Embed Size (px)
Citation preview

long-range allosteric effect in gene transcriptional regulation
Ming LiGraduate School, CAS
Zhong-can Ou-yangInstitute of Theoretical Physics, CAS
International Symposium on the Recent Progress in Quantitative and Systems Biology , Dec 9-11, The Chinese University of Hong Kong

Outline
Stressed state of DNA in vivo
Model of topologically constrained DNA and duplex separation
Long-range Allosteric Effect and Database investigation: a case study

Part I
Stressed state of DNA in vivo

4 types of nucleotides: Adenine, ThymineGuanine, Cytosine
Waston-Crick base pair:A-T, G-C
Intrinsic right-handed helix, stiff polymer
B-DNA: uniform, sequence-independent
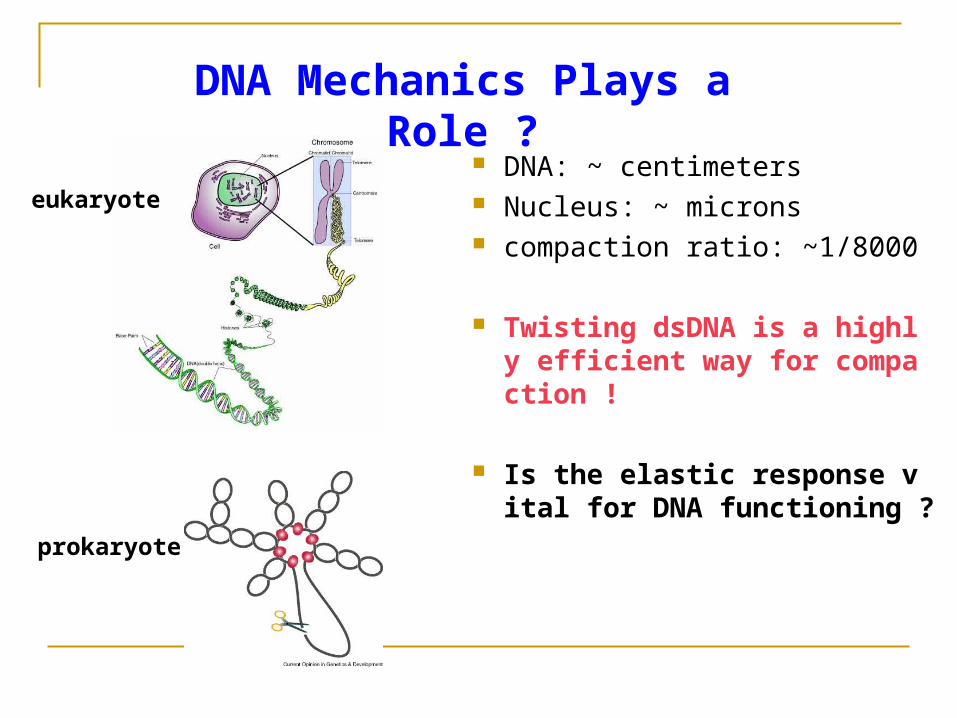
DNA: ~ centimeters Nucleus: ~ microns compaction ratio: ~1/8000
Twisting dsDNA is a highly efficient way for compaction !
Is the elastic response vital for DNA functioning ?
DNA Mechanics Plays a Role ?
eukaryote
prokaryote

Hierarchical architecture
DNA Stressed
S/MARs:Boundaries of topologically-independe
nt domains ?
bases of chromatin loops S/MAR
(Scaffold/Matrix Attachment Region)
Chromosome AssemblyChromatin Loop Model

DNA segment per nucleosome: ~167 bp
The segment is actually undertwisted :
one helical turn unwound per nucleosome.
torsional stress generated
Genomic DNA is potentially under unwinding stress

Sequence Heterogeneity Sequence Heterogeneity ? ? Structure Heterogeneity Structure Heterogeneity
bubble
cruciform
Chirality Variable Unstacking

Unwinding can promote local strand-separation (bubble) as well as global axis-supercoil
ing

Standard B-DNA
Local bubble
Unwinding stress in vivo Plays the Key Rolein Bubble formation !
In summary:

Part II
Benham Model of topologically constrained DNA &&sequence-dependent duplex separation

Lk : linking number, number of helical turns Lk0 : ‘linking number’ of relaxed DNA (uniform B-DNA) Lk0= N/10.5
σ : superhelical density. (Lk – Lk0)/ Lk0 σ< 0, negative supercoiling σ> 0, positive supercoiling
For eukaryotes, DNA is always unwound to a degree σ~ - 0.06 (1/167)
Characterizing the degree of unwinding …

Supercoiling free energy ( when σ << 1 )
2
22
1
qRTNG
21( )
2G K Lk
-0.06
10.5
92002200 q
Circular DNA Fully-stretched linearDNA with fixed ends
: bp number per helical turn of B-DNA
q is determined by the bending and twisting stiffness of dsDNA, as well as the topological constraints imposed on dsDNA
For more details on related DNA mechanics, see Ming Li, AAPPS Bulletin, Vol.16, No.3, 34-39

Bubble Formation is Sequence Dependent !
2N configurations{…10111111100…}
local bubble
a : initiation energy of bubble formation
jn = 0 … base paried
jn = 1 … base unparied
j : rewinding angle of the denatured region
ATb GCb : base unparing energy

When there are small fraction of unwound region, the supercoiling energy form slightly changes as:
N
jjnn
1
A
nTw total change in twisting turns
upon bubble formation
N
j
jjn
1 2
21 2
1( )
2
qRT nH
A
Bauer WR, Benham CJ., J Mol Biol. 1993, 234(4):1184-96.

Benham Model
total energy
Topological constraint long-range coupling between any two sites ! twisting energy of the two strands in bubble regions
unpairing energy in bubble (sequence dependent ) nucleation (initiation) energy for bubble formation (there can be multiple bubbles on a single DNA)
4321 HHHHH
21 2
1( )
2
qRT nH
A
j
jjnc
H 22 2
j
jjbnH 3
j
jj nnaarH )1( 14

8.10a 1molkcal
58.3c 21 radmolkcal
255.0ATb 1molkcal
301.1GCb 1molkcal
Parameter (illustrative) values under physiological condition
(the qualitative results shown later are actually quite insensitive to those values )

Unwounding Probability Profile
{ }
{ }
( 1)Hj
sj H
s
e n
pe
for any DNA sequence
Pi and Pj are tightly correlated due to the global topological constraints ( H1 !) imposed on the dsDNA, i.e., bubbles can be competitive in releasing the imposed twisting stress.

Can there be anything new (besides the individual denaturing events) introduced by topological constraints ? Long-range Allosteric Effect : the tele-communication between site A and B(protein binding onto the denatured site A may re-close the bubble and induce a new bubble at site B)
A
B
(DNA)
(protein)
B
A

It’s difficult to detect such a phenomenon in vivo by experiments, and it’s also almost impossible to directly ‘calculate’ such an effect quantitatively for real cases by taking account of every molecular detail (one can be drown in the details).
Beginning with everything , ending with nothing !
Anyway, How can one do anything meaningful ? --- Bioinformatics to rescue !
Bioinformatics offers an alternative approach:exploring the biological data to find the statistically significant patterns which may cast some light on the understanding of the underlying molecular mechanism

Part III
Long-range Allosteric Effect and Database investigation: a (bioinformatic) case study on SMAR function

D. menalongaster Histone gene cluster
5- —H3—H4—H2A—H2B—H1— -3MAR MAR
H3
H4
H2A
H2B
H1
SMAR SMAR
Convergently transcribed gene pairs: H4/H2A, H2B/H1
Coordinate transcription
relation between these two aspects?

S/MAR is detected between H1 and H3 by biochemical experiments
(S/MARt-DB: SM0000037 )
Some SMARs are observed as stress-induced unwound elements ( necessary but insufficient for chromatin loop formation? )
Bubble position coincides with annotated SMAR location
http://smartdb.bioinf.med.uni-goettingen.de/
SMARtDB/browse/index.html

SMAR binding to matrix: recovering the supercoiling stress on the intervening dsDNA
New unwound regions
downstream to convergently- transcribed gene pairs
relation to gene transcription , or even to the coordinate transcription of the whole gene cluster ?
5—H3—H4—H2A—H2B—H1—3
Competition Between Bubbles : Long Range Allosteric Effect (LRAE)

Adapted from: Wang, J.C. 1991. DNA topoisomerases: why so many? Journal of Biological Chemistry 266:6659-6662.
Twin-model of supercoiling domain in gene transcription
SMAR: retaining the negative supercoils around the unwound regions?? Buffer of the generated positive supercoils ??

Homologous gene locus:D.hydei Histone gene cluster
No record in SMARt DB; a prediction
Further support is given when doing the homology analysis: the intergenic sequences differ, but the LRAE is similar

The results are insensitive to parameter
perturbation

More examples supporting the existence of LRAE :
Sorghum v.s. Rice : Sh2/A1 (homologous locu
s)
SM0000063 SM0000064 SM0000065 SM0000066 SM0000067 SM0000068 SM0000069 SM0000070 SM0000071
SM0000032 SM00000170Sorghum v.s. Maize : Adh1 (homologous locus)
SM0000011 SM0000012hsIFNA
SM0000023 SM0000028 SM0000029IGF2
IFNA2 SM0000074 SM0000075
SM0000080 SM0000081cspB
S/MARt-DB is still under construction. Meaningful statistics should be given when there are enough records available.

It’s possible to reveal LRAE by sequence analysis combined with mechanics investigation in a bioinformatic way
LRAE is hopefully an effective regulatory mechanism in gene transcription
(e.g. SMAR can function via LRAE)
Summary

Thanks For Your Attention !





























