic livebirths, the impact of maternal-age-related chromosomenondisjunction on human reproductive health is evidentfrom studies of spontaneous pregnancy losses and humanoocytes. A recent investigation of over 1,700 karyotypedproducts of conception found that 14% of pregnancy lossesin women <24 years of age were due to a trisomy, and thatproportion rose to approximately 38% in women 40–44 years of age (Yusuf and Naeem 2004). Pellestor et al.(2003) found similar rates of aneuploidy among 1,367karyotyped oocytes from 520 women: 8.5% of oocytesfrom women <24 years had aneuploidy compared with39.5% of those from women between the ages of 40–44.Thus, it is thought that an increase in aneuploidy is themajor underlying factor responsible for the increased infer-tility observed among women with advancing age (Hassoldand Chiu 1985; Hassold and Hunt 2001; Pellestor et al.2002).
The basis of the maternal age eVect on aneuploidyremains one of the most important questions in medicalgenetics. A number of hypotheses center on general aspectsof ovarian function such as maternal age-related changesassociated with oocyte pool size or hormone function (e.g.,Eichenlaub-Ritter and Boll 1989; Gaulden 1992; Warbur-ton 2005). These general hypotheses have encouragedresearchers to seek underlying mechanisms by assessingspeciWc oocyte components for their vulnerability to mater-nal aging including mitochondria, the spindle apparatus,and sister cohesion protein complexes (e.g., de Bruin et al.2004; Eichenlaub-Ritter et al. 2004; Hodges et al. 2005;Schon et al. 2000; Steuerwald et al. 2005). Others haveinvestigated indicators of oocyte pool size, such as hor-mone proWles or antral follicle counts, to determine ifwomen who have had a nondisjunction event have smalleroocyte reserves than do controls (e.g., Freeman et al. 2000;van Montfrans et al. 2002; Warburton 2005). In addition,we have recently examined recombination patterns knownto be more common among nondisjoined chromosomes 21within the context of maternal age in order to shed light onthe interaction between these two established risk factors(Lamb et al. 2005; Oliver et al. 2008).
Historically, interest in the relationship between mater-nal age and chromosome nondisjunction has led investiga-tors to examine the eVect of grand-maternal age (Aagesenet al. 1984; Greenberg 1963; Malini and Ramachandra2006; Papp et al. 1977; Penrose 1964; Richards 1970; Stol-ler and Collmann 1969). The most commonly proposed
mechanism for a grand-maternal age eVect involves nondis-junction in a grand-maternal oocyte leading to a trisomicembryo. Loss of the extra chromosome in a proportion ofcells in that embryo would result in a mosaic oVspring (themother) without the DS phenotype, but with the capabilityof producing a trisomic child, assuming trisomic cellsremained in her gonads. Thus, risk factors associated withnondisjunction would apply to the grandmother of the pro-band with trisomy as well as to the mother. These factorscould be maternal-age related or not.
The use of chromosome 21-speciWc DNA markers todetermine the parental origin of the extra chromosome hasenabled investigators to reWne studies of the maternal ageeVect by including only cases of maternal origin. Such pop-ulation-based studies have reported that approximately90% of errors are maternal (e.g., Gomez et al. 2000; Mik-kelsen et al. 1995; Sherman et al. 2005; Yoon et al. 1996).In addition, speciWc pericentromeric chromosome 21 mark-ers allow cases to be categorized as the result of an error ineither meiosis I or meiosis II. Early on, investigatorsexpected the maternal age eVect to be limited to maternalmeiosis I (MMI) cases because MMI begins during the fetallife of the mother and is completed decades later at the timeof ovulation. In contrast, maternal meiosis II (MMII) is ini-tiated and completed in 3–4 days at the time of ovulation. Iftrue, the ratio of MMI to MMII would be predicted toincrease with increasing maternal age. Although both MMIand MMII errors are observed in trisomies of mothers of allages and overall show an average 3:1 ratio (Antonarakiset al. 1992; Muller et al. 2000; Sherman et al. 2005; Yoonet al. 1996), this speciWc hypothesis has not been addressed.In this study, we were able to examine pattern of MMI toMMII errors among trisomic outcomes to shed light on theinXuence of maternal age. Interestingly, Lamb et al. (1996)found speciWc diVerences in recombination patternsbetween MMI and MMII errors. They suggested that someMMII errors may not be the “classical” malsegregation ofsister chromatids during meiosis II, but instead may becaused by errors initiated in meiosis I. Thus, as is true formost other autosomal chromosomes (Hassold and Hunt2001) the majority of chromosome 21 maternal meioticerrors may begin during meiosis I,.
To better understand the eVect of maternal age on non-disjunction of chromosome 21 over the entire reproductivelife span of a woman, we have conducted a multi-year,multi-site case–control study (Freeman et al. 2007). Repre-senting six sites nation-wide and 16 years of recruitment,this population-based, case–control study is the largestcompilation of molecular, clinical, and epidemiologicaldata on trisomy 21 characterized by the origin of the chro-mosomal error. Evidence from this data set sheds additionallight on the relationship between maternal age and meioticnondisjunction.
123

Hum Genet
Subjects and methods
Subjects
Data from two population-based, case–control studies arepresented here. The studies diVer from each other withrespect to the time frame of data collection and the geo-graphic areas covered, but are otherwise nearly identical inmethodology. Both projects were based at Emory Univer-sity in Atlanta, Georgia and were approved by the appropri-ate Institutional Review Boards at all participating sites.Study personnel obtained informed consent from all partic-ipating families. From 1989 to 1999, investigators at EmoryUniversity directed the Atlanta Down Syndrome Project(ADSP) in cooperation with the Centers for Disease Con-trol and Prevention (CDC). Eligible cases were identiWedthrough a birth defect surveillance system, the MetropolitanAtlanta Congenital Defects Program (MACDP) (Correa-Villasenor et al. 2003), and included all live born infantswith documented trisomy 21 or mosaic trisomy 21 born inthe Wve-county Atlanta metropolitan area. Controls wereidentiWed through hospital records and randomly selectedfrom among newborns without birth defects from the samegeographical population. Trained interviewers administeredquestionnaires by telephone or in person to mothers ofcases and controls and obtained blood samples from caseinfants, their mothers, and their fathers when available. TheADSP completed enrollment in 2000. Further details of theADSP are available in earlier publications (Yang et al.1999; Yoon et al. 1996).
Between 2001 and 2004, Emory expanded the study toinclude Wve other sites nationwide, all of which had wellestablished birth defect surveillance systems. These sitesascertained cases and controls either statewide (Arkansas,Iowa, New Jersey) or within deWned geographic areaswithin the state (California, New York). Together, the sixsites represented approximately 11% of annual births in theUS. As with the ADSP, the National Down Syndrome Pro-ject (NDSP) enrolled cases and controls on a populationbasis and collected both questionnaire data and biologicalsamples. All six NDSP sites Wnished their recruitmenteVorts by May 2005. A detailed description of the designand implementation of the NDSP is available (Freemanet al. 2007).
In both studies an eligible case was deWned as a live borninfant with standard trisomy 21 or mosaic trisomy 21 bornto a mother living in one of the study areas at the time of thebirth. For practical reasons, NDSP-eligible cases were fur-ther limited by their “recruitability”. Recruitable familieswere those in which the mother spoke either English orSpanish and the infant was alive and available so that a bio-logical sample could be collected at enrollment. For thepresent report, eligible and recruitable NDSP cases are simply
referred to as eligible. Combining all sites and all birthyears, the ADSP and NDSP identiWed 1,881 eligible liveborn cases of trisomy 21 or mosaic trisomy 21 and 2,293controls (Table 1). As in ADSP, study personnel inter-viewed mothers of cases and controls by telephone or inperson to obtain information on maternal age, grand-mater-nal age, and ethnicity (race/ethnicity). In addition, for bothparticipating and non-participating mothers, independentinformation regarding maternal age and ethnicity was avail-able from birth records. Coding of ethnicity varied some-what from site to site, but for this report we reduced thegroups to (1) white non-Hispanic, (2) black non-Hispanic,(3) Hispanic, (4) American Indian/Alaskan Native, (5)Asian, (6) other, and (7) unknown. Among participatingmothers, we found good agreement between self-reportedethnicity and ethnicity from birth records (white 96%, black95%, Hispanic 98%). In order to be able to include ourentire sample of eligible families for these analyses, weused ethnicity from birth records. For the general popula-tion, maternal age at the birth of the infant came from vitalstatistics obtained from the same geographic areas of sur-veillance during the same years of data collection.
For both studies, the minimum requirements for a casefamily to be deWned as “enrolled” were a completed mater-nal questionnaire and biological samples from at least themother and the infant with DS. The minimum requirementfor a control family was the completion of a maternal ques-tionnaire. In total, 1,215 case families and 1,375 controlfamilies successfully enrolled in the ADSP and NDSP (par-ticipation rates: cases 64.6%; controls 60.0%). Table 1 pre-sents total population data and a breakdown of thesenumbers by study (ADSP or NDSP) and by NDSP site andincludes mean maternal age at the birth of the child forcases and controls.
Laboratory methods
Each participating family was genotyped for a panel ofchromosome 21-speciWc polymorphic markers (primarilysmall tandem repeat polymorphisms, STRPs) that span 21q(Freeman et al. 2007). The contribution of parental markeralleles to the infant with DS was used to establish theparental origin of the nondisjunction error and was basedon at least one informative marker. In a minority of cases,the father was unavailable and the origin of the error wasbased on information from the mother and infant only. Inthese cases, at least eight markers had to be informative.We inferred that an error was maternal when all markerswere consistent with a maternal origin. A paternal error wasinferred when at least two markers were inconsistent with amaternal origin. There were 20 ADSP cases and 125 NDSPcases for whom we could not determine the parental origin.Probands who were determined to be mosaic trisomy 21 by
123
Hum Genet
cytogenetic analysis were not analyzed using genetic mark-ers, as dosage cannot be accurately determined. Thus, dataon mosaics are only included among “eligible” cases.
Once parental origin was established, a core set of mark-ers located in the pericentromeric region was used to inferthe type of nondisjunction error (MI or MII). SpeciWcally, ifparental heterozygosity was retained in the trisomicoVspring (“nonreduction”), we concluded that nondisjunc-tion occurred during meiosis I with failure of homologs toseparate properly. If parental heterozygosity was reduced tohomozygosity (“reduction”), we inferred an MII error. Asnoted in the Introduction, there is evidence to suggest someportion of the MII errors may actually originate during mei-osis I in which homologs fail to segregate properly fol-lowed by an error in meiosis II in which sister chromatidsfail to separate; hence, our assay based on pericentromericalleles in the oVspring will only identify the contribution
from one parental allele. Thus, we deWne MMII cases asmaternally-derived cases in which chromosome 21-speciWcpericentromeric markers from the mother are identical bydescent.
The last category of errors includes post-zygotic mitoticerrors. These are characterized as such when all informativemarkers in the parent of origin are reduced to homozygosityalong the entire chromosome. The criterion to establish anerror as mitotic included having at least eight informativehomozygous markers spanning the length of 21q. Suchcases could also be inferred to be MII errors with no recom-bination. We do not have a method to accurately distin-guish these two types of errors. We expected these cases tobe maternal-age independent and, in fact, found this to bethe case: the mean maternal age of so-called mitotic cases didnot diVer from that of controls (Table 2). However, we alsoexpected that there should be equal numbers of “maternal” and
Table 1 Sample size, mean maternal age and standard deviation (SD) for the Atlanta Down Syndrome Project (ADSP), National Down SyndromeProject (NDSP) and for total birth population during the study period
Maternal age was missing for 20 cases and 1 control
Study site (years of enrollment)
Study sample
Eligible Enrolled Participation rate (%)
N Maternal age (mean § SD)
N Maternal age (mean § SD)
ADSP–GA1/89–12/99
Case 400 30.5 § 7.2 308 30.0 § 7.0 77.0
Control 577 27.2 § 6.2 398 27.2 § 6.2 69.0
Population N = 447,436; maternal age = 27.4 § 6.1
NDSP Total Case 1,481 33.2 § 7.2 907 33.1 § 7.1 61.4
Control 1,716 28.1 § 6.2 977 28.8 § 6.2 56.9
Population N = 1,460,083; maternal age = 28.3 § 6.2
NDSP–GA1/01–9/04
Case 206 33.5 § 6.7 154 33.4 § 6.6 74.8
Control 239 28.3 § 5.8 170 28.9 § 5.8 71.1
Population N = 204,696; maternal age = 28.1 § 6.2
NDSP–AR10/00–9/03
Case 96 30.1 § 8.6 68 30.0 § 8.5 70.8
Control 117 25.2 § 6.1 75 26.3 § 6.2 64.1
Population N = 110,007; maternal age = 25.3 § 5.7
NDSP–CA1/01-6/03
Case 498 33.3 § 7.4 267 33.0 § 7.4 53.6
Control 604 27.7 § 6.4 261 28.1 § 6.5 43.2
Population N = 563,211; maternal age = 28.3 § 6.3
NDSP–IA1/01–12/03
Case 126 31.7 § 6.8 75 31.8 § 6.7 59.5
Control 150 27.2 § 5.4 81 28.0 § 5.0 54.0
Population N = 113,295; maternal age = 27.1 § 5.7
NDSP–NJ1/01-6/04
Case 400 34.2 § 6.8 252 34.2 § 6.6 63.0
Control 457 29.3 § 6.0 299 30.0 § 6.1 65.4
Population N = 329,587; maternal age = 29.4 § 6.1
NDSP–NY10/00–9/03
Case 155 33.4 § 7.1 91 33.1 § 6.6 58.7
Control 149 29.2 § 6.2 91 29.2 § 6.2 61.1
Population N = 139,287; maternal age = 29.1 § 6.2
ADSP + NDSP Total Case 1,881 32.7 § 7.3 1,215 32.3 § 7.2 64.6
Control 2,293 27.9 § 6.2 1,375 28.3 § 6.2 60.0
Population N = 1,907,519; maternal age = 28.1 § 6.2
123

Hum Genet
“paternal” errors among such cases. Instead, we observedabout three times as many “maternal” cases as paternalcases (26 maternal and 8 paternal cases). Thus, some casesthat we classiWed as being due to mitotic errors actuallymay be due to meiotic errors. Nevertheless, if a case wasdetermined to be a mitotic error using the criteria above, itwas not included in any analyses of meiotic errors.
Statistical analysis
In order to test the validity of combining the ADSP andNDSP populations, we investigated diVerences in maternalage using t tests and in ethnicity using analysis of varianceadjusting for covariates (ANCOVA). SigniWcant resultsfrom ANOVAs were follow-up by post-hoc pairwise com-parisons using Tukey’s Studentized range test. Linearregression was used to test for an association betweenmaternal age and birth year. Logistic regression was used todetermine if there was an association between the type ofnondisjunction error and ethnicity.
After establishing that our two study populations couldbe combined when adjustments for ethnicity and year ofbirth were made (see “Results”), our objective was toinvestigate the maternal age eVect on each type of nondis-junction error, similar to our previous study on a signiW-cantly smaller data set (Yoon et al. 1996). Our basicapproach was logistic regression for comparison of DS caseand control groups as well as for comparison of MMI andMMII groups. First, we compared mothers with each spe-ciWc nondisjunction error (i.e., MMI or MMII error) to con-trol mothers to obtain odds ratios (ORs) for being in aspeciWc maternal age group adjusting for ethnicity and birthyear of the infant. Case-population analysis, adjusting forbirth year only, was done to obtain rate ratios (RRs). Mater-nal ages were divided into six groups: ·19, 20–24, 25–29,30–34, 35–39, and ¸40 years of age and age 20–24 wasused as the referent group. Second, to ask if maternal ages
diVered by origin of the error, we compared MMI andMMII groups and included maternal age as a continuousvariable and its square to account for non-linear eVects aspredictors. All models were adjusted for maternal ethnicity(white, black, Hispanic and other) and birth year (continu-ous variable).
We also investigated grand-maternal age as a risk factorfor nondisjunction. For these analyses, we used enrolledcases and controls. Similar to others, we hypothesized thatthe grand-maternal age eVect may be more detectableamong younger mothers who had a nondisjunction errorcompared with older mothers, as the younger mother’s ownmaternal age-related factors would be minimized. Thus, westratiWed enrolled cases and controls by maternal age (<30and ¸30 years) and conducted logistic regression usinggrand-maternal age as a continuous variable and maternalrace and maternal age as covariates in the model.
All analyses were done using SAS V9.
Results
Study sample characteristics
Before investigating the speciWc eVect of maternal age onnondisjunction, we Wrst examined the mean maternal ageand ethnic composition of the ADSP, NDSP, and the com-bined study samples for their potential confounding eVects.For these analyses, we used all eligible cases and controlsto get the most accurate population-based estimates. Wefound that the mean maternal age overall increased fromthe ADSP (1989–2000) to the NDSP (2001–2004) for bothcases and controls (Table 1). Adjusting for case/control sta-tus, there was a statistically signiWcant increase in maternalage for each unit increase in birth year (p < 0.0001; partialr2 = 0.02) Interestingly, there was a signiWcant interactionbetween case/control status and birth year (p = 0.0012;
Table 2 Origin of chromosome error, mean maternal age and standard deviation (SD) for enrolled participants in ADSP, NDSP, and control populations
Origin ADSP NDSP ADSP + NDSP
N % Maternal age N % Maternal age N % Maternalage
Maternal 255 88.5 30.5 § 7.0 729 93.2 33.6 § 6.8 984 92.0 32.8 § 7.0
MI 201 79.8 30.6 § 6.8 529 74.7 33.4 § 6.6 730 76.0 32.6 § 6.8
MII 51 20.2 30.3 § 7.8 179 25.3 34.2 § 7.3 230 24.0 33.3 § 7.6
Paternal 20 6.9 25.8 § 5.2 32 4.1 29.7 § 6.6 52 4.9 28.2 § 6.3
MI 9 45.0 25.6 § 5.6 13 41.9 30.3 § 7.0 22 42.3 28.4 § 6.7
MII 11 55.0 25.9 § 5.1 18 58.1 28.9 § 6.3 29 55.8 27.8 § 6.0
Mitotic 13 4.5 30.0 § 5.6 21 2.7 26.9 § 6.0 34 3.2 28.1 § 5.9
Total of cases 288 30.1 § 6.9 782 33.3 § 6.9 1,070 32.4 § 7.0
Controls 398 27.2 § 6.2 977 28.8 § 6.2 1,375 28.3 § 6.2
Population 447,436 27.4 § 6.1 1,460,083 28.3 § 6.2 1,907,519 28.1 § 6.2
Totals may be greater than the sum of sum categories because the origin of the nondisjunction error could not be determined in all cases. The proportion of MI and MII are among the parental subtypes
123

Hum Genet
partial r2 = 0.002) which indicates a steeper increase overtime in mean maternal age among cases than among controls(Fig. 1). The increase for controls across the two studieswas approximately one-half that observed in cases.
It was also evident from the changing demographics ofAtlanta over time and the diVerent demographics at eachNDSP site that ethnicity would vary between the ADSP andNDSP. The ADSP had a higher proportion of cases andcontrols who were black and a signiWcantly smaller propor-tion of Hispanics than did the NDSP. Comparison of meanmaternal ages indicated variation by ethnic group (Table 3).In both the ADSP and NDSP, white mothers tended to beolder than their black or Hispanic counterparts. SpeciW-cally, for both cases and controls, white mothers werefound to be signiWcantly older than black mothers(p < 0.01) and Hispanic mothers (p < 0.01); blacks and His-panics were not signiWcantly diVerent from each other(p > 0.05). These results were the same if all eligible sub-jects or if only enrolled subjects were used.
We also tested ethnicity as a predictor variable for case/control status when the model was adjusted for maternalage and birth year. The combined data set of all eligiblecases and controls was used for this analysis. The casemothers more often self-reported as Hispanics than aswhites compared to control mothers (adjusted OR = 1.3;95% CI = 1.1–1.6). In addition, the case mothers less oftenself-reported as blacks than whites compared to controlmothers, although this reduction was not statistically sig-niWcant (adjusted OR = 0.9; 95% CI = 0.7–1.1).
Maternal age by type of nondisjunction error
Parental origin
We examined parental origin of the nondisjunction error andits relationship to maternal age. As we required biological sam-ples to determine parental origin, we compared mean maternalages among enrolled cases to mean maternal ages of enrolled
Fig. 1 Comparison of mean maternal ages at the time of birth by the infant’s birth year. Case maternal age at birth of infant with trisomy 21, Control mater-nal age at birth of infant without trisomy 21, Population maternal ages at birth of infants in the population from with cases and controls were drawn
20
22
24
26
28
30
32
34
36
1988 1990 1992 1994 1996 1998 2000 2002 2004 2006
Birthyear
Mea
n M
ater
nal
Ag
e
Cases Controls Population
Table 3 Maternal race/ethnicity, mean maternal age and standard deviation (SD) for all eligible cases and controls
Study sample White Black Hispanic Other
N % Maternal age N % Maternal age N % Maternal age N % Maternal age
ADSP
Cases 199 49.7 31.8 § 6.2 153 38.2 29.1 § 8.1 27 6.7 27.2 § 7.3 21 5.2 32.3 § 6.2
Controls 269 46.6 29.4 § 5.5 253 43.8 24.7 § 6.2 34 5.9 26.4 § 5.7 21 3.6 29.3 § 5.5
NDSP
Cases 636 42.9 34.0 § 6.4 185 12.5 32.3 § 8.1 574 38.8 32.8 § 7.8 86 5.8 32.9 § 6.4
Controls 772 45.0 29.7 § 5.8 258 15.0 26.8 § 6.6 559 32.6 26.2 § 6.0 127 7.4 29.6 § 5.2
Total
Cases 835 44.4 33.5 § 6.4 338 18.0 30.8 § 8.3 601 31.9 32.5 § 7.8 107 5.7 32.8 § 6.4
Controls 1,041 45.4 29.6 § 5.8 511 22.3 25.7 § 6.5 593 25.9 26.2 § 5.9 148 6.4 29.6 § 5.2
123

Hum Genet
controls. After adjusting for birth year and ethnicity, we foundthat the association between advanced maternal age and DSwas only present in cases of DS of maternal origin (p < 0.0001in each study sample) and was not observed in meiotic errorsof paternal origin or in post-zygotic mitotic errors (p > 0.10 foreach study sample) (Table 2). As advanced maternal age wasrestricted to cases that were due to maternal nondisjunction, wehypothesized that the proportion of maternal versus paternaland mitotic errors would diVer between the two study periods.This would be expected, as we established that the meanmaternal age increased from the earlier to the later study peri-ods (Fig. 1). Indeed, we found a signiWcantly increased propor-tion of cases of maternal origin in NDSP compared to ADSP(93.2% vs. 88.5%, p = 0.01; Table 2).
Because the proportions of ethnic groups in the ADSP/NDSP also varied over time, we examined the associationbetween ethnicity and origin of the meiotic error. Wehypothesized that any diVerence in the proportions of mater-nal errors compared to other errors could be explained bythe overall increase in the age of mothers over time, not bythe changing proportions of ethnic groups. To test this, wemodeled origin of the error (maternal vs. other errors) as theoutcome variable and ethnicity and maternal age as the inde-pendent variables and found no signiWcant diVerence in theproportion of maternal errors by ethnic group (p > 0.10).
Stage of origin of maternal meiotic error
Our next objective was to examine the eVect of maternalage on the risk for MMI and MMII errors using twodiVerent approaches. First, using logistic regression (see
“Statistical analysis”), we found a statistically signiWcantincrease in maternal age for both MMI and MMII casescompared to controls (Table 4). For example, compared tomothers of controls, mothers of infants with trisomy 21 dueto MMI nondisjunction were four times more likely to be35–39 years old than 20–24 years old at the birth of theindex case. Similarly, mothers of infants with trisomy 21due to MMII nondisjunction were Wve times more likely tobe 35–39 years old than 20–24 years old at the birth of theindex case. The ORs for being in the oldest age group, ¸40,were 8.5 and 15.1 for MMI and MMII errors, respectively.In general, the RRs tended to show a similar maternal ageeVect, although they were somewhat greater than the ORsin the older age groups.
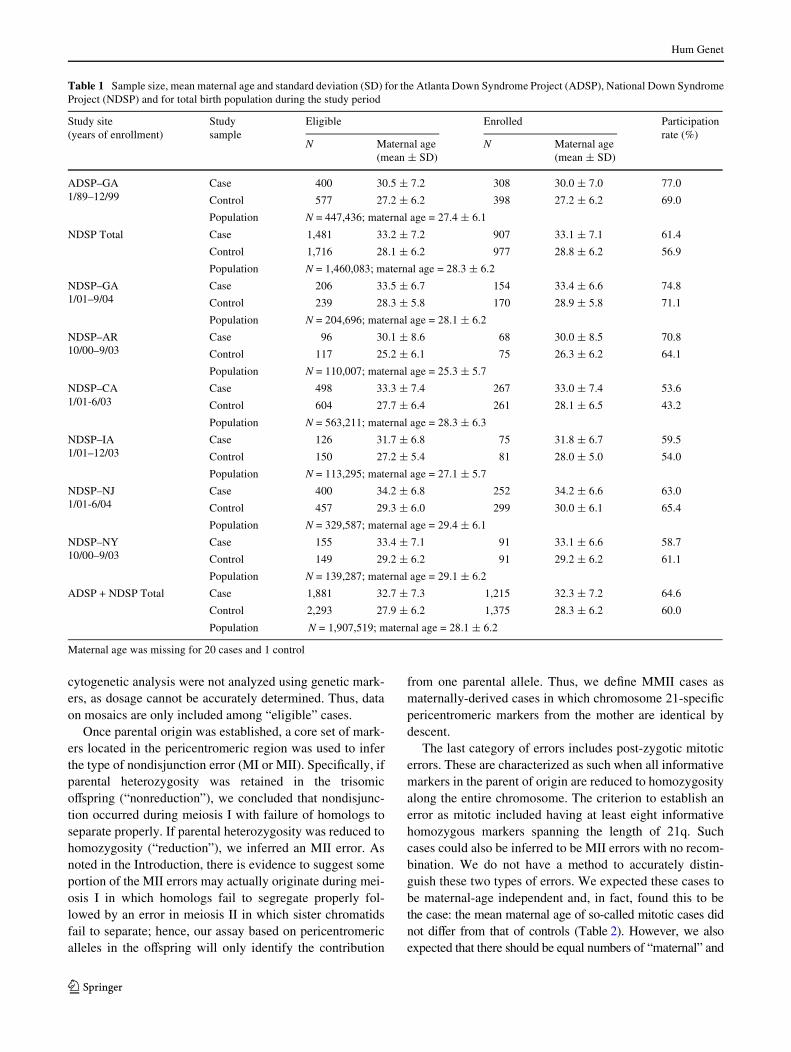
Because the ORs and RRs indicated that the frequencydistribution of maternal ages may diVer between motherswith an MMI error and those with an MMII error, our sec-ond approach was to directly compare the frequency distri-bution of maternal age for these two groups (Fig. 2).Although mean maternal age did not diVer between errortypes (Table 2), the frequency distribution did diVer signiW-cantly (�7
2 =16.6, p = 0.01; Fig. 2). By observation(Fig. 2) and by comparison of OR/RRs (Table 4), we foundthere was an increased proportion of MMII cases comparedto MMI cases among both women <15 and those 40–45 years of age. Furthermore, by simply calculating theMMI to MMII ratio in each age group without covariateadjustment, the same pattern emerged: for the six maternalage groups, ·19, 20–24, 25–29, 30–34, 35–39, and¸40 years, the ratio was 2.1, 3.8, 3.5, 4.7, 2.9, and 2.3,respectively (taken from Table 4).
Table 4 Estimated adjusted odds ratio (OR) and rate ratio (RR) for mothers of infants with trisomy 21(MMI or MMII errors) being in a speciWcage group compared to mothers of controls and the general population
Referent group are mothers age 20–24 years. Enrolled participants in ADSP and NDSP were useda Odds ratio adjusted for ethnicity/race of mother and for birth year of infantb Rate ratio adjusted for birth year of infant
Maternal cases Age group N of cases N of controls Case–control analysis Case-population analysis
ORa 95% CI RRb 95% CI
MI ·19 31 122 0.9 0.5–1.4 0.8 0.5–1.3
20–24 84 281 Ref. – Ref. –
25–29 106 355 1.0 0.7–1.4 1.0 0.8–1.4
30–34 189 396 1.6 1.2–2.2 1.9 1.4–2.4
35–39 193 170 4.0 2.8–5.5 3.8 2.9–4.9
¸40 127 51 8.5 5.6–12.9 11.6 8.8–15.3
MII ·19 15 122 1.7 0.8–3.4 1.6 0.8–3.0
20–24 22 281 Ref. – Ref. –
25–29 30 355 1.1 0.6–1.9 1.1 0.6–1.9
30–34 40 396 1.4 0.8–2.4 1.5 0.9–2.5
35–39 67 170 5.4 3.2–9.3 5.0 3.1–8.1
¸40 56 51 15.1 8.4–27.3 19.2 11.7–31.5
123
Hum Genet
Grand-maternal age eVect
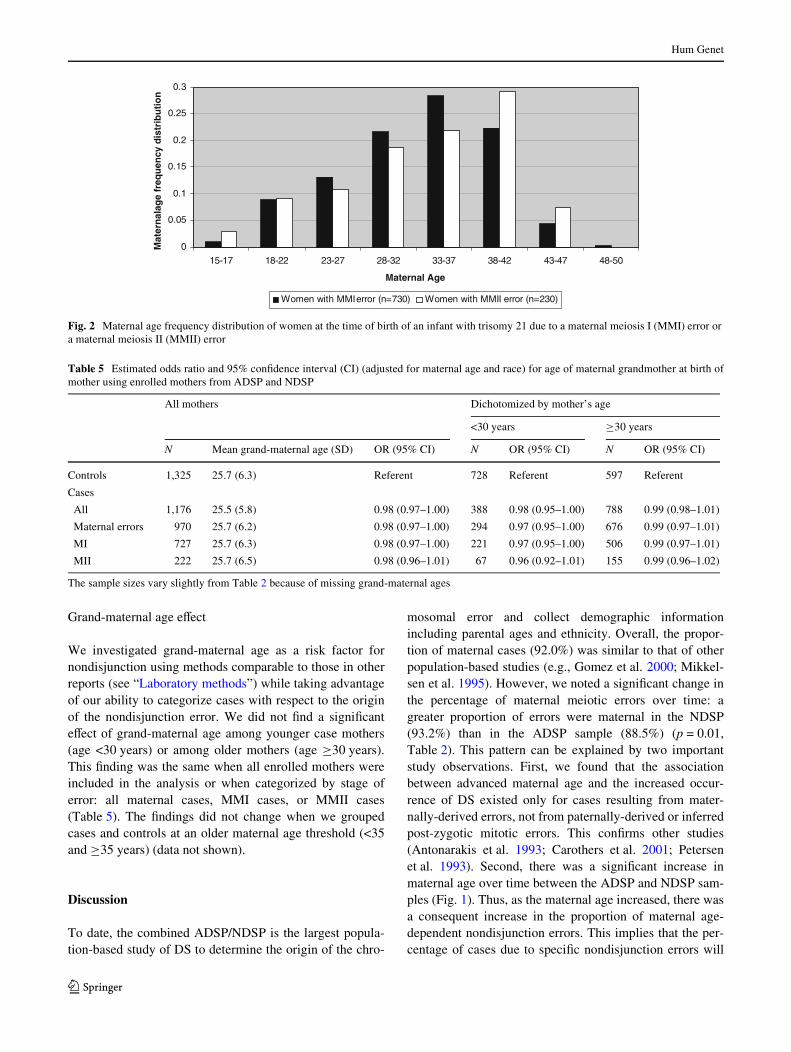
We investigated grand-maternal age as a risk factor fornondisjunction using methods comparable to those in otherreports (see “Laboratory methods”) while taking advantageof our ability to categorize cases with respect to the originof the nondisjunction error. We did not Wnd a signiWcanteVect of grand-maternal age among younger case mothers(age <30 years) or among older mothers (age ¸30 years).This Wnding was the same when all enrolled mothers wereincluded in the analysis or when categorized by stage oferror: all maternal cases, MMI cases, or MMII cases(Table 5). The Wndings did not change when we groupedcases and controls at an older maternal age threshold (<35and ¸35 years) (data not shown).
Discussion
To date, the combined ADSP/NDSP is the largest popula-tion-based study of DS to determine the origin of the chro-
mosomal error and collect demographic informationincluding parental ages and ethnicity. Overall, the propor-tion of maternal cases (92.0%) was similar to that of otherpopulation-based studies (e.g., Gomez et al. 2000; Mikkel-sen et al. 1995). However, we noted a signiWcant change inthe percentage of maternal meiotic errors over time: agreater proportion of errors were maternal in the NDSP(93.2%) than in the ADSP sample (88.5%) (p = 0.01,Table 2). This pattern can be explained by two importantstudy observations. First, we found that the associationbetween advanced maternal age and the increased occur-rence of DS existed only for cases resulting from mater-nally-derived errors, not from paternally-derived or inferredpost-zygotic mitotic errors. This conWrms other studies(Antonarakis et al. 1993; Carothers et al. 2001; Petersenet al. 1993). Second, there was a signiWcant increase inmaternal age over time between the ADSP and NDSP sam-ples (Fig. 1). Thus, as the maternal age increased, there wasa consequent increase in the proportion of maternal age-dependent nondisjunction errors. This implies that the per-centage of cases due to speciWc nondisjunction errors will
Fig. 2 Maternal age frequency distribution of women at the time of birth of an infant with trisomy 21 due to a maternal meiosis I (MMI) error ora maternal meiosis II (MMII) error
0
0.05
0.1
0.15
0.2
0.25
0.3
15-17 18-22 23-27 28-32 33-37 38-42 43-47 48-50
Maternal Age
Mat
ern
alag
e fr
equ
ency
dis
trib
uti
on
Women with MMI error (n=730) Women with MMII error (n=230)
Table 5 Estimated odds ratio and 95% conWdence interval (CI) (adjusted for maternal age and race) for age of maternal grandmother at birth ofmother using enrolled mothers from ADSP and NDSP
The sample sizes vary slightly from Table 2 because of missing grand-maternal ages
All mothers Dichotomized by mother’s age
<30 years ¸30 years
N Mean grand-maternal age (SD) OR (95% CI) N OR (95% CI) N OR (95% CI)
Controls 1,325 25.7 (6.3) Referent 728 Referent 597 Referent
Cases
All 1,176 25.5 (5.8) 0.98 (0.97–1.00) 388 0.98 (0.95–1.00) 788 0.99 (0.98–1.01)
Maternal errors 970 25.7 (6.2) 0.98 (0.97–1.00) 294 0.97 (0.95–1.00) 676 0.99 (0.97–1.01)
MI 727 25.7 (6.3) 0.98 (0.97–1.00) 221 0.97 (0.95–1.00) 506 0.99 (0.97–1.01)
MII 222 25.7 (6.5) 0.98 (0.96–1.01) 67 0.96 (0.92–1.01) 155 0.99 (0.96–1.02)
123

Hum Genet
vary with the maternal age structure of a population, andcomparisons from one study to another need to be inter-preted carefully.
The documented association between advanced maternalage and both MMI and MMII cases conWrms our previouswork (Yoon et al. 1996) and that of others (Antonarakiset al. 1992; Muller et al. 2000). Thus, the arrest of meiosisand its resumption after many years may compromise theability of the oocyte to complete both stages of meiosisproperly. Many hypotheses have been suggested to explainthe maternal age eVect and most imply an age-related deg-radation of the meiotic machinery. Recent studies haveindicated changes in gene expression in younger comparedwith older oocytes in both mouse (Hamatani et al. 2004;Pan et al. 2008) and human studies (Steuerwald et al.2007). Gene proWles that were altered by age included thoseinvolved in cell cycle regulation, cytoskeletal structure,energy pathways, transcription control, and stressresponses. Such changes could play a role in the meioticspindle abnormalities observed frequently in oocytes ofolder mothers (Battaglia et al. 1996; Eichenlaub-Ritteret al. 2004) and/or in the deterioration of sister chromatid orcentromere cohesion complexes as seen in mice by Hodgeset al. (2005). Further, checkpoint systems that monitorspindle assembly and chromosome movement may not beeVective in older oocytes (e.g., Hodges et al. 2002; LeM-aire-Adkins et al. 1997; Vogt et al. 2008).
With this large ADSP/NDSP data set, we dissected thematernal age inXuence further and found diVerences in theratio of MMI to MMII cases across the maternal age contin-uum. At all ages, MMI errors exceeded MMII errors. How-ever, the ratio of MMI to MMII was less in the youngestand the oldest maternal age groups compared with that inthe other maternal age groups. This decreased MMI toMMII ratio was particularly noticeable for women ¸40.Thus, although there are more MMI than MMII errorsacross all maternal age groups, perhaps additional factorsmore often present at the beginning and/or the end of repro-ductive life lead to an increase in meiotic errors in whichsister chromatids fail to separate properly. Hodges et al.(2002) provide strong evidence from the mouse model thatoocyte growth in an altered environment leads to anincrease in the failure of chromosomes to move toward theequator during MI (congression failure). They further showthat congression failure at MI can increase the risk for pre-mature sister chromatid segregation (PSCS) in both MI andMII. There are many factors that may be involved in thecontrol of oocyte growth. These could include the complexorchestration of signaling from the hypothalamic-pituitary-ovarian (HPO) axis as well as others involved in folliculo-genesis. Factors common to both early and late reproductivelife may involve altered hormone proWles (e.g., increasedFSH, cycle variability). Results from Hodges et al. (2002)
implicated both oocyte-somatic cell communication and analtered endocrine environment as factors that increase con-gression failure.
Studies of human oocytes are also consistent with anincreased maternal age being associated with both MMIand MMII errors (for review, see Pellestor et al. 2005). In astudy of 309 karyotypically abnormal human oocytesobserved at meiosis II metaphase, Pellestor et al. (2003)identiWed both whole chromosome nondisjunction andPSCS. Interestingly, they found that, during meiosis I, sin-gle chromatid aneuploidy occurred more frequently thandid whole chromosome aneuploidy among the 309 oocytesand had a stronger correlation with maternal age. PSCS atmeiosis I could lead to the error being classiWed as MMI orMMII depending on the action of the chromatids. From ouranalyses, we cannot determine the underlying mechanismfor meiosis II errors. That is, we cannot distinguish MIIerrors that result from PSCS at meiosis I, whole chromo-some nondisjunction at meiosis I followed by a reductionaldivision at meiosis II or a “classical” meiosis II error inwhich chromatids fail to separate properly after completinga successful meiosis I division. Maternal-age risk factorsare most likely associated with one or more of these mecha-nisms.
Our recent data that examined recombination proWlesalong nondisjoined chromosomes 21 by type of nondisjunc-tion error and maternal age provide additional insight intomechanisms underlying nondisjunction. These studies wereperformed on a subset of cases from the population-basedstudies presented here (Lamb et al. 2005; Oliver et al.2008). Among MMI cases, we found that the majority ofnondisjoined chromosomes 21 were associated with eithera lack of an exchange or a telomeric exchange and thatthese patterns inXuenced the risk for nondisjunction irre-spective of maternal age. In contrast, we found that the non-disjoined chromosomes 21 that were categorized as MMIIerrors and had a pericentromeric exchange were enrichedamong older women with this type of error. These data sug-gested a maternal age-dependent mechanism (Oliver et al.2008). In Oliver et al., we oVered two alternative explana-tions for this observation: (1) a pericentromeric exchangeinitiates or exacerbates the susceptibility to maternal agerisk factors, perhaps leading to an increase in PSCS, or (2) apericentromeric exchange protects the bivalent against age-related risk factors allowing proper segregation of homo-logues at meiosis I, but not segregation of sisters at meiosisII. The former explanation would represent a two-hitmodel: the Wrst hit being the pericentromeric recombinantevent and the second hit would involve any number of mei-otic-related structures or proteins that degrade with oocyteage (Lamb et al. 1996). The latter explanation implies thattrue MII errors may occur among older women only if biv-alents are protected from age-related factors in some way.
123

Hum Genet
This protective factor could be a proximal recombinantevent which then allows the sister cohesion complex toremain intact along most of the chromosome arm. Otherprotective factors could include genetic variants that reduceage-related degradation of meiotic structures or environ-mental factors that create an optimal environment duringthe arrested state of the oocyte, to name a few.
Lastly, we tested the hypothesis that the age of thematernal grandmother of the child with trisomy 21 aVectsthe risk for a nondisjunction error. Results from past studiesare conXicting, potentially due to diVerences in design andsampling strategies (Aagesen et al. 1984; Greenberg 1963;Malini and Ramachandra 2006; Papp et al. 1977; Penrose1964; Richards 1970; Stoller and Collmann 1969). How-ever our failure to Wnd a relationship between nondisjunc-tion and grand-maternal age is strong evidence against suchan eVect for the following reasons: we had adequate num-bers of cases and controls representing the same popula-tions in the same time frames, we documented standardtrisomy 21 by karyotype and included only maternally-derived cases in the comparison.
Although this study has many strengths, it is importantto outline its limitations. First, due to limited resources, theNDSP could only recruit mothers who spoke either Englishor Spanish. Further, case and control families whose infantdied or was placed for adoption prior to enrollment werenot recruited. These factors probably have limited impacton the results of this study. The most important limitationwas that we were not able to include pregnancies with tri-somy 21 that were either spontaneously lost or terminated.We included only live births which represent no more than10–20% of conceptions with trisomy 21(for review, seeHassold and Hunt 2001), thus, our results must be inter-preted with this in mind. For example, we discovered thatthe mean age of mothers at the time of birth increased overthe time period of the study. Martin et al. (2005) presenteda similar increase in mean maternal age based on NationalVital Statistics data. We found this increase occurred inmothers of both cases and controls; although the slope ofthe increase for cases was steeper than that for controls.The steeper increase in maternal age of infants with trisomy21 from 1989–2004 could be due to increased prenatalscreening being oVered to younger women (maternal serumscreening) beginning in the mid 1990s. It is possible thatpositive screens among these women led to conWrmatorytesting and pregnancy termination, thus reducing the pro-portion of mothers eligible for our study. Although onlyspeculation, there may have been increased acceptance ofan infant with trisomy 21, perhaps more often among olderwomen. Potentially, this could also inXuence this increasedslope. Nevertheless, these inXuences should not aVect ourinterpretation of the comparison of MI to MII errors, aswomen are blind to the type of nondisjunction error. The
participation rates varied by study site; however, there wereno signiWcant diVerences in mean maternal age for enrolledand non-enrolled cases or controls. Thus, the associationswith maternal age should be representative of the popula-tion of eligible live-born cases.
In summary, in this large population-based study, wehave conWrmed our previous Wndings (Yoon et al. 1996).SpeciWcally, the signiWcant association between advancedmaternal age and chromosome 21 nondisjunction wasrestricted to errors in the egg; the association was notobserved in paternal or in post-zygotic mitotic errors. Fur-ther, an almost three-fold higher proportion of MMI errorsover MMII errors is present at all maternal ages; however,we note that this ratio decreases for mothers <19 years andthose ¸40 years at the time of their infant’s birth. The nextlogical step will be to use both origin of the meiotic errorand recombination proWles along the nondisjoined chromo-somes 21 to classify types of nondisjunction errors.Although such parameters are still only surrogates for theexact type of error, they will provide more homogeneousgroups in which to detect maternal-age associated risk fac-tors.
Acknowledgments We gratefully acknowledge the many familiesnationwide whose participation has made this study possible. In addi-tion, we want to thank all personnel at each NDSP site and their asso-ciated birth surveillance teams who made this project a success. Lastly,we would like to thank Larry Edmonds and Dr. Paula Yoon who sharedtheir experience with the National Birth Defects Prevention Study.Their help was invaluable. This work was supported by NIH R01HD38979 and by the technical assistance of the General Clinical Re-search Center at Emory University (NIH/NCRR M01 RR00039).
References
Aagesen L, Grinsted J, Mikkelsen M (1984) Advanced grandmaternalage on the mother’s side—a risk of giving rise to trisomy 21. AnnHum Genet 48:297–301
Antonarakis SE, Avramopoulos D, Blouin JL, Talbot CC Jr, SchinzelAA (1993) Mitotic errors in somatic cells cause trisomy 21 inabout 4.5% of cases and are not associated with advanced mater-nal age. Nature Genet 3:146–150
Antonarakis SE, Petersen MB, McInnis MG, Adelsberger PA, Schin-zel AA, Binkert F, Pangalos C, Raoul O, Slaugenhaupt SA, HafezM (1992) The meiotic stage of nondisjunction in trisomy 21:determination by using DNA polymorphisms. Am J Hum Gen50:544–550
Battaglia DE, Goodwin P, Klein NA, Soules MR (1996) InXuence ofmaternal age on meiotic spindle assembly in oocytes from natu-rally cycling women. Human Reprod 11:2217–2222
Book JA, Fraccaro M, Lindsten J (1959) Cytogenetical observations inMongolism. Acta Paediatr 48:453–468
CanWeld MA, Honein MA, Yuskiv N, Xing J, Mai CT, Collins JS, De-vine O, Petrini J, Ramadhani TA, Hobbs CA, Kirby RS (2006)National estimates and race/ethnic-speciWc variation of selectedbirth defects in the United States, 1999–2001. Birth Defects ResA Clin Mol Teratol 76:747–756
Carothers AD, Castilla EE, Dutra MG, Hook EB (2001) Search for eth-nic, geographic, and other factors in the epidemiology of Down
123

Hum Genet
syndrome in South America: analysis of data from the ECLAMCproject, 1967–1997. Am J Med Genet 103:149–156
Correa-Villasenor A, Cragan J, Kucik J, O’Leary L, SiVel C, WilliamsL (2003) The metropolitan Atlanta congenital defects program:35 years of birth defects surveillance at the centers for diseasecontrol and prevention. Birth Defects Res A Clin Mol Teratol67:617–624
de Bruin JP, Dorland M, Spek ER, Posthuma G, van Heaften M, Loo-man CW, Te Velde ER (2004) Age-related changes in the ultra-structure of the resting follicle pool in human ovaries. BiolReprod 70:419–424
Eichenlaub-Ritter U, Boll I (1989) Nocodazole sensitivity, age-relatedaneuploidy, and alterations in the cell cycle during maturation ofmouse oocytes. Cytogenet Cell Genet 52:170–176
Eichenlaub-Ritter U, Vogt E, Yin H, Gosden R (2004) Spindles, mito-chondria and redox potential in ageing oocytes. Reprod BiomedOnline 8:45–58
Ford CE, Jones KW, Miller OJ, Mittwoch U, Penrose LS, Ridler M,Shapiro A (1959) The chromosomes in a patient showing bothMongolism and the Klinefelter syndrome. Lancet 1:709–710
Freeman SB, Allen EG, Oxford-Wright CL, Tinker SW, Druschel C,Hobbs CA, O’Leary LA, Romitti PA, Royle MH, Torfs CP, Sher-man SL (2007) The National Down Syndrome Project: design andimplementation. Public Health Rep 122:62–72
Freeman SB, Yang Q, Allran K, Taft LF, Sherman SL (2000) Womenwith a reduced ovarian complement may have an increased riskfor a child with Down syndrome. Am J Hum Genet 66:1680–1683
Gaulden ME (1992) Maternal age eVect: the enigma of Down syn-drome and other trisomic conditions. Mutat Res 296:69–88
Gomez D, Solsona E, Guitart M, Baena N, Gabau E, Egozcue J, Cabal-lin MR (2000) Origin of trisomy 21 in Down syndrome casesfrom a Spanish population registry. Ann Genet 43:23–28
Greenberg RC (1963) Two factors inXuencing the births of Mongols toyounger mothers. Med OV 109:62–64
Hamatani T, Falco G, Carter MG, Akutsu H, Stagg CA, Sharov AA,Dudekula DB, VanBuren V, Ko MS (2004) Age-associated alter-ation of gene expression patterns in mouse oocytes. Human MolGenet 13:2263–2278
Hassold T, Chiu D (1985) Maternal age-speciWc rates of numericalchromosome abnormalities with special reference to trisomy.Hum Genet 70:11–17
Hassold T, Hunt P (2001) To err (meiotically) is human: the genesis ofhuman aneuploidy. Nature Rev Genet 2:280–291
Hodges CA, Ilagan A, Jennings D, Keri R, Nilson J, Hunt PA (2002)Experimental evidence that changes in oocyte growth inXuencemeiotic chromosome segregation. Hum Reprod 17:1171–1180
Hodges CA, Revenkova E, Jessberger R, Hassold TJ, Hunt PA (2005)SMC1beta-deWcient female mice provide evidence that cohesinsare a missing link in age-related nondisjunction. Nat Genet37:1351–1355
Jacobs PA, Baikie AG, Court Brown WM, Strong JA (1959) The so-matic chromosomes in Mongolism. Lancet 1:710
Lamb NE, Freeman SB, Savage-Austin A, Pettay D, Taft L, Hersey J,Gu Y, Shen J, Saker D, May KM, Avramopoulos D, Petersen MB,Hallberg A, Mikkelsen M, Hassold TJ, Sherman SL (1996) Sus-ceptible chiasmate conWgurations of chromosome 21 predisposeto non-disjunction in both maternal meiosis I and meiosis II. Na-ture Genet 14:400–405
Lamb NE, Yu K, ShaVer J, Feingold E, Sherman SL (2005) Associa-tion between maternal age and meiotic recombination for trisomy21. Am J Hum Genet 76:91–99
Lejeune J (1959) Le Mongolism. Premier exemple d’aberration autos-omique humaine. Annals of Genetics 1:41–49
LeMaire-Adkins R, Radke K, Hunt PA (1997) Lack of checkpoint con-trol at the metaphase/anaphase transition: a mechanism of meiotic
nondisjunction in mammalian females. J Cell Biol 139:1611–1619
Malini SS, Ramachandra NB (2006) InXuence of advanced age ofmaternal grandmothers on Down syndrome. BMC Med Genet 7:4
Martin JA, Hamilton BE, Sutton PD, Ventura SJ, Menacker F, MunsonML (2005) Births: Wnal data for 2003. Natl Vital Stat Rep54(2):1–116
Mikkelsen M, Hallberg A, Poulsen H, Frantzen M, Hansen J, PetersenMB (1995) Epidemiology study of Down’s syndrome in Den-mark, including family studies of chromosomes and DNA mark-ers. Develop Brain Dysfunct 8:4–12
Muller F, RebiVe M, Taillandier A, Oury JF, Mornet E (2000) Parentalorigin of the extra chromosome in prenatally diagnosed fetal tri-somy 21. Hum Genet 106:340–344
Mutton D, Alberman E, Hook EB (1996) Cytogenetic and epidemio-logical Wndings in Down syndrome, England and Wales 1989 to1993. National Down syndrome Cytogenetic Register and theAssociation of Clinical Cytogeneticists. J Med Genet 33:387–394
Oliver TR, Feingold E, Yu K, Cheung V, Tinker S, Yadav-Shah M,Masse N, Sherman SL (2008) New insights into human nondis-junction of chromosome 21 in oocytes. PLoS Genet 4:e1000033
Pan H, Ma P, Zhu W, Schultz RM (2008) Age-associated increase inaneuploidy and changes in gene expression in mouse eggs. DevBiol 316:397–407
Papp Z, Varadi E, Szabo Z (1977) Grandmaternal age at birth of par-ents of children with trisomy 21. Hum Genet 39:221–224
Pellestor F, Anahory T, Hamamah S (2005) EVect of maternal age onthe frequency of cytogenetic abnormalities in human oocytes. Cy-togenet Genome Res 111:206–212
Pellestor F, Andreo B, Arnal F, Humeau C, Demaille J (2002) Mecha-nisms of non-disjunction in human female meiosis: the co-existenceof two modes of malsegregation evidenced by the karyotyping of1397 in-vitro unfertilized oocytes. Hum Reprod 17:2134–2145
Pellestor F, Andreo B, Arnal F, Humeau C, Demaille J (2003) Maternalaging and chromosomal abnormalities: new data drawn from invitro unfertilized human oocytes. Hum Genet 112:195–203
Penrose LS (1933) The relative eVects of paternal and maternal age inMongolism. J Genet 27:219–224
Penrose LS (1934) The relative aetiological importance of birth order andmaternal age in Mongolism. Proc R Soc B Biol Sci 115:431–450
Penrose LS (1964) Genetical aspects of mental deWciency. Proceed-ings of the international Copenhagen congress on the scientiWcstudy of mental retardation, pp 165–172
Petersen MB, Antonarakis SE, Hassold TJ, Freeman SB, ShermanSL, Avramopoulos D, Mikkelsen M (1993) Paternal nondis-junction in trisomy 21: excess of male patients. Human MolGenet 2:1691–1695
Richards BW (1970) Observations on mosaic parents of mongol prop-ositi. J Ment DeWc Res 14:342–346
Schon EA, Kim SH, Ferreira JC, Magalhaes P, Grace M, Warburton D,Gross SJ (2000) Chromosomal non-disjunction in human oo-cytes: is there a mitochondrial connection? Hum Reprod 15(Suppl2):160–172
Sherman SL, Freeman SB, Allen EG, Lamb NE (2005) Risk factors fornondisjunction of trisomy 21. Cytogenet Genome Res 111:273–280
Steuerwald NM, Bermudez MG, Wells D, Munne S, Cohen J (2007)Maternal age-related diVerential global expression proWles ob-served in human oocytes. Reprod Biomed Online 14:700–708
Steuerwald NM, Steuerwald MD, Mailhes JB (2005) Post-ovulatoryaging of mouse oocytes leads to decreased MAD2 transcripts andincreased frequencies of premature centromere separation andanaphase. Mol Hum Reprod 11:623–630
Stoller A, Collmann RD (1969) Grandmaternal age at birth of mothersof children with Down’s syndrome (ONGOLISM). J Ment DeWcRes 13:201–205
123

Hum Genet
van Montfrans JM, van HooV MH, Martens F, Lambalk CB (2002)Basal FSH, estradiol and inhibin B concentrations in women witha previous Down’s syndrome aVected pregnancy. Human Reprod17:44–47
Vogt E, Kirsch-Volders M, Parry J, Eichenlaub-Ritter U (2008) Spin-dle formation, chromosome segregation and the spindle check-point in mammalian oocytes and susceptibility to meiotic error.Mutat Res 651:14–29
Warburton D (2005) Biological aging and the etiology of aneuploidy.Cytogenet Genome Res 111:266–272
Yang Q, Sherman SL, Hassold TJ, Allran K, Taft L, Pettay D, KhouryMJ, Erickson JD, Freeman SB (1999) Risk factors for trisomy 21:
maternal cigarette smoking and oral contraceptive use in a popu-lation-based case–control study. Genet Med 1:80–88
Yoon PW, Freeman SB, Sherman SL, Taft LF, Gu Y, Pettay D, Flan-ders WD, Khoury MJ, Hassold TJ (1996) Advanced maternal ageand the risk of Down syndrome characterized by the meiotic stageof chromosomal error: a population-based study. Am J Hum Gen-et 58:628–633
Yusuf RZ, Naeem R (2004) Cytogenetic abnormalities in products ofconception: a relationship revisited. Am J Reprod Immunol52:88–96
123