Embed Size (px)
Citation preview

Manipulation of light wavelength at appropriate growth stageto enhance biomass productivity and fatty acid methyl ester yieldusing Chlorella vulgaris
Dae Geun Kim a, Changsu Lee a, Seung-Moon Park b, Yoon-E Choi c,d,⇑a Department of Bioprocess Engineering, Chonbuk National University, 567 Baekje-daero, Deokjin-gu, Jeonju-si, Jeollabuk-do 561-756, Republic of Koreab Division of Biotechnology, College of Environmental and Bioresource Sciences, Chonbuk National University, Iksan-si, Jeollabuk-do 570-752, Republic of Koreac LED Agri-bio Fusion Technology Research Center, Chonbuk National University, 79 Gobong-ro, Iksan-si, Jeollabuk-do 570-752, Republic of Koread Department of Bioactive Material Sciences, Chonbuk National University, 567 Baekje-daero, Deokjin-gu, Jeonju-si, Jeollabuk-do 561-756, Republic of Korea
h i g h l i g h t s
! LEDs are suitable light source of microalgal cultivations.! Blue light LED illumination led to significantly increased cell size.! Red light LED illumination led to small-sized cell with active divisions.! Innovative process with wavelength shift increased biomass and FAME.
a r t i c l e i n f o
Article history:Received 20 December 2013Received in revised form 21 February 2014Accepted 22 February 2014Available online 3 March 2014
Keywords:LEDWavelength shiftChlorella vulgarisMicroalgal cultivationBiodiesel
a b s t r a c t
LEDs light offer several advantages over the conventional lamps, thereby being considered as the optimallight sources for microalgal cultivation. In this study, various light-emitting diodes (LEDs) especially redand blue color with different light wavelengths were employed to explore the effects of light source onphototrophic cultivation of Chlorella vulgaris. Blue light illumination led to significantly increased cellsize, whereas red light resulted in small-sized cell with active divisions. Based on the discovery of theeffect of light wavelengths on microalgal biology, we then applied appropriate wavelength at differentgrowth stages; blue light was illuminated first and then shifted to red light. By doing so, biomass and lipidproductivity of C. vulgaris could be significantly increased, compared to that in the control. These resultswill shed light on a novel approach using LED light for microalgal biotechnology.
! 2014 Elsevier Ltd. All rights reserved.
1. Introduction
Due to the higher price of petroleum as well as environmentalconcerns regarding to the steep rise of carbon dioxide as a green-house gas, alternative bio-fuels have been received considerableattention worldwide (Hill et al., 2006). Among the various biomasssources, microalgae are considered to be one of the most promisingfeedstocks for biodiesel, due to their rapid growth and high lipidcontent. For these reasons, the development of biodiesel originatedfrom microalgae has become a hot topic in recent years (Chisti,2007).
Microalga could also offer several additional advantages notonly limited to microalgal biodiesel but also include a wealthy ofthe other benefits. Firstly, microalgae could biologically captureand fix carbon dioxide during the process of photosynthesis. Sec-ondly, an enormous variety of compounds with high values includ-ing antioxidants and potential medications could be also producedfrom microalgal biomass. Finally, microalgae could utilized as envi-ronmental agents, since microalga are capable of rapid uptake ofnitrogen and phosphate in the wastewater. Therefore, microalgalbiomass hold a great promise for future biotechnological applica-tions and recently it has become reality with intensive effort andinvestment around the world.
In order to utilize microalgal biomass for above-mentioned pur-poses, it must be essential to obtain sufficient microalgal biomass.Therefore, microalgal cultivation is one of the most importantprocesses for the use of microalgae to produce any substances of
http://dx.doi.org/10.1016/j.biortech.2014.02.0780960-8524/! 2014 Elsevier Ltd. All rights reserved.
⇑ Corresponding author at: LED Agri-bio Fusion Technology Research Center,Chonbuk National University, 79 Gobong-ro, Iksan-si, Jeollabuk-do 570-752,Republic of Korea. Tel.: +82 63 850 0773.
E-mail address: [email protected] (Yoon-E Choi).
Bioresource Technology 159 (2014) 240–248
Contents lists available at ScienceDirect
Bioresource Technology
journal homepage: www.elsevier .com/locate /bior tech

interest, particularly for biodiesel. Moreover, it has been well ac-cepted that the cultivation process alone claims high productioncost, restricting further industrialization or commercializationusing microalgae. So far, most of research for the production of bio-diesel using microalgal biomass was limited in the small scale.However, in order to achieve economic feasibility, efficient masscultivation strategy with high lipid productivity remains a keychallenge to microalgal biotechnology (Uduman et al., 2010).
As with photoautotrophic organisms, microalgae carry out thephotosynthesis as a main metabolism for the acquisition of organicmaterials by using energy from light sources such as sunlight orartificial light. Therefore, multiple lines of evidences suggested thatlight is the most significant factor governing the entire process ofmciroalgal cultivation. Based on different approaches to obtainlight source, the strategy for microalgal cultivations could bebroadly categorized into open-pond or photobioreactor. Whileopen-pond system utilize free sunlight, microalgal cultivation inthe closed photobioreactor with artificial light source have theindisputable advantages in that biomass productivity could be sig-nificantly increased, particularly for value added products, com-pared to those in open pond system. However, again, the supplyof artificial light causes the most expensive cost to operate thephotobioreactor. Moreover, light energy penetrated into microalgalsuspensions was significantly decreased along with increase oflight path-length at high microalgal cells concentrations, whichacts as a bottleneck for microalgal cultivation. Therefore, it willbe crucial to developing efficient process to maximize the utiliza-tion of light energy, thereby improving economic feasibility in mic-roalgal cultivation process.
Recently, light-emitting diodes (LED) has been emerged as areplacement of traditional artificial light source. Compared to theconventional tubular fluorescence lamps, the recent developedLEDs light make significant advances in narrowing a specific wave-length with low power consumption. Additionally, LEDs are suit-able light source of photobioreactor (PBR) for indoor masscultivation due to their small chip size and long duration (Zhaoet al., 2011). Due to these advantages, now, the applications ofLEDs are actively extended to the field of microalgal cultivation(Wang et al., 2007). Because LEDs could provide a particular wave-length to illuminate microalgal culture, it will be pertinent to selectLEDs for the purpose of adequate manipulation of microalgal culti-vation. In accordance with this notion, a number of studies haveaddressed that there are optimum wavelengths for each of micro-algal species, though contradictory results have been obtainedabout the influence of specific wavelength of LED on microalgalgrowth. Whereas a red light was the most effective for Botryococcusbraunii (Baba et al., 2012), a blue light led to the best biomass pro-ductivity for Nannochloropsis sp. (Das et al., 2011). Similar contra-dictory results regarding specific wavelength for microalgalcultivation were obtained previously (Baba et al., 2012; Lee andPalsson, 1994). Hence, we still have limited reports regardingLED applications into the field of microalgal cultivations.
Furthermore, although some researchers attempted to utilizeLEDs for microalgal growth, there has been no research to investi-gate and consider biological responses of microalgal cells upon thespecific wavelength of LED illuminations, including the cellularphysiology or gene expressions. To date, very few studies have onlyfocused on the installment of simple LED illuminations for micro-algal growth without knowing detailed cellular mechanisms uponspecific LED wavelengths. The knowledge on these will become ofgreat interest to the microalgal bioengineers and could be directlyapplied into microalgal biotechnology, such as mass cultivation inthe photobioreactors.
Therefore, in this study, we examined the effect of specificwavelength of LEDs on microalgal biology including growth, cellphysiology, fatty acid biosynthesis, and gene expressions using
Chlorella vulgaris. Indeed, it turned out that red and blue wave-length has a specific influence on C. vulgaris biology. Consideringthe effect of LED wavelengths in particular, we then tried to manip-ulate red or blue wavelength illumination at appropriate stage ofmicroalgal cell growth. These results demonstrated that adequatemanipulations of LED wavelengths during the microalgal cellgrowth could lead to significant increase in biomass and lipidproductivity. Here, we report a novel discovery of specific influenceof LED wavelengths on microalgal biology and propose the putativeprocess based on LED wavelength shift for the enhanced produc-tion of biomass as well as lipid using C. vulgaris.
2. Methods
2.1. Strain and culture condition
C. vulgaris was obtained from the UTEX culture collection.Inoculums were regularly prepared using modified JM medium(Thompson et al., 1988) in 500 mL Erlenmeyer flasks at25 ± 0.5 "C under cool-white fluorescent lights (approximately50 lmol m"2 s"1). For the all experiments, cells were cultivatedin 1000 mL Erlenmeyer flasks containing 600 mL JM medium. Theinitial cell density was adjusted to be 106 cells mL"1 and tempera-ture was maintained at 26 ± 0.5 "C. Continuous illumination wassupplied at an average light intensity of 100 lmol m"2 s"1 undereither monochromatic blue (kmax = 430–465 nm) or red (630–665 nm) light. To ensure sufficient aeration, cultures were bubbledwith sterilized-filtered air at a flow rate of 100 mL min"1.
2.2. Analysis
The UV/vis spectrophotometer (T60 U, korea) was used for mea-suring both optical density and the level of ROS staining.
To analyze chlorophylls, cells were centrifuged, washed twicewith de-ionized (DI) water, and the pellet was dried for 24 h. Then,equal amount of dried cells was re-suspended in 100% MeOH at50 "C for 50 min. After centrifugation at 12,000g for 5 min, super-natant was collected and absorbance was measured at 650 and665 nm, respectively. The content of chlorophyll a, chlorophyll b,and the total chlorophyll content (mg/L) was calculated using thefollowing equations:
Chlorophyll a ¼ 16:5$ A665" 8:3$ A650
Chlorophyll b ¼ 33:8$ A650" 12:5$ A665
Chlorophyllðaþ bÞ ¼ 25:5$ A650þ 4:0$ A665
2.3. Growth measurements
Algal growth was measured with optical density at 660 nm witha UV/vis spectrophotometer (T60 U, korea). The number of cellswas determined by direct counting with a haemocytometer(Hausser Scientific, Horsham, PA) under an OPTINITY microscope(KB-500, korea). The dry cell weight was measured by filteringthe algal suspension through a pre-dried and pre-weighed,0.45 lm cellulose nitrate membrane filter (Whatman, USA) anddrying in an oven at 80 "C for 24 h. The specific growth rate (l)was calculated based on the following equation.
l ¼ ðlnX1 " lnX0=t1 " t0Þ
where X1 and X0 were the biomass concentration (g L"1) at time t1
and t0, respectively.
D.G. Kim et al. / Bioresource Technology 159 (2014) 240–248 241

The size of Chlorella was measured with OPTINITY microscopecamera (C30, Korea) from more than one hundred microalgal cellsrandomly selected.
2.4. Transcription analysis
Microalgal cultivations were performed for 3 days under eitherwhite, red, or blue illumination. Total RNA was extracted from100 mg of algal cultures with TRIzol reagent (Invitrogen) inaccordance with the manufacturer’s protocols. Nucleic acidconcentrations were measured spectrophotometrically at 260 nm.The integrity and purity of the RNA were determined by 260/280 nm ratios and separated by electrophoresis on a 1% agaroseformaldehyde gel. Reverse transcription (RT) was carried out usinga capacity RNA-to-cDNA kit (Applied Biosytems). Four photosyn-thesis-related genes and one cell division-related gene in a fresh-water algae C. vulgaris were selected as target genes. These selectgenes are: (i) psaB (NCBI GeneID: 809130), which codes for thephotosystem I (PS I) reaction centre protein (ii) chlB (GeneID:809139), which codes for light-independent protochlorophyllidereductase subunit B (iii) rbcL (accession number: AF499684),which codes for the large subunit of rubisco (iv) psbA, which codesfor the photosystem II (PS II) reaction protein D1 (v) minD, whichcodes for cell division inhibitor protein. QRT-PCR analysis was per-formed with SYBR-Green# as the fluorescent. Gene specific primersfor psaB gene (psaB-F1 and psaB-R1), chlB gene (chlB-F1 and chlB-R1), rbcL gene (rbcL-F1 and rbcL-R1), psbA gene (psbA-F1 andpsbA-R1), and minD (minD-F1 and minD-R1) were used for givingrise to amplicons. The expression of each gene was normalized toendogenous 18s rRNA gene expression with specific pair of primers(18s-F1 and 18s-R1). The gene expression was calibrated using2"DDCt method. The range of expression was calibrated using2"DDCt"s–2"DDCt+s, where s is the standard deviation of DCt value(Ct = Threshold Cycle). All primers used for PCR are listed inTable 1.
2.5. FAME analysis
After cultivation, algal biomass was harvested by centrifugation(7000 rpm, 3 min) and then washed twice with DI water. The cellpellet was freeze-dried at "40 "C for 3 days and used for lipidextraction. Total lipids were extracted from 10 mg of lyophilizedbiomass with a chloroform–methanol (2:1 v/v) solvent mixtureaccording to the modified Folch method (Folch et al., 1957). Ex-tracted lipids were then converted into fatty acid methyl esters(FAMEs) via the transesterification reaction (with methanol andsulfuric acid as a catalyst) at 105 "C for 20 min. The FAMEs in theorganic phase were analyzed by gas chromatography (HP6890,Agilent, USA), with a flame ionized detector (FID) and an INNO-WAX capillary column (Agilent, USA, 30 m $ 0.32 mm $ 0.5 lm).
The identification and quantification of fatty acids were deter-mined by comparison with the retention times and peak areas ofstandards.
2.6. ROS staining
In situ detection of the superoxide radical was performed byalgae cultures with nitroblue tetrazolium (NBT) (N6876, Sigma–Aldrich) staining following the protocol described previously(Rao and Davis, 1999). Three biological replications were per-formed. Cultures grown in JM media for 3 days were centrifugedand re-suspended in 0.2% NBT. After incubation for 12 h, cells werere-centrifuged and re-suspended in 10 ml of 50% glacial acetic acid.For quantification, sonicator was used to lyse the stained cells andthe extent of ROS was measured with the absorbance at 560 nm.
2.7. Statistical analysis
All experiments were performed with three replications. Dataobtained were analysis by one-way ANOVA following Duncan’smultiple range test with the significance level (P < 0.05). The SPSSversion 17 (SPSS Inc., Chicago, IL, USA) program was used for allstatistical analyses.
3. Results and discussion
3.1. Effect of LED wavelength on algae growth
To test the effects of specific wavelength on the growth of mic-roalgae, ubiquitous microalga C. vulgaris was chosen and its growthcharacteristics cultivated under different three wavelengths of LEDwere thoroughly examined. Microalgae have chlorophylls andaccessory pigments with maximum absorption peaks in the redor blue part of light spectrum. For this reason, three differentwavelengths (450, 660 and 400–700 nm) representing blue(450 nm), red (660 nm), and white (400–700 nm) as a control wereselected to determine the effects of major wavelengths necessaryfor the photosynthesis on microalgal biology. Results showed thatspecific wavelength could indeed affect microalgal growth(Fig. 1a). Cell numbers were highly dependent on light wavelength,since C. vulgaris produced the highest and the lowest cell numberunder the red and blue light, respectively. The cell density ofC. vulgaris grown under red light was 1.5-fold higher than thoseunder blue light at the certain time point after the inoculation(Fig. 1a). These results are consistent with other microalgal speciesin that red wavelength turned out to be optimal for both Spirulinaplatensis and Chlorella pyrenoidosa (Wang et al., 2007). Matthijset al. (1996) reported that red light led to enhanced chlorophyllpigmentation, which could explain the results of a positive effectof red light. Since red light had a similar influence on microalgalgrowth in both C. pyenoidosa and S. platensis (Wang et al., 2007),the responses upon light wavelength might be evolutionary well-conserved between different microalgal species. On the other hand,white light gave rise to middle extent of cell number, whereas bluelight consistently decrease the level of cell number. One possiblespeculation is that blue light could suppress microalgal celldivision, thereby resulting in lower level of cell number. Furtherstudy might be necessary to determine cellular mechanisms ofmicroalgae upon different wavelengths of light.
3.2. The effect of blue wavelength on the microalgal cell size
Interestingly, microalgal cell morphology, e.g. cell size, ofC. vulgaris was altered significantly in response to different wave-length of light. Cell size of C. vulgaris cultivated under blue light
Table 1All primers used in this study.
Target Primer name Nucleotide sequence (50-30)
chlB chlB-F1 50-TGTTGCTGTTACACCGATGGG-30
chlB-R1 50-GAGTGGCATCTCCAAAGACT-30
psbA psbA-F1 50-GGTTATACAACGGTGGTCCT-30
psbA-R1 50-CCTTGACCGATTGGGTAGAT-30
psaB psaB-F1 50-ATTCCACGTTGCTTGGCAAGG-30
psaB-R1 50-GCTGGTTGGAACGATGGTTGA-30
rbcL rbcL-F1 50-GGACGACTGTATGGACTGATG-30
rbcL-R1 50-CACGTATGCTGGTGGAATACG-30
minD minD-F1 50-GGCTTGGTTATCGTGTTGCTC-30
minD-R1 50-ACTGCCTCTTGTGCAGATGCA-30
18s RNA 18s-F1 50-ACTTCACGAATCGCATGGCCT-30
18s-R1 50-GACTTGCCCTCCAATTGATCC-30
242 D.G. Kim et al. / Bioresource Technology 159 (2014) 240–248

was consistently larger than those cultivated under red light. How-ever, middle extent of cell size was observed under the white light(Fig. 1b). Measurement of the cell size of C. vulgaris revealed thatmicroalgal cells grown under blue light have approximately a60–70% increase in the diameter, compared to those under redlight (Fig. 1b).
To verify these observations further, two independent analyseswere performed. First method was based on the correlationbetween optical density and cell number (Choi et al., 2011). Thecorrelation graphs between optical densities (OD) and cell numberwere plotted to check any change in the correlation between blueand red light. Consistent with the observation of increase in cellsizes under blue light, the correlation curve was significantlyaltered by the blue light (Fig. 1c). Higher OD values were consis-tently obtained at certain cell number under the blue wavelength,compared to those under red wavelength (Fig. 1c). The differencein the correlations between blue and red light clearly reflects thefact that there were significant changes in morphological feature(differences in cell size) by changing light wavelength.
Additionally, the relative dry weight (g/cell) by taking cellnumbers into consideration was also calculated. To thoroughlycompare the effect of either blue or red light, eight (430, 435,440, 445, 450, 455, 460, and 465 nm) and five (645, 650, 655,660, and 665 nm) different LED wavelengths were selected fromblue and red light, respectively. Expectedly, dry weight per cellscould be fit into two categories: either blue or red light. Again, it
is evident that the dry weight per cell of C. vulgaris was alteredby the illumination of different wavelength of light (Fig. 1d).Approximately, 30% increase in the dry weight per cell of C. vulgariswas observed by changing the wavelength from red to blue(Fig. 1d). Previous reports suggested similar effect of wavelengthon microalgal cells, though the evidences were fragmental withoutproviding a conclusive result. Lee and Palsson (1994) demon-strated that red light led to smaller cells than those under thefull-spectrum white light due to the early autospore release uponred light. Chlorella cells cultivated under blue light produced largerand fewer autospores (Kowallik, 1963; Pirson and Kowallik, 1964),whereas blue light delayed the Chlamydomonas reinhardtii cell divi-sion (Münzner and Voigt, 1992; Oldenhof et al., 2004b). Interest-ingly, Oldenhof et al. (2004a, 2006) recently demonstrated thatC. reinhardtii cells could be altered in cell cycle by shifting lightwavelength to either blue or red (Oldenhof et al., 2004a, 2006).Furthermore, red or blue light could affect the division of chloro-plast nucleoids and replication of chloroplast DNA in Dunaliellasalina (Zachleder et al., 1989). Even though all of these previousstudies were not intended to perform systematic comparison be-tween blue and red light, they partially provided the evidences thatcell division could be accelerated or delayed upon red and bluelight. It is likely that differential rate of cell divisions or cell cycleprogression under either blue or red light might be underlyingmechanism for the observation of different cell size upon red orblue light. Consistent with these results, the data in this study
Fig. 1. Effects of three different LED wavelength including red (-s-), white (-d-) and blue (-.-) on C. vulgaris. (a) Cell density; (b) cell size; (c) correlation between cellnumber and OD (optical density). Different letters over the bar represent significant difference (P < 0.05); (d) dry weight per cells cultured under different wavelength. Eightblue (left) and five red wavelengths (right) were selected.
D.G. Kim et al. / Bioresource Technology 159 (2014) 240–248 243

clearly indicate that C. vulgaris morphology of cell size was signif-icantly transformed into bigger cells by the exposure of blue light,compared to those under red light.
3.3. Investigation of possible molecular mechanisms of different C.vulgaris responses under red, blue and white light
In order to determine molecular mechanisms of different C. vul-garis responses under red, blue and white light, we next examinedthe expression of genes possibly responsible for increased cell sizeupon blue light. Based on altered cell morphology and possibleinfluence over the photosynthesis under the different wavelengths,genes belonging to functional categories involved in either photo-synthesis or cell division were selected. qRT-PCR was used toinvestigate the relative expression of these genes under the condi-tions between white, red, and blue illumination. Of select genes,we could successfully identify the genes, which displayed differen-tial expressions under white, red, and blue illumination. Identifiedgenes were psaB encoding for the photosystem I (PS I) reactioncenter protein, chlB for light-independent protochlorophyllidereductase subunit B, rbcL for the large subunit of rubisco, psbAfor the photosystem II (PS II) reaction protein D1, and minD for celldivision inhibitor protein.
Except for minD and psaB, all of genes displayed similar patternsof expression in that the abundances of gene transcripts were grad-ually decreased with the change of light wavelength from blue,white and red (Fig. 2). Compared with those under red or white,mRNA expressions of chlB, psbA, and rbcL were significantly higher(P < 0.05) under the blue light. chlB is involved in light-indepen-dent reaction for chlorophyll biosynthesis (Suzuki and Bauer,1992) and rbcL encodes the large subunit of rubisco, which is thekey enzyme of the calvin cycle (Qian et al., 2009). psbA is requiredfor photosystem II encoding protein D1 (Suzuki and Bauer, 1992).Since mRNA expressions of chlB, psbA, and rbcL were increasedupon the exposure to blue light in C. vulgaris, blue light might stim-ulate the activation of enzyme(s) linked with photosynthesis.Without doubt, blue light is more energetic radiation than red lightdue to its shorter wavelength causing more stress to C. vulgariscells (Jeong et al., 2013; Menon et al., 2013). Due to this reason,it is likely that C. vulgaris cells must adapt stressful radiation ofblue light, which might require high level of expression of photo-synthesis-related genes such as chlB, psbA, and rbcL. In accordancewith this notion, it has been well accepted that photosynthesis,particularly PS II, play an important role in regulating or adjustingthe light harvesting capacity (Briantais et al., 1979; Chow et al.,1990; Sukenik et al., 1987). For this reason, it might be possibleto detect the increase of gene expressions involved in photosynthe-sis, particularly PS II, under the blue illumination, as observed inthis study (Fig. 2).
On the other hand, psaB encodes a protein of the PS I reactioncenter involved in binding of electron transfer components(Murray et al., 2006). The results showed that gene (psaB) encodingPS I protein transcribed differently from those linked with PS II.Since both blue and red light could led to increased transcript forpsaB, compared to those under full-spectrum white light, it is likelythat psaB expression could be increased upon exposure of either oftwo preferential wavelength of photosynthesis; red and blue light.
In addition, minD encodes the cell division inhibitor (Wakasugiet al., 1997). Transcription of minD was significantly suppressedunder the red light, compared to those under both blue and white.Consistent with the patterns of minD gene expression, increasedand decreased cell division was detected under the red and bluelight, respectively, thereby rendering differential C. vulgaris cellsizes upon exposure of red and blue light. Therefore, since minDexpression and C. vulgaris cell sizes were well-correlated eachother under the red and blue light, these data provides molecular
evidence that minD serves as a negative regulator of cell sizes upondifferent wavelength of light in C. vulgaris.
3.4. Determination of ROS stress level upon different wavelength
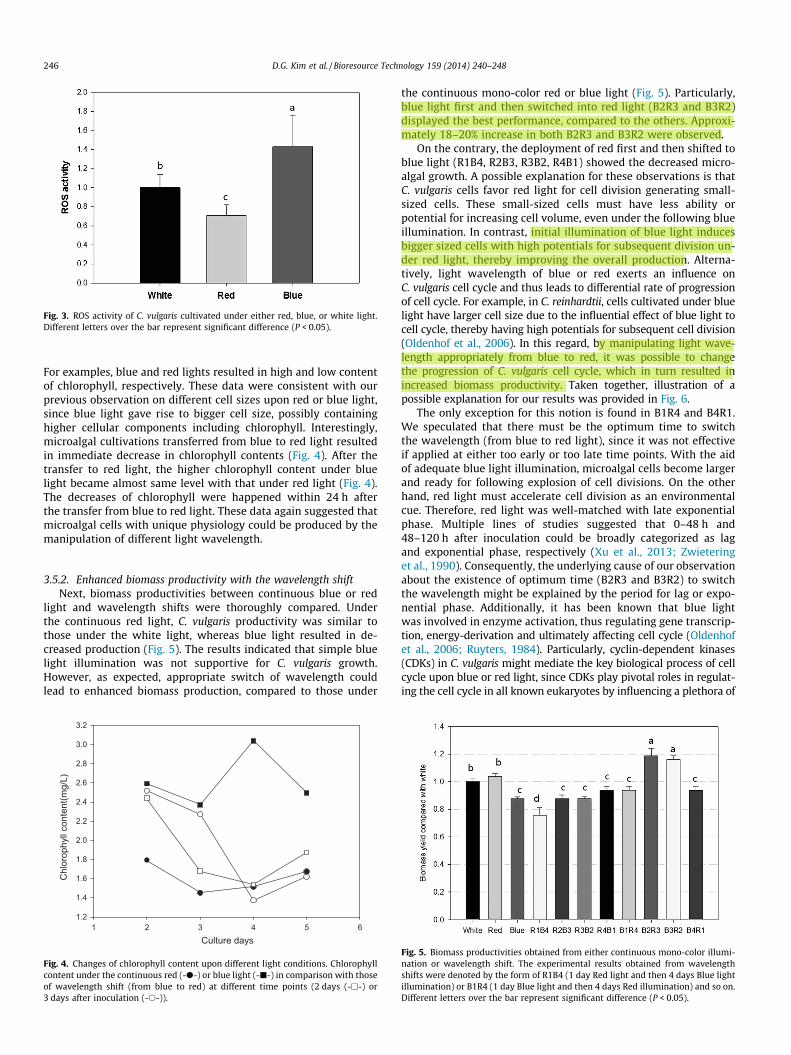
Because, theoretically, blue light has a feature of high energeticwavelength, it is reasonable to hypothesize that C. vulgaris cellscould accumulate higher extent of stress under the blue light. Tofurther understanding of responsive stress mechanisms in microal-gal cells upon specific wavelength of light, particularly blue light,we examined the extent of cellular stress via the measurement ofreactive oxygen species (ROS). Generally, ROS are the typicalmolecular mediators of various stresses in the cell of a wide varietyof living species (Apel and Hirt, 2004; Hamid Badawi et al., 2004).To detect and quantify ROS generation in C. vulgaris cells upon dif-ferent light wavelength, the standard protocol of nitroblue tetrazo-lium (NBT) staining was utilized. The reactive oxygen species (ROS)activity of C. vulgaris cultivated under three different wavelength oflight (red, blue and white) were thoroughly compared with threebiological replications (Fig. 3). Quantification demonstrated thatcultivation under blue light cells displayed significantly (P < 0.05)higher extent of ROS than those under white or red light (Fig. 3).The analyses on the level of ROS demonstrated that cells grown un-der blue light had around a 40% increase of ROS, whereas cellsgrown under red light showed more than 30% decrease of ROS level(Fig. 3). These data clearly indicated that blue wavelength led to in-crease the level of cellular ROS, whereas red light decreased theROS level in C. vulgaris.
Previously, it has been reported that shorter wavelengths suchas blue and green light deliver more energy for photosynthesis,thereby causing photo-inhibition (Das et al., 2011; Jeong et al.,2013). On the other hand, red light has an opposite feature of long-er wavelength, possibly alleviating the effect of photo-inhibition(Matthijs et al., 1996). However, there has been no direct or indi-rect evidence demonstrating the involvement of ROS for these mic-roalgal phenotypes under different wavelengths of light. In thisstudy, we first demonstrated that stress mechanism(s) associatedwith ROS could help explain the photo-inhibition of microalgaeupon blue light. Moreover, ROS level was well-correlated with dif-ferential C. vulgaris cell sizes upon exposure of red and blue light(Figs. 1d and 3). Consequently, it is conceivable that increasedROS generation via blue light illumination may be responsible forincreased cell size morphology in C. vulgaris. Similarly, microalgaHaematococcus pluvialis has a distinctive lifecycle, because it exhib-its small-sized green cells under favorable environment and big-ger-sized resting cells under unfavorable condition (Choi et al.,2011). Though C. vulgaris has not much variation in cell size likeH. pluvialis, the results indicated that blue light induces oxidativestress in C. vulgaris cells thereby causing the cellular transforma-tion into bigger cells to cope with the unfavorable condition of highenergy irradiance from blue light.
3.5. Innovative wavelength shift approach to enhance productivities ofC. vulgaris culture
From the above results, we obtained novel results regarding tophenotypic characteristics upon different light wavelengths anddiscovered the possible mechanisms behind these phenotypiccharacteristics by confirming differential gene expression as wellas proving altered ROS levels. Notably, cell morphology of sizeswas significantly changed depending on the wavelength of light.Red light enhanced cell division giving rise to small cells, whereasblue light led to increased cell size.
Since specific wavelength of light stimulates particular charac-teristics of microalgal physiology of differential cell sizes, wethen next attempted to utilize these cellular features upon LED
244 D.G. Kim et al. / Bioresource Technology 159 (2014) 240–248

wavelength into the biotechnological applications. We hypothe-sized that different cell sizes possibly reflect different cellularpotentials containing different extent of nutrients or proteins forfollowing cell divisions. Based on that, we can speculate that gen-eration of microalgal cells with bigger size during the beginningstage of cultivation might help promote subsequent cell divisionin the later growth stage, thereby leading to enhanced productivi-ties for biomass as well as total FAME. This hypothesis promptedus to propose that if we could manipulate microalgal cell sizesproperly via the appropriate illumination of LED wavelength, wemight be able to further enhance microalgal productivities.
In order to test the hypothesis, we adopted adequate manipula-tion of LED wavelength illumination as the way to generate differ-ent size of microalgal cells at different growth time points. Theexperiments were performed via the shift of light wavelength at
different time points. The set of experiments was consisted of fivedifferent light illuminations: continuous blue or red light; bluelight first and then shifted into red light or vice versa. Light wave-length was shifted either blue or red after 24, 48, 72, and 96 h ofcultivation. The experimental results obtained from wavelengthshifts were designated as the form of R1B4 (1 day Red light andthen 4 days Blue light illumination) or B1R4 (1 day Blue light andthen 4 days Red illumination) and so on.
3.5.1. Change of chlorophyll contents under the wavelength shiftChlorophyll content as an indicator of cell physiology was first
examined. The change of chlorophyll contents of C. vulgaris cellsupon different wavelength of red or blue light or under thewavelength shifts were shown in Fig. 4. Notably, the content ofC. vulgaris chlorophyll was highly dependent on light wavelength.
(A) (chlB)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
(C) (psaB)
Rela
tive
trans
crip
tiona
l abu
ndan
ce
0
1
2
3
4
5
(B) (psbA)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
(E) (minD)
White Red Blue0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
(D) (rbcL)
White Red Blue0.0
0.5
1.0
1.5
2.0
2.5
Fig. 2. Expression of ChlB (A), psbA (B), psaB (C), rbcL (D) and mind (E) from C. vulgaris cultivated under either red, blue, or white light.
D.G. Kim et al. / Bioresource Technology 159 (2014) 240–248 245
For examples, blue and red lights resulted in high and low contentof chlorophyll, respectively. These data were consistent with ourprevious observation on different cell sizes upon red or blue light,since blue light gave rise to bigger cell size, possibly containinghigher cellular components including chlorophyll. Interestingly,microalgal cultivations transferred from blue to red light resultedin immediate decrease in chlorophyll contents (Fig. 4). After thetransfer to red light, the higher chlorophyll content under bluelight became almost same level with that under red light (Fig. 4).The decreases of chlorophyll were happened within 24 h afterthe transfer from blue to red light. These data again suggested thatmicroalgal cells with unique physiology could be produced by themanipulation of different light wavelength.
3.5.2. Enhanced biomass productivity with the wavelength shiftNext, biomass productivities between continuous blue or red
light and wavelength shifts were thoroughly compared. Underthe continuous red light, C. vulgaris productivity was similar tothose under the white light, whereas blue light resulted in de-creased production (Fig. 5). The results indicated that simple bluelight illumination was not supportive for C. vulgaris growth.However, as expected, appropriate switch of wavelength couldlead to enhanced biomass production, compared to those under
the continuous mono-color red or blue light (Fig. 5). Particularly,blue light first and then switched into red light (B2R3 and B3R2)displayed the best performance, compared to the others. Approxi-mately 18–20% increase in both B2R3 and B3R2 were observed.
On the contrary, the deployment of red first and then shifted toblue light (R1B4, R2B3, R3B2, R4B1) showed the decreased micro-algal growth. A possible explanation for these observations is thatC. vulgaris cells favor red light for cell division generating small-sized cells. These small-sized cells must have less ability orpotential for increasing cell volume, even under the following blueillumination. In contrast, initial illumination of blue light inducesbigger sized cells with high potentials for subsequent division un-der red light, thereby improving the overall production. Alterna-tively, light wavelength of blue or red exerts an influence onC. vulgaris cell cycle and thus leads to differential rate of progressionof cell cycle. For example, in C. reinhardtii, cells cultivated under bluelight have larger cell size due to the influential effect of blue light tocell cycle, thereby having high potentials for subsequent cell division(Oldenhof et al., 2006). In this regard, by manipulating light wave-length appropriately from blue to red, it was possible to changethe progression of C. vulgaris cell cycle, which in turn resulted inincreased biomass productivity. Taken together, illustration of apossible explanation for our results was provided in Fig. 6.
The only exception for this notion is found in B1R4 and B4R1.We speculated that there must be the optimum time to switchthe wavelength (from blue to red light), since it was not effectiveif applied at either too early or too late time points. With the aidof adequate blue light illumination, microalgal cells become largerand ready for following explosion of cell divisions. On the otherhand, red light must accelerate cell division as an environmentalcue. Therefore, red light was well-matched with late exponentialphase. Multiple lines of studies suggested that 0–48 h and48–120 h after inoculation could be broadly categorized as lagand exponential phase, respectively (Xu et al., 2013; Zwieteringet al., 1990). Consequently, the underlying cause of our observationabout the existence of optimum time (B2R3 and B3R2) to switchthe wavelength might be explained by the period for lag or expo-nential phase. Additionally, it has been known that blue lightwas involved in enzyme activation, thus regulating gene transcrip-tion, energy-derivation and ultimately affecting cell cycle (Oldenhofet al., 2006; Ruyters, 1984). Particularly, cyclin-dependent kinases(CDKs) in C. vulgaris might mediate the key biological process of cellcycle upon blue or red light, since CDKs play pivotal roles in regulat-ing the cell cycle in all known eukaryotes by influencing a plethora of
Fig. 3. ROS activity of C. vulgaris cultivated under either red, blue, or white light.Different letters over the bar represent significant difference (P < 0.05).
Culture days1 2 3 4 5 6
Chl
orop
hyll
cont
ent(m
g/L)
1.2
1.4
1.6
1.8
2.0
2.2
2.4
2.6
2.8
3.0
3.2
Fig. 4. Changes of chlorophyll content upon different light conditions. Chlorophyllcontent under the continuous red (-d-) or blue light (-j-) in comparison with thoseof wavelength shift (from blue to red) at different time points (2 days (-h-) or3 days after inoculation (-s-)).
Fig. 5. Biomass productivities obtained from either continuous mono-color illumi-nation or wavelength shift. The experimental results obtained from wavelengthshifts were denoted by the form of R1B4 (1 day Red light and then 4 days Blue lightillumination) or B1R4 (1 day Blue light and then 4 days Red illumination) and so on.Different letters over the bar represent significant difference (P < 0.05).
246 D.G. Kim et al. / Bioresource Technology 159 (2014) 240–248

gene transcriptions. In this context, Oldenhof et al. (2004a, 2006) re-ported that, in microalga C. reinhardtii, there is corresponding in-crease in CDK activity under blue light, which in turn affectmultiple downstream genes for cell division or cell cycle progression.Because of the importance of appropriate timing of CDK activation toensure the commitment to cell cycle, it is not surprising to notice thatthere is the optimal range of period to illuminate blue or red light forproper CDK function to improve biomass productivity. Further studymight be necessary to pinpoint the involvement of CDKs in alterationof microalgal biology upon different wavelength of light as observedin this study.
To prove further, statistical analysis was also performed andfurther suggested that B2R3 and B3R2 have the highest productiv-ities, compared to the others (P < 0.05). Based on that, we con-cluded that the novel strategy for microalgal cultivation via themanipulation of light wavelength could be successfully demon-strated in C. vulgaris. Since the method based on the appropriatemanipulation of light wavelength could be easily expanded tothe other species of microalgae, we believe that this approach willopen the way to achieve further enhancement for microalgalbiotechnology.
3.5.3. Enhancement of biodiesel production with the wavelength shiftWe further tested whether wavelength shift could increase fatty
acid production for biodiesel. Table 2 summarizes the fatty acidsprofiles and total FAME (TFAME) obtained from different light illu-minations: continuous blue or red light; R1B4–R4B1; B1R4–B4R1.Fatty acid profiles were not altered significantly with the changeof the wavelength, which is consistent with the previous report(Tang et al., 2011). However, TFAME was highly dependent uponlight illumination. It should be noteworthy that overall FAMEyield(s) could be significantly increased via the switch of wave-length, possibly owing to the efficient growth with the help ofsophisticated manipulation of light wavelength. Under the contin-uous illumination of either white, red, or blue light, approximatesimilar extent of TFAME were obtained (9.61%, 10.65% and11.07%, respectively). However, TFAME obtained by appropriatewavelength shift (from blue to red) was higher than those of
continuous light illuminations due to the increased biomass pro-ductivities (Table 2). Taken together, appropriate manipulation ofLED wavelength in consideration of microalgal biology could im-prove both microalgal growth as well as biodiesel production.We believe that a novel discovery revealed in this study will helppush the limit for microalgal biotechnology leveraging its futureapplication.
4. Conclusion
In this study, the effect of specific light wavelength on C. vulga-ris biology was thoroughly investigated, particularly with red orblue light. Based on the discovery of the effect of light wavelengthson microalgal biology, the appropriate wavelength at different
Fig. 6. Putative model explaining different morphological progressions of C. vulgaris upon different wavelength of light. Blue light can increase cell sizes, whereas red lightenhanced cell divisions. Consequently, we proposed a novel strategy for microalgal cultivation based on light wavelength change (from blue to red). By doing so, microalgalcells with bigger size can be generated during the beginning stage of cultivation. Bigger microalgal cells must have a huge potential for cell division. Thus following red lightillumination then help promote subsequent cell division in the later growth stage, thereby leading to enhanced productivities for biomass as well as lipid production. (Forinterpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
Table 2Fatty acids profiles (% of total lipids) of the C. vulgaris under either continuous mono-color illumination or wavelength shifts.
Fatty acid Light condition
Red R1B4* R2B3* R3B2* R4B1* White
C10:0 1.51 0.77 0.73 0.83 0.78 0.51C14:0 1.11 1.23 1.32 1.42 1.10 1.63C16:0 16.53 14.60 15.01 14.20 15.26 14.38C17:1 11.23 12.88 12.62 13.10 12.27 12.28C18:0 7.13 5.98 6.09 3.70 7.21 2.94C18:2 10.33 7.83 8.76 7.76 8.51 9.40C18:3 29.70 33.29 32.51 33.47 33.41 30.30TFAME/DW (%) 10.65 11.44 12.07 9.80 11.33 9.61
Blue* B1R4* B2R3* B3R2* B4R1*
C10:0 1.15 1.31 1.05 1.70 1.10C14:0 1.36 0.77 1.00 0.68 1.06C16:0 14.49 16.26 15.78 16.54 15.28C17:1 13.35 11.45 11.34 11.36 12.64C18:0 3.55 10.86 6.46 8.99 4.19C18:2 8.19 10.09 9.32 9.79 10.41C18:3 32.57 30.15 31.05 30.01 29.99TFAME/DW (%) 11.07 14.11 13.09 13.40 12.24
* The experimental results obtained from wavelength shifts were designated as theform of R1B4 (1 day Red light and then 4 days Blue light illumination) or B1R4(1 day Blue light and then 4 days Red illumination) and so on.
D.G. Kim et al. / Bioresource Technology 159 (2014) 240–248 247

growth stages was conceived as a novel strategy for C. vulgaris cul-tivation. With our innovative biotechnology process of wavelengthshift, we could achieve further increased productions in both bio-mass and FAME. Consequently, these results will have significantimpact on future development of microalgal biotechnology.
Acknowledgements
This research was financially supported by the Ministry ofTrade, Industry & Energy (MOTIE), Korea Institute for Advancementof Technology (KIAT) and Honam Institute for Regional ProgramEvaluation through the Leading Industry Development for Eco-nomic Region. This work was also supported by the Advanced Bio-mass R&D Center (ABC) of Korea Grant funded by the Ministry ofEducation, Science and Technology (ABC-2012-055032). In addi-tion, this work was also supported by the Industrial TechnologyResearch Infrastructure Program (N0000004) funded by the Minis-try of Trade, Industry and Energy (MTE, Korea).
References
Apel, K., Hirt, H., 2004. Reactive oxygen species: metabolism, oxidative stress, andsignal transduction. Annu. Rev. Plant Biol. 55, 373–399.
Baba, M., Kikuta, F., Suzuki, I., Watanabe, M.M., Shiraiwa, Y., 2012. Wavelengthspecificity of growth, photosynthesis, and hydrocarbon production in the oil-producing green alga Botryococcus braunii. Bioresour. Technol. 109, 266–270.
Briantais, J.-M., Vernotte, C., Picaud, M., Krause, G., 1979. A quantitative study of theslow decline of chlorophyll a fluorescence in isolated chloroplasts. Biochim.Biophys. Acta 548, 128–138.
Chisti, Y., 2007. Biodiesel from microalgae. Biotechnol. Adv. 25, 294–306.Choi, Y.-E., Yun, Y.-S., Park, J.M., Yang, J.-W., 2011. Multistage operation of airlift
photobioreactor for increased production of astaxanthin from Haematococcuspluvialis. J. Microbiol. Biotechnol. 21, 1081–1087.
Chow, W.S., Melis, A., Anderson, J.M., 1990. Adjustments of photosystemstoichiometry in chloroplasts improve the quantum efficiency ofphotosynthesis. Proc. Natl. Acad. Sci. 87, 7502–7506.
Das, P., Lei, W., Aziz, S.S., Obbard, J.P., 2011. Enhanced algae growth in bothphototrophic and mixotrophic culture under blue light. Bioresour. Technol. 102,3883–3887.
Folch, J., Lees, M., Stanley, G.H.S., 1957. A simple method for the isolation andpurification of total lipides from animal tissues. J. Biol. Chem. 226, 497–509.
Hamid Badawi, G., Yamauchi, Y., Shimada, E., Sasaki, R., Kawano, N., Tanaka, K.,Tanaka, K., 2004. Enhanced tolerance to salt stress and water deficit byoverexpressing superoxide dismutase in tobacco (Nicotiana tabacum)chloroplasts. Plant Sci. 166, 919–928.
Hill, J., Nelson, E., Tilman, D., Polasky, S., Tiffany, D., 2006. Environmental, economic,and energetic costs and benefits of biodiesel and ethanol biofuels. Proc. Natl.Acad. Sci. 103, 11206–11210.
Jeong, H., Lee, J., Cha, M., 2013. Energy efficient growth control of microalgae usingphotobiological methods. Renew. Energ. 54, 161–165.
Kowallik, W., 1963. Die Zellteilung vonChlorella im Verlaufe einer Farblichtkultur.Planta 60, 100–108.
Lee, C.G., Palsson, B.Ø., 1994. High-density algal photobioreactors using light-emitting diodes. Biotechnol. Bioeng. 44, 1161–1167.
Matthijs, H.C., Balke, H., Van Hes, U.M., Kroon, B., Mur, L.R., Binot, R.A., 1996.Application of light-emitting diodes in bioreactors: flashing light effects and
energy economy in algal culture (Chlorella pyrenoidosa). Biotechnol. Bioeng. 50,98–107.
Menon, K.R., Balan, R., Suraishkumar, G., 2013. Stress induced lipid production inChlorella vulgaris: relationship with specific intracellular reactive species levels.Biotechnol. Bioeng. 110, 1627–1636.
Münzner, P., Voigt, J., 1992. Blue light regulation of cell division in Chlamydomonasreinhardtii. Plant physiol. 99, 1370–1375.
Murray, J.W., Duncan, J., Barber, J., 2006. CP43-like chlorophyll binding proteins:structural and evolutionary implications. Trends Plant Sci. 11, 152–158.
Oldenhof, H., Bišová, K., van den Ende, H., Zachleder, V., 2004a. Effect of red and bluelight on the timing of cyclin-dependent kinase activity and the timing of celldivision in Chlamydomonas reinhardtii. Plant Physiol. Biochem. 42, 341–348.
Oldenhof, H., Zachleder, V., van den Ende, H., 2004b. Blue light delays commitmentto cell division in Chlamydomonas reinhardtii. Plant Biol. 6, 689–695.
Oldenhof, H., Zachleder, V., van den Ende, H., 2006. Blue- and red-light regulation ofthe cell cycle in Chlamydomonas reinhardtii (Chlorophyta). Eur. J. Phycol. 41,313–320.
Pirson, A., Kowallik, W., 1964. Spectral responses to light by unicellular plants.Photochem. photobiol. 3, 489–497.
Qian, H., Chen, W., Li, J., Wang, J., Zhou, Z., Liu, W., Fu, Z., 2009. The effect ofexogenous nitric oxide on alleviating herbicide damage in Chlorella vulgaris.Aquat. Toxicol. 92, 250–257.
Rao, M.V., Davis, K.R., 1999. Ozone-induced cell death occurs via two distinctmechanisms in Arabidopsis: the role of salicylic acid. Plant J. 17, 603–614.
Ruyters, G., 1984. Effects of blue light on enzymes. In: Senger, H. (Ed.), Blue lighteffects in Biological Systems. Springer Verlag, Berlin, pp. 283–301.
Sukenik, A., Bennett, J., Falkowski, P., 1987. Light-saturated photosynthesis-limitation by electron transport or carbon fixation. Biochim. Biophys. Acta891, 205–215.
Suzuki, J.Y., Bauer, C.E., 1992. Light-independent chlorophyll biosynthesis:involvement of the chloroplast gene chlL (frxC). Plant Cell 4, 929–940.
Tang, H., Chen, M., Garcia, M., Abunasser, N., Ng, K., Salley, S.O., 2011. Culture ofmicroalgae Chlorella minutissima for biodiesel feedstock production. Biotechnol.Bioeng. 108, 2280–2287.
Thompson, A.S., Rhodes, J.C., Pettman, I., 1988. Natural environmental researchcouncil culture collection of algae and protozoa: catalogue of strains. Freshw.Biol. Ambleside, 164.
Uduman, N., Qi, Y., Danquah, M.K., Forde, G.M., Hoadley, A., 2010. Dewatering ofmicroalgal cultures: a major bottleneck to algae-based fuels. J. Renew. Sust.Energ. 2, 012701.
Wakasugi, T., Nagai, T., Kapoor, M., Sugita, M., Ito, M., Ito, S., Tsudzuki, J., Nakashima,K., Tsudzuki, T., Suzuki, Y., 1997. Complete nucleotide sequence of thechloroplast genome from the green alga Chlorella vulgaris: the existence ofgenes possibly involved in chloroplast division. Proc. Natl. Acad. Sci. 94, 5967–5972.
Wang, C.-Y., Fu, C.-C., Liu, Y.-C., 2007. Effects of using light-emitting diodes on thecultivation of Spirulina platensis. Biochem. Eng. J. 37, 21–25.
Xu, B., Cheng, P., Yan, C., Pei, H., Hu, W., 2013. The effect of varying LED light sourcesand influent carbon/nitrogen ratios on treatment of synthetic sanitary sewageusing Chlorella vulgaris. World J. Microbiol. Biotechnol., 1–12.
Zachleder, V., Kuptsova, E.S., Los, D.A., Cepák, V., Kubín, Š., Shapiguzov, J.M.,Semenenko, V.E., 1989. Division of chloroplast nucleoids and replication ofchloroplast DNA during the cell cycle of Dunaliella salina grown under blue andred light. Protoplasma 150, 160–167.
Zhao, Y.J., Hui, Z., Chao, X., Nie, E., Li, H.J., He, J., Zheng, Z., 2011. Efficiency of two-stage combinations of subsurface vertical down-flow and up-flow constructedwetland systems for treating variation in influent C/N ratios of domesticwastewater. Ecol. Eng. 37, 1546–1554.
Zwietering, M., Jongenburger, I., Rombouts, F., Van’t Riet, K., 1990. Modeling of thebacterial growth curve. Appl. Environ. Microbiol. 56, 1875–1881.
248 D.G. Kim et al. / Bioresource Technology 159 (2014) 240–248





























