Embed Size (px)
Citation preview

JOURNAL OF VIROLOGY, Aug. 2010, p. 8193–8201 Vol. 84, No. 160022-538X/10/$12.00 doi:10.1128/JVI.00685-10Copyright © 2010, American Society for Microbiology. All Rights Reserved.
Lentiviral Vif Degrades the APOBEC3Z3/APOBEC3H Protein of ItsMammalian Host and Is Capable of Cross-Species Activity�
Rebecca S. LaRue,1,2,3,4 Joy Lengyel,1,2,3 Stefan R. Jonsson,1,2,3,5
Valgerdur Andresdottir,5 and Reuben S. Harris1,2,3,4*Department of Biochemistry, Molecular Biology and Biophysics,1 Institute for Molecular Virology,2 Center for Genome Engineering,3
and Comparative and Molecular Biology Graduate Program,4 University of Minnesota, Minneapolis, Minnesota 55455, andUniversity of Iceland, Institute for Experimental Pathology Keldur v/Vesturlandsveg, 112 Reykjavík, Iceland5
Received 30 March 2010/Accepted 25 May 2010
All lentiviruses except equine infectious anemia virus (EIAV) use the small accessory protein Vif to coun-teract the restriction activity of the relevant APOBEC3 (A3) proteins of their host species. Prior studies havesuggested that the Vif-A3 interaction is species specific. Here, using the APOBEC3H (Z3)-type proteins fromfive distinct mammals, we report that this is generally not the case: some lentiviral Vif proteins are capable oftriggering the degradation of both the A3Z3-type protein of their normal host species and those of several othermammals. For instance, SIVmac Vif can mediate the degradation of the human, macaque, and cow A3Z3-typeproteins but not of the sheep or cat A3Z3-type proteins. Maedi-visna virus (MVV) Vif is similarly promiscuous,degrading not only sheep A3Z3 but also the A3Z3-type proteins of humans, macaques, cows, and cats. Incontrast to the neutralization capacity of these Vif proteins, human immunodeficiency virus (HIV), bovineimmunodeficiency virus (BIV), and feline immunodeficiency virus (FIV) Vif appear specific to the A3Z3-typeprotein of their hosts. We conclude, first, that the Vif-A3Z3 interaction can be promiscuous and, second,despite this tendency, that each lentiviral Vif protein is optimized to degrade the A3Z3 protein of its mam-malian host. Our results thereby suggest that the Vif-A3Z3 interaction is relevant to lentivirus biology.
Lentiviruses are a unique class of complex retroviruses thatencode a variety of accessory proteins in addition to the re-quired Gag, Pol, and Env proteins. The archetypal lentivirus,human immunodeficiency virus type 1 (HIV-1), infects hu-mans, but other members include simian immunodeficiencyvirus (SIV), bovine immunodeficiency virus (BIV), maedi-visna virus (MVV), caprine arthritis-encephalitis virus(CAEV), equine infectious anemia virus (EIAV), and felineimmunodeficiency virus (FIV), which infect monkeys, cattle,sheep, goats, horses, and cats, respectively. The HIV-1 acces-sory protein viral infectivity factor (Vif) has been extensivelystudied because of its essential function in inhibiting the cel-lular antiretroviral human APOBEC3G (A3G) protein (43).HIV-1 Vif binds to human A3G (and other A3 proteins) andserves as an adaptor to link it to an ELOC-based E3 ubiquitinligase complex (30, 51, 52). A3G is then polyubiquitinated anddegraded by the cellular proteasome (7, 15, 29, 30, 43, 46, 52).
Due to the potential therapeutic value of disrupting thishost-pathogen interaction, a significant amount of work hasbeen invested in defining the important contact residues be-tween A3G and HIV-1 Vif. Primate A3G homologs have beenuseful tools in this effort, as many fail to be neutralized byHIV-1 Vif despite a relatively high degree of sequence simi-larity. For example, while HIV-1 Vif effectively neutralizeshuman A3G, it does not neutralize African green monkey A3Gor rhesus macaque A3G despite 77% and 75% identity, re-spectively (4, 26, 27, 41, 51). The differential capacity of the
HIV-1 and SIVagm Vif proteins to degrade the A3G proteinsof their hosts led to demonstrations that residue 128 is a keydeterminant: D128 made each A3G protein susceptible toHIV-1 Vif and K128 made each A3G protein susceptible toSIVagm Vif (4, 26, 41, 51). This apparent on/off switch ledto the prevailing model that the Vif-A3 interaction is speciesspecific. However, even early data sets showed at least twohints that the story was more complex. First, the identity of theA3G residue 128 (K or D) does not diminish the interactionwith the Vif proteins of SIVmac or HIV-2 (41, 51). Second,SIVmac Vif was shown to potently counteract the A3G proteinsfrom rhesus macaque (as expected) but also those from hu-man, African green monkey, and chimpanzee (27). Therefore,the implication from these studies is that the full nature of theA3-Vif interaction has yet to be elucidated.
Although A3G has clearly served as the prototype for un-derstanding the A3-Vif interaction, a growing number of stud-ies indicate that other A3s are also capable of restricting len-tivirus replication and interacting with Vif. A3G is one of sevenhuman A3 proteins (A3A to -H) encoded in tandem on chro-mosome 22 (7, 16, 49). All but A3A have been implicated inthe restriction of HIV-1 replication (reviewed in references 1,10, and 45). For instance, human A3H has been shown torestrict HIV-1 replication and is susceptible to degradation byHIV-1 Vif (8, 37, 47). A3H is a Z3-type DNA deaminasecharacterized by a conserved threonine and a valine, in addi-tion to the canonical H-x1-E-x23–28-C-x2–4-C zinc-coordinatingmotif (23). The Z3-type deaminase is unique in that only onecopy exists in all mammals whose genomes have been se-quenced. It is encoded by a five-exon gene located at the distalend of each mammal’s A3 locus (adjacent to CBX7). Addi-tional observations suggest that the Z3-type deaminases ap-
* Corresponding author. Mailing address: University of Minnesota,Department of Biochemistry, Molecular Biology and Biophysics, 321Church Street S.E., 6-155 Jackson Hall, Minneapolis, MN 55455.Phone: (612) 624-0457. Fax: (612) 625-2163. E-mail: [email protected].
� Published ahead of print on 2 June 2010.
8193
on February 9, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

pear to have the capacity to restrict the Vif-deficient lentivi-ruses of their hosts. For example, African green monkey A3Hcan restrict the replication of SIVagm and is susceptible todegradation by SIVagm Vif, and the cat A3Z3 can restrict thereplication of FIV and is susceptible to degradation by FIV Vif(33, 37, 48).
Here, we take advantage of the fact that all sequenced mam-mals have a single A3Z3-type protein to test the hypothesisthat these proteins are of general relevance to lentivirus re-striction and to clarify the species-specific nature of the mam-malian A3Z3/lentiviral Vif relationship. First, we ask if human,rhesus macaque, cow, sheep, and cat A3Z3-type proteins areall capable of retrovirus restriction. Second, we ask whetherthey are susceptible to Vif-mediated degradation in a host-specific manner. We show that each lentiviral Vif protein canindeed neutralize the Z3-type A3 protein of its host species.However, we were surprised to find that several of the Vifproteins, particularly SIVmac and MVV Vif, can neutralize abroad number of A3Z3 proteins irrespective of the species oforigin and overall degree of similarity. These data indicate thatthe A3-Vif interaction is more promiscuous than previouslyappreciated. Such broad functional flexibility may be relevantto understanding past retroviral zoonoses and predicting po-tential future events. We conclude that the A3Z3-Vif interac-tion is conserved on a macroscopic level, consistent with animportant role in viral replication and particularly in specieslike artiodactyls and felines with fewer A3 proteins.
MATERIALS AND METHODS
Sequence alignments. Mammalian A3Z3 protein sequences were aligned andscored using the default matrix BLOSUM62 by protein BLAST (2, 14). The follow-ing A3Z3 sequences were compared and tested in this study: human A3H (gbEU861361), rhesus macaque A3H (gb DQ507277), cow A3Z3 (gb EU864536),sheep A3Z3 (gb EU864543), and cat A3Z3 (gb EU011792). Protein sequences forthe lentiviral Vif proteins were aligned using T_coffee version 5.31 (35). AllGenBank accession numbers for the lentiviral Vif protein sequences arelisted in the “Vif expression plasmids” section below.
APOBEC3 expression plasmids. The cow A3Z1, A3Z2, A3Z3, and A3Z2-Z3and sheep A3Z3 genes have been described previously (17, 23). These cDNAswere subcloned from the pEGFP-N3 expression vector by being digested withKpnI and SalI and inserted into a similarly digested pcDNA3.1 eukaryotic ex-pression vector with no stop codon. This expression vector added three consec-utive hemagglutinin (HA) epitope tags to the C-terminal domain (CTD) of theencoded A3 protein. All of the remaining mammalian A3Z3 genes were alsoinserted into the pcDNA3.1 expression vector, and details are described below.The human A3H (haplotype II) and rhesus macaque A3H cDNAs were kindlyprovided by M. Emerman (37, 38). These A3H sequences contained an N-terminal domain (NTD) tag that was removed by PCR amplification with theseprimers: human, forward, 5�-NNN NGA GCT CGG TAC CAC CAT GGC TCTGTT AAC AGC CGA AAC-3�, and reverse, 5�-NNN GTC GAC TCC GGACTG CTT TAT CCT CTC AAG CC-3�, and rhesus macaque, forward, 5�-NNNNGA GCT CGG TAC CAC CAT GGC TCT GCT AAC AGC-3�, and reverse,5�-NNN GTC GAC TCC TCT TGA GTT GCG TAT TGA CGA TG-3�. Theuntagged human A3H (haplotype II) was amplified with the same forwardprimer and a reverse primer with a stop codon, 5�-NNN NNG TCG ACT CAGGAC TGC TTT ATC CTC TCA AGC CGT C-3�. To ensure the same sequencefor the cat A3Z3 gene from reference 33, RNA was extracted from Crandell-Rees feline kidney (CRFK) cells and reverse transcribed and 3� rapid amplifi-cation of cDNA ends (RACE) was performed on the cDNA as described pre-viously (23). The following primers were used to amplify the cat A3Z3 genewithout a stop codon: forward, 5�-NNN NGA GCT CAG GTA CCA CCA TGAATC CAC TAC AGG AA-3�, and reverse, 5�-NNN NGT CGA CTT CAA GTTTCA AAT TTC TGA A-3�. The mouse A3Z2-Z3 and human A3G expressionplasmids were described elsewhere (11, 13, 17).
Vif expression plasmids. The lentiviral Vifs chosen for codon optimization(GenScript Corporation) match HIV-1IIIB (gb EU541617), SIVmac239 (gb
AY588946), BIVBIM127 (gb M32690), MVV Icelandic strain 1514 (gb M60610),and FIVNSCU (gb M25381). All of the Vifs were amplified by PCR from thepUC57 vector (GenScript Corporation) and cloned into the pVR1012 vector(Vical Co.). The HIV-1LAI Vif in the pVR1012 vector was a generous gift fromX. F. Yu, and it is not codon optimized. All Vifs were amplified by PCR anddesigned to add a c-myc sequence to the CTD, digested with NotI or SalI andBamHI, and cloned into the pVR1012 vector. The following primers were usedfor the lentiviral Vifs: HIV-1IIIB Vif, forward, 5�-CAC AAC AAG GTG GGCGCA GCG GCG TAC CTT GCA CTG GCC-3�, and reverse, 5�-GGA TCCCTA CAG ATC CTC TTC TGA GAT GAG TTT TTG TTC GTG GCC ATTCAT TGT-3�; HIV-1LAI Vif, forward, 5�-GTC GAC GCC ACC ATG GAAAAC AGA TGG-3�, and reverse, 5�-NNN NGG ATC CCT ACA GAT CCTCTT CTG AGA TGA GTT TTT GTT CGT GTC CAT TCA TTG T-3�; SIVmac
Vif, forward, 5�-AAG TAC CAG GTT CCT GCT GCG GCG TAT CTG GCACTC AAA-3�, and reverse, 5�-GGA TCC CTA CAG ATC CTC TTC TGA GATGAG TTT TTG TTC GGC CAG GAT ACC CAG-3�; BIV Vif, forward, 5�-CTCTAC CCC ACG CCA CGC CGC GGC GCG GCT GGC AGC TCT G-3�, andreverse, 5�-GGA TCC CTA CAG ATC CTC TTC TGA GAT GAG TTT TTGTTC AGG GTG TCC GCT CAG-3�; MVV Vif, forward, 5�-AAC ACT AACCCC AGA GCC GCG GCG AGA CTT GCC CTG CTT-3�, and reverse,5�-GGA TCC CTA CAG ATC CTC TTC TGA GAT GAG TTT TTG TTC CTCAAA AAT GCT CTC-3�; and FIV Vif, forward, 5�-AAC AGC CCA CCA CAGGCC GCG GCG CGG CTG GCC ATG CTG-3�, and reverse, 5�-GGA TCCCTA CAG ATC CTC TTC TGA GAT GAG TTT TTG TTC CAG GGA GCCAGA CCA-3�.
Vif mutants were constructed using standard site-directed mutagenesis proto-cols (Stratagene). The following primers were used on the codon-optimizedlentiviral Vifs: HIV-1IIIB Vif (SLQ to AAA), forward, 5�-CAC AAC AAG GTGGGC GCA GCG GCG TAC CTT GCA CTG GCC-3�, and reverse, 5�-GGCCAG TGC AAG GTA CGC CGC TGC GCC CAC CTT GTT GTG-3�; SIVmac
Vif (SLQ to AAA), forward, 5�-AAG TAC CAG GTT CCT GCT GCG GCGTAT CTG GCA CTC AAA-3�, and reverse, 5�-TTT GAG TGC CAG ATACGC CGC AGC AGG AAC CTG GTA CTT-3�; BIV Vif (SLQ to AAA),forward, 5�-CTC TAC CCC ACG CCA CGC CGC GGC GCG GCT GGC AGCTCT G-3�, and reverse, 5�-CAG AGC TGC CAG CCG CGC CGC GGC GTGGCG TGG GGT AGA G-3�; MVV Vif (SLQ to AAA), forward, 5�-AAC ACTAAC CCC AGA GCC GCG GCG AGA CTT GCC CTG CTT-3�, and reverse,5�-AAG CAG GGC AAG TCT CGC CGC GGC TCT GGG GTT AGT GTT-3�;and FIV Vif (TLQ to AAA), forward, 5�-AAC AGC CCA CCA CAG GCCGCG GCG CGG CTG GCC ATG CTG-3�, and reverse, 5�-CAG CAT GGCCAG CCG CGC CGC GGC CTG TGG TGG GCT GTT-3�. The followingprimers were used for HIV-1LAI Vif (SLQ to AAA): forward, 5�-GAC ATAACA AGG TAG GAG CTG CTG CAT ACT TGG CAC TAG CA-3�, andreverse, 5�-TGC TAG TGC CAA GTA TGC AGC AGC TCC TAC CTT GTTATG TC-3�.
HIV-GFP infectivity assays. Human 293T cells were cultured in Dulbeccomodified Eagle medium (DMEM) supplemented with 10% fetal bovine serumand 0.5% penicillin-streptomycin. At 50% confluence, cells were transfected(TransIt; Mirus) with 1 �g HIV-green fluorescent protein (GFP) cocktail [0.44�g of CS-CG (32), 0.28 �g of pRK5/Pack1(Gag-Pol), 0.14 �g pRK5/Rev, 0.14 �gof pMDG (vesicular stomatitis virus G protein [VSV-G]-Env)], 100 ng of A3 orempty vector, and 20 ng of codon-optimized Vif or 400 ng of wild-type HIV-1LAI
Vif with corresponding empty vector. The ratio of A3 to Vif constructs was basedon whether or not the Vif was codon optimized. All experiments were performedin triplicate except where noted in the legends.
After 48 h, virus-containing supernatants were harvested and purified bycentrifugation to remove any remaining producer cells. Producer cells wereprocessed for Western analysis and GFP� flow cytometry (Cell Lab QuantaSC-MPL; Beckman Coulter) to measure protein expression levels and transfec-tion efficiencies, respectively. The purified supernatants were placed on fresh293T target cells. After 72 h, target cells were harvested and infectivity (GFP)was measured by flow cytometry. Data were analyzed using FlowJo flow cytom-etry analysis software, version 8.7.1.
Western blotting of cell lysates and viral particles. Producer cell lysates wereharvested and pooled from triplicate experiments. Lysates and viral particlepellets were resuspended in 2� Laemmli sample buffer (25 mM Tris, pH 6.8, 8%glycerol, 0.8% SDS, 2% 2-mercaptoethanol, 0.02% bromophenol blue) andheated at 98°C degrees for 10 min, and a fraction of the sample was run on a 12%SDS-PAGE gel. Protein was transferred to a polyvinylidene difluoride (PVDF)membrane (Millipore). Membranes with cell lysates were first probed with anti-c-myc (Sigma-Aldrich) for detection of Vif expression and then incubated with ahorseradish peroxidase (HRP)-conjugated secondary antibody and developedusing a chemiluminescent HRP antibody detection reagent (HyGLO; Denville
8194 LARUE ET AL. J. VIROL.
on February 9, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

Scientific). Blots were then stripped (0.2 M glycine, 1.0% SDS, 1.0% Tween 20,pH 2.2), and this protocol was repeated with an anti-HA antibody (Covance) todetect A3 proteins and with antitubulin (Bio-Rad) to monitor protein loading.To detect human A3H, we used an anti-human monoclonal A3H antibody kindlyprovided by M. Emerman (24).
RESULTS
Mammalian A3Z3 proteins are relatively conserved in com-parison to lentiviral Vifs. All mammals have only one Z3-typeA3 domain (Fig. 1A). We previously described the Z3-type A3proteins of cattle and sheep and calculated evidence for posi-tive selection in nonhuman lineages (23). Primate and catA3Z3 proteins also show evidence for positive selection (23,
33, 38). Accordingly, these mammalian A3Z3 proteins showsignificant variation, ranging from 48% to 88% identity and62% to 92% similarity (Fig. 1B). The percent identity andsimilarity of the mammalian A3Z3 protein sequences correlatewith those for evolutionarily related species, where human andrhesus macaque A3Z3 proteins are 83% identical (90% simi-lar) and cow and sheep A3Z3 proteins are 88% identical (92%similar) (Fig. 1A and B). The cat A3Z3 protein is the mostremoved, ranging from 51% to 52% identity (65% to 66%similarity) with the human and rhesus macaque A3Z3s and62% identity (74% to 77% similarity) with the cow and sheepA3Z3s. Although A3Z3 proteins (or A3 proteins in general) donot share high sequence identity, as is the case for essentialcellular genes like those that encode the core of the ribosome,there are clearly conserved residues and motifs such as theconserved zinc-binding domain found in all A3 proteins andresidues unique to A3Z3 proteins (22).
In contrast, the Vif proteins of lentiviruses are vastly differ-ent in primary amino acid sequence. Alignments of knownlentiviral Vif protein sequences show less than 30% identity,even between evolutionarily related viruses (36, 42). However,despite low sequence identity between different Vifs, the pro-teins share a highly conserved S/TLQY/RLA motif (Fig. 1C)(36, 42). This virus-specific motif in HIV-1, HIV-2, differentprimate SIVs, BIV, and MVV Vifs is essential for bindingELOC, which is part of a ubiquitin ligation complex (ELOB/C,CUL2 or CUL5, and RBX) capable of degrading A3s (25).The primate lentiviral Vifs (HIV and SIV) are distinct fromthe other mammalian Vifs in that they share a zinc-bindingdomain, HX5CX17-18CX3–5H (HCCH), which binds to CUL5(25). BIV and MVV Vif lack this domain and therefore do notcoimmunoprecipitate with the CUL5 protein when expressedin human cells (25). MVV Vif, on the other hand, coimmuno-precipitates the CUL2 protein, indicating that nonhuman Vifsmay utilize a different Cullin (25). Due to broad conservationof the Vif S/TLQY/RLA motif, we constructed triple aminoacid substitution derivatives (S/TLQ to AAA) of each lentiviralVif to use as controls. This mutant is unable to bind ELOC,and it is a common negative control for many HIV-1 and SIVVif studies (e.g., references 20, 25, 44, and 52).
Representative mammalian A3Z3 proteins can restrict HIV.Every completely annotated mammalian A3 locus to date hasonly one copy of a Z3-type A3 gene (A3H-like). We hypothe-sized that all A3Z3-type proteins are neutralized by their host-specific lentiviral Vif. A3Z3 proteins from human, rhesus ma-caque, and cat were selected because these mammals areinfected with a Vif-containing lentivirus and are capable ofrestricting HIV or its host lentivirus in the absence of a func-tional Vif protein (8, 12, 24, 33, 34, 37, 48, 54). The A3Z3proteins of cow and sheep were also chosen for the represen-tative panel because these artiodactyls are also infected withVif-containing lentiviruses. Previous studies in our lab haveshown that the cow and sheep A3Z2-Z3 can restrict HIV (17).The cow and sheep A3 proteins are also naturally expressed assingle domain variants (23), making them more ideal for com-parisons with the Z3-type A3s of other mammals.
First, we performed a series of single-cycle HIV-GFP infec-tivity assays to test whether the single domain cow A3 proteins(A3Z1, A3Z2, and A3Z3) could restrict an HIV-based virusconstruct. The HIV-GFP viral cocktail was transfected into
FIG. 1. Comparison of A3Z3 and Vif sequences. (A) The related-ness of the mammals relevant to this study. The numbers at the branchnodes are approximate divergence times in millions of years. (Adaptedfrom reference 22). (B) Percent sequence identity and similarity be-tween all the mammalian A3Z3 proteins. Abbreviations for differentmammals: Hs, human; Rh, rhesus macaque; Bt, cow; Oa, sheep; Fc,cat. (C) Alignment of the ELOC binding motifs of the lentiviral Vifsused in this study. The numbers are the first and last residue numbers.
VOL. 84, 2010 APOBEC3-Vif INTERACTION PROFILES 8195
on February 9, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

293T cells alongside an empty pcDNA3.1 vector or one of thecow A3 proteins in the presence of empty pVR012 vector, BIVVif-myc, or the BIV VifSLQ-AAA-myc. A portion of the pro-ducer cell virus-containing supernatant was used to infect fresh293T cells to measure infectivity by GFP� flow cytometry. Wewere surprised that HIV-GFP was restricted equally well bythe cow single domain A3Z3 protein and by the double domainA3Z2-Z3 protein (Fig. 2A). The cow A3Z1 protein showed noanti-HIV activity, despite being a very active DNA deaminase(23). As controls used previously by many labs, human A3Gand mouse A3 both restrict the infectivity of HIV-GFP (e.g.,references 17, 21, 27, 50, and 55).
In parallel experiments, we examined the Vif susceptibilityof the cow A3 proteins and found that BIV Vif restores theinfectivity of viruses produced in the presence of cow A3Z3and A3Z2-Z3 but not those produced with human A3G ormouse A3 (Fig. 2A). These data suggested that BIV Vif mightinteract only with the cow Z3-type deaminase domain. How-ever, immunoblots of the proteins expressed in the producercell extracts showed that the Z2-type domain was also clearlysusceptible to Vif-mediated degradation despite the fact thatthis domain is not antiviral (Fig. 2B).
As anticipated by the infectivity data, BIV Vif also degradedthe cow A3Z3 and A3Z2-Z3 proteins. None of the cow A3s aredegraded in the presence of mutant BIV VifSLQ-AAA-myc (Fig.2). Overall, these experiments demonstrated that BIV Vif trig-gers the degradation of both Z2- and Z3-type A3s and that Viffunction requires the conserved SLQ motif (and therefore
likely also the interaction with ELOC). In addition, the resultsshow that BIV Vif either does not interact with or does nottrigger the degradation of the Z1-type A3.
BIV Vif function appears species specific in contrast to thatof the related MVV Vif protein. We next asked whether BIVVif could neutralize other mammalian A3Z3 proteins besidescow A3Z3. HIV-GFP was produced in the presence of a di-verse set of mammalian A3Z3 proteins, and as describedabove, the producer cells were analyzed by immunoblottingand the resulting viral supernatants were analyzed by infectingsusceptible target cells. The A3Z3 proteins showed a range ofHIV-GFP restriction phenotypes, with rhesus macaque A3Happearing to have the most potency and the sheep A3Z3 theleast (Fig. 3A). Some of these differences may be due in part toslight variations in protein expression. However, consistentwith the species-specific hypothesis, BIV Vif rescued the in-fectivity only of viruses produced in the presence of cow A3Z3.This counterrestriction effect correlated with greatly reducedcellular cow A3Z3 protein levels. In contrast, even the sheepA3Z3, which is 92% similar to the cow A3Z3, showed no signof being degraded.
To ask whether these observations would be reciprocal andpossibly generalizable, we turned to the most closely relatedlentivirus-host interaction, that between MVV Vif and thesheep A3s. The predicted evolutionary distance between thesheep and cow is similar to that between the human and rhesusmacaque, approximately 28 million years (Fig. 1A) (3). BIVand MVV have similar genome organizations (36), and it isreasonable to presume that these viruses arose from a commonancestor (9, 19). However, in comparison to the species-spe-cific activity of BIV Vif, MVV Vif counteracted the restrictionactivity of nearly all of the A3Z3 proteins (Fig. 3B). MVV Vifneutralized the sheep and cow A3Z3 proteins with nearly100% efficiency, and it was also very effective at degrading thehuman, rhesus macaque, and cat A3Z3 proteins. These effectswere equally apparent at the protein level in immunoblots ofproducer cell extracts. Interestingly, despite MVV Vif havingno effect on the restriction capability of human A3G and themouse A3, protein degradation was also detected for theseintended non-single domain A3Z3 controls (human A3G �Z2Z1 and mouse A3 � Z2Z3). These phenotypes are notobserved with the MVV VifSLQ-AAA-myc construct, but thismay be due to the apparent instability caused by the SLQmutations (Fig. 3B). Taken together, these data demonstratethat MVV Vif lacks both species specificity and Z-type speci-ficity, based on its capability of triggering the degradation of abroad array of mammalian A3 proteins that contain differentcombinations of the Z-type domains.
MVV Vif-mediated degradation of the A3s is not simply anartifact of degrading all cellular proteins, because tubulin lev-els were unaffected and toxicity was not observed (Fig. 3B anddata not shown). Such promiscuity was unanticipated, so wenext asked whether MVV Vif has a preference for degradingsheep A3Z3. We analyzed the infectivity of HIV-GFP pro-duced in the presence of sheep A3Z3, human A3H, mouse A3,or human A3G and the range of MVV Vif levels (0, 5, 20, and50 ng plasmid per transfection). Even the lowest MVV Viflevels were sufficient to rescue HIV-GFP infectivity by coun-teracting sheep A3Z3 or human A3H (data not shown). How-ever, immunoblot analysis of the producer cell lysates showed
FIG. 2. Antiretroviral properties of cow A3 proteins and their sen-sitivity to BIV Vif. (A) Relative infectivity of HIV-GFP produced inthe presence of the indicated A3s and a vector control (open bars),BIV Vif (black bars), or BIV VifSLQ-AAA (gray bars). The doublevector control (empty pcDNA3.1 and pVR1012 vectors) was normal-ized to 1, and the error bars report the standard errors of the meansfrom 3 independent experiments. (B) Immunoblots showing the pro-ducer cell levels of A3 proteins (anti-HA), Vif (anti-myc), and tubulin.�, vector control; �, wild-type Vif; m, VifSLQ-AAA mutant.
8196 LARUE ET AL. J. VIROL.
on February 9, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

hsA3H rhA3H btA3Z3 fcA3Z3oaA3Z3 mmA3none
+- mVif
1.2
1.0
0.8
0.6
0.4
0.2
0
+- m +- m +- m +- m +- m +- m
36
28
Rel
ativ
e In
fect
ivity
No VifBIV VifBIV Vif SLQ->AAA
hsA3H rhA3H btA3Z3 fcA3Z3oaA3Z3 mmA3none
+- mVif
1.2
1.0
0.8
0.6
0.4
0.2
0
+- m +- m +- m +- m +- m +- m
Rel
ativ
e In
fect
ivity
hsA3G
+- m
36
28
A
B
No VifMVV VifMVV Vif SLQ->AAA
BIV Vif-myc
A3-HA
α tub
MVV Vif-myc
A3-HA
α tub
FIG. 3. Sensitivity of mammalian A3Z3 proteins to BIV and MVV Vif. (A) Relative infectivity of HIV-GFP produced in the presence of theindicated A3s and a vector control (open bars), BIV Vif (black bars), or BIV VifSLQ-AAA (gray bars). The double vector control was normalizedto 1, and the error bars report the standard errors of the means from 3 independent experiments. Producer cell immunoblots are shown below forA3 proteins (anti-HA), Vif (anti-myc), and tubulin. The mouse A3 signals were faint on the original full immunoblot, and so an image of a longerexposure was inserted in the identical position for presentation. (B) Relative infectivity of HIV-GFP produced in the presence of the indicated A3sand a vector control (open bars), MVV Vif (black bars), or MVV VifSLQ-AAA (gray bars). The figure is organized and labeled as described above.
8197
on February 9, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

that only the sheep A3Z3 was efficiently degraded at the lowestconcentrations, suggesting that MVV Vif has some preferencefor sheep A3Z3.
HIV-1 Vif neutralization of human A3H and other mamma-lian A3Z3 proteins. Prior reports have indicated that HIV-1Vif fails to neutralize rhesus macaque A3H or cat A3Z3 (34,48). Recent studies with HIV-1 Vif and human A3H (haplo-type II) have yielded conflicting results, with some showingneutralization and others not (8, 12, 24, 37, 54). To furtherinvestigate the potential interaction between HIV-1 Vif andhuman A3H and, if apparent, to ask whether it is speciesspecific, we did a series of HIV-GFP infection experiments asdescribed above with HIV-1 Vif from the LAI strain and ourpanel of mammalian A3Z3s. As observed in a prior study withHIV-1LAI Vif (24), HIV-GFP infectivity was partly restoredand the level of cellular human A3H slightly diminished (Fig.4A). HIV-GFP infectivity in the presence of the cow, sheep,and mouse A3s was also improved significantly by expressingHIV-1LAI Vif, but this effect was not accompanied by a signif-icant change in cellular protein levels. It is unclear whether allof these effects were dependent on the conserved SLQ motif,because the mutated protein did not express well in compari-son to the wild-type protein. Overall, however, the phenotypesattributable to HIV-1LAI Vif were quite modest in comparisonto results with other lentiviral Vif proteins or data with HIV-1Vif and human A3G (Fig. 2 and 3) (see below and also priorstudies, e.g., references 6, 29 to 31, 44, 46, 52, and 53).
We therefore wondered whether the C-terminal HA epitopetag might be protecting human A3H from Vif-mediated deg-radation. To address this point, we compared the sensitivitiesof human A3H-HA and untagged A3H to neutralization byHIV-1LAI Vif (Fig. 4B). In both instances, the infectivity ofHIV-GFP recovered significantly and this effect correspondedto the near disappearance of the A3H-specific band from thecell lysate immunoblot. A similarly large amount of A3H deg-radation was apparent in analogous experiments with HIV Viffrom strain IIIB (data not shown). The only significant differ-ence between the immunoblot experiments shown in Fig. 4Aand those in Fig. 4B is the fact that the former required the useof an anti-HA monoclonal antibody to compare all of the A3sand the latter used an A3H-specific monoclonal antibody todetect the human protein (kindly provided by M. Emerman[24]). To further investigate this suspected difference in anti-body sensitivity, the blot in Fig. 4B was stripped and probedwith the anti-HA antibody and a faint band was detected forthe HA-tagged A3H expressed with HIV Vif (data not shown).So the difference in A3 degradation is most likely linked toantibody sensitivity (with the anti-HA monoclonal antibodybeing more sensitive). Nevertheless, the sum of our resultscombined to indicate that HIV-1 Vif has a notable preferencefor human A3H over other Z3-type A3s. The modest neutral-ization of two of the other A3s may be related to the degra-dation-independent inhibition noted previously for HIV-1 Vif(18, 39, 40).
FIG. 4. Sensitivity of mammalian A3Z3 proteins to HIV-1LAI Vif. (A) Relative infectivity of HIV-GFP produced in the presence of theindicated A3s and a vector control (open bars), HIV-1LAI Vif (black bars), or HIV-1LAI VifSLQ-AAA (gray bars). The double vector control wasnormalized to 1, and the error bars report the standard errors of the means from 3 independent experiments. Producer cell immunoblots are shownbelow for A3 proteins (anti-HA), Vif (anti-myc), and tubulin. (B) The relative infectivity of HIV-GFP produced in the presence of untagged humanA3H (haplotype II) or A3H-HA and a vector control (open bars), HIV-1LAI Vif (black bars), or HIV-1LAI VifSLQ-AAA (gray bars). Producer cellimmunoblots are shown below for A3H (anti-A3H), Vif (anti-myc), and tubulin.
8198 LARUE ET AL. J. VIROL.
on February 9, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
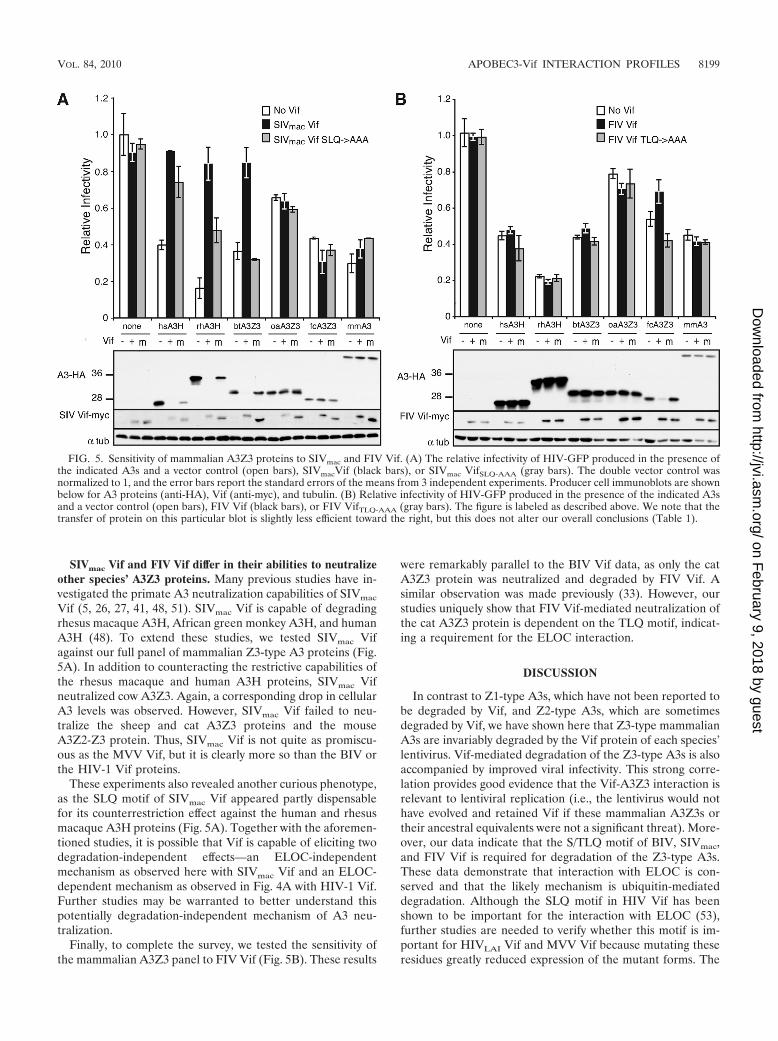
SIVmac Vif and FIV Vif differ in their abilities to neutralizeother species’ A3Z3 proteins. Many previous studies have in-vestigated the primate A3 neutralization capabilities of SIVmac
Vif (5, 26, 27, 41, 48, 51). SIVmac Vif is capable of degradingrhesus macaque A3H, African green monkey A3H, and humanA3H (48). To extend these studies, we tested SIVmac Vifagainst our full panel of mammalian Z3-type A3 proteins (Fig.5A). In addition to counteracting the restrictive capabilities ofthe rhesus macaque and human A3H proteins, SIVmac Vifneutralized cow A3Z3. Again, a corresponding drop in cellularA3 levels was observed. However, SIVmac Vif failed to neu-tralize the sheep and cat A3Z3 proteins and the mouseA3Z2-Z3 protein. Thus, SIVmac Vif is not quite as promiscu-ous as the MVV Vif, but it is clearly more so than the BIV orthe HIV-1 Vif proteins.
These experiments also revealed another curious phenotype,as the SLQ motif of SIVmac Vif appeared partly dispensablefor its counterrestriction effect against the human and rhesusmacaque A3H proteins (Fig. 5A). Together with the aforemen-tioned studies, it is possible that Vif is capable of eliciting twodegradation-independent effects—an ELOC-independentmechanism as observed here with SIVmac Vif and an ELOC-dependent mechanism as observed in Fig. 4A with HIV-1 Vif.Further studies may be warranted to better understand thispotentially degradation-independent mechanism of A3 neu-tralization.
Finally, to complete the survey, we tested the sensitivity ofthe mammalian A3Z3 panel to FIV Vif (Fig. 5B). These results
were remarkably parallel to the BIV Vif data, as only the catA3Z3 protein was neutralized and degraded by FIV Vif. Asimilar observation was made previously (33). However, ourstudies uniquely show that FIV Vif-mediated neutralization ofthe cat A3Z3 protein is dependent on the TLQ motif, indicat-ing a requirement for the ELOC interaction.
DISCUSSION
In contrast to Z1-type A3s, which have not been reported tobe degraded by Vif, and Z2-type A3s, which are sometimesdegraded by Vif, we have shown here that Z3-type mammalianA3s are invariably degraded by the Vif protein of each species’lentivirus. Vif-mediated degradation of the Z3-type A3s is alsoaccompanied by improved viral infectivity. This strong corre-lation provides good evidence that the Vif-A3Z3 interaction isrelevant to lentiviral replication (i.e., the lentivirus would nothave evolved and retained Vif if these mammalian A3Z3s ortheir ancestral equivalents were not a significant threat). More-over, our data indicate that the S/TLQ motif of BIV, SIVmac,and FIV Vif is required for degradation of the Z3-type A3s.These data demonstrate that interaction with ELOC is con-served and that the likely mechanism is ubiquitin-mediateddegradation. Although the SLQ motif in HIV Vif has beenshown to be important for the interaction with ELOC (53),further studies are needed to verify whether this motif is im-portant for HIVLAI Vif and MVV Vif because mutating theseresidues greatly reduced expression of the mutant forms. The
FIG. 5. Sensitivity of mammalian A3Z3 proteins to SIVmac and FIV Vif. (A) The relative infectivity of HIV-GFP produced in the presence ofthe indicated A3s and a vector control (open bars), SIVmacVif (black bars), or SIVmac VifSLQ-AAA (gray bars). The double vector control wasnormalized to 1, and the error bars report the standard errors of the means from 3 independent experiments. Producer cell immunoblots are shownbelow for A3 proteins (anti-HA), Vif (anti-myc), and tubulin. (B) Relative infectivity of HIV-GFP produced in the presence of the indicated A3sand a vector control (open bars), FIV Vif (black bars), or FIV VifTLQ-AAA (gray bars). The figure is labeled as described above. We note that thetransfer of protein on this particular blot is slightly less efficient toward the right, but this does not alter our overall conclusions (Table 1).
VOL. 84, 2010 APOBEC3-Vif INTERACTION PROFILES 8199
on February 9, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

conservation of the Vif-A3Z3 interaction also suggests theexistence of at least one common underlying structural deter-minant, because the lentiviral Vifs are much more divergent atthe amino acid level than the A3Z3s of their mammalian host(Fig. 1).
A second major conclusion to stem from our studies is thatsome lentiviral Vif proteins lack species specificity (summa-rized in Table 1). The best example is MVV Vif, which triggersthe near-complete degradation of the sheep, cow, and catA3Z3 proteins and partial degradation of the human and rhe-sus macaque A3H proteins. MVV Vif also restores or im-proves HIV infectivity in the presence of each mammalianA3Z3 protein. In contrast, the closest relative to this protein,BIV Vif, appears to neutralize only the cow A3Z3 protein. Asecond example is SIVmac Vif, which triggers the degradationof the rhesus macaque, human, and cow A3Z3 proteins andprovides a corresponding improvement in virus infectivity.Again, in contrast, the closest relative to this protein, HIV-1Vif, appears fairly specific to the human A3H protein. It isprobable that had we expanded the survey to include an evenbroader range of mammalian A3s, each lentiviral Vif would beable to trigger the degradation of multiple A3s (i.e., the Vif-A3Z3 interaction appears to be far from species specific). It isfurther probable that Vifs from different lentiviral strainswithin a species will possess a range of A3 neutralization ca-pabilities, as has been indicated by the different strains ofHIV-1 (28).
Another apparent trend in our data set is that Vif may havethe capacity to antagonize the restriction activity of a givenA3Z3 either without triggering its degradation or by utilizing adifferent degradation pathway. First, we observed an increasein HIV-GFP infectivity when HIV-1LAI Vif was coexpressedwith cow A3Z3, sheep A3Z3, or mouse A3Z2-Z3, even in theabsence of detectable protein degradation. It may be possiblethat HIV-1 Vif is interacting in a way that hinders the A3protein’s ability to restrict but is not efficiently recruiting orcorrectly positioning the cellular degradation machinerycomplex (18, 39, 40). Second, we observed that the SIVmac
VifSLQ-AAA mutant is capable of triggering partial degrada-tion of the human and rhesus macaque A3H proteins. It istherefore possible that Vif may bind to A3Z3 proteins andrecruit an alternative cellular complex to induce degrada-tion. Other possibilities are that degradation may depend inpart on species-specific interactions or that the Vif variants
analyzed here may not be fully representative of the manyother variants that exist naturally.
The A3Z3-type deaminase domain can be expressed on itsown and show antiviral activity. It can also be expressed as partof a double-domain deaminase (Z2-Z3) in many mammals,including cattle, sheep, pigs, cats, and mice. The cow, sheep,and cat A3Z2-Z3 proteins all restrict HIV and/or FIV and areneutralized by its host-specific Vif (17, 33, 34). We show in thisstudy that the cow A3Z2 protein is degraded in the presence ofBIV Vif, even though it does not restrict HIV (Fig. 2). It istherefore possible that Vif is able to simultaneously interactwith both types of Z domains (the A3Z2-Z3 protein), resultingin a synergistic neutralization effect. Additional studies will beneeded to systematically address this possibility and test alter-natives such as additive neutralization or competitive binding.
Finally, our data clearly demonstrate that the Vif-A3Z3 in-teraction is not species specific but more likely to be hostoptimized and considerably promiscuous. The idea of a spe-cies-specific A3-Vif interaction arose following seminal workthat mapped a key residue in A3G, D128, which appeared todictate the susceptibility of this protein to Vif-mediated deg-radation (4, 26, 27, 41, 51). Here, our broad survey suggeststhat lentiviral Vifs are optimized to interact with their hostspecies’ A3Z3 proteins because in all instances the lentiviralVifs can effectively degrade and neutralize the A3Z3 of theirhost. However, we also observe that this interaction can behighly versatile and, in multiple instances, Vif is able to rec-ognize and degrade A3 proteins of other species, some fromwhich it is far removed (at least in evolutionary terms [Fig.1A]). Therefore, while Vif invariably functions as an A3 an-tagonist in its host species, its ability to neutralize A3s of otherspecies provides a mechanism to overcome this key barrier tozoonotic transmission. Additional comparative studies arelikely to help map critical interacting regions and inform effortsto develop novel antiviral drugs that leverage the Vif-A3 axis.
ACKNOWLEDGMENTS
We thank J. Albin, J. Hultquist, and V. Pathak for comments on themanuscript; M. Emerman for key reagents; X-F. Yu for sharing un-published data; and J. Weaver for providing technical assistance.
R. LaRue is a student in the Comparative Molecular Bioscienceprogram and is supported in part by a studentship from the MNAgricultural Experiment Station (MAES) and College of VeterinaryMedicine (CVM). This work was funded by a grant from the NationalInstitutes of Health AI064046.
We dedicate this paper to David Derse, who enthusiastically sup-ported our nonhuman A3 studies over the years.
REFERENCES
1. Albin, J. S., and R. S. Harris. 2010. Interactions of host APOBEC3 restric-tion factors with HIV-1 in vivo: implications for therapeutics. Expert Rev.Mol. Med. 12:e4.
2. Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990.Basic local alignment search tool. J. Mol. Biol. 215:403–410.
3. Bininda-Emonds, O. R., M. Cardillo, K. E. Jones, R. D. MacPhee, R. M.Beck, R. Grenyer, S. A. Price, R. A. Vos, J. L. Gittleman, and A. Purvis. 2007.The delayed rise of present-day mammals. Nature 446:507–512.
4. Bogerd, H. P., B. P. Doehle, H. L. Wiegand, and B. R. Cullen. 2004. A singleamino acid difference in the host APOBEC3G protein controls the primatespecies specificity of HIV type 1 virion infectivity factor. Proc. Natl. Acad.Sci. U. S. A. 101:3770–3774.
5. Bogerd, H. P., R. L. Tallmadge, J. L. Oaks, S. Carpenter, and B. R. Cullen.2008. Equine infectious anemia virus resists the antiretroviral activity ofequine APOBEC3 proteins through a packaging-independent mechanism.J. Virol. 82:11889–11901.
6. Conticello, S. G., R. S. Harris, and M. S. Neuberger. 2003. The Vif protein
TABLE 1. Qualitative summary of Vif-mediated A3 degradationand rescue of HIV-GFP infectivity from A3 restriction
A3Degradationa Rescue of infectionb
BIV MVV HIV SIVmac FIV BIV MVV HIV SIVmac FIV
btA3Z3 �� �� � �� � ** ** * ** �oaA3Z3 � �� � � � � ** ** � �hsA3H � �� � �� � � * ** ** �rhA3H � �� � �� � � * � ** �fcA3Z3 � �� � � �� � * � � *mmA3 � � � � � � � * � �hsA3G � �� ��d NDc ND � � **d ND ND
a A3 degradation: ��, 50 to 100%; �, 5 to 49%; �, 0 to 4.9%.b HIV infectivity: **, complete rescue; *, partial rescue; �, no rescue.c ND, not done.d Data not shown.
8200 LARUE ET AL. J. VIROL.
on February 9, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

of HIV triggers degradation of the human antiretroviral DNA deaminaseAPOBEC3G. Curr. Biol. 13:2009–2013.
7. Conticello, S. G., C. J. Thomas, S. K. Petersen-Mahrt, and M. S. Neuberger.2005. Evolution of the AID/APOBEC family of polynucleotide (deoxy)cyti-dine deaminases. Mol. Biol. Evol. 22:367–377.
8. Dang, Y., L. M. Siew, X. Wang, Y. Han, R. Lampen, and Y. H. Zheng. 2008.Human cytidine deaminase APOBEC3H restricts HIV-1 replication. J. Biol.Chem. 283:11606–11614.
9. Gifford, R. J., A. Katzourakis, M. Tristem, O. G. Pybus, M. Winters, andR. W. Shafer. 2008. A transitional endogenous lentivirus from the genome ofa basal primate and implications for lentivirus evolution. Proc. Natl. Acad.Sci. U. S. A. 105:20362–20367.
10. Goila-Gaur, R., and K. Strebel. 2008. HIV-1 Vif, APOBEC, and intrinsicimmunity. Retrovirology 5:51.
11. Hache, G., M. T. Liddament, and R. S. Harris. 2005. The retroviral hyper-mutation specificity of APOBEC3F and APOBEC3G is governed by theC-terminal DNA cytosine deaminase domain. J. Biol. Chem. 280:10920–10924.
12. Harari, A., M. Ooms, L. C. Mulder, and V. Simon. 2009. Polymorphisms andsplice variants influence the antiretroviral activity of human APOBEC3H.J. Virol. 83:295–303.
13. Harris, R. S., S. K. Petersen-Mahrt, and M. S. Neuberger. 2002. RNAediting enzyme APOBEC1 and some of its homologs can act as DNAmutators. Mol. Cell 10:1247–1253.
14. Henikoff, S., and J. G. Henikoff. 1992. Amino acid substitution matrices fromprotein blocks. Proc. Natl. Acad. Sci. U. S. A. 89:10915–10919.
15. Iwatani, Y., D. S. Chan, L. Liu, H. Yoshii, J. Shibata, N. Yamamoto, J. G.Levin, A. M. Gronenborn, and W. Sugiura. 2009. HIV-1 Vif-mediated ubiq-uitination/degradation of APOBEC3G involves four critical lysine residuesin its C-terminal domain. Proc. Natl. Acad. Sci. U. S. A. 106:19539–19544.
16. Jarmuz, A., A. Chester, J. Bayliss, J. Gisbourne, I. Dunham, J. Scott, and N.Navaratnam. 2002. An anthropoid-specific locus of orphan C to U RNA-editing enzymes on chromosome 22. Genomics 79:285–296.
17. Jonsson, S. R., G. Hache, M. D. Stenglein, S. C. Fahrenkrug, V. Andresdot-tir, and R. S. Harris. 2006. Evolutionarily conserved and non-conservedretrovirus restriction activities of artiodactyl APOBEC3F proteins. NucleicAcids Res. 34:5683–5694.
18. Kao, S., R. Goila-Gaur, E. Miyagi, M. A. Khan, S. Opi, H. Takeuchi, and K.Strebel. 2007. Production of infectious virus and degradation ofAPOBEC3G are separable functional properties of human immunodefi-ciency virus type 1 Vif. Virology 369:329–339.
19. Katzourakis, A., M. Tristem, O. G. Pybus, and R. J. Gifford. 2007. Discoveryand analysis of the first endogenous lentivirus. Proc. Natl. Acad. Sci. U. S. A.104:6261–6265.
20. Kobayashi, M., A. Takaori-Kondo, Y. Miyauchi, K. Iwai, and T. Uchiyama.2005. Ubiquitination of APOBEC3G by an HIV-1 Vif-Cullin5-Elongin B-Elongin C complex is essential for Vif function. J. Biol. Chem. 280:18573–18578.
21. Kobayashi, M., A. Takaori-Kondo, K. Shindo, A. Abudu, K. Fukunaga, andT. Uchiyama. 2004. APOBEC3G targets specific virus species. J. Virol.78:8238–8244.
22. LaRue, R. S., V. Andresdottir, Y. Blanchard, S. G. Conticello, D. Derse, M.Emerman, W. C. Greene, S. R. Jonsson, N. R. Landau, M. Lochelt, H. S.Malik, M. H. Malim, C. Munk, S. J. O’Brien, V. K. Pathak, K. Strebel, S.Wain-Hobson, X. F. Yu, N. Yuhki, and R. S. Harris. 2009. Guidelines fornaming nonprimate APOBEC3 genes and proteins. J. Virol. 83:494–497.
23. LaRue, R. S., S. R. Jonsson, K. A. Silverstein, M. Lajoie, D. Bertrand, N.El-Mabrouk, I. Hotzel, V. Andresdottir, T. P. Smith, and R. S. Harris. 2008.The artiodactyl APOBEC3 innate immune repertoire shows evidence for amulti-functional domain organization that existed in the ancestor of placen-tal mammals. BMC Mol. Biol. 9:104.
24. Li, M. M., L. I. Wu, and M. Emerman. 2010. The range of humanAPOBEC3H sensitivity to lentiviral Vif proteins. J. Virol. 84:88–95.
25. Luo, K., Z. Xiao, E. Ehrlich, Y. Yu, B. Liu, S. Zheng, and X. F. Yu. 2005.Primate lentiviral virion infectivity factors are substrate receptors that as-semble with cullin 5-E3 ligase through a HCCH motif to suppressAPOBEC3G. Proc. Natl. Acad. Sci. U. S. A. 102:11444–11449.
26. Mangeat, B., P. Turelli, S. Liao, and D. Trono. 2004. A single amino aciddeterminant governs the species-specific sensitivity of APOBEC3G to Vifaction. J. Biol. Chem. 279:14481–14483.
27. Mariani, R., D. Chen, B. Schrofelbauer, F. Navarro, R. Konig, B. Bollman,C. Munk, H. Nymark-McMahon, and N. R. Landau. 2003. Species-specificexclusion of APOBEC3G from HIV-1 virions by Vif. Cell 114:21–31.
28. Marin, M., S. Golem, K. M. Rose, S. L. Kozak, and D. Kabat. 2008.Human immunodeficiency virus type 1 Vif functionally interacts withdiverse APOBEC3 cytidine deaminases and moves with them betweencytoplasmic sites of mRNA metabolism. J. Virol. 82:987–998.
29. Marin, M., K. M. Rose, S. L. Kozak, and D. Kabat. 2003. HIV-1 Vif proteinbinds the editing enzyme APOBEC3G and induces its degradation. Nat.Med. 9:1398–1403.
30. Mehle, A., J. Goncalves, M. Santa-Marta, M. McPike, and D. Gabuzda.
2004. Phosphorylation of a novel SOCS-box regulates assembly of the HIV-1Vif-Cul5 complex that promotes APOBEC3G degradation. Genes Dev. 18:2861–2866.
31. Mehle, A., B. Strack, P. Ancuta, C. Zhang, M. McPike, and D. Gabuzda.2004. Vif overcomes the innate antiviral activity of APOBEC3G by promot-ing its degradation in the ubiquitin-proteasome pathway. J. Biol. Chem.279:7792–7798.
32. Miyoshi, H., U. Blomer, M. Takahashi, F. H. Gage, and I. M. Verma. 1998.Development of a self-inactivating lentivirus vector. J. Virol. 72:8150–8157.
33. Munk, C., T. Beck, J. Zielonka, A. Hotz-Wagenblatt, S. Chareza, M. Bat-tenberg, J. Thielebein, K. Cichutek, I. G. Bravo, S. J. O’Brien, M. Lochelt,and N. Yuhki. 2008. Functions, structure, and read-through alternative splic-ing of feline APOBEC3 genes. Genome Biol. 9:R48.
34. Munk, C., J. Zielonka, H. Constabel, B. P. Kloke, B. Rengstl, M. Battenberg,F. Bonci, M. Pistello, M. Lochelt, and K. Cichutek. 2007. Multiple restric-tions of human immunodeficiency virus type 1 in feline cells. J. Virol. 81:7048–7060.
35. Notredame, C., D. G. Higgins, and J. Heringa. 2000. T-Coffee: a novelmethod for fast and accurate multiple sequence alignment. J. Mol. Biol.302:205–217.
36. Oberste, M. S., and M. A. Gonda. 1992. Conservation of amino-acid se-quence motifs in lentivirus Vif proteins. Virus Genes 6:95–102.
37. OhAinle, M., J. A. Kerns, M. M. Li, H. S. Malik, and M. Emerman. 2008.Antiretroelement activity of APOBEC3H was lost twice in recent humanevolution. Cell Host Microbe 4:249–259.
38. OhAinle, M., J. A. Kerns, H. S. Malik, and M. Emerman. 2006. Adaptiveevolution and antiviral activity of the conserved mammalian cytidine deami-nase APOBEC3H. J. Virol. 80:3853–3862.
39. Opi, S., S. Kao, R. Goila-Gaur, M. A. Khan, E. Miyagi, H. Takeuchi, and K.Strebel. 2007. Human immunodeficiency virus type 1 Vif inhibits packagingand antiviral activity of a degradation-resistant APOBEC3G variant. J. Virol.81:8236–8246.
40. Santa-Marta, M., F. A. da Silva, A. M. Fonseca, and J. Goncalves. 2005.HIV-1 Vif can directly inhibit apolipoprotein B mRNA-editing enzyme cat-alytic polypeptide-like 3G-mediated cytidine deamination by using a singleamino acid interaction and without protein degradation. J. Biol. Chem.280:8765–8775.
41. Schrofelbauer, B., D. Chen, and N. R. Landau. 2004. A single amino acid ofAPOBEC3G controls its species-specific interaction with virion infectivityfactor (Vif). Proc. Natl. Acad. Sci. U. S. A. 101:3927–3932.
42. Shacklett, B. L., and P. A. Luciw. 1994. Analysis of the vif gene of felineimmunodeficiency virus. Virology 204:860–867.
43. Sheehy, A. M., N. C. Gaddis, J. D. Choi, and M. H. Malim. 2002. Isolationof a human gene that inhibits HIV-1 infection and is suppressed by the viralVif protein. Nature 418:646–650.
44. Sheehy, A. M., N. C. Gaddis, and M. H. Malim. 2003. The antiretroviralenzyme APOBEC3G is degraded by the proteasome in response to HIV-1Vif. Nat. Med. 9:1404–1407.
45. Smith, J. L., W. Bu, R. C. Burdick, and V. K. Pathak. 2009. Multiple ways oftargeting APOBEC3-virion infectivity factor interactions for anti-HIV-1drug development. Trends Pharmacol. Sci. 30:638–646.
46. Stopak, K., C. de Noronha, W. Yonemoto, and W. C. Greene. 2003. HIV-1Vif blocks the antiviral activity of APOBEC3G by impairing both its trans-lation and intracellular stability. Mol. Cell 12:591–601.
47. Tan, L., P. T. Sarkis, T. Wang, C. Tian, and X. F. Yu. 2009. Sole copy ofZ2-type human cytidine deaminase APOBEC3H has inhibitory activityagainst retrotransposons and HIV-1. FASEB J. 23:279–287.
48. Virgen, C. A., and T. Hatziioannou. 2007. Antiretroviral activity and Vifsensitivity of rhesus macaque APOBEC3 proteins. J. Virol. 81:13932–13937.
49. Wedekind, J. E., G. S. Dance, M. P. Sowden, and H. C. Smith. 2003. Mes-senger RNA editing in mammals: new members of the APOBEC familyseeking roles in the family business. Trends Genet. 19:207–216.
50. Wiegand, H. L., B. P. Doehle, H. P. Bogerd, and B. R. Cullen. 2004. A secondhuman antiretroviral factor, APOBEC3F, is suppressed by the HIV-1 andHIV-2 Vif proteins. EMBO J. 23:2451–2458.
51. Xu, H., E. S. Svarovskaia, R. Barr, Y. Zhang, M. A. Khan, K. Strebel, andV. K. Pathak. 2004. A single amino acid substitution in human APOBEC3Gantiretroviral enzyme confers resistance to HIV-1 virion infectivity factor-induced depletion. Proc. Natl. Acad. Sci. U. S. A. 101:5652–5657.
52. Yu, X., Y. Yu, B. Liu, K. Luo, W. Kong, P. Mao, and X. F. Yu. 2003. Inductionof APOBEC3G ubiquitination and degradation by an HIV-1 Vif-Cul5-SCFcomplex. Science 302:1056–1060.
53. Yu, Y., Z. Xiao, E. S. Ehrlich, X. Yu, and X. F. Yu. 2004. Selective assemblyof HIV-1 Vif-Cul5-ElonginB-ElonginC E3 ubiquitin ligase complex througha novel SOCS box and upstream cysteines. Genes Dev. 18:2867–2872.
54. Zhen, A., T. Wang, K. Zhao, Y. Xiong, and X. F. Yu. 2010. A single aminoacid difference in human APOBEC3H variants determines HIV-1 Vif sen-sitivity. J. Virol. 84:1902–1911.
55. Zielonka, J., I. G. Bravo, D. Marino, E. Conrad, M. Perkovic, M. Battenberg,K. Cichutek, and C. Munk. 2009. Restriction of equine infectious anemiavirus by equine APOBEC3 cytidine deaminases. J. Virol. 83:7547–7559.
VOL. 84, 2010 APOBEC3-Vif INTERACTION PROFILES 8201
on February 9, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from





























