Embed Size (px)
Citation preview

• Lecture 6
• Intracellular Compartments
• and Protein Sorting

Membrane-enclosedcompartments
Proteins:enzymes,transporter andsurface markers
10,000-20,000proteins deliveredto different compartments

Major intracellular compartmentsVital chemical reactions take place in or on membrane surface
Compartments increase surface and isolate reactions

Microtubules helpthe localizationof the ER and theGolgi apparatus
Bacteria have noInternal membranes
Eukaryotic cells are 1000-10,000 timesgreater--need internalmembranes

Development ofplastids

topology

Topologically equivalent spaces are shown in red

Gated transport
Transmembrane transport
Vesicular transport
Transport is highly regulated


Signal sequence and signal patch



50 nucleoporinsOctagonal
Variable numbers ofpores (3000-4000)depending on TXN
100 histone moleculesper minute per pore
6 large and small ribosomalsubunits per minute per pore
SEM
“Basket”


Results from injection:
<5000 Daltons: fast diffusion17 Kd: 2 minutes>60 Kd: cannot enter
Channel is 9 nm in diameter15 nm long
Ribosome 30 nm
DNA, RNA polymerases100-200 Kd subunits

One or two short sequencesRich in positively charged aaLys, Arg
Immunofluorescencemicrographs showing T-antigen localization
An experiment usingrecombinant DNA technique

Gold particles coatedwith nuclear localizationsignals
Pore dilates to 26 nm
Not throughlipid bilayer
Folded confomration
Nuclear importReceptors!!!
Bind to nucleoporins
FG-repeats

Nuclear export signalsNuclear export receptors
Nuclear transport receptors (karyopherins)
A single pore complex conducts traffic in both directions
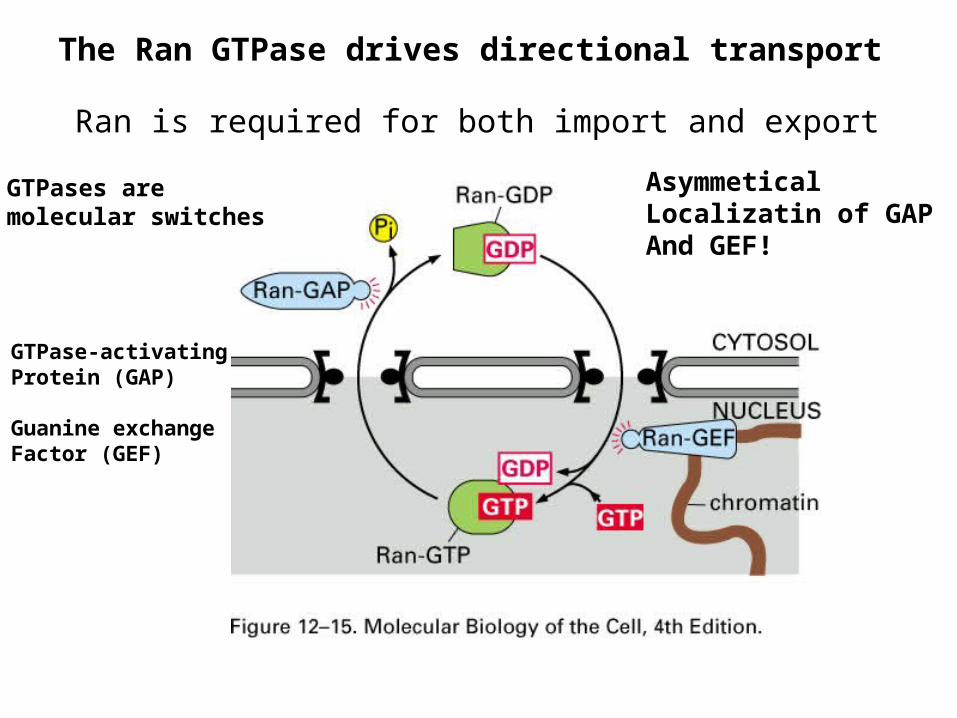
The Ran GTPase drives directional transport
Ran is required for both import and export
GTPases aremolecular switches
GTPase-activatingProtein (GAP)
Guanine exchangeFactor (GEF)
AsymmeticalLocalizatin of GAPAnd GEF!

Ran-GTP causes cargorelease of import receptor
Ran-GTP causes cargobinding of export receptor
Free export receptorsreturn to the nucleusGTP-bound import receptors return to the cytosol

Nuclear localization of TXN factors control gene expression

The nuclear lamina
Meshwork of interconnected protein subunits, nuclear lamins
Intermediate filament proteins, interact with nuclear pore complexesand integral membrane proteins, chromatin

NLS is not cleaved offafter transport--repeatedimport


The subcompartments of mitochondria and chloroplasts

Mitochondrial proteinsare first fully synthesized:different from proteinstransported into ER
Signal sequence:Amphipathic helix
Signal peptidase

Translocases of the outer and inner mito membranes

Proteins transiently spanning the inner and outer membranesduring their translocation into the matrix
Precursor proteins remain unfoldedbefore transport

Protein import by mitochondria

ATP hydrolyses at two sites plus a H+ gradient across inner membrane
Release from cytosolic hsp 70
Further translocation through TIM requires H+ gradient
Signal peptide is positively charged
Release from mito hsp70

Two models of how mito hsp70 could drive protein transport

Proteins destined for various mito space

Two signal sequencesare required for proteinsdirected to the thylakoidmembrane in chloroplasts
Four routesinto the thylakoidspace
Signal sequences formito and chloroplastsare different
GTP and ATP

Urate oxidase
Peroxisomes have one single membrane
No DNA or ribosomes
Catalase and urate oxidaseOxidative reactions not taken over by mito
RH2+O2->R+H2O2
H2O2+R’H2->R’+2H2OCatalase
Urate oxidate(R=uric acid)
-oxidation
biosynthesis of plasmalogens
photorespiration
glyoxylate cycle

A model of how new peroxisomes are produced
From preexistingperoxisomes
growth
fission
Transport mechanismsunknown: no unfolding necessary
23 peroxinsSimilar to nuclear transport

Summary
1. Cells are highly compartmentalized; proteins are sortedto different compartments;
2. Nuclear transport, nuclear pore, nucleoporins, NLS;3. Ran GTPase control direction;4. Nuclear lamina, nuclear lamins;5. Mitochondria transport, signal sequence, TOM, TIM,
energy;6. Chloroplast transport, thylakoid;7. Peroxisomes, structure, function, transport, biogenesis.






























