Embed Size (px)
Citation preview

doi:10.1152/ajpcell.00446.2010 300:C466-C476, 2011. First published 15 December 2010;Am J Physiol Cell Physiol
Teresa D. Tetley, Miguel C. Seabra and Alistair N. HumeGiulia Bolasco, Dhani C. Tracey-White, Tanya Tolmachova, Andrew J. Thorley,epithelium structure in miceLoss of Rab27 function results in abnormal lung
You might find this additional info useful...
for this article can be found at:Supplemental materialhttp://ajpcell.physiology.org/content/suppl/2011/03/03/ajpcell.00446.2010.DC1.html
32 articles, 16 of which can be accessed free at:This article cites http://ajpcell.physiology.org/content/300/3/C466.full.html#ref-list-1
including high resolution figures, can be found at:Updated information and services http://ajpcell.physiology.org/content/300/3/C466.full.html
can be found at:AJP - Cell Physiologyabout Additional material and information http://www.the-aps.org/publications/ajpcell
This infomation is current as of April 25, 2011.
American Physiological Society. ISSN: 0363-6143, ESSN: 1522-1563. Visit our website at http://www.the-aps.org/.a year (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2011 by the
is dedicated to innovative approaches to the study of cell and molecular physiology. It is published 12 timesAJP - Cell Physiology
on April 25, 2011
ajpcell.physiology.orgD
ownloaded from

Loss of Rab27 function results in abnormal lung epithelium structure in mice
Giulia Bolasco,1 Dhani C. Tracey-White,1 Tanya Tolmachova,1 Andrew J. Thorley,2 Teresa D. Tetley,2
Miguel C. Seabra,1,3,4 and Alistair N. Hume1
1Molecular Medicine, 2Pharmacology and Toxicology, National Heart and Lung Institute, Imperial College London, London,United Kingdom; 3Instituto Gulbenkian de Ciência, Oeiras; and 4Faculdade de Ciências Médicas, Universidade Nova deLisboa, Lisboa, Portugal
Submitted 3 November 2010; accepted in final form 13 December 2010
Bolasco G, Tracey-White DC, Tolmachova T, Thorley AJ,Tetley TD, Seabra MC, Hume AN. Loss of Rab27 function resultsin abnormal lung epithelium structure in mice. Am J Physiol CellPhysiol 300: C466–C476, 2011. First published December 15, 2010;doi:10.1152/ajpcell.00446.2010.—Rab27 small GTPases regulate se-cretion and movement of lysosome-related organelles such as T cellcytolytic granules and platelet-dense granules. Previous studies indicatedthat Rab27a and Rab27b are expressed in the murine lung suggesting thatthey regulate secretory processes in the lung. Consistent with thosestudies, we found that Rab27a and Rab27b are expressed in cell types thatcontain secretory granules: alveolar epithelial type II (AEII) and Claracells. We then used Rab27a/Rab27b double knockout (DKO) mice toexamine the functional consequence of loss of Rab27 proteins in themurine lung. Light and electron microscopy revealed a number ofmorphological changes in lungs from DKO mice when compared withthose in control animals. In aged DKO mice we observed atrophy of thebronchiolar and alveolar epithelium with reduction of cells numbers,thinning of the bronchiolar epithelium and alveolar walls, and enlarge-ment of alveolar airspaces. In these samples we also observed increasednumbers of activated foamy alveolar macrophages and granulocyte con-taining infiltrates together with reduction in the numbers of Clara cellsand AEII cells compared with control. At the ultrastructural level weobserved accumulation of cytoplasmic membranes and vesicles in Claracells. Meanwhile, AEII cells in DKO accumulated large mature lamellarbodies and lacked immature/precursor lamellar bodies. We hypothesizethat the morphological changes observed at the ultrastructural level inDKO samples result from secretory defects in AEII and Clara cells andthat over time these defects lead to atrophy of the epithelium.
Rab27 proteins; intracellular transport; alveolar epithelium type IIcell; Clara cell
SMALL GTPASES of the Rab family regulate intracellular transportin eukaryotic cells (5, 22, 24). Analyses of mammalian ge-nomes indicate that this family comprises �60 members thathave intracellular compartment-specific localization and func-tion. Rabs act as molecular switches cycling between an activeGTP and inactive GDP bound states. Active Rabs recruiteffectors to the membrane upon which they reside, and thesetransduce functions in vesicle formation, transport, docking, orfusion. After nucleotide hydrolysis, inactive Rab is then re-trieved to the cytosol in complex with Rab GDP dissociationinhibitor (GDI) in readiness for reutilization in further roundsof transport. Nascent Rabs are posttranslationally modified bycovalent attachment of geranylgeranyl groups to carboxyl-terminus cysteine residues, which is essential for their targetingto specific intracellular membranes.
While a subset of “housekeeping” Rabs are expressed ubiq-uitously, Rab27 GTPases (Rab27a and Rab27b) are expressedin cell types with specialized secretory functions (6, 29). Inthese cells, Rab27 localizes to, and regulates, the motility andexocytosis of lysosome-related organelles (LROs) and secre-tory granules; e.g., melanosomes in melanocytes, lytic granulesin cytotoxic T cells (CTL), and dense granules in platelets (1,8, 11, 25, 28, 32). To facilitate these diverse functions Rab27interacts with eleven known downstream effectors (5, 13). Forinstance, in melanocytes, we and others (9, 21) have shownthat Rab27a-GTP sequentially recruits effector Melanphilin/Slac2-a and molecular motor myosin 5a to melanosomesthereby allowing their retention in actin-rich peripheral den-drites. Meanwhile, in CTLs, Rab27a mediates granule dockingand exocytosis via effector Munc13–4 (17). Differential en-gagement of effectors is in part due to their cell type-specificpattern of expression (10). In some cell types, Rab27a andRab27b isoforms are expressed together, and recent data sug-gest that they may perform sequential roles in granule dockingand exocytosis via engagement of different effectors (14, 20).
Previous analysis revealed that Rab27 proteins are expressedin the murine lung, suggesting that they play an important rolein lung cell types with specialized secretory functions (6, 29).Within the conducting airways (bronchioles), Clara cells per-form an important role in production and release of lung liningfluid components; e.g., surfactant protein A, Clara cell-specificprotein (CCSP). While in the gas exchange, (alveolar) epithe-lium type II (AEII) cells perform a similar function producingsurfactant protein and phospholipid components (3, 15). Liningfluid proteins and lipids perform an essential role in the innateimmune response to invading pathogens; e.g., defensins andcollectins, and reduction of surface tension that maintains thealveolar structure; e.g., surfactant proteins and lipids. Interest-ingly, within AEII cells a subset of surfactant components arestored in lysosome-related organelles (LROs), known as lamel-lar bodies (LB), that release their cargo into the airspace uponstimulation (4).
Several studies provide evidence for an important role forRab GTPases in the physiological function of the lung. Forexample, Rab3D has been found to localize to a population ofLBs in AEII cells, although its precise function in this contextremains unknown (30). Rab14, meanwhile, was found partiallylocalized to LBs, and knockdown resulted in partial inhibitionof their evoked release suggesting a role for Rab14 in thisprocess (7). Most recently, analysis of the chocolate mutantmouse and the ruby mutant rat that contain Rab38 genemutations revealed a role for this protein in surfactant exocy-tosis and LB structure (18, 19). In this study, we investigatedthe cell-type specific and intracellular localization of Rab27
Address for reprint requests and other correspondence: A. N. Hume, Schoolof Biomedical Sciences, Queen’s Medical Centre, Nottingham NG7 2UH, UK(e-mail: [email protected]).
Am J Physiol Cell Physiol 300: C466–C476, 2011.First published December 15, 2010; doi:10.1152/ajpcell.00446.2010.
0363-6143/11 Copyright © 2011 the American Physiological Society http://www.ajpcell.orgC466
on April 25, 2011
ajpcell.physiology.orgD
ownloaded from

proteins in the murine lung as well as the effects of loss ofRab27 upon morphology of the pulmonary epithelium.
MATERIALS AND METHODS
Chemicals and antibodies. Unless otherwise stated all chemicalswere obtained from Sigma-Aldrich (Poole, UK). The following anti-bodies were used for immunoblotting: rabbit polyclonals anti-Rab27aand anti-Rab27b raised against the COOH-terminus hypervariableregion of each protein at 1:1,000 (Davids Biotechnologie, Regens-berg, Germany) and rabbit anti-Calnexin Stressgen (SPA-860D, Bio-Whittaker; Wokingham, UK) at 1:10,000. For immunohistochemistry,rabbit anti-pro-surfactant protein C (SPC) antibody (Abcam ab28744,Cambridge, UK) was used at 1:1,000, rabbit anti-Clara cell-specificprotein (CCSP) (UK cat no. 07-623, Upstate, Dundee) was at 1:1,000,goat anti-SPC (Research Diagnostics RTSURFCCabG) was at 1:100,and rabbit polyclonal anti-Rab27a and anti-Rab27b antibodies (asabove) were each at 1:100.
Mouse strains. All mice were treated humanely and in accordancewith the UK Home Office regulations under project license PPL70/7078 at the Central Biomedical Services of Imperial College,London, UK. C57BL/6J wild-type mice were purchased from B&KUniversal Limited (Hull, UK). Rab27a-deficient mice, ashen(Rab27aash/ash) mice were described previously (31) and the genera-tion of Rab27b KO mice and double Rab27KO was described else-where (27). All strains were maintained on a C57/Bl/6 background.
Immunoblotting. Immunoblotting was performed as described pre-viously (12). Perfused lung lysates were prepared using a Polytronhomogenizer and an appropriate volume (10� vol/wt) of lysis buffer(150 mM NaCl, 20 mM Tris·HCl, pH 7.5, 1 mM DTT, 1% CHAPS,and 1� PI cocktail) followed by incubation on ice for 15 min. Nucleiand debris were then harvested by centrifugation (3,000 g at 4°C for10 min), and the protein content of the PNS was quantified using BCAprotein assay kit (Pierce, UK).
Real-time PCR analysis. Total RNA from resected human lungtissue obtained from 19 transplant donors was reverse transcribedusing a reaction mix of 1� RT buffer (500 �M each dNTP, 3 mMMgCl2, 75 mM KCl, 50 mM Tris·HCl, pH 8.3), 20 units of RNasinRnase inhibitor (Promega, Madison, WI), 10 mM dichloro-diphenyl-trichloroethane (DDT), 100 units of Superscript II RNase H-reversetranscriptase (Invitrogen, Uppsala, Sweden), and 250 ng of randomhexamers (Promega). First-strand cDNA synthesis was carried out ina final volume of 20 �l, incubating at 20°C for 10 min and 42°C for30 min, and inactivating reverse transcriptase by heating at 99°C for5 min and cooling at 5°C for 5 min.
Real-time PCR were performed using the 7000 Abi Prism (AppliedBiosystems, Foster City, CA) with optimized PCR conditions. Thereaction was carried out in a 96-well plate adding 3 �l of dilutedtemplate cDNA to a final reaction volume of 25 �l. The PCR mastermix was assembled with TaqMan Universal Master Mix Reagents(Applied Biosystems) and each Taqman Gene Expression Assay,Hs00608302 (Applied Biosystems) for human Rab27a, Hs01072206(Applied Biosystems) for Rab27b. Each target assay was replicatedthree times and performed in multiplex reaction with the 18S rRNAendogenous control gene (4310893E, Applied Biosystems). The ther-mal cycling conditions comprised an initial denaturation step at 95°Cfor 10 min, and 50 cycles at 95°C for 15 s and 65°C for 1 min.Real-time quantitative values were obtained from the Ct number atwhich the increase in signal associated with exponential growth ofPCR products starts to be detected. Results, expressed as amount intarget genes (Rab27a, Rab27b) expression relative to the referencegene (18s rRNA), were calculated with the �Ct method. Briefly, the�Ct value of the samples was determined by subtracting the averageCt value of the target gene from the average Ct value of the 18s rRNAgene.
Lung histology and morphometry. Male mice were terminallyanesthetized by intraperitoneal injection of ketamine-xylazine (100
and 12 mg/kg, respectively) and heparin (300 U/ml). Animals wereperfused with PBS through the right ventricle of the heart until thelungs were visually free of blood. The trachea was then exposed, anda Luer cannula (BD Insyte; 20 gauge 1.1 � 30 mm) was inserted andsecured with surgical thread. The lungs and heart were then removedand fixed by careful inflation with 10% formalin neutral buffersolution via the trachea at a constant hydrostatic pressure of 30cmH2O at the height of the carina in the upright position for 15 min.The lungs were further incubated overnight in fixative, and the rightlung was embedded in paraffin. After deparaffinization and rehydra-tion, 4-�m sections were stained using hematoxylin and eosin.Stained sections were then observed using a Zeiss Axiovert 200inverted microscope and images captured using a Hamamatsu OrcaER CCD. For mean linear intercept analysis of the integrity ofalveolar epithelium, 10 randomly acquired low-magnification (20�)images for each of 3 age-matched animals were overlaid with a gridcomprising 10 equally spaced horizontal lines, and the number oftimes that the epithelium intersected the grid was recorded for eachimage. The mean was determined and plotted as a percentage ofage-matched control (wild-type, WT) samples. For measurement ofbronchiolar thickness, bronchiolar epithelium areas were identifiedon the basis of intensity; the area was then calculated by subtrac-tion of the luminal-airway area from the total bronchiolar area. Thebronchiolar perimeter was then determined by taking the averageof the inner and outer perimeter measurements (determined witharea measurements). The area was then divided by the perimeter togive a thickness measurement (area/length of perimeter). Thismeasurement was performed on 10 randomly selected bronchioles
Fig. 1. Rab27a and Rab27b proteins are expressed in the lungs of mice andhumans. A: lung lysates were immunoblotted using Rab27a and Rab27bisoform-specific antibodies. Lysates from control animals (Rab27aash/ash andRab27BKO) lacking Rab27a and Rab27b, respectively, confirm the specificityof the antibody staining. Additionally, lysates were blotted with calnexin-specific antibodies as a control for equal loading. B: real-time PCR wasperformed to measure the expression of Rab27a- and Rab27b-specific tran-scripts in total cDNA prepared from samples of resected human lung. Signalwas normalized using 18s rRNA as control and expression level is displayedas �Ct value (inversely related to expression level).
C467RAB27 FUNCTION IN LUNG EPITHELIUM
AJP-Cell Physiol • VOL 300 • MARCH 2011 • www.ajpcell.org
on April 25, 2011
ajpcell.physiology.orgD
ownloaded from
from each of 3 age-matched animals. P values were derived usingthe Student’s t-test.
Immunohistochemistry. Immunofluorescence was performed on ei-ther deparaffinized, formalin-fixed paraffin sections or frozen sec-tions. Sections were deparaffinized using Histoclear and then takenthrough reducing alcohols to water, and rinsed in 1� PBS. For antigenretrieval, samples were boiled for 9 min in 10 mM sodium citratebuffer (pH 6.0) and cooled for 5–10 min in water. Slides were washedin 1� PBS and incubated with blocking buffer (1� PBS, 0.025%Triton X-100, 1% BSA, 10% donkey serum) for 1 h and thenincubated overnight at 4°C with primary antibody diluted in blockingbuffer. The following day slides were washed three times withwashing buffer (1� PBS, 0.1% Triton X-100), incubated with Al-exa488- or Alexa568-conjugated secondary antibodies, and finallywashed as before and mounted using Immunofluor mountant (MPBiomedicals, Cambridge, UK). Nuclei were visualized using 4,6-diamidino-2-phenylindole (DAPI) stain. For immunofluorescence offrozen sections, perfused lungs were manually inflated via the tracheausing a 1:1 mixture of OCT (Cellpath, Powys, UK) and PBS, andlungs were then embedded in OCT, frozen using liquid N2, and storedat �80°C. Semithin sections (5 �m) were cut at �20°C using acryostat (Bright Instruments, Huntingdon, UK) and fixed with 4%paraformaldehyde (PFA) (wt/vol), 1� PBS for 20 min at ambienttemperature and then processed for blocking and immunolabeling asdescribed for the deparaffinized sections. Images of fluorescent anti-body-stained sections were obtained using either wide-field ZeissAxiovert 200 controlled by Simple PCI acquisition software withimages recorded using a Hamamatsu EM-CCD or a Zeiss LSM-510inverted confocal microscope (Zeiss, Welwyn Garden City, UK).Images were edited using Adobe Photoshop CS software. For AEIIcell counting 10 randomly collected images of alveolar areas of the
lungs of 3 age-matched, sex-matched mice stained with pro-SPCantibody were imported into Volocity 5.4 image analysis software,and AEII cells were identified on the basis of staining intensity abovebackground as well as area measurements. P values were derivedusing the Student’s t-test.
Conventional electron microscopy. Lungs of C57BL/6 mice wereinfused and fixed with a mixture of 2% (wt/vol) PFA, 2% (wt/vol)glutaraldehyde (TAAB) in 0.1 M sodium cacodylate buffer (Agar),pH 7.4, postfixed with 1% (wt/vol) OsO4 supplemented with 1.5%(wt/vol) potassium ferrocyanide, dehydrated in ethanol, and infiltratedwith propylene oxide (Agar)/Epon (Agar) (1:1) followed by Eponembedding. Ultrathin sections were cut with an Ultracut S microtome(Leica), counter-stained with lead citrate, and observed with a trans-mission electron microscope (TEM) Jeol 1010. Images were obtainedusing a Gatan ORIUS CCD camera. For measurement of lamellarbody area, electron microscope images were imported into Image Jsoftware, the perimeter of each organelle was defined manually, andthe area was measured using the measure tool within the software.
Ultracryotomy and immunogold labeling. Lungs of C57BL/6 micewere infused and fixed with 2% (wt/vol) PFA and 0.1% (wt/vol)glutaraldehyde in 0.1 M sodium phosphate buffer (pH 7.1). Sampleswere cut in 0.5-mm3 blocks, embedded in 12% gelatin, and infused in2.3 M sucrose (26). Mounted gelatin blocks were frozen in N2, andultrathin (50 nm) cryosections were cut at �120°C with an Ultracryo-microtome (Leica). Sections were retrieved in 1.15 M (wt/vol) su-crose-2% (vol/vol) methylcellulose solution and processed for immu-nolabeling. After the sections were blocked with 0.5% (wt/vol) BSA,single immunolabeling was performed on the sections in a humidchamber with primary antibodies, and protein A was coupled to10-nm diameter gold particles (PAG-10 nm).
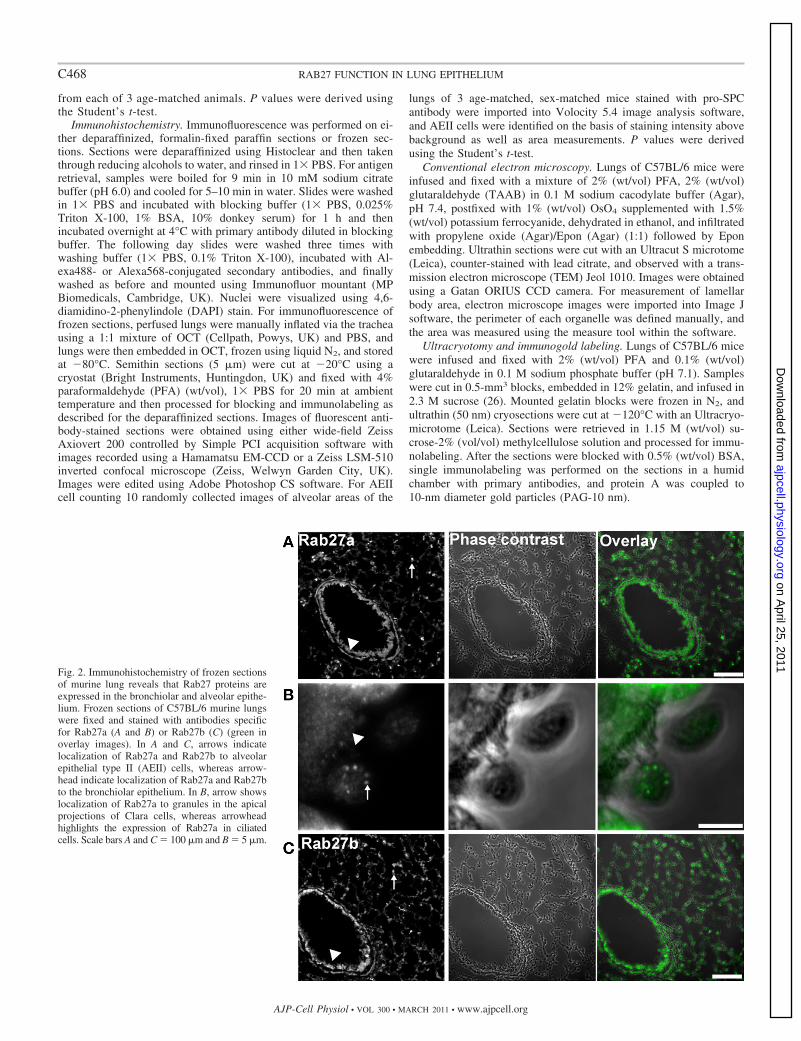
Fig. 2. Immunohistochemistry of frozen sectionsof murine lung reveals that Rab27 proteins areexpressed in the bronchiolar and alveolar epithe-lium. Frozen sections of C57BL/6 murine lungswere fixed and stained with antibodies specificfor Rab27a (A and B) or Rab27b (C) (green inoverlay images). In A and C, arrows indicatelocalization of Rab27a and Rab27b to alveolarepithelial type II (AEII) cells, whereas arrow-head indicate localization of Rab27a and Rab27bto the bronchiolar epithelium. In B, arrow showslocalization of Rab27a to granules in the apicalprojections of Clara cells, whereas arrowheadhighlights the expression of Rab27a in ciliatedcells. Scale bars A and C � 100 �m and B � 5 �m.
C468 RAB27 FUNCTION IN LUNG EPITHELIUM
AJP-Cell Physiol • VOL 300 • MARCH 2011 • www.ajpcell.org
on April 25, 2011
ajpcell.physiology.orgD
ownloaded from

RESULTS
Rab27A and Rab27B are expressed in the murine lung. As afirst step to investigate the role of Rab27 proteins in thefunction of the pulmonary epithelium, we used isoform-spe-cific Rab27 antibodies to test the expression of Rab27a andRab27b proteins in murine lung lysates. In control (WT) lunglysate we observed that both isoforms are expressed (Fig. 1A).The specificity of Rab27a and Rab27b signal was confirmed byblotting of lysate derived from mutant mice lacking individu-ally Rab27a (Rab27aash/ash) or Rab27b (Rab27bKO), respec-tively. Moreover, quantitative real-time PCR analysis ofRab27a and Rab27b mRNA levels in resected human lungtissue samples showed that both genes are significantly ex-pressed in the lung (Fig. 1B).
Rab27 proteins are expressed in Clara cells and AEII cells.We then used immunofluorescence microscopy to reveal thedistribution of Rab27 isoforms in frozen sections of C57Bl/6mouse lung. This showed that Rab27a is highly expressed inthe bronchiolar epithelium (Fig. 2A, arrowhead) where it is
present in both ciliated and Clara cells (Fig. 2B, arrowhead andarrow, respectively). In high-magnification images of bron-chiolar epithelium, Rab27a was found to be particularly appar-ent in apical vesicles in Clara cells that may represent secretoryvesicles (Fig. 2B, arrow). Within the alveolar epithelium,Rab27a was found to be highly expressed within a subset ofcells (Fig. 2A, arrow). Double immunolabeling of surfactantprotein C (SPC), which is specifically expressed in AEII cells,revealed that Rab27a-positive cells are AEII cells (Fig. 3A).High-magnification images of AEII cells showed that whileSPC is present in punctate structures (presumably LBs),Rab27a is present on small vesicles throughout the cytoplasmthat do not significantly colocalize with LBs (Fig. 3B). Similaranalysis using specific antibodies indicated that Rab27b is alsoexpressed in ciliated and Clara cells of the bronchiolar epithe-lium (Fig. 2C, arrowhead), as well as a subset of cells in thealveolar epithelium (Fig. 2C, arrow). Double immunolabelingof frozen sections with SPC-specific antibodies confirmed thata subset of these Rab27b-positive cells are AEII cells (Fig. 3C,
Fig. 3. Rab27 proteins are localized to cytoplasmic vesicles in AEII cells. Frozen sections of C57BL/6 murine lungs were fixed and stained with antibodiesspecific for Rab27a (A, B) or Rab27b (C, D) (green in overlay images), surfactant protein C (SPC, red in overlay images), and DAPI (blue in overlay images)and analyzed using confocal immunofluorescence microscopy. Low-magnification images show Rab27a (A) and Rab27b (C) expression in AEII cells (arrows).High-magnification images show the intracellular distribution of Rab27a (B) and Rab27b (D) in AEII. Scale bars A and C � 25 �m, B and D � 5 �m.
C469RAB27 FUNCTION IN LUNG EPITHELIUM
AJP-Cell Physiol • VOL 300 • MARCH 2011 • www.ajpcell.org
on April 25, 2011
ajpcell.physiology.orgD
ownloaded from
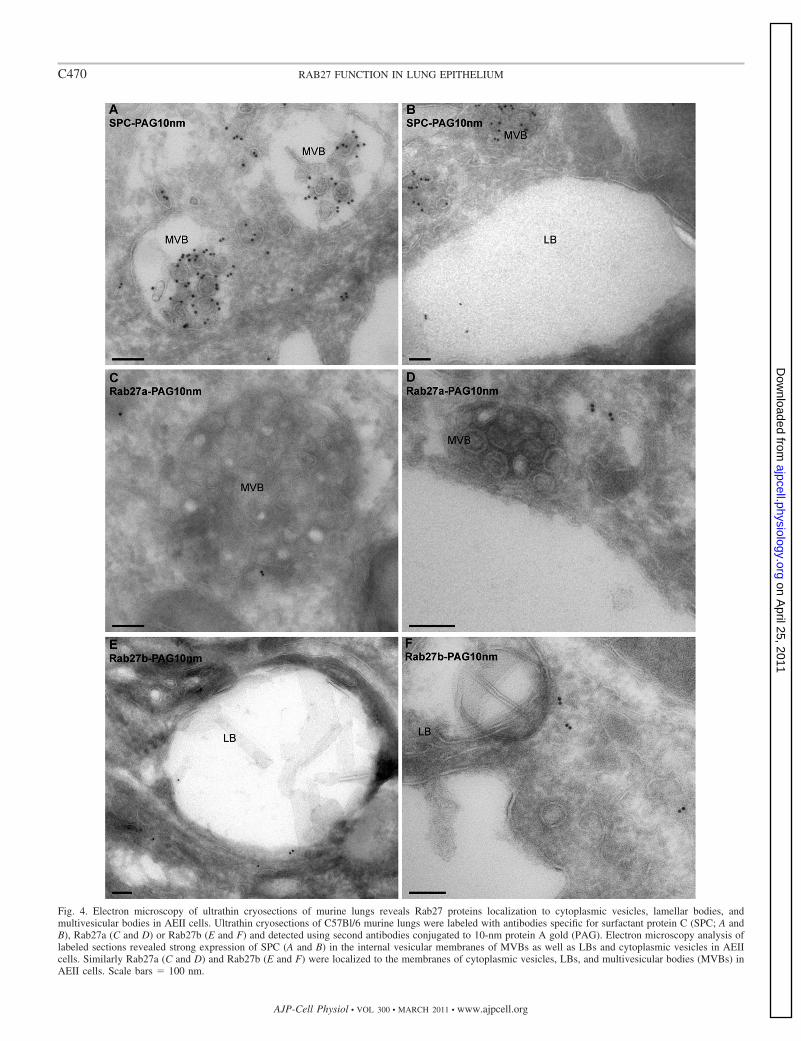
Fig. 4. Electron microscopy of ultrathin cryosections of murine lungs reveals Rab27 proteins localization to cytoplasmic vesicles, lamellar bodies, andmultivesicular bodies in AEII cells. Ultrathin cryosections of C57Bl/6 murine lungs were labeled with antibodies specific for surfactant protein C (SPC; A andB), Rab27a (C and D) or Rab27b (E and F) and detected using second antibodies conjugated to 10-nm protein A gold (PAG). Electron microscopy analysis oflabeled sections revealed strong expression of SPC (A and B) in the internal vesicular membranes of MVBs as well as LBs and cytoplasmic vesicles in AEIIcells. Similarly Rab27a (C and D) and Rab27b (E and F) were localized to the membranes of cytoplasmic vesicles, LBs, and multivesicular bodies (MVBs) inAEII cells. Scale bars � 100 nm.
C470 RAB27 FUNCTION IN LUNG EPITHELIUM
AJP-Cell Physiol • VOL 300 • MARCH 2011 • www.ajpcell.org
on April 25, 2011
ajpcell.physiology.orgD
ownloaded from

arrows). High-magnification images of double-labeled sectionsreveal little colocalization of Rab27b with SPC-positive LBs(Fig. 3D). This analysis also showed that Rab27b is stronglyexpressed on cytoplasmic granules in an additional cell typepresent throughout the alveolar epithelium (Fig. 3C, arrowheadand see online supplementary Fig. S1A at AJP-Cell Physiolwebsite). The specificity of antibody staining was confirmed bystaining frozen sections of lung from double knockout (DKO)mice (see online supplementary Fig. S2).
Rab27 proteins associate with cytoplasmic vesicles, LBs,and multivesicular bodies in AETII cells and apical secretorygranules in Clara cells. We then used cryoimmunoelectronmicroscopy to investigate further the identity of Rab27-posi-tive structures within AEII cells in ultrathin cryosections of thelungs of C57BL/6 mice. Staining with SPC-specific antibodies(Fig. 4, A and B) showed that this protein is present in theinternal membranes of multivesicular bodies (MVBs), LBprecursor vesicles, and mature LBs. Similar analysis showedthat Rab27a is associated with MVBs and vesicles presentthroughout the cytoplasm (Fig. 4, C and D). Furthermore,Rab27a did not show strong colocalization with LBs. Mean-while, Rab27b was localized to cytoplasmic vesicles and insome cases to the limiting membrane of LBs (Fig. 4, E and F).Consistent with immunofluorescence, immunoelectron micros-copy analysis confirmed the existence of a second class ofRab27b-expressing cell type within the capilliary network ofthe alveolar wall. High-magnification images revealed thatthese cells are eosinophils, based on the presence of cytoplas-mic crystalloid granules that were labeled with anti-Rab27bantibodies (see online supplementary Fig. S1, B–E). Mean-while, in the bronchiolar epithelium, examination of immuno-gold-labeled cryosections revealed that Rab27a is present on thelimiting membrane of secretory CCSP granules in Clara cells (Fig.5, A and B). We also observed gold particles in Clara cells incryosections labeled with Rab27b-specific antibodies (data notshown).
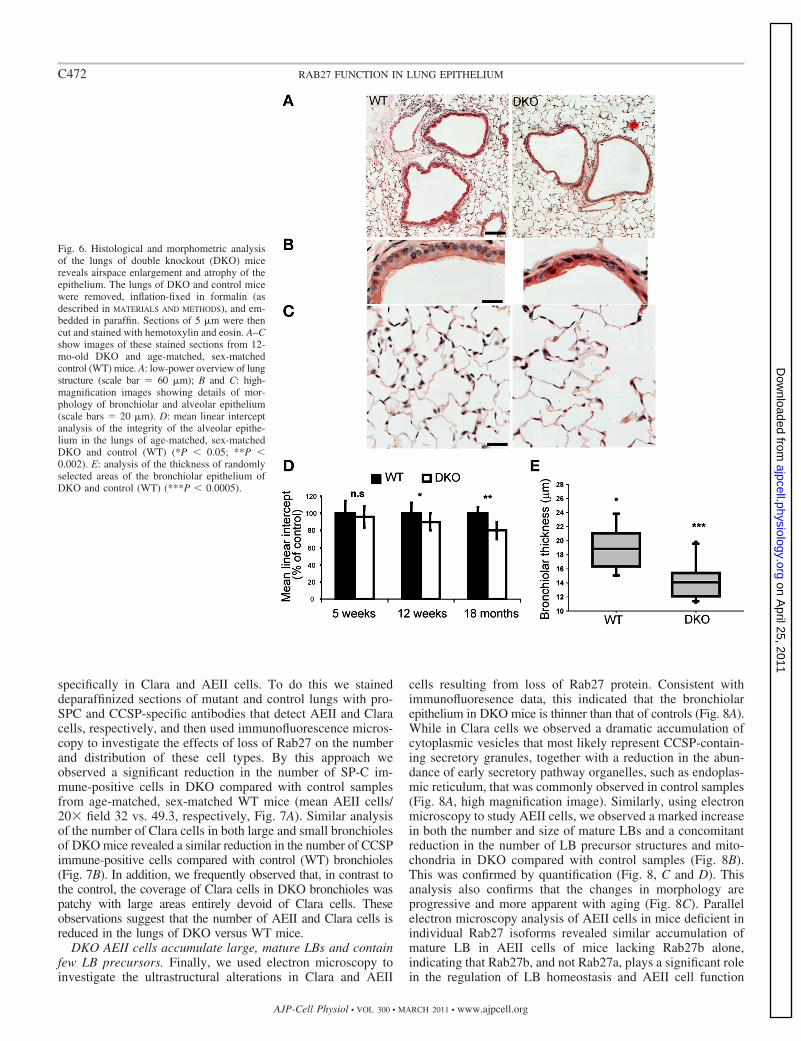
Loss of Rab27 protein results in atrophy of the pulmonaryepithelium. Based on the previous observation that Rab27proteins are expressed in the murine lung, we next aimed toinvestigate the function of Rab27 in this context. As a first stepwe examined the morphology of lungs from mice lacking bothRab27 isoforms (DKO). As shown in Fig. 6A, we observedthinning of the bronchiolar epithelium as well as markedenlargement of the alveolar air spaces in lungs from the DKOcompared with control (WT) samples. These morphologicalalterations were clearly observed in high-magnification imagesof bronchioles (Fig. 6B), which show disorganization andshortening of bronchiolar cilia, while alveolar areas clearlyshow enlargement of alveolar spaces and thinning of thealveolar epithelium (Fig. 6C). Quantification using mean linearintercept measurement (Fig. 6D) indicated that airspace en-largement in the DKO reaches significance in the lungs of 12wk- and 18-mo-old mice. This also indicated that airspaceenlargement becomes more evident with increasing age. Mean-while, measurement of the thickness of the epithelium inmultiple bronchioles confirmed that there is significant thin-ning of the epithelium in DKO compared with age-matched,sex-matched control (WT) samples (Fig. 6E). In addition tothese changes, we also observed increased incidence of acti-vated foamy macrophages (see online supplementary Fig. S3A,arrows) and cellular infiltrates containing granulocytes (sup-plementary Fig. S3, B and C, arrows) in 12-mo-old DKO micecompared with those of age-matched, sex-matched controls(WT) (4/5 DKO vs. 2/9 control samples). Morphological anal-yses of the lungs of mice lacking individual Rab27 isoformsindicate that Rab27b plays a more significant role in mainte-nance of the integrity of the pulmonary epithelium than doesRab27a (data not shown).
The numbers of cells expressing AEII and Clara markerproteins are reduced in the lungs of DKO mice. Given that lossof Rab27 proteins resulted in generalized atrophy of the pul-monary epithelium, we investigated the effect of loss of Rab27
Fig. 5. Cryoimmunoelectron microscopy of murine lungs reveals Rab27a localization to secretory/CCSP granules within Clara cells. Ultrathin cryosections ofC57Bl/6 murine lungs were labeled with antibody specific for Rab27a (A and B) and detected using protein A conjugated to 10-nm gold particles (PAG). Electronmicroscopy analysis at higher magnification (B) of labeled sections revealed expression of Rab27a (A and B) in membrane of Clara cell granules. Mito,mitochondria; CCSP, Clara cell secreted protein granule. Scale bars � 500 nm.
C471RAB27 FUNCTION IN LUNG EPITHELIUM
AJP-Cell Physiol • VOL 300 • MARCH 2011 • www.ajpcell.org
on April 25, 2011
ajpcell.physiology.orgD
ownloaded from
specifically in Clara and AEII cells. To do this we staineddeparaffinized sections of mutant and control lungs with pro-SPC and CCSP-specific antibodies that detect AEII and Claracells, respectively, and then used immunofluorescence micros-copy to investigate the effects of loss of Rab27 on the numberand distribution of these cell types. By this approach weobserved a significant reduction in the number of SP-C im-mune-positive cells in DKO compared with control samplesfrom age-matched, sex-matched WT mice (mean AEII cells/20� field 32 vs. 49.3, respectively, Fig. 7A). Similar analysisof the number of Clara cells in both large and small bronchiolesof DKO mice revealed a similar reduction in the number of CCSPimmune-positive cells compared with control (WT) bronchioles(Fig. 7B). In addition, we frequently observed that, in contrast tothe control, the coverage of Clara cells in DKO bronchioles waspatchy with large areas entirely devoid of Clara cells. Theseobservations suggest that the number of AEII and Clara cells isreduced in the lungs of DKO versus WT mice.
DKO AEII cells accumulate large, mature LBs and containfew LB precursors. Finally, we used electron microscopy toinvestigate the ultrastructural alterations in Clara and AEII
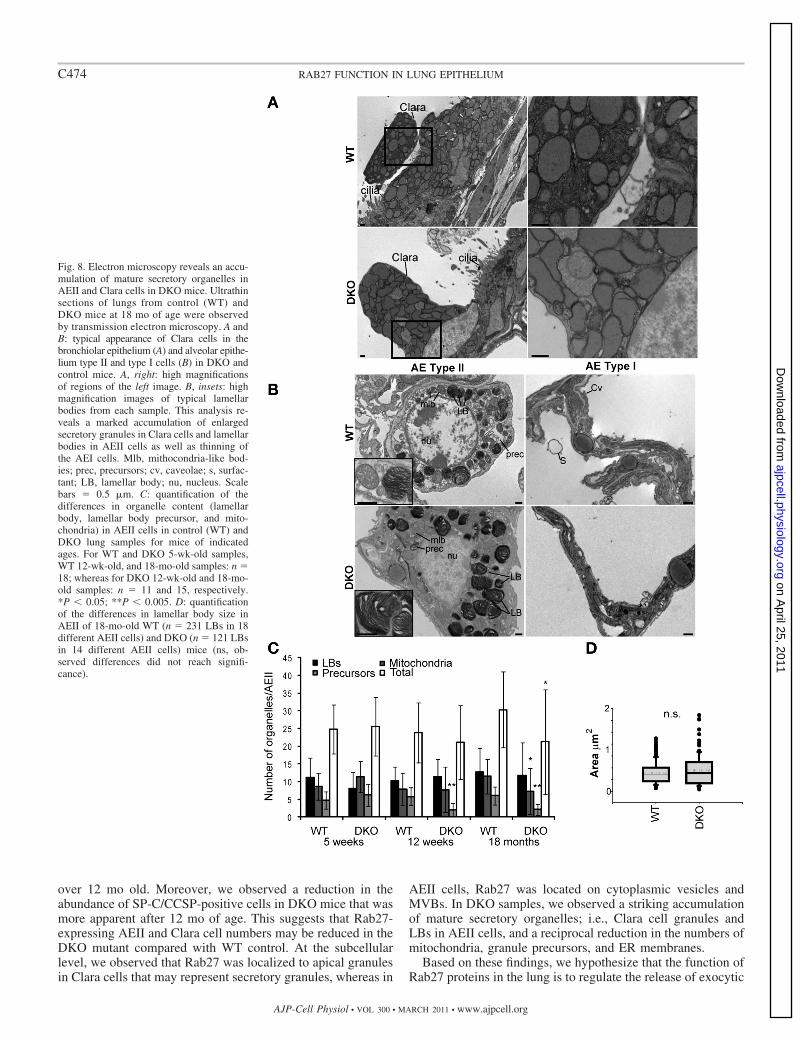
cells resulting from loss of Rab27 protein. Consistent withimmunofluoresence data, this indicated that the bronchiolarepithelium in DKO mice is thinner than that of controls (Fig. 8A).While in Clara cells we observed a dramatic accumulation ofcytoplasmic vesicles that most likely represent CCSP-contain-ing secretory granules, together with a reduction in the abun-dance of early secretory pathway organelles, such as endoplas-mic reticulum, that was commonly observed in control samples(Fig. 8A, high magnification image). Similarly, using electronmicroscopy to study AEII cells, we observed a marked increasein both the number and size of mature LBs and a concomitantreduction in the number of LB precursor structures and mito-chondria in DKO compared with control samples (Fig. 8B).This was confirmed by quantification (Fig. 8, C and D). Thisanalysis also confirms that the changes in morphology areprogressive and more apparent with aging (Fig. 8C). Parallelelectron microscopy analysis of AEII cells in mice deficient inindividual Rab27 isoforms revealed similar accumulation ofmature LB in AEII cells of mice lacking Rab27b alone,indicating that Rab27b, and not Rab27a, plays a significant rolein the regulation of LB homeostasis and AEII cell function
Fig. 6. Histological and morphometric analysisof the lungs of double knockout (DKO) micereveals airspace enlargement and atrophy of theepithelium. The lungs of DKO and control micewere removed, inflation-fixed in formalin (asdescribed in MATERIALS AND METHODS), and em-bedded in paraffin. Sections of 5 �m were thencut and stained with hemotoxylin and eosin. A–Cshow images of these stained sections from 12-mo-old DKO and age-matched, sex-matchedcontrol (WT) mice. A: low-power overview of lungstructure (scale bar � 60 �m); B and C: high-magnification images showing details of mor-phology of bronchiolar and alveolar epithelium(scale bars � 20 �m). D: mean linear interceptanalysis of the integrity of the alveolar epithe-lium in the lungs of age-matched, sex-matchedDKO and control (WT) (*P � 0.05; **P �0.002). E: analysis of the thickness of randomlyselected areas of the bronchiolar epithelium ofDKO and control (WT) (***P � 0.0005).
C472 RAB27 FUNCTION IN LUNG EPITHELIUM
AJP-Cell Physiol • VOL 300 • MARCH 2011 • www.ajpcell.org
on April 25, 2011
ajpcell.physiology.orgD
ownloaded from

(data not shown). Ultrastructural analysis of AEI cells con-firmed that there is marked thinning of the alveolar epitheliumin DKO compared with control lungs (Fig. 8B).
DISCUSSION
In this study we used light and electron microscopy toinvestigate the localization and function of Rab27 proteins inthe lung. Our main findings are that Rab27a and Rab27b areexpressed in Clara and ciliated cells in bronchioles as well asAEII cells in the alveolar region. Rab27b is additionallystrongly expressed in cells in the parenchyma that we havedefined as eosinophils based on our finding that they contain
eosinophil-specific crystalloid granules. In particular, we findthat Rab27b is associated with cytoplasmic face of thesegranules. These observations are largely consistent with previ-ous analysis of expression of GFP-Rab27a transgenic proteinand Rab27b-driven lacZ expression in the lungs of mice (6,29). However, our finding of high Rab27b expression ineosinophils is surprising given that previous studies indicatedthat Rab27a, not Rab27b, is expressed in circulating eosino-phils in humans (2). Meanwhile, our analysis of the structure ofthe lungs of DKO mice revealed a generalized atrophy of boththe alveolar and bronchiolar epithelium together with mildairspace enlargement that was particularly apparent in mice
Fig. 7. Immunohistochemistry reveals a reduction in the number of cells positive for AEII and Clara marker proteins in the lungs of DKO mice. Paraffin sectionsof lungs from 18-mo-old control (WT) and DKO mice were dewaxed and stained with ProSPC- (A) or CCSP (B)-specific antibodies to detect AEII and Claracells and DAPI to detect cell nuclei. A: left, fluorescence and phase contrast images of the number and distribution of Pro-SPC-positive AEII cells and themorphology of lung parenchyma in mutant and control samples. Right, quantification of the Pro-SPC data showing the number of AEII cells present in 10randomly selected low-magnification fields from mutant and control samples. *P value �0.0001. B: fluorescence micrographs show the typical distribution ofClara cells in the large and small bronchiolar airways in the lungs of 18-mo-old control (WT) and DKO mice. Scale bars � 20 �m.
C473RAB27 FUNCTION IN LUNG EPITHELIUM
AJP-Cell Physiol • VOL 300 • MARCH 2011 • www.ajpcell.org
on April 25, 2011
ajpcell.physiology.orgD
ownloaded from
over 12 mo old. Moreover, we observed a reduction in theabundance of SP-C/CCSP-positive cells in DKO mice that wasmore apparent after 12 mo of age. This suggests that Rab27-expressing AEII and Clara cell numbers may be reduced in theDKO mutant compared with WT control. At the subcellularlevel, we observed that Rab27 was localized to apical granulesin Clara cells that may represent secretory granules, whereas in
AEII cells, Rab27 was located on cytoplasmic vesicles andMVBs. In DKO samples, we observed a striking accumulationof mature secretory organelles; i.e., Clara cell granules andLBs in AEII cells, and a reciprocal reduction in the numbers ofmitochondria, granule precursors, and ER membranes.
Based on these findings, we hypothesize that the function ofRab27 proteins in the lung is to regulate the release of exocytic
Fig. 8. Electron microscopy reveals an accu-mulation of mature secretory organelles inAEII and Clara cells in DKO mice. Ultrathinsections of lungs from control (WT) andDKO mice at 18 mo of age were observedby transmission electron microscopy. A andB: typical appearance of Clara cells in thebronchiolar epithelium (A) and alveolar epithe-lium type II and type I cells (B) in DKO andcontrol mice. A, right: high magnificationsof regions of the left image. B, insets: highmagnification images of typical lamellarbodies from each sample. This analysis re-veals a marked accumulation of enlargedsecretory granules in Clara cells and lamellarbodies in AEII cells as well as thinning ofthe AEI cells. Mlb, mithocondria-like bod-ies; prec, precursors; cv, caveolae; s, surfac-tant; LB, lamellar body; nu, nucleus. Scalebars � 0.5 �m. C: quantification of thedifferences in organelle content (lamellarbody, lamellar body precursor, and mito-chondria) in AEII cells in control (WT) andDKO lung samples for mice of indicatedages. For WT and DKO 5-wk-old samples,WT 12-wk-old, and 18-mo-old samples: n �18; whereas for DKO 12-wk-old and 18-mo-old samples: n � 11 and 15, respectively.*P � 0.05; **P � 0.005. D: quantificationof the differences in lamellar body size inAEII of 18-mo-old WT (n � 231 LBs in 18different AEII cells) and DKO (n � 121 LBsin 14 different AEII cells) mice (ns, ob-served differences did not reach signifi-cance).
C474 RAB27 FUNCTION IN LUNG EPITHELIUM
AJP-Cell Physiol • VOL 300 • MARCH 2011 • www.ajpcell.org
on April 25, 2011
ajpcell.physiology.orgD
ownloaded from

granules in specialized secretory cell types such as Clara andAEII cells. This suggestion is consistent with the localizationdata, as well as the observed accumulation of end-stage secre-tory organelles in these cell types in mice lacking Rab27proteins. Further support for this idea comes from the largenumber of other studies that have linked Rab27 function toregulated exocytosis in a variety of cell types, including CTLs,platelets, and neutrophils (5, 13). Leading from this, we hy-pothesize that chronic disruption of exocytosis in Clara andAEII cells is the cause of pulmonary epithelium atrophy inDKO. Consistent with this, the reduction in the number ofgranule precursor structures, early secretory pathway organ-elles, and mitochondria point to a mechanism whereby accu-mulation of end-stage organelles feeds back into to a reductionin metabolic activity in these cells. Given that other cell types;e.g., bronchiolar ciliated cells and AEI cells, are thought to bederived from these Clara cells and AEII cells, reduced meta-bolic, and possibly proliferative, activity in these cells may gosome way to explaining the widespread atrophy of the lungtissue observed in the DKO mice (16). Interestingly, thechanges in lung architecture observed in DKO are similar tothose that occur during natural aging, suggesting that the DKOmay represent a useful model for the study of accelerated agingof the lung (23). The relatively mild phenotype observed in theDKO mutant lungs suggests that the function of Rab27 proteinsis partially redundant. In line with this idea it has been reportedthat the closely protein Rab3d, whose function is also linked toexocytosis, is expressed in the pulmonary epithelium (30).Also consistent with the relatively mild nature of the DKOmutant phenotype, we did not observe significant differences inthe levels of secreted SP-A, SP-D, SP-C, and CCSP in thebronchoalveolar lavage fluid of DKO and WT animals (datanot shown).
Future studies should address the functional consequences ofthese changes in lung structure in the DKO mice; they shouldalso identify the cargo whose exocytosis is regulated by Rab27and the molecular mechanisms by which this occurs.
ACKNOWLEDGMENTS
We thank Lorraine Lawrence for technical assistance in the preparation ofparaffin-embedded and frozen sections and Dr. Martin Spitaler for assistancewith microscopy.
The current address for A. N. Hume: School of Biomedical Sciences,Queen’s Medical Centre, Nottingham NG7 2UH, UK; and current address forG. Bolasco: EMBL Mouse Biology Unit, Campus A. Buzzati-Traverso, Via E.Ramarini, 32, 00015 Monterotondo (RM), Italy.
GRANTS
This work was supported by Wellcome Trust Programme Grant (Reference075498/Z/04/Z) to M. C. Seabra.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the author(s).
REFERENCES
1. Bahadoran P, Aberdam E, Mantoux F, Busca R, Bille K, Yalman N,de Saint-Basile G, Casaroli-Marano R, Ortonne JP, Ballotti R.Rab27a: A key to melanosome transport in human melanocytes. J CellBiol 152: 843–850, 2001.
2. Coughlin JJ, Odemuyiwa SO, Davidson CE, Moqbel R. Differentialexpression and activation of Rab27A in human eosinophils: relationship toblood eosinophilia. Biochem Biophys Res Commun 373: 382–386, 2008.
3. Davis CW, Dickey BF. Regulated airway goblet cell mucin secretion.Annu Rev Physiol 70: 487–512, 2008.
4. Dietl P, Haller T. Exocytosis of lung surfactant: from the secretoryvesicle to the air-liquid interface. Annu Rev Physiol 67: 595–621, 2005.
5. Fukuda M. Regulation of secretory vesicle traffic by Rab small GTPases.Cell Mol Life Sci 65: 2801–2813, 2008.
6. Gomi H, Mori K, Itohara S, Izumi T. Rab27b is expressed in a widerange of exocytic cells and involved in the delivery of secretory granulesnear the plasma membrane. Mol Biol Cell 18: 4377–4386, 2007.
7. Gou D, Mishra A, Weng T, Su L, Chintagari NR, Wang Z, Zhang H,Gao L, Wang P, Stricker HM, Liu L. Annexin A2 interactions withRab14 in alveolar type II cells. J Biol Chem 283: 13156–13164, 2008.
8. Haddad EK, Wu X, Hammer JA, 3rd, Henkart PA. Defective granuleexocytosis in Rab27a-deficient lymphocytes from Ashen mice. J Cell Biol152: 835–842, 2001.
9. Hammer JA, 3rd, Wu XS. Rabs grab motors: defining the connectionsbetween Rab GTPases and motor proteins. Curr Opin Cell Biol 14: 69–75,2002.
10. Hume AN, Collinson LM, Hopkins CR, Strom M, Barral DC, Bossi G,Griffiths GM, Seabra MC. The leaden gene product is required withRab27a to recruit myosin Va to melanosomes in melanocytes. Traffic 3:193–202, 2002.
11. Hume AN, Collinson LM, Rapak A, Gomes AQ, Hopkins CR, andSeabra MC. Rab27a regulates the peripheral distribution of melanosomesin melanocytes. J Cell Biol 152: 795–808, 2001.
12. Hume AN, Tarafder AK, Ramalho JS, Sviderskaya EV, and SeabraMC. A coiled-coil domain of melanophilin is essential for Myosin Varecruitment and melanosome transport in melanocytes. Mol Biol Cell 17:4720–4735, 2006.
13. Izumi T. Physiological roles of Rab27 effectors in regulated exocytosis.Endocr J 54: 649–657, 2007.
14. Johnson JL, Brzezinska AA, Tolmachova T, Munafo DB, Ellis BA,Seabra MC, Hong H, Catz SD. Rab27a and Rab27b regulate neutrophilazurophilic granule exocytosis and NADPH oxidase activity by indepen-dent mechanisms. Traffic 11: 533–547, 2010.
15. Mason RJ. Biology of alveolar type II cells. Respirology 11, Suppl:S12–S15, 2006.
16. Morrisey EE, Hogan BL. Preparing for the first breath: genetic andcellular mechanisms in lung development. Dev Cell 18: 8–23, 2010.
17. Neeft M, Wieffer M, de Jong AS, Negroiu G, Metz CH, van Loon A,Griffith J, Krijgsveld J, Wulffraat N, Koch H, Heck AJ, Brose N,Kleijmeer M, van der Sluijs P. Munc13–4 is an effector of rab27a andcontrols secretion of lysosomes in hematopoietic cells. Mol Biol Cell 16:731–741, 2005.
18. Osanai K, Higuchi J, Oikawa R, Kobayashi M, Tsuchihara K, IguchiM, Huang J, Voelker DR, Toga H. Altered lung surfactant system in aRab38-deficient rat model of Hermansky-Pudlak syndrome. Am J PhysiolLung Cell Mol Physiol 298: L243–L251, 2010.
19. Osanai K, Oikawa R, Higuchi J, Kobayashi M, Tsuchihara K, IguchiM, Jongsu H, Toga H, Voelker DR. A mutation in Rab38 small GTPasecauses abnormal lung surfactant homeostasis and aberrant alveolar struc-ture in mice. Am J Pathol 173: 1265–1274, 2008.
20. Ostrowski M, Carmo NB, Krumeich S, Fanget I, Raposo G, Savina A,Moita CF, Schauer K, Hume AN, Freitas RP, Goud B, Benaroch P,Hacohen N, Fukuda M, Desnos C, Seabra MC, Darchen F, AmigorenaS, Moita LF, Thery C. Rab27a and Rab27b control different steps of theexosome secretion pathway. Nat Cell Biol 12: 19–30, 2010.
21. Seabra MC, Coudrier E. Rab GTPases and myosin motors in organellemotility. Traffic 5: 393–399, 2004.
22. Seabra MC, Mules EH, Hume AN. Rab GTPases, intracellular trafficand disease. Trends Mol Med 8: 23–30, 2002.
23. Sharma G, Goodwin J. Effect of aging on respiratory system physiologyand immunology. Clin Interv Aging 1: 253–260, 2006.
24. Stenmark H. Rab GTPases as coordinators of vesicle traffic. Nat Rev MolCell Biol 10: 513–525, 2009.
25. Stinchcombe JC, Barral DC, Mules EH, Booth S, Hume AN, Ma-chesky LM, Seabra MC, Griffiths GM. Rab27a is required for regulatedsecretion in cytotoxic T lymphocytes. J Cell Biol 152: 825–834, 2001.
26. Tokuyasu KT. A technique for ultracryotomy of cell suspensions andtissues. J Cell Biol 57: 551–565, 1973.
27. Tolmachova T, Abrink M, Futter CE, Authi KS, and Seabra MC.Rab27b regulates number and secretion of platelet dense granules. ProcNatl Acad Sci USA 104: 5872–5877, 2007.
28. Tolmachova T, Anders R, Abrink M, Bugeon L, Dallman MJ, FutterCE, Ramalho JS, Tonagel F, Tanimoto N, Seeliger MW, Huxley C,Seabra MC. Independent degeneration of photoreceptors and retinal
C475RAB27 FUNCTION IN LUNG EPITHELIUM
AJP-Cell Physiol • VOL 300 • MARCH 2011 • www.ajpcell.org
on April 25, 2011
ajpcell.physiology.orgD
ownloaded from

pigment epithelium in conditional knockout mouse models of choroider-emia. J Clin Invest 116: 386–394, 2006.
29. Tolmachova T, Anders R, Stinchcombe J, Bossi G, Griffiths GM,Huxley C, Seabra MC. A general role for Rab27a in secretory cells. MolBiol Cell 15: 332–344, 2004.
30. van Weeren L, de Graaff AM, Jamieson JD, Batenburg JJ, ValentijnJA. Rab3D and actin reveal distinct lamellar body subpopulations in alveolarepithelial type II cells. Am J Respir Cell Mol Biol 30: 288–295, 2004.
31. Wilson SM, Yip R, Swing DA, O’Sullivan TN, Zhang Y, Novak EK,Swank RT, Russell LB, Copeland NG, Jenkins NA. A mutation inRab27a causes the vesicle transport defects observed in ashen mice. ProcNatl Acad Sci USA 97: 7933–7938, 2000.
32. Wu X, Rao K, Bowers MB, Copeland NG, Jenkins NA, HammerJA, 3rd. Rab27a enables myosin Va-dependent melanosome captureby recruiting the myosin to the organelle. J Cell Sci 114: 1091–1100,2001.
C476 RAB27 FUNCTION IN LUNG EPITHELIUM
AJP-Cell Physiol • VOL 300 • MARCH 2011 • www.ajpcell.org
on April 25, 2011
ajpcell.physiology.orgD
ownloaded from





























