Embed Size (px)
Citation preview

Mandibular Corpus Strain in Primates: Further Evidencefor a Functional Link Between Symphyseal Fusionand Jaw-Adductor Muscle Force
WILLIAM L. HYLANDER,1* MATTHEW J. RAVOSA,2CALLUM F. ROSS,3 AND KIRK R. JOHNSON1
1 Department of Biological Anthropology and Anatomy, Duke UniversityMedical Center, Durham, North Carolina 277102 Department of Cell and Molecular Biology, Northwestern UniversityMedical School, Chicago, Illinois 606113 Department of Anatomical Sciences, Health Sciences Center, University ofNew York at Stony Brook, Stony Brook, New York 11794
KEY WORDS symphyseal fusion; in vivo bone strain; functionalmorphology; jaw-adductor muscle force; mastication; primates
ABSTRACT Previous work indicates that compared to adult thick-tailedgalagos, adult long-tailed macaques have much more bone strain on thebalancing-side mandibular corpus during unilateral isometric molar biting(Hylander [1979a] J. Morphol. 159:253–296). Recently we have confirmed inthese same two species the presence of similar differences in bone-strainpatterns during forceful mastication. Moreover, we have also recordedmandibular bone strain patterns in adult owl monkeys, which are slightlysmaller than the galago subjects. The owl monkey data indicate the presenceof a strain pattern very similar to that recorded for macaques, and quiteunlike that recorded for galagos. We interpret these bone-strain patterndifferences to be importantly related to differences in balancing-side jaw-adductor muscle force recruitment patterns. That is, compared to galagos,macaques and owl monkeys recruit relatively more balancing-side jaw-adductor muscle force during forceful mastication.
Unlike an earlier study (Hylander [1979b] J. Morphol. 160:223–240), we areunable to estimate the actual amount of working-side muscle force relative tobalancing-side muscle force (i.e., the W/B muscle force ratio) in these speciesbecause we have no reliable estimate of magnitude, direction, and preciselocation of the bite force during mastication. A comparison of the masticationdata with the earlier data recorded during isometric molar biting, however,supports the hypothesis that the two anthropoids have a small W/B jaw-adductor muscle force ratio in comparison to thick-tailed galagos. These dataalso support the hypothesis that increased recruitment of balancing-sidejaw-adductor muscle force in anthropoids is functionally linked to theevolution of symphyseal fusion or strengthening. Moreover, these data refutethe hypothesis that the recruitment pattern differences between macaquesand thick-tailed galagos are due to allometric factors. Finally, although theevolution of symphyseal fusion in primates may be linked to increased stressassociated with increased balancing-side muscle force, it is currently unclearas to whether the increased force is predominately vertically directed,
Grant sponsor: NIH; Grant number: DE04531; Grant number: DE05595: Grant sponsor: NSF; Grant number: SBR-9420764.*Correspondence to: Professor William L. Hylander, Department of Biological Anthropology and Anatomy, Duke University Medical
School, Box 3170, Durham, NC 27710. E-mail: [email protected] 16 July 1997; accepted 30 July 1998.
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 107:257–271 (1998)
r 1998 WILEY-LISS, INC.

transversely directed, or is a near equal combination of these two forcecomponents (cf. Ravosa and Hylander [1994] In Fleagle and Kay [eds.]:Anthropoid Origins. New York: Plenum, pp. 447–468). Am J Phys Anthropol107:257-271, 1998. r 1998 Wiley-Liss, Inc.
The evolution of symphyseal fusion inmammals, i.e., the complete or partial ossifi-cation of the left and right dentaries, hasoccurred independently in many mamma-lian lineages (cf. Beecher, 1977). Further-more, symphyseal fusion has also evolvednumerous times in various primate lin-eages, and the functional significance of thistrait is a much debated topic.
In 1975 it was suggested that in primatessymphyseal fusion and increased amountsof balancing-side jaw-adductor muscle forceare functionally linked (Hylander, 1975).That is, symphyseal fusion functions to pre-vent structural failure of the symphysis bystrengthening it so as to more effectivelycounter increased amounts of symphysealstress caused by increased recruitment ofbalancing-side jaw-adductor muscle forceduring unilateral mastication and biting (cf.Hylander, 1977; Ravosa and Hylander, 1994).
A few years after hypothesizing that sym-physeal fusion and balancing-side jaw adduc-tor muscle force are linked, it was discoveredthat long-tailed macaques (Macaca fascicu-laris) have about 1.5 to two times more bonestrain along the working-side corpus thanalong the balancing-side corpus, whereasthick-tailed galagos (Otolemur crassicauda-tus) have about seven times more strainalong the working side corpus (Hylander,1979a). It was also demonstrated during thelate 1970s that the unfused symphysis ofthick-tailed galagos is highly mobile with arelatively low concentration of supportiveconnective tissues, and therefore structur-ally is much weaker than the completelyfused symphysis found in all extant anthro-poids1 (Beecher, 1977, 1979).
The differences in strain patterns betweenmacaques and galagos were interpreted toindicate that compared to those prosimian
primates with unfused and structurallyweaker symphyses, anthropoid primates re-cruit relatively more balancing-side jaw-adductor muscle force during unilateral mo-lar biting. These data in and of themselves,however, provided no clue as to the actualamount of working-side adductor muscleforce relative to balancing-side adductormuscle force (i.e., the W/B muscle forceratio) for these species. In that same year ananalysis of bending moments combined withsimultaneous transducer biting data andmandibular bone strain data recorded fromthe lower border of the mandible was used tocalculate a first-approximation of the W/Bratio for vertically directed adductor muscleforce during isometric molar biting(Hylander, 1979b). The data indicated thatfor macaques this ratio is about 1.4 whereasfor galagos it is about 3.0. These data pro-vided further support for the hypothesisthat compared to those prosimians withhighly mobile, unfused, and structurallyweaker symphyses, extant anthropoids re-cruit relatively more balancing-side jaw-adductor muscle force during unilateral iso-metric biting.
Although the mandibular bone strain dataprovide support for the argument that sym-physeal fusion and increased balancing-sidejaw-adductor muscle recruitment patternsare functionally linked, there are at leasttwo problems with this line of support. First,since the galago subjects were very reluc-tant to chew various food items, this argu-ment is based on data recorded during epi-sodes of isometric molar biting on a forcetransducer, rather than during mastication.As primate jaw-adductor muscle recruit-ment patterns during mastication differ con-siderably from episodes of isometric unilat-eral molar biting, and as data recordedduring mastication are arguably more func-tionally relevant, a better (or at least addi-tional) test of what we will refer to as thesymphyseal fusion-muscle recruitment hy-
1Some extant prosimians have rather robust partially-fusedsymphyses (Beecher, 1977, 1979) and there may not be majordifferences in strength between these symphyses and the com-pletely fused symphysis of anthropoids (Ravosa and Hylander,1994).
258 W.L. HYLANDER ET AL.

pothesis involves analyzing mandibular bonestrain patterns recorded during mastica-tion. Second, and more importantly, allome-tric factors may influence jaw-adductormuscle recruitment patterns, and thereforeperhaps the correlation between symphy-seal morphology and the relative amount ofbalancing-side muscle force in macaquesand galagos is a spurious one. The rationalefor the allometric argument is as follows.
Larger mammals may have proportion-ally less total physiological cross-sectionalarea to their jaw-adductor muscles andtherefore relatively less capability to gener-ate adductor muscle force (cf. Scapino, 1981).If so, perhaps larger mammals recruit morebalancing-side jaw-adductor muscle force soas to develop equivalent amounts of biteforce (Scapino, 1981; Hylander, 1985; Ra-vosa, 1991, 1996). Thus, long-tailed ma-caques, which are two to four times largerthan thick-tailed galagos, may simply re-cruit more balancing-side muscle force be-cause of allometric constraints, and if so,symphyseal fusion or strengthening in pri-mates may not be functionally linked to anincrease in balancing-side jaw-adductormuscle force during mastication, but insteadto some other mechanical requirement (cf.Hylander and Johnson, 1997).
The purpose of this paper is to test furtherthe symphyseal fusion-muscle recruitmenthypothesis. We propose to address the appar-ent problems in the earlier study by analyz-ing mandibular mid-corpus strains recordedfrom long-tailed macaques and thick-tailedgalagos during mastication, because, as al-ready noted, this behavior is arguably morefunctionally relevant than isometric molarbiting on a force transducer. We also proposeto test the competing and what we will referto as the allometric constraint-muscle recruit-ment hypothesis. This hypothesis can betested by analyzing jaw-adductor musclerecruitment patterns in the above ma-caques, galagos, and a third primate speciesthat has a relatively weak, highly mobileunfused symphysis and whose body size isequal to or greater than Macaca fascicu-laris. Alternatively, this hypothesis can betested by analyzing jaw-adductor musclerecruitment patterns in macaques and gala-
gos and a primate species that has a com-pletely fused symphysis and whose body sizeis equal to or smaller than adult Otolemurcrassicaudatus.
There are no known extant primate spe-cies that are as large as (or larger than)adult long-tailed macaques and have rela-tively weak, highly mobile unfused symphy-ses.2 There are, however, numerous anthro-poid species that have fused mandibularsymphyses and are as small as (or smallerthan) adult thick-tailed galagos. We haveselected adult owl monkeys (Aotus trivirga-tus), which are slightly smaller than thick-tailed galagos, to serve as experimental sub-jects.
The symphyseal fusion-muscle recruit-ment hypothesis predicts that the two pri-mates with fused symphyses (macaques andowl monkeys) will have relatively high lev-els of balancing-side mid-corpus mandibularbone strain, whereas the greater galagoswill have relatively low levels of balancing-side mid-corpus bone strain. Therefore, thishypothesis predicts that the W/B ratio formandibular strain values are relatively smallin macaques and owl monkeys, and rela-tively large in galagos. In contrast, the allo-metric constraint-muscle recruitment hy-pothesis predicts that the largest species,long-tailed macaques, will have the highestlevels of balancing-side bone strain, the inter-mediate-sized species, thick-tailed galagos,will have intermediate levels of balancing-side bone strain, and the smallest species,owl monkeys, will have the lowest levels ofbalancing-side bone strain. Therefore, theallometric hypothesis predicts that the W/Bratio for mandibular strain values are rela-tively small in macaques, intermediate ingalagos, and relatively large in owl mon-keys.
MATERIALS AND METHODSSubjects
The following primates served as subjects:four adult female (macaques 1, 2, 3, and 5)
2Although some adult indriids are as large as an adultlong-tailed macaque, their unfused (or partially fused) symphy-ses are very well buttressed (Ravosa, 1991) with a dense concen-tration of fibrous connective tissue, some of which is eithercalcified, ossified, or chondrified (Beecher, 1977).
259MANDIBULAR STRAIN AND SYMPHYSEAL FUSION

and one adult male (macaque 4) Macacafascicularis; four adult male (galagos 3, 5, 6,and 7) Otolemur crassicaudatus; and oneadult female (owl monkey 2) and one adultmale (owl monkey 1) Aotus trivirgatus.
Experiments
No new experiments were initiated forthis study. Instead, this study is based on arecent analysis of data originally derivedfrom a large and diverse series of experi-ments that were designed for other pur-poses. For macaques, the data are derivedfrom studies described in Hylander (1979a,1986), Hylander and Crompton (1986), andHylander et al.(1987). For owl monkeys, thedata are derived from studies described inRoss (1993) and Ross and Hylander (1996).For galagos, the data are derived from experi-ments described in Hylander (1979a) andRavosa and Hylander (1993). In one experi-ment rosette strain gages were bonded bilat-erally, i.e., one rosette on the left and one onthe right side of the mandible (macaque 4,exp. E). In the remaining experiments therosette strain gages were bonded unilater-ally (Fig. 1). In four experiments two ro-settes (rosette 1 and rosette 2) were bondedunilaterally along the macaque mandibularcorpus immediately adjacent to one another(macaque 2, exps. A and B; macaque 3, exp.A; macaque 4, exp. A). With the exception ofmacaque 2, Exp. C, all rosettes in all sub-
jects were bonded along the upper or middleportion of the lower half of the mandibularcorpus, i.e., at or slightly below the mid-corpus level. In this exception, the rosettewas bonded very close to the lower border ofthe mandible. Rosette alignment and posi-tion was determined from radiographs and/ordirect measurements during the bondingprocedure.
Rosette bonding and bone strainrecording procedures
The procedures and apparatus for bond-ing miniature 120-ohm stacked rosettes tocortical bone with a cyanoacrylate adhesiveare described elsewhere (Hylander, 1979a,1984, 1986; Hylander and Johnson, 1989).These procedures were performed with thesubjects under general anesthesia using ei-ther fluothane or a combination of nitrousoxide and methoxyflurane (Hylander, 1979a),or the subjects were heavily sedated with acombination of acepromazine and ketamine(Connolly and Quimby, 1978). In all instancesthe surgical site was infiltrated subcutane-ously with a local anesthetic (lidocaine HCl)containing epinephrine (1:100,000) so as toprovide both profound anesthesia and ad-equate hemostasis.
Recording procedure
Each of the strain-gage elements of eachrosette formed one arm of a Wheatstonebridge. Bridge excitation for each elementwas 1 volt. The voltage output from each ofthe strain-gage elements was first condi-tioned and amplified and then ordinarilyrecorded at 15 inches per second with amultiple-channel FM tape recorder. In sev-eral instances, however, the data were re-corded on a six-channel chart recorder(Hylander, 1979a).
Prior to recovery from the general anesthe-sia or sedation, the subject was placed in arestraining chair (macaques) or restrainingsling-suit (galagos and owl monkeys) espe-cially designed to permit normal head, neck,and jaw movements during mastication(Hylander, 1979a; Ross and Hylander, 1996).Once the animal was fully alert, usually 3–6hours later, it was fed pieces of apple skinand hard or tough foods, i.e., monkey chow,
Fig. 1. Drawing of a macaque mandible indicatingthe approximate location of the rosettes. For each ma-caque experiment the rosette was positioned within theperimeter of the oval located below the postcanine teethalong the lower half of the mandible. Rosette position forthe galagos and owl monkeys was confined, due to sizeconsiderations, to the second-molar mid-corpus region.See text for discussion.
260 W.L. HYLANDER ET AL.

dried apricot, popcorn kernels, dried gelatincandy (dried gummy bears) and prune seeds.
Immediately before and after each se-quence of mastication the zero level of strainwas determined when the subject’s man-dible was at rest. The subject was fed vari-ous foods in an intermittent and randomfashion and data were recorded until eithersufficient data were obtained or the animalrefused to eat any additional food. Duringthe recording sessions we noted directly onthe voice track of the tape recorder whatfoods were eaten and whether the subjectchewed on the left or right side (cf. Hylander,1979a and Ross and Hylander, 1996). Chew-ing side identification was accomplished pri-marily by noting the position of the tongueand food bolus during jaw opening, i.e.,noting whether the food was placed on theleft or right side prior to the next powerstroke. Macaques typically chewed on onlyone side throughout an entire sequence. Incontrast, owl monkeys and galagos typicallyengaged in three or four power strokes onone side of the jaw, then shifted to theopposite side for a similar number of powerstrokes, and they continued to repeat thispattern throughout the chewing sequence.At the conclusion of the recording sessionthe animal was sedated, each rosette wasremoved, the surgical incision was resu-tured, an antibiotic was administered, andthe animal was returned to its cage. Healingof the surgical site was uneventful in allsubjects.
Strain analysis
With the exception of data drawn directlyfrom Tables 8 and 9 in Hylander (1979a),which were not recorded on a FM taperecorder, all bone strain recordings werereproduced for visual examination by play-ing the raw strain data from the tape re-corder into a six-channel chart recorder. Asdescribed elsewhere (Hylander and Johnson,1997), the raw strains of all chewing cyclesfor each selected sequence were played si-multaneously from the tape recorder into a16-channel analog-to-digital converter, andthe digitized values were written to the harddisk of a microcomputer. Each channel wassampled and digitized at a rate of 500 Hz
with a channel separation time of 0.125 ms.The digitized data files were used for subse-quent strain calculations and analysis. Foreach food and subject we selected two chew-ing sequences for analysis. For macaquesthis consisted of one sequence of chewing onthe left and one of chewing on the right. Forgalagos and owl monkeys, however, mostsequences contained both left and rightchews. Therefore, when possible, we se-lected one sequence that began with chewson the left and one that began with chews onthe right.
The digitized raw strain values were fil-tered at 40 Hz using a digital low-passButterworth filter. The magnitude of themaximum and minimum principal strains(e1 and e2) and maximum shear strains(gmax) and the direction of e1 were calcu-lated in 2 ms intervals. For purposes ofanalysis, gmax was used as an indicator ofpeak strain (Hylander and Johnson, 1989).Descriptive statistics of e1, e2, and gmaxwere calculated for peak strain, and for the25%, 50% and 75% level of peak strain forboth loading and unloading of each powerstroke of mastication and incision. We alsocalculated the ratio of working-side gmaxvalues/balancing-side gmax values (W/Bstrain) from the peak mean values. For thisstudy we will only report on the descriptivestatistics for the magnitude of gmax and theW/B gmax ratios at peak strain duringmastication.
RESULTS
Typically, some subjects simply refused toeat certain foods. With the exception of onegalago (galago 5) and one macaque (ma-caque 1), all subjects chewed at least one ofthe hard or tough foods. Furthermore, themacaques and owl monkeys readily chewedapple skin. In contrast, none of the galagoschewed apple skin, although they all readilychewed apple pulp. As the only food thatgalago 5 chewed was apple pulp, we willpresent the strain data recorded from thissubject during the chewing of this food. Datarecorded from the other primates during thechewing of apple pulp, however, will not bepresented. As in our most recent studies, we
261MANDIBULAR STRAIN AND SYMPHYSEAL FUSION
will present strain data recorded duringboth the chewing of apple skin and the chew-ing of comparatively hard or tough foods(Hylander and Johnson, 1997). For reasonsto be discussed later, however, we intend tofocus most of our attention on data recordedduring the chewing of hard or tough foods.
Strain magnitude and W/B strain ratios
The largest mandibular strains in all threespecies are associated with the chewing ofhard or tough food items (Tables 1, 2, and 3).Furthermore, with the exception of the oneexperiment where the rosette was posi-
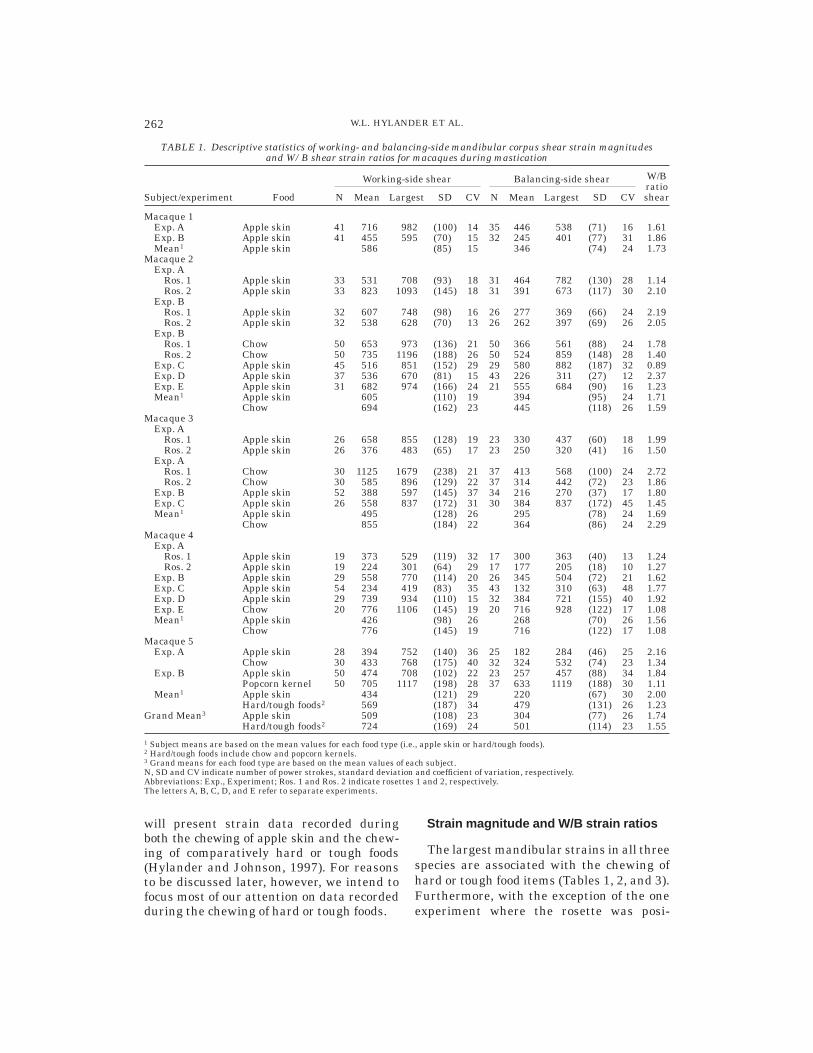
TABLE 1. Descriptive statistics of working- and balancing-side mandibular corpus shear strain magnitudesand W/B shear strain ratios for macaques during mastication
Subject/experiment Food
Working-side shear Balancing-side shear W/BratioshearN Mean Largest SD CV N Mean Largest SD CV
Macaque 1Exp. A Apple skin 41 716 982 (100) 14 35 446 538 (71) 16 1.61Exp. B Apple skin 41 455 595 (70) 15 32 245 401 (77) 31 1.86Mean1 Apple skin 586 (85) 15 346 (74) 24 1.73
Macaque 2Exp. A
Ros. 1 Apple skin 33 531 708 (93) 18 31 464 782 (130) 28 1.14Ros. 2 Apple skin 33 823 1093 (145) 18 31 391 673 (117) 30 2.10
Exp. BRos. 1 Apple skin 32 607 748 (98) 16 26 277 369 (66) 24 2.19Ros. 2 Apple skin 32 538 628 (70) 13 26 262 397 (69) 26 2.05
Exp. BRos. 1 Chow 50 653 973 (136) 21 50 366 561 (88) 24 1.78Ros. 2 Chow 50 735 1196 (188) 26 50 524 859 (148) 28 1.40
Exp. C Apple skin 45 516 851 (152) 29 29 580 882 (187) 32 0.89Exp. D Apple skin 37 536 670 (81) 15 43 226 311 (27) 12 2.37Exp. E Apple skin 31 682 974 (166) 24 21 555 684 (90) 16 1.23Mean1 Apple skin 605 (110) 19 394 (95) 24 1.71
Chow 694 (162) 23 445 (118) 26 1.59Macaque 3
Exp. ARos. 1 Apple skin 26 658 855 (128) 19 23 330 437 (60) 18 1.99Ros. 2 Apple skin 26 376 483 (65) 17 23 250 320 (41) 16 1.50
Exp. ARos. 1 Chow 30 1125 1679 (238) 21 37 413 568 (100) 24 2.72Ros. 2 Chow 30 585 896 (129) 22 37 314 442 (72) 23 1.86
Exp. B Apple skin 52 388 597 (145) 37 34 216 270 (37) 17 1.80Exp. C Apple skin 26 558 837 (172) 31 30 384 837 (172) 45 1.45Mean1 Apple skin 495 (128) 26 295 (78) 24 1.69
Chow 855 (184) 22 364 (86) 24 2.29Macaque 4
Exp. ARos. 1 Apple skin 19 373 529 (119) 32 17 300 363 (40) 13 1.24Ros. 2 Apple skin 19 224 301 (64) 29 17 177 205 (18) 10 1.27
Exp. B Apple skin 29 558 770 (114) 20 26 345 504 (72) 21 1.62Exp. C Apple skin 54 234 419 (83) 35 43 132 310 (63) 48 1.77Exp. D Apple skin 29 739 934 (110) 15 32 384 721 (155) 40 1.92Exp. E Chow 20 776 1106 (145) 19 20 716 928 (122) 17 1.08Mean1 Apple skin 426 (98) 26 268 (70) 26 1.56
Chow 776 (145) 19 716 (122) 17 1.08Macaque 5
Exp. A Apple skin 28 394 752 (140) 36 25 182 284 (46) 25 2.16Chow 30 433 768 (175) 40 32 324 532 (74) 23 1.34
Exp. B Apple skin 50 474 708 (102) 22 23 257 457 (88) 34 1.84Popcorn kernel 50 705 1117 (198) 28 37 633 1119 (188) 30 1.11
Mean1 Apple skin 434 (121) 29 220 (67) 30 2.00Hard/tough foods2 569 (187) 34 479 (131) 26 1.23
Grand Mean3 Apple skin 509 (108) 23 304 (77) 26 1.74Hard/tough foods2 724 (169) 24 501 (114) 23 1.55
1 Subject means are based on the mean values for each food type (i.e., apple skin or hard/tough foods).2 Hard/tough foods include chow and popcorn kernels.3 Grand means for each food type are based on the mean values of each subject.N, SD and CV indicate number of power strokes, standard deviation and coefficient of variation, respectively.Abbreviations: Exp., Experiment; Ros. 1 and Ros. 2 indicate rosettes 1 and 2, respectively.The letters A, B, C, D, and E refer to separate experiments.
262 W.L. HYLANDER ET AL.

tioned very near the lower border of themandible (macaque 2, exp. C), the largestaverage maximum shear strains (gmax) oc-cur along the working side. In addition, theaverage W/B gmax ratios for the two anthro-poid species tend to be much smaller than
the W/B ratios for galagos. This is primarilybecause the average balancing-side strainsin owl monkeys and macaques (836 µe and501 µe, respectively) are very much largerthan those of galagos (216 µe). Species differ-ences in both the absolute strains and in the
TABLE 2. Descriptive statistics of working- and balancing-side mandibular corpus shear strain magnitudesand W/B shear strain ratios for galagos during mastication
Subject/food
Working-side shear Balancing-side shear W/BshearratioN Mean Largest SD CV N Mean Largest SD CV
Galago 3Chow 18 1459 2653 (755) 52 19 330 746 (171) 52 4.42
Galago 5Apple1 36 325 607 (138) 42 23 62 172 (41) 66 5.24
Galago 6Gelatin candy 1 13 1171 1575 (227) 19 10 85 100 (14) 16 13.78Gelatin candy 2 13 839 1082 (231) 28 23 72 110 (18) 25 11.65Dried prune 29 565 1254 (321) 57 11 66 123 (35) 53 8.56Mean2
Hard/tough foods3 858 (260) 35 74 (22) 32 11.33Galago 7
Gelatin candy 1 50 1163 1563 (213) 18 35 227 370 (51) 22 5.12Gelatin candy 2 19 1385 2161 (335) 24 21 259 390 (68) 26 5.35Mean3
Hard/tough foods3 1274 (274) 21 243 (60) 24 5.24Grand mean4
Hard/tough foods3 1197 (430) 36 216 (84) 36 7.001 This subject chewed apple pulp but not apple skin.2 Subject means are based on the mean values for each food type (i.e., apple skin or hard/tough foods).3 Hard/tough foods include gelatin candy, dried prune and chow.4 Grand means for each food type are based on the mean values of each subject.N, SD and CV indicate number of power strokes, standard deviation and coefficient of variation, respectively.Gelatin candy 1 and 2 refer to different chewing sequences associated with this food.
TABLE 3. Descriptive statistics of working- and balancing-side mandibular corpus shear strain magnitudesand W/B shear strain ratios for owl monkeys during mastication
Subject/food
Working-side shear Balancing-side shear W/BratioshearN Mean Largest SD CV N Mean Largest SD CV
Owl Monkey 1Apple skin 22 961 1372 (269) 28 14 889 1343 (288) 32 1.08Gelatin candy 1 30 1263 1629 (208) 16 15 894 1248 (289) 32 1.41Gelatin candy 2 20 1289 1570 (205) 16 19 905 1338 (230) 25 1.42Dried prune 36 1333 2333 (361) 27 26 1236 2197 (409) 33 1.08Mean1
Apple skin 961 (269) 28 889 (288) 32 1.08Hard/tough foods2 1295 (258) 20 1012 (309) 30 1.31
Owl Monkey 2Apple skin 26 504 824 (152) 30 39 377 661 (110) 29 1.34Gelatin candy 50 938 1229 (127) 14 50 711 935 (89) 13 1.32Dried prune 29 793 1105 (187) 24 23 727 956 (107) 15 1.09Dried apricot 43 747 1085 (161) 22 37 543 852 (178) 33 1.38Mean1
Apple skin 504 (152) 30 377 (110) 29 1.34Hard/tough foods2 826 (158) 20 660 (125) 20 1.26
Grand mean3
Apple skin 733 (211) 29 633 (199) 31 1.21Hard/tough foods2 1061 (208) 20 836 (217) 25 1.28
1 Subject means are based on the mean values for each food type (i.e., apple skin or hard/tough foods).2 Hard/tough foods include gelatin candy, dried prune and dried apricot.3 Grand means for each food type are based on the mean values of each subject.N, SD and CV indicate number of power strokes, standard deviation and coefficient of variation, respectively.Gelatin candy 1 and 2 refer to different chewing sequences associated with this food.
263MANDIBULAR STRAIN AND SYMPHYSEAL FUSION
W/B ratios can be readily seen in Tables 1, 2,and 3. For example, the grand mean W/Bratios for owl monkeys and macaques dur-ing the chewing of hard or tough foods areabout 1.3 and 1.6, respectively. In contrast,the grand mean ratio for galagos during thechewing of hard or tough foods is 7.0.
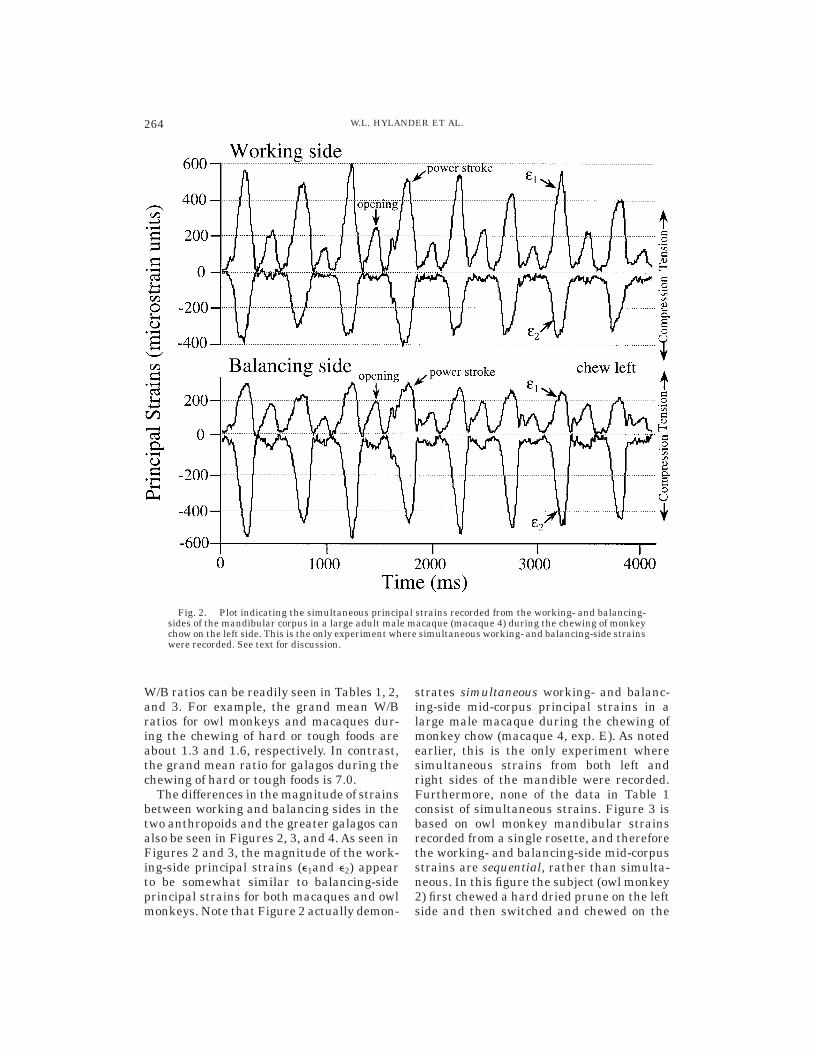
The differences in the magnitude of strainsbetween working and balancing sides in thetwo anthropoids and the greater galagos canalso be seen in Figures 2, 3, and 4. As seen inFigures 2 and 3, the magnitude of the work-ing-side principal strains (e1and e2) appearto be somewhat similar to balancing-sideprincipal strains for both macaques and owlmonkeys. Note that Figure 2 actually demon-
strates simultaneous working- and balanc-ing-side mid-corpus principal strains in alarge male macaque during the chewing ofmonkey chow (macaque 4, exp. E). As notedearlier, this is the only experiment wheresimultaneous strains from both left andright sides of the mandible were recorded.Furthermore, none of the data in Table 1consist of simultaneous strains. Figure 3 isbased on owl monkey mandibular strainsrecorded from a single rosette, and thereforethe working- and balancing-side mid-corpusstrains are sequential, rather than simulta-neous. In this figure the subject (owl monkey2) first chewed a hard dried prune on the leftside and then switched and chewed on the
Fig. 2. Plot indicating the simultaneous principal strains recorded from the working- and balancing-sides of the mandibular corpus in a large adult male macaque (macaque 4) during the chewing of monkeychow on the left side. This is the only experiment where simultaneous working- and balancing-side strainswere recorded. See text for discussion.
264 W.L. HYLANDER ET AL.
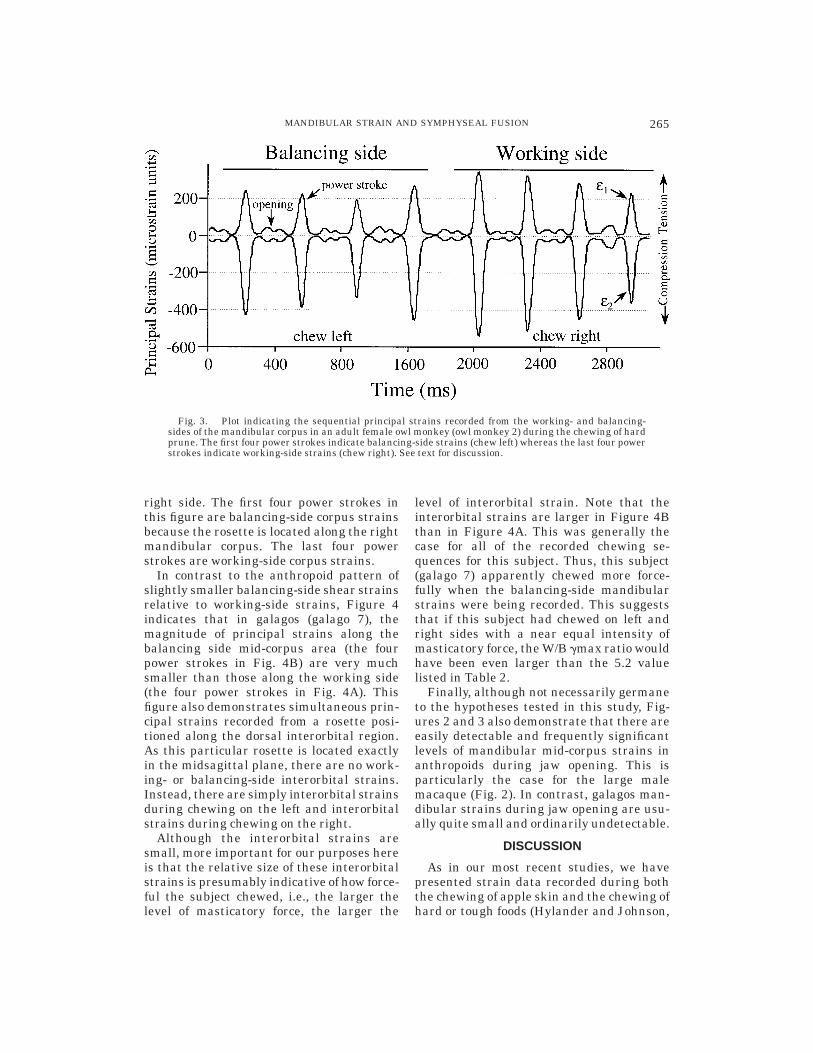
right side. The first four power strokes inthis figure are balancing-side corpus strainsbecause the rosette is located along the rightmandibular corpus. The last four powerstrokes are working-side corpus strains.
In contrast to the anthropoid pattern ofslightly smaller balancing-side shear strainsrelative to working-side strains, Figure 4indicates that in galagos (galago 7), themagnitude of principal strains along thebalancing side mid-corpus area (the fourpower strokes in Fig. 4B) are very muchsmaller than those along the working side(the four power strokes in Fig. 4A). Thisfigure also demonstrates simultaneous prin-cipal strains recorded from a rosette posi-tioned along the dorsal interorbital region.As this particular rosette is located exactlyin the midsagittal plane, there are no work-ing- or balancing-side interorbital strains.Instead, there are simply interorbital strainsduring chewing on the left and interorbitalstrains during chewing on the right.
Although the interorbital strains aresmall, more important for our purposes hereis that the relative size of these interorbitalstrains is presumably indicative of how force-ful the subject chewed, i.e., the larger thelevel of masticatory force, the larger the
level of interorbital strain. Note that theinterorbital strains are larger in Figure 4Bthan in Figure 4A. This was generally thecase for all of the recorded chewing se-quences for this subject. Thus, this subject(galago 7) apparently chewed more force-fully when the balancing-side mandibularstrains were being recorded. This suggeststhat if this subject had chewed on left andright sides with a near equal intensity ofmasticatory force, the W/B gmax ratio wouldhave been even larger than the 5.2 valuelisted in Table 2.
Finally, although not necessarily germaneto the hypotheses tested in this study, Fig-ures 2 and 3 also demonstrate that there areeasily detectable and frequently significantlevels of mandibular mid-corpus strains inanthropoids during jaw opening. This isparticularly the case for the large malemacaque (Fig. 2). In contrast, galagos man-dibular strains during jaw opening are usu-ally quite small and ordinarily undetectable.
DISCUSSION
As in our most recent studies, we havepresented strain data recorded during boththe chewing of apple skin and the chewing ofhard or tough foods (Hylander and Johnson,
Fig. 3. Plot indicating the sequential principal strains recorded from the working- and balancing-sides of the mandibular corpus in an adult female owl monkey (owl monkey 2) during the chewing of hardprune. The first four power strokes indicate balancing-side strains (chew left) whereas the last four powerstrokes indicate working-side strains (chew right). See text for discussion.
265MANDIBULAR STRAIN AND SYMPHYSEAL FUSION

1997). The data recorded during the chew-ing of apple skin facilitate the direct compari-son of mandibular corpus strains with strainsrecorded from other regions of the face dur-ing the chewing of apple skin (e.g., Hylanderet al., 1991). Furthermore, these data alsofacilitate the interspecific comparison ofmandibular corpus strains as presumablythe mechanical properties of apple skin arevirtually identical and therefore the chew-ing of apple skin requires similar levels ofmasticatory force prior to swallowing. Forthis study, however, we intend to focus ourattention largely on data recorded duringthe chewing of hard or tough foods for thefollowing reasons.
First, as already noted, none of the gala-gos chewed apple skin. Second, data associ-ated with the chewing of hard or tough foodsare important because structural adapta-tions to counter masticatory stress are pre-sumably in response to the more forcefulmasticatory loading regimes. Third, and
most importantly, earlier work has estab-lished that W/B masseter muscle force ratiosare frequently influenced by overall mastica-tory force levels (Hylander et al., 1992;Hylander and Johnson, 1997). In most ma-caques the lowest W/B masseter force ratios(which begin to approach 1.0) are associatedwith the more forceful power strokes. Thus,as the magnitude of masticatory force levelsfrequently influence W/B ratios for jaw-adductor muscle force, in a comparativestudy such as this all subjects should berequired to chew in a relatively forcefulmanner so as to insure maximum recruit-ment levels of balancing-side jaw-adductormuscle force.
A test of the allometric constraint-and symphyseal fusion-muscle
recruitment hypotheses
The data presented in Tables 1, 2, and 3clearly refute the allometric constraint-muscle recruitment hypothesis and strongly
Fig. 4. Plot indicating sequential and simultaneousprincipal strains recorded from the interorbital regionand the right side of the mandibular corpus in an adultmale thick-tailed galago (galago 7) during the chewingof hard prune. A: Working-side mandibular strains(chew right). B: Balancing-side mandibular strains (chewleft). Note that the interorbital strains are much larger
during chewing on the left than chewing on the right,indicating that for these chewing cycles the most force-ful chews are associated with chewing on the left. Inspite of this outcome, the balancing-side strains forchewing on the left are still much smaller than theworking-side strains during chewing on the right. Seetext for discussion.
266 W.L. HYLANDER ET AL.

support the symphyseal fusion-muscle re-cruitment hypothesis in that compared togalagos, 1) the average amount of balancing-side strain is much larger in both macaquesand owl monkeys, and 2) the W/B strainratios for owl monkeys and macaques arevery similar to one another, i.e., about 1.3and 1.6, respectively, whereas the W/B strainratio for thick-tailed galagos is much larger,i.e., 7.0. Furthermore, the W/B strain ratiosfor macaques and galagos during mastica-tion are very similar to those during isomet-ric transducer biting (1.8 and 7.1, respec-tively) (Hylander, 1979a).
Allometry, muscle recruitment,and mandibular strain
The observations that the smallest andlargest primates in this sample (owl mon-keys and macaques, respectively) have W/Bstrain ratio values more similar to one an-other, and the intermediate-sized galago spe-cies has W/B strain ratios considerably largerindicates that allometric constraints, as out-lined in the Introduction, are of negligibleimportance within this particular group ofprimates. Moreover, even when consideringthe W/B strain ratios for only the two anthro-poids, there is no evidence to support theallometric constraint hypothesis because thishypothesis predicts that the W/B strain ra-tios should be smallest in macaques. In-stead, the data indicate that the smallestoverall ratios are found in the owl monkeys(hard/tough foods: about 1.6 and 1.3 formacaques and owl monkeys; apple skin:about 1.7 and 1.2 for macaques and owlmonkeys).
A comparison of interspecific mandibularstrains during the chewing of certain foods,however, reveals possible allometric effects.Presumably the dried gelatin candies eatenby two of the galagos (galagos 6 and 7) andthe two owl monkeys have very similarmechanical properties. If so, as the mechani-cal properties of food items largely deter-mine how forceful an animal must chew orbite so as to break this item down, presum-ably the level of bite force distributed alongpremolar and molar shearing crests andcrushing and grinding surfaces in these twospecies are more or less the same during thechewing of this food. As the owl monkeymandible is about the same size as (or
perhaps slightly smaller than) the galagomandible and the gelatin candy eaten bythese two species is the same size and shape,it is not surprising that the average working-side mandibular strains for this food aresimilar (owl monkeys, 1107 µe; galagos, 1140µe). There is, however, about five times morestrain along the balancing-side corpus inowl monkeys than in galagos (805 µe and161 µe, respectively). Rather than beinglinked to allometric constraints, these largedifferences in balancing-side strains arelikely functionally linked to differences inbalancing-side jaw-adductor muscle forceand symphyseal morphology.
Although the hard and/or tough foodseaten by macaques (popcorn kernels andmonkey chow) differ in their mechanicalproperties from those eaten by owl monkeys(gelatin candy and dried prunes and apri-cots), presumably the mechanical propertiesof the apple skin that was eaten by these twospecies are very similar. The average work-ing-side mandibular strains during apple-skin chewing in macaques (509 µe) aresmaller than those recorded from owl mon-keys (733 µe). On average during the chew-ing of apple skin the working-side (andbalancing-side) mandibular strains in ma-caques are about 69% (and 48%) of thecorresponding values recorded from owl mon-keys. Presumably these differences in meanvalues suggest that the owl monkey man-dible is strained and stressed significantlymore than the macaque mandible duringthe chewing of apple skin, although there isconsiderable overlap in strain values be-tween these two species. Furthermore, thesedifferences are probably partially due tomasticatory forces of similar magnitude act-ing on mandibles that are very dissimilar insize. It is also possible, however, that thesestrain differences are also influenced by therelative size of the ingested food items. Aswe did not adjust or scale the relative size ofthe pieces of apple skin offered to the varioussubjects, we are unable to partition whatportion of the differences in strain are due tomandibular size differences, and what por-tion is due to initial food size differences.
Finally, there is at least one additionalallometric factor that influences balancing-side jaw-adductor recruitment patterns.Compared to small free-ranging mamma-
267MANDIBULAR STRAIN AND SYMPHYSEAL FUSION

lian herbivores, large free-ranging herbi-vores tend to eat a greater proportion offoods composed of hard and/or tough difficult-to-chew materials (Bell, 1971; Rensberger,1973). A literature survey extends this find-ing to extant primates (Kay, 1973; Kay andHylander, 1978; Beecher, 1983; Ravosa, 1991,1996). As demonstrated experimentally inmacaques, a dietary shift to harder and/ortougher foods is likely associated with notonly an absolute increase in total mastica-tory force, but a disproportionate increase inthe recruitment of balancing-side jaw-adduc-tor muscle force relative to working-side jawmuscle force (Hylander, 1979a; Hylander etal., 1992). Thus, although allometric con-straints may indeed influence balancing-side muscle recruitment patterns, this mayhave more to do with the mechanical proper-ties of selected food items, and less so withthe constraints on jaw-muscle force produc-tion associated with increasing body size.This food selection factor, as argued else-where, may be importantly related to whysymphyseal fusion is associated with largebody size in some mammalian lineages(Scapino, 1981; Ravosa and Hylander, 1994).
Increased vertical and transverse muscleforce and symphyseal fusion
If symphyseal fusion in primates is indeedfunctionally linked to an increase in balanc-ing-side jaw-adductor muscle force, is thislinkage confined mainly to the vertical com-ponent of jaw-adductor muscle force, is itconfined mainly to the transverse compo-nent of this muscle force, or are both forcecomponents of near equal importance? Al-though we are unable to answer these inter-related questions at this time, there areimportant implications associated with thesequestions as these two force componentstend to load the symphysis in two verydifferent ways.
An increase in vertically directed jaw-adductor muscle force causes an increase invertically directed bite force, i.e., an increasein crushing or vertical shear along the teeth.Furthermore, as vertical force from the bal-ancing-side muscles is transferred to theworking side, this causes an increase indorsoventral shear stress throughout thesymphysis, as well as increased bending of
the symphysis due to twisting of the balanc-ing-side mandibular corpus about its longaxis (Hylander, 1984, 1985). Thus, perhapssymphyseal fusion in primates is an adap-tive response to counter some combinationof dorsoventral shear and bending of thesymphysis (cf. Hylander, 1975, 1979a, 1979b;Beecher, 1977, 1983; Ravosa, 1991, 1996;Ravosa and Hylander, 1994; and Ravosa andSimons, 1994), and if so, fusion is function-ally linked to increasing the amount of verti-cally directed bite force, i.e., increased crush-ing and vertical shearing along thepostcanine dentition.
In contrast to the consequences of anincrease in vertically directed jaw-adductormuscle force, an increase in transverselydirected muscle force causes an increase inthe transverse component to the bite force,i.e., an increase in grinding or transverseshear along the teeth. Moreover, as in-creased transversely directed muscle force isalso transferred across the symphysis fromthe balancing- to the working-side corpus,this causes an increase in lateral transversebending or wishboning of the symphysis(Hylander and Johnson, 1994). Thus, per-haps symphyseal fusion in primates is anadaptive response to counter wishboning ofthe symphysis (Ravosa and Hylander, 1994),and if so, fusion is functionally linked toincreasing the amount of transversely di-rected bite force, i.e., increased grinding orvertical shearing along the postcanine denti-tion.
Unfortunately our strain data are unableto resolve the issue regarding the relativeimportance of these force components onsymphyseal fusion. Although symphysealstrains in macaques have been partiallycharacterized, indicating that the macaquemandibular symphysis experiences varyingdegrees of dorsoventral shear, bending dueto twisting of the mandibular corpora abouttheir long axes, and wishboning (Hylander,1984, 1985), we have no symphyseal straindata for galagos or owl monkeys. Despitethis lack of data, however, we can reason-ably assume that increased balancing-sidejaw-adductor activity in these two speciescontributes to increased vertical bite forcemagnitudes (cf. Hylander, 1977), and there-fore this increased activity causes varying
268 W.L. HYLANDER ET AL.

amounts of dorsoventral shear along theirsymphyses. In addition, due to the geometryof jaw-adductor muscles in galagos and owlmonkeys, it is also likely that their symphy-ses are bent due to twisting of the mandibu-lar corpora about their long axes (cf.Hylander, 1979a). What we do not know,however, is whether the mandibular symphy-sis of owl monkeys and galagos experienceswishboning.
As noted earlier, although we originallycalculated descriptive statistics for straindirections, these data are not reported here.This is because doing so would have lengthedthis manuscript considerably while duplicat-ing much of what has already been pub-lished (Hylander, 1979a; Hylander et al.,1987). Furthermore, and more importantly,these data are unable to help resolve theissue regarding symphyseal loading pat-terns in galagos and owl monkeys. This isbecause due to the small size of the galagoand owl monkey mandibles and the rela-tively large size of the strain gage rosettes,we were forced to record strain from a veryrestricted area of the corpus in these sub-jects. This in turn meant that we wereunable to test various hypotheses regardingloading patterns of the corpus by examiningchanges in strain directions relative to alter-ations in rosette position. Although our lim-ited directional data on owl monkeys andgalagos are consistent with the hypothesesthat their working-side corpora are directlysheared and twisted about their long axeswhereas their balancing-side corpora aredirectly sheared, twisted about their longaxes, and bent in the parasagittal plane, it isunclear as to whether or not the owl monkeyand galago mandible experiences wishbon-ing.
We intend to pursue the issue of symphy-seal loading indirectly by doing a detailedEMG analysis of the superficial and deepportions of the masseter muscle during thechewing of hard and/or tough foods in ba-boons, macaques, thick-tailed galagos andowl monkeys (Hylander, Ravosa, Ross, Wall,and Johnson, n.d.). This analysis will enableus to determine the recruitment patterns ofone portion of the jaw-adductor muscle massthat has a predominantly vertical orienta-tion, the superficial masseter, and one with
a relatively larger and arguably predomi-nant transverse orientation, the deep masse-ter. These data should give us additionalinsights as to whether there are importantand major differences in recruitment pat-terns in these muscles, and therefore insymphyseal loading patterns, within theseprimates. We are particularly interested inthe relative magnitude of working- and bal-ancing-side jaw-adductor muscles, i.e., W/Bratios of superficial and deep masseter EMGactivity, as well as differential timing pat-terns of the various parts of the deep andsuperficial masseter. Most notably, duringthe power stroke of mastication, do onlyanthropoids have relatively small W/B ra-tios for the superficial and/or deep masse-ters, and do owl monkeys, but not galagos,exhibit the late activity of the balancing-sidedeep masseter which has been shown tocontribute to wishboning stresses in thesymphysis of macaques and baboons(Hylander and Johnson, 1994)?
Finally, although symphyseal fusion hasevolved independently in many mammalianlineages, we have intentionally focused ourattention on the evolution of symphysealfusion in primates. This is primarily for theobvious reason that our data are drawnexclusively from various species of primates.There is, however, at least one notable prob-lem with the hypothesis linking symphysealfusion or strengthening and increased bal-ancing-side muscle force. EMG data re-corded from the jaw adductors of the NorthAmerican opossum (Didelphis marsupialis)suggest that during mastication, the overallbalancing-side muscle force is perhaps aslarge as the working-side muscle force(Crompton, 1985, 1995; and Crompton andHylander, 1986). The reason such a findingis a problem for this hypothesis is simplybecause opossums have a mobile unfusedsymphysis, rather than a rigid fully fusedsymphysis.
Similar to our EMG study of primate jawmuscles, a quantitative analysis of peakEMG activity of the superficial and deepmasseter in adult opossums is currently inprogress (Hylander, Johnson, and Cromp-ton, n.d.). If this analysis confirms thatjaw-muscle recruitment patterns in opos-sums are more similar to anthropoid pri-
269MANDIBULAR STRAIN AND SYMPHYSEAL FUSION

mates than they are to greater galagos, thiswill indicate that symphyseal strengthening(fusion) and increased balancing-side muscleforce are not functionally linked in all mam-malian lineages.
CONCLUSIONS
This study tested two competing hypoth-eses about recruitment patterns of balancing-side jaw-adductor muscles in primates dur-ing mastication. One hypothesis suggeststhat increased balancing-side jaw-adductorforce is functionally linked to the evolutionof symphyseal strengthening or fusion,whereas the other claims that this forceincrease is due to allometric constraintsassociated with the evolution of increasingbody size. The mandibular bone strain datarecorded from long-tailed macaques, thick-tailed galagos, and owl monkeys, clearlyrefute the allometric constraint-muscle re-cruitment hypothesis and strongly supportthe symphyseal fusion-muscle recruitmenthypothesis. The data indicate that anthro-poids both larger and smaller than thick-tailed galagos have balancing-side muscleforce recruitment patterns that are quitesimilar to one another, and both are quiteunlike the pattern found in galagos.
Currently it is unclear as to whethersymphyseal fusion in primates is function-ally linked to an increase in vertically di-rected or transversely directed muscle andbite forces. If it is linked to increased verti-cally directed forces, then presumably sym-physeal fusion is an adaptive response tocounter some combination of dorsoventralshear and bending of the symphysis due totwisting of the mandibular corpora abouttheir long axes. If it is linked to transverselydirected forces, than presumably symphy-seal fusion is an adaptive response to counterlateral transverse bending or wishboning.
ACKNOWLEDGMENTS
This study was supported by the Depart-ment of Biological Anthropology andAnatomy, Duke University. It was also sup-ported by research grants from NIH (MERITAward DE04531 to WLH), (DE05595 to MJRand WLH) and NSF (SBR-9420764 to WLH)(BNS-91–00523 to CFR and WLH). We thankDr. Christine E. Wall for her significant help
on various aspects of this manuscript. Wealso thank Dr. Don Schmechel for allowingus to work with the thick-tailed galagos inhis care.
LITERATURE CITED
Beecher RM (1977) Function and fusion at the mandibu-lar symphysis. Am. J. Phys. Anthropol. 47:325–336.
Beecher RM (1979) Functional significance of the man-dibular symphysis. J. Morphol. 159:117–130.
Beecher RM (1983) Evolution of the mandibular symphy-sis in Notharctinae (Adapidae, Primates). Int. J.Primatol. 4:99–112.
Bell RHV (1971) A grazing ecosystem in the Serengeti.Scientific Am. 225:86–93.
Connolly R, and Quimby FW (1978) Acepromazine-ketamine anesthesia in the rhesus monkey (Macacamulatta). Lab. Anim. Sci. 28:72–74.
Crompton AW (1985) Origin of the mammalian temporo-mandibular joint. In DS Carlson, JA McNamara, andKA Ribbons (eds.): Developmental Aspects of Temporo-mandibular Joint Disorders. Monograph #16, Cranio-facial Growth Series, Center for Human Growth andDevelopment, University of Michigan, Ann Arbor,Michigan, pp. 1-18.
Crompton AW (1995) Masticatory function in nonmam-malian cynodonts and early mammals. In JJ Thoma-son (ed.): Functional Morphology in Vertebrate Paleon-tology, Cambridge: Cambridge University Press, pp.55-75.
Crompton AW, and Hylander WL (1986) Changes inmandibular function following the acquisition of adentary-squamosal jaw articulation. In N Hotten III,PD Maclean, JJ Roth and EC Roth (eds.): The Ecologyand Biology of Mammal-like Reptiles, WashingtonD.C.: Smithsonian Press, pp. 263-282.
Hylander WL (1975) Incisor size and diet in anthropoidswith special reference to cercopithecidae. Science 189:1095–1098.
Hylander WL (1977) In vivo bone strain in the mandibleof Galago crassicaudatus. Am. J. Phys. Anthropol.46:309–326.
Hylander WL (1979a) Mandibular function in Galagocrassicaudatus and Macaca fascicularis: An in vivoapproach to stress analysis of the mandible. J. Mor-phol. 159:253–296.
Hylander WL (1979b) The functional significance ofprimate mandibular form. J. Morphol. 160:223–240.
Hylander WL (1984) Stress and strain in the mandibu-lar symphysis of primates: A test of competing hypoth-eses. Am. J. Phys. Anthropol. 64:1–46.
Hylander WL (1985) Mandibular function and biome-chanical stress and scaling. Am. Zool. 25:315–330.
Hylander WL (1986) In vivo bone strain as an indicatorof masticatory bite force in Macaca fascicularis. Arch.Oral Biol. 31:149–157.
Hylander WL, and Crompton AW (1986) Jaw move-ments and patterns of mandibular bone strain duringmastication in the monkey Macaca fascicularis. Arch.Oral Biol. 31:841–848.
Hylander WL, and Johnson KR (1989) The relationshipbetween masseter force and masseter electromyo-gram during mastication in the monkey Macaca fas-cicularis. Arch. Oral Biol. 34:713–722.
Hylander WL, and Johnson KR (1993) Modelling rela-tive masseter force from surface electromyogramsduring mastication in non-human primates. ArchOral Biol. 38:233–240.
Hylander WL, and Johnson KR (1994) Jaw musclefunction and wishboning of the mandible during mas-
270 W.L. HYLANDER ET AL.

tication in macaques and baboons. Am. J. Phys.Anthropol. 94:523–547.
Hylander WL, and Johnson KR (1997) In vivo bonestrain patterns in the zygomatic arch of macaques andthe significance of these patterns for functional inter-pretations of craniofacial form. Am. J. Phys. Anthro-pol. 102:203–232.
Hylander WL, Johnson KR, and Crompton AW (1987)Loading patterns and jaw movements during mastica-tion in Macaca fascicularis: A bone-strain, electromyo-graphic, and cineradiographic analysis. Am. J. Phys.Anthropol. 72:287–314.
Hylander WL, Johnson KR, and Crompton AW (1992)Muscle force recruitment and biomechanical model-ing: An analysis of masseter muscle function in Ma-caca fascicularis. Am. J. Phys. Anthropol. 88:365–387.
Hylander WL, Johnson KR, and Crompton AW (1998)Masseter force in the North American opossum: Test-ing the symphyseal fusion-muscle recruitment hypoth-esis. Arch. Oral Biol. (in review).
Hylander WL, Picq PG, and Johnson KR (1991) Mastica-tory-stress hypotheses and the supraorbital region ofprimates. Am. J. Phys. Anthropol. 86:1–36.
Hylander WL, Ravosa MJ, Ross CF, Wall CE, andJohnson KR (1997) The relationship between symphy-seal fusion and jaw-adductor muscle force in pri-mates: An EMG study. Am. J. Phys. Anthropol. (inreview).
Kay RF (1973) Mastication, molar tooth structure anddiet in primates. Ph.D. Thesis, Yale University.
Kay RF, and Hylander WL (1978) The dental structureof mammalian folivores with special reference toprimates and phalangeroidea (Marsupialia). In G
Montgomery (ed.): The Ecology of Arboreal Folivores.Washington, D.C.: Smithsonian Institution Press, pp.173–191.
Ravosa MJ (1991) Structural allometry of the prosimianmandibular corpus and symphysis. J. Hum. Evol.20:3–20.
Ravosa MJ (1996) Mandibular form and function inNorth American and European Adapidae and Omomy-idae. J. Morphol. 229:171–196.
Ravosa MJ, and Hylander WL (1993) Functional analy-sis of the galago circumorbital region. Am. J. Phys.Anthropol. 90:Suppl. 16:168.
Ravosa MJ, and Hylander WL (1994) Function andfusion of the mandibular symphysis in primates:stiffness or strength? In JG Fleagle and RF Kay (eds.):Anthropoid Origins. New York: Plenum, pp. 447–468.
Ravosa MJ, and Simons EL (1994) Mandibular growthand function in Archeolemur. Am. J. Phys. Anthropol.95:63–76.
Rensberger JM (1973) Sanctimus (Mammalia, Roden-tia) and the phyletic relationships of the large arika-reean geomyoids. J. Paleontol. 47:835–853.
Ross CF (1993) The Functions of the Postorbital Septumand Anthropoid Origins. Ph.D. Thesis, Duke Univer-sity.
Ross CF, and Hylander WL (1996) In vivo and in vitrobone strain in the owl monkey circumorbital regionand the function of the postorbital septum. Am. J.Phys. Anthropol. 101:183–215.
Scapino RP (1981) Morphological investigations intofunctions of the jaw symphysis in carnivorans. J.Morphol. 167:339–375.
271MANDIBULAR STRAIN AND SYMPHYSEAL FUSION





























