Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHE~~I~TRY
Vol. 239, No. 9, September 1964
Printed in U.S.A.
Lipid Metabolism of Euglena gracilis*
D. HULANICKA, J. ERWIN,~ AND K. BLOCH
From the James Bryant Conant Laboratory, Harvard University, Cambridge 38, Massachusetts
(Received for publication, March 6, 1964)
In both plants and animals pathways exist for the conversion of oleic acid to polyenoic acids (1, 2), but these pathways lead to different end products in the two kingdoms. In green plants the double bond system is extended toward the methyl end of the molecule to form linoleic and linolenic acids (9,12, lBocta- decatrienoic acid) (1,3,4). The ciliated protozoa, the amoebae, and some other animal protists share the ability of plants to convert oleic acid to linoleic acid, but all additional double bonds enter in the direction of the carboxyl group of the molecule (5, 6). In higher animals, linoleic acid is not synthesized, but when obtained from the diet, linoleic acid is desaturated to 6,9,12- octadecatrienoic acid and then converted to arachidonic acid by chain elongation and further carboxyl-directed desaturation (2).
The phytoflagellate Euglena gracilis is a protist which exhibits the metabolic behavior of a plant in the light and of an animal in the dark, and it has been shown that this organism synthe- sizes cu-linolenic acid as well as CzO polyenoic acids of the animal type (7, 8). Grown in the dark as a strict heterotroph, this organism contains large amounts of polyenoic CzO acids, including arachidonic acid, and little or no linolenic acid. On the other hand, Euglena cells growing under photoauxotrophic conditions are rich in linolenic acid and contain relatively small amounts of the C& polyenoic acids (7). The physiological significance of the synthesis of linolenic acid in photosynthetic Euglena has previously been discussed (8).
We now wish to describe in more detail the identification of some of the polyenoic acids that occur in photoauxotrophic (grown in light, green) and heterotrophic (grown in dark, etio- lated) euglenids and the pathways by which these acids are formed. The lipid types synthesized by Euglena in the light and in the dark have also been examined. Related studies were carried out with the colorless euglenid Astasia longa.
EXPERIMENTAL PROCEDURE
E. gracilis Z was grown in stationary culture at room tempera- ture on a synthetic medium containing sucrose (9). The cells were inoculated into “low form” 2500-ml Erlenmeyer flasks containing 500 ml of medium, and the cultures were carefully shielded from light. A. Zonga was raised on a similar synthetic medium (10) in the daylight. For photosynthetic growt,h of Euglena, cells were cultured either on the sucrose-containing medium or on an inorganic medium containing small amounts
* Supported by grants-in-aid from the United States Public Health Service, the National Science Foundation, the Life In- surance Medical Research Fund, and the Eugene P. Higgins Trust Fund of Harvard University.
t Present address, Department of Microbiology, School of Medicine, State University of New York, Upstate Medical Center, Syracuse, New York.
of citrate (11) in 4000-ml columns illuminated with banks of daylight fluorescent lamps providing saturating illumination. Citrate serves as a chelating agent but not as a ~carbon source for these flagellates. When cells were grown in the light, the cultures were continuously flushed with a mixture of either air- 5% COz or air-0.50/, CO* under sterile conditions. Cells were harvested in the logarithmic phase of growth by centrifugation at low speed.
Saponijcation and Isolation of Fatty Acids-Cells were sus- pended in a solution of 10 % KOH in 50% aqueous methanoi and digested by heating under reflux for 2 to 3 hours. Non- saponifiable materials were removed by extraction with petro- leum ether, and the fatty acids were extracted with ethyl ether after acidification of the aqueous phase. The acids were con- verted into the methyl esters by treatment with diazomethane and analyzed by gas-liquid chromatography on a column packed with polydiethylene glycol succinate on Chromosorb W, 60 to 80 mesh, and operated at 165-180”. The instrument used was Research Specialties Company model 600 with a &ionization detector. The radioactivity of individual methyl esters was determined as previously described (12). Acids were identified by comparing retention times of the peaks with those of known standards. The values given are retention times relative to a value of 1.0 for methyl stearate.
Separation of Saturated and Unsaturated Fatty Acids-Methyl esters of fatty acids were separated into saturated and unsatu- rated fractions by silicic acid chromatography of their mercuric acetate adducts (12). Mono-, di-, tri-, and polyunsaturated esters were separated by the following adaptation of thin layer chromatography (13). A solution of 1.32 g of AgN03 in 16 ml of distilled water was added to 6.6 g of silica gel G (E. Merck, Darmstadt, Germany), and the mixture was applied to the glass plates. The plates were dried for several hours at 60” and then at 100” for 30 minutes immediately before use. The chromato- grams were developed at room temperature with the solvent n-pentane-ethyl ether-acetic acid (70:30 :0.4) by the ascending technique. The developed plates were sprayed with 2,7-di- chlorofluoresceine and spots were visualized under ultraviolet light. Fractions were identified by comparison with spots of authentic palmitic, stearic, oleic, linoleic, linolenic, and arachi- donic acids (obtained from Applied Science Laboratories, State College, Pennsylvania). By this technique, saturated, mono- unsaturated, and dienoic acids were well separated from each other and from polyenoic acids, the distances between fractions ranging from 1 to 2 cm. Separation between tri- and polyenoic acids was less efficient but could be improved by continuing development for 30 minutes aft,er the solvent front had reached the edge of the plate. When the isolated fatty acids were radio- active they were visualized by autoradiography of the thin layer
2778
by guest on August 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

September 1964 D. Hulanicka, J. Erwin, and K. Bloch 2779
chromatograms. A thin sheet of polyethylene was placed be- tween the x-ray film and the plate to avoid any reaction between the silver nitrate and t.he film.
Further Identijication of Unsaturated Fatty Acids-Individual components were collected aft,er gas-liquid chromatography, dis- solved in absolute methanol, and reduced catalytically in the presence of a platinum catalyst. The reduction products were identified by gas-liquid chromatography. To determine the position of the double bonds in the unsaturated fatty acids, the individual methyl esters were subjected to oxidation by per- manganate-periodate (14). The oxidation products were ester- fied and the dimethyl esters of the dicarboxylic acids and the monomethyl esters of the monocarboxylic acids were identified by gas-liquid chromatography.
Because the retention times of some of the monocarboxylic acid esters and of some of the dicarboxylic acid esters are very similar (e.g. methyl undecanoate and dimethyl glutarate) the ester mixture was first fractionated by chromatography on silicic acid. The monocarboxylic esters were eluted with 5% ether in pentane and the dicarboxylic esters with ether.
For establishing the position of the double bond nearest to the methyl end of the molecule in some of the Czo polyenoic fatty acids the methyl esters were subjected to ozonolysis (15). Ozone was passed into purified dichloromethane for 30 minutes at -60” and the concentration of dissolved ozone was checked by titration with sodium thiosulfate. The fatty acid sample (200 t.o 500 pg) was introduced, and the ozone solution was kept first at -60” for 2 hours and then for 1 hour at room tempera- ture. The ozonides were reduced by treatment with zinc powder in glacial acetic acid at 40” for 15 minutes. The volatile alde- hydes were trapped in an aqueous solution of 2,4-dinitrophenyl- hydrazine and the dinitrophenylhydrazones were extracted with benzene. The dried benzene extracts were reduced in volume and the residues were extracted with methanol-ethyl ether, 1: 1. The hydrazones were then chromatographed on silica gel G thin layer plates. The plates were developed at room temperature with isopropyl ether in n-pentane (1: l), dried, and redeveloped with the same solvent. The separated dinitrophenylhydrazones were eluted from the plates with methanol-ether, 1 :l. The dinitrophenylhydrazone of propionaldehyde was identified by cochromatography with an authentic sample.
Potassium acetate-l-i*C and sodium acetate-3H were purchased from the New England Nuclear Corporation. Linoleic acid-l- i*C and linolenic acid-1-l*C were obtained from the California Corporation for Biochemical Research. 11,14-Eicosadienoic and 8,11,14-eicosatrienoic acids labeled with 3H, and 6,9,12-octa- decatrienoic acid-1-l*C were prepared biosynthetically as de- scribed below.
Preparation of 6,9, I.%Octadecatrienoic Acid-1-‘*C-A flask containing 500 ml of culture medium (9) and 9.4 PC of l-l*C- linoleic acid was inoculated with Tetrahymena pyriformis, and the cells were harvested after 12 hours. After extraction and saponification of the lipids, the isolated fatty acids were esterified and separated by thin layer chromatography (silver nitrate- silica gel) as described above. An aliquot of the trienoic acid fraction was subjected to gas-liquid chromatography and shown to contain a single peak with the retention time of 6,9,12-Cl8 (1.63). The acid recovered (2.74 mg) contained 2 X lo6 d.p.m. or 10% of the radioactivity added to the Tetrahymena culture in the form of l-14C-linoleic acid.
Preparation of Tritium-labeled 1 I , 1 .&Eicosadienoic and 8,li ,
1CEicosatrienoic Acids-E. gracilis Z was grown in the dark in 1 liter of medium (9) with the addition of 2 mc of 3H-acetate. The cells were harvested 20 hours later and the isolated fatty acids were fractionated first by thin layer chromatography and then by gas-liquid chromatography. Two fatty acid fractions were collected, 11,14-eicosadienoic acid (retention time, 2.8; 2.5 X lo6 d.p.m.) and 8,11,14-eicosatrienoic acid (retention time, 3.4; 1.2 x lo6 d.p.m.).
Lipid Analysis-To facilitate lipid analysis, E. gracilis was cultured in the presence of acetate-l-l%, either in the light or in the dark. After harvesting and washing, the cells were extracted three times with 200 ml portions of chloroform-methanol, 2: 1 (v/v). The extracts were filtered and the filtrates were pooled. The cell residue was saved for determination of bound fatty acids. After evaporation the extracted lipids were redissolved in 10 ml of chloroform-methanol (1: 1) and chromatographed on silicic acid (16). The neutral, “pigment,” and phospholipid fractions were further separated by thin layer chromatography (5). Fractions were visualized on thin layer plates by auto- radiography, and the various lipid types were identified by com- parison of RF values with those of standards and by spraying of the spots with specific reagents. Dragendorff reagent was used for visualizing lecithin; ninhydrin for phosphatidylethanolamine and for phosphatidylserine; and 2,7-dichlorofluoresceine and iodine for glycerides. For determining the galactose content of lipid fractions, samples were hydrolyzed in acid (1 ml of 2 N
HCI + 0.2 ml of methanol + 0.1 ml of ethyl ether) for 90 min- utes. The hydrolysates were lyophilized and the residues were extracted with water. Galactose was determined by assay with galactose oxidase (Worthington Biochemical Corporation, Free- hold, New Jersey).
RESULTS
Determination of Double Bond Structures (Table I)
Monounsaturated Acids-The fraction having a retention time of 0.69 on gas chromatograms yielded palmitic acid on catalytic reduction and migrated as a monoenoic acid on thin layer plates. For further examination this hexadecenoic acid was isolated from a larger batch of green Euglena cells. When degraded oxidatively, the acid yielded both CT and CS dicarboxylic acids. The hexadecenoic acid fraction is therefore a mixture of 9-hexa- decenoic acid and of the A7 isomer. Judging from the yield of the two dicarboxylic acids, the relative amounts of these isomeric hexadecenoic acids are approximately equal, at least in green Euglena.
Di- and Polyunsaturated Acids-The procedures customary for structural analysis of polyunsaturated fatty acids have been followed in this investigation. The location of double bonds nearest to the carboxyl group and of those nearest to the methyl terminus of the fatty acid chain is given by the chain lengths of the dicarboxylic acid and monocarboxylic acid fragments, re- spectively, obtained on oxidative degradation of the molecule. The total number of double bonds is deduced from the behavior of the fatty acid on thin layer chromatograms. Structural assignments can then be made if it is assumed that the unsatu- rated fatty acids of E. gracilis have the usual methylene-inter- rupted double bond system. For example, the acid with a re- tention time of 2.8 relative to methyl stearate yields on reduction eicosanoic acid and migrates as a dienoic acid on thin layer chromatograms. Oxidation yields a C1l dicarboxylic acid and
by guest on August 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from
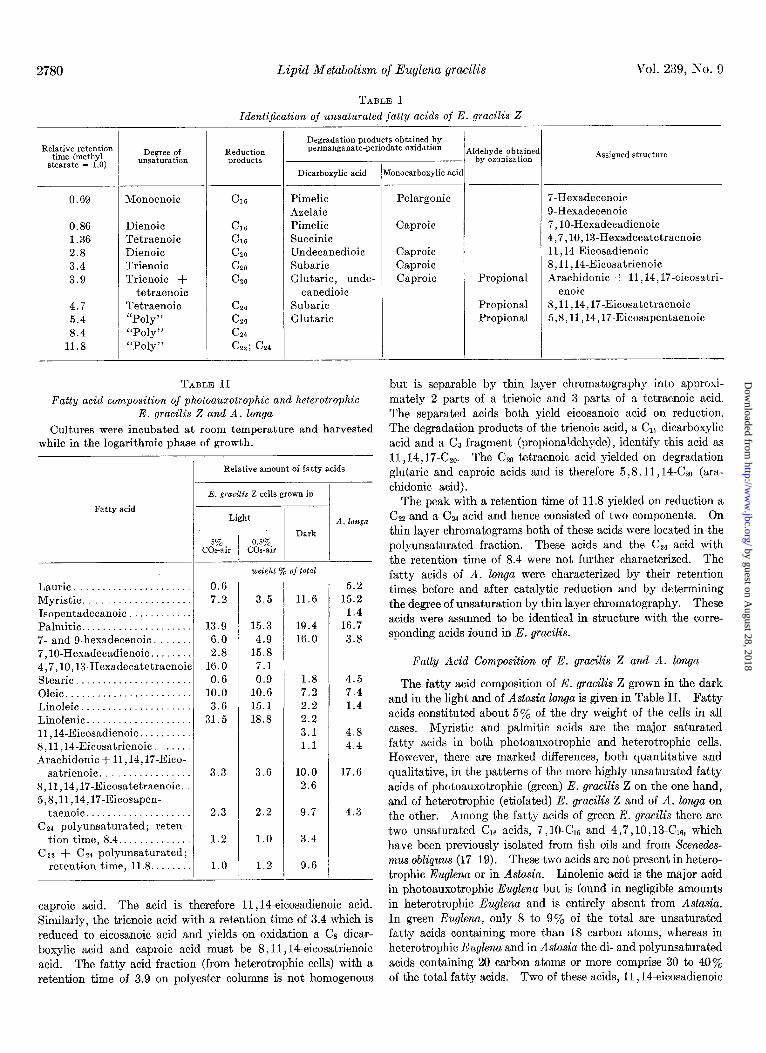
2780 Lipid Metabolism of Euglena gracilis Vol. 239, No. 9
TABLE I
Identi$cation of unsaturated fatty acids of E. grads Z
Degradation products obtained by permanganate-periodate oxidation A Relative retention
time (methyl stearate = 1.0)
ddehyde obtainer by ozonization Assigned structure d Degree of Reduction
unsaturation products
0.69 Monoenoic
Dienoic Tetraenoic Dienoic Trienoic Trienoic +
tetraenoic Tetraenoic “Poly” “Poly” “Poly”
_(
1
-
Dicarboxylic acid [onocarboxylic a&
Pimelic Azelaic Pimelic Succinic Undecanedioic Subaric Glutaric, unde-
canedioic Subaric Glutaric
Pelargonic
Caproic
Caproic Caproic Caproic
7-Hexadecenoic 9.Hexadecenoic 7, IO-Hexadecadienoic 4,7,10,13-Hexadecatetraenoic 11,14-Eicosadienoic 8,11,14-Eicosatrienoic Arachidonic + 11,14,17-eicosatri-
enoic 8,11,14,17-Eicosatetraenoic 5,8,11,14,17-Eicosapentaenoic
0.86 1.36 2.8 3.4 3.9
4.7 5.4 a.4
11.8
Propional
Propional Propional
-
but is separable by thin layer chromatography into approxi- mately 2 parts of a trienoic and 3 parts of a tetraenoic acid. The separated acids both yield eicosanoic acid on reduction. The degradation products of the trienoic acid, a Cl1 dicarboxylic acid and a CB fragment (propionaldehyde), identify this acid as 11,14,17-CzO. The Czo tetraenoio acid yielded on degradation glutaric and caproic acids and is therefore 5,8,11,14-CZO (ara- chidonic acid).
The peak with a retention time of 11.8 yielded on reduction a Czz and a Cza acid and hence consisted of two components. On thin layer chromatograms both of these acids were located in the polyunsaturated fraction. These acids and the C24 acid with the retention time of 8.4 were not further characterized. The fatty acids of A. Zonga were characterized by their retention times before and after catalytic reduction and by determining the degree of unsaturation by thin layer chromatography. These acids were assumed to be identical in structure with the corre- sponding acids found in E. gracilis.
Fatty Acid Composition of E. gracilis Z and A. longa
The fatty acid composition of E. gracilis Z grown in the dark and in the light and of Astasia Zonga is given in Table II. Fatty acids constituted about 5% of the dry weight of the cells in all cases. Myristic and palmitic acids are the major saturated fatty acids in both photoauxotrophic and heterotrophic cells. However, there are marked differences, both quantitative and qualitative, in the patterns of the more highly unsaturated fatty acids of photoauxotrophic (green) E. gracilis Z on the one hand, and of heterotrophic (etiolated) E. gracilis Z and of A. longa on the other. Among the fatty acids of green E. gracilis there are two unsaturated Cl6 acids, 7,l O-C& and 4,7,10, 13-C16, which have been previously isolated from fish oils and from Scenedes- mus obliquus (17-19). These two acids are not present in hetero- trophic Euglena or in Astasia. Linolenic acid is the major acid in photoauxotrophic Euglena but is found in negligible amounts in heterotrophic Euglena and is entirely absent from Astasia. In green Euglena, only 8 to 9% of the total are unsaturated fatty acids containing more than 18 carbon atoms, whereas in heterotrophic Euglena and in Astasia the di- and polyunsaturated acids containing 20 carbon atoms or more comprise 30 to 40% of the total fatty acids. Two of these acids, 11,14-eicosadienoic
TABLE II
Fatty acid composition of photoauxotrophic and heterotrophic E. yracilis Z and A. longa
Cultures were incubated at, room temperature and harvested while in the logarithmic phase of growth.
Relative amount of fatty acids
Fatty acid
E. gracilis Z cells grown in
Light
Dark 5% 0.5%
CO&r CO*-air
Laurie ..................... Myristic ................... Isopentadecanoic ........... Palmitic. ................... 7- and 9-hexadecenoic ...... 7,10-Hexadecadienoic. ...... 4,7,10,WHexadecatetraenoi Stearic ..................... Oleic ....................... Linoleic .................... Linolenic ................... lI,14-Eicosadienoic. ........ 8,11,14-Eicosatrienoic . ..... Arachidonic + 11,14,17-Eico-
satrienoic. ............... 8,11,14,17-Eicosatetraenoic. 5,8,11,14,17-Eicosapen-
taenoic ................... Cs4 polyunsaturated; reten-
tion time, 8.4 ............. C22 + C24 polyunsaturated;
retention time, 11.8 .......
-
!C
-
0.6 7.2
13.9 6.0 2.8
16.0 0.6
10.0 3.6
31.5
5.2 3.5 11.6 15.2
1.4 15.3 19.4 16.7 4.9 16.0 3.8
15.8 7.1 0.9 1.8 4.5
10.6 7.2 7.4 15.1 2.2 1.4 18.8 2.2
3.1 4.8 1.1 4.4
3.3 3.6 10.0 17.6 2.6
2.3 2.2 9.7 4.3
1.2 1.0 3.4
1.0 1.2 9.6
A. longa
caproic acid. The acid is therefore 11,1/i-eicosadienoic acid. Similarly, the trienoic acid with a retention time of 3.4 which is reduced to eicosanoic acid and yields on oxidation a CS dicar- boxylic acid and caproic acid must be 8,11,14-eicosatrienoic acid. The fatty acid fraction (from heterotrophic cells) with a retention time of 3.9 on polyester columns is not homogenous
by guest on August 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

September 1964 D. Hulanicka, J. Erwin, and K. Bloch 2781
acids and 8,11,14-eicosatrienoic acids, which are probably pre- cursors of arachidonic acid (see below), could not be detected in green Euglena. The fatty acid fraction with a retention time of 3.9 was shown by degradation to be a mixture of arachidonic acid and 11,14,17-eicosatrienoic acid. Because these two acids have identical retention times, their relative contribution to this fraction is difficult to assess, but from arguments to be presented below, we consider it likely that in green Euglena the major com- ponent of the mixture is the 11,14,17-triene and that in hetero- trophic Euglena and in Astasia this fraction consists primarily of arachidonic acid. In one case the fatty acids of heterotrophic Euglena were found to contain arachidonic acid and the 11,14, 17-C&, acid in a ratio of 3:2. It will also be noted (Table II) that there are significant differences in the concentration of some of the polyunsaturated fatty acids between green Euglena cells grown in atmospheres of 5% COz-air and in 0.5% COTair. In particular, there is a substantial increase of the Cl6 tetraenoic acid and of linolenic acid in the cells grown in the presence of the higher COz concentration. Since several laboratories have shown that COz enhances the photosynthetic evolution of oxygen (26-22), this CO2 effect on the synthesis of certain polyunsatu- rated fatty acids supports the suggestion that linolenic acid plays a role in reactions of plant photosynthesis that lead to oxygen evolution (7).
The fatty acid patterns and some of the lipids of green and etiolated E. gracilis have recently been investigated by Rosen- berg (23). In his report the absence of linolenic acid in etiolated Euglena (7, 8) is confirmed, but no mention is made of the ac- cumulation of Go, C&2, and Cz4 polyunsaturated acids in hetero- trophic cells. The results of Rosenberg are difficult to compare with ours since growth conditions were not specified and since the double bond structures of the unsaturated fatty acids were not determined.
Biosynthesis of Unsaturated Acids in Heterotrophic Euglenids
The structure of the major unsaturated fatty acids found in plants suggests that the further desaturation of oleic acid to polyenoic acids occurs chiefly, if not exclusively, by placement of double bonds in the direction of the methyl end of the molecule. In higher animals the further desaturation of oleic acid is a minor reaction (2) but, to the extent that it occurs, it proceeds in the direction of the carboxyl group of the fatty acids. In photo- auxotrophic E. gracilis, the pattern of biosynthesis seems to be primarily that of plants, since the major product of desaturation of oleic acid is linolenic acid.
In heterotrophic euglenids, and to some extent in Euglena grown in the light on organic media, the fatty acid spectrum is extremely complex, particularly in the region of polyunsaturated fatty acids containing 20 carbon atoms or more (8, 24). In considering pathways to the various types of fatty acid, in the light of their chemical structure and of knowledge gained from other organisms, two major principles of biosynthesis may be assumed to operate: chain elongation by addition of CZ units, and extension of double bond systems to form dienes and poly- enes of the divinylmethane type. In a given case it must then be decided whether a particular double bond is introduced before or after the fatty acid chain is lengthened. For example, 8,11, 1Beicosatrienoic acid can arise from linoleic acid’ by way of 6,9,12-octadecatrienoic acid, i.e. by desaturation followed by Cz addition, or by way of 11, lbeicosadienoic acid, i.e. by the con- verse sequence of steps.
3,6,9,12,X-&, 8,11,14,17-CZO 5,8,11,14-C&
5,8,11,14,17-Czo
FIG. 1. Pathways for the biosynthesis of Cl8 and Go polyun- saturated fatty acids in E. gracilis. The open arrow indicates the major route in photoauxotrophic cells. The relative impor- tance of the various fatty acid transformations in Euglena cells grown in the dark is indicated by the width of the black arrows. Question marks refer to reactions for which the evidence is indi- rect.
In order to distinguish between and to assess the relative significance of alternate pathways (see Fig. l), cells were grown in the presence of labeled linoleic acid and of the isomeric lino- lenic acids. The incorporation of radioactivity into the fatty acids of heterotrophic Euglena is shown in Table III, and the corresponding experiments with Astasia, in Table IV. The labeled acids were readily incorporated (10 to 30%) and metabo- lized by the organisms. Radioactivity from the fatty acids supplied to the medium was not extensively randomized as a result of degradation into smaller units in any of these experi- ments since carbon from linoleic acid-l-14C was not appreciably incorporated into oleic acid, stearic acid, or shorter chain acids. The observed labeling pattern, therefore, reflects direct conver- sion of the substrates rather than reutilization of degradation products.
The experiment with linoleic acid-l-l% shows that the 9,12- CU dienoic acid is effectively converted to all polyunsaturated fatty acids containing 18 or more carbon atoms (Table III). Linoleic acid can clearly serve as the common and initial pre- cursor, whatever the sequence of chain elongations and desatura- tions leading to the structurally diverse polyunsaturated prod- ucts.
Starting from linoleic acid, one of the pathways leads to linolenic acid by methyl-directed desaturation. In heterotrophic Euglena, this is a minor route, in agreement with the low lino- lenate content of these cells. The total activities and relative specific activities are several times greater in 11,lCeicosadienoic acid, indicating that in cells grown in the dark, chain elongation of linoleate is preferred over direct desaturation. Since the 6,9,12-isomer of linolenate does not accumulate in Euglena either in the light or in the dark, the role played by this acid in the formation of the CzO trienes and tetraenes cannot be assessed.
With linolenic acid-I-14C as the substrate there is incorporation
1 It will also be assumed that the initial desaturation of oleate produces exclusively linoleic acid, even though the formation of the isomeric 6,9X& acid, known to occur in higher animals (2) cannot be ruled out.
by guest on August 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2782 Lipid Metabolism of Euglena gracilis
TABLE III
Vol. 239, No. 9
Radioactivity in fatty acids of E. gracilis Z grown in dark in presence of labeled precursors
One liter of sucrose-containing medium (9) was inoculated with E. gracilis and the culture was incubated at room temperature in the dark for 3 days. The radioactive substrate, dissolved in 0.1 ml of ethanol, was then added to the culture and incubation continued for 36 hours under the same conditions. Total fatty acids were isolated from the harvested and washed cells and were separated as de- scribed in “Experimental Procedure.”
Organism incubated with
Linoleic acid-l-W, 4.4 X 106 Linolenic acid-l-W, 4.4 X 106 6,9,12-Octadecatrienoic d.p.m. d.p.m. acid-l-%, 0.94 X 106 d.p.m.
Fatty acids
Total d.p.m. RSA*
0.5 0.6 1.8 0.6
422.0 79.0
Total d.p.m.
52 90 52 55
110 2600
RSA*
0.3 0.8 4.0 2.2
27.0 165.0
122.1 90 11.4 129.0 40 8.5 160.5 1020 38.2
258.1 4960 352.0 334.0 93 15
248.0 3012 401.2
Total d.p.m. RSA*
Myristic ............................................ 78 Palmitic ............................................ 98 Stearic ............................................. 52 Oleic ............................................... 42 Linoleic ............................................ 5032 Linolenic ........................................... 870 6,9,12-Octadecatrienoic. ............................ 11,14-Eicosadienoic. ................................ 2426 8,11,14-Eicosatrienoic . ............................. 1142 Arachidonic + 11,14,17-eicosatrienoic . ............. 9916 8,11,14,17-Eicosatetraenoic ......................... 5,8,11,14,17-Eicosapentaenoic ...................... 8748 C24 polyunsaturated; retention time, 8.4 ............. 3442 Cs2 polyunsaturated + C24 polyunsaturated; reten-
tiontime, 11.8 .................................... 4120
* Relative specific activity.
2 14
204 124
3530 160 452 864
130 144
62
3.3 2.6
49.5 1.9 2.2 3.6
0.8 8.4
1.6
TABLE IV
Distribution of radioactivity in fatty acids of A. longa The culture conditions were the same as described for E. gracilis inTable III, except that the cultures were not protected from light.
I Organisms incubated with
Fatty acids
-
.- Linolenic acid-l-W, 4.4 X 106
d.p.m. Linoleic acid-l-W, 4.4 X 106
d.p.m. 6,9,12-Octadecatrienoic
acid-l-W 0 94 X 106 d.p.m. , .
Total d.p.m. RSA’
.- Total d.p.m. RSA* Total d.p.m. RSA*
Myristic ............................................ 44 Palmitic ............................................ 81 Palmitoleic ......................................... 47 Stearic ............................................. 69 Oleic ............................................... 69 Linoleic ............................................ 2,225 Linolenic ........................................... 6,9,12-Octadecatrienoic ............................ 11,14-Eicosadienoic ................................. 3,120 8,11,14-Eicosatrienoic . ............................. 7,020 Arachidonic + 11,14,17-eicosatrienoic ............... 19,730 5,8,11,14,17-Eicosapentaenoic . ..................... 10,550 Cs4 polyunsaturated; retention time, 8.4 ............. 14,720 Ct2 + C&4 polyunsaturated; retention time, 11.8. .... 12,680
* Relative specific activity.
0.5 0.6 4.5 3.7 1.8
620.0
28 70 26 38 20 84
2,022
0.16 0.28 0.81 0.48 0.19 6.4
1,220 116 2.1 224 46 0.8 208 1,406 5.2 690 14,204 250.0 362 673 405.0 208 8,346 401.0
4 0.0 20 0.1 16 1.3
2,222 59.3 58 3.5
300 21.1 1,082 20.8
256 18.3 234 11.7
74 4.6
into the fraction containing arachidonic acid and 11,14,17-C& tions is (retention time, 3.9), into 5,8,11,14,17-Car,, and into the C&a + Cz4 fraction (retention time, 11.8). Thus, even though the Linolenate 2 11,14,17-C~0--,8,11,14,17-Cz0+5,8,11,14,17-Cza
synthesis of linolenic from linoleic acid is relatively slow in heterotrophic Euglena, the acid is efficiently elongated and fur- i.e. chain elongation followed by two carboxyl-directed desatura- ther desaturated to CzO polyunsaturated acids when added to the tions. There is no direct evidence for the alternative route, growth medium. The most likely pathway for these transforma- desaturation of the Cis acid followed by CB addition, because Cis
by guest on August 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

September 1964 D. Hulaniclca, J. Erwin, and K. Bloch 2783
tetra- and pentaenoic acids have not been found in Euglena. Indications for the existence of alternate routes will be discussed below.
In the experiment with the labeled linolenic acid, the specific activity calculated for the fraction containing 11,14,17-CzO and arachidonic acid is quite low by comparison with the values for the CZo pentaenoic acid or the Czz and Cs4 acids. The likely explanat,ion is that radioact,ivity in this fraction is associated only with the 11,14,17-CzO triene component, since a conversion of linolenate to arachidonate is improbable on structural grounds.
In the experiment with 6,9,12-octadecatrienoic acid-l-i4C (Table III), the total activities and relative specific activities of other fatty acids were relatively low. The data indicate that some conversion to 8,11,14-CzO and arachidonic acid occurred but that the 6,9,12-isomer of 1inolenat.e is a much less efficient precursor of these acids than linoleic acid. Even if 6,9, 12-Cle were synt,hesized by Euglena, which is not certain, it could not contribute to a major extent to the formation of Go polyun- saturated fatty acids in this organism. The acid is more effec- tively utilized by the euglenid mut.ant A. Zonga (Table IV).
Experiment with Linolenic-l-14C and -i1,14-Czo-3H, and 8,11,14-C&H Acids
The experiments described above show that in etiolated E. gracilis linoleic acid can enter two pathways to polyunsaturated acids, one proceeding by way of linolenic acid to 5,8,11,14,17- Cza and longer chain fatty acids, and another pathway involving initial Cz addition, followed by carboxyl-directed desaturations to form arachidonic acid and the uncharacterized acids of greater chain length. In order to test which of these routes predomi- nates when the cells are grown in the dark, experiments were performed with two acids that serve as independent indicators of the two pathways. In one of these experiments, the precursors were linolenic acid-l-14C and tritium-labeled 11,14-eicosadienoic acid, and in a second experiment they were linolenic-1-i4C and tritium-labeled 8,11,14-eicosatrienoic acids (Table V). The fatty acids isolated from the cells were assayed and the ratio 3H : 14C was calculated in each case.
The interpretation of some of these data is complicated by the fact that one of the key chromatographic fractions (retention time, 3.9) contains a mixture of arachidonic acid and 11,14,17- eicosatrienoic acid. For the reasons stated earlier, we assume that in the experiments in Table V, the 14C content of this frac- tion represents the 11,14,17-CzO triene, which is logically derived from linolenic acid, and that the tritium content is due to arachidonic acid, a logical product of the 8,11,14-C& acid. The 1% data of Table V confirm those of Table III in showing that the major pathway for linolenic acid in heterotrophic Euglena is Cz addition to 11,14,17-CZO followed by the introduction of two double bonds in the direction of the carboxyl group, pro- ducing5,8,11,14,17-Czo. The data, however, do not eliminate an alternate route to the C&o pentaene by way of 6,9,12,15-C& and 3,6,9,12,15-C!rs, a pathway discussed by Klenk (25). The tritium dat,a of Table V show that the 11,14-C& acid formed by chain elongation of linoleic acid (Table III) is desaturated in the direction of the carboxyl group to form 8,11,14-C&0 and arachi- donic acid, and that there is another branch from 8,11,14-C&0 leading to 8,11,14,17-CZO (methyl-directed desaturation) and then to 5,8,11,14, 17-CzO (carboxyl-directed desaturation). In both double labeling experiments, the 3H:14C ratios were about twice as great in the fraction containing arachidonic acid and
TABLE V
Distribution of radioactivity among fatty acids of E. gracilis Z in “double labeling” experiments
For each of the two experiments 1 liter of sucrose-containing medium (9) was inoculated with E. gracilis and the culture was incubated at room temperature in the dark for 3 days. An ethanol solution of the two radioactive substrates was added to the cul- ture and incubation was continued for 36 hours under the same conditions. Total fatty acids were isolated from the harvested and washed cells and were separated as described in “Experi- mental Procedure.”
Fatty acids isolated
Linoleic Linolenic . 11,14-Eicosadienoic. 8,11,14-Eicosatrienoic Arachidonic + 11,14,
17.eicosatrienoic. 8,11,14,17-Eicosatet-
raenoic 5,8,11,14,17-Eicosa-
pentaenoic. .
:.
‘I
.I
Organisms incubated with
Linolenic acid-l-‘&C Linolenic-1-Y and and 11,14-eicosadienoic
acid-3H* 8,11,14-eicosatrienoic
acid-aHt
Total activity Total activity ‘H:“C - ‘H:W
1°C “C aH
d.p.m.
105 1 63 10,845 I 0
251 16,544 64 ’ 351
1,072 1.170
492 j 391
3,050 835
2
L
0.6 0.0 l6.7 5.4
I
a.p.m. 215 7
2,079 0 454 800 110 3,097
0.0
0.0
1.8 28.1
1.09 3,452 7,347 2.1
0.6 1,325 1,256 0.95
0.27 6,237 474 0.08
* Linolenic acid-l-l%, 2.8 X 106d.p.m., plus 11,14-C20-3H, 2.2 x lo6 d.p.m.
t Linolenic acid-l-W, 1.1 X 10” d.p.m., plus 8,11,14-C&H, 1.1 X lo6 d.p.m.
11,14, 17-CzO as they were in the 8,11,14,17-C& tetraene frac- tion. These findings suggest that the pathway from ll,14C20 via 8,11,14-CzO to arachidonic acid is preferred over the route leading to the 8,11,14,17-C&0 tetraene; i.e. double bond intro- duction in the direction of the carboxyl group predominates in the synthesis of polyunsaturated fatty acids by heterotrophic cells.
The most reliable information provided by the 3H:14C ratios (Table V) relates to the origin of the 8,11,14, 17-CZO and 5,8, ll,14,17-C20 acids. Both acids contain tritium as well as W, and therefore both arise from linolenic acid and also independ- ently from the 11,14-C& diene by way of the 8,11, 14CZ0 triene. The two pathways seem to contribute equally to the formation of the 8,11,14, 17-CzO tetraene (which is a very minor constituent of the Euglena lipids), since the 3H : 14C ratio is near unity in this acid. On the other hand, the 5,8,11,14,17-C& pentaenoic acid, which accounts for about 10% of the total fatty acids of heterotrophic EugZena, must arise chiefly from the 14C-labeled Iinolenic acid, since the 3H :i4C ratio is substantially higher in the Czo tetraene (precursor) than it is in the CZO pentaene (product). Another metabolite of linolenate and precursor for the CZO pentaene must be postulated, and this could be 3,6,9,12,15-C&, a hypothetical desaturation product of linolenate. Other trans- formations which are feasible on structural grounds are 11,14-
c20 + 11,14,17-C20 and arachidonic acid + 5,8,11,14,17-C20. These reactions are probably of lesser significance in hetero-
by guest on August 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2784 Lipid Metabolism of Euglena gracilis Vol. 239, No. 9
TABLE VI
Lipid fractions of E. gracilis grown in light and in dayk
Cel:;xx;; in Cells grown in the light on inorganic medium
Lipid fraction Relative Fatty Relative Fatty amount* acid amount acid from
f$;
70 of total 70 of total rg/mg 1. Neutral lipids. 20.5 36 8.2 10 -1 2. “Pigment” fraction. . 5.9 13 70.4 66 67.5 3. Phospholipid fraction.. - . 18.7 27 16.4 20 4. “Polar” lipid fraction. 55.6 24 4.3 4 -
* Calculated from dry weights. t None of the lipid fractions of Euglena grown in the dark con-
tained galactose. $ Dash indicates that galactose is not detectable.
trophic Euglena since the predominant pattern in these cells is carboxyl-directed desaturation.
Cl6 Unsaturated Fatty Acids
Euglena contains hexadecenoic acids of varying degrees of unsaturation and with double bond structures which are con- sistent with the following sequence of reactions.
7-c& + 7,lO.CM 4 +4,7,10,13-CM
In heterotrophic cells the concentration of Cl6 monoene is high while the Cl6 diene and Cl6 tetraene are either absent, or present only in trace amounts. The pattern is reversed in photoauxo- trophic cells, the Cl6 tetraene comprising as much as 16% of the total fatty acids (Table II). It also seems significant that under conditions favoring optimal photosynthesis-in cells grown in 5yo COa-air-the content of Cl6 tetraene is twice as high as in cells grown in an atmosphere containing only 0.5% cot.
The Cl6 monoene fraction was shown by chemical degradation to be a mixture of the A7 and As isomers. Since the mechanism for the synthesis of monounsaturated fatty acids in Euglena is unknown, the possibility must be considered that the 7-C& acid arises by P-oxidation of oleate. By analogy the 4,7,10,13-C& tetraene could be an oxidation product of 6,9,12,15-Cra, an acid which has been isolated by Klenk and Brockerhoff (26). It is curious that in the case of the C16 acids photoauxotrophic metabo- lism in Euglena favors the formation of a polyunsaturated deriva- tive in which the double bond system is extended maximally in the directions both of the carboxyl group and of the terminal methyl group of the molecule. By contrast, the light-stimulated desaturations of the C1a acids extend the initial double bond system only in the direction of the methyl group of the molecule.
Biosynthesis of Polyunsaturated Fatty Acids in A. longa
The fatty acid composition of this colorless euglenid is quali- tatively the same as that of etiolated Euglena except for the complete absence of linolenic acid and 8,11,14,17-CZa in Astasia (Table I). The conversion patterns of radioactive linoleate and of linolenate were also similar in the two organisms, apart from the apparent inability of Astasia to convert added linoleic acid to linolenate. Nevertheless, although the formation of lino- lenate cannot be demonstrated in Astasia, dietary linolenate is
metabolized by this organism in the same manner as it is by etiolated cells of wild-type Euglena. Astasia is also capable of converting the 6,9,12 isomer of linolenate to arachidonic acid, presumably by way of 8,11,14-C&a. The fact that Astasia contains the 5,8,ll,14,17-C20 pentaene and forms this acid from linoleic acid and from 6,9,12-C&, although less efficiently than from linolenate, suggests the existence of a route to the CzO pentaene either from arachidonate or by way of 8,11,14,17-CZo. In any event, the formation of the 5,8,11,14,17-C& acid by Astasia indicates that this euglenid mutant has retained the ability to carry out both methyl-directed and carboxyl-directed desaturations which are characteristic of wild-type Euglena.
Lipid Composition of E. gracilis
Growth conditions profoundly affect not only the fatty acid composition but also the relative amounts of the various lipid fractions of E. gracilis (Table VI). In photoauxotrophically grown cells, 70% of the total lipid is associated with the “pig- ment” fraction, while in etiolated cells more than half of the total lipids are in the “polar” lipid fraction. In green cells there is very little of this “polar” lipid, while the “pigment” fraction is virtually absent in etiolated cells. The percentage of phos- pholipids is essentially the same in both green and etiolated cells, but the latter contain approximately twice as much neutral lipid as the former. The distinction between “polar” lipids and phospholipids is probably arbitrary for the following reason. The “polar” lipids, Fraction V as isolated by the procedure of Nelson and Freeman and stated by these authors to contain lecithin and sphingomyelin (16), behaved as a single component on thin layer chromatograms when developed with a solvent system capable of separating various phospholipids. The R, of the fraction was identical with that of lecithin and gave a posi- tive test with the Dragendorff reagent. The “polar” lipid is therefore a lecithin with an increased affinity for silicic acid, conceivably because of its high content of polyunsaturated fatty acids (see below). The “polar” lipids are therefore more prop- erly classified as a subfraction of the phospholipids, and on this basis, the total phospholipid content of etiolated cells is 74.3% compared to only 20.7% in green cells.
When analyzed by thin layer chromatography, the neutral lipids from green cells were found to consist primarily of di- glycerides, while in etiolated cells, triglycerides were the major component. The diglycerides of green cells may be the pre- cursors of the galactolipids rather than of triglycerides as dis- cussed below.
In photoauxotrophic cells the “pigment” fraction was found to be a composite of several pigments and of six different lipids separable by thin layer chromatography. These lipids have not been further examined except that they all yielded galactose on acid hydrolysis. The presence of galactosylglycerides in Euglena grown in the light has been reported by Carter et al. (27). In heterotrophic Euglena, the “pigment,” fraction is a minor com- ponent, containing little pigment and only a single lipid com- ponent, which failed to yield galactose on hydrolysis. This finding confirms the absence of galactose-containing lipids in heterotrophic Euglena previously observed by Carter et al. (27).
The major phospholipid of green Euglena is phosphatidyl- serine; phosphatidylcholine and phosphatidylethanolamine are minor components. By contrast, the phospholipid fraction of heterotrophic cells (Fraction 3, Table VI) contains primarily phosphatidylethanolamine with lesser amounts of phosphatidyl-
by guest on August 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from
September 1964 D. Hulanicka, J. Erwin, and K. Bloch 2785
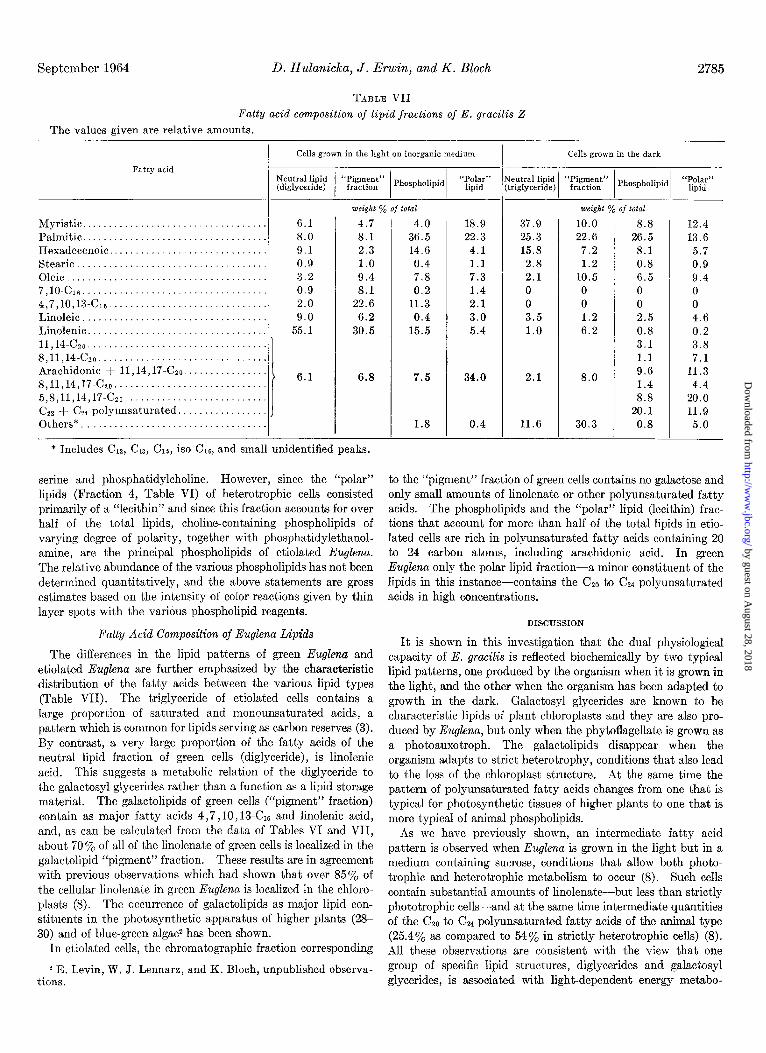
TABLE VII
Fatty acid composition of lipid fractions of E. gracilis Z
The values given are relative amounts.
I Cells grown in the light on inorganic medium Cells grown in the dark
Myristic. ........................ Palmitic. ........................ Hexadecenoic .................... Stearic .......................... Oleic ............................ 7,lO.C~ ......................... 4,7,10,13-c16 .................... Linoleic ......................... Linolenic ........................ 11,14-czo ........................ 8,11,14-c~~ ...................... Arachidonic + 11,14,17-CZO. ..... 8,11,14,17-czo ................... 5,8,11,14,17-czo ................. CZZ + C24 polyunsaturated ....... Others*. ........................
. 6.1 4.7 4.0 18.9 37.9 10.0 . 8.0 8.1 36.5 22.3 25.3 22.6
9.1 2.3 14.6 4.1 15.8 7.2 0.9 1.0 0.4 1.1 2.8 1.2 3.2 9.4 7.8 7.3 2.1 10.5 0.9 8.1 0.2 1.4 0 0 2.0 22.6 11.3 2.1 0 0
. 9.0 6.2 0.4 3.0 3.5 1.2 . 55.1 30.5 15.5 5.4 1.0 6.2
6.8
. . . .
* Includes Clz, C13, C15, iso-C,e, and small unidentified peaks.
7.5
1.8
serine and phosphatidylcholine. However, since the “polar” lipids (Fraction 4, Table VI) of heterotrophic cells consisted primarily of a “lecithin” and since this fraction accounts for over half of the total lipids, choline-containing phospholipids of varying degree of polarity, together wit.h phosphatidylethanol- amine, are the principal phospholipids of etiolated Euglena. The relative abundance of the various phospholipids has not been determined quantitatively, and the above statements are gross estimates based on the intensity of color reactions given by thin layer spots with the various phospholipid reagents.
Fatty Acid Composition of Euglena Lipids
The differences in the lipid patterns of green Euglena and etiolated Euglena are further emphasized by the characteristic distribution of the fatty acids between the various lipid types (Table VII). The triglyceride of etiolated cells contains a large proportion of sat.urated and monounsaturated acids, a pattern which is common for lipids serving as carbon reserves (3). By contrast, a very large proportion of the fatty acids of the neutral lipid fraction of green cells (diglyceride), is linolenic acid. This suggests a metabolic relation of the diglyceride to the galactosyl glycerides rather than a function as a lipid storage material. The galactolipids of green cells (“pigment” fraction) contain as major fatty acids 4,7,10, 13-CX and linolenic acid, and, as can be calculated from the data of Tables VI and VII, about 70 y0 of all of the linolenate of green cells is localized in the galactolipid “pigment” fraction. These results are in agreement with previous observations which had shown that over 85% of the cellular linolenate in green Euglena is localized in the chloro- plasts (8). The occurrence of gala&lipids as major lipid con- stituents in the photosynthetic apparatus of higher plants (2% 30) and of blue-green algae2 has been shown.
In etiolated cells, the chromatographic fraction corresponding
2 E. Levin, W. J. Lennarz, and K. Bloch, unpublished observa- tious.
34.0
0.4
2.1
11.6
8.0
30.3
8.8 12.4 26.5 13.6 8.1 5.7 0.8 0.9 6.5 9.4 0 0 0 0 2.5 4.6 0.8 0.2 3.1 3.8 1.1 7.1 9.6 11.3 1.4 4.4 8.8 20.0
20.1 11.9 0.8 5.0
to the “pigment” fraction of green cells contains no galactose and only small amounts of linolenate or other polyunsaturated fatty acids. The phospholipids and the “polar” lipid (lecithin) frac- tions that account for more than half of the total lipids in etio- lated cells are rich in polyunsaturated fatty acids containing 20 to 24 carbon atoms, including arachidonic acid. In green Euglena only the polar lipid fraction-a minor constituent of the lipids in this instance-contains the Czo to C&4 polyunsaturated acids in high concentrations.
DISCUSSION
It is shown in this investigation that the dual physiological capacity of E. gracilis is reflected biochemically by two typical lipid patterns, one produced by the organism when it is grown in the light, and the other when the organism has been adapted to growth in the dark. Galactosyl glycerides are known to be characteristic lipids of plant chloroplasts and they are also pro- duced by Euglena, but only when the phytoflagellate is grown as a photoauxotroph. The galactolipids disappear when the organism adapts to strict heterotrophy, conditions that also lead to the loss of the chloroplast structure. At the same time the pattern of polyunsaturated fatty acids changes from one that is typical for photosynthetic tissues of higher plants to one that is more typical of animal phospholipids.
As we have previously shown, an intermediate fatty acid pattern is observed when Euglena is grown in the light but in a medium containing sucrose, conditions that allow both photo- trophic and heterotrophic metabolism to occur (8). Such cells contain substantial amounts of linolenate-but less than strictly phototrophic cells-and at the same time intermediate quantities of the CtO to C, polyunsaturated fatty acids of the animal type (25.4% as compared to 54% in strictly heterotrophic cells) (8). All these observations are consistent with the view that one group of specific lipid structures, diglycerides and galactosyl glycerides, is associated with light-dependent energy metabo-
by guest on August 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2786 Lipid Metabolism of Euglena gracilis Vol. 239, No, 9
lism and that phosphatidylcholine and phosphatidylethanol- amine, lipids of the type found in a,nimal mitochondria, occur in association with heterotrophic energy metabolism (oxidative phosphorylation). At the morphological level the light-dark transition of Euglena is expressed by the disappearance of chloroplasts and by a marked increase in mitochondrial struc- tures (31).
The lipid types and fatty acid structures of Euglena are not novel but are found elsewhere in either animal or plant tissues (32, 33). The unique feature of lipid metabolism in Euglena is the dual nature of the biosynthetic pattern which follows that of plants in the light and of animals in the dark, without change of genotype.
The pathways for the formation of polyunsaturated fatty acids in Euglena as indicated by the patterns of double bonds and by the results of tracer experiments are summarized in Fig. 1. Apparently the reactions of the linolenate, or plant pathway, and of the arachidonate, or animal pathway, exist at all times, i.e. both in photoauxotrophic and in heterotrophic cells. How- ever, the relative rates at which the two types of pathways operate differ markedly in the light and in the dark. Photo- auxotrophic cells do produce C&, CZz, and CZ4 polyunsaturated fatty acids, including arachidonate (8, 24), but the amounts produced in the light are very much smaller than in heterotrophic cells. Conversely, heterotrophic cells contain small amounts of linolenate along with very much larger amounts of C&o to CZ4 polyunsaturated fatty acids of the animal type. This low content of linolenate in cells grown in the dark is not necessarily a true measure of the extent of linolenate synthesis. As the experiments reported here indicate, linolenate synthesis probably continues in the dark at a substantial rate, but instead of accu- mulating it is now further desaturated in the direction of the carboxyl group and its chain is lengthened to form 5,8,11,14,17- C 20, either by way of 11,14, 17-CQo or by way of Cl8 tetra- and Cl* pentaenoic acids. In either event, the reactions charac- teristically promoted by growth in the dark are chain elongation and introduction of additional double bonds in the carboxyl direction.
Arachidonic acid can apparently be synthesized by Euglena
from linoleic acid by two routes: either by way of 6,9, 12-C1s (y- linolenic acid) and homo-y-linolenic acid (8,11, 14-Czo) , the route demonstrated in vertebrates (2)) or by initial chain elonga- tion to 11,14-C&0 followed by two carboxyl-directed desatura- tions. The latter sequence has recently been shown in rat liver enzymes by Stoffel (34), and evidence for the existence of this route in a soil amoeba has been presented by Korn (35). The relative contribution of the two pathways for the synthesis of arachidonic acid in Euglena cannot be precisely evaluated on the basis of the present data.
The photoauxotrophic pattern of polyunsaturated fatty acid synthesis undergoes two major modifications in the dark, en- hancement of chain elongation to fatty acids containing 20 or more carbon atoms and introduction of additional double bonds, primarily in the direction of the carboxyl group of the molecule. These two reactions appear to be suppressed by light or light- induced reactions of the chloroplast.
SUMMARY
The lipid and fatty acid composition of Euglena gracilis Z has
been investigated. Cells grown as st,rict photoauxotrophs and
cells grown heterotrophically after dark-adaptation were ana- lyzed.
The major polyunsaturated fatty acids of photoauxotrophic cells are linolenic acid and 4,7,10,16-hexadecatetraenoic acid. The principal lipids in these cells are a diglyceride, galacto- lipids, and a phospholipid fraction containing chiefly phospha- tidylserine.
The principal polyunsaturated fatty acids of heterotrophic cells are arachidonic acid and (or) 11,14,17-eicosatrienoic acid, 5,8,11,14,17-eicosapentaenoic acid, and uncharacterized C 22 and CQ4 polyunsaturated acids. Heterotrophic cells contain triglycerides and large amounts of phospholipids, but lack galactolipids.
Some of the pathways to polyunsaturated fatty acids have been investigated in heterotrophic Euglena and in Astasia longa. Except for the formation of a C16 tetraenoic acid, the biosynthetic pathways are qualitatively the same in the light and in the dark. However in heterotrophic Euglena two types of reactions pre- dominate, chain elongation of linoleate and linolenate, and introduction of additional double bonds in the direction of the carboxyl groups of the CZo acids. On dark adaptation the lipids and fatty acids of Euglena change from a “plant” to an “animal”’ pattern.
Acknowledgments-The authors wish to thank Dr. L. Provasoli of Haskins Laboratories for a gift of the culture of Astasia Zonga, and Dr. G. Holz of the State University of New York, Upstate Medical Center, Syracuse, for the culture of EugZena gracilis Z.
REFERENCES
1. SIMMONS, R. O., AND QUACKENBUSH, F. N., J. Am. Oil Chem- ists’ SW., 31, 441 (1962).
2.
3.
4. 5. 6.
7.
MEAD, J. F., in K. BLOCH (Editor), Lipide metabolism, John Wiley and Sons, Inc., New York, 1960 p. 41.
HILDITCH, T. P., The chemical constitution of natural fafats, John Wiley and Sons, Inc., New York, 1956, p. 146.
DEBUCH, H.. 2. Naturforsch.. Pt. b. 16. 246 (1961). ERWIN, j., AND BLOC& K., j. Biol: Chem., i38, i618 (1963). ERWIN, J., HULANICKA, D., AND BLOCH, K., J. Comp. Bio-
them. and Physiol., in press. ERWIN, J., AND BLOCH, K., Biochem. and Biophys. Research
Communs., 9, 103 (1962). 8. 9.
ERWIN, J., AND BLOCH, K., Biochem. Z., 338, 496 (1963). HUTNER, S. H., BACH, M. K., AND Ross, G. I. M., J. Protozool.,
3, 101 (1956). 10. PADILLA, G. M., AND JAMES, T. W., Exptl. Cell Research, 20,
401 (1960). 11. CRAMER, M., AND MYERS, G., Arch. Mikrobiol., 17, 384 (1952). 12. GOLDFINE, H., AND BLOCH, K., J. Biol. Chem., 236, 2596 (1961). 13. MANGOLD, M. K., J. Am. Oil Chemists’ Sot., 28, 78 (1961). 14. VAN RUDLOFF, E., Can. J. Chem., 34, 1413 (1952). 15. KLEIN. F., AND DE JONG, K.. Rec. trav. chim.. ‘76. 1285 (1956). 16. NELSON, 6. J., AND FREEMAN, N. K., J. l&ok Chem:, 234,
1375 (1959). 17.
18.
PASCHKE, R. F., AND WHEELER, D. H., J. Am. Oil Chemists’ Sot., 31, 81 (1954).
19.
20.
21.
22.
SCHLENK, H., MANGOLD, H. K., GELLERMANN, J. L., LINK, W. E.. MORISSETTE. R. A.. HOLMAN. R. T.. AND HAYES, H., J. ‘Am. Oil Chenksts’ So;., 37, 547 ‘(1960). ’
KLENK, E., AND KNIPPRATH, W., 2. physiol. Chem., 317, 243 (1959).
WARBURG, O., AND KRIPPAHL, G., 2. Naturforsch., Pt. b, 13. 509 (1958).
ABELES, F. B., BROWN, A. H., AND MAYNE, B. C., Plant Phys- iol., 36, 202 (1961).
STERN, B. K., AND VENNESLAND, B., J. Biol. Chem., 237, 596 (1963).
by guest on August 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

September 1964 D. Hulanicka, J. Erwin, and K. Bloch 2787
23. ROSENBERG, A., Biochemistry, 2, 1148 (1963). 24. KORN, E. D., Biochem. and Biophys. Research Communs., 14,
1 (1963). 25. KLENK, E., Ezperientia, 17, 199 (1961). 26. KLENH, E., AND BROCKERHOFF, H., Z. vhusiol. Chem.. 307.
272 (i957 j . _” I
27. CARTER, H. E., OHNO, K., NOJIMA, S., TIPTON, C. L., AND STANAZEV, N. Z., J. Livid Research. 2. 215 (1961).
32. SHORLAND, F. B., in M. FLORKIN AND H. S. MASON (Editors), Comparative biochemistry, Vol. d, Academic Press, Inc., New York, 1962, p. 41.
28. BENSON, A.‘A., W&ER, R., FERRARI,‘R: A., AND KILLER, J. 33. KLENK, E., KNIPPRATH, W., EBERHAGEN, D., AND KOOF, H. A., J. Am. Chem. Sot., 80, 4740 (1958). P., 2. physiol. them., 334, 44 (1963).
29. ZILL, L. D., AND HARMON, E. A., Biochim. et Biophys. Acta, 34. STOFFEL, W., Z. physiol. Chem., 336, 71 (1963). 67, 573 (1962). 35. KORN, E. D., J. Biol. Chem., 239, 396 (1964).
30. SASTRY, P. S., AND KATES, M., Biochim. et Biophys. Acta, 70, 214 (1963).
31. WOLKEN, J. J., Euglena: an experimental organism for bio- chemical and biophysical studies, Rutgers University Press, New Brunswick,.N1 J., 1961.
by guest on August 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

D. Hulanicka, J. Erwin and K. BlochEuglena gracilisLipid Metabolism of
1964, 239:2778-2787.J. Biol. Chem.
http://www.jbc.org/content/239/9/2778.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/239/9/2778.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on August 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from





























