Embed Size (px)
Citation preview

Marine Pollution Bulletin 61 (2010) 309–322
Contents lists available at ScienceDirect
Marine Pollution Bulletin
journal homepage: www.elsevier .com/locate /marpolbul
Biogeochemical typology and temporal variability of lagoon waters in a coralreef ecosystem subject to terrigeneous and anthropogenic inputs (New Caledonia)
R. Fichez a,e,*, S. Chifflet a,e, P. Douillet a,e, P. Gérard b,e, F. Gutierrez c, A. Jouon b, S. Ouillon d, C. Grenz a,e
a IRD, Universidad Autonoma Metropolitana Iztapalapa, DCBS, Departamento de Hidrobiología, Av. San Rafael Atlixco 186, Col. Vicentina C.P. 09340, Iztapalapa, México D.F., Mexicob Centre IRD de Nouméa, BPA5, 98848 Nouméa, New Caledoniac Universidad Autonoma Metropolitana Iztapalapa, DCBS, Departamento de Hidrobiologia, Av. San Rafael Atlixco 186, Col. Vicentina C.P. 09340, Iztapalapa, México D.F., Mexicod Université de Toulouse, IRD, UMR LEGOS, 14 Av. Edouard Belin, 31400 Toulouse, Francee Université de la Méditerranée, UMR CNRS 6535 LOPB, Rue de la Batterie des Lions, 13007 Marseille, France
a r t i c l e i n f o a b s t r a c t
Keywords:Coral reef lagoonNutrientsHydrologyTypologyTemporal variabilityTrophic status
0025-326X/$ - see front matter � 2010 Elsevier Ltd. Adoi:10.1016/j.marpolbul.2010.06.021
* Corresponding author at: IRD, Universidad AutonoDCBS, Departamento de Hidrobiología, Av. San Rafael A09340, Iztapalapa, México D.F., Mexico. Tel.: +52 58 047 38.
E-mail address: [email protected] (R. Fichez).
Considering the growing concern about the impact of anthropogenic inputs on coral reefs and coral reeflagoons, surprisingly little attention has been given to the relationship between those inputs and the tro-phic status of lagoon waters. The present paper describes the distribution of biogeochemical parametersin the coral reef lagoon of New Caledonia where environmental conditions allegedly range from pristineoligotrophic to anthropogenically influenced. The study objectives were to: (i) identify terrigeneous andanthropogenic inputs and propose a typology of lagoon waters, (ii) determine temporal variability ofwater biogeochemical parameters at time-scales ranging from hours to seasons. Combined ACP-clusteranalyses revealed that over the 2000 km2 lagoon area around the city of Nouméa, ‘‘natural” terrigeneousversus oceanic influences affecting all stations only accounted for less than 20% of the spatial variabilitywhereas 60% of that spatial variability could be attributed to significant eutrophication of a limited num-ber of inshore stations. ACP analysis allowed to unambiguously discriminating between the natural tro-phic enrichment along the offshore–inshore gradient and anthropogenically induced eutrophication.High temporal variability in dissolved inorganic nutrients concentrations strongly hindered their useas indicators of environmental status. Due to longer turn over time, particulate organic material and morespecifically chlorophyll a appeared as more reliable nonconservative tracer of trophic status. Results fur-ther provided evidence that ENSO occurrences might temporarily lower the trophic status of the NewCaledonia lagoon. It is concluded that, due to such high frequency temporal variability, the use of biogeo-chemical parameters in environmental surveys require adapted sampling strategies, data managementand environmental alert methods.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
The impact of anthropogenically enhanced runoff and chemicalinputs to the coastal zone is acknowledged as a worldwide scale is-sue (see reviews in Sheppard (2000)). Coral reef environments arestrongly affected by global change and due to prevailing oligo-trophic conditions they are commonly considered as sensitiveenvironments easily altered by even moderate anthropogenic in-puts (Adey et al., 2000). As a consequence, community structurealterations observed during the past two decades have often beenempirically related to anthropogenic alteration of environmentalconditions. Considering this repeated claim, the paucity of existing
ll rights reserved.
ma Metropolitana Iztapalapa,tlixco 186, Col. Vicentina C.P.
4 46 00x2754; fax: +52 58 04
recent papers dealing with the spatial and temporal variability ofhydrology in coral reef lagoons is anachronistic. Moreover, mostof the available recent studies on lagoon water chemical statusdeal with atoll reef environments subject to insignificant anthro-pogenic inputs (Charpy-Roubaud et al., 1990; Delesalle and Sour-nia, 1992; Charpy et al., 1997; Le Borgne et al., 1997; Pages et al.,2001; Dufour et al., 2001) whereas environmental assessmentstudy of barrier reef lagoons exposed to significant terrigeneousand anthropogenic inputs prove to be surprisingly scarce (Erfte-meijer and Herman, 1994; Morrison and Naqasima, 1999; Muslimand Jones, 2003). Despite the high number of publications dealingwith coastal environment alterations, few really focused on thedistribution and temporal variability of biogeochemical parame-ters in the water (Larned, 1998; Liston et al., 1992; Muslim andJones, 2003) and a lot of datasets have been published and dis-cussed in books or grey literature reports (Haynes et al., 2001;Furnas, 2003). The Great Barrier Reef region received some signif-icant attention on environmental status assessment and temporal

310 R. Fichez et al. / Marine Pollution Bulletin 61 (2010) 309–322
variability (Devlin and Brodie, 2005; Fabricius et al., 2005; Mosset al., 2005; Udy et al., 2005; Woodridge et al., 2006; Brodieet al., 2007) and similar efforts yielding information on environ-mental typology has to be applied to other reef areas worldwide.Such an evolution is urgently needed as relationship between coralreef health and terrigeneous or anthropogenic inputs have oftenbeen stated when proper information on environmental statuswas so evidently missing.
As part of a several year programme studying environmental is-sues in Pacific high island coral reef lagoons, an intensive study onwater biogeochemistry was conducted to precisely define environ-mental status in the southern part of the New Caledonia west coastlagoon close to the city of Nouméa. The objectives defined were:
� To define the distribution of biogeochemical parameters in alagoon subject to combined oceanic, terrigeneous and anthro-pogenic influences.� To propose a simplified typology of lagoon waters.� To assess the magnitude of temporal variability of water bio-
geochemical parameters at time-scales ranging from hours toseasons considering that this aspect has rarely been investi-gated despite the major constraints it exerts on the implemen-tation of environmental surveys (see also Le Borgne et al.(2010); Torreton et al. (2010)).
2. Material and methods
2.1. Study site
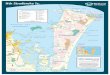
New Caledonia is a French overseas country (Jost, 1998) locatedin the Pacific Ocean between latitudes 19� and 23� south and lon-gitudes 163� and 168� east, just to the north of the Tropic of Cap-ricorn, (Fig. 1). The coral barrier reef of New Caledonia delimits alarge lagoon. Water temperature in the lagoon tends to be 1–2 �Clower in winter and 1–2 �C higher in summer than in the ocean
Fig. 1. Study site – location and bathymetric map of the study site in the
(Rougerie, 1986; Le Borgne et al., 2010). Salinity is generally closeto ocean salinity but in protected bays it may decrease due to occa-sional rainfalls or increase during period of droughts which areespecially pronounced during ENSO events (Ouillon et al., 2005,2010). On the southern part of the west coast where the studywas conducted, rainfalls are moderate and it is extremely rare forestuarine plumes to extend as far as the reef which is more than10 km away from the coast (Labrosse et al., 2000). The semi-diur-nal tide, with amplitudes of 1.5 m at spring tide and 0.3 m at neaptide, propagates from the south to the north and its influence onlagoon circulation is largely depending on the geomorphology(Douillet, 1998). Trade winds favour rapid renewal rate of lagoonwater and recent hydrodynamic modelling work calculated anaverage residence time of no more than 11 days for the south-westpart of the lagoon that covers an area of 2000 km2 (Bujan et al.,2000; Jouon et al., 2006; Ouillon et al., 2010). Lagoon water qualityhence is considered as chiefly determined by ocean water inflow,land originating inputs and anthropogenic inputs mainly originat-ing from industrial and waste water discharges and mining activi-ties (Labrosse et al., 2000).
2.2. Sampling
The sampling was conducted on a total of 23 cruises coveringthe 1997–2001 period (Table 1) and over 33 stations located alongland-reef transects were sampled from March 1997 to November1999. This data set was mainly used to establish seasonal variabil-ity for a limited number of stations typical of the main lagoon sub-systems. In complement, surveys were conducted over periods oftime of 1–7 days to assess high frequency variability. Finally, astronger sampling effort was invested in September 2000 (87 sam-pling stations) and March 2001 (86 sampling stations) duringcruises dedicated to assessing the distribution of chemical param-eters at the scale of the whole lagoon.
lagoon of New Caledonia, distribution of cruises sampling stations.

Table 1Time table for the sampling cruises conducted over 5 years in the south-west lagoonof New Caledonia. T = temporal survey of 32 sampling stations; HR-S = high resolu-tion survey of 89 sampling stations.
1997 1998 1999 2000 2001
January T TFebruary TMarch T T HR-SApril T TMay T TJune TJuly TAugust T TSeptember T HR-SOctober T TNovember T T TDecember T T
R. Fichez et al. / Marine Pollution Bulletin 61 (2010) 309–322 311
2.3. CTD profiles
Salinity profiles were obtained from a SeaBird SBE 19 CTDequipped with additional turbidity (Seapoint optical back scatter)and in situ fluorescence (Wet Lab Wetstar) sensors. Salinity wasobtained with a precision of 0.001 while calibrated back scatteringturbidity sensors provided NTU data with a precision of 0.1 NTU. Incoral reef lagoons, where various origins of particles provide a mul-ti-modal particle size distribution, it has been demonstrated thatback scattering is linearly correlated to suspended particulate mat-ter concentrations of less than 10 g L�1 with a correspondence of1 NTU for 1 mg with negligible variability between individualnephelometers (Larcombe et al., 1995; Bunt et al., 1999; Ouillonet al., 2004). In situ fluorescence is presented as arbitrary fluores-cence units with a precision of 0.01 units. The correspondencefrom in situ fluorescence to chlorophyll a (R2 > 0.7) or pheopigment(R2 > 0.5) concentrations was calculated using coincidental discretesub-surface sampling, sampled water filtration on Whatman GF/Ffilters and in vitro fluorescence measurements.
2.4. Particulate and dissolved material
Discrete water samples were taken using a Niskin bottle at 3 mdepth. Samples for the fluorometric determination of ammonia(Kérouel and Aminot, 1997; Holmes et al., 1999) were taken as soonas the sampling bottle was retrieved and the reagent was instantlyinjected onboard in order to limit sample contamination samplesfor the analysis of other dissolved nutrients were stored unfilteredin the deep-freeze. Samples for the determination of particulate or-ganic material were filtered on Whatman GF/F filters and stored inthe deep-freeze. Nitrate + nitrite (reported as NO3
�) concentrationswere determined at nanomolar concentrations (Raimbault et al.,1990), phosphate (PO4
3�) concentrations were determined accord-ing to Murphy and Riley (1962) and silicates (Si(OH)4) according toFanning and Pilson (1973) all analyses being conducted on a Bran-Luebbe III continuous flow autoanalyzer. Total dissolved nitrogen(DN) and phosphorus (DP) concentrations were determines as ni-trate and phosphate concentrations after wet oxidation of watersamples. Particulate and dissolved organic nitrogen (PON) andphosphorus (POP) were analysed as nitrate and phosphate aftersubjecting loaded filters to wet oxidation (Raimbault et al., 1990).Active chlorophyll a and pheopigment concentrations were deter-mined by the in vitro fluorometric technique (Lorenzen, 1966).
2.5. Data analysis
Data interpretation called for a reliable multi-parametricapproach of the complex hydrochemical data set generated by
the two cruises of September 2000 and March 2001. Our goalwas to identify the main sources of influence, assess the existingcorrelations between environmental variables and establish a sta-tistically supported water typology. Data analysis was conductedusing a combination of principal component analysis (PCA) andcluster analysis (Legendre and Legendre, 1998). The PCA was con-ducted on the correlation matrixes (i.e. standardized variables) of12 variables (Sal, turb, Cha, Pheo, NH4
+, NO3�, PO4
3�, Si(OH)4, DN,DP, POP, and PON) versus 87 (September 2000) or 82 (March2001) sampling stations. Additional classification was obtainedby conducting an agglomerative cluster analysis on the correlationmatrixes resulting from the PCA (Webster and Oliver, 1991; Lebartet al., 1995). Combined PCA and cluster analysis classification re-sults were plotted on biplots representing the two first principalcomponent axes PC1 and PC2 as they jointly accounted for morethan 70% of spatial variability. Indexes of average water residencetime such as ‘‘water age” or ‘‘local e-flushing time” were calculatedfor each sampling station and used in the PCA analysis as explan-atory variables.
3. Results and discussion
3.1. Distribution of environmental variables
The distribution of sub-surface (2–4 m depth averaged) turbid-ity in September 2002 (Fig. 2a) showed highest suspended loads inthe Bay of Sainte-Marie, and the Bay of Dumbea, maximum valuesof more than 3.5 NTU being reached in the inner parts of KoutioBay and Grande Rade. Turbidity levels of more than 2.5 NTU wererecorded in Boulari Bay close to the estuary of La Coulée River butno significant turbidity increase was observed at the mouth of LesPirogues River. Turbidity of less than 0.5 were recorded in mostpart of the middle lagoon except at station M38 in the back reefarea were an average turbidity of 1.2 NTU was recorded. This valuehowever only corresponded to a 3 m surface layer lying over a20 m deep layer of water with a turbidity of 0.5 NTU. Turbidity lev-els in March 2001 (Fig. 2e) were comparable to those measured inSeptember 2000 except for a peak over 4 NTU in the north-westback reef area and at the mouth of Les Pirogues River. In this lattercase it is important to note that, due to the low sampling resolutionin this area, data interpolation tends to overestimate the spreadingof the actual plume.
In September 2000 and March 2001, chlorophyll a concentra-tions (Fig. 2b and f) of more than 0.8 lg L�1 were measured in Sain-te-Marie Bay as well as in Grande Rade and Koutio Bay, the twourbanised embayments in Dumbea Bay. Lower chlorophyll concen-trations were measured at the mouth of rivers (Port Laguerre, LaCoulée, Les Pirogues) suggesting moderate nutrient inputs fromthe river. Higher concentrations (0.4–0.8 lg L�1) recorded closeto the Dumbea estuary was very likely to originate from the exten-sion of Koutio Bay inputs. Chlorophyll a concentrations wereslightly higher in March 2001 than in September 2000 in the mid-dle lagoon with concentrations between 0.2 and 0.5 lg L�1. Thechlorophyll a versus pheopigment ratio was remarkably constantat 1.1 with a standard deviation of 0.12 in September and at 1.34with a standard deviation of 0.36 in March with no significantmodification in this average ratio even under chloropigmentenrichment conditions.
Whatever the period, PON (Fig. 2c and g) and POP (Fig. 2d and h)distributions were very similar to the distribution of chlorophyll a.In September 2000 concentrations in PON ranged between 0.5 and0.9 lM in the lagoon and increased to more than 1.9 lM in theinner part of the three urbanised bays located around the city ofNouméa. Similarly, POP concentrations ranged between 0.02 and0.06 lM in the lagoon and increased to more than 0.16 lM in the

Fig. 2. Spatial distribution of particulate material – distribution of turbidity, chlorophyll a, particulate organic nitrogen (PON) and phosphorus (POP) in the New Caledonialagoon around the city of Nouméa during two cruises conducted in September 2000 and March 2001.
312 R. Fichez et al. / Marine Pollution Bulletin 61 (2010) 309–322

Fig. 3. Spatial distribution of dissolved material – distribution of ammonia (NH4+), nitrate + nitrite (NO3), dissolved total nitrogen (DN) and silicate (Si(OH)4) in the New
Caledonia lagoon around the city of Nouméa during two cruises conducted in September 2000 and March 2001.
R. Fichez et al. / Marine Pollution Bulletin 61 (2010) 309–322 313

314 R. Fichez et al. / Marine Pollution Bulletin 61 (2010) 309–322
inner parts of the three bays. In March 2001, PON and POP concen-trations in the middle lagoon were globally higher than in Septem-ber 2000 commonly reaching values between 0.7 and 1.5 lM and0.04 and 0.08 lM for PON and POP, respectively. Similarly, concen-trations in the coastal urbanised bays were higher in March 2001than in September 2000. A slight increase in POP and PON concen-trations was measured in the north-west part of the lagoon whencompared with the south-east part, a situation that could be due toa conjunction of higher water residence time and inputs of com-bined terrigeneous and anthropogenic inputs.
During the September 2000 cruise, ammonia concentrations(Fig. 3A) of more than 0.7 lM were measured at the four innersampling points in Sainte-Marie Bay and in the very inner part ofKoutio Bay. NH4
+ concentrations rapidly decreased to less than0.2 lM in the south-east part of the lagoon and to less than0.3 lM in its north-west part. The situation in March 2001(Fig. 3E) was entirely different as NH4
+ concentrations were below0.2 lM through the whole lagoon except for a small increase to0.3 lM in the inner part of Boulari Bay.
In September 2000, highest nitrate concentrations (Fig. 3B)were measured in Sainte-Marie Bay and in the inner part of Dum-bea Bay including Koutio Bay and the waters around the mouth ofDumbea River. High concentrations were measured at two sam-pling sites in the north-west back reef area, hence far away from
Table 2Characteristics of the first five components of the principal component analysis (PCA)on the standardized 12 variables � 87 sampling points matrix for seawater param-eters in the south-west lagoon of New Caledonia for September 2000 and March 2001.
Component rank 1 2 3 4 5
September 2000Eigenvalue 7.55 1.86 0.87 0.66 0.39Proportion of variance (%) 62.92 15.5 7.29 5.52 3.21Cumulative proportion (%) 62.92 78.43 85.71 91.24 94.45
March 2001Eigenvalue 7.86 1.30 0.82 0.74 0.38Proportion of variance (%) 65.49 10.85 6.86 6.19 3.20Cumulative proportion (%) 65.49 76.35 83.21 89.39 92.59
-0.8 -0.4 0 0.4 0.8
-0.8
-0.4
0
0.4
0.8
Sal
NO3
NH4PO4TurbChl
POPPheo
NOPDP
Si(OH)4
DN
Factor 2 - 15.5 %
Factor 1 - 62.9 %
September 2000
Water local e-flushing time
Water age
a
Fig. 4. Principal component analysis variables – representation of the variables as a funcMarch 2001 sampling cruises.
any source of terrigeneous or anthropogenic inputs. NO3� concen-
trations were below 0.03 lM in most other parts of the lagoon. InMarch, NO3
� concentrations (Fig. 3F) of more than 0.1 lM weremeasured in Koutio Bay. Outside this bay, NO3
� concentrationswere mostly below 0.04 lM except for the middle part of the la-goon where concentrations of more than 0.08 lM were recordedat three stations with a peak at 0.12 lM at station M33.
The distribution pattern of total dissolved nitrogen (DN) in Sep-tember 2000 (Fig. 3C) and March 2001 (Fig. 3G) was very similar tothe distribution of NH4
+ especially when considering the discrep-ancy between the south-east and north-east part of the lagoon inSeptember 2000 and the strong global impoverishment in March2001.
Si(OH)4 concentrations in the middle lagoon were mostly be-tween 1 and 2 lM in September 2000 (Fig. 3A) and increased tovalues ranging between 3 and 5 in March 2001. Significant coastalincrease in Si(OH)4 concentrations to values of more than 6 lMwere exclusively measured in Dumbea and Boulari Bays, the twopartially sheltered bays subject to significant river inputs. Riversare major providers of silica to the lagoon as Si(OH)4 concentra-tions of more than 400 lM were measured in the freshwaters ofthe three main rivers. The March 2001 cruise occurred during therainy season and after significant rainfalls and the joint decreasein salinity and increase of Si(OH)4 concentrations in lagoon watersis a clear consequence of river inputs.
3.2. Water typology and main influencing factors
The results from a principal component analysis (PCA) com-bined to a cluster analysis provided a synthetic classification ofthe water bodies in response to the respective importance of themain sources of influence. Results from the PCA (Table 2) showedaxes 1 and 2 accounted for 78% and 74% of the total variability inSeptember 2000 and March 2001, respectively, and hence couldbe unambiguously considered as the two main structuring factors.
In September 2000 (Fig. 4a), axis 1, which accounted for 63% ofthe variability alone, was correlated in decreasing order with: chlo-rophyll a and pheopigments (r = 0.95); POP (r = 0.93); turbidity,
Factor 2 - 10.9 %
Factor 1 - 65.5 %-0.8 -0.4 0 0.4 0.8
0
0.4
0.8
DN DP
SalNH4
NO3
Si(OH)4
Turb
PO4POP
Chl
PheoPON
March 2001
-0.4
-0.8
Water local e-flushing time
Water age
b
tion of axes 1 and 2 from the principal component analysis for September 2000 and

R. Fichez et al. / Marine Pollution Bulletin 61 (2010) 309–322 315
PO43� and PON (r = 0.92), turbidity (r = 0.90) and NH4
+ (r = 0.87).Being essentially correlated with trophic status parameters, axis1 could be considered as indicative of an enrichment gradient fromoligotrophic on the negative side to eutrophic on the positive side.Axis 2 which accounted for 15.5% of the variability was correlatedin decreasing order with: salinity (r = 0.82); DN (r = �0.62); sili-cates (r = �0.57) and NO3
� (r = 0.54). Axis 2 could be related toan oceanic versus river gradient from oceanic salted waters onthe positive end to slightly desalted waters with increased riverborne silica, DN, and NO3
� on the negative end. It must be men-tioned here that inverse salinity gradients might occasionally de-velop inshore when strong draught conditions associated toENSO events occur.
In March 2001 (Fig. 4b), axis 1 which accounted for 63.9% of thevariability alone was positively (0.84 < r < 0.95) correlated with:PO4
3�, PON, POP, DP, chlorophyll a, pheopigments and turbidity,
Fig. 5. Water typology – results from the combined principal component analysis and clu
again indicating an enrichment gradient from oligotrophic on thenegative end to eutrophic on the positive end. Axis 2 which ac-counted for 10.2% of the variability was correlated with NO3
�
(r = �0.71), silicates (r = �0.51) and salinity (r = 0.44), indicating agradient from oceanic salted waters on the positive end to slightlydesalted waters and increased river borne Si and N on the negativeend. The PCA distribution of environmental variables was stronglyconvergent for both sampling occurrence and also converged withresults from a recent PCA analysis conducted on environmentalvariables in the Great Barrier Reef region (Fabricius et al., 2005)that identified two major influencing factors strongly comparableto those identified in the New Caledonia lagoon.
The combined PCA/cluster analysis for September 2000 (Fig. 5a)allowed for the distinction of six classes. Class 1, 2 and 4, respec-tively accounting for 41, 25 and 6 stations over a total of 87 sta-tions (88%) sampled during the cruise, were disposed along a
ster analysis for September 2000 and March 2001 sampling cruises whole data set.

316 R. Fichez et al. / Marine Pollution Bulletin 61 (2010) 309–322
diagonal axis (grossly from stations A03 to D48). The sampling sta-tions gathered in class 1 were essentially located in the south-wes-tern offshore part of the lagoon and close to reef passages,indicating a major oceanic influence. Stations from class 4 wereexclusively located in the inshore part of the bay of Dumbea andtheir PCA projection is a consequence of: (i) a negative correlationwith axis 2 corresponding to fresh water inputs from the DumbeaRiver, (ii) a positive correlation with axis 1 corresponding to amoderate enrichment in trophic status. Stations from class 2 whichwere distributed in-between were located in the north-easternpart of the lagoon, in the open bay of Boulari and in the entranceof other bays, indicating intermediate conditions. This diagonalordination therefore can be clearly attributed to a natural contin-uum between oceanic and inshore (freshwater + terrigeneousnutrients) sources of influence corresponding to an inland agingof the water bodies as shown by the water residence time indexesused as explanatory variables (Fig. 4a) (see also Ouillon et al.(2010)). Therefore, even though the biogeochemical compositionof 88% of the water samples collected in the lagoon was exclusivelycommanded by this oceanic to inshore continuum, which thereforeappeared as a driving constrains acting at the scale of the whole la-goon, it only explained for a modest fraction of the overall spatialvariability. Class 3, 5 and 6 respectively gathered 8, 5 and 2 sam-pling stations that strongly departed from the previously identifiedoceanic to inshore continuum. Such a distribution mainly resultedfrom a strong positive correlation with axis 1 corresponding toenrichment in trophic status. The distribution of Sainte-MarieBay stations (prefix N) along the trophic gradient was consistentwith their location along the entrance (N28) to the inner part(N04 and N05) of the bay transect. Stations N04 and N05 locatedin the most inshore part of Sainte-Marie Bay could be consideredas strongly influenced by anthropogenic eutrophication due to in-puts from poorly to untreat waste water discharges. Finally, fouramong the five stations that formed class 5 were located in Dum-bea Bay close to the city of Nouméa and the Dumbea river estuarytherefore experiencing combined terrigeneous and anthropogenicinputs.
The combined PCA/cluster analysis for March 2001 (Fig. 5b)showed a slightly higher dispersion of the data set along axis 1and yielded a six classes typology very comparable to September2000 typology. Class 1, 2 and 4 with 45, 11 and 4 stations, respec-tively, over a total of 86 stations (70%) were organised along a diag-onal offshore versus inshore continuum similar to the oneidentified in September 2000. Class 3, 5 and 6 respectively gath-ered 18, 3 and 1 stations, respectively, strongly departed fromthe offshore versus inshore continuum due to a significant influ-ence of the eutrophication factor, with stations from the inshoreparts of Sainte-Marie Bay (N04 and N05) and Koutio Bay (D64and D65) again appearing as the most strongly impacted.
The typology strongly converged for the two sampling occa-sions even though they corresponded to very different seasonswith significant changes in the absolute value of each parameter.The most important feature in that typology relates to the fact thata very limited number of artificially eutrophized stations (12–30%)accounted for most (�80%) of the spatial variability. Scientistsworking in meso-eutrophic temperate marine environments mightconsider the observed absolute level of trophic enrichment asinsignificant but our results demonstrate that even when deliveredin moderate amount nutrient anthropogenic inputs generate a verysignificant departure from natural equilibrium hence playing adeterminant role in structuring environmental conditions andassociated biota responses. Such a major specificity of eutrophica-tion processes in coral reef oligotrophic ecosystems still has to befully perceived by the scientific community as well as by environ-mentally concerned decision makers. Additionally, the typologypermitted to identify key variables that were most closely linked
to the two major influencing factors and could be specifically stud-ied in term of temporal variability. With a variable versus factorcorrelation of more than 0.9 in both occasions against axis 1, chlo-rophyll a unambiguously stood as a good indicator of trophic sta-tus. With a variable versus factor correlation of 0.7 in September2000 and 0.44 in March 2001 against axis 2, salinity stood as oneof the best indicator of oceanic versus freshwater influences, sili-cates also appearing as a potentially valuable indicator of such aninfluence. Beside those two essential indicators, variability of someother parameters commonly used in environmental studies (tur-bidity, inorganic nutrients, and particulate organic matter) wasalso investigated.
3.3. Vertical structure
High resolution information on the vertical distribution of keyparameters (temperature, salinity, turbidity, and in situ chloropig-ment related fluorescence) in the water column is a prerequisite toassess the representativity of discrete water sampling. Comparisonof profile measured at different time required the conversion ofdata to relative data expressed as
½X�rd ¼ ½X�md=V ½X�prof ð1Þ
where ½X�rd was the calculated relative concentration at depth d,½X�md was the measured concentration at depth d and ½X�prof wasthe average concentration over the whole vertical profile, verticalheterogeneity being evidenced by the divergence from a ratio valueof 1. Profiles recorded at a single sampling station during the wholesurvey were plotted together as a function of depth (Fig. 6). Onlythree data sets are presented corresponding to stations that wereidentified previously as representative of oceanic influence (stationsA03), intermediate lagoon (station D41), and eutrophic areas (N04)groups.
Temperature and salinity displayed a very moderate heteroge-neity with insignificant stratification. Turbidity distributionshowed frequent occurrence of a significant bottom nepheloidlayer reaching values of 4–6 times the profile averaged turbidityvalue and generally limited to a few meters above the benthicboundary layer. In the inner and outer lagoon stations, a log-linearregression provided the best fit with R2 values of 0.24 and 0.45,respectively. For chloropigments, despite occasional surface max-ima, the most common trend observed in the outer and intermedi-ate lagoon corresponded to an increase with depth inchloropigment concentrations. Curve fit from log-linear and linearregressions yielded very similar results. This vertical trend was notobserved at inshore stations certainly because shallow water con-ditions did not allow for a significant vertical gradient. Consideringthe weak density stratification and the prevailing euphotic condi-tions (nonpresented irradiance profiles), the increase in chloropig-ment concentration with depth could be explained by photo-inhibition in the upper layers and/or nutrient inputs from the ben-thic system. Very little is known about photo-inhibition but thatprocess would not be of significance beyond a few meter depths.Furthermore, the second hypothesis is strongly supported by re-cent work on benthic metabolism and biogeochemical modellingin the New Caledonia lagoon that concluded that pelagic primaryproduction was strongly controlled by benthic recycling and nutri-ent release in the water column (Bujan et al., 2000; Grenz et al.,2003; Pinazo et al., 2004).
3.4. Temporal variability
The study of short term temporal variability combining verticalprofiling and discrete water sampling was first conducted on se-lected typical sampling stations with a 30 min frequency over peri-

0
10
20
30
40
0,9 1 1,1
0
10
20
30
40
0,9 1 1,1
y = 12,4Ln(x) + 19,02R2 = 0,4538
0
10
20
30
40
0 1 2 3 4 5 6
y = 24,7Ln(x) + 18,39R2 = 0,5347
0
10
20
30
40
0 1 2 3
0
10
20
30
40
0,9 1 1,1
0
10
20
30
40
0,9 1 1,1
y = 7,74Ln(x) + 10,78R2 = 0,2411
0
10
20
30
40
0 1 2 3 4
y = 22,17Ln(x) + 10,315R2 = 0,4005
0
10
20
30
40
0 1 2
0
10
20
30
40
0,9 1 1,1
0
10
20
30
40
0,9 1 1,1
0
10
20
30
40
0 1 2 3 4 5
0
10
20
30
40
0 1 2
Temperature / profile averaged temperature
Salinity / profile averaged salinity
Turbidity / profile averaged turbidity
Chloropigments / profile averaged chloropigments
Outer lagoon A03
Inner lagoon D41
Eutrophic bay N04
Fig. 6. Vertical distribution – vertical distribution of temperature, salinity, turbidity and chloropigments calculated from in situ fluorescence. Values correspond to the ratio ofraw data at a specific depth versus data averaged over the whole profile (see Eq. (1) in the text). Profiles recorded at a single sampling station on each occasions during the5 years survey are plotted together as a function of depth. Significant best fit linear and log-linear regression curves and equations are reported.
R. Fichez et al. / Marine Pollution Bulletin 61 (2010) 309–322 317
ods of 25 h (Fig. 7). Little variability was observed in the middle la-goon (stations M33 and A24), demonstrating day/night trends to
be insignificant. However, the occasional passage of 3–5 m thicksub-surface lenses of slightly desalted and chloropigment enriched

D47D47
M33
A24
N04
M33
A24
Elapsed time (h) Elapsed time (h)
Dep
th (
m)
Dep
th (
m)
Dep
th (
m)
Dep
th (
m)
0 5 10 15 20 25
-10
-8
-6
-4
-2
0 .30 .40 .50 .60 .70 .80 .91 .0
N04 35.90
35.80
35.85
-10
-8
-6
-4
-2
-10
-8
-6
-4
-2
0 5 10 15 20 25
0.700.650.600.550.500.450.400.350.30
35.7035.6535.6035.5535.5035.4535.40
0 5 10 15 20 250 5 10 15 20 25
-5
-10
-15
-20
-5
-10
-15
-20
0 5 10 15 20 250 5 10 15 20 25
-10
-8
-6
-4
-2
0.05
0.04
0.03
0.02
0.01
35.60
35.58
35.56
-5
-10
-15
-20
-25
-5
-10
-15
-20
-25
0 5 10 15 20 25 0 5 10 15 20 25
0.21
0.19
0.17
0.15
0.13
0.11
0.09
0.07
0.05
0.03
35.38
35.36
35.34
35.32
35.30
35.28
35.26
35.24
35.22
Fig. 7. Analysis of high frequency temporal variability – interpolated plot of chloropigment (in situ fluorometry derived) and salinity profiles (30 min frequency) over 25 htime periods in four typical sampling stations (beware of different color/value scales). Black bars represent obscurity periods (irradiance < 1 lE m�2 s�1). Station A24 = outerlagoon (September 2000), station M33 = middle lagoon (June 2003), station D47 = inner bay (October 1998), and station N04 = eutrophic bay (October 1998).
318 R. Fichez et al. / Marine Pollution Bulletin 61 (2010) 309–322
water were observed on eight occurrences over 25 h at the mostoffshore station (A24). Previous results from hydrodynamic model-ling strongly suggest that surface desalted water lenses could orig-inate from the southern part of New Caledonia mainland (Douillet,1998; Douillet et al., 2001), their conservation as identifiablehydrologic subsystem being only possible under conditions ofmoderate wind stress and wave energy. The represented situationhowever can be considered as infrequent as the outer lagoon isgenerally subject to sustained trade winds resulting in strong
hydrodynamic conditions, rapid renewal of water and water col-umn homogeneity (Bujan et al., 2000; Douillet et al., 2001; Pinazoet al., 2004). Inshore, stations D47 in front of the Dumbea riverestuary and N04 in the inner part of eutrophized Sainte-MarieBay exhibited a much stronger temporal variability. In DumbeaBay, the influence of Dumbea River (0.5 m3 s�1 on average) gener-ated some moderate variations in salinity with no evident link to aday/night cycle while chloropigments clearly increased frommorning to twilight and decreased from twilight to morning. In

R. Fichez et al. / Marine Pollution Bulletin 61 (2010) 309–322 319
Sainte-Marie Bay salinity varied moderately due to the absence ofsignificant fresh water inputs (0.15 m3 s�1 on average) but the day/night cycle of chloropigment biomass was strongly marked espe-cially at depth with a doubling in concentrations at the end ofthe afternoon. As bays appeared more subject to short term vari-ability, some additional information was obtained by deployingmoored buoys recording environmental parameters at 3 m depthwith a 10 min frequency and over periods of several days (Fig. 8).At station D47, in front of the Dumbea River, we observed signifi-cant decreases in salinity with a 12 h period during the 2 first daysfollowed by a long period with lower and more erratic variations.The 12 h period clearly related to the influence of spring tide, min-imum salinity values being reached at low tide (nonpresentedpressure data). Turbidity irregularly increased from 1 to 9 NTUduring the 2 first days corresponding to the low salinity pulsesand decreased within a 1–3 NTU range afterward. Chloropigmentconcentrations were around 0.35 lg L�1 during the initial periodof salinity pulses and high turbidity and increased to a maximumof 0.8 lg L�1 afterward with hourly variations reaching a threefoldfactor. At station N20 in Sainte-Marie Bay, rapid salinity variationsof ca. 0.15 did not directly correlated with the bi-diurnal tidal cyclebut most likely revealed current driven local movement of waterbodies. Turbidity and chloropigment concentrations varied veryrapidly in the range 1–2.5 NTU and 0.35–0.97 lg L�1, respectively,both parameters showing no correlation with salinity. That intenseshort term variability of environmental parameters in inshore coralreef lagoon waters (see also Ouillon et al. (2010)) was interpretedas a direct consequence of the increase in spatial variability in themost wind sheltered coastal areas (Figs. 2 and 3). In those places,slow water renewal and significant terrestrial and anthropogenic
34.8
34.9
35.0
35.1
35.2
35.3
35.4
35.5
Salinity
0
2
4
6
8
10
Turbidity(NTU)
0.00
0.02
0.04
0.06
0.08
0.10
Time
Fluorometry
15/06 17/06 19/06 21/06 23/06
15/06 17/06 19/06 21/06 23/06
15/06 17/06 19/06 21/06 23/06
Station D47
Fig. 8. Analysis of high frequency temporal variability – mooring records (10 min frequenriver inputs (Dumbea Bay, station D46) and in a bay subject to eutrophication (Sainte-M
inputs generated strong gradients on short distances and even alimited displacement of water masses proved to generate signifi-cant short term eulerian variability.
The analysis of monthly variability of environmental variableswas conducted over the 1997–1999 period on a total of 32 sam-pling stations (Table 1) temporal variability trends between sta-tions with very different average concentration levels werecompared by considering relative concentrations according to:
½X�rt ¼ ½X�mt=½X�surv ð2Þ
where ½X�rt was the calculated relative concentration at samplingtime t, ½X�mt was the measured concentration at sampling time tand ½X�surv was the average concentration over the whole profile,temporal variability being evidenced by the divergence from a ratiovalue of 1. Results from the sampling stations classified as middlelagoon from the PCA typology (class 1, n = 14) were plotted andcompared to data from stations influenced by terrigeneous (D47)and anthropogenic (D01 and N04) inputs (Fig. 9). Only chlorophylla, ammonia and nitrates were retained as they were identified bythe PCA as the most structuring non conservative variables. In themiddle lagoon, chlorophyll a showed a relatively unambiguous sea-sonal variability with a maximum value at the end of May, hencejust before reaching minimum water temperature (southern hemi-sphere), corresponding to minimum values in NH4
+ (recycled N) andmaximum values in NO3
� (new N). Such a seasonal trend in chloro-phyll a concentrations has already been reported for the New Cale-donia lagoon (Binet and Le Borgne, 1996; Le Borgne et al., 2010) aswell as in the oceanic province around New Caledonia (Dandonneauand Gohin, 1984) the later author attributing the phytoplanktonbloom to the rise of the nutricline due to winter cooling of surface
35.3
35.4
35.5
35.6
Salinity
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Turb
idity
(NTU
)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
22/06 24/06 26/06 28/06
22/06 24/06 26/06 28/06
22/06 24/06 26/06 28/06
Time
Fluo
rom
etry
Station N20
cy) of salinity, turbidity and in situ fluorescence at 3 m depth in a bay influenced byarie Bay, station N20) in June 2001.

A
0,0
1,0
2,0
3,0
4,0
27/09/1997
26/11/1997
25/01/1998
26/03/1998
25/05/1998
24/07/1998
22/09/1998
21/11/1998
20/01/1999
0
1
2
3
4
Lagoon
D47
D01
N04
B
0,0
1,0
2,0
3,0
4,0
27/09/1997
26/11/1997
25/01/1998
26/03/1998
25/05/1998
24/07/1998
22/09/1998
21/11/1998
20/01/1999
LagoonD47D01N04
C
0
1
2
3
4
5
6
7
18/08/1997
17/10/1997
16/12/1997
14/02/1998
15/04/1998
14/06/1998
13/08/1998
12/10/1998
11/12/1998
09/02/1999
Lagoon
D47
D01
N04
Rel
ativ
e [C
hlor
ophy
ll a]
(L
agoo
n, D
47, D
01)
Rel
ativ
e [C
hlor
ophy
ll a]
(N
04)
Rel
ativ
e [N
H4]
Rel
ativ
e [N
O3]
Fig. 9. Analysis of low frequency temporal variability trends – distribution of relative (see Eq. (2) in the text) chlorophyll a (A), NH4+ (B), and NO3
� (C) over the period October1997–January 1999. Average values + standard deviation from middle lagoon stations are plotted together with values for stations singled out by the principal componentanalysis as significantly impacted by anthropogenic inputs (Fig. 5).
Table 3Comparison of short term versus long term variability using variation percentage(100 � standard deviation/average) calculated from 24 h surveys with a hourlysampling frequency and 1 year survey with a monthly sampling frequency in threespecific lagoon areas as defined from Fig. 5.
Lagoon Terrigeneous EutrophicA03/A24 D47 N04
Day Year Day Year Day Year
NH4+ (lM) 72.92 96.33 nd 55.58 55.58 75.93
NO3� (lM) 27.94 114.87 58.78 157.90 65.08 131.55
PO43� (lM) 21.11 78.04 28.26 67.65 47.16 76.41
Si (lM) 52.51 20.56 35.34 52.39 61.22 57.09DN (lM) 10.45 12.52 7.99 12.85 13.63 15.34DP (lM) 18.40 27.96 10.33 32.60 29.84 37.48PON (lM) 7.22 30.39 8.75 51.94 19.00 36.10POP (lM) 10.65 81.76 8.07 38.57 20.96 51.24POC (lM) 24.79 16.18 12.77 25.13 18.94 37.80Chlorophyll a 19.15 51.06 7.90 55.83 21.07 98.21
320 R. Fichez et al. / Marine Pollution Bulletin 61 (2010) 309–322
waters. Even though that hypothesis might appear to be supportedby the observed concurrent increase in chlorophyll a and NO3
�
(Fig. 9b and c), the occurrence of a similar cycle in NO3� concentra-
tions in estuarine systems with ambient concentrations severalfolds higher than in the middle lagoon cast some doubts on its al-leged oceanic origin. The detailed analysis of such a process wasnot within the scope of this article but the study of the forces driv-ing such an unusual seasonal trend should require some futureattention as it might relate to the coincident occurrence of episodesof calm weather allowing for the build up of phytoplankton bio-mass. That seasonal trend was also observed for stations D47 andD01 but with some significantly higher variability especially atD01. Variability at station N04 was important and did not matchthe seasonal trend observed for other stations. In that later case,the relatively constant daily supply of anthropogenic nutrientapparently results in a non season-dependent variability. Chloro-phyll a concentrations during the 1998–1999 warm seasons weresignificantly higher than during the 1997–1998 warm seasons. Sucha difference could be related to the occurrence of a severe El Niñoepisode in 1997 resulting in a very dry period in the south-westernpart of the Pacific Ocean, the return to normal conditions resultingin strong rainy episodes in January 1999. The 1997 ENSO eventproved to have a detectable impact on temperature and salinityvariations in the New Caledonia lagoon (Ouillon et al., 2005). Thegeneral trophic level of the lagoon is naturally low due to its rapidflushing by oligotrophic oceanic waters (Jouon et al., 2006) but ishowever sustained by terrigeneous inputs. In such an environmen-tal context, the occurrence of El Niño events provoking severe peri-ods of droughts in the western part of the Pacific Ocean mightsignificantly lower the inputs of terrigeneous nutrients hencedecreasing phytoplankton nutrient sources. Beyond the direct effecton plankton population, such a decrease would result in the foodlimitation of lagoon suspension feeders. Such a stressing factorwould combine with other stresses that have been listed as poten-tial triggering factors to coral bleaching and the various epidemiccrises recorded in coral reef environments during ENSO events(Lafferty et al., 2004)
Using the variation coefficient (100 � standard deviation/aver-age) to compare hourly versus monthly variability (Table 3)showed that the daily variability was generally close to monthlyvariability for dissolved material and significantly lower thanmonthly variability for particulate material, a result that mostlyconverges with the reported pattern of phytoplankton in theNew Caledonia lagoon (Torreton et al., 2010). Those results demon-
strate that the variability at various time-scales of biogeochemicalparameters is a constraining factor that must be properly assessedwhen designing environmental monitoring projects.
4. Conclusions
Under pristine conditions, waters in the lagoon of New Caledo-nia would be organised along a continuum gradient between twosources: (i) water with an oceanic signature and (ii) coastal wateridentified by slightly lower salinities and higher Si(OH)4 concentra-tions together with a modest enrichment in nutrients and particu-late organic material. The Dumbea Bay combining long waterresidence time together with significant river inputs clearly standat the extreme end of the terrigeneous influence gradient. Resultsshowed that such a natural continuum would generate low vari-ability in water chemistry and trophic status, most of the outer la-goon stations experiencing very similar environmental conditions.That absence of a strong environmental constraining gradient inthe outer lagoon could be considered as a factor favouring theexpression of a strong biodiversity as spatially heterogeneous fac-tors mostly related to habitat complexity could express themselvesas the main environmental constraints hence yielding strong biotaspatial variability.
Apart from this naturally occurring gradient, the parametersmeasured allowed for the unambiguous identification of an

R. Fichez et al. / Marine Pollution Bulletin 61 (2010) 309–322 321
anthropogenically driven eutrophication gradient mostly related tountreated sewage release. Eutrophication hotspots were con-strained to urbanised embayments such as Koutio Bay, GrandeRade and Sainte-Marie Bay, where the most specific signatureswere measured. The multivariate analysis of the dataset clearly al-lowed discriminating between natural eutrophication of agedwater bodies and additional eutrophication due to anthropogenicinputs. Using such data treatment approaches is essential to accu-rately interpreting biota response to alleged environmental stress,a step which has already been applied to the New Caledonia lagoon(Dumas et al., 2007; Sasal et al., 2007). It is also important to stressthat eutrophication does not relate to bulk concentrations but is adynamic process leading toward the increase from an initial tro-phic status. In this regard, eutrophication of oligotrophic coral reeflagoons must be fully acknowledged as a very specific process inwhich even slight increases in trophic status may result in very sig-nificant shifts in the structure and composition of pelagic primaryproducers (Jacquet et al., 2006) consecutively impacting the wholebiogeochemical cycle.
Except for canyons where a sharp stratification in two distinctlayers might occasionally occur due to open ocean water intrusion,water bodies from the open lagoon were generally feebly stratifiedwith moderate to no temperature and salinity gradients. Chloro-phyll a concentrations did not always obey such a vertical stabilityrule as concentrations often significantly increased with depth,certainly as a consequence of nutrient release at the water–sedi-ment interface. That clearly indicated a strong relationship be-tween pelagic production and benthic recycling. Salinity andtemperature gradients were more pronounced inshore, in bayswhere calmer hydrodynamic and wave regimes prevailed, butdue to their shallowness no clear chlorophyll a vertical gradientcould be observed.
Short term (�hourly) variability was low in the outer part of thecoral reef lagoon but strongly increased inshore mainly due to thedisplacement of water bodies with contrasted biogeochemical sig-natures. Hourly variability was generally close to monthly variabil-ity for dissolved material and lower than monthly variability forparticulate material. Despite such variability, we observed that se-vere dry weather conditions due to the 1997 El Niño significantlydepleted the lagoon trophic status. Therefore, ENSO might beresponsible for food limitation of lagoon suspension feeders, a fac-tor that could play an additional role in coral bleaching and thevarious epidemic crises recorded in coral reef environments duringENSO events.
Finally, the general strong significance of short term variabil-ity has rarely been documented and represents a factor thatneed to be seriously assessed before any conclusion on environ-mental degradation could be drawn. On an environmental man-agement point of view, such variability represents a stronglimitation to the definition of reliable biogeochemical indicatorsof environmental status and the implementation of environmen-tal monitoring programmes in coral reef lagoons would requirethree major steps: (i) selecting biogeochemical parametersadapted to environmental stress, (ii) establishing a proper defini-tion of local guidelines for each selected biogeochemical param-eter, (iii) defining alert/control strategies adapted to evidencedtemporal variability.
Acknowledgements
This work was financially supported by the French Institut deRecherche pour le Développement (IRD) and by the French Pro-gramme National Environnement Côtier (PNEC). We express spe-cial thanks to the staff of IRD Research Vessels Alis, Coris andAldric and to the technical staff of the IRD Centre in Nouméa.
References
Adey, W.H., McConnaughey, T.A., Small, A.M., Spoon, D.M., 2000. Coral reefs:endangered, biodiverse, genetic resources. In: Sheppard, C. (Ed.), Seas at theMillennium, An Environmental Evaluation, vol. 3. Elsevier, Amsterdam, pp. 33–42.
Binet, D., Le Borgne, R., 1996. La station côtière de Nouméa dix ans d’observationssur l’hydrologie et le pelagos du lagon sud-ouest de Nouvelle-Calédonie.Archives Sciences de la Mer Biologie Marine n�2, Editions IRD, Nouméa, p. 37.
Brodie, J., De’ath, G., Devlin, M., Furnas, M., Wright, M., 2007. Spatial and temporalpatterns of near-surface chlorophyll a in the Great Barrier Reef lagoon. Mar.Freshwater Res. 58, 1–12.
Bujan, S., Grenz, C., Fichez, R., Douillet, P., 2000. Evolution saisonnière du cyclebiogéochimique dans le lagon sud-ouest de Nouvelle-Calédonie. Applicationd’un modèle compartimental. C.R. Acad. Sci. Paris, Life Sci., Ecol. 323, 225–233.
Bunt, J.A.C., Larcombe, P., Jago, C.F., 1999. Quantifying the response of opticalbackscatter devices and transmissometers to variations in suspendedparticulate matter. Cont. Shelf Res. 19, 1199–1220.
Charpy, L., Dufour, P., Garcia, N., 1997. Particulate organic matter in sixteenTuamotu atoll lagoons (French Polynesia). Mar. Ecol. Progr. Ser. 151, 55–65.
Charpy-Roubaud, C.J., Charpy, L., Crémoux, J.L., 1990. Nutrient budget of thelagoonal waters in an open central South Pacific atoll (Tikehau, Tuamotu,French Polynesia). Mar. Biol. 107, 67–73.
Dandonneau, Y., Gohin, F., 1984. Meridional and seasonal variations of the seasurface chlorophyll concentration in the southwestern tropical Pacific (14 to32�S, 160 to 175�E). Deep Sea Res. 31, 1377–1393.
Delesalle, B., Sournia, A., 1992. Residence time of water and phytoplankton biomassin coral reef lagoons. Cont. Shelf Res. 12, 939–949.
Devlin, M.J., Brodie, J., 2005. Terrestrial discharge into the Great Barrier Reef lagoon:nutrient behavior in coastal waters. Mar. Pollut. Bull. 51, 9–22.
Douillet, P., 1998. Tidal dynamics of the south-west lagoon of New Caledonia:observations and 2D numerical modelling. Oceanol. Acta 21, 69–79.
Douillet, P., Ouillon, S., Cordier, E., 2001. A numerical model for fine suspendedsediment transport in the southwest lagoon of New Caledonia. Coral Reefs 20,361–372.
Dufour, P., Andréfouët, S., Charpy, L., Garcia, N., 2001. Atoll morphometry controlslagoon nutrient regime. Limnol. Oceanogr. 46, 456–461.
Dumas, P., Kulbicki, M., Fichez, R., Chifflet, S., Ferraris, J., 2007. Environmentalfactors influencing urchin spatial distributions on New Caledonian disturbedcoral reefs. J. Exp. Mar. Biol. Ecol. 344, 88–100.
Erftemeijer, P.L.A., Herman, P.M.J., 1994. Seasonal changes in environmentalvariables, biomass, production and nutrient contents in two contrastingtropical intertidal seagrass beds in South Sulawesi, Indonesia. Oecologia 99,45–59.
Fabricius, K., De’ath, G., McCook, L., Turak, E., Williams, D.McB., 2005. Changes inalgal, coral and fish assemblages along water quality gradients on the inshoreGreat Barrier Reef. Mar. Pollut. Bull. 51, 384–398.
Fanning, O., Pilson, M.E.Q., 1973. On the spectrophotometric determination ofdissolved silica in natural waters. Anal. Chem. 45, 136–140.
Furnas, M., 2003. Catchments and Corals: Terrestrial Runoff to the Great BarrierReef. Australian Institute of Marine Science and CRC Reef Research Centre,Townsville. p. 334.
Grenz, C., Denis, L., Boucher, G., Chauvaud, L., Clavier, J., Fichez, R., Pringault, O.,2003. Spatial variability in sediment oxygen consumption under winterconditions in a lagoonal system in New Caledonia (South Pacific). J. Exp. Mar.Biol. Ecol. 285–286, 33–47.
Haynes, D., Brodie, J., Christie, C., Devlin, M., Michalek-Wagner, K., Morris, S.,Ramsay, M., Storrie, J., Waterhouse, J., Yorkston, H., 2001. Great Barrier ReefWater Quality: Current Issues. Great Barrier Reef Marine Park Authority,Townsville.
Holmes, R.M., Aminot, A., Kérouel, R., Bethanie, A., Hooher, A., Peterson, B.J., 1999. Asimple and precise method for measuring ammonium in marine and freshwaterecosystems. Can. J. Aquat. Sci. 56, 1801–1808.
Jacquet, S., Delesalle, B., Torréton, J.P., Blanchot, J., 2006. Response of phytoplanktoncommunities to increased anthropogenic influences (southwestern lagoon, NewCaledonia). Mar. Ecol. Progr. Ser. 320, 65–78.
Jost, C., 1998. The French-speaking Pacific: Population, Environment andDevelopment Issues. Géopacifique Association, Nouméa. p. 271.
Jouon, A., Douillet, P., Ouillon, S., Fraunié, P., 2006. Calculations of hydrodynamictime parameters in a semi-opened coastal zone using a 3D hydrodynamicmodel. Cont. Shelf Res. 26, 1395–1415.
Kérouel, R., Aminot, A., 1997. Fluorometric determination of ammonia in sea andestuarine waters by direct segmented flow analysis. Mar. Chem. 57, 265–275.
Labrosse, P., Fichez, R., Farman, R., Adams, T., 2000. New Caledonia. In: Sheppard, C.(Ed.), Seas at the Millenium, An Environmental Evaluation, vol. 2. Elsevier,Amsterdam, pp. 723–736.
Lafferty, K.D., Porter, J.W., Ford, S.E., 2004. Are diseases increasing in the ocean?Annu. Rev. Ecol. Evol. Syst. 35, 31–54.
Larcombe, P., Ridd, P.V., Wilson, B., Prytz, A., 1995. Factors controlling suspendedsediment on inner-shelf coral reefs, Townsville, Australia. Coral Reefs 14, 163–171.
Larned, S.T., 1998. Nitrogen- versus phosphorus-limited growth and sources ofnutrients for coral reef macroalgae. Mar. Biol. 132, 409–421.
Le Borgne, R., Rodier, M., Le Bouteiller, A., Kulbicki, M., 1997. Plankton biomass andproduction in an open atoll lagoon: Uvea, New Caledonia. J. Exp. Mar. Biol. Ecol.212, 187–210.

322 R. Fichez et al. / Marine Pollution Bulletin 61 (2010) 309–322
Le Borgne, R., Douillet, P., Fichez, R., Torréton, J.-P., 2010. Hydrography and planktontemporal variabilities at different time scales in the southwest lagoon of NewCaledonia: a review. Mar. Pollut. Bull. 61, 297–308.
Lebart, L., Morineau, A., Piron, M., 1995. Statistique ExploratoireMultidimensionnelle. Dunod, Paris. p. 456.
Legendre, P., Legendre, L., 1998. Numerical Ecology: Developments inEnvironmental Modeling. Elsevier Science, Amsterdam. p. 853.
Liston, P., Furnas, M.J., Mitchell, A.W., Drew, E.A., 1992. Local and mesoscalevariability of surface water temperature and chlorophyll in the northern GreatBarrier Reef, Australia. Cont. Shelf Res. 12, 907–921.
Lorenzen, C.J., 1966. A method for the continuous measurement of in vivochlorophyll concentration. Deep Sea Res. 13, 223–227.
Morrison, R.J., Naqasima, M.R., 1999. Fiji’s Great Astrolabe Lagoon: baseline studyand management issues for a pristine marine environment. Ocean CoastalManage. 42, 617–636.
Moss, A., Brodie, J., Furnas, M., 2005. Water quality guidelines for the Great BarrierReef World Heritage Area: a basis for development and preliminary values. Mar.Pollut. Bull. 51, 76–88.
Murphy, P., Riley, J.P., 1962. A modified single solution method for thedetermination of phosphate in natural waters. Anal. Chim. Acta 27, 31–36.
Muslim, I., Jones, G., 2003. The seasonal variation of dissolved nutrients, chlorophylla and suspended sediments at Nelly Bay, Magnetic Island. Estuar. Coast. ShelfSci. 57, 445–455.
Ouillon, S., Douillet, P., Andréfouët, S., 2004. Coupling satellite data with in situmeasurements and numerical modeling to study fine suspended sedimenttransport: a study for the lagoon of New Caledonia. Coral Reefs 23, 109–122.
Ouillon, S., Douillet, P., Fichez, R., Panché, J.Y., 2005. Enhancement of regionalvariations in salinity and temperature in a coral reef lagoon, New Caledonia. C.R.Geosci. 337, 1509–1517.
Ouillon, S., Douillet, P., Lefebvre, J.P., Le Gendre, R., Jouon, A., Bonneton, P.,Fernandez, J.M., Chevillon, C., Magand, O., Lefèvre, J., Le Hir, P., Laganier, R.,Dumas, F., Marchesiello, P., Andréfouët, S., Panché, J.Y., Fichez, R., 2010.
Circulation and suspended sediment transport in a coral reef lagoon: thesouthwest lagoon of New Caledonia. Mar. Pollut. Bull. 61, 269–296.
Pages, J., Andréfouet, S., Délesalle, B., Prasil, V., 2001. Hydrology and trophic state inTakapoto atoll lagoon: comparison with other Tuamotu lagoons. Aquat. LivingResour. 14, 183–193.
Pinazo, C., Bujan, S., Douillet, P., Fichez, R., Grenz, C., Maurin, A., 2004. Impact ofwind and freshwater inputs on phytoplankton biomass in the coral reef lagoonof New Caledonia during the summer cyclonic period: a coupled 3Dbiogeochemical modelling approach. Coral Reefs 23, 281–296.
Raimbault, P., Slawyk, G., Coste, B., Fry, J., 1990. Feasibility of measuring an automatedcolorimetric procedure for the determination of seawater nitrate in the 0 to 100nM range: examples from field and culture. Mar. Biol. 104, 347–351.
Rougerie, F., 1986. Le lagon sud-ouest de Nouvelle-Calédonie: spécificitéhydrologique, dynamique et productivité. ORSTOM, Études et thèses, Nouméa,p. 224.
Sasal, P., Mouillot, D., Fichez, R., Chifflet, S., Kulbicki, M., 2007. The use of fishparasites as biological indicators of coral-reef lagoon environmental conditions:a case study of Apogonidae parasites in the lagoon of New Caledonia. Mar.Pollut. Bull. 54, 1697–1706.
Sheppard, C., 2000. Seas at the Millennium, An Environmental Evaluation. Elsevier,Amsterdam. p. 2352.
Torreton, J.-P., Rochelle Newall, E., Pringault, O., Jacquet, S., Faure, V., Briand, E.,2010. Variability of primary and bacterial production in a coral reef lagoon(New Caledonia). Mar. Pollut. Bull. 61, 335–348.
Udy, J., Gall, M., Longstaff, B., Moore, K., Roelfsema, C., Spooner, D.R., Albert, S.,2005. Water quality monitoring: a combined approach to investigategradients of change in the Great Barrier Reef. Aust. Mar. Pollut. Bull. 51,224–238.
Webster, R., Oliver, M.A., 1991. Geostatistics for Environmental Scientists. Wileyand Sons, Chichester. p. 271.
Woodridge, S., Brodie, J., Furnas, M., 2006. Exposure of inner-shelf reefs to nutrientenriched runoff entering the Great Barrier Reef lagoon: post-European changesand the design of water quality targets. Mar. Pollut. Bull. 52, 1467–1479.